Induction of apoptosis by streptochlorin isolated from Streptomyces sp. in human leukemic U937 cells Cheol Park a , Hee Jae Shin b , Gi-Young Kim c , Taeg Kyu Kwon d , Taek-Jeong Nam e , Se-Kwon Kim f , Jaehun Cheong g , Il-Whan Choi h, * , Yung Hyun Choi a, * a Department of Biochemistry, Dongeui University College of Oriental Medicine, Department of Biomaterial Control (BK21 Program), Dongeui University Graduate School, Yangjung-dong San 45, Busanjin-gu, Busan 614-052, South Korea b Marine Natural Product Chemistry Laboratory, Ocean Research and Development Institute, Ansan 425-600, South Korea c Faculty of Applied Marine Science, Cheju National University, Jeju 690-756, South Korea d Department of Immunology, School of Medicine, Keimyung University, Taegu 700-712, South Korea e Department of Food and Life Science, Pukyong National University, Busan 608-737, South Korea f Department of Chemistry and Marine Bioprocess Research Center, Pukyong National University, Busan 608-737, South Korea g Department of Molecular Biology, Pusan National University, Busan 609-735, South Korea h Department of Microbiology, College of Medicine and Center for Viral Disease Research, Inje University, Busan 614-735, South Korea article info Article history: Received 24 March 2008 Accepted 23 June 2008 Available online 1 July 2008 Keywords: Streptochlorin U937 Apoptosis Bcl-2 Caspase-3 . abstract Streptochlorin is a small molecule that produced by marine Streptomyces sp. that is known to have anti- angiogenic and anti-cancer properties. However, the mechanism by which streptochlorin functions is not well understood. In this study, we investigated the pro-apoptotic effect of streptochlorin in human leu- kemic U937 cells. Streptochlorin treatment resulted in concentration- and time-dependent growth inhi- bition by inducing apoptosis. The increase in apoptosis that was induced by streptochlorin was correlated with down-regulation of anti-apoptotic Bcl-2 expression, up-regulation of pro-apoptotic Bax and FasL, a decrease in the mitochondrial membrane potential (MMP), activation of caspases and degradation of poly-(ADP-ribose)polymerase and phospholipase C-c1 protein. In addition, the cytotoxic effects and apoptotic characteristics induced by streptochlorin were significantly inhibited by z-DEVD-fmk, a cas- pase-3 inhibitor, which demonstrates the important role that caspase-3 played in the process. Further- more, Bcl-2 overexpression significantly reversed the streptochlorin-induced growth inhibitory effects via inhibition of the MMP collapse and caspases activation and effectively attenuated the apoptotic response to streptochlorin. However, the elevated levels of FasL expression induced by streptochlorin were not reduced by Bcl-2 overexpression. Taken together, these findings demonstrate that the pro-apop- totic effect of streptochlorin is mediated through activation of caspases and mitochondria in U937 cells. Ó 2008 Elsevier Ltd. All rights reserved. 1. Introduction Apoptosis is the active process of programmed cell death that occurs during many important physiological conditions, such as embryonic development and tissue remodeling. However, most cancer cells block apoptosis, which allows them to survive despite undergoing genetic and morphologic transformations. Apoptotic cells are characterized by several unique features, including cell shrinkage, chromatin condensation, DNA fragmentation, cell sur- face expression of phosphatidylserine, and membrane blebbing (Okada and Mak, 2004; Jin and el-Deiry, 2005; Han et al., 2008). A central component of the apoptotic machinery is a family of cy- stein-containing aspartate-specific proteases, termed caspase. Caspases are present in cells as inactive proenzymes, with the ac- tive tetramer being formed as a result of removal of the prodomain and cleavage between the large and small subunits (Hengartner, 2000; Suda et al., 1993). Caspase activity is either directly or indi- rectly responsible for the cleavage of several intracellular proteins that are characteristically proteolysed during apoptosis. It has been reported that two major pathways, the Fas/tumor necrosis factor (TNF) death receptor (extrinsic) pathway and the mitochondria- dependent (intrinsic) pathway, lead to activation of caspases and 0887-2333/$ - see front matter Ó 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.tiv.2008.06.010 Abbreviations: DAPI, 4,6-diamidino-2-phenylindile; DEVD, Asp-Glu-Val-Asp; DMSO, dimethyl sulfoxide; ECL, enhanced chemiluminescence; EtBr, ethidium bromide; FasL, Fas ligand; FBS, fetal bovine serum; IETD, Ile-Glu-Thr-Asp; JC-1, 5,5 0 , 6,6 0 -tetrachloro-1,1 0 ,3,3 0 -tetraethyl-imidacarbocyanine iodide; LEHD, Leu-Glu-His- Asp; MMP, mitochondrial membrane potential; PARP, poly-(ADP-ribose)polymerase; PBS, phosphate-buffered saline; PCR, polymerase chain reaction; PI, propidium iodide; PLC, phospholipase C; pNA, p-nitroaniline; TNF, tumor necrosis factor. * Corresponding authors. Tel.: +82 51 850 7413; fax: +82 51 853 4036 (Y.H. Choi), tel.: +82 51 890 6461; fax: +82 51 891 6004 (I.W. Choi). E-mail addresses: [email protected] (I.-W. Choi), [email protected] (Y.H. Choi). Toxicology in Vitro 22 (2008) 1573–1581 Contents lists available at ScienceDirect Toxicology in Vitro journal homepage: www.elsevier.com/locate/toxinvit

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Toxicology in Vitro 22 (2008) 1573–1581
Contents lists available at ScienceDirect
Toxicology in Vitro
journal homepage: www.elsevier .com/locate / toxinvi t
Induction of apoptosis by streptochlorin isolated from Streptomyces sp.in human leukemic U937 cells
Cheol Park a, Hee Jae Shin b, Gi-Young Kim c, Taeg Kyu Kwon d, Taek-Jeong Nam e, Se-Kwon Kim f,Jaehun Cheong g, Il-Whan Choi h,*, Yung Hyun Choi a,*
a Department of Biochemistry, Dongeui University College of Oriental Medicine, Department of Biomaterial Control (BK21 Program), Dongeui University Graduate School,Yangjung-dong San 45, Busanjin-gu, Busan 614-052, South Koreab Marine Natural Product Chemistry Laboratory, Ocean Research and Development Institute, Ansan 425-600, South Koreac Faculty of Applied Marine Science, Cheju National University, Jeju 690-756, South Koread Department of Immunology, School of Medicine, Keimyung University, Taegu 700-712, South Koreae Department of Food and Life Science, Pukyong National University, Busan 608-737, South Koreaf Department of Chemistry and Marine Bioprocess Research Center, Pukyong National University, Busan 608-737, South Koreag Department of Molecular Biology, Pusan National University, Busan 609-735, South Koreah Department of Microbiology, College of Medicine and Center for Viral Disease Research, Inje University, Busan 614-735, South Korea
a r t i c l e i n f o
Article history:Received 24 March 2008Accepted 23 June 2008Available online 1 July 2008
Keywords:StreptochlorinU937ApoptosisBcl-2Caspase-3.
0887-2333/$ - see front matter � 2008 Elsevier Ltd. Adoi:10.1016/j.tiv.2008.06.010
Abbreviations: DAPI, 4,6-diamidino-2-phenylindiDMSO, dimethyl sulfoxide; ECL, enhanced chemilubromide; FasL, Fas ligand; FBS, fetal bovine serum; IET6,60-tetrachloro-1,10 ,3,30-tetraethyl-imidacarbocyanineAsp; MMP, mitochondrial membrane potential; PARP, pPBS, phosphate-buffered saline; PCR, polymerase chiodide; PLC, phospholipase C; pNA, p-nitroaniline; TNF
* Corresponding authors. Tel.: +82 51 850 7413; faxtel.: +82 51 890 6461; fax: +82 51 891 6004 (I.W. Ch
E-mail addresses: [email protected] (I.-W. Choi), ch
a b s t r a c t
Streptochlorin is a small molecule that produced by marine Streptomyces sp. that is known to have anti-angiogenic and anti-cancer properties. However, the mechanism by which streptochlorin functions is notwell understood. In this study, we investigated the pro-apoptotic effect of streptochlorin in human leu-kemic U937 cells. Streptochlorin treatment resulted in concentration- and time-dependent growth inhi-bition by inducing apoptosis. The increase in apoptosis that was induced by streptochlorin was correlatedwith down-regulation of anti-apoptotic Bcl-2 expression, up-regulation of pro-apoptotic Bax and FasL, adecrease in the mitochondrial membrane potential (MMP), activation of caspases and degradation ofpoly-(ADP-ribose)polymerase and phospholipase C-c1 protein. In addition, the cytotoxic effects andapoptotic characteristics induced by streptochlorin were significantly inhibited by z-DEVD-fmk, a cas-pase-3 inhibitor, which demonstrates the important role that caspase-3 played in the process. Further-more, Bcl-2 overexpression significantly reversed the streptochlorin-induced growth inhibitory effectsvia inhibition of the MMP collapse and caspases activation and effectively attenuated the apoptoticresponse to streptochlorin. However, the elevated levels of FasL expression induced by streptochlorinwere not reduced by Bcl-2 overexpression. Taken together, these findings demonstrate that the pro-apop-totic effect of streptochlorin is mediated through activation of caspases and mitochondria in U937 cells.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
Apoptosis is the active process of programmed cell death thatoccurs during many important physiological conditions, such asembryonic development and tissue remodeling. However, mostcancer cells block apoptosis, which allows them to survive despite
ll rights reserved.
le; DEVD, Asp-Glu-Val-Asp;minescence; EtBr, ethidiumD, Ile-Glu-Thr-Asp; JC-1, 5,50 ,iodide; LEHD, Leu-Glu-His-
oly-(ADP-ribose)polymerase;ain reaction; PI, propidium, tumor necrosis factor.
: +82 51 853 4036 (Y.H. Choi),oi)[email protected] (Y.H. Choi).
undergoing genetic and morphologic transformations. Apoptoticcells are characterized by several unique features, including cellshrinkage, chromatin condensation, DNA fragmentation, cell sur-face expression of phosphatidylserine, and membrane blebbing(Okada and Mak, 2004; Jin and el-Deiry, 2005; Han et al., 2008).A central component of the apoptotic machinery is a family of cy-stein-containing aspartate-specific proteases, termed caspase.Caspases are present in cells as inactive proenzymes, with the ac-tive tetramer being formed as a result of removal of the prodomainand cleavage between the large and small subunits (Hengartner,2000; Suda et al., 1993). Caspase activity is either directly or indi-rectly responsible for the cleavage of several intracellular proteinsthat are characteristically proteolysed during apoptosis. It has beenreported that two major pathways, the Fas/tumor necrosis factor(TNF) death receptor (extrinsic) pathway and the mitochondria-dependent (intrinsic) pathway, lead to activation of caspases and

Table 1Gene-specific primers for RT-PCR
Name Sequence of primers
TRAIL Sence 50-ATG GCT ATG ATG GAG TCC AG-30
Antisence 50-TTG TCC TGC ATC TGC TTC AGC-30
DR4 Sence 50-CAG AAC GTC CTG GAG CCT GTA AC-30
Antisence 50-ATG TCC ATT GCC TGA TTC TTT GTG-30
DR5 Sence 50-GGG AAG AAG ATT CTC CTG AGA TGT G-30
Antisence 50-ACA TTG TCC TCA GCC CCA GGT CG-30
Fas Sence 50-TCT AAC TTG GGG TGG CTT TGT CTT C-30
Antisence 50-GTG TCA TAC GCT TTC TTT CCA T-30
FasL Sence 50-GGA TTG GGC CTG GGG ATG TTT CA-30
Antisence 50-AGC CCA GTT TCA TTG ATC ACA AGG-30
Bax Sence 50-ATG-GAC-GGG-TCC-GGG-GAG-30
Antisence 50-TGG-AAG-AAG-ATG-GGC-TGA-30
Bcl-2 Sence 50-CAG-CTG-CAC-CTG-ACG-30
Antisence 50-GCT-GGG-TAG-GTG-CAT-30
Bcl-xL Sence 50-CAG CTG CAC CTG ACG-30
Antisence 50-GCT GGG TAG GTG CAT-30
GAPDH Sence 50-CGG-AGT-CAA-CGG-ATT-TGG-TCG-TAT-30
Antisence 50-AGC-CTT-CTC-CAT-GGT-GGT-GAA-GAC-30
1574 C. Park et al. / Toxicology in Vitro 22 (2008) 1573–1581
consequent apoptosis in mammalian cells (Fulda and Debatin,2006; Wajant et al., 2005). The Fas/TNF death receptor pathwayis initiated by binding of a ligand to the Fas/TNF death receptoron the cell surface, which then activates caspase-8 and apoptoticcell death. However, changes in the mitochondrial integrity in re-sponse to a broad range of physical and chemical stimuli can trig-ger the intrinsic pathway of apoptosis (Chowdhury et al., 2006;Fulda and Debatin, 2006). It is believed that the release of cyto-chrome c from the mitochondria into the cytosol is a key eventin the intrinsic pathway (Mow et al., 2001; Mohamad et al.,2005). Once in the cytosol, cytochrome c can activate caspase-9,which in turn cleaves and activates the executioner, caspase-3.After activation, several specific substrates for caspase-3 includingpoly-(ADP-ribose)polymerase (PARP) and phospholipase C (PLC)-c1 are cleaved, which eventually leads to apoptosis (Lazebnik etal., 1994; Bae et al., 2000). Consistent with the role that the mito-chondria plays in the control of cell death, survival or apoptoticfactors such as the Bcl-2 family act on the organelle to prevent orfacilitate the release of apoptogenic factors (Yin, 2000; Zinkel etal., 2006; Jeong and Seol, 2008). Many recent studies have shownthat many chemopreventive and/or chemotherapeutic agents cancause tumor cell death via the induction of apoptosis, which isthe preferred method of managing cancer. Therefore, the inductionof apoptotic cell death is an important mechanism in the anti-can-cer properties of many drugs.
Recent studies have shown that marine microorganisms are anovel and rich source of bioactive compounds due to their poten-tial pharmacological activities. However, it is believed that mole-cules produced by those microorganisms have therapeuticproperties that have not yet been discovered (Schweder et al.,2005; Singh and Pelaez, 2008). In addition, studies evaluating thecomponents of marine microorganisms have shown that manyare not general cytotoxic agents, but are instead targeted towardsspecific cellular or biochemical events and may therefore be usefulas anti-microbial, anti-cancer or anti-inflammatory agents (Pro-ksch et al., 2002; Singh and Pelaez, 2008). We previously screenedmarine natural products for the ability to induce apoptosis in can-cer cells and found that streptochlorin isolated from Streptomycessp. exhibited selective cytotoxicity against several cancer cell lines(Choi et al., 2007; Shin et al., 2007). However, the mechanism bywhich streptochlorin exerts its function is not yet fully understood.Therefore, we used the human leukemic U937 cell line as a modelsystem to investigate the effects of streptochlorin-induced apopto-sis. We found that streptochlorin-induced apoptosis is accompa-nied by modulation of the Fas/Fas ligand (FasL) system, Bcl-2family members, mitochondrial dysfunction and activation ofcaspases. In addition, blocking of caspase-3 activation and Bcl-2overexpression reduced streptochlorin-induced apoptosis inU937 cells.
Fig. 1. Growth inhibition in response to streptochlorin treatment of U937 cells.U937 cells were plated at a concentration of 1 � 105 cells per 60-mm plate and thenincubated for 24 h, after which they were treated with various concentrations ofstreptochlorin for the indicated times. (A) Cell number and (B) viability weredetermined by hemocytometer counts of tryphan blue-excluding cells and an MTTassay, respectively. Each time point represents the means ± SD of three independentexperiments. The significance was determined by a Student’s t-test (*p < 0.05 vs.vehicle control).
2. Materials and methods
2.1. Cell culture and viability assay
U937 cells were obtained from the American type culture col-lection (Rockville, MD), and cultured in RPMI 1640 medium sup-plemented with 10% heat-inactivated fetal bovine serum (FBS),2 mM glutamine, 100 U/mL penicillin, and 100 lg/mL streptomy-cin (Gibco BRL, Gaithersburg, MD) at 37� and 5% CO2. The Bcl-2overexpressing U937 (U937/Bcl-2) cells were a generous gift fromDr. T.K. Kwon (Department of Immunology, Keimyung UniversitySchool of Medicine, Taegu, Korea) and maintained in a mediumcontaining 0.7 lg/mL geneticin (G418 sulfate). Streptochlorin iso-lated from Streptomyces sp. (strain 04DH110) was prepared as de-scribed previously (Shin et al., 2007) and dissolved in dimethyl
sulfoxide (DMSO) as a stock solution at 1 mg/mL concentration,and stored in aliquots at �20 �C. Cell number and its viability weredetermined by tryphan blue exclusion assay and MTT assay,respectively.

C. Park et al. / Toxicology in Vitro 22 (2008) 1573–1581 1575
2.2. Nuclear staining with DAPI
After treating U937 cells with streptochlorin, the cells were har-vested, washed in ice-cold phosphate-buffered saline (PBS) andfixed with 3.7% paraformaldehyde (Sigma Chemical Co., St. Louis,MO) in PBS for 10 min at room temperature. The fixed cells werewashed with PBS and stained with a 4,6-diamidino-2-phenylindile(DAPI, Sigma) solution for 10 min at room temperature. The nucle-ar morphology of the cells was examined by fluorescent micros-copy (Carl Zeiss, Germany).
2.3. DNA fragmentation assay
The cells were treated with different concentrations of strepto-chlorin and lysed on ice in a buffer containing 10 mM tris–HCl(pH 7.4), 150 mM NaCl, 5 mM EDTA, and 0.5% triton X-100 for30 min. The lysates were vortexed and cleared by centrifugationat 10,000g for 20 min. The fragmented DNA in the supernatantwas extracted using an equal volume of neutral phenol:chloro-form:isoamylalcohol (25:24:1, v/v/v) and analyzed electrophoret-ically on 1.0% agarose gel containing ethidium bromide (EtBr,Sigma).
Fig. 2. Induction of apoptosis by streptochlorin treatment of U937 cells. After being treatinverted microscopy (A, magnification, �200) or fixed and stained with DAPI so the numagnification, �400). (C) To analyze the DNA fragmentation, the genomic DNA was extraTo quantify the degree of apoptosis induced by streptochlorin, the cells were evaluateddegradation, using a flow cytometer. Each point represents the mean of representativeStudent’s t-test (*p < 0.05 vs. vehicle control).
2.4. Flow cytometry analysis for measurement of sub-G1 phase
The cells were harvested and washed once with PBS, fixed inice-cold 70% ethanol and stored at 4 �C. Prior to analysis, the cellswere washed once again with PBS, suspended in 1 mL of a cold pro-pidium iodide (PI, Sigma) solution containing 100 lg/mL RNase A,50 lg/mL PI, 0.1% (w/v) sodium citrate, and 0.1% (v/v) NP-40, andfurther incubated on ice for 30 min in the dark. Flow cytometricanalyzes were carried out using a flow cytometer (FACS Caliber,Becton Dikinson, San Jose, CA) and CellQuest software was usedto determine the relative DNA content based on the presence ofa red fluorescence.
2.5. RNA extraction and reverse transcription-PCR
The total RNA was prepared using an RNeasy kit (Qiagen, La Jol-la, CA) and primed with random hexamers to synthesize the com-plementary DNA using AMV reverse transcriptase (AmershamCorp., Arlington Heights, IL) according to the manufacturer’sinstructions. A polymerase chain reaction (PCR) was carried outusing a Mastercycler (Eppendorf, Hamburg, Germany) with theprimers shown in Table 1. The following conditions were used
ed with streptochlorin for five days, the cells were sampled and either examined byclear morphology could be photographed under fluorescence using a blue filter (B,cted, electrophoresed in a 1.0% agarose gel and then visualized by EtBr staining. (D)for sub-G1 DNA content, which represents the fractions undergoing apoptotic DNAexperiments performed at least three times. The significance was determined by a

1576 C. Park et al. / Toxicology in Vitro 22 (2008) 1573–1581
for the PCR reactions: 1 � (94 �C for 3 min); 35 � (94 �C for 45 s;58 �C for 45 s; and 72 �C for 1 min) and 1 � (72 �C for 10 min).The amplification products obtained by PCR were separated elec-trophoretically on a 1.0% agarose gel and visualized by EtBrstaining.
2.6. Protein extraction and Western blotting
The cells were harvested and lysed. The protein concentrationswere measured using a Bio Rad protein assay (BioRad Lab., Hercu-les, CA) according to the manufacturer’s instructions. For Westernblot analysis, an equal amount of protein was subjected to elec-trophoresis on SDS–polyacrylamide gel and transferred by elec-troblotting to a nitrocellulose membrane (Schleicher andSchuell, Keene, NH). The blots were probed with the desired anti-bodies for 1 h, incubated with the diluted enzyme-linked second-ary antibody and visualized by enhanced chemiluminescence(ECL) according to the recommended procedure (AmershamCorp., Arlington Heights, IL). The primary antibodies were pur-chased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA) andCell Signaling Technology, Inc. (Boston, MA). The peroxidase-la-beled donkey anti-rabbit immunoglobulin and peroxidase-labeledsheep anti-mouse immunoglobulin were purchased from Amer-sham Corp.
2.7. Assay of caspase-3, -8 and -9 activity
The enzymatic activity of the caspases induced by streptochlo-rin was assayed using a colorimetric assay kit according to themanufacturer’s protocol (R&D Systems, Minneapolis, MN). Briefly,the cells were lysed in a lysis buffer for 30 min on an ice bath.The lysed cells were centrifuged at 12,000g for 10 min, and100 lg of the protein was incubated with 50 lL of a reaction bufferand 5 lL of the colorimetric tetrapeptides, Asp-Glu-Val-Asp
Fig. 3. Effects of streptochlorin on the levels of the death receptor and Bcl-2 family membwas isolated and reverse-transcribed. The resulting cDNA was subjected to PCR using theagarose gel and then visualized by EtBr staining. GAPDH was used as the internal control.after which they were transferred onto nitrocellulose membranes. The membranes wereECL detection system. Actin was used as the internal control.
(DEVD)-p-nitroaniline (pNA) for caspase-3, Ile-Glu-Thr-Asp(IETD)-pNA for caspase-8 and Leu-Glu-His-Asp (LEHD)-pNA forcaspase-9, respectively, at 37 �C for 2 h. The optical density of thereaction mixture was quantified spectrophotometrically at a wave-length of 405 nm.
2.8. Mitochondrial membrane potential (MMP, DWm) assay
The mitochondrial membrane potential (MMP, DWm) of intactcells was measured by flow cytometry with the lipophilic cationicprobe 5,50,6,60-tetrachloro-1,10,3,30-tetraethyl-imidacarbocyanineiodide (JC-1, Sigma). JC-1 is a ratiometric, dual-emission fluores-cent dye that is internalized and concentrated by respiring mito-chondria and can reflect changes in MMP in living cells. There aretwo excitation wavelengths, 527 nm (green) for the monomerform and 590 nm (red) for the J-aggregate form. With normalmitochondrial function, MMP is high and the red fluorescence ispredominant. However, when there is mitochondrial injury,MMP is reduced, leading to an increase in green fluorescence(Salvioli et al., 1997). Quantitation of red and green fluorescentsignals reflects whether mitochondria are damaged. For thisstudy, the cells were trypsinized and the cell pellets were resus-pended in 500 lL of PBS, incubated with 10 lM JC-1 for 20 min at37 �C. The cells were subsequently washed once with cold PBS,suspended in a total volume of 500 lL and analyzed using flowcytometry.
2.9. Statistical analysis
The data is expressed as a mean ± SD. A statistical comparisonwas performed using one-way ANOVA followed by a Fisher test.The significant differences between the groups were determinedusing an unpaired Student’s t-test. A p value <0.05 was consideredsignificant.
ers in U937 cells. (A) After incubation with streptochlorin for five days, the total RNAindicated primers. The reaction products were subjected to electrophoresis in a 1.0%(B) The cells were lysed and the cellular proteins were then separated in SDS–PAGE,
then probed with the indicated antibodies and the proteins were visualized using an
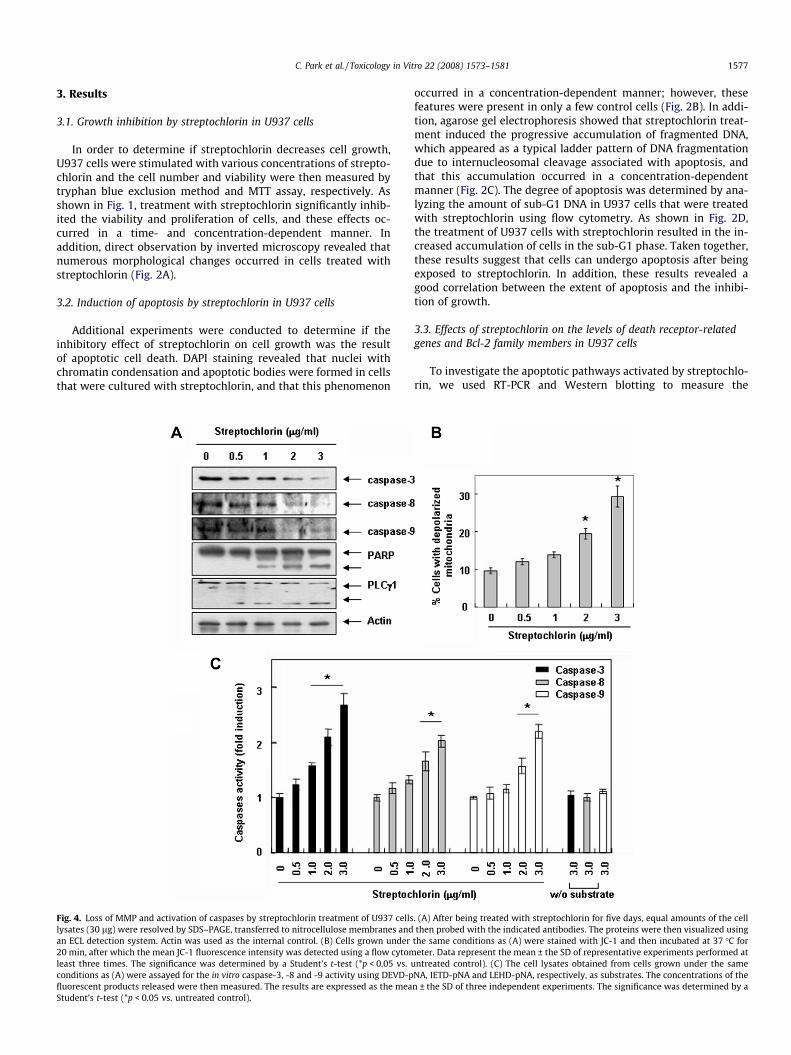
C. Park et al. / Toxicology in Vitro 22 (2008) 1573–1581 1577
3. Results
3.1. Growth inhibition by streptochlorin in U937 cells
In order to determine if streptochlorin decreases cell growth,U937 cells were stimulated with various concentrations of strepto-chlorin and the cell number and viability were then measured bytryphan blue exclusion method and MTT assay, respectively. Asshown in Fig. 1, treatment with streptochlorin significantly inhib-ited the viability and proliferation of cells, and these effects oc-curred in a time- and concentration-dependent manner. Inaddition, direct observation by inverted microscopy revealed thatnumerous morphological changes occurred in cells treated withstreptochlorin (Fig. 2A).
3.2. Induction of apoptosis by streptochlorin in U937 cells
Additional experiments were conducted to determine if theinhibitory effect of streptochlorin on cell growth was the resultof apoptotic cell death. DAPI staining revealed that nuclei withchromatin condensation and apoptotic bodies were formed in cellsthat were cultured with streptochlorin, and that this phenomenon
Fig. 4. Loss of MMP and activation of caspases by streptochlorin treatment of U937 cellslysates (30 lg) were resolved by SDS–PAGE, transferred to nitrocellulose membranes andan ECL detection system. Actin was used as the internal control. (B) Cells grown under20 min, after which the mean JC-1 fluorescence intensity was detected using a flow cytomleast three times. The significance was determined by a Student’s t-test (*p < 0.05 vs. uconditions as (A) were assayed for the in vitro caspase-3, -8 and -9 activity using DEVD-pfluorescent products released were then measured. The results are expressed as the meaStudent’s t-test (*p < 0.05 vs. untreated control).
occurred in a concentration-dependent manner; however, thesefeatures were present in only a few control cells (Fig. 2B). In addi-tion, agarose gel electrophoresis showed that streptochlorin treat-ment induced the progressive accumulation of fragmented DNA,which appeared as a typical ladder pattern of DNA fragmentationdue to internucleosomal cleavage associated with apoptosis, andthat this accumulation occurred in a concentration-dependentmanner (Fig. 2C). The degree of apoptosis was determined by ana-lyzing the amount of sub-G1 DNA in U937 cells that were treatedwith streptochlorin using flow cytometry. As shown in Fig. 2D,the treatment of U937 cells with streptochlorin resulted in the in-creased accumulation of cells in the sub-G1 phase. Taken together,these results suggest that cells can undergo apoptosis after beingexposed to streptochlorin. In addition, these results revealed agood correlation between the extent of apoptosis and the inhibi-tion of growth.
3.3. Effects of streptochlorin on the levels of death receptor-relatedgenes and Bcl-2 family members in U937 cells
To investigate the apoptotic pathways activated by streptochlo-rin, we used RT-PCR and Western blotting to measure the
. (A) After being treated with streptochlorin for five days, equal amounts of the cellthen probed with the indicated antibodies. The proteins were then visualized using
the same conditions as (A) were stained with JC-1 and then incubated at 37 �C foreter. Data represent the mean ± the SD of representative experiments performed atntreated control). (C) The cell lysates obtained from cells grown under the same
NA, IETD-pNA and LEHD-pNA, respectively, as substrates. The concentrations of then ± the SD of three independent experiments. The significance was determined by a

1578 C. Park et al. / Toxicology in Vitro 22 (2008) 1573–1581
expression of the death receptors and corresponding pro-apoptoticligands (TRAIL, DR4, DR5, Fas and FasL), as well as the expression ofthe Bcl-2 family members (Bax, Bcl-2, Bcl-xL and Bid). As shown inFig. 3, the transcriptional and translational levels of FasL and pro-apoptotic Bax expression were markedly increased in the strepto-chlorin-treated U937 cells, whereas the levels of anti-apoptoticBcl-2 expression were significantly inhibited in response to thestreptochlorin treatment in a concentration-dependent manner.However, the levels of TRAIL, DR4, DR5, Fas, Bcl-xL and Bid expres-sion were relatively unchanged in response to streptochlorintreatment.
3.4. Loss of MMP and activation of caspase by streptochlorin
The role of the mitochondria in streptochlorin-induced apopto-sis was further investigated by examining the effect of streptochlo-rin on the MMP and caspase activity in U937 cells. Exposure ofU937 cells to various concentrations of streptochlorin led to a sig-nificant reduction in the level of MMP, and this reduction occurredin a dose-dependent manner (Fig. 4A). In addition, streptochlorintreatment resulted in down-regulation of pro-caspase-3, -8 and-9 (Fig. 4B). In order to further quantify the proteolytic activation
Fig. 5. Inhibition of streptochlorin-induced apoptosis of U937 cells by a caspase-3 inhibwith 3.0 lg streptochlorin for five days. (A) Equal amounts of cell lysates (30 lg) were resanti-caspase-3, anti-PARP and anti-PLC-c1 antibodies. Actin was used as an internal co(A) were assayed for in vitro caspase-3 activity using DEVD-pNA as a substrate and therepresent the average of two independent experiments and the results are expressed as tstained with DAPI for 10 min and then photographed with a fluorescence microscope usDNA and separating it by electrophoresis in a 1.0% agarose gel containing EtBr. (E) Cells gra flow cytometer. Results represent the mean ± the SD of triplicate determinations. The
of the caspases, the concentration of protein in the lysates of cellstreated with streptochlorin was normalized and then assayed fortheir caspase-3, -8 and -9 activity using DEVD-pNA, IETD-pNAand LEHD-pNA as substrates, respectively. As shown in Fig. 4C,treatment with streptochlorin increased the activity of the caspas-es in a concentration-dependent manner (approximately 2.5-foldafter five days of exposure to 3.0 lg/mL streptochlorin). Subse-quent Western blot analysis revealed that progressive proteolyticcleavage of the PARP and PLC-c1 proteins, which are downstreamtargets of the activated caspase-3 (Lazebnik et al., 1994; Bae et al.,2000), occurred in U937 cells that were treated with streptochlo-rin, and that this cleavage was induced in a concentration-depen-dent manner (Fig. 4A).
3.5. Inhibition of streptochlorin-induced apoptosis by a caspase-3inhibitor
Caspase-3 is one of the key proteases responsible for the cleav-age of PARP and PLC-c1 and subsequent apoptosis. Therefore, weused a general and potent inhibitor of caspase-3, z-DEVD-fmk, tofurther evaluate the significance of caspase-3 activation in re-sponse to streptochlorin treatment. As shown in Fig. 5A and B,
itor. Cells were treated with z-DEVD-fmk (50 lM) for 1 h prior to being challengedolved by SDS–PAGE, transferred to nitrocellulose membranes, and then probed withntrol. (B) The cell lysates obtained from cells grown under the same conditions asconcentrations of the fluorescent products released were then measured. The datahe mean ± the SD of three independent experiments. (C) The cells were sampled anding a blue filter. (D) DNA fragmentation was analyzed by extracting the fragmentedown under the same conditions as (A) were evaluated for sub-G1 DNA content usingsignificance was determined by a Student’s t-test (*p < 0.05 vs. vehicle control).

C. Park et al. / Toxicology in Vitro 22 (2008) 1573–1581 1579
pre-treatment of the cells with z-DEVD-fmk (50 lM) attenuatedcaspase-3 activation as well as the degradation of PARP and PLC-c1. Furthermore, z-DEVD-fmk prevented chromatin condensation,DNA fragmentation and the increase in the sub-G1 population thatwas observed in streptochlorin-treated cells (Fig. 5C–E).
3.6. Inhibition of streptochlorin-induced apoptosis by overexpressionof Bcl-2
Because the levels of anti-apoptotic Bcl-2 protein were mark-edly down-regulated in streptochlorin-treated cells, the effect ofhigh intracellular levels of Bcl-2 on streptochlorin-induced apopto-sis was evaluated using U937/vector and U937/Bcl-2 cells that con-stitutively expressed high levels of Bcl-2. Western blot analyzesrevealed that treatment with streptochlorin significantly inducedthe cleavage of caspases, PARP and PLC-c1 in U937/vector cells,whereas the treatment of cells that overexpressed Bcl-2 did not in-duce the cleavage of these molecules (Fig. 6A). However, the in-creased levels of FasL protein induced by treatment of U937/vector cells with streptochlorin were not reduced in U937/Bcl-2cells. Next, cell lysates containing equal amounts of total proteinfrom cells treated with streptochlorin were assayed to determinetheir in vitro caspase activity. As shown in Fig. 6B and C, treatmentwith streptochlorin significantly increased the activity of caspase-3, -8, and -9 and induced a loss of MMP in U937/vector cells, butnot in U937/Bcl-2 cells. Additionally, treatment of U937/vectorcells with streptochlorin significantly induced the accumulationof cells with sub-G1 DNA content, increased DNA fragmentationand increased the appearance of apoptotic bodies (Fig. 7A–C), how-ever, it did not alter Bcl-2 overexpression.
Fig. 6. Bcl-2 overexpression inhibits streptochlorin-induced activation of caspases andtreated with 3.0 lg of streptochlorin for five days, after which they were lysed and thproteins were transferred onto nitrocellulose membranes, which were then probed withobtained from cells grown under the same conditions as (A) were assayed for the in vitromeasured. The results are expressed as the mean ± the SD of three independent experimwhich the mean JC-1 fluorescence intensity was detected using a flow cytometer. Data rtimes. The significance was determined by a Student’s t-test (*p < 0.05 vs. untreated con
4. Discussion
Although recent studies have shown that streptochlorin cansuppress the growth of cultured cancer cells in vitro (Shin et al.,2007), the signaling pathway by which this occurs has not beenelucidated. In the present study, we showed that streptochlorintreatment induced mitochondrial damage and apoptosis in humanleukemia U937 cells. Furthermore, a caspase-3 inhibitor signifi-cantly attenuated this streptochlorin-induced apoptosis, andBcl-2 overexpression was found to decrease caspase activationdownstream of the mitochondria, thereby inhibiting the strepto-chlorin-mediated apoptosis (see Fig. 7).
It has recently been proposed that mitochondria represent apossible target for anti-cancer drug induced apoptosis (Mow etal., 2001; Mohamad et al., 2005). Since the discovery of Bcl-2, sev-eral mechanisms for the anti-apoptotic properties of this proteinhave been proposed (Murphy et al., 1996; Kluck et al., 1997). Theanti-apoptotic function of Bcl-2 against pro-apoptotic Bax may beexplained by its ability to control several key steps involved indeath signaling. Bcl-2 family significantly regulates apoptosiseither as an activator (Bax) or as an inhibitor (Bcl-2 or Bcl-xL);therefore, it has been suggested that the Bax/Bcl-2 ratio is a keyfactor in regulation of the apoptotic process (Thees et al., 2005;Zinkel et al., 2006). The results of this study indicated that strepto-chlorin treatment results in significant cell growth inhibition andthe induction of apoptosis in U937 cells. Therefore, we evaluatedthe effects of streptochlorin on the levels of proteins in the Bcl-2family in U937 cells to test the mechanism by which streptochlo-rin-induced apoptosis occurs. Our data demonstrated that strepto-chlorin-induced apoptosis was related to augmented levels of the
mitochondrial dysfunction in U937 cells. (A) U937/vector or U937/Bcl-2 cells weree cellular proteins (30 lg) were then separated by SDS–PAGE. Next, the separated
the indicated antibodies. Actin was used as an internal control. (B) The cell lysatescaspase activity. The concentrations of the fluorescent products released were thenents. (C) The cells were stained with JC-1 and incubated at 37 �C for 20 min, after
epresent the mean ± the SD of representative experiments performed at least threetrol).
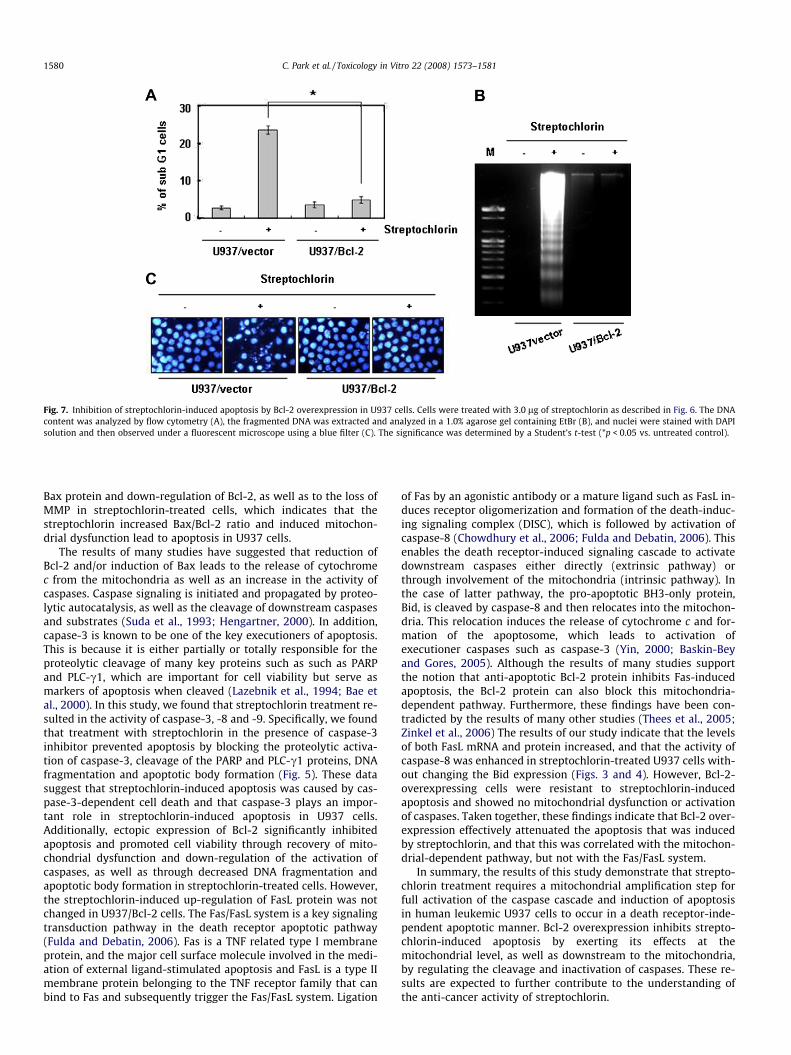
Fig. 7. Inhibition of streptochlorin-induced apoptosis by Bcl-2 overexpression in U937 cells. Cells were treated with 3.0 lg of streptochlorin as described in Fig. 6. The DNAcontent was analyzed by flow cytometry (A), the fragmented DNA was extracted and analyzed in a 1.0% agarose gel containing EtBr (B), and nuclei were stained with DAPIsolution and then observed under a fluorescent microscope using a blue filter (C). The significance was determined by a Student’s t-test (*p < 0.05 vs. untreated control).
1580 C. Park et al. / Toxicology in Vitro 22 (2008) 1573–1581
Bax protein and down-regulation of Bcl-2, as well as to the loss ofMMP in streptochlorin-treated cells, which indicates that thestreptochlorin increased Bax/Bcl-2 ratio and induced mitochon-drial dysfunction lead to apoptosis in U937 cells.
The results of many studies have suggested that reduction ofBcl-2 and/or induction of Bax leads to the release of cytochromec from the mitochondria as well as an increase in the activity ofcaspases. Caspase signaling is initiated and propagated by proteo-lytic autocatalysis, as well as the cleavage of downstream caspasesand substrates (Suda et al., 1993; Hengartner, 2000). In addition,capase-3 is known to be one of the key executioners of apoptosis.This is because it is either partially or totally responsible for theproteolytic cleavage of many key proteins such as such as PARPand PLC-c1, which are important for cell viability but serve asmarkers of apoptosis when cleaved (Lazebnik et al., 1994; Bae etal., 2000). In this study, we found that streptochlorin treatment re-sulted in the activity of caspase-3, -8 and -9. Specifically, we foundthat treatment with streptochlorin in the presence of caspase-3inhibitor prevented apoptosis by blocking the proteolytic activa-tion of caspase-3, cleavage of the PARP and PLC-c1 proteins, DNAfragmentation and apoptotic body formation (Fig. 5). These datasuggest that streptochlorin-induced apoptosis was caused by cas-pase-3-dependent cell death and that caspase-3 plays an impor-tant role in streptochlorin-induced apoptosis in U937 cells.Additionally, ectopic expression of Bcl-2 significantly inhibitedapoptosis and promoted cell viability through recovery of mito-chondrial dysfunction and down-regulation of the activation ofcaspases, as well as through decreased DNA fragmentation andapoptotic body formation in streptochlorin-treated cells. However,the streptochlorin-induced up-regulation of FasL protein was notchanged in U937/Bcl-2 cells. The Fas/FasL system is a key signalingtransduction pathway in the death receptor apoptotic pathway(Fulda and Debatin, 2006). Fas is a TNF related type I membraneprotein, and the major cell surface molecule involved in the medi-ation of external ligand-stimulated apoptosis and FasL is a type IImembrane protein belonging to the TNF receptor family that canbind to Fas and subsequently trigger the Fas/FasL system. Ligation
of Fas by an agonistic antibody or a mature ligand such as FasL in-duces receptor oligomerization and formation of the death-induc-ing signaling complex (DISC), which is followed by activation ofcaspase-8 (Chowdhury et al., 2006; Fulda and Debatin, 2006). Thisenables the death receptor-induced signaling cascade to activatedownstream caspases either directly (extrinsic pathway) orthrough involvement of the mitochondria (intrinsic pathway). Inthe case of latter pathway, the pro-apoptotic BH3-only protein,Bid, is cleaved by caspase-8 and then relocates into the mitochon-dria. This relocation induces the release of cytochrome c and for-mation of the apoptosome, which leads to activation ofexecutioner caspases such as caspase-3 (Yin, 2000; Baskin-Beyand Gores, 2005). Although the results of many studies supportthe notion that anti-apoptotic Bcl-2 protein inhibits Fas-inducedapoptosis, the Bcl-2 protein can also block this mitochondria-dependent pathway. Furthermore, these findings have been con-tradicted by the results of many other studies (Thees et al., 2005;Zinkel et al., 2006) The results of our study indicate that the levelsof both FasL mRNA and protein increased, and that the activity ofcaspase-8 was enhanced in streptochlorin-treated U937 cells with-out changing the Bid expression (Figs. 3 and 4). However, Bcl-2-overexpressing cells were resistant to streptochlorin-inducedapoptosis and showed no mitochondrial dysfunction or activationof caspases. Taken together, these findings indicate that Bcl-2 over-expression effectively attenuated the apoptosis that was inducedby streptochlorin, and that this was correlated with the mitochon-drial-dependent pathway, but not with the Fas/FasL system.
In summary, the results of this study demonstrate that strepto-chlorin treatment requires a mitochondrial amplification step forfull activation of the caspase cascade and induction of apoptosisin human leukemic U937 cells to occur in a death receptor-inde-pendent apoptotic manner. Bcl-2 overexpression inhibits strepto-chlorin-induced apoptosis by exerting its effects at themitochondrial level, as well as downstream to the mitochondria,by regulating the cleavage and inactivation of caspases. These re-sults are expected to further contribute to the understanding ofthe anti-cancer activity of streptochlorin.

C. Park et al. / Toxicology in Vitro 22 (2008) 1573–1581 1581
Acknowledgements
This research was supported by a Grant (M2007-03) from Mar-ine Bioprocess Research Center of the Marine Bio 21 Center fundedby the Ministry of Land, Transport and Maritime, Republic of Korea.
References
Bae, S.S., Perry, D.K., Oh, Y.S., Choi, J.H., Galadari, S.H., Ghayur, T., Ryu, S.H., Hannun,Y.A., Suh, P.G., 2000. Proteolytic cleavage of phospholipase C-c1 duringapoptosis in Molt-4 cells. FASEB Journal 14, 1083–1092.
Baskin-Bey, E.S., Gores, G.J., 2005. Death by association: BH3 domain-only proteinsand liver injury. The American Journal of Physiology – Gastrointestinal andLiver Physiology 289, G987–G990.
Choi, I.K., Shin, H.J., Lee, H.S., Kwon, H.J., 2007. Streptochlorin, a marine naturalproduct, inhibits NF-jB activation and suppresses angiogenesis in vitro. Journalof Microbiology and Biotechnology 17, 1338–1343.
Chowdhury, I., Tharakan, B., Bhat, G.K., 2006. Current concepts in apoptosis: thephysiological suicide program revisited. Cellular and Molecular Biology Letters11, 506–525.
Fulda, S., Debatin, K.M., 2006. Extrinsic versus intrinsic apoptosis pathways inanticancer chemotherapy. Oncogene 25, 4798–4811.
Han, S.I., Kim, Y.S., Kim, T.H., 2008. Role of apoptotic and necrotic cell death underphysiologic conditions. BMB Reports 41, 1–10.
Hengartner, M.O., 2000. The biochemistry of apoptosis. Nature 407, 770–776.Jeong, S.Y., Seol, D.W., 2008. The role of mitochondria in apoptosis. BMB Reports 41,
11–22.Jin, Z., el-Deiry, W.S., 2005. Overview of cell death signaling pathways. Cancer
Biology and Therapy 4, 139–163.Kluck, R.M., Bossy-Wetzel, E., Green, D.R., Newmeyer, D.D., 1997. The release of
cytochrome c from mitochondria: a primary site for Bcl-2 regulation ofapoptosis. Science 275, 1132–1136.
Lazebnik, Y.A., Kaufmann, S.H., Desnoyers, S., Poirier, G.G., Earnshaw, W.C., 1994.Cleavage of poly(ADP-ribose) polymerase by a proteinase with properties likeICE. Nature 371, 346–347.
Mohamad, N., Gutierrez, A., Nunez, M., Cocca, C., Martin, G., Cricco, G., Medina, V.,Rivera, E., Bergoc, R., 2005. Mitochondrial apoptotic pathways. Biocell 29, 149–161.
Mow, B.M., Blajeski, A.L., Chandra, J., Kaufmann, S.H., 2001. Apoptosis and theresponse to anticancer therapy. Current Opinion in Oncology 13, 453–462.
Murphy, A.N., Bredesen, D.E., Cortopassi, G., Wang, E., Fiskum, G., 1996. Bcl-2potentiates the maximal calcium uptake capacity of neural cell mitochondria.Proceedings of the National Academy of Sciences of the United States ofAmerica 93, 9893–9898.
Okada, H., Mak, T.W., 2004. Pathways of apoptotic and non-apoptotic death intumour cells. Nature Reviews Cancer 4, 592–603.
Proksch, P., Edrada, R.A., Ebel, R., 2002. Drugs from the seas – current status andmicrobiological implications. Applied Microbiology and Biotechnology 59, 125–134.
Salvioli, S., Ardizzoni, A., Franceschi, C., Cossarizza, A., 1997. JC-1, but not DiOC6(3)or rhodamine 123, is a reliable fluorescent probe to assess delta psi changes inintact cells: implications for studies on mitochondrial functionality duringapoptosis. FEBS Letters 411, 77–82.
Schweder, T., Lindequist, U., Lalk, M., 2005. Screening for new metabolites frommarine microorganisms. Advances in Biochemical Engineering andBiotechnology 96, 1–48.
Shin, H.J., Jeong, H.S., Lee, H.S., Park, S.K., Kim, H.M., Kwon, H.J., 2007. Isolation andstructure determination of streptochlorin, an antiproliferative agent from amarine-derived Streptomyces sp. 04DH110. Journal of Microbiology andBiotechnology 17, 1403–1406.
Singh, S.B., Pelaez, F., 2008. Biodiversity, chemical diversity and drug discovery.Progress in Drug Research 65, 143–174.
Suda, T., Takahashi, T., Golstein, P., Nagata, S., 1993. Molecular cloning andexpression of the Fas ligand, a novel member of the tumor necrosis factorfamily. Cell 75, 1169–1178.
Thees, S., Hubbard, G.B., Winckler, J., Schultz, C., Rami, A., 2005. Specific alteration ofthe Bax/Bcl2 ratio and cytochrome c without execution of apoptosis in thehippocampus of aged baboons. Restorative Neurology and Neuroscience 23, 1–9.
Wajant, H., Gerspach, J., Pfizenmaier, K., 2005. Tumor therapeutics by design:targeting and activation of death receptors. Cytokine and Growth FactorReviews 16, 55–76.
Yin, X.M., 2000. Signal transduction mediated by Bid, a pro-death Bcl-2 familyproteins, connects the death receptor and mitochondria apoptosis pathways.Cell Research 10, 161–167.
Zinkel, S., Gross, A., Yang, E., 2006. Bcl-2 family in DNA damage and cell cyclecontrol. Cell Death and Differentiation 13, 1351–1359.
Related Documents











