Basic nutritional investigation Influence of dietary blueberry and broccoli on cecal microbiota activity and colon morphology in mdr1a / mice, a model of inflammatory bowel diseases Gunaranjan Paturi Ph.D. a, * , Tafadzwa Mandimika Ph.D. b , Christine A. Butts Ph.D. b , Shuotun Zhu Ph.D. c , Nicole C. Roy Ph.D. d, e , Warren C. McNabb Ph.D. d, e , Juliet Ansell D.Phil. b, e a The New Zealand Institute for Plant and Food Research Limited, Private Bag 92169, Auckland 1142, New Zealand b The New Zealand Institute for Plant and Food Research Limited, Private Bag 11600, Palmerston North 4442, New Zealand c School of Medical Sciences, The University of Auckland, Auckland 1142, New Zealand d AgResearch Grasslands, Palmerston North 4442, New Zealand e Riddet Institute, Massey University, Palmerston North 4442, New Zealand article info Article history: Received 11 November 2010 Accepted 29 July 2011 Keywords: Bacterial translocation Colon inflammation Functional foods Gut health Large intestine Short-chain fatty acids abstract Objective: Enteric microbiota has been shown to be associated with various pathological conditions such as inflammatory bowel disease (IBD). This study aimed to determine the anti-inflammatory colonic effects of blueberries and broccoli in mdr1a / mice (IBD mouse model) through modifi- cation of microbiota composition in the gastrointestinal tract. Methods: The mdr1a / mice were fed either a control diet or the control diet supplemented with either 10% blueberry or broccoli for 21 wk. We investigated the influence of these diets on cecal microbiota and organic acids, colon morphology, and bacterial translocation to mesenteric lymph nodes. Results: In comparison to mice fed the control diet, blueberry and broccoli supplementation altered cecum microbiota similarly with the exception of Faecalibacterium prausnitzii, which was found to be significantly lower in broccoli-fed mice. High concentrations of butyric acid and low concen- trations of succinic acid were observed in the cecum of broccoli-fed mice. Blueberry- and broccoli- supplemented diets increased colon crypt size and the number of goblet cells per crypt. Only the broccoli-supplemented diet significantly lowered colonic inflammation compared to mice fed the control diet. Translocation of total microbes to mesenteric lymph nodes was lower in broccoli-fed mice compared to blueberry and control diet groups. Conclusion: Dietary blueberries and/or broccoli altered the composition and metabolism of the cecal microbiota and colon morphology. Overall, these results warrant further investigation through clinical studies to establish whether the consumption of blueberries and/or broccoli is able to alter the composition and metabolism of large intestine microbiota and promote colon health in humans. Ó 2012 Elsevier Inc. All rights reserved. Introduction Inflammatory bowel disease (IBD) is characterized by chronic relapsing inflammation of the gastrointestinal (GI) tract and includes the closely related pathologies: ulcerative colitis (UC) and Crohn’s disease (CD). The host’s immune system, genotype, and enteric microbiota are all known factors involved in the pathogenesis of IBD. Even though the exact etiology of IBD remains unknown, one possible cause of the disease is thought to be an aberrant immune response to commensal microbiota of the GI tract. In healthy adults, the GI tract harbors a diverse microbial community consisting predominantly of the phyla Firmicutes and Bacteroidetes as well as smaller populations of Proteobacteria, Actinobacteria, Fusobacteria, and Verrucomicrobia [1]. The microbiota of healthy individuals are generally stable over time [2], whereas UC and CD patients exhibit temporal changes in commensal microbiota characterized by depletion in members of the phyla Firmicutes and Bacteroidetes [3]. There are no findings of a specific pathogen as a causative agent, but an association between pathogenic microorganisms such as Salmonella spp. and Campylobacter spp. in patients with a relapse * Corresponding author. Tel.: þ64 9 926 3515; fax: þ64 9 925 7001. E-mail address: [email protected] (G. Paturi). 0899-9007/$ - see front matter Ó 2012 Elsevier Inc. All rights reserved. doi:10.1016/j.nut.2011.07.018 Contents lists available at ScienceDirect Nutrition journal homepage: www.nutritionjrnl.com Nutrition 28 (2012) 324–330

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

lable at ScienceDirect
Nutrition 28 (2012) 324–330
Contents lists avai
Nutrition
journal homepage: www.nutr i t ionjrnl .com
Basic nutritional investigation
Influence of dietary blueberry and broccoli on cecal microbiota activity andcolon morphology in mdr1a�/� mice, a model of inflammatory bowel diseases
Gunaranjan Paturi Ph.D. a,*, Tafadzwa Mandimika Ph.D. b, Christine A. Butts Ph.D. b,Shuotun Zhu Ph.D. c, Nicole C. Roy Ph.D. d,e, Warren C. McNabb Ph.D. d,e, Juliet Ansell D.Phil. b,e
a The New Zealand Institute for Plant and Food Research Limited, Private Bag 92169, Auckland 1142, New Zealandb The New Zealand Institute for Plant and Food Research Limited, Private Bag 11600, Palmerston North 4442, New Zealandc School of Medical Sciences, The University of Auckland, Auckland 1142, New ZealanddAgResearch Grasslands, Palmerston North 4442, New ZealandeRiddet Institute, Massey University, Palmerston North 4442, New Zealand
a r t i c l e i n f o
Article history:Received 11 November 2010Accepted 29 July 2011
Keywords:Bacterial translocationColon inflammationFunctional foodsGut healthLarge intestineShort-chain fatty acids
* Corresponding author. Tel.: þ64 9 926 3515; fax:E-mail address: [email protected]
0899-9007/$ - see front matter � 2012 Elsevier Inc. Adoi:10.1016/j.nut.2011.07.018
a b s t r a c t
Objective: Enteric microbiota has been shown to be associated with various pathological conditionssuch as inflammatory bowel disease (IBD). This study aimed to determine the anti-inflammatorycolonic effects of blueberries and broccoli in mdr1a�/� mice (IBD mouse model) through modifi-cation of microbiota composition in the gastrointestinal tract.Methods: The mdr1a�/� mice were fed either a control diet or the control diet supplemented witheither 10% blueberry or broccoli for 21 wk. We investigated the influence of these diets on cecalmicrobiota and organic acids, colon morphology, and bacterial translocation to mesenteric lymphnodes.Results: In comparison to mice fed the control diet, blueberry and broccoli supplementation alteredcecum microbiota similarly with the exception of Faecalibacterium prausnitzii, which was found tobe significantly lower in broccoli-fed mice. High concentrations of butyric acid and low concen-trations of succinic acid were observed in the cecum of broccoli-fed mice. Blueberry- and broccoli-supplemented diets increased colon crypt size and the number of goblet cells per crypt. Only thebroccoli-supplemented diet significantly lowered colonic inflammation compared to mice fed thecontrol diet. Translocation of total microbes to mesenteric lymph nodes was lower in broccoli-fedmice compared to blueberry and control diet groups.Conclusion: Dietary blueberries and/or broccoli altered the composition and metabolism of thececal microbiota and colon morphology. Overall, these results warrant further investigationthrough clinical studies to establish whether the consumption of blueberries and/or broccoli is ableto alter the composition and metabolism of large intestine microbiota and promote colon health inhumans.
� 2012 Elsevier Inc. All rights reserved.
Introduction
Inflammatory bowel disease (IBD) is characterized by chronicrelapsing inflammation of the gastrointestinal (GI) tract andincludes the closely related pathologies: ulcerative colitis (UC)and Crohn’s disease (CD). The host’s immune system, genotype,and enteric microbiota are all known factors involved in thepathogenesis of IBD. Even though the exact etiology of IBDremains unknown, one possible cause of the disease is thought to
þ64 9 925 7001.o.nz (G. Paturi).
ll rights reserved.
be an aberrant immune response to commensal microbiota ofthe GI tract. In healthy adults, the GI tract harbors a diversemicrobial community consisting predominantly of the phylaFirmicutes and Bacteroidetes as well as smaller populations ofProteobacteria, Actinobacteria, Fusobacteria, and Verrucomicrobia[1]. The microbiota of healthy individuals are generally stableover time [2], whereas UC and CD patients exhibit temporalchanges in commensal microbiota characterized by depletion inmembers of the phyla Firmicutes and Bacteroidetes [3]. There areno findings of a specific pathogen as a causative agent, but anassociation between pathogenic microorganisms such asSalmonella spp. and Campylobacter spp. in patients with a relapse

Table 1Composition of experimental diets (g/kg)
Ingredients Control Blueberry Broccoli
Casein* 200 200 200Choline bitartratey 2 2 2DL-methioniney 3 3 3Ethoxyquiny 0.1 0.1 0.1Sucrosez 500 400 400Dextrinx 150 150 150Arbocel, non-nutritive bulk{ 50 50 50Corn oiljj 50 50 50Mineral mix** 35 35 35Vitamin mix** 10 10 10Blueberries d 100 d
Broccoli d d 100
G. Paturi et al. / Nutrition 28 (2012) 324–330 325
of IBD has been observed [4]. Differences in microbiota compo-sition between UC and CD have been observed [5]; however, theinterplay between changes in microbiota and the pathogenesisof these diseases remains unknown.
In the GI tract, commensalmicrobiota constantly interactwithintestinal epithelial cells, a process that is essential for thedevelopment of a healthy GI tract [6] and functioning of theimmune system [7]. Epithelial cells covering the GI tract act asa barrier by separating the enteric microbiota and host tissue andalso play a key role in providing the first line of defense againstpathogenic bacteria while maintaining GI homeostasis. A dys-functional intestinal barrier could lead to bacterial translocationfrom the lumen through themesenteric lymphnodes (MLNs) andvia systemic circulation to infiltrate the liver and spleen, poten-tially causing sepsis or organ failure [8]. However, the role ofintestinal barrier function in IBD is still under debate; whetherbarrier dysfunction is a result of intestinal inflammation ora primary causative factor of the disease remains unanswered.
Fruit and vegetables are a good source of phytochemicals suchas phenolic acids, flavonoids, minerals, and vitamins. A recentstudy reported the potential anti-inflammatory effect of poly-phenol metabolites derived from colon microbiota [9]. Blue-berries are a rich source of anthocyanins and polyphenols;fermentation by microbiota may affect their phenolic composi-tion and antioxidant capacity, thereby enhancing their anti-inflammatory potential. Osman et al. [10] observed the beneficialeffects of blueberries in an experimental colitis model resultingin significant improvement in colon health. Dietary poly-saccharides found in fruit and vegetables resist digestion in theupper GI tract and reach the large intestine relatively intact,where they act as a substrate for bacterial fermentation. Short-chain fatty acids (SCFAs) are the end products of this anaerobicbacterial fermentation, and butyric acid in particular has beenfound to be most effective in enhancing intestinal barrier func-tion [11]. In our previous study, we observed the modification ofmicrobiota composition by broccoli, which is a good source offermentable dietary fiber [12]. Examining the influence of blue-berries and broccoli on large intestinal health in an IBD modelmay demonstrate the potential benefits of fruit and vegetables inthe amelioration of intestinal inflammation and provide infor-mation about possible dietary alternatives for IBD patients.
In the current study, multidrug resistance gene deficient(mdr1a�/�) mice were used as an IBD model due to its sponta-neous development of colon inflammation despite the presenceof an intact immune system [13]. The lack of mdr1 pump (Abcb1transporters) in mdr1a�/� mice makes them unable to expelxenobiotics from inside epithelial cells into the intestinal lumen,resulting in damage to the intestinal barrier. The hypothesis ofthis study was that fruit and vegetable–based diets providefermentation substrates for the large intestine microbiota andthe resultant fermentation products can reduce inflammationand promote colon health. In the present study, we evaluated thepotential of diets supplementedwith either blueberry or broccolito modify microbiota composition and metabolic processes,resulting in reduction of colon inflammation and improvementof large intestine functionality in an IBD mouse model.
* Alacid, Lactic casein 30 mesh, New Zealand Milk Products Ltd., Wellington,New Zealand.
y Sigma-Aldrich, Sydney, Australia.z Caster sugar, Chelsea, New Zealand Sugar Company Ltd., Auckland, New
Zealand.x Wheaten cornstarch, Golden Harvest, Primary Foods Ltd., Auckland, New
Zealand.{ Arbocel B 600, J. Rettenmaier & Sohne GmbH þ Co., Rosenberg, Germany.jj Tradewinds, Davis Trading, Palmerston North, New Zealand.** Prepared in-house based on the AIN-76A diet formulation [33].
Materials and methods
Blueberry and broccoli samples
Blueberries (Vaccinium corymbosum L hybrid “Marimba,” “Misty,” and“O’Neal”) and green broccoli immature florets (Brassica oleracea L var italica)were obtained from growers in the Hawke’s Bay and Manawatu regions of NewZealand. The nutrient profile of blueberries and broccoli is available in New
Zealand FOOD files 2010 Manual [14]. Plant material was freeze dried, milledthrough a 1-mmmesh fitted to a grindingmill, and stored at 4�C until further use.The diets were prepared at the start of the study and were stored at �20�C untilrequired.
Mice and experimental design
Male mdr1a�/� mice (5–6 wk old) purchased from Taconic Farms, Inc.(Germantown, NY, USA) were housed in individual shoebox-style cages con-taining pressed wood chips with a plastic tube for environmental enrichment.The roomwas temperature controlled (22–24�C, humidity of 60 � 5%) with 12 hlight/dark cycle. All the experimental procedures for this study were approved bythe AgResearch Grasslands Animal Ethics Committee (Palmerston North, NewZealand) according to the Animal Welfare Act 1999.
After acclimatization for 1 wk, mice were randomly allocated into experi-mental groups (n ¼ 12 per group), control (AIN-76A diet), blueberry (10% blue-berry), and broccoli (10% broccoli) and were fed for 21 wk under conventionalhousing conditions. Mice were given ad libitum access to diet and waterthroughout the experiment. The components in the experimental diets areshown in Table 1. After 21 wk of feeding, mice were euthanized by CO2
asphyxiation. The cecum contents were collected and stored at �80�C, whichwere used for real-time polymerase chain reaction (PCR) quantification andorganic acids analysis. MLNs were collected and then stored at �80�C for real-time PCR quantification. Colon tissues were excised and fixed in 10% formalinfor histological analysis.
Microorganisms
All the culture media to grow bacterial strains were purchased from Oxoid(Adelaide, Australia). The following bacterial strains representative of each groupwere used to construct standard curves for real-time PCR quantification: Bac-teroides fragilis NZRM 964, Bacteroides vulgatus ATCC 8482, Enterococcus faecalisAGR 991, and Clostridium perfringens ATCC 13124 were grown in brain heartinfusion broth; Lactobacillus reuteri DPC 16 in de Man-Rogosa-Sharpe broth;Bifidobacterium adolescentis ATCC 15703 in de Man-Rogosa-Sharpe broth sup-plemented with 0.05% cysteine (Sigma-Aldrich, Sydney, Australia); Escherichiacoli ATCC 35150 in tryptone soy broth and Faecalibacterium prausnitzii DSM 17677in reinforced clostridial medium. All strains were grown anaerobically for 24 h at37�C in gas jars using a GasPak system (Oxoid) except E. coli, which was grownaerobically (37�C for 24 h). The bacterial cell count was estimated using a Neu-bauer hemocytometer.
Genomic DNA extraction
Bacterial genomic DNA from cecum contents was extracted by homogenizingwith 0.1 mm zirconia-silica beads (BioSpec Products Inc., Bartlesville, OK, USA) inASL buffer and then following the QIAamp DNA stool mini kit protocol (Qiagen,Melbourne, Australia). Bacterial genomic DNA from MLNs was extracted byhomogenizing the tissue in 180 mL of ATL buffer (Qiagen) and then following the
Table 2Primers used for real-time PCR quantification
Target bacteria Primer sequence (5’ / 3’) Annealingtemperature (�C)
Reference
Total bacteria F: TCCTACGGGAGGCAGCAGTR: GGACTACCAGGGTATCTAATCCTGTT
60 [34]
Bacteroides-Prevotella-Porphyromonas group F: GGTGTCGGCTTAAGTGCCATR: CGGA(C/T)GTAAGGGCCGTGC
63 [35]
Bifidobacterium spp. F: TCGCGTC(C/T)GGTGTGAAAGR: CCACATCCAGC(A/G)TCCAC
63 [35]
Clostridium perfringens group F: ATGCAAGTCGAGCGA(G/T)GR: TATGCGGTATTAATCT(C/T)CCTTT
55 [35]
Enterococcus spp. F: CCCTTATTGTTAGTTGCCATCATTR: ACTCGTTGTACTTCCCATTGT
64 [35]
Lactobacillus spp. F: CGATGAGTGCTAGGTGTTGGAR: CAAGATGTCAAGACCTGGTAAG
60 [36]
Bacteroides vulgatus F: GCATCATGAGTCCGCATGTTCR: TCCATACCCGACTTTATTCCTT
63 [37]
Escherichia coli F: CATGCCGCGTGTATGAAGAAR: CGGGTAACGTCAATGAGCAAA
60 [38]
Faecalibacterium prausnitzii F: GGAGGAAGAAGGTCTTCGGR: AATTCCGCCTACCTCTGCACT
60 [39]
Table 3Body weight and food intake of mice fed experimental diets
Diet Body weight (g) Food intake (g)*
Baseline Week 11 Week 21 Week 11 Week 21
Control 26.0 � 0.5 27.5 � 0.7 28.1 � 0.9 28.7 � 0.7 20.6 � 0.9Blueberry 25.4 � 0.6 27.3 � 0.9 28.6 � 1.1 30.2 � 0.8 20.2 � 1.2Broccoli 25.3 � 0.5 28.5 � 0.7 29.5 � 0.9 29.5 � 0.7 20.7 � 0.9P values 0.66 0.51 0.59 0.43 0.94
The data are mean of n ¼ 12 (control), n ¼ 8 (blueberry) and n ¼ 12 (broccoli).Data expressed as mean � standard error of mean
* Food intake was measured twice during the experiment at week 11 and 21.
G. Paturi et al. / Nutrition 28 (2012) 324–330326
protocol for isolation of genomic DNA from Gram-positive bacteria (QIAamp DNAmini kit; Qiagen). To construct standard curves, freshly grown bacterial strainswere resuspended in 180 mL of lysozyme (20 mg/mL in water) and incubated for18 h at 37�C. Subsequently, genomic DNA from bacterial isolates was extractedaccording to the protocol for isolation of genomic DNA from Gram-positivebacteria (QIAamp DNA mini kit).
Real-time PCR quantification
The primers used to construct standard curves from reference bacterialstrains and quantifying microbiota in cecum and MLNs are shown in Table 2. Asdescribed earlier [12], real-time PCR quantification was carried out usinga LightCycler 480 instrument (Roche Diagnostics, Mannheim, Germany) induplicates with a reaction volume of 20 mL, consisting of 10 mL of LightCycler 480SYBR Green I Master mix, 0.5 mM of each primer, and 5 mL of DNA template orwater. The melting curve analysis was performed from 60 to 95�C (0.1�C persecond) with continuous fluorescence acquisition. The standard curves weregenerated using a 10-fold dilution series (1 �109 colony forming units [CFU]/ml)of genomic DNA from each reference bacterial strain.
Organic acids
The SCFAs (acetic, butyric, formic, and propionic acids), lactic acid and suc-cinic acid concentrations in cecum contents of eachmousewere quantified by gaschromatography (GC) [15]. Briefly, cecum contents were homogenized in 0.01 Mphosphate-buffered saline and 2-ethylbutyric acid added as an internal standard(5 mM). This was followed by centrifugation (3000 � g for 5 min, 4�C), and thenthe supernatant was mixed with hydrochloric acid and diethyl ether. Afterfurther centrifugation (3000 � g for 10 min, 4�C), the upper diethyl ether phasewas collected and mixed with derivatizing agent, N-tert-butyldimethylsilyl-N-methyltrifluoroacetamide with 1% tert-butyldimethylchlorosilane (Sigma-Aldrich). The samples were left for 48 h at room temperature to allow completederivatization and were then analyzed using a Shimadzu gas chromatographsystem (GC-17A) fittedwith a HP-1 column (10m� 0.53 mm� 2.65 mm) (AgilentTechnologies, Santa Clara, CA, USA).
Assessment of colon inflammation
The colon inflammation in mice was evaluated according to the methoddescribed earlier [16], with few modifications. For each mouse an intact piece ofcolon tissue was fixed in 10% neutral buffered formalin, embedded in a paraffinblock, and sliced into sections of 5 mm thickness. The sections were stained withhematoxylin and eosin for light microscopic examination of inflammation. Colontissue sections were scored (0 ¼ no injury to 10 ¼ maximum injury) for 12inflammation parameters, which were categorized into inflammatory cell infil-tration (lymphocytes/plasma cells, monocytes/macrophages, and neutrophils)and tissue architecture damage (aberrant crypts, crypt abscess, crypt hyperplasia,crypt injury, crypt loss, loss of goblet cells, lymphoid aggregates, submucosalthickening, and surface loss). The histological injury score (HIS) was calculatedas: HIS ¼ ([inflammatory cell infiltration] � 3) þ tissue architecture damage. Thesum of inflammatory cell infiltration was multiplied by 3 to give more weight tothis value, which represents the major feature of inflammation observed.
Analysis of colon crypt depth and goblet cells
Formalin-fixed colon tissue from each mouse was embedded in a paraffinblock and processed to generate 5-mm-thick transverse sections. For microscopicexamination, sections were stained with hematoxylin and eosin, and alcian blue.As described previously by Paturi et al. [12], the crypt depths in the colon weremeasured using Image-Pro Plus analysis software (4.1 version) at �20 magnifi-cation in a bright field microscope (Model Axiophot; Carl Zeiss Inc., Gottingen,Germany). Intact colon crypts were used for 30 random measurements. Thenumber of goblet cells per crypt was counted in 30 crypts using the same imageof the histological section.
Statistical analysis
The data were analyzed using one-way analysis of variance followed by post-hoc analysis by least significant difference test. The body weight gain for eachmouse was analyzed at week 11 and 21. A value of P < 0.05 was consideredstatistically significant. All analyses were carried out using GenStat 11th edition(VSN International, Hemel Hempstead, UK).
Results
Body weight and food intake
The mean body weights and food intakes were not signifi-cantly different between mice fed the experimental diets(Table 3). The body weight gain did not differ significantlybetween the experimental diets at week 11 (P ¼ 0.14) or 21 (P ¼0.17) (data not shown).
Quantification of cecum microbiota
There was no significant effect of experimental diets on totalbacteria (P ¼ 0.14), Bacteroides-Prevotella-Porphyromonas group
Control Blueberry BroccoliB
acteroid
es-P
revotella-P
orphyrom
onas
grou
p (L
og10
CFU
/g)
0
2
4
6
8
10
12
Control Blueberry Broccoli
Bifid
obacteriu
m s
pp. (
Log 1
0 C
FU/g
)
0
2
4
6
8
10
12
Control Blueberry Broccoli
C. perfrin
gens g
roup
(Log
10 C
FU/g
)
0
2
4
6
8
10
12
Control Blueberry Broccoli
Enterococcus s
pp. (L
og10
CFU
/g)
0
2
4
6
8
10
12
Control Blueberry BroccoliLactobacillu
s s
pp. (
Log 1
0 C
FU/g
)
0
2
4
6
8
10
12
Control Blueberry Broccoli
Tota
l bac
teria
(Log
10 C
FU/g
)
0
2
4
6
8
10
12 a a a a a a
a b b a b b a b b
a a a
Control Blueberry Broccoli
F. prausnitzii (L
og10
CFU
/g)
0
2
4
6
8
10
12 a a b
Control Blueberry Broccoli
B. vulg
atus (L
og10
CFU
/g)
0
2
4
6
8
10
12
Control Blueberry Broccoli
E. coli (L
og10
CFU
/g)
0
2
4
6
8
10
12 a b ba a a
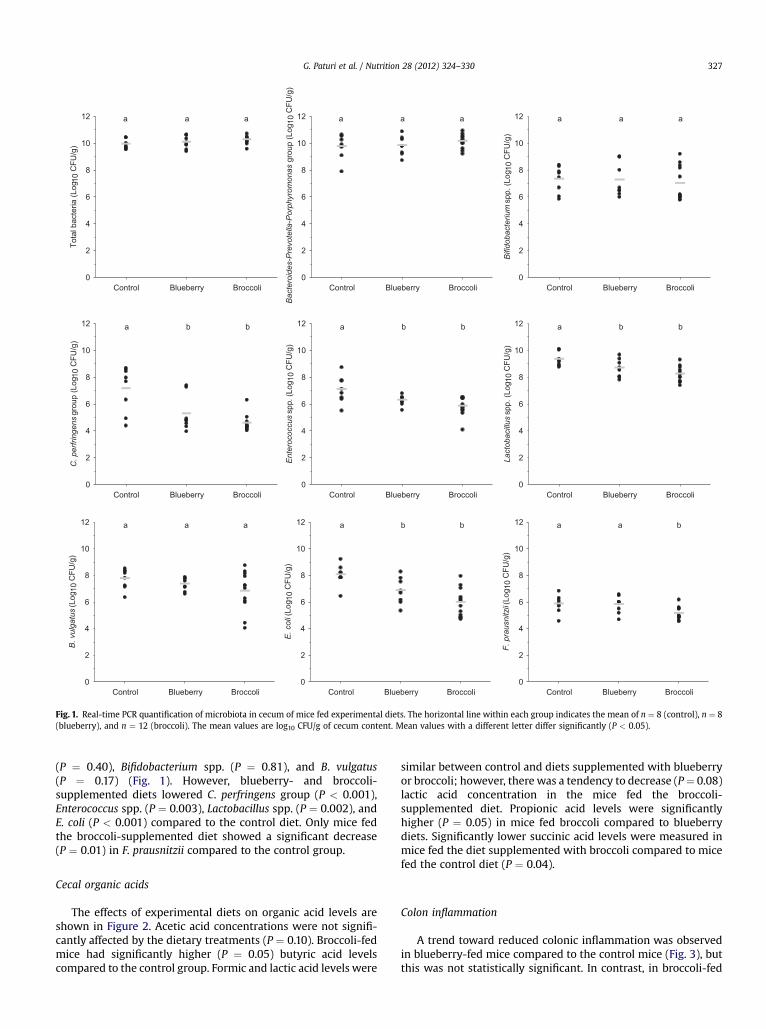
Fig. 1. Real-time PCR quantification of microbiota in cecum of mice fed experimental diets. The horizontal line within each group indicates the mean of n ¼ 8 (control), n ¼ 8(blueberry), and n ¼ 12 (broccoli). The mean values are log10 CFU/g of cecum content. Mean values with a different letter differ significantly (P < 0.05).
G. Paturi et al. / Nutrition 28 (2012) 324–330 327
(P ¼ 0.40), Bifidobacterium spp. (P ¼ 0.81), and B. vulgatus(P ¼ 0.17) (Fig. 1). However, blueberry- and broccoli-supplemented diets lowered C. perfringens group (P < 0.001),Enterococcus spp. (P ¼ 0.003), Lactobacillus spp. (P ¼ 0.002), andE. coli (P < 0.001) compared to the control diet. Only mice fedthe broccoli-supplemented diet showed a significant decrease(P ¼ 0.01) in F. prausnitzii compared to the control group.
Cecal organic acids
The effects of experimental diets on organic acid levels areshown in Figure 2. Acetic acid concentrations were not signifi-cantly affected by the dietary treatments (P ¼ 0.10). Broccoli-fedmice had significantly higher (P ¼ 0.05) butyric acid levelscompared to the control group. Formic and lactic acid levels were
similar between control and diets supplemented with blueberryor broccoli; however, there was a tendency to decrease (P¼ 0.08)lactic acid concentration in the mice fed the broccoli-supplemented diet. Propionic acid levels were significantlyhigher (P ¼ 0.05) in mice fed broccoli compared to blueberrydiets. Significantly lower succinic acid levels were measured inmice fed the diet supplemented with broccoli compared to micefed the control diet (P ¼ 0.04).
Colon inflammation
A trend toward reduced colonic inflammation was observedin blueberry-fed mice compared to the control mice (Fig. 3), butthis was not statistically significant. In contrast, in broccoli-fed

Acetic Butyric Formic Lactic Propionic Succinic
Org
anic
aci
ds (µ
mol
/g)
0
20
40
60
80Control Blueberry Broccoli
a abb
a
a aa
a
a
a a
a
abb
a
a
ab
b
Fig. 2. Organic acids in the cecum of mice fed experimental diets. The data aremean of n ¼ 10 (control), n ¼ 8 (blueberry), and n ¼ 12 (broccoli). The mean valuesare mmol/g of cecum content. Error bars represent standard error of the mean. Meanvalues with a different letter differ significantly (P < 0.05).
G. Paturi et al. / Nutrition 28 (2012) 324–330328
mice, a significant reduction (P ¼ 0.05) in colonic inflammationwas observed compared to those fed the control diet.
Colon crypt depth and goblet cells
As shown in Figure 4, diets supplemented with either blue-berry or broccoli affected the colon morphology, as evidenced byelongated crypts (P ¼ 0.002) and also an increased number ofgoblet cells per crypt (P ¼ 0.005).
Bacterial translocation
The translocation of total bacteria to MLNs was similarbetween mice fed the blueberry-supplemented and control diets
Control Blueberry Broccoli
His
tolo
gica
l inj
ury
scor
e
0
10
20
30
40
50
60
70
b
ab
a
Fig. 3. Histological injury score in colon of mice fed experimental diets. The dataare mean of n ¼ 12 (control), n ¼ 8 (blueberry), and n ¼ 12 (broccoli). Error barsrepresent standard error of the mean. Mean values with a different letter differsignificantly (P < 0.05).
(Fig. 5). However, mice fed the broccoli-supplemented diet hadsignificantly less bacteria present in their MLNs (P < 0.001).
Discussion
In the current study, diets supplemented with either blue-berry or broccoli altered the cecum microbiota, particularlyspecific bacteria that are known to be associatedwith IBD (Fig.1).Certain opportunistic pathogenic bacteria are able to prevail inhigher numbers when there is an unstable environment in the GItract as a result of disease. For example, clones resemblingC. perfringenswere detected in the ileal pouch of UC patients [17].E. faecalis, a commensal bacterium of humans and animals, hasshown the capability to induce IBD in IL-10 gene-deficient mice[18], suggesting that certain enteric microbiota tend to be moreopportunistic and can induce colon inflammation. In the presentstudy, a significant reduction in C. perfringens group andEnterococcus spp. was observed in mice fed diets supplementedwith either blueberry or broccoli (Fig. 1). However, the associa-tion between these bacteria and a reduction in intestinalinflammation needs further investigation. Additionally, adherentinvasive E. coli have been implicated with IBD, which is known tocolonize the mucosa through adhering to intestinal epithelialcells. Kotlowski et al. [19] reported a 3 to 4 log increase of E. coli inthe intestinal tissues of patients with IBD. In the present study,blueberry- and broccoli-supplemented diets inhibited thegrowth of E. coli. In particular, broccoli-fed mice showed a 2 logdecrease in the cecal E. coli population compared to the controldiet (Fig. 1). The influence of these diets (blueberry and broccoli)in lowering E. coli may be beneficial to the host. In an earlierstudy using control diets only, we observed an increase in E. coliinmdr1a�/�mice at 25 wk of age comparedwith prior to onset ofinflammation at 12 wk of age [20].
Microbes belonging to Bifidobacterium spp. and Lactobacillusspp. are generally regarded as safe, and their increase in the GItract is considered beneficial to the host. Blueberry- and broccoli-supplemented diets showed no effect on Bifidobacterium spp,whereas both blueberry and broccoli diets significantly reducedLactobacillus spp. (Fig. 1). Lactic acid is one of the fermentationend products of several commensal bacteria in the GI tractincluding bifidobacteria and lactobacilli. In a densely colonizedecosystem, metabolic cross-feeding between bacteria is vital insubstrate breakdown. For example, bacterial strains isolatedfrom human feces, Anaerostipes caccae and Eubacterium hallii areable to convert acetic and lactic acids into butyric acid [21]. Highfecal lactic acid concentrations in patients with severe UC havebeen observed [22]. In this study, the broccoli-supplemented dietdecreased the Lactobacillus spp, and corresponding low lacticacid levels were observed (Fig. 2). The decrease in lactic acidconcentration can be considered beneficial to the host duringintestinal inflammation due to the role of lactic acid as a favoredcosubstrate for sulfate-reducing bacteria, which releasehydrogen sulfide known to be toxic for intestinal epithelial cells[23]. An excess of hydrogen sulfide or an impaired detoxificationmechanism can damage the colon mucosa and in fact highconcentrations of hydrogen sulfide have been implicated in theetiology of UC [24].
We focused on organic acids analysis due to the impact ofthese bacterial metabolites on host physiological functions. Inparticular, a recent study demonstrated the anti-inflammatoryproperties of acetic and propionic acids in addition to butyricacid [25]. A significant decrease in histological injury scoreobserved in the colon ofmice fed the broccoli-supplemented dietcould be a consequence of the higher overall concentrations of

Control Blueberry Broccoli
Cry
pt d
epth
(µm
)
0
50
100
150
200
250
300A B
bb
a
Control Blueberry Broccoli
Gob
let c
ells
/cry
pt
0
5
10
15
20
25
a
bb
Fig. 4. Mice fed experimental diets showing variation in (A) colon crypt depth and (B) goblet cells/crypt. The data are mean of n ¼ 11 (control), n ¼ 7 (blueberry), and n ¼ 12(broccoli). Error bars represent standard error of the mean. Mean values with a different letter differ significantly (P < 0.05).
G. Paturi et al. / Nutrition 28 (2012) 324–330 329
acetic, butyric, and propionic acids in the cecum (Figs. 2 and 3).The larger crypts observed in mice fed diets supplemented witheither blueberry or broccoli could be a result of lower succinicacid levels (Figs. 2 and 4A), due to the inhibitory effect of succinicacid on colonic epithelial cell proliferation and reducing cryptsize [26]. Dysplastic or loss of crypts in colon is an indicativemeasure of inflammatory colonic damage and altered colonfunctionality; the larger crypts observed in mice fed blueberry-and broccoli-supplemented diets suggest that broccoli andblueberry have anti-inflammatory effects, which result inimprovement/maintenance of colonic health [27,28].
It is well known that butyric acid is considered an importantsource of energy to colonocytes. Mice fed the broccoli-supplemented diet showed a significant decrease in F. prausnit-zii, a known butyric acid–producing bacteria (Fig. 1). Therefore,the elevated butyric acid levels observed in broccoli-fed mice(Fig. 2) must have been due to an increase in other butyric-acid-producing bacteria or a result of metabolic cross-feedingbetween bacteria. IBD is characterized by increased intestinal
Control Blueberry Broccoli
Tota
l bac
teria
(Log
10 C
FU/g
)
0
1
29
10a
a
b
Fig. 5. Real-time PCR quantification of total bacteria in mesenteric lymph nodes ofmice fed experimental diets. The data are mean of n ¼ 8 (control), n ¼ 8 (blueberry),and n ¼ 12 (broccoli). Error bars represent standard error of the mean. Mean valueswith a different letter differ significantly (P < 0.05).
permeability, where luminal bacteria breach the intestinalbarrier and enter the blood circulation, causing systemicinflammation; butyric acid has been shown to significantlyreduce bacterial internalization and translocation across T84 cellmonolayers [29]. In this study, we also observed an increase incecal butyric acid concentration accompanied by less bacterialtranslocation to MLNs in mice fed broccoli-supplemented diet(Figs. 2 and 5). Broccoli-fed mice may have resulted in improvedmaintenance of GI epithelial proliferation and inhibition ofpathogenic bacterial growth along with a subsequent reductionin bacterial translocation [10].
As part of GI homeostasis, the mucus layer covering the GItract acts as a protective barrier against a harsh luminal envi-ronment. Intestinal diseases can alter the mucus layer; forexample, in UC patients, thin and discontinuous mucus layerswith a low goblet cell density were found [30]. A defectivemucosal barrier function resulting from abnormal synthesis and/or processing of mucins is a characteristic of IBD [31]. Dey et al.[32] observed extensive loss of goblet cells and massive inflam-matory cell aggregates in the colon of an ulcerative colitis mousemodel, whereas treatment by dietary phenethylisothiocyanateproduced by Brassica food plants reduced the mucosal inflam-mation and depletion of goblet cells. In the current study, blue-berry- and broccoli-supplemented diets contributed towarda significant increase in goblet cells per crypt (Fig. 4B), which canbe considered beneficial due to the ability of these cells to secretemucin.
Conclusion
Currently, there is insufficient literature to propose an asso-ciation between diet and IBD; polyphenols and dietary poly-saccharides in fruit and vegetables can be effective in promotingGI health benefits. Dietary blueberries and/or broccoli alteredcecal microbiota composition and metabolism, and colonmorphology. Future studies are necessary to unravel the under-lying mechanisms by which broccoli, and to a lesser extentblueberries, were beneficial to intestinal inflammation.
Acknowledgments
The authors acknowledge the assistance of Sheridan Martell,Hannah Smith, Halina Stoklosinski, and Andrew McLachlan(Plant and Food Research). E. faecalis and F. prausnitzii culture

G. Paturi et al. / Nutrition 28 (2012) 324–330330
stocks were received as a gift from AgResearch culture collection.This study was supported by a Plant and Food Research GutActivity Strategic Science Investment, and Nutrigenomics NewZealand, a collaboration between AgResearch, Plant and FoodResearch, and The University of Auckland, funded by the NewZealand Foundation for Research, Science, and Technology.
References
[1] Eckburg PB, Bik EM, Bernstein CN, Purdom E, Dethlefsen L, Sargent M, et al.Diversity of the human intestinal microbial flora. Science 2005;308:1635–8.
[2] Zoetendal EG, Akkermans AD, De Vos WM. Temperature gradient gelelectrophoresis analysis of 16S rRNA from human fecal samples revealsstable and host-specific communities of active bacteria. Appl EnvironMicrobiol 1998;64:3854–9.
[3] Frank DN, St Amand AL, Feldman RA, Boedeker EC, Harpaz N, Pace NR.Molecular-phylogenetic characterization of microbial community imbal-ances in human inflammatory bowel diseases. Proc Natl Acad Sci USA2007;104:13780–5.
[4] MylonakiM, Langmead L, Pantes A, Johnson F, Rampton DS. Enteric infectionin relapse of inflammatory bowel disease: importance of microbiologicalexamination of stool. Eur J Gastroenterol Hepatol 2004;16:775–8.
[5] Swidsinski A, Loening-Baucke V, Vaneechoutte M, Doerffel Y. ActiveCrohn’s disease and ulcerative colitis can be specifically diagnosed andmonitored based on the biostructure of the fecal flora. Inflamm Bowel Dis2008;14:147–61.
[6] Ley RE, Peterson DA, Gordon JI. Ecological and evolutionary forces shapingmicrobial diversity in the human intestine. Cell 2006;124:837–48.
[7] Artis D. Epithelial-cell recognition of commensal bacteria and maintenanceof immune homeostasis in the gut. Nat Rev Immunol 2008;8:411–20.
[8] Turner JR. Molecular basis of epithelial barrier regulation: from basicmechanisms to clinical application. Am J Pathol 2006;169:1901–9.
[9] Larrosa M, Luceri C, Vivoli E, Pagliuca C, Lodovici M, Moneti G, et al. Poly-phenolmetabolites fromcolonicmicrobiota exert anti-inflammatoryactivityon different inflammation models. Mol Nutr Food Res 2009;53:1044–54.
[10] Osman N, Adawi D, Ahrne S, Jeppsson B, Molin G. Probiotics and blueberryattenuate the severity of dextran sulfate sodium (DSS)-induced colitis. DigDis Sci 2008;53:2464–73.
[11] Peng L, Li Z-R, Green RS, Holzman IR, Lin J. Butyrate enhances the intestinalbarrier by facilitating tight junction assembly via activation of AMP-activatedprotein kinase in Caco-2 cell monolayers. J Nutr 2009;139:1619–25.
[12] Paturi G, Butts C, Monro J, Nones K, Martell S, Butler R, et al. Cecal andcolonic responses in rats fed 5 or 30% corn oil diets containing either 7.5%broccoli dietary fiber or microcrystalline cellulose. J Agric Food Chem2010;58:6510–5.
[13] Panwala CM, Jones JC, Viney JL. A novel model of inflammatory boweldisease: mice deficient for the multiple drug resistance gene, mdr1a,spontaneously develop colitis. J Immunol 1998;161:5733–44.
[14] Huffman L, Sivakumaran S, Sharp K. New Zealand FOODfiles 2010 Manual.Palmerston North, New Zealand: The New Zealand Institute for Plant andFood Research Limited; 2010.
[15] Richardson AJ, Calder AG, Stewart CS, Smith A. Simultaneous determinationof volatile and non-volatile acidic fermentation products of anaerobes bycapillary gas chromatography. Lett Appl Microbiol 1989;9:5–8.
[16] Nones K, Dommels YEM, Martell S, Butts C, McNabb WC, Park ZA, et al. Theeffects of dietary curcumin and rutin on colonic inflammation and geneexpression in multidrug resistance gene-deficient (mdr1a(�/�)) mice,a model of inflammatory bowel diseases. Br J Nutr 2009;101:169–81.
[17] Falk A, Olsson C, Ahrne S, Molin G, Adawi D, Jeppsson B. Ileal pelvic pouchmicrobiota from two former ulcerative colitis patients, analysed by DNA-based methods, were unstable over time and showed the presence ofClostridium perfringens. Scand J Gastroenterol 2007;42:973–85.
[18] Balish E, Warner T. Enterococcus faecalis induces inflammatory boweldisease in interleukin-10 knockout mice. Am J Pathol 2002;160:2253–7.
[19] Kotlowski R, Bernstein CN, Sepehri S, Krause DO. High prevalence ofEscherichia coli belonging to the B2þD phylogenetic group in inflamma-tory bowel disease. Gut 2007;56:669–75.
[20] Nones K, Knoch B, Dommels YEM, Paturi G, Butts C, McNabb WC, et al.Multidrug resistance gene deficient (mdr1a�/�) mice have an alteredcaecal microbiota that precedes the onset of intestinal inflammation. J ApplMicrobiol 2009;107:557–66.
[21] Duncan SH, Louis P, Flint HJ. Lactate-utilizing bacteria, isolated from humanfeces, that produce butyrate as a major fermentation product. Appl EnvironMicrobiol 2004;70:5810–7.
[22] Vernia P, Caprilli R, Latella G, Barbetti F, Magliocca FM, Cittadini M. Fecallactate and ulcerative colitis. Gastroenterology 1988;95:1564–8.
[23] Marquet P, Duncan SH, Chassard C, Bernalier-Donadille A, Flint HJ. Lactatehas the potential to promote hydrogen sulphide formation in the humancolon. FEMS Microbiol Lett 2009;299:128–34.
[24] Rowan FE, Docherty NG, Coffey JC, O’Connell PR. Sulphate-reducingbacteria and hydrogen sulphide in the aetiology of ulcerative colitis. Br JSurg 2009;96:151–8.
[25] Tedelind S, Westberg F, Kjerrulf M, Vidal A. Anti-inflammatory propertiesof the short-chain fatty acids acetate and propionate: A study with rele-vance to inflammatory bowel disease. World J Gastroenterol 2007;13:2826–32.
[26] Inagaki A, Ichikawa H, Sakata T. Inhibitory effect of succinic acid onepithelial cell proliferation of colonic mucosa in rats. J Nutr Sci Vitaminol2007;53:377–9.
[27] Yasui Y, Hosokawa M, Mikami N, Miyashita K, Tanaka T. Dietary astaxanthininhibits colitis and colitis-associated colon carcinogenesis in mice viamodulation of the inflammatory cytokines. Chem Biol Interact 2011;193:79–87.
[28] Andujar I, Recio MC, Giner RM, Cienfuegos-Jovellanos E, Laghi S,Muguerza B, et al. Inhibition of ulcerative colitis in mice after oral admin-istration of a polyphenol-enriched cocoa extract is mediated by the inhibi-tion of STAT1 and STAT3 phosphorylation in colon cells. J Agric Food Chem2011;59:6474–83.
[29] Lewis K, Lutgendorff F, Phan V, Soderholm JD, Sherman PM, McKay DM.Enhanced translocation of bacteria across metabolically stressed epitheliais reduced by butyrate. Inflamm Bowel Dis 2010;16:1138–48.
[30] Strugala V, Dettmar PW, Pearson JP. Thickness and continuity of theadherent colonic mucus barrier in active and quiescent ulcerative colitisand Crohn’s disease. Int J Clin Pract 2008;62:762–9.
[31] Kim Y, Ho S. Intestinal goblet cells and mucins in health and disease: recentinsights and progress. Curr Gastroenterol Rep 2010;12:319–30.
[32] Dey M, Kuhn P, Ribnicky D, Premkumar V, Reuhl K, Raskin I. Dietary phe-nethylisothiocyanate attenuates bowel inflammation in mice. BMC ChemBiol 2010;10:4.
[33] Reeves PG, Nielsen FH, Fahey GC. AIN-93 purified diets for laboratoryrodents: final report of the American Institute of Nutrition ad hoc writingcommittee on the reformulation of the AIN-76A rodent diet. J Nutr1993;123:1939–51.
[34] Nadkarni MA, Martin FE, Jacques NA, Hunter N. Determination of bacterialload by real-time PCR using a broad-range (universal) probe and primersset. Microbiology 2002;148:257–66.
[35] Rinttila T, Kassinen A, Malinen E, Krogius L, Palva A. Development of anextensive set of 16S rDNA-targeted primers for quantification of pathogenicand indigenous bacteria in faecal samples by real-time PCR. J ApplMicrobiol 2004;97:1166–77.
[36] Fu CJ, Carter JN, Li Y, Porter JH, Kerley MS. Comparison of agar plate andreal-time PCR on enumeration of Lactobacillus, Clostridium perfringensand total anaerobic bacteria in dog faeces. Lett Appl Microbiol 2006;42:490–4.
[37] Wang R-F, Cao W-W, Cerniglia CE. PCR detection and quantification ofpredominant anaerobic bacteria in human and animal fecal samples. ApplEnviron Microbiol 1996;62:1242–7.
[38] Huijsdens XW, Linskens RK, Mak M, Meuwissen SGM, Vandenbroucke-Grauls CMJE, Savelkoul PHM. Quantification of bacteria adherent togastrointestinal mucosa by real-time PCR. J Clin Microbiol 2002;40:4423–7.
[39] Ramirez-Farias C, Slezak K, Fuller Z, Duncan A, Holtrop G, Louis P. Effect ofinulin on the human gut microbiota: stimulation of Bifidobacterium ado-lescentis and Faecalibacterium prausnitzii. Br J Nutr 2009;101:541–50.
Related Documents











