121 Mutation Research, 51 (1978) 121--132 © Elsevier/North-Holland Biomedical Press INCREASED SENSITIVITY OF UV-REPAIR-DEFICIENT HUMAN CELLS TO DNA BOUND PLATINUM PRODUCTS WHICH UNLIKE THYMINE DIMERS ARE NOT RECOGNIZED BY AN ENDONUCLEASE EXTRACTED FROM MICROCOCCUS L UTEUS H.N.A. FRAVAL, C.J. RAWLINGS and J.J. ROBERTS Institute of Cancer Research, Royal Cancer Hospital, Pollards Wood Research Station, Nightingales Lane, Chalfont St. Giles, Bucks. HP8 4SP (Great Britain) (Received 28 October 1977) (Revision received 8 February 1978) (Accepted 28 February 1978) Summary We have studied the response of human cells in cultdre to cis platinum[II] diammine dichloride (cis Pt[II] ) induced DNA damage. The survival data, mea- sured as a function of cis Pt[II] dose were similar in a normal cell line (Human foetal lung) compared to a UV-sensitive, thymine dimer excision repair-defi- cient cell line (Xeroderma pigmentosum). However, there was a marked differ- ence between the two cell lines when binding to DNA was plotted against dose of cis Pt[II] given for 1 h. When these findings were expressed as cell survival versus binding to DNA, a 4.1-fold difference between the slopes of the survival curves for the two cell lines was obtained. These findings are consistent with the notion that normal cells are able to excise cis Pt[II] induced damage from their genome and thus increase their ability to survive as compared to excision- deficient cells. An endonuclease preparation from Micrococcus luteus is able to recognise UV damage in DNA, but did not recognise cis Pt[II] induced damage. These results possibly indicate differences in the pathways of repair of damage caused by the two agents. Introduction The antitumour properties of cis Pt [II], currently undergoing extensive clin- ical trials [27,35,36], are thought to be related to the ability of this compound to bind to DNA and inhibit the normal progression of DNA synthesis [11,32]. The DNA adducts formed following cis Pt[II] treatment are still in dispute and for this reason, detailed analysis of the cellular ability to remove specific cis Pt[II] damage from the genome has not been forthcoming [18,20]. Beck and Brubaker [3], and Drobnik et al. [9] have suggested on the basis of

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

121
Mutation Research, 51 (1978) 121--132 © Elsevier/North-Holland Biomedical Press
INCREASED SENSITIVITY OF UV-REPAIR-DEFICIENT HUMAN CELLS TO DNA BOUND PLATINUM PRODUCTS WHICH UNLIKE THYMINE DIMERS ARE NOT RECOGNIZED BY AN ENDONUCLEASE EXTRACTED FROM MICROCOCCUS L UTEUS
H.N.A. FRAVAL, C.J. RAWLINGS and J.J. ROBERTS
Institute of Cancer Research, Royal Cancer Hospital, Pollards Wood Research Station, Nightingales Lane, Chalfont St. Giles, Bucks. HP8 4SP (Great Britain)
(Received 28 October 1977) (Revision received 8 February 1978) (Accepted 28 February 1978)
Summary
We have studied the response of human cells in cultdre to cis platinum[II] diammine dichloride (cis Pt[II] ) induced DNA damage. The survival data, mea- sured as a function of cis Pt[II] dose were similar in a normal cell line (Human foetal lung) compared to a UV-sensitive, thymine dimer excision repair-defi- cient cell line (Xeroderma pigmentosum). However, there was a marked differ- ence between the two cell lines when binding to DNA was plot ted against dose of cis Pt[II] given for 1 h. When these findings were expressed as cell survival versus binding to DNA, a 4.1-fold difference between the slopes of the survival curves for the two cell lines was obtained. These findings are consistent with the notion that normal cells are able to excise cis Pt[II] induced damage from their genome and thus increase their ability to survive as compared to excision- deficient cells. An endonuclease preparation from Micrococcus luteus is able to recognise UV damage in DNA, but did not recognise cis Pt[II] induced damage. These results possibly indicate differences in the pathways of repair of damage caused by the two agents.
Introduction
The ant i tumour properties of cis Pt [II], currently undergoing extensive clin- ical trials [27,35,36], are thought to be related to the ability of this compound to bind to DNA and inhibit the normal progression of DNA synthesis [11,32]. The DNA adducts formed following cis Pt[II] t reatment are still in dispute and for this reason, detailed analysis of the cellular ability to remove specific cis Pt[II] damage from the genome has not been forthcoming [18,20].
Beck and Brubaker [3], and Drobnik et al. [9] have suggested on the basis of

122
experiments with various DNA-repair-deficient mutants of Escherichia coli that the cis Pt[II] induced damage was reversible and that this might be related to an excision or recombination repair facility. By contrast, when platinum- treated transforming DNA was assayed in host cells of different repair capacity, it was concluded by Munchausen [21] that the excision-repair pathway did not modify the biological activity of the DNA in this system. Treatment of Chinese hamster V79 cells with cis Pt[II] has been shown not to lead to the rapid for- mation of single-strand breaks or alkali-labile sites in cellular DNA [31]. On the other hand some indirect evidence for the presence of an excision-repair pro- cess in Chinese hamster cells was obtained. Thus t ime-dependent changes in the sedimentation characteristics of DNA from cis Pt[II] treated cells in alkaline sucrose gradients could be due to the loss of interstrand cross links during sev- eral hours by a DNA-repair process [31].
In order to obtain additional evidence for the existence of an excision-repair process operating on cis Pt[II] damaged DNA, we have compared the ability of a normal human cell line and a cell line known to be deficient in the excision of UV-induced thymine dimers [6] and certain chemically induced damage [5,17, 28,30], to survive a given binding of cis Pt[II] to their genome. We have dem- onstrated that normal cells are approximately 4 times more efficient than our Xeroderma pigmentosum XP 12 BE cells in surviving a given cis Pt[II] binding level to DNA and therefore conclude that this is likely to be due to the greater ability of normal cells to excise cis Pt[II] induced damage from the genetic material.
To compare the ability of one step of the excision-repair mechanism for UV- induced damage to recognize cis Pt[II] damage, we have determined the activ- ity of a crude extract of Micrococcus luteus to recognize cis Pt[II] damage. The crude extract has been shown to contain endonuclease activity [4] which is able to insert single-strand breaks in the DNA of UV-irradiated cells. However, we report here that we have not been able to demonstrate a comparable ability of the extract to recognize cis Pt[II] induced damage. We therefore speculate that a different pathway may exist in normal cells for its removal.
Materials and methods
Cell Strains Human foetal lung cells (FL/A) were cultured in our laboratory from pri-
mary amniotic explants. Frozen stocks of a DNA excision-repair deficient human skin fibroblast cell line of complimentat ion group A were obtained from the American type Culture Collection, Rockville, Md. (U.S.A.) and were designated XP 12 BE (CRL 1223). Chinese hamster V79-379A is an established cell line in our laboratory.
Media and cell culture Both human cell lines were grown at 37°C as monolayer cultures in sealed
Nunclon 800-ml plastic flasks (A/S Nunc, Denmark) in Gibco-modified Eagle minimal essential medium (MEM) ( F l l ) supplemented with 15% foetal calf serum (Gibco Bio-cult, Glasgow, U.K.), benzyl penicillin (200 i.U/ml), strep- tomycin sulphate (200 #g/ml) and sodium bicarbonate (2.2 g/l). Recently con- fluent monolayer cell cultures were subcultured by decanting the medium,

123
washing the cells with 20 ml PBS followed by the addition of 0.25% trypsin. Excess trypsin was immediately removed leaving only a film of trypsin still in contact with the cells. After 10 min at 37°C cells were stripped from the sur- face and resuspended in fresh medium giving a dilution factor of between 2 and 4. The cell suspension was re-inoculated into new flasks which were aspi- rated before sealing with 5% CO2 in air.
Chinese hamster V79 cells were grown in suspension culture as previously described [ 24].
Chemicals Cis Pt[II] was generously given to use by Johnson Matthey Research Centre.
[14C]Thymide 61.5 mCi/mMol and [3H]thymidine 20--25 Ci/mMol were ob- tained from the Radiochemical Centre, Amersham, (U.K.).
Cis Pt [II] treatment Cells which were approaching or had recently attained confluence were
stripped into a small volume of 0.25% trypsin and were subsequently resus- pended in Gibco F14 MEM (suspension medium) supplemented as described above.
(A) Monolayer treatment. Cells in suspension medium were appropriately diluted using Gibco F14 MEM supplemented with 7.5% foetal calf serum plus antibiotics (dilution medium). 1 ml of cell suspension was added to 9-cm petri dishes (Sterilin Ltd., U.K.) containing 9 ml Gibco F l l MEM. After allowing 4--6 h to attach to the vessel surface, the cells were t}eated with various doses of a freshly prepared solution of cis Pt[II] in dimethyl sulphoxide (DMSO). The DMSO concentration never exceeded 0.1% (v/v). After 1 h the medium was removed from the cells which were then rinsed once with 10 ml PBS and refed with 15 ml fresh medium (Gibco F l l ) .
(B) Suspension treatment. Stirred suspension cultures (5 × l 0 s cells/ml) were treated with various concentrations of cis Pt[II] in DMSO for 1 h at 37 ° C. Cells were pelleted by centrifugation at 200 rpm in an MSE bench centrifuge at 37 ° C, resuspended in fresh dilution medium and serially diluted prior to adding to 9-cm dishes for estimation of colony-forming ability. The remaining cells (10 s per treatment) required for binding studies were washed once with PBS, pelleted, flash frozen and stored at --20 ° C for subsequent isolation of the DNA.
UV trea tmen t 106 cells growing as a monolayer in 9-cm petri dishes were washed once with
PBS after removing the medium and exposed to UV irradiation (predominantly 254 nm) using a low pressure mercury germicidal lamp. (Ultra Violet Products Inc., San Gabriel, U.S.A.) at an incident intensity of 1 J/m2/sec. The treated monolayer was trypsinised, resuspended in dilution medium, serially diluted and plated on to 9-cm dishes containing feeder layers prepared the previous day.
Feeder layer and survival assay A cell suspension of the appropriate cell line (2 × 10 s cells/ml) was irradiated
with 7.5 krad and was subsequently added to plates (10 s cells/plate) 18 h

124
before an experiment. Appropriate numbers of treated cells (to give 50 colo- nies/plate) were added in 1 ml of medium to these plates which were then incubated in a water-saturated atmosphere of 5% CO2 in air at 37°C. After 7 days all plates were refed with fresh medium and reincubated for a further 7 days. The medium was then decanted and the plates, washed once with PBS, were fixed with methanol and stained with Giemsa. The capacity of cells to survive a t reatment was judged by their ability to form colonies containing 50 cells or more. Survival was expressed as a percentage of the control survival. Plating efficiencies for FL/A varied from 46 to 72% and for XP 12 BE from 15 to 20%.
DNA extraction The method for DNA extraction was modified from that of Kirby and Cook
[13]. A frozen pellet of 108 cells was resuspended in 12 ml of a solution con- taining 6% w/v p-aminosalicylic acid, 6% w/v sec-butanol, 1% w/v NaC1 and 1% triisopropylnaphthalene sulphonic acid (sodium salt) (PAS solution). The resultant viscous suspension was added to 10 ml of a phenol solution (contain- ing 500 g phenol, 75 ml m-cresol, 55 ml H20 and 0.5 g 8-hydroxyquinoline) and the resultant solution was shaken vigorously. Following centrifugation at 10 000 g for 20 min at room temperature the clear PAS layer was removed together with the interface and was added to 5 ml of phenol solution. After vigorous shaking the emulsion was recentrifuged. The DNA was precipitated from the resulting PAS layer with two volumes of 2-ethoxyethanol. The fibrous precipitate was twice washed with 75% ethanol, absolute ethanol, and ether in turn and dried in a vacuum dessicator. The DNA was hydrolysed in 0.3 ml of 1 N HC1 at 70°C for 2 h and the DNA content was calculated in mg/ml using absorption spectroscopy at 260 nm (e = 8900). In all cases the extracted DNA, measured as a function of the ratio absorption at 260 nm/280 nm was 1.8 + 0.1, was judged to be pure.
Estimation of platinum A Perkin Elmer Atomic Absorption spectrometer (Model 306) was standard-
ized using a 0.2 pg/ml platinum chloride stock solution. The cis Pt[II] con- tained in 50-#1 aliquots of DNA hydrolysates was determined and expressed as nmoles Pt/g DNA.
Preparation of Micrococcus luteus endonuclease extract The M. luteus extract was prepared according to the method of Carrier and
Setlow [4]. We have only purified the enzyme as far as fraction II in order to retain the maximum endonuclease activity. The crude extract was stored in vials in liquid nitrogen.
Detection of endonuelease-sensitive sites For each experiment two stirred suspension cultures of Chinese hamster
V79-379A cells were labelled overnight with either 0.2 pCi/ml [14C]thymidine 61.5 mCi/mmol or 0.5 pCi/ml [3H]thymidine 0.4 Ci/mmol. Cells labelled with [14C]thymidine were treated with either UV or cis Pt[II] as described above. An equal number of [3H]thymidine-labelled cells was mixed with these treated cells and provided an internal control for the experiment. The DNA from this mixed culture was extracted with PAS solution followed by a single depro-

125
teination with phenol solution and gentle shaking. The aqueous PAS (nucleic acid solution) was carefully removed and dialysed (Visking, size 5, Medicell International, U.K.) against 2 1 of enzyme buffer {containing 5 mM K2HPO4, 0.5 mM EDTA Na2, and 10% glycerol) at 4°C overnight.
Enzyme treatment of the DNA dialysate was carried out at 35°C for 60 min in the dark using 0.4 ml dialysate, 0.05 ml buffer (containing 0.5 M Tris, 0.1 M mercaptoethanol, 0.1 M EDTA Na2 and 60 pg/ml salmon sperm DNA pH 8.0}, 0.15 ml M. luteus extract, (or 0.15 ml of enzyme buffer for the controls}. After incubation, 100 pl was removed and placed on the lysing layer (0.4 ml contain- ing 0.5 M NaOH, 0.4 M NaC1, 0.2% sodium lauryl n-sarcosinate) of a 11.7 ml 5--25% alkaline sucrose gradient for 100 min at 20°C [32]. Following centrifu- gation at 15 000 rpm for 16 h at 20°C in a SW 40Ti rotor on a Beckman L265B ultracentrifuge, the gradients were fractionated (0.4 ml) and the radio- activity assayed by scintillation counting [26].
The number-average molecular weight Mn of DNA was calculated from the sedimentation profiles using the gradient calibrations previously described [26]. The number of breaks introduced into DNA was calculated according to the method of Paterson et a1.[22] or Waters and Regan [34].
Results
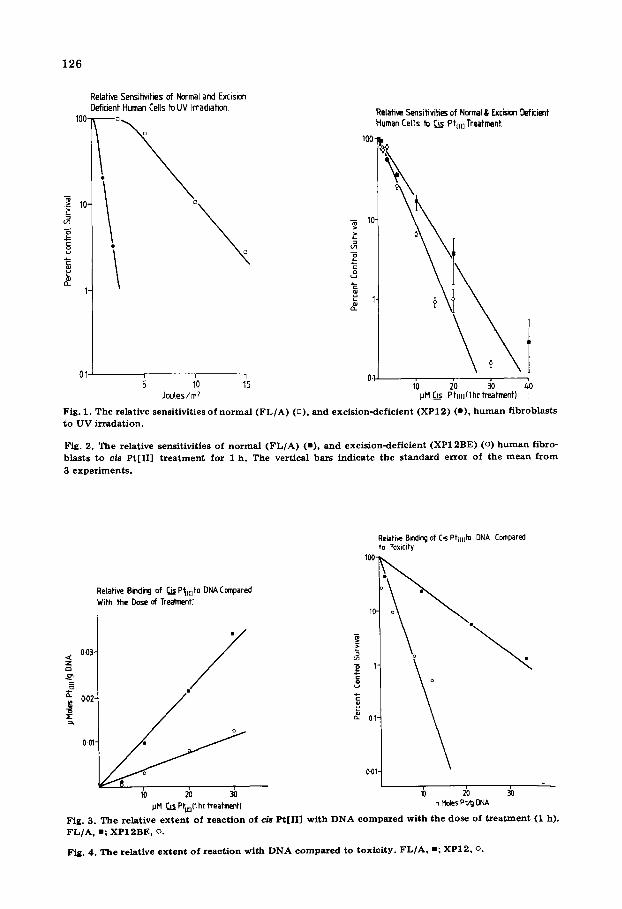
To establish the relative sensitivities of normal foetal lung and Xeroderma cells, survival curves were obtained following treatment with various doses of U.V. (Fig. 1). Clearly XP 12 BE cells showed a marked increase in sensitivity compared to FL/A cells. The Do value of 3.1 J /m 2 (the dose required to reduce the survival from f to 0.37 f on the exponential part of the survival curve) ob- tained for FL/A cells was reduced to 0.55 J /m 2 in XP 12 BE cells. These Do values were similar to those obtained by Kraemer et al. [14] for a comparison of XP 12 BE with normal cells. The XP 12 BE excision-repair-deficient cells are therefore 5.6 times more sensitive to UV-irradiation compared to normal cells. This difference is consistent with the evidence that XP 12 BE cells are deficient in excision-repair of UV-damage to the DNA. Since FL/A cells appear to dem- onstrate a similar ability to survive UV irradiation compared to normal skin cells, and because they are relatively easy to culture, they were used in this s tudy where a large number of ceils were required, in preference to adult skin fibroblasts.
Treatment of the two cell lines with cis Pt[II] for 1 h resulted in the survival curves shown in Fig. 2. Whereas FL/A cells showed a Do dose of 6.4 pM cis Pt [II], the survival curve for XP 12 BE cells had a Do dose of 4.4 pM. If these results are expressed as the ratio
D 0 value of FL/A cells D0 value of XP 12 BE cells
they can be directly compared to results obtained for UV irradiation and other chemicals. A value of 1.45 was obtained for cis Pt[II] compared to 5.6 for UV irradiation. This result indicated that cis Pt[II] might no t be recognized by the DNA-excision-repair mechanism deficient in XP 12 BE and so lead to only a small decrease in the Do value in the absence of this repair system. To establish
1 2 6
Relat~e Sensitivities of Normal and Excision Deficient Human Celts to UV Irradiation.
-~ 100 l
.-- 10-
o .1= § g g e~
0"1 1'0 1~5
Joules/rn 2
Relative Sensitivities of N~mal & Excision 0eficient Human CeUs b ~ ?flit] Treatment
100.
10
1-
0ti t 10 2'0 3'0 gO
UH ~ Pt[Hlllhrtreatment)
Fig. 1 . T h e re la t ive sensi t iv i t ies o f n o r m a l ( F L / A ) (D), a n d e x c i s i o n - d e f i c i e n t ( X P 1 2 ) (o) , h u m a n f i b r o b l a s t s t o U V i r r a d a t i o n .
Fig. 2. The re la t ive sens i t iv i t ies o f n o r m a l ( F L / A ) (=), a n d e x c i s i o n - d e f i c i e n t ( X P 1 2 B E ) (o) h u m a n f ib ro - b l a s t s t o cis P t [ I I ] t r e a t m e n t f o r 1 h . The ver t i ca l ba r s i n d i c a t e t h e s t a n d a r d e r r o r o f t he m e a n f r o m
3 e x p e r i m e n t s .
0.03.
0~2- "5
Relative Binding of ~ Ptlo]to DNA Compared With the Dose d Treah~ent-
Retative Bindhg of Cis Ptnl]to DNA Compared to Toxicity
100-
10-
-
.>
o 001-
001-
lb zb ~ ~ 2~ pl'l ~ Ptr.](1 he treatmenfl n HoLes Pt/g ONA
Fig. 3. The relat ive extent o f react ion of c/~ P t [ I I ] w i th DNA compared w i th the dose of t reatment (1 h). F L / A , =; X P I 2 B E , o.
Fig. 4 . T h e re la t ive e x t e n t o f r e a c t i o n w i t h D N A c o m p a r e d t o t o x i c i t y . F L / A , m; X P 1 2 , o.
106 M~ ,
10-
127
107 I
5-
10"
o~
108 I
Fraction No.
MW
10"
I S"
r~ r~ C~ C~ 2o-
10-
106 107 108 I I I
([ _ t ~.~ Mn= 3,1xI06 d CON / % I0 Jm'2UV °-'°
1 *ENZYME
' s e d ~ ~ ' s ' ' (O) dR Mn=22x10 d CON [ \ - x 7 16Jm'2UV o-o / _ _ ~ ~ ~ I ~ _ M n 2 7 10 d ENZYME
s e d ~ 10 20 30 Fraction no.
-10
5 ! "r
fl: C5 m ro
-20 ~_ o~
"10
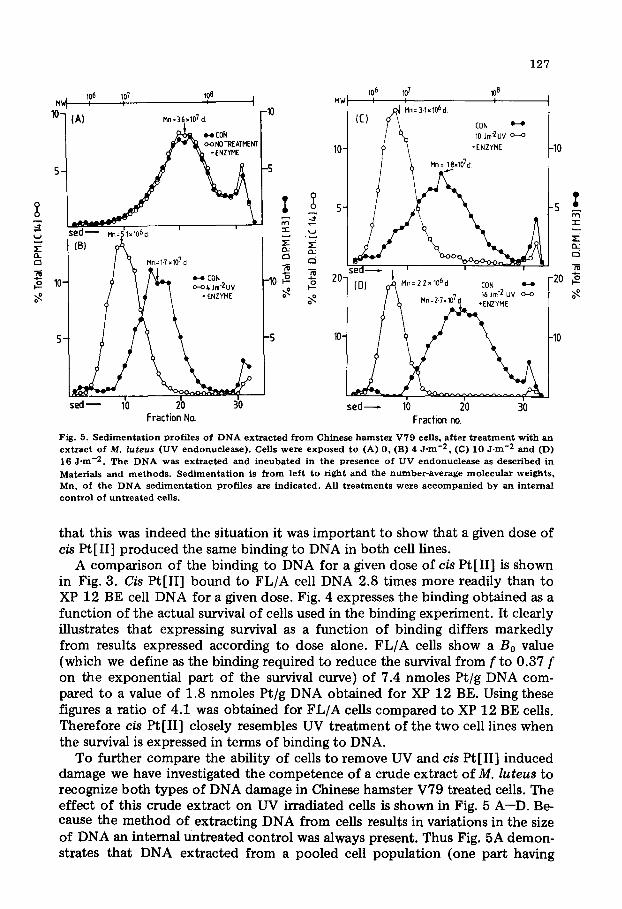
Fig. 5. S e d i m e n t a t i o n prof i les o f D N A e x t r a c t e d f r o m Chinese h a m s t e r V79 ceils, a f t e r t r e a t m e n t w i th an ex t r ac t of M. luteus (UV endonue lease ) . Cells we re exposed to (A) 0, (B) 4 J -m -2 , (C) 10 J . m -2 and (D) 16 J ' m -2 . The D N A was e x t r a c t e d a nd i n c u b a t e d in the presence of UV endonuc l ea se as descr ibed in Materials and m e t h o d s . S e d i m e n t a t i o n is f r o m lef t t o r ight and the n u m b e r - a v e r a g e m o l e c u l a r weights , Mn, of the D N A s e d i m e n t a t i o n prof i les are ind ica ted . All t r e a t m e n t s were a c c o m p a n i e d b y an in te rna l c o n t r o l of u n t r e a t e d cells.
that this was indeed the situation it was important to show that a given dose of cis Pt[II] produced the same binding to DNA in both cell lines.
A comparison of the binding to DNA for a given dose of cis Pt[II] is shown in Fig. 3. Cis Pt[II] bound to FL/A cell DNA 2.8 times more readily than to XP 12 BE cell DNA for a given dose. Fig. 4 expresses the binding obtained as a function of the actual survival of cells used in the binding experiment. It clearly illustrates that expressing survival as a function of binding differs markedly from results expressed according to dose alone. FL/A cells show a B0 value (which we define as the binding required to reduce the survival from f to 0.37 f on the exponential part of the survival curve) of 7.4 nmoles Pt/g DNA com- pared to a value of 1.8 nmoles Pt/g DNA obtained for XP 12 BE. Using these figures a ratio of 4.1 was obtained for FL/A cells compared to XP 12 BE cells. Therefore cis Pt[II] closely resembles UV treatment of the two cell lines when the survival is expressed in terms of binding to DNA.
To further compare the ability of cells to remove UV and cis Pt[II] induced damage we have investigated the competence of a crude extract o fM. luteus to recognize both types of DNA damage in Chinese hamster V79 treated cells. The effect of this crude extract on UV irradiated cells is shown in Fig. 5 A--D. Be- cause the method of extracting DNA from cells results in variations in the size of DNA an internal untreated control was always present. Thus Fig. 5A demon- strates that DNA extracted from a pooled cell population (one part having
1 2 8
50. c
o
\ t~
t~
= 2S"
o
/ o /
o 1'o io UV Dose(Jm "2)
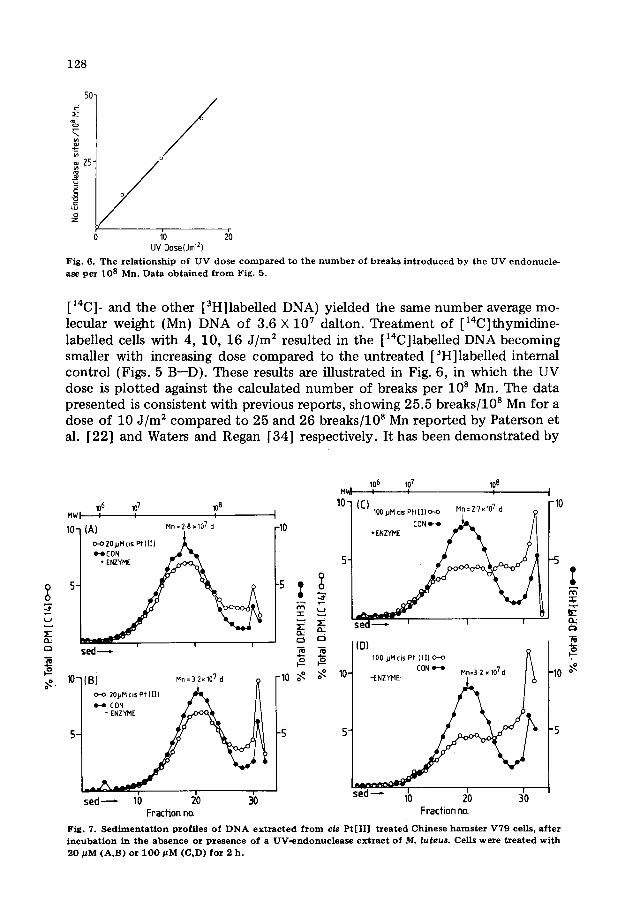
Fig. 6. The r e l a t i o n s h i p o f U V d o s e c o m p a r e d t o t h e n u m b e r of b r e a k s i n t r o d u c e d b y the U V e n d o n u c l e - ase p e r 108 Mn. D a t a o b t a i n e d f r o m Fig. 5.
[14C]- and the other [3H]labelled DNA) yielded the same number average mo- lecular weight (Mn) DNA of 3.6 X 107 dalton. Treatment of [14C]thymidine- labelled cells with 4, 10, 16 J /m 2 resulted in the [14C]labelled DNA becoming smaller with increasing dose compared to the untreated [~H]labelled internal control (Figs. 5 B--D). These results are illustrated in Fig. 6, in which the UV dose is plotted against the calculated number of breaks per 108 Mn. The data presented is consistent with previous reports, showing 25.5 breaks/10 s Mn for a dose of 10 J /m 2 compared to 25 and 26 breaks/10 s Mn reported by Paterson et al. [22] and Waters and Regan [34] respectively. It has been demonstrated by
106 107 108 M'~I i i i
106 107 10 s 10- (C) lOOpMcis PtlII)O-O Mn=27x107 d Q Mwl ! i I
10 ~ (A) Mn : 2 8 xl07 d 10 ,,ENZYME CONH ~ / /
s e d - - ' i ,
s e d - - ' (D)
10- (B) M°=32,1o7 d
A - ENZYME
;ed ~ 10 20 30
-10 o'-? o'-? 10-
100 uMcis Pt l ID o-o
-ENZYME CON H Mn=32 x 107 d I t ~
-S 5
s e d - - - 1~) iO 3*0
"10
"5
• 10 ~
Fraction no. Fraction no. Fig. 7. S e d i m e n t a t i o n p ro f i l e s o f D N A e x t r a c t e d f r o m cis P t [ I I ] t r e a t e d C h i n e s e h a m s t e r V 7 9 cells, af ter i n c u b a t i o n in t h e a b s e n c e o r p r e s e n c e o f a U V - e n d o n u c l e a s e e x t r a c t o f M. luteus . Cel l s w e r e t r e a t e d w i t h 2 0 MM (A,B) o r 1 0 0 / ~ M (C,D) f o r 2 h .

129
these authors that this represented a recognition of 100% of the UV-induced thymine dimers. Clearly the crude M. luteus extract used by us is able to recog- nise UV-induced DNA damage.
We then substi tuted cis Pt[II] for UV treatment. Fig. 7 A--B and C--D com- pare the size of DNA obtained in the absence and presence of t reatment with the M. luteus extract. Clearly the peak DNA Mn in both 20 pM and 100 pM cis Pt[II] treated cells is not altered after incubation with the M. luteus extract. However, since cis Pt[II] causes inter DNA-strand cross-links the size of the DNA was greater than that in the untreated internal controls. Treatment with 20 pM and 100 pM cis Pt[II] for 2 h would have resulted in approximately 4 and 10 lesions per l 0 s Mn DNA respectively [32]. If only a small proport ion of these had been recognized this should have resulted in a decrease in the value of Mn. However, since this was not the case we conclude that cis Pt[II] lesions are not recognized by the M. luteus extract.
Discussion
The similar relationship between the dose of cis Pt[II] and the survival of either foetal lung or XP cells led us initially to conclude that cis Pt[II] was related to the class of agents, proposed by Regan and Setlow [24], which invoke normal DNA-repair synthesis in XP cells. Certainly the cis Pt[II] survival curve for XP 12 BE was inconsistent with the demonstrat ion of increased toxic- ity of a number of agents in this cell line [5,17,28,30]. However, a comparison of the extent of reaction of cis Pt[II] with DNA in these two cells revealed a considerable difference in binding for a given dose. Thus when the extent of reaction of cis Pt[II] with DNA was compared to the survival obtained, a marked difference in sensitivity between the two cell lines was apparent. This result demonstrates the importance of considering binding vs. survival data when comparing the action of cis Pt[II] on more than one cell line.
XP 12 BE cells belong to complimentat ion group A and as such have been shown to display a partial defect in post-replication repair capacity [16] as well as a marked deficiency in excision-repair of various agents (as measured by repair-replication, unscheduled DNA synthesis, and kinetics of adduct removal from DNA). However, XP variants, which are characteristically deficient in post- replication repair and yet have a normal excision-repair capacity, do not demon- strate a large increase in toxici ty to both UV and chemical agents compared to normal cells [ 16,17]. It is reasonable to predict therefore that the binding versus survival results obtained for XP 12 BE and normal foetal lung cells after cis Pt[II] t reatment cannot be explained by a defect in post-replication repair alone. On this basis we assume that the majority of the increase in toxici ty of XP 12 BE cells following cis Pt[II] t reatment is due to a deficient excision-repair capac- ity. Further studies on the kinetics of removal of platinum lesions and possibly some estimation of repair synthesis are required to confirm this assumption.
Although XP cells are apparently more sensitive to cis Pt[II] t reatment com- pared to normal cells it does not necessarily follow that both UV and cis Pt[II] induced damage is repaired by the same pathway. Indeed, we have shown that an endonuclease, isolated from M. luteus, is able to recognise UV induced dam- age but not cis Pt[II] damage in DNA extracted from Chinese hamster V79

130
cells after t reatment with cis Pt[II] . A similar result has been obtained after t reatment with 7-bromomethylbenz(a)anthracene (7-BMBA) in these cells (un- published data) and after N-acetoxy-2-acetylaminofluorene (AAAF) treatment of normal human fibroblasts [1 ]. It is clearly not possible to draw firm conclu- sions with regard to repair processes in mammalian cells from studies using a crude microbial enzyme extract. Nevertheless these observations could indicate that if the chemically-induced lesions are excised by a similar mechanism to that which excises UV-induced thymine dimers then a different endonu- clease(s) is required to incise the chemically modified DNA in the first step of any excision-repair process. Alternatively, the failure of the UV endonuclease to recognize chemically induced damage could indicate that such lesions are removed from DNA by a quite distinct excision process. These alternatives are consistent with a number of lines of evidence. Dipple and Roberts [8] have found that rodent cells and Hela cells are able to excise 7-BMBA induced dam- age from their DNA at approximately the same rate, whereas by contrast it has been shown that UV-induced thymine dimers are not removed, or only slowly removed, by rodent cells [7,23]. Ikenaga et al. [12] demonstrated that the removal of the various adducts of 4-nitroquinoline-l-oxide bound to cellular DNA was comparable in human and mouse cells. Similarly, Amacher et al. [2] were able to show that whereas mouse cells were able to remove AAAF-induced lesions from DNA, they only removed relatively few thymine dimers. In addition the kinetics of thymine dimer removal were markedly different to those for AAAF products indicating that the two agents were removed by different path- ways. Following a dose of UV irradiation which saturated unscheduled DNA syn- thesis Ahmed and Setlow [1] found that subsequent t reatment with AAAF fur- ther increased the amount of induced unscheduled DNA synthesis, and the extent of repair-replication was additive. Finally, the relative sensitivities and excision-repair capacities of a number of XP cell lines to UV-irradiation induced damage did not parallel their sensitivities and excision-repair of 7-BMBA induced damage [19]. Clearly then, the UV-thymine dimer excision repair may differ from that involved in the excision of a whole spectrum of chemically induced damage.
The above interpretation of the data also implies that if a separate process is involved in the removal of cis Pt[II] induced damage, from that required for the removal of thymine dimers then that process is also deficient in XP 12 BE cells. Hence XP 12 BE could therefore be deficient in a number of different repair pathways. Thus, in addition to being deficient in thymine dimers exci- sion-repair, XP cells have been shown to lack ability to remove DNA protein cross-links [ 10], to have an altered apurinic DNA endonuclease activity [ 15] and to have low levels of photoreactivating enzyme [29].
Acknowledgements
We are grateful to the Rustenberg Platinum Mines Ltd. and Johnson Matthey Research Center for their generous support; and for grants from the Medical Research Council and the Cancer Research Campaign. Our thanks to Susan Young for technical assistance.

131
References
1 A h m e d , F.E. and R.B. Se t low, Dif fe ren t ra te l imi t ing steps in excis ion repa i r of u l t raviole t - an d N-ace- t o x y - 2 - a c e t y l a m i n o f l u o r e n e - d a m a g e d DNA in n o r m a l h u m a n f ibroblasts , Proc. Natl . Acad . Sci.
(U.S.A.) , 74 ( 1 9 7 7 ) 1 5 4 8 - - 1 5 5 2 . 2 A m a c h e r , D.E. , J .A . El l iot t a nd M.W. L i e b e r m a n , Di f fe rences in r e m o v a l of a c e t y l a m i n o f l u o r e n e an d
p y r i m i d i n e d imer s f r o m the DNA of cu l t u r ed m a m m a l i a n cells, Proc. Natl. Acad . Sci. (U.S.A.) , 74
( 1 9 7 7 ) 1 5 5 3 - - 1 5 5 7 . 3 Beck, D.J. and R .R. Brubaker , Effec t of cis p l a t i n u m [ I I ] d i a m m i n o d ichlor ide on wild t y p e an d D N A
repa i r def ic ien t m u t a n t s of E. coli, J. Bacteriol . , 116 (1973) 1 2 4 7 - - 1 2 5 2 . 4 Carrier, W.L. and R.B. Set low, Endonuc lease f r o m Micrococcus luteus wh ich has ac t iv i ty t o w a r d s UV
i r rad ia ted DNA: pur i f i ca t ion a nd p roper t i e s , J. Bacter ioL, 102 (1970) 1 7 8 - - 1 8 6 . 5 Cleaver, J .E. and D. Boo t sma , X e r o d e r m a p i g m e n t o s u m : Biochemica l an d genet ic character is t ics , Ann .
Rev. Genet . , 9 ( 1 9 7 5 ) 19- -38 . 6 Cleaver, J .E. , X e r o d e r m a p i g m e n t o s u m a h u m a n disease in which an initial stage of D N A repai r is
defec t ive , Proc. Natl . Acad . Sci. (U.S.A.) , 63 (1969) 4 2 8 - - 4 3 5 . 7 Cleaver, J .E. , Repa i r processes for p h o t o c h e m i c a l d a m a g e in m a m m a l i a n cells, in: J .T. L e t t et al.
(Eds.) , Advances in Ra d i a t i on Biology, Vol. 4, A c a d e m i c Press, New York , 1974 , pp . 1- -75 . 8 Dipple, A. a n d J.J . Rober t s , Excis ion of 7 - b r o m o m e t h y l b e n z [ a ] a n t h r a c e n e - - D N A ad d u c t s in repli-
ca t ing m a m m a l i a n cells, B iochemis t ry , 16 ( 1 9 7 7 ) 1 4 9 9 - - 1 5 0 3 . 9 Drobnfk , J., M. Urb~inkov~ a nd A. Krekulovd , The e f fec t of c i s -d ich lorod iamminep la t inum[I I ] on
Escherichia coli B, The role of fil, exr a nd hcr marke r s , Mu ta t i on Res. , 17 ( 1 9 7 3 ) 13 - -20 . 10 Fornace , A.J. and K.W. K o h n , D N A - - p r o t e i n cross-l inking b y UV rad ia t ion in n o r m a l h u m a n an d
x e r o d e r m a p i g m e n t o s u m fibroblasts , Biochim. Biophys. Acta , 435 ( 1 9 7 6 ) 9 5 - - 1 0 3 . 11 Harde r , H.C., R .G. Smi th a nd A.F. L e r o y , T e m p l a t e - p r i m e r inac t iva t ion b y c i s -a n d trans-dichlorodi-
a m m i n e p l a t i n u m for h u m a n D N A p o l y m e r a s e ~1 fl and Rausche r m u r i n e l e u k a e m i a virus reverse t ranscr ip tase , as a m e c h a n i s m of c y t o t o x i c i t y , Cancer Res., 36 (1976) 3 8 2 1 - - 3 8 2 9 .
12 Ikenaga , M., H. T a k e b e and Y. Ishii, Excis ion repai r of DNA base d a m a g e in h u m a n cells t r ea t ed w i th the chemicM ca rc inogen 4 -n i t roqu ino l ine - l -ox ide , M u t a t i o n Res. , 43 (1977) 4 1 5 - - 4 2 7 .
13 Ki rby , K.S. and E.A. Cook , I so la t ion of DNA f r o m m a m m a l i a n tissues, B iochem. J. , 104 (1967) 2 5 4 - - 257.
14 K r a e m e r , K.H. , A.P. Andrews , S.F. Bar re t t and J .H. Robbins , Co lony - fo rming abi l i ty of ul t raviolet- i r - r ad ia t ed X e r o d e r m a p i g m e n t o s u m f ibroblas ts f r o m d i f fe ren t DNA-repa l r c o m p l e m e n t a t i o n groups , Biochim. Biophys. Acta , 442 (1976) 147 - -153 .
15 Kuhn le in , U., E.E. P e n h o e t a nd S. L inn , An a l te red apur in ic D N A endonuc lease ac t iv i ty in g roup A and group D X e r o d e r m a p i g m e n t o s u m fibroblasts , Proc. Natl . Acad. Sci. (U.S.A.) , 73 (1976) 1 1 6 9 - - 1 1 7 3 .
16 L e h m a n n , A.R. , S. Kirk-Bell, C.F. Ar ie t t , S.A. Ha rc ou r t , E.A. de Weerd-Kaste le in , W. Keijzer an d P. Hal l -Smith , Repa i r of UV light d a m a g e in a va r i e ty of h u m a n f ibroblas t cell strains, Cancer Res., 37 (1977) 9 0 4 - - 9 1 0 .
17 Maher , V.M., J .J . McCormick , P.L. Grove r and P. Sims, Effec ts of DNA repa i r and the c y t o t o x i c i t y and m u t a g e n i c i t y of po lyeyc l ic h y d r o c a r b o n der ivat ives in n o r m a l and X e r o d e r m a p i g m e n t o s u m hu- m a n f ibroblasts , Mu ta t i on Res., 43 (1977) 1 1 7 - - 1 3 8 .
18 Mansy, S., B. Rosenbe rg a nd A.J. T h o m s o n , Binding of cis an d trans d i c h l o r o d i a m m i n e p l a t i n u m to nucleosides , 1. L o c a t i o n of the b ind ing sites, J. Am. Chem. Soc., 95 (1973) 1 6 3 3 - - 1 6 4 0 .
19 McCaw, B.A., A. Dipple , S. Young and J .J . Robe r t s , Excis ion of h y d r o c a r b o n - D N A ad d u c t s and con- s equen t cell survival of n o r m a l a nd repa i r -defec t ive h u m a n cells, Chem.-Biol . In te rac t . , In press.
20 Morris, C.R. and G.R. Gale, I n t e r a c t i o n s of an a n t i t u m o r p l a t i n u m c o m p o u n d wi th d e o x y r i b o n u c l e i c acid, h is tones , L -amino acids, po ly -L-amino acids, nucleos ides and nuc leo t ides , Chem.-Biol . In te rac t . , 7 ( 1 9 7 3 ) 3 0 5 - - 3 1 5 .
21 Munchausen , L .L. , The chemica l a nd biological e f fec ts of cis d i c h l o r o d i a m m i n e p l a t i n u m [ I I ] an anti- t u m o r agent , on DNA, Proe. Natl . Acad . Sci. (U.S.A.) , 71 ( 1 9 7 4 ) 4 5 1 9 - - 4 5 2 2 .
22 Pa terson , M.C., P.H.M. L o h m a n a n d M.L. S luy te r , Use of a UV endonuc lease f r o m Micrococeus luteus to m o n i t o r the progress of D N A repa i r on UV i r rad ia ted h u m a n cells, Mu ta t i on Res., 19 (1973) 2 4 5 - - 256.
23 Regan , J .D. , J .E. T rosko a nd W.L. Carrier, Ev idence for excis ion of UV induced p y r i m i d i n e d imer s f rom the D N A of h u m a n cells in v i t ro , Biophys . J., 8 ( 1 9 6 8 ) 3 1 9 - - 3 2 5 .
24 Regan, J .D. and R.B. Se t low, T w o f o r m s of repa i r in the D N A of h u m a n cells d a m a g e d by chemica l carc inogens and m u t a g e n s , Cancer Res., 34 ( 1 9 7 4 ) 3 3 1 8 - - 3 3 2 5 .
25 Rober t s , J .J . , J .M. Pascoe, J .E. Plant , J .E. S t u r r o c k a nd A.R. Cra tho rn , Quan t i t a t i ve aspects of the repai r of a lky i a t ed D N A in cu l tu red m a m m a l i a n cells, 1. The e f fec t of H e L a and Chinese h a m s t e r cell survival of a lky la t ion of cellular m a c r o m o l e c u l e s , Chem.-Biol . In te rac t . , 3 ( 1 9 7 1 ) 29- -47 .
26 Rober t s , J .J . , F. Friedlos, H.W. van den Berg a nd D.J. Kixkland, Inh ib i t ion by caffe ine of post-repli- ca t ion repa i r in Chinese h a m s t e r cells t r e a t e d w i th 7 - h r o m o m e t h y l b e n z [ a ] a n t h r a c e n e : e n h a n c e m e n t of

1 3 2
t o x i c i t y , c h r o m o s o m e d a m a g e a n d i n h i b i t i o n of l i ga t i on o f n e w l y - s y n t h e s i s e d D N A , Chem. -B io l . I n t e r ac t . , 17 ( 1 9 7 7 ) 2 6 5 - - 2 9 0 .
27 R o z e n z w e i g , M., D.D. y o n H o f f , J .S . P e n t a a n d F.M. Muggia , Cl in ica l s t a t u s o f cis d i a m m i n e d i eh lo ro - p l a t i n u m [ I I ] , J . Clin, H e m a t o l . Onco l . , 7 ( 1 9 7 7 ) 6 7 2 - - 6 8 0 .
2 8 S t i ch , H . F . , R . H . C . San a n d Y. K a w a z o e , I n c r e a s e d sens i t iv i ty o f X e r o d e r m a p i g m e n t o s u m cells to s o m e c h e m i c a l c a r c i n o g e n s a n d m u t a g e n s , M u t a t i o n Res . , 17 ( 1 9 7 3 ) 1 2 7 - - 1 3 7 .
29 S u t h e r l a n d , B.M,, M. R ice a n d E .K. Wagner , X e r o d e r m a p i g m e n t o s u m cells c o n t a i n l o w levels o f p h o - t o r e a c t i v a t i n g e n z y m e , P roc . Nai l . A c a d . Sci. (U.S .A. ) , 7 2 ( 1 9 7 5 ) 1 0 3 - - 1 0 7 .
3 0 T a k e b e , H. , J . F u r u y a m a , Y. Miki a n d S. K o n d o , H igh sens i t iv i ty o f X e r o d e r m a p i g m e n t o s u m cells t o t he c a r c i n o g e n 4 - r d t r o q u i n o l i n e - l - o x i d e , M u t a t i o n Res . , 15 ( 1 9 7 2 ) 9 8 - - 1 0 0 .
31 V a n d e n Berg , H.W. a n d J . J . R o b e r t s , I n v e s t i g a t i o n i n t o the m e c h a n i s m of a c t i o n o f a n t i t u m o u r pla t i - n u m c o m p o u n d s : T ime- a n d dose - d e p e n d e n t c h a n g e s in t he a lka l ine suc rose g r a d i e n t s e d i m e n t a t i o n p ro f i l e s o f D N A f r o m h a m s t e r cells t r e a t e d w i t h cis p l a t i n u m d i a m m i n e d i c h l o r i d e , Chem. -B io l . In te r - ac t . , 11 ( 1 9 7 5 ) 4 9 3 - - 4 9 9 .
3 2 V a n d e n Berg, H.W. a n d J . J . R o b e r t s , I n h i b i t i o n b y c a f f e i n e o f p o s t - r e p l i c a t i o n r e p a i r in Chinese h a m - s t e r cells t r e a t e d w i th c/s p l a t i n u m [ I I ] d i a m m i n e d i ch lo r ide : t h e e x t e n t o f p l a t i n u m b i n d i n g t o t e m - p l a t e D N A in r e l a t i o n t o t h e size o f l o w m o l e c u l a r w e i g h t n a s c e n t D N A , Chem. -B io l . I n t e r ac t . , 12 ( 1 9 7 6 ) 3 7 5 - - 3 9 0 .
3 3 V a n d e n Berg, H.W., H . N . A . F rava l a n d J . J . R o b e r t s , R e p a i r o f D N A d a m a g e d b y n e u t r a l p l a t i n u m c o m p l e x e s , J . Clin. H e m a t o l . Oneo l . , 7 ( 1 9 7 7 ) 3 4 9 - - 3 7 3 .
3 4 Waters , R . a n d J .D. R e g a n , R e c o m b i n a t i o n o f U V - i n d u c e d p y r i m i d i n e d i m e r s in h u m a n f ib rob la s t s , B i o c h e m . B i o p h y s . Res. C o m m u n . , 7 2 ( 1 9 7 6 ) 8 0 3 - - 8 0 7 .
3 5 Wi l t shaw, E. a n d T. K r o n e r , Phase II s t u d y o f cis d i c h l o r o d i a m m i n e p l a t i n u m [ I I ] CNS 1 1 9 8 7 5 in a d v a n c e d a d e n o c a r c i n o m a of t h e o v a r y , C a n c e r T r e a t m e n t R e p o r t s , 6 0 ( 1 9 7 6 ) 5 5 - - 6 0 .
36 Wit tes , R .E. , E. Cv i tkov ic , I .H. K r a k o f f a n d E.W. S t r o n g , The ro le o f cis d i a m m i n e d i c h l o r o p l a t i n u m [ I I ] in t he t r e a t m e n t of h e a d a n d n e c k c a n c e r , J . Cl in ica l H e m a t o l . Oneo l . , 7 ( 1 9 7 7 ) 7 1 1 - - 7 1 6 .
Related Documents











