Influence of temperature on gonadal development of Ruditapes philippinarum (Adams and Reeve, 1850) with special reference to ingested food and energy balance Marina Delgado a, ⁎ , Alejandro Pérez Camacho b,1 a Institut de Recerca i Tecnologia Agroalimentaries, Centre de Aqüicultura, Carretera del Poble Nou, Km 5.5. E-43540 Sant Carles de la Rápita, Tarragona, Spain b Instituto Español de Oceanografía, Muelle de Ánimas s/n, E-15001 A Coruña, Spain Received 20 October 2005; received in revised form 14 November 2006; accepted 17 November 2006 Abstract This study evaluated the influence of temperature on gonadal development in Ruditapes philippinarum under controlled dietary conditions, in order to distinguish between the effect of temperature alone and the associated effect of temperature on ingestion. The experiment was therefore divided into two groups: group L (at temperatures of 14 °C and 18 °C, with a daily ingestion rate of 470–550 μg organic weight of phytoplankton per g clam live weight) and group H (at temperatures of 18 °C and 22 °C, with a daily ingestion rate of 1000–1100 μg organic weight of phytoplankton per g clam live weight). Both ingestion and respiration rates were calculated for each of the diets and temperatures. Gonadal development of the different groups of clams was monitored over the 70-day experimental period by means of image analysis techniques and calculation of the soft tissue dry weight of a standard clam. Experimental results show that when the amount of food ingested is similar a difference of temperature between 14 °C and 18 °C, or between 18 °C and 22 °C, has no significant effect on the rate of gonadal development, the effect of temperature possibly being masked by the differences in the respective energy balances. When a higher temperature coincides with a higher ingestion rate, as is often the case with naturally-occurring populations of bivalves, then gonadal development is greater, and faster, at the higher temperature, although this is mainly due to the greater amount of food ingested. When, in experimental (or environmental) conditions, a high temperature (18 °C) is associated with a low ingestion rate, a situation of negative energy balance arises, associated with a slow rate of gonadal development that takes place at the cost of the animal's energy reserves. When the temperature is reduced to 14 °C, there is a decrease in the ingestion rate and lower energy consumption, leading to a situation of positive energy balance. In these conditions gonadal development is slow and the surplus of available energy is, presumably, used to accumulate energy reserves, as is often the case during the stage of growth prior to the onset of gonadal development in many bivalves that inhabit geographical areas with noticeable seasonal variations. © 2006 Elsevier B.V. All rights reserved. Keywords: Energy balance; Gonadal development; Image analysis; Ingested food Ruditapes philippinarum; Temperature Aquaculture 264 (2007) 398 – 407 www.elsevier.com/locate/aqua-online ⁎ Corresponding author. Tel.: +34 977745427; fax: +34 977714138. E-mail addresses: [email protected] (M. Delgado), [email protected] (A. Pérez Camacho). 1 Tel.: +34 981205362; fax: +34 981229077. 0044-8486/$ - see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.aquaculture.2006.11.009

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

2007) 398–407www.elsevier.com/locate/aqua-online
Aquaculture 264 (
Influence of temperature on gonadal development of Ruditapesphilippinarum (Adams and Reeve, 1850) with special reference
to ingested food and energy balance
Marina Delgado a,⁎, Alejandro Pérez Camacho b,1
a Institut de Recerca i Tecnologia Agroalimentaries, Centre de Aqüicultura, Carretera del Poble Nou, Km 5.5. E-43540Sant Carles de la Rápita, Tarragona, Spain
b Instituto Español de Oceanografía, Muelle de Ánimas s/n, E-15001 A Coruña, Spain
Received 20 October 2005; received in revised form 14 November 2006; accepted 17 November 2006
Abstract
This study evaluated the influence of temperature on gonadal development in Ruditapes philippinarum under controlled dietaryconditions, in order to distinguish between the effect of temperature alone and the associated effect of temperature on ingestion.The experiment was therefore divided into two groups: group L (at temperatures of 14 °C and 18 °C, with a daily ingestion rate of470–550 μg organic weight of phytoplankton per g clam live weight) and group H (at temperatures of 18 °C and 22 °C, with adaily ingestion rate of 1000–1100 μg organic weight of phytoplankton per g clam live weight). Both ingestion and respiration rateswere calculated for each of the diets and temperatures. Gonadal development of the different groups of clams was monitored overthe 70-day experimental period by means of image analysis techniques and calculation of the soft tissue dry weight of a standardclam.
Experimental results show that when the amount of food ingested is similar a difference of temperature between 14 °C and18 °C, or between 18 °C and 22 °C, has no significant effect on the rate of gonadal development, the effect of temperature possiblybeing masked by the differences in the respective energy balances.
When a higher temperature coincides with a higher ingestion rate, as is often the case with naturally-occurring populations ofbivalves, then gonadal development is greater, and faster, at the higher temperature, although this is mainly due to the greateramount of food ingested.
When, in experimental (or environmental) conditions, a high temperature (18 °C) is associated with a low ingestion rate, asituation of negative energy balance arises, associated with a slow rate of gonadal development that takes place at the cost of theanimal's energy reserves. When the temperature is reduced to 14 °C, there is a decrease in the ingestion rate and lower energyconsumption, leading to a situation of positive energy balance. In these conditions gonadal development is slow and the surplus ofavailable energy is, presumably, used to accumulate energy reserves, as is often the case during the stage of growth prior to theonset of gonadal development in many bivalves that inhabit geographical areas with noticeable seasonal variations.© 2006 Elsevier B.V. All rights reserved.
Keywords: Energy balance; Gonadal development; Image analysis; Ingested food Ruditapes philippinarum; Temperature
⁎ Corresponding author. Tel.: +34 977745427; fax: +34 977714138.E-mail addresses: [email protected] (M. Delgado), [email protected] (A. Pérez Camacho).
1 Tel.: +34 981205362; fax: +34 981229077.
0044-8486/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.aquaculture.2006.11.009

399M. Delgado, A. Pérez Camacho / Aquaculture 264 (2007) 398–407
1. Introduction
Seasonal fluctuations in environmental parametersdetermine to a great extent the reproductive cycle ofbivalves. Temperature has traditionally been assigned amajor role, although the existence of sufficient nutri-tional reserves in the animal or the abundance of food inthe natural environment also play a major part in theevolution of the reproductive cycle (Sastry, 1970, 1979;Gimazane, 1972; Pérez-Camacho et al., 2003).
Environmental factors, by acting on the componentsof the energy balance equation (Bayne and Newell,1983), determine the amount of energy that is availablefor reproduction and/or somatic growth, and can have aconsiderable impact on reproduction. Environmentalstress can also limit energy reserves and lead to theadoption of strategies such as delaying the production ofgametes or reabsorbing already formed gametes, de-signed to ensure the survival of the adult specimens at thecost of a reduced level of reproductive activity (Gagdiland Bossert, 1970; Callow, 1979).
Most studies that have evaluated the influence ofenvironmental parameters on gonadal development ofbivalves in the natural environment suffer from atemporal and spatial coincidence of factors such astemperature and food availability that make it difficult tostudy their separate effects. With specific reference to R.philippinarum, there is usually an a posteriori relationestablished between the environmental conditions (tem-perature, chlorophyll, salinity) and gametogenesis,spawning period or conditioning index (Holland andChew, 1974; Beninger and Lucas, 1984; Laruelle et al.,1994; Xie and Burnell, 1994).
In the case of studies performed under controlledexperimental conditions the effect of these environmen-tal factors is not usually sufficiently well separated, theirsuperposition masking the interpretation of the resultsand preventing the effect of each of the factors on thereproductive cycle from being evaluated independently.An increase in temperature speeds up most of thephysiological processes, including clearing and inges-tion rates as well as those for respiration and growth(Walne, 1972; Griffiths and Griffiths, 1987; Hoffman,1983). This notwithstanding, in many studies of theinfluence of temperature on reproduction in bivalves thesame amount of food is usually offered to the specimens,regardless of the experimental temperature, and ignoringthe effect of temperature on the various physiologicalrates, in particular the rate at which food is ingested(Saucedo et al., 2001; Martínez and Pérez, 2003).
When experiments are conducted at a constant tem-perature the amount of food available conditions the
ingestion rate of bivalves. There is usually a direct re-lation between ingestion rates and the amount of foodpresent in the water, as has been shown by several dif-ferent studies carried out by our research group (Beiraset al., 1993; Albentosa et al., 1996; Pérez-Camacho et al.,2002; Delgado and Pérez-Camacho, 2002). However,what is commonly controlled in studies on the influenceof the amount of food on reproductive cycles is thequantity of food offered, rather than the amount of foodthat the bivalves actually ingest (Muranaka and Lannan,1984; Chaparro, 1990).
If the purpose of a study is to evaluate the effect ofenvironmental factors on reproduction, then it isessential to control food ingestion and metabolic energyconsumption, since these two parameters are the prin-cipal indicators of the amount of energy available forsomatic and/or gonadal growth.
In accordance with the above, in the present study onthe influence of temperature on the reproductive processof R. philippinarum it was proposed to study this factoron its own, eliminating the associated effect of tem-perature on ingestion. In order to do so, ingestion wascontrolled in our experiments, which were conducted attemperatures of 14 °C, 18 °C and 22 °C, and foodavailability was regulated to ensure that the ingestion offood by clams kept at the different temperatures re-mained the same, regardless of temperature. At the sametime gonadal development was monitored by means ofimage analysis techniques and the calculation of softtissue dry weight of a standard clam.
2. Materials and methods
2.1. Biological material
The clams (R. philippinarum) were gathered from abeach in the Ría de Arosa (NW Spain), with specimensmeasuring between 33 and 44 mm being selected for thepurposes of the experiment. The initial average length ofthe clams was 36.3±0.22 mm for experimental condi-tions 18L, 18H and 22H, and 38.8±0.22 mm for ex-perimental condition 14L.
2.2. Design and experimental conditions
The experiment was performed in 12 l plastic tanks,containing natural sea-water filtered through 1 μm in aflow-through circuit at a variable rate of l/hr, dependingon the different ingestion rates.Water salinity was 33‰.
The aim of the experiment was to verify the effect oftemperature on gonadal development over temperaturesranging between 14 °C and 22 °C, which are those
400 M. Delgado, A. Pérez Camacho / Aquaculture 264 (2007) 398–407
found on the Galician coast (NW Spain) during thereproductive season of R. philippinarum.
The clams were fed on a diet of the microalgae Iso-chrysis galbana clone T-ISO. The microalgae wereinitially cultured in 6 l jars and then transferred to 1000 ltanks. Walne medium (Walne, 1966) and industrialfertilizer were used for the jar and tank cultivation,respectively. The microalgae were harvested during thestationary growth phase.
The food was introduced into the circulating waterwith a variable-flow peristaltic pump, at a concentrationof approximately 100,000 cells of I.galbana/ml.
Since one of the experimental requirements was tomaintain identical ingestion rates (IR) at different tem-peratures, and taking into account the influence oftemperature on the IR of the clams, given the noticeabledifference in IR between clams kept at 14 °C and thosekept at 22 °C, which is also repeated in metabolic energyconsumption (Fig. 1), if the clams kept at 22 °C has thesame IR as those kept at 14 °C then this would lead to anextremely negative energy balance in the former, whichwould have a considerable effect on gonadal develop-ment (Delgado and Pérez-Camacho, 2005).
In order to minimise the effect of the differencesbetween energy balances at the different temperatures,the comparison of temperatures was divided into twosections, 14 °C–18 °C and 18 °C–22 °C. Thus, twolevels of IR were assayed, referred to as low (L) andhigh (H), whilst three different temperatures were used,14 °C, 18 °C and 22 °C, giving a total of 4 experimentalconditions: 14L, 18L, 18H and 22H (with 120 speci-mens per experimental condition).
The daily food ration was 750 μg organic weight ofphytoplankton per g clam live weight for experimental
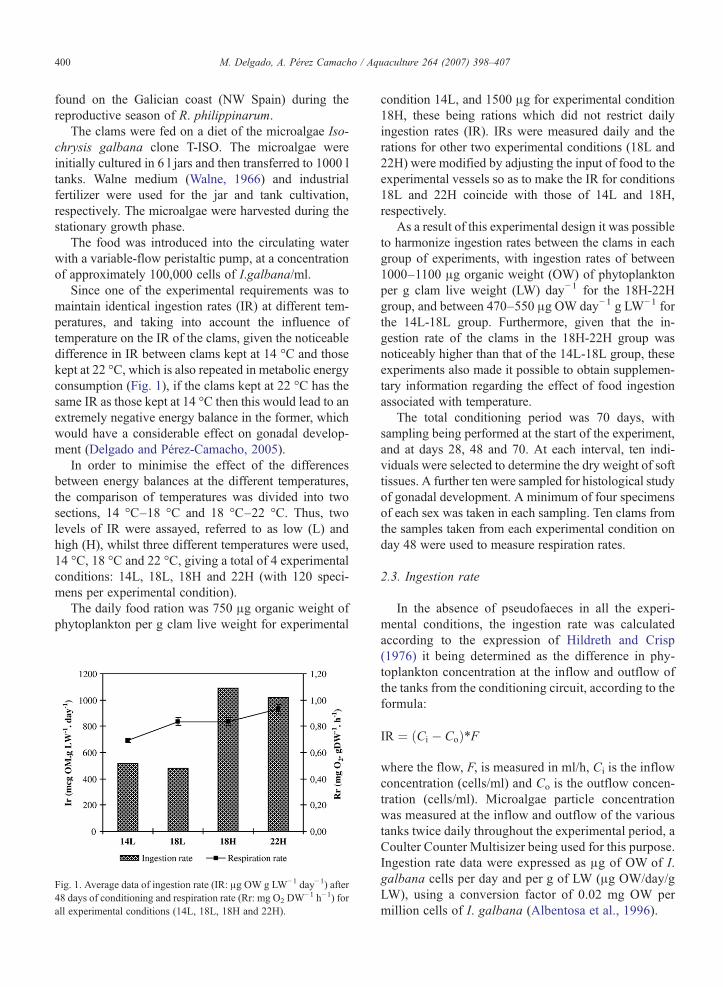
Fig. 1. Average data of ingestion rate (IR: μg OW g LW−1 day−1) after48 days of conditioning and respiration rate (Rr: mg O2 DW
−1 h−1) forall experimental conditions (14L, 18L, 18H and 22H).
condition 14L, and 1500 μg for experimental condition18H, these being rations which did not restrict dailyingestion rates (IR). IRs were measured daily and therations for other two experimental conditions (18L and22H) were modified by adjusting the input of food to theexperimental vessels so as to make the IR for conditions18L and 22H coincide with those of 14L and 18H,respectively.
As a result of this experimental design it was possibleto harmonize ingestion rates between the clams in eachgroup of experiments, with ingestion rates of between1000–1100 μg organic weight (OW) of phytoplanktonper g clam live weight (LW) day−1 for the 18H-22Hgroup, and between 470–550 μg OW day−1 g LW−1 forthe 14L-18L group. Furthermore, given that the in-gestion rate of the clams in the 18H-22H group wasnoticeably higher than that of the 14L-18L group, theseexperiments also made it possible to obtain supplemen-tary information regarding the effect of food ingestionassociated with temperature.
The total conditioning period was 70 days, withsampling being performed at the start of the experiment,and at days 28, 48 and 70. At each interval, ten indi-viduals were selected to determine the dry weight of softtissues. A further ten were sampled for histological studyof gonadal development. A minimum of four specimensof each sex was taken in each sampling. Ten clams fromthe samples taken from each experimental condition onday 48 were used to measure respiration rates.
2.3. Ingestion rate
In the absence of pseudofaeces in all the experi-mental conditions, the ingestion rate was calculatedaccording to the expression of Hildreth and Crisp(1976) it being determined as the difference in phy-toplankton concentration at the inflow and outflow ofthe tanks from the conditioning circuit, according to theformula:
IR ¼ ðCi � CoÞ*F
where the flow, F, is measured in ml/h, Ci is the inflowconcentration (cells/ml) and Co is the outflow concen-tration (cells/ml). Microalgae particle concentrationwas measured at the inflow and outflow of the varioustanks twice daily throughout the experimental period, aCoulter Counter Multisizer being used for this purpose.Ingestion rate data were expressed as μg of OW of I.galbana cells per day and per g of LW (μg OW/day/gLW), using a conversion factor of 0.02 mg OW permillion cells of I. galbana (Albentosa et al., 1996).

401M. Delgado, A. Pérez Camacho / Aquaculture 264 (2007) 398–407
2.4. Respiration rate
Respiration rates were measured by the rate ofdecline in oxygen partial pressure recorded by means ofoxygen sensors (510 BOD probe (YSI)) connected tooxygen meters (YSI Model 58).
Clams were placed in 700 ml respirometers filledwith air-saturated seawater. Eight replicates for eachtemperature were set up. An additional control flaskwithout animals was set at each temperature to checkvariation in oxygen concentration. The rate of oxygenconsumption (Rr) was calculated as follows:
Rr ¼ V=½t⁎ðCo−CtÞ�ðmgO2=hÞ
where V is the volume of the respirometer, t is themeasurement period and Co and Ct are the initial andfinal oxygen concentration in the respirometers.
2.5. Maturity indices: Soft tissue dry weight of a standardclam
In each sampling, the clams were measured andweighed, and their sex determined through microscopicexamination of a tissue smear. Thereafter, the adductormuscles were cut and the clams placed on their ventralsurface, allowing them to drain for 5 min. The softtissues were separated and freeze dried, taking thefreeze-dried weight as dry weight (DW).
At the start of the experiment, no statistically sig-nificant differences in DW were observed betweenspecimens from the four experimental conditions 18L,18H, 22H and 14L (ANOVA, pN0.05). The temporalevolution of the soft-tissue dry weight (DW) withreference to a specimen of constant size, should nor-mally correspond to the accumulation or loss of re-productive tissue (Pérez-Camacho, 1980; Beningerand Lucas, 1984; Pérez-Camacho et al., 2003);moreover, and to avoid variability caused by size dif-ferences, DW was corrected to a standard-sized in-dividual (LW=12 g: mean weight of the clams with amean length of 36.4 mm in the initial sampling), usingthe linear regression LW=a+bDW, calculated for eachsampling.
2.6. Maturity indices: Gonadal occupation index
A conventional histology protocol was followed. Thesoft tissues were fixed with Bouin's fixative (Bancroftand Stevens, 1996), sealed in paraffin, and 4 μm sliceswere taken. Para-Pak® Trichrome stain was used(Meridian Bioscience, Inc.).
For each specimen six fields of vision of the gonadwere chosen at random, corresponding to three differentdepths in the body of the clam. MicroImage software(Olympus) was used to process and analyze the imagesobtained, from both males and females, as described byDelgado and Pérez-Camacho (2003). These authorsdefine the gonadal occupation index (GOI) as follows:
GOI ¼ ðarea occupied by gametes=
area of the analyzed fieldÞ*100Þ
Each specimen was assigned a mean value for GOI,obtained from the 6 images analyzed in each case.
2.7. Statistical analysis
Comparisons between the soft tissue dry weight,respiration rate and gonadal occupation index under thedifferent experimental conditions were established byANOVA for a minimum significance level of 95%.Multiple comparisons between experimental conditionswere performed with the multiple rank test, using theleast significant difference (LSD)method. Cochran's testwas used to guarantee the homogeneity of the variances.Parameters expressed as percentages were modified byangular transformation (arcsine√%). TheMann-Whitneyand Kruskal-Wallis tests were used when the homoge-neity of the variances could not be ensured in two samplecomparisons. All the statistical analyses were performedwith Statgraphics plus 5.1 software, according to themethods described by Snedecor and Cochran (1980) andZar (1974).
3. Results
3.1. Ingestion rate
During the whole of the conditioning period theingestion rate of the clams in group H (18 °C and 22 °C)was much higher than those in group L (14 °C and18 °C). The mean values of the ingestion rates after48 days (Fig. 1) were similar within each experimentalgroup, fluctuating between 1000 and 1100 μg OW gLW−1 day−1 in group H, and between 470 and 550 μgOW g LW−1.day−1in group L. The ANOVA performedon the data corresponding to mean ingestion rates for 12-day periods, until the moment of the third sampling (day48), shows statistically significant differences (ANOVA,F: 78.96, n:16, pb0.001). The multiple range test (LSD)shows significant differences ( pb95%) between groupsH and L, whilst the differences within either group arenot statistically significant.
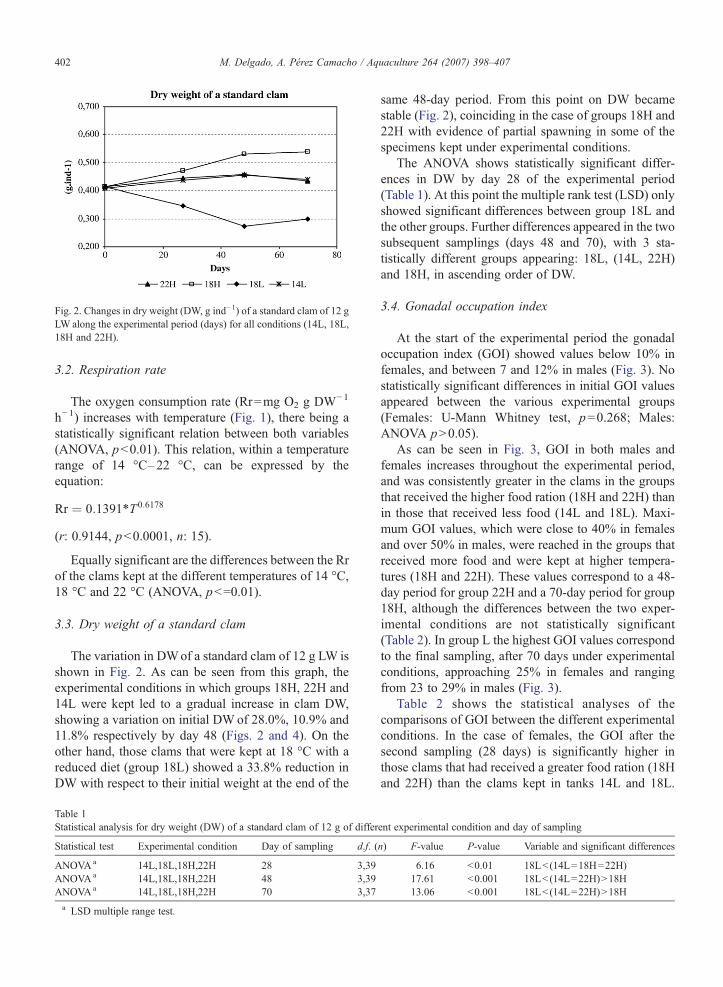
Fig. 2. Changes in dry weight (DW, g ind−1) of a standard clam of 12 gLW along the experimental period (days) for all conditions (14L, 18L,18H and 22H).
402 M. Delgado, A. Pérez Camacho / Aquaculture 264 (2007) 398–407
3.2. Respiration rate
The oxygen consumption rate (Rr=mg O2 g DW−1
h−1) increases with temperature (Fig. 1), there being astatistically significant relation between both variables(ANOVA, pb0.01). This relation, within a temperaturerange of 14 °C–22 °C, can be expressed by theequation:
Rr ¼ 0:1391*T0:6178
(r: 0.9144, pb0.0001, n: 15).
Equally significant are the differences between the Rrof the clams kept at the different temperatures of 14 °C,18 °C and 22 °C (ANOVA, pb=0.01).
3.3. Dry weight of a standard clam
The variation in DWof a standard clam of 12 g LW isshown in Fig. 2. As can be seen from this graph, theexperimental conditions in which groups 18H, 22H and14L were kept led to a gradual increase in clam DW,showing a variation on initial DW of 28.0%, 10.9% and11.8% respectively by day 48 (Figs. 2 and 4). On theother hand, those clams that were kept at 18 °C with areduced diet (group 18L) showed a 33.8% reduction inDW with respect to their initial weight at the end of the
Table 1Statistical analysis for dry weight (DW) of a standard clam of 12 g of differ
Statistical test Experimental condition Day of sampling d.f. (
ANOVAa 14L,18L,18H,22H 28 3,39ANOVAa 14L,18L,18H,22H 48 3,39ANOVAa 14L,18L,18H,22H 70 3,37
a LSD multiple range test.
same 48-day period. From this point on DW becamestable (Fig. 2), coinciding in the case of groups 18H and22H with evidence of partial spawning in some of thespecimens kept under experimental conditions.
The ANOVA shows statistically significant differ-ences in DW by day 28 of the experimental period(Table 1). At this point the multiple rank test (LSD) onlyshowed significant differences between group 18L andthe other groups. Further differences appeared in the twosubsequent samplings (days 48 and 70), with 3 sta-tistically different groups appearing: 18L, (14L, 22H)and 18H, in ascending order of DW.
3.4. Gonadal occupation index
At the start of the experimental period the gonadaloccupation index (GOI) showed values below 10% infemales, and between 7 and 12% in males (Fig. 3). Nostatistically significant differences in initial GOI valuesappeared between the various experimental groups(Females: U-Mann Whitney test, p=0.268; Males:ANOVA pN0.05).
As can be seen in Fig. 3, GOI in both males andfemales increases throughout the experimental period,and was consistently greater in the clams in the groupsthat received the higher food ration (18H and 22H) thanin those that received less food (14L and 18L). Maxi-mum GOI values, which were close to 40% in femalesand over 50% in males, were reached in the groups thatreceived more food and were kept at higher tempera-tures (18H and 22H). These values correspond to a 48-day period for group 22H and a 70-day period for group18H, although the differences between the two exper-imental conditions are not statistically significant(Table 2). In group L the highest GOI values correspondto the final sampling, after 70 days under experimentalconditions, approaching 25% in females and rangingfrom 23 to 29% in males (Fig. 3).
Table 2 shows the statistical analyses of thecomparisons of GOI between the different experimentalconditions. In the case of females, the GOI after thesecond sampling (28 days) is significantly higher inthose clams that had received a greater food ration (18Hand 22H) than the clams kept in tanks 14L and 18L.
ent experimental condition and day of sampling
n) F-value P-value Variable and significant differences
6.16 b0.01 18Lb (14L=18H=22H)17.61 b0.001 18Lb (14L=22H)N18H13.06 b0.001 18Lb (14L=22H)N18H
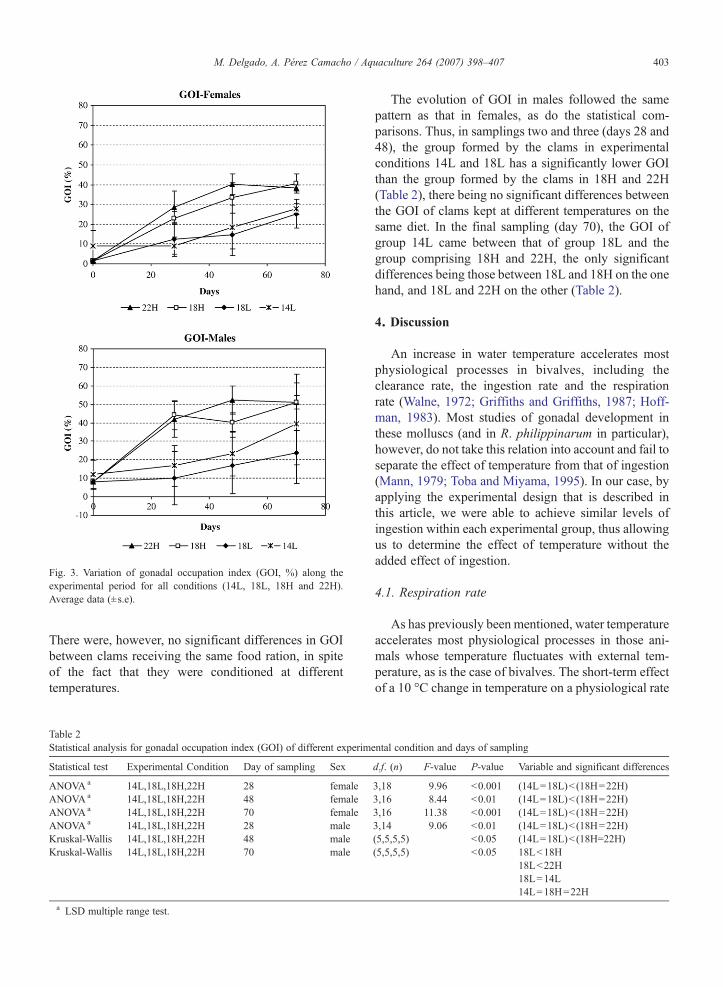
Fig. 3. Variation of gonadal occupation index (GOI, %) along theexperimental period for all conditions (14L, 18L, 18H and 22H).Average data (±s.e).
403M. Delgado, A. Pérez Camacho / Aquaculture 264 (2007) 398–407
There were, however, no significant differences in GOIbetween clams receiving the same food ration, in spiteof the fact that they were conditioned at differenttemperatures.
Table 2Statistical analysis for gonadal occupation index (GOI) of different experime
Statistical test Experimental Condition Day of sampling Sex
ANOVAa 14L,18L,18H,22H 28 femaleANOVAa 14L,18L,18H,22H 48 femaleANOVAa 14L,18L,18H,22H 70 femaleANOVAa 14L,18L,18H,22H 28 maleKruskal-Wallis 14L,18L,18H,22H 48 maleKruskal-Wallis 14L,18L,18H,22H 70 male
a LSD multiple range test.
The evolution of GOI in males followed the samepattern as that in females, as do the statistical com-parisons. Thus, in samplings two and three (days 28 and48), the group formed by the clams in experimentalconditions 14L and 18L has a significantly lower GOIthan the group formed by the clams in 18H and 22H(Table 2), there being no significant differences betweenthe GOI of clams kept at different temperatures on thesame diet. In the final sampling (day 70), the GOI ofgroup 14L came between that of group 18L and thegroup comprising 18H and 22H, the only significantdifferences being those between 18L and 18H on the onehand, and 18L and 22H on the other (Table 2).
4. Discussion
An increase in water temperature accelerates mostphysiological processes in bivalves, including theclearance rate, the ingestion rate and the respirationrate (Walne, 1972; Griffiths and Griffiths, 1987; Hoff-man, 1983). Most studies of gonadal development inthese molluscs (and in R. philippinarum in particular),however, do not take this relation into account and fail toseparate the effect of temperature from that of ingestion(Mann, 1979; Toba and Miyama, 1995). In our case, byapplying the experimental design that is described inthis article, we were able to achieve similar levels ofingestion within each experimental group, thus allowingus to determine the effect of temperature without theadded effect of ingestion.
4.1. Respiration rate
As has previously been mentioned, water temperatureaccelerates most physiological processes in those ani-mals whose temperature fluctuates with external tem-perature, as is the case of bivalves. The short-term effectof a 10 °C change in temperature on a physiological rate
ntal condition and days of sampling
d.f. (n) F-value P-value Variable and significant differences
3,18 9.96 b0.001 (14L=18L)b (18H=22H)3,16 8.44 b0.01 (14L=18L)b (18H=22H)3,16 11.38 b0.001 (14L=18L)b (18H=22H)3,14 9.06 b0.01 (14L=18L)b (18H=22H)(5,5,5,5) b0.05 (14L=18L)b (18H=22H)(5,5,5,5) b0.05 18Lb18H
18Lb22H18L=14L14L=18H=22H

Fig. 4. Gonadal occupation index (GOI, %) for males and females anddry weight (DW, %) increment, after 48 days and for all experimentalconditions (14L, 18L, 18H and 22H). EB: Energy balance.
404 M. Delgado, A. Pérez Camacho / Aquaculture 264 (2007) 398–407
can be measured by the thermal coefficient Q10 (Newelland Branch, 1980; Randal et al., 1997).
The metabolic rates of most animals with variablebody temperature increase two-to threefold for eachincrease of 10 °C in external temperature. However, themetabolic rates of some ectoderms show a high degreeof independence from temperature. Thus, certain inter-tidal invertebrates, which undergo major fluctuations inexternal temperature with the ebb and flow of the tides,have metabolic rates whose Q10 comes close to 1.0,their metabolic rates therefore changing very little withvariations in temperature of up to 20 °C (Randal et al.,1997).
In our experiment, the respiration rate measured attemperatures between 14 °C and 22 °C show a Q10 ofapproximately 1.65. In this case, the clams weregathered from the inter-tidal zone, which forms part ofthe natural habitat of R. philippinarum, this origin,according to Randal et al. (1997), being a possiblereason for the low value of their thermal coefficient.
Since oxygen consumptions were measured 28 daysafter the experiment commenced, another possible ex-planation for this low value for Q10 in this species couldbe thermal acclimatisation (Bayne and Newell, 1983).According to these authors, in situations of prolongedexposure to a change in temperature, the organism canadjust its oxygen consumption rate to a level which iscomparable to that it had before the change in tem-perature occurred.
4.2. Soft tissue dry weight of a standard clam
The change in clamDWover time varied according tothe different environmental conditions under which theexperiments took place (see Figs. 1 and 2). Thus, after48 days under experimental conditions, the DW of astandard 12 g LW clam increased by 28.0% in exper-imental conditions 18H, by 10.72% in 14L and 11.8% in22H, whilst it decreased by 33.8% in 18L (Fig. 4).
As has already been stated, in addition to its relationwith food ingestion, temperature has an influence onmetabolic energy consumption. Within the range oftemperatures assayed in our experiments, the oxygenconsumption rate is directly proportional to temperature(Fig. 1), in accordance with results described by Bodoyet al. (1986), Laing and Child (1996) and Isono et al.(1998) for this particular species.
Thus, the difference in DW between the clams ingroups 18H and 22H (Fig. 2), kept at 18 °C and 22 °Crespectively, and with the same level of ingestion, can beexplained by the increase in energy consumption thatoccurs when there is an increase in temperature (Fig. 1).
For the same reason, the DW values for groups 14L and22H are similar, even though the clams in 14L ingestedonly half the amount of food as ingested by those in 22H(it being possible, in this case, to consider the existenceof a situation of physiological stress, as described byToba and Miyama, in 1995, in R. philippinarum whentemperatures rise above 23 °C). Similarly, the differentrates of metabolic energy consumption would explainthe different results recorded for 14L and 18L, twogroups with similar levels of ingestion. Finally, thedifference in DW between 18L and 18H can be ex-plained by the difference in food ingested, with the lattergroup of clams ingesting twice as much as the former(Figs. 1 and 2).
The use of DWas an indicator of the degree of sexualmaturity of the clams can be somewhat problematic, aswe have already pointed out in a previous study of theeffect of food ingestion on sexual maturation in R. de-cussatus (Pérez-Camacho et al., 2003). If this criterionwere to be applied, we would be led to think that theclams in groups 14L and 18L differed widely in theirdegree of sexual maturity (although in general this doesnot correspond with the results of the GOI, which showsimilar states of gonadal development, as shown byFigs. 3 and 4). Likewise, the difference in DW betweenthe clams in groups 18H and 22H would indicate thatthey too have a different degree of sexual maturity(when their GOI is in fact similar). On the other hand,we would be led to believe that 14L and 22H, whichshare the same DW value, also have a similar degree ofgonadal development, whilst this clearly contradicts thefindings from their GOI (Figs. 3 and 4). Only in thecases of 14L and 18H, which would correspond to anexperimental (or environmental) situation in which norestrictions are placed on food ingestion, is DW a validindicator of the degree of sexual maturity, its values

405M. Delgado, A. Pérez Camacho / Aquaculture 264 (2007) 398–407
coinciding with those of the GOI (Figs. 3 and 4). Theseresults confirm the findings of our experiments onthe influence of diet on gonadal development inR. decussatus conditioned at a constant temperature of18 °C (Pérez-Camacho et al., 2003; Delgado and Pérez-Camacho, 2003). In the latter case, variations in DW in astandard clam were in direct relation to the food ration,and were a good indicator of the change in gonadaldevelopment and spawning over time, although only insituations of positive energy balance. However, in certainconditions, it is possible for a situation of negativeenergy balance, producing a decrease in DW, to co-existwith a process of gonadal development (Pérez-Camachoet al., 2003; Delgado and Pérez-Camacho, 2003).
4.3. Gonadal development
Sastry (1968), in his study of Aequipecten irradians,asserts that a minimum threshold temperature (which heplaces at 15 °C) is needed for reproductive activity tocommence, but that a higher temperature (20 °C) isrequired for the completion of gonadal development inthe said species. Navarro and Iglesias (1995) place thetemperature threshold for the onset of gonadal devel-opment in R. decussatus between 13.5–14.5 °C.
The evolution of GOI in our experiments shows thatthe gonad of R. philippinarum develops, at the veryleast, in a temperature gradient from 14 °C to 22 °C(Figs. 3 and 4). In the absence of factors (whetherexperimental or environmental) limiting ingestion, thehigher the temperature, the greater the amount ofingested food and the metabolic energy consumption(Fig. 1). At high temperatures this results in a rapidprocess of sexual maturation (experimental conditions18H and 22H), and a greater or lesser increase in DW,depending on the energy balance. At 14 °C the process ofsexual maturing is much slower (Fig. 3), possibly, to alarge extent, as a result of a reduced food intake (Fig. 1).
In our opinion, the effect of an increase in temperatureon gonadal development is fundamentally due to theincrease in ingestion rate that normally accompanies this,since when food restrictions ensure that ingestion ratesstay the same (18H and 22H; 14L and 18L), the degree ofgonadal development is the same at temperatures of18 °C and 22 °C, as is also the case at 14 °C and 18 °C.Similarly, Toba and Miyama (1995) conclude that, fortemperatures ranging from 10–27 °C, temperature isprobably not a limiting factor in the reproductive activityof R. philippinarum. Also, Navarro et al. (2000) allot amajor role in the gonadal development of Argopectenpurpuratus to the quality of the diet, and reject the effectof temperature within the range studied (16–20 °C).
Other authors, such as Mann (1979) or Wilson andSimons (1985), however, find a direct relation betweentemperature and the rate of gametogenesis. Mann (1979)suggests that in R. philippinarum gametogenesis de-pends directly on temperature and exposure time. Alongthe same lines, Utting and Doyou (1992) establish aminimum of 480 day degrees (number of days×degreesabove a critical minimum threshold of 10 °C) of con-ditioning for spawning to take place in R. philippi-narum. However, these results take neither energyconsumption nor ingested food into account, and arenot applicable to those situations, such as can arise undercertain natural conditions, in which food availability maybe limited.
When food availability is limited, a reduction infertility may occur which is proportional to the decreasein energy available for producing gametes, as has beenshown by Bayne et al. (1978) in Mytilus edulis. Thisbehavior would seem to be confirmed by the data fromour experiment corresponding to the clams conditionedat 18 °C on two different diets (H and L), which indicatea direct relation between the ingested food, the quantityof gonad developed and the degree of gonadal devel-opment, thus confirming the results of our previousstudy of R. decussatus (Delgado and Pérez-Camacho,2003). Navarro and Iglesias (1995) also suggest thatthere have to be an excess of energy available (fromeither stored reserves or available nutrients) for thespecific effects associated with temperature to appear inthe reproductive process of R. decussatus.
If we focus our attention to the energy balance, in-stead of ingestion, and compare two experimental groupswhich, although they ingested widely differing amountsof food, have similar energy balances, as is the case of14L and 22H in our experiment, we can see that althoughDW varies along very similar lines for both tempera-tures, sexual maturity is much greater at the highertemperature. This result leads one to think that temper-ature may exert a certain influence on the accumulationand utilization of energy reserves in bivalves, and thatwhilst at 22 °C the energy available is devoted to gonadaldevelopment and maintaining metabolic energy con-sumption, at 14 °C a large proportion of this energy isstored as an energy reserve, which would foreseeably betransformed into gonad when the temperature increases,as happens in certain species after the transition fromwinter to spring (Loosanoff, 1953; Sastry, 1968; Shafeeand Daoudi, 1991; Ponurovsky and Yakovlev, 1992).
From the above, and to summarize, we can define aseries of situations that set a framework for the repro-ductive behaviour of R. philippinarum with respect totemperature and diet. Thus, when a higher temperature

406 M. Delgado, A. Pérez Camacho / Aquaculture 264 (2007) 398–407
coincides with a greater ingestion of food (as is the caseof experimental conditions 18L versus 22H, and 14Lversus 18H in our experiment, and as commonly occursin natural populations of bivalves), gonadal developmentis much greater, and faster, at the higher temperature.
If the amount of food ingested is similar, then tem-perature differences of 4° (from 14 °C to 18 °C, andfrom 18 °C to 22 °C) have no significant impact on therate of gonadal development (probably as a result of thepossible effect of the increase in temperature on gonadaldevelopment being compensated for by the decrease inenergy available for reproduction when metabolic ex-penditure is also increased and the food ingestion rateremains the same).
If under experimental (or environmental) conditionsa high temperature is associated with a low rate ofingestion, as the result of limited food availability (ex-perimental condition 18L), then a situation of negativeenergy balance occurs, together with a slowing down ofgonadal development, and the resulting utilization of theanimal’s energy reserves for gonadal development.
When the temperature decreases to 14 °C there is adecrease in the ingestion rate, but at the same time energyconsumption also decreases, leading to a situation ofpositive energy balance (experimental condition 14L).Under these conditions, gonadal development is slow,and the available surplus energy is presumably used toaccumulate energy reserves, as occurs in many bivalvesthat live in regions with high seasonal variations duringthe stage preceding the commencement of gonadaldevelopment.
Acknowledgements
We are grateful to Carlota Rodríguez García andConcepción Fernández Pena for technical assistance. M.Delgado was supported by a post-doctoral grant fromthe Ministry of Education and Science of the Spanishgovernment (2002–2004). The authors would like tothank two anonymous reviewers whose commentsnotably improved the manuscript.
References
Albentosa, M., Pérez-Camacho, A., Labarta, U., Fernández Reiríz, M.J.,1996. Evaluation of live microalgal diets for the seed culture ofRuditapes decussatus using physiological and biochemical para-meters. Aquaculture 148, 11–23.
Bancroft, J.D., Stevens, A., 1996. Theory and practice of histologicaltechniques. Churchill Livingstone (Ed), New York.
Bayne, B.L., Newell, R.C., 1983. Physiological energetics of MarineMolluscs. In: Wilbur, K.M. (Ed.), The Mollusca, vol. 4. Saleuddin,Academic Press, New York, pp. 407–515.
Bayne, B.L., Holland, D.L., Moore, M.N., Lowe, D.M., Widdows, J.,1978. Further studies on the effects of stress in the adult on theeggs of Mytilus edulis. J. Mar. Biol. Assoc. U.K. 58, 825–841.
Beiras, R.A., Pérez Camacho, A., Albentosa, M., 1993. Influence offood concentration on energy balance and growth performance ofVenerupis pullastra seed reared in an open-flow system. Aquacul-ture 116, 353–365.
Beninger, P.G., Lucas, A., 1984. Seasonal variations in condition,reproductive activity, and gross biochemical composition of twospecies of adult clam reared in a common habitat: Tapes decussatus(L.) (Jeffreys) and Tapes philippinarum (Adams and Reeve). J. Exp.Mar. Biol. Ecol. 79, 19–37.
Bodoy, A., Maitre-Allain, T., Riva, A., 1986. Comparaison de larespiration chez Ruditapes decussatus (L.) et R. philippinarum(Adams and Reeve) en fonction de la temperature. Vie Milieu 36,83–89.
Callow, P., 1979. The cost of reproduction—A physiologicalapproach. Biol. Rev. 54, 23–40.
Chaparro, O.R., 1990. Effect of temperature and feeding onconditioning of Ostrea chilensis (Philippi, 1845) reproductors.Aquac. Fish. Manage. 21, 399–405.
Delgado, M., Pérez-Camacho, A., 2002. Efectos de la ración dealimento en el desarrollo gonadal de la almeja Ruditapesdecussatus (L.). Bol. Inst. Esp. Oceanogr. 18 (1–4), 293–300.
Delgado, M., Pérez-Camacho, A., 2003. A study of gonadaldevelopment in Ruditapes decussatus (L.) (Mollusca: bivalvia),using image analysis: influence of food ration and energy balance.J. Shellfish Res. 22 (2), 435–441.
Delgado, M., Pérez-Camacho, A., 2005. Histological study of gonadaldevelopment of Ruditapes Decussatus (L.) and their relationshipwith food amount. Sci. Mar. 69 (1), 87–97.
Gagdil, M., Bossert, W.H., 1970. Life historical consequences ofnatural selection. Am. Nat. 104, 1–25.
Gimazane, J.P., 1972. Étude experimentale de l’action de quelquesfacteurs externes sur la reprise de l’activité génitale de la coque,Cerastoderma edule (L.), Mollusque Bivalve. C. R .Soc. Biol.166, 587–589.
Griffiths, C.L., Griffiths, R.J., 1987. Bivalvia. In: Pandian, T.J.,Vernberg, F.J. (Eds.), Animal Energetics, vol. 2. Academic Press,New York, pp. 1–88.
Hoffman, K.H., 1983.Metabolic and enzyme adaptation to temperatureand pressure. In: Hochachka, P.W. (Ed.), The Mollusca, vol. 2.Academic Press, New York, pp. 220–256.
Holland, D.A., Chew, K.K., 1974. Reproductive cycle of the manilaclam (Venerupis japonica) from Hood Canal, Washington. Proc.Natl. Shellfish Ass. 64, 53–58.
Hildreth, D.I., Crisp, D.J., 1976. A corrected formula for calculation offiltration rate of bivalve molluscs in an experimental flowingsystem. J. Mar. Biol. Assoc. U.K. 56, 111–120.
Isono, R.S., Kita, J., Kishida, C., 1998. Upper temperature effect onrates of growth and oxygen consumption of the Japanese littleneckclam, Ruditapes philippinarum. Nippon Suisan Gakkaishi 64 (3),373–376.
Laing, I., Child, A.R., 1996. Comparative tolerance of small juvenilepalourdes (Tapes decussatus L.) and manila clams (Tapesphilippinarum Adams and Reeve) to low temperature. J. Exp.Mar. Biol. Ecol. 195, 267–285.
Laruelle, F., Guillou, J., Paulet, Y.M., 1994. Reproductive pattern ofthe clams Ruditapes decussatus and Ruditapes philippinarum onintertidal flats in Brittany. J. Mar. Biol. Assoc. U.K. 172, 69–96.
Loosanoff, V.L., 1953. Reproductive cycle in Cyprina islandica. Biol.Bull. 104, 146–155.

407M. Delgado, A. Pérez Camacho / Aquaculture 264 (2007) 398–407
Mann, R., 1979. The effect of temperature on growth, physiology, andgametogenesis in the manila clam Tapes philippinarum. J. Exp.Mar. Biol. Ecol. 38, 121–133.
Martínez, G., Pérez, H., 2003. Effect of different temperature regimeson reproductive conditioning in the scallop Argopecten purpur-atus. Aquaculture 228, 153–167.
Muranaka, M.S., Lannan, J.E., 1984. Broodstock management ofCrassostrea gigas: enviromental influences on broodstock condi-tioning. Aquaculture 39, 217–228.
Navarro, E., Iglesias, J.I.P., 1995. Energetics of reproduction related toenvironmental variability in bivalve molluscs. Haliotis 24, 43–55.
Navarro, J.M., Leiva, G.E., Marínez, G., Aguilera, C., 2000.Interactive effects of diet and temperature on the scope for growthArgopecten purpuratus during reproductive conditioning.J. Exp. Mar. Biol. Ecol. 247, 67–83.
Newell, R.C., Branch, G.M., 1980. The effects of temperature on themaintenance of metabolic energy balance in marine invertebrates.Adv. Mar. Biol. 17, 329–396.
Pérez-Camacho, A., 1980. Biología de Venerupis pullastra (Montagu,1803) y Venerupis decussata (Linné, 1767) (Mollusca: Bivalvia)con especial referencia a factores determinantes de la producción.Bol. Inst. Esp. Oceanogr. 281, 353–358.
Pérez-Camacho, A., Delgado, M., Albentosa, M., 2002. Influencia deltamaño y la concentración de alimento sobre las tasas deaclaramiento e ingestión de la almeja Ruditapes decussatus (L.).Bol. Inst. Esp. Oceanogr. 18 (1–4), 315–320.
Pérez-Camacho, A., Delgado, M., Labarta, U., Fernández-Reiriz, M.J.,2003. Energy balance, gonadal development and biochemicalcomposition in the clam Ruditapes decussatus (L.). Mar. Ecol.,Prog. Ser. 258, 133–145.
Ponurovsky, S.K., Yakovlev, Y.M., 1992. The reproductive biology ofthe Japanese littleneck, Tapes philippinarum (A. Adams and Reeve,1850) (Bivalvia: Veneridae). J. Shellfish Res. 11 (2), 265–277.
Randal, D., Burggen, W., French, K., 1997. Eckert animal physiology:mechanisms and adaptations. W.H. Freeman and Co., New York.
Sastry, A.N., 1968. The relationship among food, temperature, andgonad development of the bay scallops Aequipecten irradiansLamarck. Physiol. Zool. 41, 44–53.
Sastry, A.N., 1970. Reproductive physiological variation in latitudi-nally separated populations of the bay scallop, Aequipectenirradians Lamarck. Biol. Bull. 138, 56–65.
Sastry, A.N., 1979. Pelecypoda (excluding Ostreidae). In: Giese, A.C.,Pearse, J.S. (Eds.), Reproduction of Marine Invertebrates, vol. 5 (5).Academic Press, New York, pp. 113–292.
Saucedo, P., Rodríguez-Jaramillo, C., Aldana-Avilés, C., Monsalvo-Spencer, P., Reynoso-Granados, T., Villarreal, H., Monforte, M.,2001. Gonadic conditioning of the calafia mother-of-pearl oyster,Pinctada mazatlantica (Hanley, 1856), under two temperaturesregimes. Aquaculture 195, 103–119.
Shafee, M.S., Daoudi, M., 1991. Seasonal changes in the biochemicalcomposition and calorific content of the black scallopChlamys varia(L.) from Lanveoc, Bay of Brest. Oceanol. Acta 4 (3), 331–341.
Snedecor, G.W., Cochran, W.G., 1980. Statistical Methods, 7th ed.Iowa State University Press, Ames.
Toba, M., Miyama, Y., 1995. Influence of temperature on the sexualmaturation in manila clam, Ruditapes philippinarum. SuisanZoshoku 43 (3), 305–314.
Utting, S.D., Doyou, J., 1992. The increased utilization of egg lipidreserves following induction of triploidy in the manila clam (Tapesphilippinarum). Aquaculture 103, 17–28.
Walne, P.R., 1966. Experiments in the large-scale culture of the larvaeof Ostrea edulis L. Fish. Invest., II XXV (5), 1–62.
Walne, P.R., 1972. The influence of current speed, body size and watertemperature on the filtration rate of five species of bivalves. J. Mar.Biol. Assoc. U.K. 52, 345–374.
Wilson, J.H., Simons, S., 1985. Gametogenesis and breeding ofOstreaedulis on the west coast of Ireland. Aquaculture 46, 307–321.
Xie, Q., Burnell, G.M., 1994. A comparative study of the gametogeniccycles of the clams Tapes philippinarum (Adams and Reeve) andTapes decussatus (Linnaeus) on the south coast of Ireland.J. Shellfish Res. 13 (2), 467–472.
Zar, J.H., 1974. Biostatistical Analysis. Prentice-Hill, EnglewoodCliffs, N.J.
Related Documents











