BIOTECHNOLOGICAL PRODUCTS AND PROCESS ENGINEERING Influence of growth regulators and elicitors on cell growth and α-tocopherol and pigment productions in cell cultures of Carthamus tinctorius L. Smita P. Chavan & Vinayak H. Lokhande & Kirti M. Nitnaware & Tukaram D. Nikam Received: 5 July 2010 / Revised: 9 November 2010 / Accepted: 15 November 2010 # Springer-Verlag 2010 Abstract The present study examined the effects of plant growth hormones, incubation period, biotic (Trametes versicolor, Mucor sp., Penicillium notatum, Rhizopus stolonifer, and Fusarium oxysporum) and abiotic (NaCl, MgSO 4 , FeSO 4 , ZnSO 4 , and FeCl 3 ) elicitors on cell growth and α-tocopherol and pigment (red and yellow) productions in Carthamus tinctorius cell cultures. The cell growth and α-tocopherol and pigment contents improved significantly on Murashige and Skoog (MS) liquid medium containing 50.0 μM α-naphthalene acetic acid (NAA) and 2.5 μM 6-Benzyladenine (BA) at 28 days of incubation period. Incorporation of T. versicolor (50 mg l −1 ) signif- icantly enhanced the production of α-tocopherol (12.7- fold) and red pigment (4.24-fold). Similarly, supplemen- tation of 30 mg l −1 T. versicolor (7.54-fold) and 70 mg l −1 Mucor sp. (7.40-fold) significantly increased the produc- tion of yellow pigment. Among abiotic elicitors, NaCl (50–70 mg l −1 ) and MgSO 4 (10–30 mg l −1 ) significantly improved production of α-tocopherol (1.24-fold) and red pigment (20-fold), whereas yellow pigment content in- creased considerably by all the abiotic elicitor treatments. Taken together, the present study reports improved productions of α-tocopherol and the pigment as a stress response of safflower cell cultures exposed to these elicitors. Keywords Carthamus tinctorius . Cell culture . Elicitors . α-Tocopherol . Red and yellow pigment Introduction Carthamus tinctorius L. (safflower, Asteraceae) is an important oilseed crop cultivated throughout the semiarid regions of the world for its high content of linoleic acid (78% of total fatty acids; Velasco et al. 2005; Hojati et al. 2010). Safflower contains α-tocopherol, which accounts for more than 95% of the total tocopherols (α, β, γ, and δ; Furuya et al. 1987). The oil from the seeds of safflower is a premium edible oil since its consumption help to lower blood cholesterol, as α-tocopherol is an efficient scavenger of activated oxygen species (Caretto et al. 2004; Velasco et al. 2005). Besides, safflower also contains red (water- insoluble) and yellow (water-soluble) pigment utilized for producing herbal medicines, food colorants, cosmetics, textile, and natural dyes (Watanabe et al. 1997; Fatahi et al. 2008; Li et al. 2009). However, by limitation to de novo synthesis of α-tocopherol and pigment productions and due to unavailability of large sources of raw plant material for the extraction, extensive research has been conducted on synthetic production of these compounds to achieve the demand of the ever-growing human population. In recent years, public interest on the use of naturally synthesized compounds has grown rapidly due to its biosafety unlike synthetic compounds (Fatahi et al. 2008). Extraction of the compounds from safflower plant is often tedious and expensive as the plant is seasonal and not annual. An alternative method for the production of these specific metabolites is to use tissue culture techni- ques, which would ensure continuous supply of uniform quality, highly specialized, natural components which otherwise would not have been possible (Caretto et al. 2004). Such processes can easily be optimized during scale- up studies for commercial exploitation (Savitha et al. 2006; Zhao et al. 2010). S. P. Chavan : V. H. Lokhande : K. M. Nitnaware : T. D. Nikam (*) Department of Botany, University of Pune, Pune 411007 Maharashtra, India e-mail: [email protected] Appl Microbiol Biotechnol DOI 10.1007/s00253-010-3014-4

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

BIOTECHNOLOGICAL PRODUCTS AND PROCESS ENGINEERING
Influence of growth regulators and elicitors on cell growthand α-tocopherol and pigment productions in cell culturesof Carthamus tinctorius L.
Smita P. Chavan & Vinayak H. Lokhande &
Kirti M. Nitnaware & Tukaram D. Nikam
Received: 5 July 2010 /Revised: 9 November 2010 /Accepted: 15 November 2010# Springer-Verlag 2010
Abstract The present study examined the effects of plantgrowth hormones, incubation period, biotic (Trametesversicolor, Mucor sp., Penicillium notatum, Rhizopusstolonifer, and Fusarium oxysporum) and abiotic (NaCl,MgSO4, FeSO4, ZnSO4, and FeCl3) elicitors on cellgrowth and α-tocopherol and pigment (red and yellow)productions in Carthamus tinctorius cell cultures. The cellgrowth and α-tocopherol and pigment contents improvedsignificantly on Murashige and Skoog (MS) liquid mediumcontaining 50.0 μM α-naphthalene acetic acid (NAA) and2.5 μM 6-Benzyladenine (BA) at 28 days of incubationperiod. Incorporation of T. versicolor (50 mg l−1) signif-icantly enhanced the production of α-tocopherol (12.7-fold) and red pigment (4.24-fold). Similarly, supplemen-tation of 30 mg l−1 T. versicolor (7.54-fold) and 70 mg l−1
Mucor sp. (7.40-fold) significantly increased the produc-tion of yellow pigment. Among abiotic elicitors, NaCl(50–70 mg l−1) and MgSO4 (10–30 mg l−1) significantlyimproved production of α-tocopherol (1.24-fold) and redpigment (20-fold), whereas yellow pigment content in-creased considerably by all the abiotic elicitor treatments.Taken together, the present study reports improvedproductions of α-tocopherol and the pigment as a stressresponse of safflower cell cultures exposed to theseelicitors.
Keywords Carthamus tinctorius . Cell culture .
Elicitors .α-Tocopherol . Red and yellow pigment
Introduction
Carthamus tinctorius L. (safflower, Asteraceae) is animportant oilseed crop cultivated throughout the semiaridregions of the world for its high content of linoleic acid(78% of total fatty acids; Velasco et al. 2005; Hojati et al.2010). Safflower contains α-tocopherol, which accounts formore than 95% of the total tocopherols (α, β, γ, and δ;Furuya et al. 1987). The oil from the seeds of safflower is apremium edible oil since its consumption help to lowerblood cholesterol, as α-tocopherol is an efficient scavengerof activated oxygen species (Caretto et al. 2004; Velasco etal. 2005). Besides, safflower also contains red (water-insoluble) and yellow (water-soluble) pigment utilized forproducing herbal medicines, food colorants, cosmetics,textile, and natural dyes (Watanabe et al. 1997; Fatahi etal. 2008; Li et al. 2009). However, by limitation to de novosynthesis of α-tocopherol and pigment productions and dueto unavailability of large sources of raw plant material forthe extraction, extensive research has been conducted onsynthetic production of these compounds to achieve thedemand of the ever-growing human population. In recentyears, public interest on the use of naturally synthesizedcompounds has grown rapidly due to its biosafety unlikesynthetic compounds (Fatahi et al. 2008).
Extraction of the compounds from safflower plant isoften tedious and expensive as the plant is seasonal andnot annual. An alternative method for the production ofthese specific metabolites is to use tissue culture techni-ques, which would ensure continuous supply of uniformquality, highly specialized, natural components whichotherwise would not have been possible (Caretto et al.2004). Such processes can easily be optimized during scale-up studies for commercial exploitation (Savitha et al. 2006;Zhao et al. 2010).
S. P. Chavan :V. H. Lokhande :K. M. Nitnaware :T. D. Nikam (*)Department of Botany, University of Pune,Pune 411007 Maharashtra, Indiae-mail: [email protected]
Appl Microbiol BiotechnolDOI 10.1007/s00253-010-3014-4

Similar attempts have been made in the past forenhanced productions of α-tocopherol and pigment fromsafflower (Furuya et al. 1987; Hanagata et al. 1992, 1993,Hanagata and Karube 1994; Wakayama et al. 1994; Gao etal. 2000; Fatahi et al. 2008; Hojati et al. 2010). However, todate, there is no information available on the influence ofbiotic and abiotic elicitors on productions of thesecompounds in cell cultures of safflower. Therefore, thepresent investigation attempts to study such effects whichwould optimize the scale-up productions of these importantcompounds and would ease the downstream processes.
Materials and methods
Seed germination and establishment of calluses and cellcultures
Certified safflower (cv. Bhima) seeds procured fromNimkar Agriculture Research Institute (NARI, Phaltan,Maharashtra, India) were used as source of plant materialfor establishment of cell cultures. The seeds (500) weresuitably disinfected (Nikam and Shitole 1999) and thenplaced in a medium containing 1.0% (w/v) sucrose and0.8% (w/v) agar–agar (Himedia Pvt., India) only. The tubeswere incubated at 25±2 °C in total absence of light forgermination of seeds. Cotyledon explants from 7 days oldaxenic seedlings were used for the establishment ofcalluses.
These explants (∼1.0 cm2) were cultured aseptically onsolid Murashige and Skoog (MS) medium (Murashige andSkoog 1962) containing 3.0% (w/v) sucrose supplementedwith cytokinins [6-benzyladenine (BA) and kinetin (Kin);1.2–20.0 μM] and auxins like α-naphthalene acetic acid(NAA), indole-3-acetic acid (IAA), and 2,4-dichlorophe-noxyacetic acid (2,4-D) in varying concentrations of 1.2–50.0 μM. The pH of the medium was adjusted to 5.8 priorto autoclaving at 121 °C for 15 min. The cultures wereincubated at 25 ± 2 °C daily for 16 h in light[40 μmol m−2 s−1 photon flux density (PFD)] and 8 h indarkness. After 4 weeks, fresh weight (FW) of the callusesformed was determined. Dry weight (DW) was recordedafter oven-drying the calluses at 60 °C for 48 h. Themoisture content (M%) of the calluses were determinedusing the equation [(FW−DW)/FW] × 100.
The friable calluses (∼0.5 g of wet biomass) werecultured in MS liquid medium (50 ml per flask)supplemented with 50.0 μM NAA + 2.5 μM BA, andthe flasks were incubated on a gyratory shaker (Steel-mate, India) at 80 rpm under the same controlledconditions as described above. This cell culture wasperiodically subcultured on the same medium at aninterval of 4 weeks.
Quantitative studies of red and yellow pigment
Pigments were extracted from dried petals of safflower(Fatahi et al. 2008) and identified using thin layerchromatography (TLC; Rudometova et al. 2001). Themaximum absorption spectra of the pigment acetoneextracts were recorded by scanning in the range of 300–600 nm wavelengths using ultraviolet (UV)–visiblespectrophotometer-1800 (Shimadzu, Kyoto, Japan), andthe data thus obtained were used for quantitation of thepigments.
Influence of growth regulators and incubation periodon cell growth and α-tocopherol and pigment accumulation
Among the various concentrations of cytokinins (1.2–20.0 μM BA and Kin) and auxins (1.2–50.0 μM IAA,NAA, and 2,4-D) tested, the media which producedoptimum cell biomass were used for the estimation of cellgrowth and α-tocopherol and pigment accumulation.Influence of the incubation period (7, 14, 21, 28, and35 days) on cell growth and accumulation of the com-pounds has also been recorded.
α-Tocopherol was extracted from cell biomass (Panfili etal. 1994) and analyzed using high-performance liquidchromatography (HPLC). The isocratic HPLC system(DIONEX 170 AU, Germany) comprised a P-680 solventdelivery pump, a BDS Hypersil C18 (250×4.6 mm, 5 μm)column (Thermo scientific, Part No. 28105-254630), and aUV detector (170 U). The mobile phase involved methanol:water (98:2 v/v) with a flow rate of 1.0 ml min−1 at 25 °C.Samples of α-tocopherol (20 μl) were injected and scannedat 220 nm. The content in the samples was quantified bycomparing the peak area with that of an α-tocopherolstandard (Cat. No. 10191, Sigma-Aldrich, USA). Thepigments were extracted from cell biomass (Gao et al.2000) and estimated using a UV–visible spectrophotometer.The α-tocopherol and pigment contents were calculated andexpressed in terms of μg g−1 FW and mg g−1 DW,respectively.
Elicitor preparation and administration
The fungal strains viz., Trametes versicolor (NCIM 993),Mucor sp. (NCIM 881), Penicillium notatum (NCIM 745),Rhizopus stolonifer (NCIM 880), and Fusarium oxysporum(NCIM 1008) were procured from National Collection ofIndustrial Microorganism (NCIM), National ChemicalLaboratory (NCL, Pune, India). The fungal strains werecultured on liquid potato-dextrose medium containingpotato (200 g l−1), dextrose (20 g l−1) and pH 5.5 withtartaric acid (10%; Rajendran et al. 1994). The cultureswere incubated for 28 days under the same controlled
Appl Microbiol Biotechnol
conditions as described above. Then the cultures wereautoclaved at 121 °C for 15 min and the fungal mycelialmats were harvested for drying in a hot air oven at 60 °Cfor 48 h. The dried fungal mats were ground to fine powderand used as biotic elicitors. Different salts such as sodiumchloride (NaCl), ferrous sulfate (FeSO4), magnesiumsulfate (MgSO4), ferric chloride (FeCl3), and zinc sulfate(ZnSO4) were used as abiotic elicitors.
C. tinctorius cell cultures (1.0 g wet biomass) wereexposed to MS liquid medium containing 50.0 μM NAA +2.5 μM BA and varying concentrations (0, 10, 30, 50, or70 mg l−1) of the biotic or abiotic elicitors. The cultureswere incubated for 4 weeks on a gyratory shaker at 80 rpmunder the same controlled conditions as described above.The cultures were then used for the quantitation of α-tocopherol and pigment contents.
Statistical analysis
The experiments were carried out in a randomized blockdesign with three independent biological replicates andrepeated thrice. Data were subjected to one-way analysis ofvariance (ANOVA) and Duncan’s multiple range test(DMRT) was performed at 5% probability level todetermine the significant differences, using statisticalsoftware SPSS 10.0.
Results
Seed germination and calluses and cell cultureestablishment
The safflower seeds exhibited 90% germination after 7 daysof incubation. At the end of the second week, callusesinduction was observed from cotyledon explants, whichincreased progressively up to 4 weeks. It was observed thatMS medium supplemented with 50.0 μM NAA and 2.5 μMBA produced calluses having the highest FW, DW, and M% (data not shown); therefore, the same medium has beenused for further studies.
Influence of growth regulators and incubation periodon cell growth and α-tocopherol and pigment accumulation
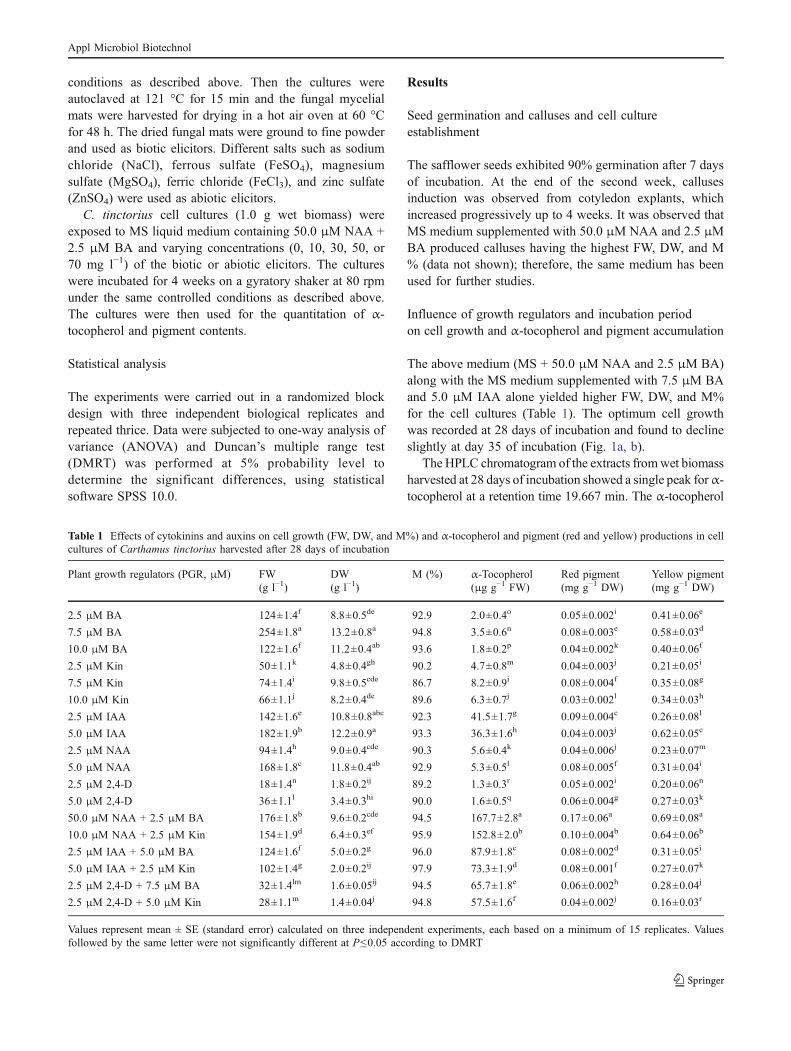
The above medium (MS + 50.0 μM NAA and 2.5 μM BA)along with the MS medium supplemented with 7.5 μM BAand 5.0 μM IAA alone yielded higher FW, DW, and M%for the cell cultures (Table 1). The optimum cell growthwas recorded at 28 days of incubation and found to declineslightly at day 35 of incubation (Fig. 1a, b).
The HPLC chromatogram of the extracts fromwet biomassharvested at 28 days of incubation showed a single peak forα-tocopherol at a retention time 19.667 min. The α-tocopherol
Table 1 Effects of cytokinins and auxins on cell growth (FW, DW, and M%) and α-tocopherol and pigment (red and yellow) productions in cellcultures of Carthamus tinctorius harvested after 28 days of incubation
Plant growth regulators (PGR, μM) FW(g l−1)
DW(g l−1)
M (%) α-Tocopherol(μg g−1 FW)
Red pigment(mg g−1 DW)
Yellow pigment(mg g−1 DW)
2.5 μM BA 124±1.4f 8.8±0.5de 92.9 2.0±0.4o 0.05±0.002i 0.41±0.06e
7.5 μM BA 254±1.8a 13.2±0.8a 94.8 3.5±0.6n 0.08±0.003e 0.58±0.03d
10.0 μM BA 122±1.6f 11.2±0.4ab 93.6 1.8±0.2p 0.04±0.002k 0.40±0.06f
2.5 μM Kin 50±1.1k 4.8±0.4gh 90.2 4.7±0.8m 0.04±0.003j 0.21±0.05i
7.5 μM Kin 74±1.4i 9.8±0.5cde 86.7 8.2±0.9i 0.08±0.004f 0.35±0.08g
10.0 μM Kin 66±1.1j 8.2±0.4de 89.6 6.3±0.7j 0.03±0.002l 0.34±0.03h
2.5 μM IAA 142±1.6e 10.8±0.8abc 92.3 41.5±1.7g 0.09±0.004c 0.26±0.08l
5.0 μM IAA 182±1.9b 12.2±0.9a 93.3 36.3±1.6h 0.04±0.003j 0.62±0.05c
2.5 μM NAA 94±1.4h 9.0±0.4cde 90.3 5.6±0.4k 0.04±0.006j 0.23±0.07m
5.0 μM NAA 168±1.8c 11.8±0.4ab 92.9 5.3±0.5l 0.08±0.005f 0.31±0.04i
2.5 μM 2,4-D 18±1.4n 1.8±0.2ij 89.2 1.3±0.3r 0.05±0.002i 0.20±0.06n
5.0 μM 2,4-D 36±1.1l 3.4±0.3hi 90.0 1.6±0.5q 0.06±0.004g 0.27±0.03k
50.0 μM NAA + 2.5 μM BA 176±1.8b 9.6±0.2cde 94.5 167.7±2.8a 0.17±0.06a 0.69±0.08a
10.0 μM NAA + 2.5 μM Kin 154±1.9d 6.4±0.3ef 95.9 152.8±2.0b 0.10±0.004b 0.64±0.06b
2.5 μM IAA + 5.0 μM BA 124±1.6f 5.0±0.2g 96.0 87.9±1.8c 0.08±0.002d 0.31±0.05i
5.0 μM IAA + 2.5 μM Kin 102±1.4g 2.0±0.2ij 97.9 73.3±1.9d 0.08±0.001f 0.27±0.07k
2.5 μM 2,4-D + 7.5 μM BA 32±1.4lm 1.6±0.05ij 94.5 65.7±1.8e 0.06±0.002h 0.28±0.04j
2.5 μM 2,4-D + 5.0 μM Kin 28±1.1m 1.4±0.04j 94.8 57.5±1.6f 0.04±0.002j 0.16±0.03r
Values represent mean ± SE (standard error) calculated on three independent experiments, each based on a minimum of 15 replicates. Valuesfollowed by the same letter were not significantly different at P≤0.05 according to DMRT
Appl Microbiol Biotechnol
content was expressed in terms of μg g−1 FW (Table 1).However, dried calluses showed negligible contents of α-tocopherol (data not shown). It was observed that the α-tocopherol content was directly proportional to the biomassin the cell cultures of C. tinctorius (Table 1; Fig. 1a).
The thin layer chromatogram showed the presence of redand yellow pigment as single spots with absorption maximaat 520 and 403 nm, respectively. Similar to α-tocopherol,maximum production of pigments was observed in thesame medium used to get maximum cell biomass (Table 1;Fig. 1b).
Influence of biotic elicitors on α-tocopherol and pigmentaccumulation
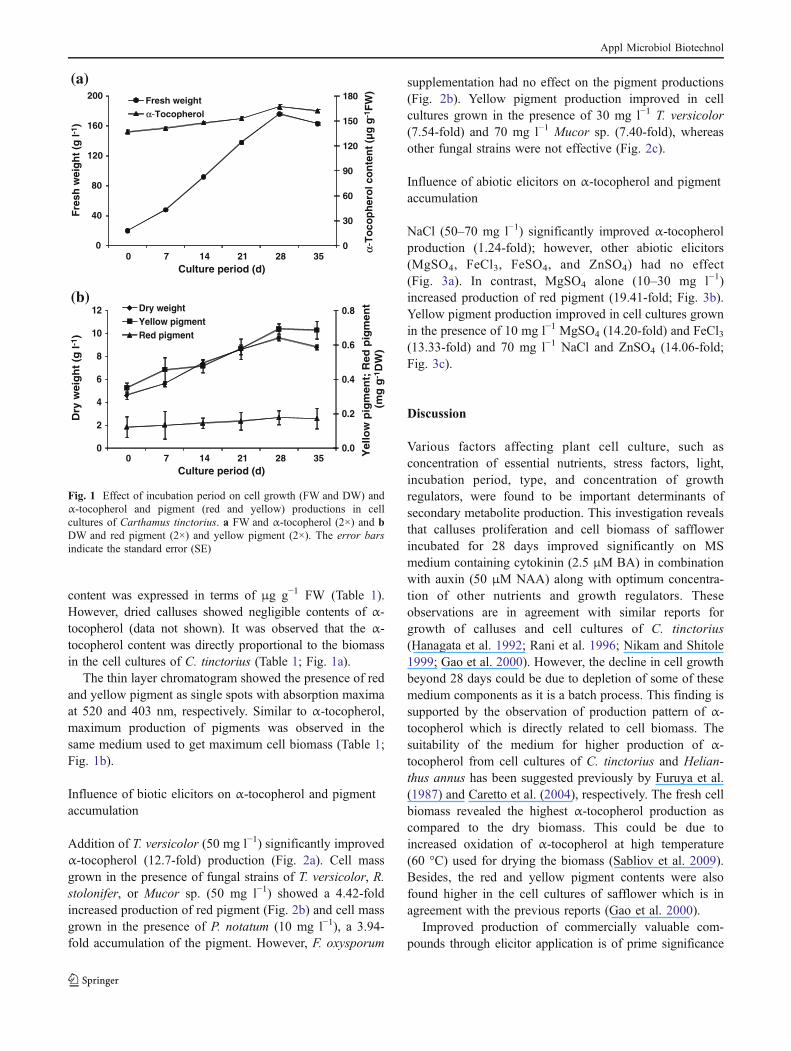
Addition of T. versicolor (50 mg l−1) significantly improvedα-tocopherol (12.7-fold) production (Fig. 2a). Cell massgrown in the presence of fungal strains of T. versicolor, R.stolonifer, or Mucor sp. (50 mg l−1) showed a 4.42-foldincreased production of red pigment (Fig. 2b) and cell massgrown in the presence of P. notatum (10 mg l−1), a 3.94-fold accumulation of the pigment. However, F. oxysporum
supplementation had no effect on the pigment productions(Fig. 2b). Yellow pigment production improved in cellcultures grown in the presence of 30 mg l−1 T. versicolor(7.54-fold) and 70 mg l−1 Mucor sp. (7.40-fold), whereasother fungal strains were not effective (Fig. 2c).
Influence of abiotic elicitors on α-tocopherol and pigmentaccumulation
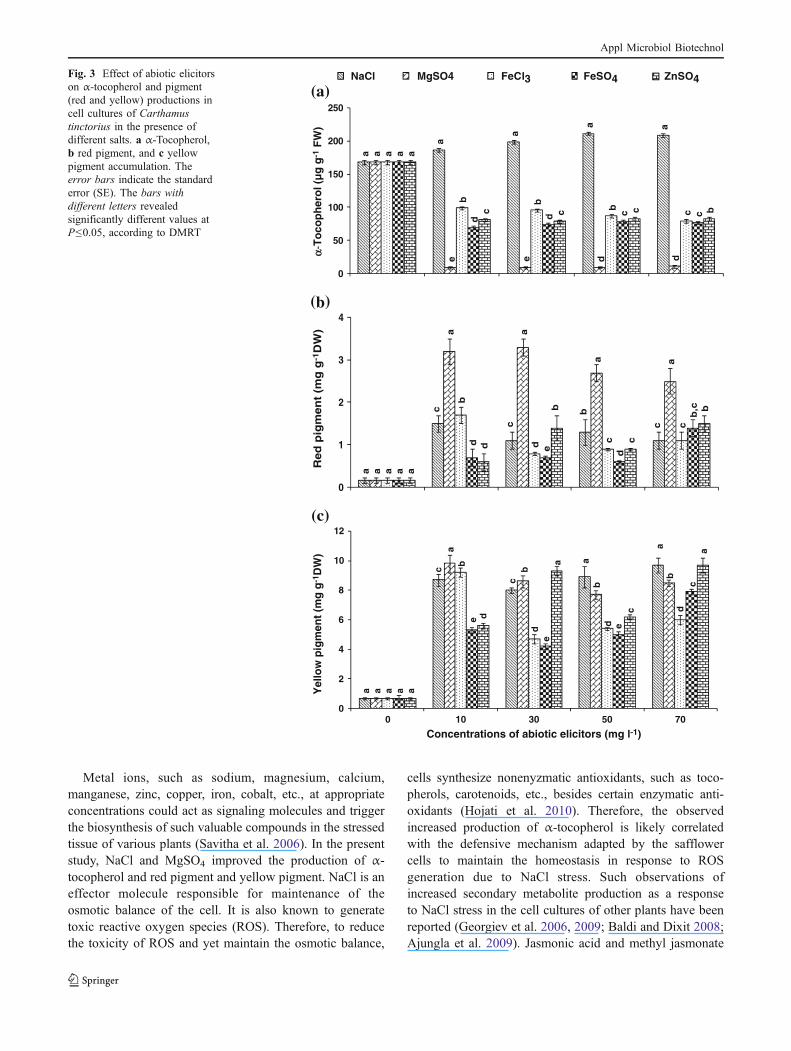
NaCl (50–70 mg l−1) significantly improved α-tocopherolproduction (1.24-fold); however, other abiotic elicitors(MgSO4, FeCl3, FeSO4, and ZnSO4) had no effect(Fig. 3a). In contrast, MgSO4 alone (10–30 mg l−1)increased production of red pigment (19.41-fold; Fig. 3b).Yellow pigment production improved in cell cultures grownin the presence of 10 mg l−1 MgSO4 (14.20-fold) and FeCl3(13.33-fold) and 70 mg l−1 NaCl and ZnSO4 (14.06-fold;Fig. 3c).
Discussion
Various factors affecting plant cell culture, such asconcentration of essential nutrients, stress factors, light,incubation period, type, and concentration of growthregulators, were found to be important determinants ofsecondary metabolite production. This investigation revealsthat calluses proliferation and cell biomass of safflowerincubated for 28 days improved significantly on MSmedium containing cytokinin (2.5 μM BA) in combinationwith auxin (50 μM NAA) along with optimum concentra-tion of other nutrients and growth regulators. Theseobservations are in agreement with similar reports forgrowth of calluses and cell cultures of C. tinctorius(Hanagata et al. 1992; Rani et al. 1996; Nikam and Shitole1999; Gao et al. 2000). However, the decline in cell growthbeyond 28 days could be due to depletion of some of thesemedium components as it is a batch process. This finding issupported by the observation of production pattern of α-tocopherol which is directly related to cell biomass. Thesuitability of the medium for higher production of α-tocopherol from cell cultures of C. tinctorius and Helian-thus annus has been suggested previously by Furuya et al.(1987) and Caretto et al. (2004), respectively. The fresh cellbiomass revealed the highest α-tocopherol production ascompared to the dry biomass. This could be due toincreased oxidation of α-tocopherol at high temperature(60 °C) used for drying the biomass (Sabliov et al. 2009).Besides, the red and yellow pigment contents were alsofound higher in the cell cultures of safflower which is inagreement with the previous reports (Gao et al. 2000).
Improved production of commercially valuable com-pounds through elicitor application is of prime significance
0.0
0.2
0.4
0.6
0.8
0
2
4
6
8
10
12
Yel
low
pig
men
t; R
ed p
igm
ent
(mg
g-1
DW
)
Dry
wei
gh
t (g
l-1 )
Dry weightYellow pigmentRed pigment
(a)
(b)
α-T
oco
ph
ero
l co
nte
nt
(µg
g-1
FW
)
0
30
60
90
120
150
180
0
40
80
120
160
200
0 7 14 21 28 35
0 7 14 21 28 35
Fre
sh w
eig
ht
(g l-
1 )
Culture period (d)
Culture period (d)
Fresh weight
α-Tocopherol
Fig. 1 Effect of incubation period on cell growth (FW and DW) andα-tocopherol and pigment (red and yellow) productions in cellcultures of Carthamus tinctorius. a FW and α-tocopherol (2×) and bDW and red pigment (2×) and yellow pigment (2×). The error barsindicate the standard error (SE)
Appl Microbiol Biotechnol

to the bioindustry (Murphy et al. 2007). It depends onparameters, such as elicitor’s specificity, concentration, andexposure time, as well as culture conditions (nutrientcomposition of the medium, growth regulators, and light)and growth stages of the cell culture (Wiktorowska et al.2010). In the present investigation, cell biomass obtained inthe presence of fungi like T. versicolor, R. stolonifer, andMucor sp. significantly improved the productions of α-tocopherol and pigment. It has been suggested that theelicitors at appropriate concentration might act as signalingmolecules which could be perceived by a receptor present onthe plasma membrane thus initiating the complex signal
transduction network involving regulation of gene expres-sion responsible for biosynthesis of target compounds (Zhaoet al. 2005; Savitha et al. 2006). Therefore, it may beinferred from the observations that the variation in yield ofα-tocopherol and pigment might be due to the diversity andspecificity associated with the interactions between fungalelicitor signals and plant cell receptors, the complexity ofelicitor signal transduction and thereafter defense responsesin plant cells (Zhao et al. 2001). Similar reports of animproved production of secondary metabolites have beenreported for cell cultures of Xanthophyllomyces dendrorhousand Rubia tinctorum (Wang et al. 2006; Orban et al. 2008).
(a)
(b)
(c)
a
d
e
c
d
a
a
b
b
c
a
c
c
a
a
a
b,c
a
a
b
a
b d
a
c
a
e
d
e
e
a d
c
d d
a
c
b
a
a
a
b
a
c
b
a
a
e
b
c
T. versicolor R. stoloniferMucor sp.P. notatumF. oxysporum
a c c c ca c c c,d
ca c c d da
a a
a
a
a
b b b b
0
500
1000
1500
2000
2500
αα-T
oco
ph
ero
l (µ
g g
-1F
W)
0
0.25
0.5
0.75
1R
ed p
igm
ent
(mg
g-1
DW
) Y
ello
w p
igm
ent
(mg
g-1
DW
)
0
1
2
3
4
5
6
0 10 30 50 70Concentrations of biotic elicitors (mg l-1)
Fig. 2 Effect of biotic elicitorson α-tocopherol and pigment(red and yellow) productions incell cultures of Carthamustinctorius in the presence ofdifferent fungi. a α-Tocopherol,b red pigment, and c yellowpigment accumulation. Theerror bars indicate the standarderror (SE). The bars withdifferent letters revealedsignificantly different values atP≤0.05, according to DMRT
Appl Microbiol Biotechnol
Metal ions, such as sodium, magnesium, calcium,manganese, zinc, copper, iron, cobalt, etc., at appropriateconcentrations could act as signaling molecules and triggerthe biosynthesis of such valuable compounds in the stressedtissue of various plants (Savitha et al. 2006). In the presentstudy, NaCl and MgSO4 improved the production of α-tocopherol and red pigment and yellow pigment. NaCl is aneffector molecule responsible for maintenance of theosmotic balance of the cell. It is also known to generatetoxic reactive oxygen species (ROS). Therefore, to reducethe toxicity of ROS and yet maintain the osmotic balance,
cells synthesize nonenyzmatic antioxidants, such as toco-pherols, carotenoids, etc., besides certain enzymatic anti-oxidants (Hojati et al. 2010). Therefore, the observedincreased production of α-tocopherol is likely correlatedwith the defensive mechanism adapted by the safflowercells to maintain the homeostasis in response to ROSgeneration due to NaCl stress. Such observations ofincreased secondary metabolite production as a responseto NaCl stress in the cell cultures of other plants have beenreported (Georgiev et al. 2006, 2009; Baldi and Dixit 2008;Ajungla et al. 2009). Jasmonic acid and methyl jasmonate
a
c
c
a
a
a
a
b
b
b
a
b
d
d
d
a
e
e
e
c
a
d
a
c
a0
2
4
6
8
10
12
0 10 30 50 70
Yel
low
pig
men
t (m
g g
-1D
W)
Concentrations of abiotic elicitors (mg l-1)
(b)
(c)
NaCl MgSO4 FeCl3 FeSO4 ZnSO4(a)
a
c
c
b
c
a
a a
a a
a
b
d c
c
a
d
e d
b,c
a
d
b
c
b
0
1
2
3
4R
ed
pig
me
nt
(mg
g-1
DW
)
a
a
a
a a
a
e e d d
a
b b
b c
a
d d c c
a
c c c b
0
50
100
150
200
250
αα-T
oco
ph
ero
l (µ
g g
-1 F
W)
Fig. 3 Effect of abiotic elicitorson α-tocopherol and pigment(red and yellow) productions incell cultures of Carthamustinctorius in the presence ofdifferent salts. a α-Tocopherol,b red pigment, and c yellowpigment accumulation. Theerror bars indicate the standarderror (SE). The bars withdifferent letters revealedsignificantly different values atP≤0.05, according to DMRT
Appl Microbiol Biotechnol

have been used widely as abiotic elicitors (Georgiev et al.2006; Baldi and Dixit 2008); however, the presentinvestigation suggests NaCl and MgSO4 as inexpensiveabiotic elicitors for improved productions of α-tocopheroland pigment in the cell cultures of safflower.
The results thus obtained on the administration of bioticand abiotic elicitors in safflower cell cultures suggest aneffective and suitable approach to overcome the limitedproductions of α-tocopherol and pigment from safflowerplants. In addition, the system will provide year-round supplyof these compounds and a healthy alternative for humanbeings. Since most of the elicitor chemicals are commerciallyavailable or can be prepared and easily administered to cellcultures in the laboratory, they are practically suitable forlarge-scale production of secondary metabolites.
Acknowledgments SPC is grateful to UGC-DRS-SAP-Phase III andASSIST and DST FIST Programme, Government of India for financialsupport. The author is also thankful to Dr. Ghosh, Dr. Suprsanna,Dr. Pal and Dr. Jadhav for their help in the correction of manuscript.
References
Ajungla L, Patil PP, BarmukhRB,NikamTD (2009) Influence of biotic andabiotic elicitors on accumulation of hyoscyamine and scopolamine inroot cultures of Datura metel L. Indian J Biotechnol 8:317–322
Baldi A, Dixit VK (2008) Enhanced artemisinin production by cell culturesof Artemisia annua. Curr Trends Biotechnol Pharm 2(2):341–348
Caretto S, Speth EB, Fachechi C, Gala R, Zacheo G, Giovinazzo G(2004) Enhancement of vitamin E production in sunflower cellcultures. Plant Cell Rep 23:174–179
Fatahi F, Carapetian J, Heidari R (2008) Spectroscopic measurement ofvaluable pigments from petals of safflower (Carthamus tinctorius L.)and their identification by TLC method. Res J Biol Sci 3(7):761–763
Furuya T, Yoshikawa T, Kimura T, Kaneko H (1987) Production oftocopherols by cell culture of safflower. Phytochemistry 26(10):2741–2747
Gao WY, Fan L, Paek KY (2000) Yellow and red pigment productionby cell cultures of Carthamus tinctorius L. in a bioreactor. PlantCell Tiss Org Cult 60:95–100
Georgiev M, Kuzeva S, Pavlov A, Kovacheva E, Ilieva M (2006)Enhanced rosmarinic acid production by Lavandula vera MMcell suspension culture through elicitation with vanadyl sulfate. ZNaturforsch 61:241–244
Georgiev MI, Weber J, Maciuk A (2009) Bioprocessing of plant cellcultures for mass production of targeted compounds. ApplMicrobiol Biotechnol 83:809–823
Hanagata N, Karube I (1994) Red pigment production by Carthamustinctorius cells in a two-stage culture system. J Biotechnol 31(1):59–65
Hanagata N, Ito A, Fukuju Y, Murata K (1992) Red pigmentformation in cultured cells of Carthamus tinctorius L. BiosciBiotechnol Biochem 56:44–47
Hanagata N, Ito A, Uehara H (1993) Behavior of cell aggregate ofCarthamus tinctorius L. cultured cells and correlation with redpigment formation. J Biotechnol 30:259–269
Hojati M, Modarres-Sanavy SAM, Karimi M, Ghanati F (2010)Responses of growth and antioxidant systems in Carthamustinctorius L. under water deficit stress. Acta Physiol Plant.doi:10.1007/s11738-010-0521-y
Li HX, Han SY, Wang XW, Maa X, Zhang K, Wang L, Maa ZZ, TuPF (2009) Effect of the carthamins yellow from Carthamustinctorius L. on hemorheological disorders of blood stasis in rats.Food Chem Toxicol 47:1797–1802
Murashige T, Skoog F (1962) A revised medium for rapid growth andbioassay with tobacco tissue cultures. Physiol Plant 15:473–497
Murphy T, Parra R, Radman R, Roy I, Antony HA, Dixon K, KeshavarzT (2007) Novel application of oligosaccharides as elicitors for theenhancement of bacitracin A production in cultures of Bacilluslicheniformis. Enzyme Microb Technol 40(6):1518–1523
Nikam TD, Shitole MG (1999) In vitro culture of safflower L. cv.Bhima: initiation, growth optimization and organogenesis. PlantCell Tiss Org Cult 55:15–22
Orban N, Boldizsar I, Szucs Z, Danos B (2008) Influence of differentelicitors on the synthesis of anthraquinone derivatives in Rubiatinctorum L. cell suspension cultures. Dyes Pigm 77(1):249–257
Panfili G, Manzi P, Pizzoferrato L (1994) High-performance liquidchromatographic method for the simultaneous determination oftocopherols, carotenes, and retinol and its geometric isomers inItalian cheeses. Analyst 119:1161–1165
Rajendran L, Suvarnalatha G, Ravishankar GA, Venkataraman LV(1994) Enhancement of anthocyanin production in callus culturesof Daucus carota L. under the influence of fungal elicitors. ApplMicrobiol Biotechnol 42:227–231
Rani KJ, Rao TN, Radhunatham G, Rao PV (1996) Studies on callusgrowth and differentiation in safflower. Indian J Genet PlantBreed 56(4):458–461
Rudometova NV, Pasovskij AP, Blohina EA (2001) Method ofisolation and identification of carthamin from safflower. Appli-cation’s perspectives in Russian food products. In: Bergman JW,Mundel HH (eds) Proc 5th Int Safflower Conf, July 23–27,Williston, North Dakota and Sidney, Montana, USA, pp 163
Sabliov CM, Fronczek C, Astete CE, Khachaturyan M, KhachatryanL, Leonardi C (2009) Effects of temperature and UV light ondegradation of α-tocopherol in free and dissolved form. J Am OilChem Soc 86:895–902
Savitha BC, Thimmaraju R, Bhagyalakshmi N, Ravishnkar GA (2006)Different biotic and abiotic elicitors influence betalain productionin hairy root cultures of Beta vulgaris in shake flask andbioreactor. Process Biochem 41:50–60
Velasco L, Perez-Vich B, Fernandez-Martinez JM (2005) Identifica-tion and genetic characterization of a safflower mutant with amodified tocopherol profile. Plant Breed 124:459–463
Wakayama S, Kusaka K, Kanehira T (1994) Kinobeon A, a novel redpigment produced in safflower tissue culture systems. ZNaturforsch 49:1–5
Wang W, Yu L, Zhou P (2006) Effects of different fungal elicitors ongrowth, total carotenoids and astaxanthin formation by Xantho-phyllomyces dendrorhous. Bioresour Technol 97:26–31
Watanabe T, Hasegawa N, Yamamota A, Nagai S, Terabe S (1997)Separation and determination of yellow and red safflowerpigments in food by capillary electrophoresis. Biosci Biotech61(7):1179–1183
Wiktorowska E, Długosz M, Janiszowska W (2010) Significantenhancement of oleanolic acid accumulation by biotic elicitorsin cell suspension cultures of Calendula officinalis L. EnzymeMicrob Technol 46(1):14–20
Zhao J, Zhu WH, Hu Q (2001) Selection of fungal elicitors to increaseindole alkaloid accumulation in Catharanthus roseus suspensioncell culture. Enzyme Microb Technol 28:666–672
Zhao J, Lawrence CD, Verpoorte R (2005) Elicitor signal transductionleading to production of plant secondary metabolites. BiotechnolAdv 23:283–333
Zhao JL, Zhou LG, Wu JY (2010) Effects of biotic and abioticelicitors on cell growth and tanshinone accumulation in Salviamiltiorrhiza cell cultures. Appl Microb Biotechnol 87:137–144
Appl Microbiol Biotechnol
Related Documents











