Influences of the corticotropic axis and sympathetic activity on neurochemical consequences of 3,4- methylenedioxymethamphetamine (MDMA) administration in Fischer 344 rats Francesca Fernandez, Sylvie Aguerre, Pierre Morme `de and Francis Chaouloff NeuroGe ´ne ´ tique et Stress, INSERM U471-INRA, Institut F. Magendie, Rue Camille Saint Sae ¨ ns, 33077 Bordeaux Ce ´ dex, France Keywords: adrenalectomy, body temperature, chlorisondamine, hippocampus, serotonergic systems, striatum Abstract The respective influences of the corticotropic axis and sympathetic activity on 3,4-methylenedioxymethamphetamine (MDMA, ecstasy) immediate effects on body temperature and long-term neurotoxicity, as assessed by decreases in hippocampal and striatal [ 3 H]5-hydroxytryptamine ([ 3 H]5-HT) reuptake, [ 3 H]paroxetine binding at 5-HT transporters (5-HTT), and 5-HT and 5-hydroxyindoleacetic acid (5-HIAA) levels, were examined in Fischer 344 rats. On each of the two injections of MDMA (5 or 10 mg/kg s.c. once a day for 2 consecutive days) body temperature rapidly increased in a dose-dependent manner. Six days after the last injection of 10 mg/kg MDMA, [ 3 H]5-HT reuptake, [ 3 H]paroxetine binding and 5-HT and 5-HIAA levels were decreased in the hippocampus and, to a lower extent, in striatum. Prior adrenalectomy (1 week beforehand), which weakened the immediate hyperthermic effect of MDMA, prevented the long-term MDMA-elicited reduction in hippocampal and striatal [ 3 H]paroxetine binding. Supplementation of adrenalectomised Fischer 344 rats with corticosterone almost reinstated the immediate hyperthermic effect of MDMA and restored MDMA-elicited reduction in hippocampal and striatal [ 3 H]paroxetine binding. In a final set of experiments, Fischer 344 rats were pretreated (30 min before each of the two injections of 10 mg/kg MDMA) with the ganglionic blocker chlorisondamine (2.5 mg/kg). This pretreatment markedly reduced the amplitudes of the immediate hyperthermia and long-term declines in hippocampal [ 3 H]5-HT reuptake and [ 3 H]paroxetine binding at 5-HTT, and in hippocampal and striatal 5-HT and 5-HIAA levels. These results suggest that sympathetic activity (possibly through its control of body temperature), but not corticotropic activity, plays a key role in MDMA-elicited neurotoxicity in Fischer 344 rats. Introduction Administration of 3,4-methylenedioxymethamphetamine (MDMA, ‘ecstasy’) to rats, guinea pigs and nonhuman primates produces, in the long term, lasting reductions in markers of serotonergic neurons (Battaglia et al., 1987; Commins et al., 1987; Battaglia et al., 1988; Ricaurte et al., 1988; De Souza et al., 1990). These markers include the concentrations of 5-hydroxytryptamine (serotonin, 5-HT) and its metabolite 5-hydroxyindoleacetic acid (5-HIAA), tryptophan hydroxylase activity and the density and the reuptake activity of plasma membrane 5-HT transporters (5-HTT). Taken with morpho- logical evidence for an MDMA-elicited degeneration of fine 5-HT nerve terminals (Commins et al., 1987; Molliver et al., 1990), it is now believed that the reductions in these markers reflect neuro- toxicity (Green et al., 1995; Sprague et al., 1998). The mechanisms through which MDMA is neurotoxic to serotonergic systems are only partly identified. There is evidence for an initial 5-HTT-mediated reuptake of MDMA into serotonergic nerve terminals followed by 5-HT release from storage vesicles (Schmidt et al., 1987; Steele et al., 1987; Rudnick & Wall, 1992). That pretreatment with a 5-HT 2A receptor antagonist prevents the long-term neurotoxic impact of MDMA (Schmidt & Kehne, 1990; Schmidt et al., 1990) suggests that direct or indirect (through MDMA metabolites) release of 5-HT and, in turn, 5-HT 2A receptor stimulation, plays a role in neurotoxicity. It has been proposed that MDMA-elicited release of dopamine (Yamamoto & Spanos, 1988; Nash, 1990; Schmidt et al., 1994; White et al., 1994) leads to its uptake by serotonergic nerve terminals where it is deaminated to generate oxidative processes (Schmidt & Kehne, 1990; Sprague et al., 1998). Furthermore, the formation of MDMA metabolites which may generate free radicals may be another mechanism leading to oxidative stress in serotonergic nerve terminals (Schmidt & Kehne, 1990; Green et al., 1995; Sprague et al., 1998). This theory has received support from the measurement of free radicals (Colado et al., 1997), and the protective effects of spin-trap reagents and metabolic antioxidants (Schmidt & Kehne, 1990; Colado & Green, 1995; Gudelsky, 1996; Aguirre et al., 1997; Colado et al., 1997). Lastly, MDMA-elicited hyperthermia, which may favour the generation of free radicals (Globus et al., 1995), has also received much attention because environment- and drug-induced decreases in core temperature prior to MDMA injection may trigger neuroprotection (Miller & O’Callaghan, 1994; Farfel & Seiden, 1995; Malberg et al., 1996; Malberg & Seiden, 1998; Colado et al., 1999). This result then suggests that drugs endowed with sympatholytic activity should bear neuroprotective properties against MDMA-elicited neurotoxicity; however, to our knowledge, such a possibility has not been tested so far. Correspondence: Dr Francis Chaouloff, as above. E-mail: [email protected] Received 13 March 2002, revised 6 June 2002, accepted 10 June 2002 doi:10.1046/j.1460-9568.2002.02110.x European Journal of Neuroscience, Vol. 16, pp. 607–618, 2002 ª Federation of European Neuroscience Societies

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

In¯uences of the corticotropic axis and sympatheticactivity on neurochemical consequences of 3,4-methylenedioxymethamphetamine (MDMA) administrationin Fischer 344 rats
Francesca Fernandez, Sylvie Aguerre, Pierre MormeÁde and Francis ChaouloffNeuroGeÂneÂtique et Stress, INSERM U471-INRA, Institut F. Magendie, Rue Camille Saint SaeÈns, 33077 Bordeaux CeÂdex, France
Keywords: adrenalectomy, body temperature, chlorisondamine, hippocampus, serotonergic systems, striatum
Abstract
The respective in¯uences of the corticotropic axis and sympathetic activity on 3,4-methylenedioxymethamphetamine (MDMA,ecstasy) immediate effects on body temperature and long-term neurotoxicity, as assessed by decreases in hippocampal and
striatal [3H]5-hydroxytryptamine ([3H]5-HT) reuptake, [3H]paroxetine binding at 5-HT transporters (5-HTT), and 5-HT and
5-hydroxyindoleacetic acid (5-HIAA) levels, were examined in Fischer 344 rats. On each of the two injections of MDMA (5 or10 mg/kg s.c. once a day for 2 consecutive days) body temperature rapidly increased in a dose-dependent manner. Six days
after the last injection of 10 mg/kg MDMA, [3H]5-HT reuptake, [3H]paroxetine binding and 5-HT and 5-HIAA levels were
decreased in the hippocampus and, to a lower extent, in striatum. Prior adrenalectomy (1 week beforehand), which weakened theimmediate hyperthermic effect of MDMA, prevented the long-term MDMA-elicited reduction in hippocampal and striatal
[3H]paroxetine binding. Supplementation of adrenalectomised Fischer 344 rats with corticosterone almost reinstated the
immediate hyperthermic effect of MDMA and restored MDMA-elicited reduction in hippocampal and striatal [3H]paroxetine
binding. In a ®nal set of experiments, Fischer 344 rats were pretreated (30 min before each of the two injections of 10 mg/kgMDMA) with the ganglionic blocker chlorisondamine (2.5 mg/kg). This pretreatment markedly reduced the amplitudes of the
immediate hyperthermia and long-term declines in hippocampal [3H]5-HT reuptake and [3H]paroxetine binding at 5-HTT, and in
hippocampal and striatal 5-HT and 5-HIAA levels. These results suggest that sympathetic activity (possibly through its control ofbody temperature), but not corticotropic activity, plays a key role in MDMA-elicited neurotoxicity in Fischer 344 rats.
Introduction
Administration of 3,4-methylenedioxymethamphetamine (MDMA,
`ecstasy') to rats, guinea pigs and nonhuman primates produces, in
the long term, lasting reductions in markers of serotonergic neurons
(Battaglia et al., 1987; Commins et al., 1987; Battaglia et al., 1988;
Ricaurte et al., 1988; De Souza et al., 1990). These markers include
the concentrations of 5-hydroxytryptamine (serotonin, 5-HT) and its
metabolite 5-hydroxyindoleacetic acid (5-HIAA), tryptophan
hydroxylase activity and the density and the reuptake activity of
plasma membrane 5-HT transporters (5-HTT). Taken with morpho-
logical evidence for an MDMA-elicited degeneration of ®ne 5-HT
nerve terminals (Commins et al., 1987; Molliver et al., 1990), it is
now believed that the reductions in these markers re¯ect neuro-
toxicity (Green et al., 1995; Sprague et al., 1998).
The mechanisms through which MDMA is neurotoxic to
serotonergic systems are only partly identi®ed. There is evidence
for an initial 5-HTT-mediated reuptake of MDMA into serotonergic
nerve terminals followed by 5-HT release from storage vesicles
(Schmidt et al., 1987; Steele et al., 1987; Rudnick & Wall, 1992).
That pretreatment with a 5-HT2A receptor antagonist prevents the
long-term neurotoxic impact of MDMA (Schmidt & Kehne, 1990;
Schmidt et al., 1990) suggests that direct or indirect (through MDMA
metabolites) release of 5-HT and, in turn, 5-HT2A receptor stimulation,
plays a role in neurotoxicity. It has been proposed that MDMA-elicited
release of dopamine (Yamamoto & Spanos, 1988; Nash, 1990;
Schmidt et al., 1994; White et al., 1994) leads to its uptake by
serotonergic nerve terminals where it is deaminated to generate
oxidative processes (Schmidt & Kehne, 1990; Sprague et al., 1998).
Furthermore, the formation of MDMA metabolites which may
generate free radicals may be another mechanism leading to oxidative
stress in serotonergic nerve terminals (Schmidt & Kehne, 1990; Green
et al., 1995; Sprague et al., 1998). This theory has received support
from the measurement of free radicals (Colado et al., 1997), and the
protective effects of spin-trap reagents and metabolic antioxidants
(Schmidt & Kehne, 1990; Colado & Green, 1995; Gudelsky, 1996;
Aguirre et al., 1997; Colado et al., 1997). Lastly, MDMA-elicited
hyperthermia, which may favour the generation of free radicals
(Globus et al., 1995), has also received much attention because
environment- and drug-induced decreases in core temperature prior to
MDMA injection may trigger neuroprotection (Miller & O'Callaghan,
1994; Farfel & Seiden, 1995; Malberg et al., 1996; Malberg & Seiden,
1998; Colado et al., 1999). This result then suggests that drugs
endowed with sympatholytic activity should bear neuroprotective
properties against MDMA-elicited neurotoxicity; however, to our
knowledge, such a possibility has not been tested so far.
Correspondence: Dr Francis Chaouloff, as above.E-mail: [email protected]
Received 13 March 2002, revised 6 June 2002, accepted 10 June 2002
doi:10.1046/j.1460-9568.2002.02110.x
European Journal of Neuroscience, Vol. 16, pp. 607±618, 2002 ã Federation of European Neuroscience Societies

It has been suggested that the corticotropic axis also plays a role in
neurotoxic effects of MDMA. Thus, MDMA stimulates cortico-
sterone release (Nash et al., 1988) and alters corticoid receptor gene
expression, especially in the hippocampus (Yau et al., 1994).
Adrenalectomy, but not adrenalectomy combined with corticosterone
supplementation, prevents or attenuates the long-term decreases in
hippocampal tryptophan hydroxylase activity (and 5-HT and 5-HIAA
levels) elicited, respectively, by single 10 or 20 mg/kg doses of
MDMA (Johnson et al., 1989). This result raises three questions: ®rst,
does the impact of adrenalectomy on these markers extend to other
indices of neurotoxicity (5-HTT binding and/or reuptake activity)?
This question is crucial as corticoids are permissive to 5-HT synthesis
(see Chaouloff, 1993), thus opening the possibility that MDMA-
elicited corticosterone release affects hippocampal 5-HT synthesis
but not the integrity of hippocampal serotonergic neurons. Second, if
corticoids mediate the long-term neurotoxic effects of MDMA, is it
possible to anticipate such a result by the measurement of some
immediate effects of MDMA, e.g. on body temperature? Third, the
study of Johnson et al. (1989) used Sprague-Dawley rats, i.e. rats
endowed with a corticotropic axis thought to be normoresponsive
compared to other strains (Aulakh et al., 1988; Sternberg et al., 1992;
Dhabbar et al., 1993). Thus it would be expected that the
corticotropic axis would impact even more on MDMA-induced
degeneration of hippocampal 5-HT neurons if this axis were
hyperactive and/or hyperreactive. The Fischer 344 (F344) rat strain,
which bears such hyperactivity and hyperesponsiveness of the
corticotropic axis (Sternberg et al., 1989; Calogero et al., 1992;
Sternberg et al., 1992; Dhabbar et al., 1993), should be suitable for
testing the latter hypothesis.
With these questions in mind, we ®rst analysed the immediate body
temperature and long-term neurochemical effects of MDMA in F344
rats. To address the issue of the long-term neurochemical conse-
quences of MDMA administration, we focused our attention on
hippocampal [3H]5-HT reuptake, hippocampal and striatal
[3H]paroxetine binding at 5-HTT, and hippocampal and striatal
5-HT and 5-HIAA levels. Then, to investigate the role of the
corticotropic axis in the long-term neurochemical effects of MDMA,
we measured the respective impacts of adrenalectomy with and
without corticosterone supplementation on the aforementioned
neurochemical variables to which we added striatal [3H]5-HT
reuptake analyses. In keeping with the observation that all these
manipulations had consequences on the immediate body temperature
responses to MDMA, we took advantage of the lack of information as
to the putative neuroprotective properties of sympatholytics (see
above) to analyse, by means of the ganglionic blocker chlorisond-
amine, the extent to which sympathetic activity had also an in¯uence
on the long-term neurochemical effects of MDMA.
Materials and methods
Animals and housing conditions
Inbred male F344 rats (IFFA CREDO, Les Oncins, France), aged
5 weeks on arrival, were housed four per cage under constant
temperature (22 6 1 °C) and a 12-h light : 12-h dark cycle (lights on
at 07.00 h), with food and water ad libitum. All rats were tested at
least 2 weeks after their arrival, except for rats undergoing sham
surgery or adrenalectomy, which were tested at least 3 weeks after
their arrival (1 week after surgery). All efforts were made to
minimize the number of animals used, and procedures were as
established by the French legislation on animal welfare (O.J. no. 87±
848).
MDMA administration
After weighing, rats were injected s.c. with either saline or MDMA,
this protocol being repeated 24 h later. Seven days after the ®rst of
the two injections, the rats were killed by cervical dislocation, the
brains removed and the hippocampi and the striata rapidly dissected
out. In the ®rst series of experiments, 5 and 10 mg/kg doses of
MDMA were used, whereas only the highest dose was used
thereafter. Note that in the last series of experiments, rats were
pretreated 30 min beforehand with saline or chlorisondamine
(2.5 mg/kg i.p.) before each of the two injections of saline or
MDMA. This dose of chlorisondamine has been shown to fully block
sympathetic activity (McCarty & Kopin, 1979) without altering the
vigilance state of the animal (Chaouloff et al., 1994).
Surgery and plasma corticosterone analyses
In one series of experiments, rats were anaesthetized with a 60-mg/kg
dose of pentobarbitone sodium, and the adrenals visualized (sham
rats) or removed (adrenalectomised rats) through bilateral incision in
the ¯anks. In a second series of experiments, a similar procedure was
followed, except that half of the adrenalectomised rats were at the
same time implanted with subcutaneous corticosterone pellets. For
this purpose, powdered corticosterone was mixed with cholesterol
(1 : 1), melted and poured into wax moulds designed for 2-week
release, as previously reported (Cador et al., 1993; Kulikov et al.,
1997). The weight of the pellet contents was adjusted to 100 mg
(50 mg corticosterone). Following surgery, the animals were returned
to their home cages with tap water containing (adrenalectomised
rats) or not (sham rats) 0.9% NaCl (saline). In the two series of
experiments, rats were injected twice with saline or MDMA 7 and
8 days after surgery, and killed another week after the ®rst injection.
In the second series of experiments, thymus weights as well as
circulating levels of corticosterone were analysed at the time of
dissection. For corticosterone analyses, trunk blood was collected in
EDTA-coated tubes and centrifuged. Plasma corticosterone levels
were assessed by competitive protein binding following extraction
with absolute ethanol using rhesus monkey serum as the source of
transcortin and [3H]corticosterone as the radioligand (Cador et al.,
1993; Kulikov et al., 1997).
Drugs and chemicals
MDMA hydrochloride was generously supplied by the National
Institute on Drug Abuse (Bethesda, MD, USA). Corticosterone,
cholesterol, pargyline hydrochloride, 5-HT hydrochloride and
5-HIAA were purchased from Sigma Chemicals (St Quentin
Fallavier, France). Fluoxetine hydrochloride and citalopram hydro-
chloride were, respectively, obtained from Mediat (Milan, Italy) and
H. Lundbeck A/S (Copenhagen, Denmark); chlorisondamine hydro-
chloride was obtained from Ciba-Geigy (now Novartis, Basel,
Switzerland). [3H]5-HT creatinine sulphate (21.4 Ci/mmol),
[3H]paroxetine (19.7 Ci/mmol) and [3H]corticosterone (76.5 Ci/
mmol) were all purchased from New England Nuclear (Paris,
France).
Neurochemical assays
[3H]5-HT reuptake assays were performed as previously described
(Pollier et al., 2000). Thus, fresh hippocampi and/or striata were
homogenized in ice-cold 0.32 M sucrose and centrifuged at 1000 g
(10 min, 4 °C). The supernatants were removed and centrifuged at
12 000 g (30 min, 4 °C): the resulting P2 pellets were then
resuspended in ice-cold 0.32 M sucrose and used for reuptake studies.
Sample aliquots of 50 mL were preincubated (5 min, 37 °C) in the
608 F. Fernandez et al.
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 607±618

presence of 350 mL of oxygenated (5% CO2/95% O2 for 30 min)
Krebs buffer containing (in mM) NaCl, 120; NaHCO3, 25; glucose,
10; KCl, 5; MgCl2, 1.2; EDTA, 0.05; CaCl2, 1.3; NaH2PO4, 1;
ascorbic acid, 0.1; and pargyline, 0.06 (glucose, ascorbic acid and
pargyline were made fresh every day). After preincubation, 50 mL of
10 nM [3H]5-HT were added, and the tubes left for further incubation
under weak agitation (10 min, 37 °C). For inhibition studies, 10 nM
[3H]5-HT were added to the buffer that contained (or not: baseline
level) 50 mL of MDMA (10-9±10-3M). All uptake assays, performed
in duplicate, were stopped by the addition of cold buffer through
Whatman GF/C glass ®bre ®lters (Poly Labo, Strasbourg, France).
The tubes and the ®lters were then washed with the buffer, and
entrapped radioactivity counted by liquid scintillation. Nonspeci®c
uptake was determined with 10 mM ¯uoxetine. Protein concentrations
were assessed using bovine serum albumin as standard (Bradford,
1976).
[3H]Paroxetine binding assays were performed as described
previously (Pollier et al., 2000), except that [3H]paroxetine was
used instead of [3H]citalopram. Following dissection, hippocampi
and striata were put on dry ice before storage at ±80 °C. Tissues were
then homogenized in 40 volumes ice-cold Tris-HCl buffer (pH 7.4),
and centrifuged (20 000 g for 10 min at 4 °C). The resulting pellets
were washed twice by resuspension in 40 volumes Tris-HCl, the last
washing lasting 1 h (35 °C) to remove endogenous 5-HT. The pellets
resulting from the third centrifugation were then stored at ±80 °C
until binding experiments. At that time, the pellets were suspended in
a 50-mM Tris±HCl buffer (pH 7.4) containing 5 mM KCl and 120 mM
NaCl, and transferred to glass tubes. The reaction was carried out for
90 min at 25 °C in the presence of 1 nM [3H]paroxetine with/without
10 mM ¯uoxetine for estimation of nonspeci®c binding. All assays
were stopped by the addition of cold buffer followed by a rapid
®ltration through Whatman GF/B glass ®bre ®lters. The ®lters were
washed twice with the buffer, and entrapped radioactivity counted by
liquid scintillation. Protein concentrations were assessed using bovine
serum albumin as standard (Bradford, 1976).
For 5-HT and 5-HIAA determinations, tissues were assayed as
previously described (Pollier et al., 2000). Except for the column and
the electrochemical detector (see below), all other materials (auto-
matic injector, cooled autosampler, pump, peak integrator, software)
were from Hewlett-Packard (series 1100; Hewlett-Packard France,
Evry, France). Tissues were stored at ±80 °C before being weighed,
sonicated in 0.4 N perchloric acid, and centrifuged (15 000 g for
10 min at 4 °C). Ten microlitres were injected in a C18 reversed-
phase Spherisorb column (150 3 4.6 mm, 5-mm spheres; Touzart et
Matignon, Courtaboeuf, France) with a mobile phase consisting of
0.05 M sodium acetate, 0.045 M citrate buffer (pH 3.8) containing
0.001% octane sulphonic acid and 15% methanol. Hippocampal and
striatal 5-HT and 5-HIAA concentrations were assessed using a
Decade electrochemical detector with capillary cell design (Antec,
Leiden, The Netherlands) set at 0.67 V (20 nA) against a Ag/AgCl
reference electrode.
Body temperature recordings
Rats were gently restrained and temperature measured by insertion of
a thermocouple, connected to a digital readout (Thermalert TH-5,
Physitemp; Phymep, Paris, France), into the rectum. Body tempera-
tures were measured three times, each separated by a 30-min interval,
before the s.c. administration of saline or MDMA. For technical
reasons, body temperature recordings in the ®rst series of experiments
(dose±responses to MDMA) lasted only for the 2 h (four recordings)
that followed MDMA injection. Note, however, that MDMA effects
on temperature are already maximal at 2 h (Dafters, 1994; Malberg
et al., 1996; Colado et al., 1997; Malberg & Seiden, 1998), including
in F344 rats (see Results). In the other series of experiments, body
temperatures were recorded up to 3 h after MDMA injection, except
for some chlorisondamine-pretreated rats which were recorded up to
7 h after MDMA injection (see Results). All these procedures were
repeated 24 h later. In all series of experiments, the ambient
temperature ranged between 22 and 23 °C and stable body tempera-
ture was reached within the 30 s that followed the probe insertion.
Statistics
All data are given as means 6 SEM. When necessary, data were log-
transformed to achieve homogeneity of variances before comparison
through one-way and multivariate analyses of variance (ANOVA)
followed, if signi®cant, by Tukey's multiple comparison test. One-
way ANOVAs included one between-factor (Dose of MDMA) whereas
multivariate ANOVAs included two between-factors, namely
Pretreatment (sham or adrenalectomy; sham, adrenalectomy or
adrenalectomy plus corticosterone; saline or chlorisondamine) and
Treatment (saline or MDMA) to which was added one within-factor
(i.e. Time) for body temperature data analyses.
Results
Immediate effects of MDMA on body temperature in F344 rats
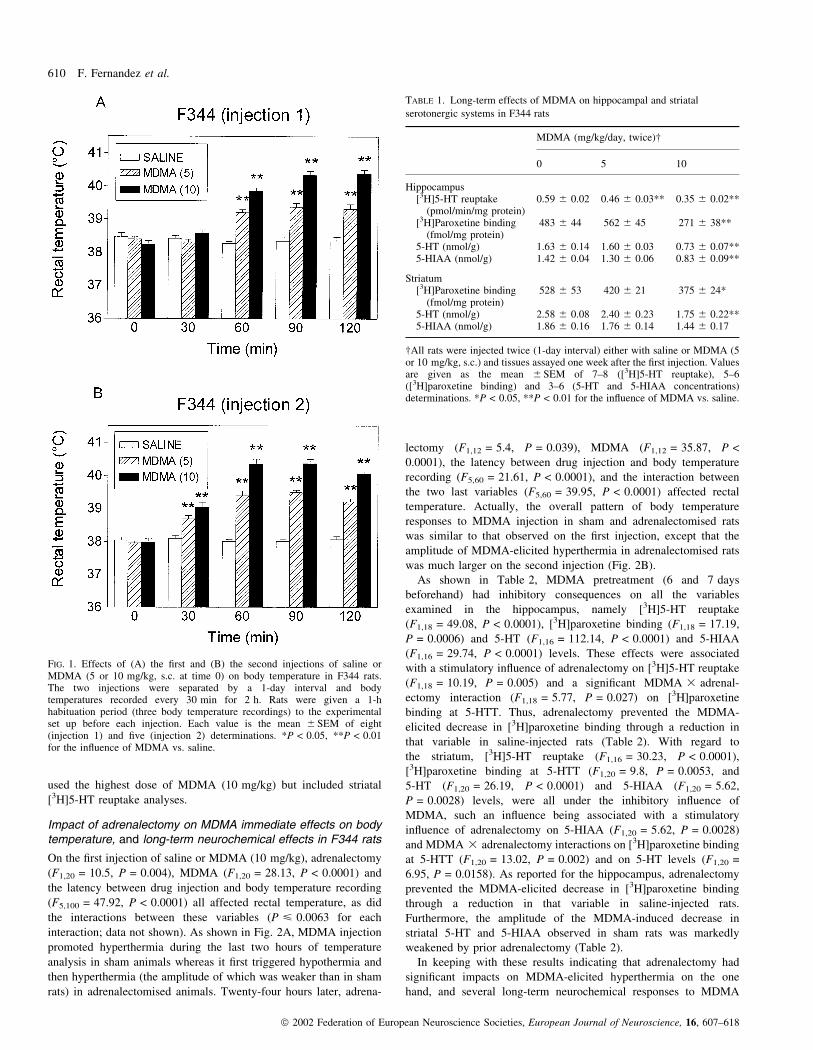
On the ®rst injection, the pretreatment (F2,21 = 45.21, P < 0.0001),
the latency between drug injection and body temperature recording
(F4,84 = 131.01, P < 0.0001) and the interaction between these
variables (F8,84 = 55.84, P < 0.0001), all affected rectal temperature.
MDMA administration was found to elicit dose-dependent, albeit
delayed, increases in body temperature in F344 rats (Fig. 1A). As
observed on the ®rst injection, the pretreatment (F2,12 = 124.13,
P < 0.0001) and the latency between drug injection and body
temperature recording (F4,48 = 119.6, P < 0.0001) affected rectal
temperature on the second day, as did the interaction between these
variables (F8,48 = 37.75, P < 0.0001). Indeed, it was found that the
second injection of MDMA, as opposed to the ®rst injection, elicited
immediate dose-dependent increases in body temperature in F344 rats
(Fig. 1B).
Long-term neurochemical effects of MDMA in F344 rats
Table 1 reports the effects of MDMA (last of the two injections
6 days beforehand) on hippocampal [3H]5-HT reuptake rates, and on
hippocampal and striatal [3H]paroxetine binding at 5-HTT and 5-HT
and 5-HIAA levels in F344 rats (striatal [3H]5-HT reuptake was not
assayed in this ®rst series of experiments). Hippocampal [3H]5-HT
reuptake and [3H]paroxetine binding were in¯uenced by the pretreat-
ment (F2,20 = 16.13, P < 0.0001 and F2,14 = 12.49, P = 0.0012,
respectively) with MDMA decreasing [3H]5-HT reuptake in a dose-
dependent manner whereas only the highest dose proved effective on
[3H]paroxetine binding (Table 1). Similarly, hippocampal 5-HT and
5-HIAA levels differred with regard to the pretreatment
(F2,7 = 22.58, P = 0.0009 and F2,6 = 25.09, P = 0.0012, respect-
ively), with the 10 mg/kg dose of MDMA decreasing these levels
(Table 1).
In striatum, [3H]paroxetine binding measurements and 5-HT
levels varied with the pretreatment (F2,9 = 5.24, P = 0.031, and
F2,15 = 4.22, P = 0.039, respectively), with the 10 but not the
5 mg/kg dose having signi®cant impacts (Table 1).
As these results indicated that F344 rats were sensitive to the long-
term neurochemical effects of MDMA, we next examined whether
the corticotropic axis mediated such a sensitivity. To do so, we only
Stress hormones, MDMA and serotonergic systems 609
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 607±618
used the highest dose of MDMA (10 mg/kg) but included striatal
[3H]5-HT reuptake analyses.
Impact of adrenalectomy on MDMA immediate effects on bodytemperature, and long-term neurochemical effects in F344 rats
On the ®rst injection of saline or MDMA (10 mg/kg), adrenalectomy
(F1,20 = 10.5, P = 0.004), MDMA (F1,20 = 28.13, P < 0.0001) and
the latency between drug injection and body temperature recording
(F5,100 = 47.92, P < 0.0001) all affected rectal temperature, as did
the interactions between these variables (P < 0.0063 for each
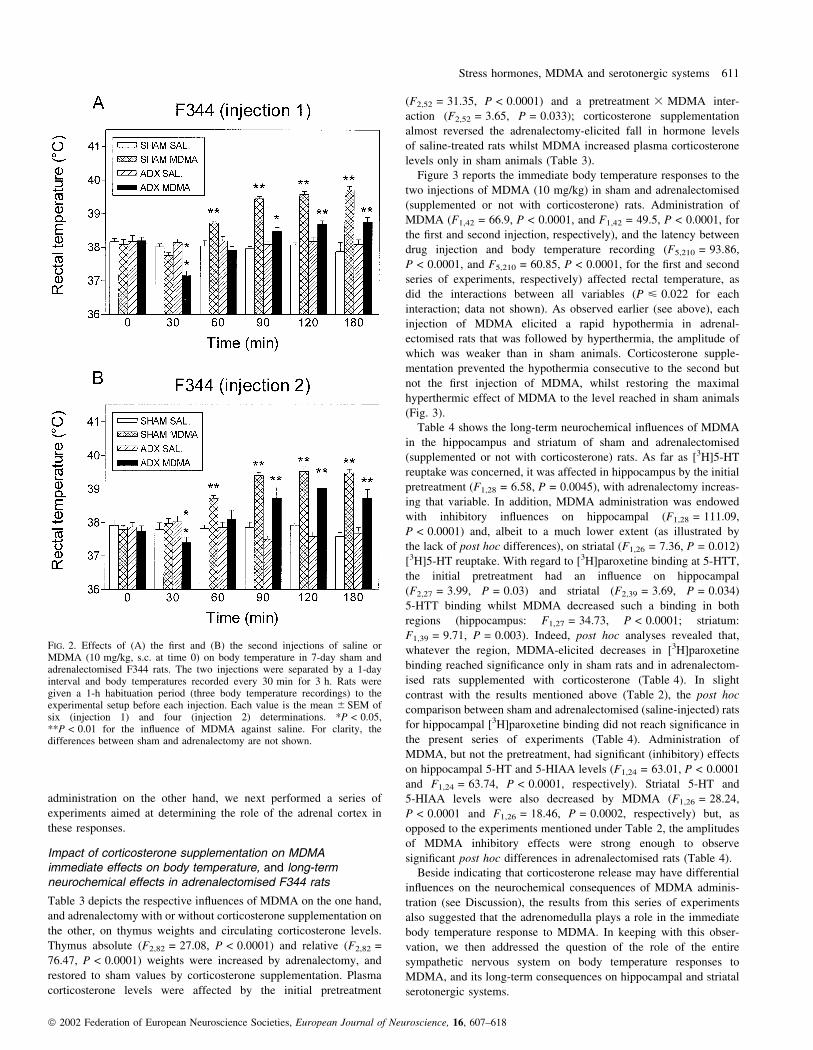
interaction; data not shown). As shown in Fig. 2A, MDMA injection
promoted hyperthermia during the last two hours of temperature
analysis in sham animals whereas it ®rst triggered hypothermia and
then hyperthermia (the amplitude of which was weaker than in sham
rats) in adrenalectomised animals. Twenty-four hours later, adrena-
lectomy (F1,12 = 5.4, P = 0.039), MDMA (F1,12 = 35.87, P <
0.0001), the latency between drug injection and body temperature
recording (F5,60 = 21.61, P < 0.0001), and the interaction between
the two last variables (F5,60 = 39.95, P < 0.0001) affected rectal
temperature. Actually, the overall pattern of body temperature
responses to MDMA injection in sham and adrenalectomised rats
was similar to that observed on the ®rst injection, except that the
amplitude of MDMA-elicited hyperthermia in adrenalectomised rats
was much larger on the second injection (Fig. 2B).
As shown in Table 2, MDMA pretreatment (6 and 7 days
beforehand) had inhibitory consequences on all the variables
examined in the hippocampus, namely [3H]5-HT reuptake
(F1,18 = 49.08, P < 0.0001), [3H]paroxetine binding (F1,18 = 17.19,
P = 0.0006) and 5-HT (F1,16 = 112.14, P < 0.0001) and 5-HIAA
(F1,16 = 29.74, P < 0.0001) levels. These effects were associated
with a stimulatory in¯uence of adrenalectomy on [3H]5-HT reuptake
(F1,18 = 10.19, P = 0.005) and a signi®cant MDMA 3 adrenal-
ectomy interaction (F1,18 = 5.77, P = 0.027) on [3H]paroxetine
binding at 5-HTT. Thus, adrenalectomy prevented the MDMA-
elicited decrease in [3H]paroxetine binding through a reduction in
that variable in saline-injected rats (Table 2). With regard to
the striatum, [3H]5-HT reuptake (F1,16 = 30.23, P < 0.0001),
[3H]paroxetine binding at 5-HTT (F1,20 = 9.8, P = 0.0053, and
5-HT (F1,20 = 26.19, P < 0.0001) and 5-HIAA (F1,20 = 5.62,
P = 0.0028) levels, were all under the inhibitory in¯uence of
MDMA, such an in¯uence being associated with a stimulatory
in¯uence of adrenalectomy on 5-HIAA (F1,20 = 5.62, P = 0.0028)
and MDMA 3 adrenalectomy interactions on [3H]paroxetine binding
at 5-HTT (F1,20 = 13.02, P = 0.002) and on 5-HT levels (F1,20 =
6.95, P = 0.0158). As reported for the hippocampus, adrenalectomy
prevented the MDMA-elicited decrease in [3H]paroxetine binding
through a reduction in that variable in saline-injected rats.
Furthermore, the amplitude of the MDMA-induced decrease in
striatal 5-HT and 5-HIAA observed in sham rats was markedly
weakened by prior adrenalectomy (Table 2).
In keeping with these results indicating that adrenalectomy had
signi®cant impacts on MDMA-elicited hyperthermia on the one
hand, and several long-term neurochemical responses to MDMA
TABLE 1. Long-term effects of MDMA on hippocampal and striatal
serotonergic systems in F344 rats
MDMA (mg/kg/day, twice)²
0 5 10
Hippocampus[3H]5-HT reuptake
(pmol/min/mg protein)0.59 6 0.02 0.46 6 0.03** 0.35 6 0.02**
[3H]Paroxetine binding(fmol/mg protein)
483 6 44 562 6 45 271 6 38**
5-HT (nmol/g) 1.63 6 0.14 1.60 6 0.03 0.73 6 0.07**5-HIAA (nmol/g) 1.42 6 0.04 1.30 6 0.06 0.83 6 0.09**
Striatum[3H]Paroxetine binding
(fmol/mg protein)528 6 53 420 6 21 375 6 24*
5-HT (nmol/g) 2.58 6 0.08 2.40 6 0.23 1.75 6 0.22**5-HIAA (nmol/g) 1.86 6 0.16 1.76 6 0.14 1.44 6 0.17
²All rats were injected twice (1-day interval) either with saline or MDMA (5or 10 mg/kg, s.c.) and tissues assayed one week after the ®rst injection. Valuesare given as the mean 6 SEM of 7±8 ([3H]5-HT reuptake), 5±6([3H]paroxetine binding) and 3±6 (5-HT and 5-HIAA concentrations)determinations. *P < 0.05, **P < 0.01 for the in¯uence of MDMA vs. saline.
FIG. 1. Effects of (A) the ®rst and (B) the second injections of saline orMDMA (5 or 10 mg/kg, s.c. at time 0) on body temperature in F344 rats.The two injections were separated by a 1-day interval and bodytemperatures recorded every 30 min for 2 h. Rats were given a 1-hhabituation period (three body temperature recordings) to the experimentalset up before each injection. Each value is the mean 6 SEM of eight(injection 1) and ®ve (injection 2) determinations. *P < 0.05, **P < 0.01for the in¯uence of MDMA vs. saline.
610 F. Fernandez et al.
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 607±618
administration on the other hand, we next performed a series of
experiments aimed at determining the role of the adrenal cortex in
these responses.
Impact of corticosterone supplementation on MDMAimmediate effects on body temperature, and long-termneurochemical effects in adrenalectomised F344 rats
Table 3 depicts the respective in¯uences of MDMA on the one hand,
and adrenalectomy with or without corticosterone supplementation on
the other, on thymus weights and circulating corticosterone levels.
Thymus absolute (F2,82 = 27.08, P < 0.0001) and relative (F2,82 =
76.47, P < 0.0001) weights were increased by adrenalectomy, and
restored to sham values by corticosterone supplementation. Plasma
corticosterone levels were affected by the initial pretreatment
(F2,52 = 31.35, P < 0.0001) and a pretreatment 3 MDMA inter-
action (F2,52 = 3.65, P = 0.033); corticosterone supplementation
almost reversed the adrenalectomy-elicited fall in hormone levels
of saline-treated rats whilst MDMA increased plasma corticosterone
levels only in sham animals (Table 3).
Figure 3 reports the immediate body temperature responses to the
two injections of MDMA (10 mg/kg) in sham and adrenalectomised
(supplemented or not with corticosterone) rats. Administration of
MDMA (F1,42 = 66.9, P < 0.0001, and F1,42 = 49.5, P < 0.0001, for
the ®rst and second injection, respectively), and the latency between
drug injection and body temperature recording (F5,210 = 93.86,
P < 0.0001, and F5,210 = 60.85, P < 0.0001, for the ®rst and second
series of experiments, respectively) affected rectal temperature, as
did the interactions between all variables (P < 0.022 for each
interaction; data not shown). As observed earlier (see above), each
injection of MDMA elicited a rapid hypothermia in adrenal-
ectomised rats that was followed by hyperthermia, the amplitude of
which was weaker than in sham animals. Corticosterone supple-
mentation prevented the hypothermia consecutive to the second but
not the ®rst injection of MDMA, whilst restoring the maximal
hyperthermic effect of MDMA to the level reached in sham animals
(Fig. 3).
Table 4 shows the long-term neurochemical in¯uences of MDMA
in the hippocampus and striatum of sham and adrenalectomised
(supplemented or not with corticosterone) rats. As far as [3H]5-HT
reuptake was concerned, it was affected in hippocampus by the initial
pretreatment (F1,28 = 6.58, P = 0.0045), with adrenalectomy increas-
ing that variable. In addition, MDMA administration was endowed
with inhibitory in¯uences on hippocampal (F1,28 = 111.09,
P < 0.0001) and, albeit to a much lower extent (as illustrated by
the lack of post hoc differences), on striatal (F1,26 = 7.36, P = 0.012)
[3H]5-HT reuptake. With regard to [3H]paroxetine binding at 5-HTT,
the initial pretreatment had an in¯uence on hippocampal
(F2,27 = 3.99, P = 0.03) and striatal (F2,39 = 3.69, P = 0.034)
5-HTT binding whilst MDMA decreased such a binding in both
regions (hippocampus: F1,27 = 34.73, P < 0.0001; striatum:
F1,39 = 9.71, P = 0.003). Indeed, post hoc analyses revealed that,
whatever the region, MDMA-elicited decreases in [3H]paroxetine
binding reached signi®cance only in sham rats and in adrenalectom-
ised rats supplemented with corticosterone (Table 4). In slight
contrast with the results mentioned above (Table 2), the post hoc
comparison between sham and adrenalectomised (saline-injected) rats
for hippocampal [3H]paroxetine binding did not reach signi®cance in
the present series of experiments (Table 4). Administration of
MDMA, but not the pretreatment, had signi®cant (inhibitory) effects
on hippocampal 5-HT and 5-HIAA levels (F1,24 = 63.01, P < 0.0001
and F1,24 = 63.74, P < 0.0001, respectively). Striatal 5-HT and
5-HIAA levels were also decreased by MDMA (F1,26 = 28.24,
P < 0.0001 and F1,26 = 18.46, P = 0.0002, respectively) but, as
opposed to the experiments mentioned under Table 2, the amplitudes
of MDMA inhibitory effects were strong enough to observe
signi®cant post hoc differences in adrenalectomised rats (Table 4).
Beside indicating that corticosterone release may have differential
in¯uences on the neurochemical consequences of MDMA adminis-
tration (see Discussion), the results from this series of experiments
also suggested that the adrenomedulla plays a role in the immediate
body temperature response to MDMA. In keeping with this obser-
vation, we then addressed the question of the role of the entire
sympathetic nervous system on body temperature responses to
MDMA, and its long-term consequences on hippocampal and striatal
serotonergic systems.
FIG. 2. Effects of (A) the ®rst and (B) the second injections of saline orMDMA (10 mg/kg, s.c. at time 0) on body temperature in 7-day sham andadrenalectomised F344 rats. The two injections were separated by a 1-dayinterval and body temperatures recorded every 30 min for 3 h. Rats weregiven a 1-h habituation period (three body temperature recordings) to theexperimental setup before each injection. Each value is the mean 6 SEM ofsix (injection 1) and four (injection 2) determinations. *P < 0.05,**P < 0.01 for the in¯uence of MDMA against saline. For clarity, thedifferences between sham and adrenalectomy are not shown.
Stress hormones, MDMA and serotonergic systems 611
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 607±618

Impact of ganglionic blockade on MDMA immediate effects onbody temperature, and long-term neurochemical effects inF344 rats
When examined up to 3 h after saline or MDMA injections (Fig. 4),
body temperature was found to be altered by chlorisondamine
pretreatment (F1,20 = 233.6, P < 0.0001 and F1,20 = 54.5,
P < 0.0001, for the ®rst and second series of injections), by
MDMA (F1,20 = 128.8, P < 0.0001 and F1,20 = 20.2, P = 0.0002,
for the ®rst and second series of injections), and by the latency
between MDMA injection and body temperature recording
(F6,120 = 23.68, P < 0.0001, and F6,120 = 11.57, P < 0.0001, for the
®rst and second series of injections). Furthermore, the interactions
between variables were all found to exert in¯uences on body
temperature (P < 0.0008 for each interaction; data not shown). Thus,
chlorisondamine and MDMA displayed, respectively, hypothermic
and hyperthermic properties throughout the two series of experiments
(Fig. 4). In addition, compared to saline administration, MDMA
administration to chlorisondamine-pretreated rats triggered a time-
dependent pattern of temperature responses, i.e. an ampli®cation of
chlorisondamine-elicited hypothermia followed by a progressive rise
in body temperature, with the amplitude of that rise decreasing on the
second MDMA injection (compared to the ®rst MDMA injection).
Because MDMA-elicited hyperthermia in chlorisondamine-pretreated
rats did not reach a plateau level at the end of the 3 h of analysis,
another experiment was set up where body temperature was
registered up to 7 h after MDMA injection (n = 4 rats/group). The
hyperthermia elicited by the ®rst injection of MDMA in chlorison-
damine-pretreated rats reached its maximum 4 h after MDMA
administration, this level remaining constant for an additional 2 h
and decreasing thereafter (MDMA-elicited hyperthermia was equiva-
lent in saline- and chlorisondamine-pretreated rats during the last 3 h
of analysis; data not shown). With regard to the second injection of
MDMA, its hyperthermic consequences in chlorisondamine-pre-
treated rats reached their maximum 3 h after MDMA administration
and vanished thereafter; indeed, neither chlorisondamine nor MDMA
had any in¯uence on body temperature for the last 3 h of analysis
(data not shown).
Table 5 shows the long-term neurochemical in¯uences of MDMA
in the hippocampus and striatum of saline- and chlorisondamine-
pretreated rats. In the hippocampus, [3H]5-HT reuptake was
in¯uenced by chlorisondamine (F1,19 = 5.72, P = 0.027) and
MDMA (F1,19 = 18.35, P = 0.0004) administration, post hoc analy-
ses showing that MDMA was endowed with a signi®cant effect in
saline-treated animals only. [3H]Paroxetine binding data con®rmed
[3H]5-HT reuptake data, as chlorisondamine (F1,27 = 13.68,
P = 0.001) and MDMA (F1,27 = 8.76, P = 0.0063) exerted strong
in¯uences whilst post hoc tests showed that the inhibitory impact of
MDMA lacked signi®cance in chlorisondamine-pretreated rats.
Hippocampal 5-HT levels were affected by chlorisondamine
(F1,20 = 4.71, P = 0.042), MDMA (F1,20 = 45.72, P < 0.0001), and
by the interaction between each treatment (F1,20 = 7.56, P = 0.012).
In addition, 5-HIAA levels were sensitive to MDMA administration
(F1,20 = 12.03, P = 0.0024) and to a chlorisondamine 3 MDMA
interaction (F1,20 = 6.04, P = 0.023). As far as striatum was
TABLE 2. Long-term effects of MDMA on hippocampal and striatal serotonergic systems in sham and adrenalectomised F344 rats
Sham Adrenalectomy
Saline MDMA (10 mg/kg) Saline MDMA (10 mg/kg)
Hippocampus[3H]5-HT reuptake (pmol/min/mg protein) 0.54 6 0.03 0.27 6 0.02** 0.60 6 0.03 0.39 6 0.04**,+
[3H]Paroxetine binding (fmol/mg protein) 354 6 10 210 6 26** 275 6 25+ 237 6 215-HT (nmol/g) 2.18 6 0.07 1.38 6 0.05** 2.12 6 0.05 1.35 6 0.10**5-HIAA (nmol/g) 1.00 6 0.05 0.75 6 0.02** 1.03 6 0.04 0.77 6 0.06**
Striatum[3H]5-HT reuptake (pmol/min/mg protein) 0.64 6 0.01 0.50 6 0.03** 0.58 6 0.03 0.50 6 0.01*[3H]Paroxetine binding (fmol/mg protein) 426 6 19 317 6 17** 355 6 16++ 363 6 105-HT (nmol/g) 2.47 6 0.11 1.73 6 0.11** 2.33 6 0.10 2.09 6 0.06+
5-HIAA (nmol/g) 1.41 6 0.07 1.07 6 0.04** 1.46 6 0.10 1.41 6 0.10++
All rats were injected twice (1-day interval) either with saline or MDMA (10 mg/kg, s.c.) and tissues assayed one week after the ®rst injection. Sham surgery oradrenalectomy were performed one week before the ®rst saline or MDMA injection. Values are given as the mean 6 SEM of 5±6 ([3H]5-HT reuptake,[3H]paroxetine binding) or 4±6 (5-HT and 5-HIAA concentrations) determinations. *P < 0.05, **P < 0.01 for the in¯uence of MDMA vs. saline. +P < 0.05,++P < 0.01 for the in¯uence of adrenalectomy.
TABLE 3. Long-term effects of MDMA on thymus weights and plasma corticosterone levels in sham, adrenalectomised and corticosterone-treated
adrenalectomised F344 rats
Sham Adrenalectomy Adrenalectomy + Corticosterone
Saline MDMA (10 mg/kg) Saline MDMA (10 mg/kg) Saline MDMA (10 mg/kg)
Thymus (mg) 420 6 13 415 6 10 524 6 13++ 511 6 13++ 433 6 23 400 6 18Thymus (mg/100 g b.w.) 201 6 6 197 6 5 298 6 10++ 309 6 10++ 221 6 12 214 6 8Plasma corticosterone (ng/mL) 45 6 10 73 6 13** 9 6 2++ 5 6 1++ 28 6 4 27 6 3++
All rats were injected twice (1-day interval) either with saline or MDMA (10 mg/kg, s.c.) and the animals killed for thymus weight and plasma corticosteroneanalyses. Sham surgery, adrenalectomy and the s.c. implantation of Cort. (50 mg) pellets were all performed 1 week before the ®rst saline or MDMA injection.Values are given as the mean 6 SEM of 14±16 (thymus weights) and 9±10 (plasma corticosterone) determinations. **P < 0.01 for the in¯uence of MDMA vs.saline; ++P < 0.01 vs. sham animals.
612 F. Fernandez et al.
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 607±618

concerned, neither [3H]5-HT reuptake nor [3H]paroxetine binding
proved sensitive to chlorisondamine and/or MDMA in these series of
experiments. However, 5-HT and 5-HIAA were in¯uenced by
MDMA treatment (5-HT: F1,20 = 22.25, P < 0.0001; 5-HIAA:
F1,26 = 17.62, P = 0.0004), and were sensitive to a chlorisond-
amine 3 MDMA interaction (5-HT: F1,20 = 4.45, P = 0.0475; 5-
HIAA: F1,20 = 4.36, P = 0.0499). Indeed, post hoc tests showed that
MDMA-elicited reductions in hippocampal and striatal 5-HT and
5-HIAA levels were either markedly reduced or prevented by
chlorisondamine pretreatment (Table 5).
Discussion
The goal of this series of experiments was to explore the roles of the
corticotropic axis and sympathetic activity in MDMA-elicited
neurotoxicity in the hippocampus and the striatum, the extent of
this neurotoxic effect being assessed by the simultaneous measure-
ment of [3H]5-HT reuptake, [3H]paroxetine binding at 5-HTT, and
5-HT and 5-HIAA levels (Molliver et al., 1990; Green et al., 1995;
Sprague et al., 1998). Furthermore, the possibility that corticotropic
activity also affects some immediate responses to MDMA, e.g.
hyperthermia, thought to be predictive of its neurotoxic impact (see
below), led us to monitor body temperature. However, because we
chose to address our questions in F344 rats, which bear a hyperactive
and hyperesponsive corticotropic axis (Sternberg et al., 1989;
Calogero et al., 1992; Sternberg et al., 1992; Dhabbar et al., 1993),
we ®rst checked by means of two doses of MDMA that this strain
displays hyperthermia and signi®cant long-term reductions in the
aforementioned neurochemical variables.
Corticosterone, sympathetic activity and MDMA-elicitedhyperthermia in F344 rats
Acute administration of MDMA has a rapid effect on the
thermoregulatory system, the amplitude of which depends on ambient
temperature and is independent of MDMA-induced changes in
locomotion (Dafters, 1994). Indeed, MDMA elicits hypothermia and
hyperthermia when ambient temperatures are, respectively, below
and above 20±22 °C (Gordon et al., 1991; Dafters, 1994; Malberg &
Seiden, 1998), the value of that `break point' depending on the rat
strain (Malpass et al., 1999) and the rat environment (familiar vs.
unfamiliar: Miller & O'Callaghan, 1994). In our hands, a dose-
dependent hyperthermia was observed on both injections in F344 rats.
However, such a hyperthermic effect of MDMA was preceded by a
hypothermic phasis if the rats were either adrenalectomised or
pretreated with the ganglionic blocker chlorisondamine. Furthermore,
corticosterone implants, at doses that reversed adrenalectomy-elicited
thymus hypertrophy (see below), proved ineffective on MDMA-
induced hypothermia in adrenalectomised F344 rats, thus indicating
that the lack of adrenomedulla, rather than the lack of adrenal cortex,
was responsible for the immediate hypothermia in adrenalectomised
rats. This result, which indicates that MDMA may rapidly stimulate
the adrenomedulla in intact animals, is in keeping with past evidence
for adrenaline-releasing effects of 5-HT indirect (5-HT reuptake
inhibitors, 5-HT releasers) and direct (5-HT1A and 5-HT2A receptor)
agonists (Chaouloff, 1993). Lastly, with regard to the long-lasting
hyperthermic effects of MDMA in F344 rats, our experiments suggest
that the sympathetic system (as shown by chlorisondamine pretreat-
ment) and, to a lesser extent, corticoids (as shown by the respective
effects of adrenalectomy and corticosterone supplementation) medi-
ate such a hyperthermia, the effects of corticoids being possibly
indirect through their control of sympathetic activity (Axelrod &
Reisine, 1984).
Neurochemical effects of MDMA in F344 rats
Administration of MDMA triggered long-term dose-dependent
decreases in hippocampal [3H]5-HT reuptake. Although MDMA
bears intrinsic inhibitory properties on [3H]5-HT reuptake (Steele
et al., 1987; Rudnick & Wall, 1992; Crespi et al., 1997), including in
F344 rats (IC50 = 0.93 6 0.08 mM, n = 4), it is unlikely that the
long-term decrease in hippocampal [3H]5-HT reuptake in MDMA-
treated rats is due to such an intrinsic effect. Thus, other variables
examined in the hippocampus, such as [3H]paroxetine binding at
5-HTT and 5-HT and/or 5-HIAA levels, also proved sensitive in the
long term to MDMA administration. However, this was true for the
FIG. 3. Effects of (A) the ®rst and (B) the second injections of saline orMDMA (10 mg/kg, s.c. at time 0) on body temperature in F344 rats bearinga sham surgery, adrenalectomy, or adrenalectomy supplemented withcorticosterone (CORT.) pellets. All pretreatments were performed 7 daysbefore the ®rst injection of saline or MDMA. The two injections of salineor MDMA were separated by a 1-day interval and body temperaturesrecorded every 30 min for 3 h. Rats were given a 1-h habituation period (3body temperature recordings) to the experimental set up before eachinjection. Each value is the mean 6 SEM of 8 determinations. *P < 0.05and **P < 0.01 for the in¯uence of MDMA against saline. For clarity, thedifferences between pretreatments are not shown.
Stress hormones, MDMA and serotonergic systems 613
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 607±618

highest dose of MDMA used whereas the 5 mg/kg dose decreased
neither 5-HTT densities nor 5-HT and 5-HIAA levels although it
diminished [3H]5-HT reuptake. Such a dissociation between MDMA
respective effects on 5-HTT density and function has also been
recently observed in animals treated with another substituted
amphetamine (namely methamphetamine: Haughey et al., 2000).
The analysis of 5-HTT densities and indole concentrations in the
striatum revealed a pattern of decreases in MDMA-treated animals
that were almost consistent with the aforementioned data in the
hippocampus. However, these decreases (taken as percentages of the
values measured in saline-treated rats) were weaker in striatum than
in hippocampus. Indeed, the high sensitivity of hippocampal, as
compared to striatal, serotonergic neurons to the long-term effects of
MDMA was observed throughout the whole study, including in the
last series of experiments (chlorisondamine pretreatments; see
below).
Role of the corticotropic axis in MDMA neurochemical effectsin F344 rats
In keeping with the results depicted above, we next examined
whether the corticotropic axis partly or totally mediated the long-term
neurochemical in¯uences of MDMA in F344 rats. One support for
such a hypothesis would have been the preliminary demonstration
that the corticotropic axis of F344 rats responds positively to MDMA
administration. Indeed, such an experiment was not set up for the
following two reasons: (i) a correct assessment of this responsiveness
would have required dose- and time-dependent responses of plasma
corticosterone to MDMA administration, an experiment rendered
impossible by the limited amount of MDMA available, and (ii) a
previous study has shown that the acute stimulation of 5-HT2A,C
receptors, i.e. the receptors that mediate MDMA-elicited cortico-
sterone release (Nash et al., 1988), is able to increase plasma
corticosterone in F344 rats (Calogero et al., 1992). Instead, we
thought that a correct estimation of the role of the corticotropic axis
in the long-term effects of MDMA should rely on the demonstration
that adrenalectomy attenuates or prevents such long-term effects in
F344 rats. In the hippocampus, MDMA-induced reductions in [3H]5-
HT reuptake and, in tissue, 5-HT and 5-HIAA levels, proved
insensitive to adrenalectomy. In striatum, an almost similar pattern of
responses could be observed although the weak in¯uence of MDMA
in that region and some heterogeneity in the data between the two
series of experiments cannot be dismissed. As concerns hippocampal
and striatal [3H]paroxetine binding, the amplitudes of MDMA-
elicited reductions in that variable were markedly weakened, if not
removed, by prior adrenalectomy. Indeed, the overall analysis of
[3H]paroxetine binding in the two series of experiments strongly
suggest that such an impact of adrenalectomy was due to intrinsic
inhibitory effects of adrenalectomy on baseline (i.e. saline-treated
rats) [3H]paroxetine binding at 5-HTT. Thus, adrenalectomy reduced,
respectively, hippocampal and striatal baseline [3H]paroxetine bind-
ing by 22% and 17% in the ®rst series of experiments, and by 14%
(trend only) and 17% in the second series of experiments although,
for an unknown reason, baseline [3H]paroxetine binding differred
between the two series of experiments. Furthermore, it was found that
corticosterone supplementation reinstated the inhibitory impact of
MDMA on hippocampal and striatal [3H]paroxetine binding, but not
the adrenalectomy-induced reduction in baseline [3H]paroxetine
binding. One explanation could be that the adrenomedulla, rather
than the adrenal cortex, controls baseline hippocampal and striatal
[3H]paroxetine binding, but the results from the chlorisondamine
experiments partly rule out such an hypothesis (see below).
Alternatively, MDMA-elicited reductions in [3H]paroxetine binding
and baseline [3H]paroxetine binding might be sensitive to low and
high corticosterone concentrations, respectively, with these high
concentrations being above those provided by our supplementation
regimen. At ®rst glance, this could be supported by the observation
that our corticosterone regimen provided circulating corticosterone
levels that did not reach those measured in sham animals. However, it
is likely that the higher circulating corticosterone levels in sham rats
are fully accounted for by a rapid release of corticosterone as they
were killed. Furthermore, in keeping with the high and low af®nities
of, respectively, mineralocorticoid receptors and glucocorticoid
receptors for corticosterone (Reul & De Kloet, 1985), and past
evidence for the exclusive presence of glucocorticoid receptors in the
TABLE 4. Long-term effects of MDMA on hippocampal and striatal serotonergic systems in sham, adrenalectomised and corticosterone-treated
adrenalectomised F344 rats
Sham Adrenalectomy Adrenalectomy + Corticosterone
Saline MDMA (10 mg/kg) Saline MDMA (10 mg/kg) Saline MDMA (10 mg/kg)
Hippocampus[3H]5-HT reuptake 0.53 6 0.02 0.31 6 0.02** 0.61 6 0.02+ 0.38 6 0.02** 0.62 6 0.04+ 0.38 6 0.02**(pmol/min/mg protein)[3H]Paroxetine binding 520 6 26 378 6 31** 449 6 19 398 6 24 465 6 27 291 6 21**,+
(fmol/mg protein)5-HT (nmol/g) 1.93 6 0.15 1.08 6 0.16** 2.08 6 0.21 1.24 6 0.07** 2.07 6 0.09 1.04 6 0.10**5-HIAA (nmol/g) 1.16 6 0.04 0.75 6 0.07** 1.31 6 0.08 0.91 6 0.04** 1.17 6 0.05 0.82 6 0.06**
Striatum[3H]5-HT reuptake 0.82 6 0.04 0.74 6 0.05 0.80 6 0.03 0.73 6 0.06 0.96 6 0.07 0.71 6 0.09**(pmol/min/mg protein)[3H]Paroxetine binding 715 6 31 550 6 34** 593 6 60+ 571 6 26 579 6 34+ 483 6 23*(fmol/mg protein)5-HT (nmol/g) 2.44 6 0.08 1.87 6 0.09** 2.55 6 0.19 1.94 6 0.15** 2.30 6 0.07 1.90 6 0.07*5-HIAA (nmol/g) 1.40 6 0.06 1.11 6 0.09* 1.59 6 0.08 1.19 6 0.08** 1.33 6 0.11 1.16 6 0.06
All rats were injected twice (1-day interval) either with saline or MDMA (10 mg/kg, s.c.) and tissues assayed one week after the ®rst injection. Sham surgery,adrenalectomy and the s.c. implantation of corticosterone (Cort., 50 mg) pellets were all performed one week before the ®rst saline or MDMA injection. Values aregiven as the mean 6 SEM of 5±6 ([3H]5-HT reuptake), 5±8 ([3H]paroxetine binding) and 5±6 (5-HT and 5-HIAA concentrations) determinations. *P < 0.05,**P < 0.01 for the in¯uence of MDMA vs. saline. +P < 0.05 vs. sham rats.
614 F. Fernandez et al.
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 607±618

thymus (Miller et al., 1990), the fact that our corticosterone
supplementation regimen fully prevented the adrenalectomy-elicited
increase in thymus weights (thus con®rming previous data: Cador
et al., 1993) may be taken as evidence for an effective reinstatement
of adequate corticosterone concentrations. Furthermore, plasma
corticosterone levels were increased by MDMA pretreatment in
sham rats, an observation either shared (Yau et al., 1994) or not
(McNamara et al., 1995; Aguirre et al., 1999) by others; actually, the
ineffectiveness of MDMA in altering thymus weights (and adrenal
weights; data not shown), and which con®rms previous data (Yau
et al., 1994), rather suggests increased fear at the time of killing and/
or increased activity of the corticotropic axis, such an increase being,
however, not sustained enough to alter target organs.
The observation that corticosteroid removal by adrenalectomy
prevented MDMA-elicited reductions in [3H]paroxetine binding, but
not in [3H]5-HT reuptake and 5-HT and 5-HIAA levels in the
hippocampus, raises the question of the net impact of adrenalectomy
on the MDMA neurotoxic effect. Although most, if not all, studies
consider the neurochemical variables used herein as valuable indices
of neurotoxicity (see Introduction), the lack of morphological and/or
immunocytochemical tools in this study impedes any clearcut answer
to that point. Glial ®brillary acidic protein overexpression may be the
simplest tool to show neurotoxicity (O'Callaghan et al., 1995),
including following MDMA injection (O'Callaghan et al., 1995;
Aguirre et al., 1997); unfortunately, adrenalectomy per se also
increases glial ®brillary acidic protein expression (O'Callaghan et al.,
1989), thus rendering inadequate that paradigm in the present study.
At the present stage, we can only underline the ®nding that the
MDMA-elicited reduction in [3H]5-HT reuptake, i.e. a measure of
membrane-bound 5-HTT, was insensitive to adrenalectomy, as were
5-HT and 5-HIAA levels, thereby suggesting that adrenalectomy
affected only cytosolic 5-HTT (assuming that [3H]paroxetine
measures both cytosolic and membrane 5-HTTs) without any
interaction with the MDMA neurodegenerative effects. Such a
statement is supported by the chlorisondamine experiments where
MDMA-elicited reductions in hippocampal [3H]5-HT reuptake,
[3H]paroxetine binding and 5-HT and 5-HIAA levels all proved
sensitive to the pretreatment (see below).
Role of sympathetic activity in MDMA neurochemical effects inF344 rats
As indicated earlier, the body temperature responses to MDMA in
adrenalectomised F344 rats, complemented or not with cortico-
sterone, point to a permissive effect of the adrenomedulla in the early
(up to 60 min) hyperthermic effects, but not the long-term
neurochemical effects, of MDMA. Furthermore, the possibility that
the mechanisms that allow MDMA to promote hyperthermia at later
times (i.e. after 60 min) also play a role in its long-term neuro-
chemical effects remained, however, open. This was supported by
pharmacological and physiological studies (Miller & O'Callaghan,
1994; Farfel & Seiden, 1995; Malberg et al., 1996; Malberg &
Seiden, 1998; Colado et al., 1999) which have suggested a link
between the hyperthermic and the neurotoxic properties of MDMA.
However, some results either do not support a direct link between
hyperthermia and neurotoxicity or suggest that hyperthermia only
partially explains neurotoxicity (Malberg et al., 1996; Aguirre et al.,
1997; Sprague et al., 1998). Moreover, the challenges (e.g. cold
exposure, amino acid and monoamine receptor ligands, catechol-
amine synthesis inhibitors) that have allowed the establishment of
that link are also endowed with peripheral and central actions which
are beyond the sole control of body temperature. Indeed, there is
evidence for an immediate stimulatory in¯uence of MDMA upon
sympathetic activity (O'Cain et al., 2000; Pedersen & Blessing,
2001), which is in keeping with the acute sympathoexcitatory
properties of 5-HT reuptake inhibitors/releasers (Chaouloff et al.,
1992) and 5-HT2A receptor agonists (Chaouloff, 1993). Con®rmingly,
the vasoconstrictor properties of MDMA have been shown to
contribute to its hyperthermic effect (Pedersen & Blessing, 2001).
In the present study, we chose to selectively address the role of
sympathetic activity by measuring the impacts of the ganglionic
blocker chlorisondamine (Gosling & Lu, 1969; Clarke et al., 1994) on
the hyperthermic and long-term neurochemical effects of MDMA. In
the hippocampus, the amplitudes of MDMA-elicited decreases in
[3H]5-HT reuptake, [3H]paroxetine binding and 5-HT and 5-HIAA
levels were markedly reduced (by 56, 63, 56 and 81%, respectively)
FIG. 4. Effects of (A) the ®rst and (B) the second injections of saline orMDMA (10 mg/kg, s.c. at time 0) on body temperature in F344 ratspretreated 30 min beforehand with saline or chlorisondamine (`CHLOR.';2.5 mg/kg i.p). The two injections of saline or MDMA were separated by a1-day interval and body temperatures recorded every 30 min for 3 h. Ratswere given a 30-min habituation period (two body temperature recordings)to the experimental set up before saline or chlorisondamine pretreatments.Each value is the mean 6 SEM of six determinations. *P < 0.05,**P < 0.01 for the in¯uence of MDMA against saline. For clarity, thedifferences between pretreatments are not shown (see text for bodytemperature data up to 7 h after MDMA injection).
Stress hormones, MDMA and serotonergic systems 615
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 607±618

by chlorisondamine pretreatment. In striatum, for an unknown reason,
neither [3H]5-HT reuptake nor [3H]paroxetine binding were
decreased to signi®cant extents by MDMA in these series of
experiments; however, con®rming the hippocampal data mentioned
above, it was observed that MDMA-elicited reductions in 5-HT and
5-HIAA levels were, respectively, diminished by 60 and 67% in
chlorisondamine-pretreated rats. Surprisingly, we also observed that
chlorisondamine pretreatment was endowed with slight but signi®-
cant intrinsic stimulatory in¯uences on baseline hippocampal [3H]5-
HT reuptake and more evidently on [3H]paroxetine binding. The
reason for such an increase is unknown, as sympathetic activity has
never been reported to affect 5-HTT binding, especially in the long
term. Indeed, one possibility would be that although chlorisondamine
is a preferential peripheral nicotinic receptor antagonist (Gosling &
Lu, 1969; Clarke et al., 1994), it also blocked central (e.g. in
hippocampus) nicotinic receptors, thereby altering, in the long-term,
[3H]paroxetine binding. This is, however, unlikely because (i) central
nicotinic receptor blockade by chlorisondamine has been shown to
only occur with high doses (10 mg/kg s.c.: Clarke, 1984; Clarke et al.,
1994) (ii) such a high dosage proved, however, ineffective on long-
term nicotinic receptor binding (Clarke et al., 1994), and (iii) acute
hippocampal nicotinic receptor blockade does not alter, per se, 5-HT
transmission as indicated by in vitro release experiments (Kenny
et al., 2000; Reuben & Clarke, 2000). Although our results show that
the ganglionic blocker chlorisondamine markedly weakened the long-
term consequences of MDMA on hippocampal and striatal seroto-
nergic neurons, we believe that the mechanisms underlying its
intrinsic effects on serotonergic systems should be investigated.
Conclusions
By means of neurochemical tools thought to provide a good estimate
of the integrity of serotonergic nerve terminals, this study strongly
suggests that sympathetic blockade, but not corticosterone removal,
diminishes the neurotoxic impact of MDMA, especially in the
hippocampus. Future work should, however, assess the extent to
which this result is supported by morphological ®ndings and how it
extends to other rat strains, including standard strains. We believe
that future work should also analyse whether the protective effect of
chlorisondamine reported above can be further increased by a longer-
lasting ganglionic blockade (indicating that the neurotoxic effect of
MDMA is fully accounted for by its hyperthermic effect) or is already
maximal (indicating that body temperature-independent mechanisms
may further protect against neurotoxicity).
Acknowledgements
This work was supported by la Mission InterministeÂrielle de Lutte contre laDrogue et la Toxicomanie (grant 99D05/AH003D to F.C), INSERM, INRA,and le Conseil ReÂgional d'Aquitaine. The authors wish to thank the NationalInstitute on Drug Abuse (Bethesda, MD, USA) for the kind gift of MDMA,and Aline Foury for corticosterone assays.
Abbreviations
5-HIAA, 5-hydroxyindoleacetic acid; 5-HT, 5-hydroxytryptamine/serotonin;5-HTT, 5-hydroxytryptamine/serotonin transporter; F344, Fischer 344;MDMA, 3,4-methylenedioxymethamphetamine.
References
Aguirre, N., Barrionuevo, M., Ramirez, M.J., Del Rio, J. & Lasheras, B.(1997) a-Lipoic acid prevents 3,4-methylenedioxymethamphetamine(MDMA)-induced neurotoxicity. Neuroreport, 10, 3675±3680.
Aguirre, N., Frechilla, D., Garcia-Osta, A., Lasheras, B. & Del Rio, J. (1999)Differential regulation by methylenedioxymethamphetamine of 5-hydroxy-tryptamine1A receptor density and mRNA expression in rat hippocampus,frontal cortex, and brainstem: the role of corticosteroids. J. Neurochem., 68,1099±1105.
Aulakh, C.S., Wozniak, K.M., Hill, J.L., Devane, C.L., Tolliver, T.J. &Murphy, D.L. (1988) Differential neuroendocrine responses to the 5-HTagonist m-chlorophenylpiperazine in Fawn-Hooded rats relative to Wistarand Sprague-Dawley rats. Neuroendocrinology, 48, 401±406.
Axelrod, J. & Reisine, T.D. (1984) Stress hormones: their interaction andregulation. Science, 224, 452±459.
Battaglia, G.S., Yeh, S.Y. & De Souza, E.B. (1988) MDMA-inducedneurotoxicity: parameters of degeneration and recovery of brain serotoninneurons. Pharmacol. Biochem. Behav., 29, 269±274.
Battaglia, G.S., Yeh, S.Y., O'Hearn, E., Molliver, M.E., Kuhar, M.J. & DeSouza, E.B. (1987) 3,4-Methylenedioxymethamphetamine and 3,4-methylenedioxyamphetamine destroy serotonin terminals in rat brain:quanti®cation of neurodegeneration by measurement of [3H]paroxetine-labeled serotonin uptake sites. J. Neurochem., 242, 911±916.
Bradford, M.M. (1976) A rapid and sensitive method for the quantitation ofmicrogram quantities of protein-dye binding. Anal. Biochem., 72, 248±254.
TABLE 5. Long-term effects of MDMA on hippocampal and striatal serotonergic systems in saline and chlorisondamine-treated F344 rats
Saline-treated Chlorisondamine-treated
Saline MDMA (10 mg/kg) Saline MDMA (10 mg/kg)
Hippocampus[3H]5-HT reuptake (pmol/min/mg protein) 0.39 6 0.02 0.24 6 0.03** 0.41 6 0.03 0.34 6 0.02+
[3H]Paroxetine binding (fmol/mg protein) 377 6 18 244 6 28* 427 6 35 398 6 26++
5-HT (nmol/g) 1.78 6 0.09 1.01 6 0.08** 1.73 6 0.08 1.41 6 0.07*,++
5-HIAA (nmol/g) 0.95 6 0.03 0.65 6 0.06** 0.89 6 0.06 0.84 6 0.05+
Striatum[3H]5-HT reuptake (pmol/min/mg protein) 0.57 6 0.01 0.51 6 0.02 0.57 6 0.02 0.54 6 0.02[3H]Paroxetine binding (fmol/mg protein) 499 6 36 501 6 61 543 6 46 602 6 475-HT (nmol/g) 2.53 6 0.08 1.84 6 0.08** 2.41 6 0.14 2.15 6 0.09+
5-HIAA (nmol/g) 1.44 6 0.08 1.00 6 0.04** 1.28 6 0.09 1.13 6 0.05
All rats were injected twice (1-day interval) either with saline or MDMA (10 mg/kg, s.c) and tissues assayed one week after the ®rst injection. Saline orchlorisondamine (2.5 mg/kg i.p.) pretreatments were performed 30 min before each of the two saline or MDMA injections. Values are given as the mean 6 SEMof 5±7 ([3H]5-HT reuptake), 7±8 ([3H]paroxetine binding) and 6 (5-HT and 5-HIAA concentrations) determinations. *P < 0.05, **P < 0.01 for the in¯uence ofMDMA vs. saline. +P < 0.05, ++P < 0.01 for the in¯uence of chlorisondamine.
616 F. Fernandez et al.
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 607±618

Cador, M., Dulluc, J. & MormeÁde, P. (1993) Modulation of the locomotorresponse to amphetamine by corticosterone. Neuroscience, 56, 981±988.
Calogero, A.E., Sternberg, E.M., Bagdy, G., Smith, C., Bernardini, R.,Aksentijevich, S., Wilder, R.L., Gold, P.W. & Chrousos, G.P. (1992)Neurotransmitter-induced hypothalamic-pituitary-adrenal axis responsive-ness is defective in in¯ammatory disease-susceptible Lewis rats: in vivo andin vitro studies suggesting globally defective hypothalamic secretion ofcorticotropin-releasing hormone. Neuroendocrinology, 55, 600±608.
Chaouloff, F. (1993) Physiopharmacological interactions between stresshormones and central serotonergic systems. Brain Res. Rev., 9, 219±233.
Chaouloff, F., Baudrie, V. & Coupry, I. (1994) Effects of chlorisondamine andrestraint on cortical [3H]ketanserin binding, 5-HT2A receptor-mediated headshakes, and behaviours in models of anxiety. Neuropharmacology, 33, 449±456.
Chaouloff, F., Gunn, S.H. & Young, J.B. (1992) Central 5-hydroxytryptamine2
receptors are involved in the adrenal catecholamine-releasing andhyperglycemic effects of the 5-hydroxytryptamine indirect agonist d-fen¯uramine in the conscious rat. J. Pharmacol. Exp. Ther., 260, 1008±1016.
Clarke, P.B.S. (1984) Chronic central nicotinic blockade after a singleadministration of the bisquaternary ganglion-blocking drug chlorisond-amine. Br. J. Pharmacol., 83, 527±535.
Clarke, P.B.S., Chaudieu, I., El-Bizri, H., Boksa, P., Quik, M., Esplin, B.A. &Capek, R. (1994) The pharmacology of the nicotinic antagonist,chlorisondamine, investigated in rat brain and autonomic ganglion. Br. J.Pharmacol., 111, 397±405.
Colado, M.I., Esteban, B., O'Shea, E., Granados, R. & Green, A.R. (1999)Studies on the neuroprotective effect of pentobarbitone on MDMA-inducedneurodegeneration. Psychopharmacology, 142, 421±425.
Colado, M.I. & Green, A.R. (1995) The spin-trap reagent a-phenyl-N-tert-butyl nitrone prevents neurodegeneration following administration of`ecstasy'. Eur. J. Pharmacol., 280, 343±346.
Colado, M.I., O'Shea, E., Granados, R., Murray, T.K. & Green, A.R. (1997) Invivo evidence for free radical involvement in the degeneration of rat brain5-HT following administration of MDMA (`ecstasy') and p-chloroamphetamine but not the degeneration following fen¯uramine. Br.J. Pharmacol., 121, 889±900.
Commins, D.L., Vosmer, G., Virus, R.M., Woolverton, W.L., Schuster, C.R.& Seiden, L.S. (1987) Biochemical and histological evidence thatmethylenedioxymethamphetamine (MDMA) is toxic to neurons in ratbrain. J. Pharmacol. Exp. Ther., 241, 338±345.
Crespi, D., Mennini, T. & Gobbi, M. (1997) Carrier-dependent and Ca2+-dependent 5-HT and dopamine release induced by (+) amphetamine, 3,4-methylendioxymethamphetamine, p-chloroamphetamine and (+) fen¯ur-amine. Br. J. Pharmacol., 121, 1735±1743.
Dafters, R.I. (1994) Effect of ambient temperature on hyperthermia andhyperkinesis induced by 3,4-methylenedioxymethamphetamine (MDMA or! ecstasy @) in rats. Psychopharmacology, 114, 505±508.
De Souza, E.B., Battaglia, G. & Insel, T.R. (1990) Neurotoxic effects ofMDMA on brain serotonin neurons: evidence from neurochemical andradioligand binding studies. In Whitaker-Azmitia, P.M. & Peroutka, S.J.(eds), The Neuropharmacology of Serotonin. Ann. NY Acad. Sci., Vol. 600.The New York Academy of Sciences, New York, pp. 682±697.
Dhabbar, F.S., McEwen, B.S. & Spencer, R.L. (1993) Stress response, adrenalsteroid receptor levels, and corticosteroid-binding globulin levels ± Acomparison between Sprague-Dawley, Fischer 344, and Lewis rats. BrainRes., 616, 89±98.
Farfel, G.M. & Seiden, L.S. (1995) Role of hypothermia in the mechanism ofprotection against serotonergic toxicity. I. Experiments using 3,4-methylenedioxy-methamphetamine, dizocilpine, CGS 19755 and NBQX.J. Pharmacol. Exp. Ther., 272, 860±867.
Globus, M.Y., Busto, R., Lin, B., Schnippering, H. & Ginsberg, M.D. (1995)Detection of free radical activity during transient global ischemia andrecirculation: effects of intraischemic brain temperature modulation. J.Neurochem., 65, 1250±1256.
Gordon, C.J., Watkinson, W.P., O'Callaghan, J.P. & Miller, D.B. (1991)Effects of 3,4-methylenedioxymethamphetamine on autonomicthermoregulatory responses of the rat. Pharmacol. Biochem. Behav., 38,339±344.
Gosling, J.A. & Lu, T.C. (1969) Uptake and distribution of some quaternaryammonium compounds in the central nervous system of the rat. J.Pharmacol. Exp. Ther., 167, 56±62.
Green, A.R., Cross, A.J. & Goodwin, G.M. (1995) Review of thepharmacology and clinical pharmacology of 3,4-methylenedioxy-
methamphetamine (MDMA or `Ecstasy'). Psychopharmacology, 119,247±260.
Gudelsky, G.A. (1996) Effect of ascorbate and cysteine on the 3,4-methylenedioxymethamphetamine-induced depletion of brain serotonin.J. Neural Transm., 103, 1397±1404.
Haughey, H.M., Fleckenstein, A.E., Metzger, R.R. & Hanson, G.R. (2000) Theeffects of methamphetamine on serotonin transporter activity: role ofdopamine and hyperthermia. J. Neurochem., 75, 1608±1617.
Johnson, M., Stone, D.M., Bush, L.G., Hanson, G.R. & Gibb, J.W. (1989)Glucocorticoids and 3,4-methylenedioxymethamphetamine (MDMA)-induced neurotoxicity. Eur. J. Pharmacol., 161, 181±188.
Kenny, P.J., File, S.E. & Neal, M.J. (2000) Evidence for a complex in¯uenceof nicotinic acetylcholine receptors on hippocampal serotonin release.J. Neurochem., 75, 2409±2414.
Kulikov, A., MormeÁde, P. & Chaouloff, F. (1997) Effects of adrenalectomyand corticosterone replacement on diurnal [3H]citalopram binding in ratmidbrain. Neurosci. Lett., 222, 127±131.
Malberg, J.E., Sabol, K.E. & Seiden, L.S. (1996) Co-administration of MDMAwith drugs that protect against MDMA neurotoxicity produces differenteffects on body temperature in the rat. J. Pharmacol. Exp. Ther., 278, 258±267.
Malberg, J.E. & Seiden, L.S. (1998) Small changes in ambient temperaturecause large changes in 3,4-methylenedioxymethamphetamine (MDMA)-induced serotonin neurotoxicity and core body temperature in the rat.J. Neurosci., 18, 5086±5094.
Malpass, A., White, J.M., Irvine, R.J., Somogyi, A.A. & Bochner, F. (1999)Acute toxicity of 3,4-methylenedioxymethylamphetamine (MDMA) inSprague-Dawley and Dark Agouti rats. Pharmacol. Biochem. Behav., 64,29±34.
McCarty, R. & Kopin, I.J. (1979) Stress-induced alterations in plasmacatecholamines and behavior of rats: effects of chlorisondamine andbretylium. Behav. Neural Biol., 27, 249±265.
McNamara, M.G., Kelly, J.P. & Leonard, B.E. (1995) Some behavioural andneurochemical aspects of subacute 3,4-methylenedioxymethamphetamineadministration in rats. Pharmacol. Biochem. Behav., 52, 479±484.
Miller, D.B. & O'Callaghan, J.P. (1994) Environment-, drug- and stress-induced alterations in body temperature affect the neurotoxicity ofsubstituted amphetamines in the C57BL/6J mouse. J. Pharmacol. Exp.Ther., 270, 752±760.
Miller, A.H., Spencer, R.L., Stein, M. & McEwen, B.S. (1990) Adrenal steroidbinding in spleen and thymus after stress or dexamethasone. Am. J. Physiol.,193, E405±E412.
Molliver, M.E., Berger, U.V., Mamounas, L.A., Molliver, D.C., O'Hearn, E. &Wilson, M.A. (1990) Neurotoxicity of MDMA and related compounds.Anatomic studies. In Whitaker-Azmitia, P.M. & Peroutka, S.J. (eds), TheNeuropharmacology of Serotonin. Ann. NY Acad. Sci., Vol. 600. The NewYork Academy of Sciences, New York, pp. 640±661.
Nash, J.F. (1990) Ketanserin pretreatment blocks MDMA-induced dopaminerelease in the striatum as measured by in vivo microdialysis. Life Sci., 47,2401±2408.
Nash, J.F., Meltzer, H.Y. & Gudelsky, G.A. (1988) Elevation of serumprolactin and corticosterone concentrations in the rat after theadministration of 3,4-methylenedioxymethamphetamine. J. Pharmacol.Exp. Ther., 245, 873±879.
O'Cain, P.A., Hletko, S.B., Ogden, B.A. & Varner, K.J. (2000) Cardiovascularand sympathetic responses and re¯ex changes elicited by MDMA. Physiol.Behav., 70, 141±148.
O'Callaghan, J.P., Brinton, R.E. & McEwen, B.S. (1989) Glucocorticoidsregulate the concentration of glial ®brillary acidic protein throughout thebrain. Brain Res., 494, 159±161.
O'Callaghan, J.P., Jensen, K.F. & Miller, D.B. (1995) Quantitative aspects ofdrug and toxicant-induced astrogliosis. Neurochem. Int., 26, 115±124.
Pedersen, N.P. & Blessing, W.W. (2001) Cutaneous vasoconstrictioncontributes to hyperthermia induced by 3,4-methylenedioxymeth-amphetamine (ecstasy) in conscious rabbits. J. Neurosci., 21, 8648±8654.
Pollier, F., Sarre, S., Aguerre, S., Ebinger, G., MormeÁde, P., Michotte, Y. &Chaouloff, F. (2000) Serotonin reuptake inhibition by citalopram in ratstrains differing for their emotionality. Neuropsychopharmacology, 22, 64±76.
Reuben, M. & Clarke, P.B.S. (2000) Nicotine-evoked [3H]5-hydroxytrypt-amine release from rat striatal synaptosomes. Neuropharmacology, 39, 290±299.
Reul, J.M.H.M. & De Kloet, E.R. (1985) Two receptor systems forcorticosterone in rat brain: microdistribution and differential occupation.Endocrinology, 117, 2505±2512.
Stress hormones, MDMA and serotonergic systems 617
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 607±618

Ricaurte, G.A., DeLanney, L.E., Wiener, S.G., Irwin, I. & Langston, J.W.(1988) 5-Hydroxyindoleacetic acid in cerebrospinal ¯uid re¯ectsserotonergic damage induced by 3,4-methylenedioxymethamphetamine inCNS of non-human primates. Brain Res., 474, 359±363.
Rudnick, G. & Wall, S.C. (1992) The molecular mechanism of ! ecstasy @[3,4-methylenedioxy-methamphetamine (MDMA) ]: serotonin transportersare targets for MDMA-induced serotonin release. Proc. Natl Acad. Sci.USA, 89, 1817±1821.
Schmidt, C.J., Abbate, G.M., Black, C.K. & Taylor, V.L. (1990) Selective5-hydroxytryptamine2 receptor antagonists protect against the neurotoxicityof methylenedioxymethamphetamine in rats. J. Pharmacol. Exp. Ther., 255,478±483.
Schmidt, C.J. & Kehne, J.H. (1990) Neurotoxicity of MDMA. Neurochemicaleffects. In Whitaker-Azmitia, P.M. & Peroutka, S.J. (eds), TheNeuropharmacology of Serotonin. Ann. NY Acad. Sci., Vol. 600. TheNew York Academy of Sciences, New York, pp. 665±680.
Schmidt, C.J., Levin, J.A. & Lovenberg, W. (1987) In vitro and in vivoneurochemical effects of methylenedioxymethamphetamine on striatalmonoaminergic systems in the rat brain. Biochem. Pharmacol., 36, 747±755.
Schmidt, C.J., Sullivan, C.K. & Fadayel, G.M. (1994) Blockade of striatal5-hydroxytryptamine2 receptors reduces the increase in extracellularconcentrations of dopamine produced by the amphetamine analogue 3,4-methylenedioxymethamphetamine. J. Neurochem., 62, 1382±1389.
Sprague, J.E., Everman, S.L. & Nichols, D.E. (1998) An integrated hypothesisfor the serotonergic axonal loss induced by 3,4-methylenedioxy-methamphetamine. Neurotoxicology, 19, 427±442.
Steele, T.D., Nichols, D.E. & Yim, G.K.W. (1987) Stereochemical effects of3,4-methylenedioxymethamphetamine (MDMA) and related amphetaminederivatives on inhibition of uptake of [3H]monoamines into synaptosomesfrom different regions of rat brain. Biochem. Pharmacol., 36,2297±2303.
Sternberg, E.M., Glowa, J.R., Smith, M.A., Calogero, A.E., Hill, J.M.,Listwak, S.J., Aksentijevich, S., Chrousos, G.P., Wilder, R.L. & Gold, P.W.(1992) Corticotropin releasing hormone related behavioral andneuroendocrine responses to stress in Lewis and Fischer 344 rats. BrainRes., 570, 54±60.
Sternberg, E.M., Hill, J.M., Chrousos, G.P., Kamilaris, T., Listwak, S.J., Gold,P.W. & Wilder, R.L. (1989) In¯ammatory mediator-induced hypothalamic-pituitary-adrenal axis activation is defective in streptococcal cell wallarthritis-susceptible Lewis rats. Proc. Natl Acad. Sci. USA, 86,2374±2378.
White, S.R., Duffy, P. & Kalivas, P.W. (1994) Methylenedioxy-methamphetamine depresses glutamate-evoked neuronal ®ring andincreases extracellular levels of dopamine and serotonin in the nucleusaccumbens in vivo. Neuroscience, 62, 41±50.
Yamamoto, B.K. & Spanos, L.J. (1988) The acute effects ofmethylenedioxymethamphetamine on dopamine release in the awake-behaving rat. Eur. J. Pharmacol., 148, 195±203.
Yau, J.L.W., Kelly, P.A.T., Sharkey, J. & Seckl, J.R. (1994) Chronic 3,4-methylenedioxymethamphetamine administration decreases glucocorticoidand mineralocorticoid receptor, but increases 5-hydroxytryptamine1C
receptor gene expression in the rat hippocampus. Neuroscience, 61,31±40.
618 F. Fernandez et al.
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 607±618
Related Documents











