Summary We estimated nitrogen (N) use by trees of three poplar species exposed for 3 years to free air CO 2 enrichment (FACE) and determined whether the CO 2 treatment affected the future N availability of the plantation. Trees were harvested at the end of the first 3-year rotation and N concentration and content of woody tissues determined. Nitrogen uptake of fine roots and litter was measured throughout the first crop rotation. The results were related to previously published variations in soil N content during the same period. We estimated retrans- location from green leaves and processes determining N mobi- lization and immobilization, such as mineralization and nitrifi- cation, and N immobilization in litter and microbial biomass. In all species, elevated CO 2 concentration ([CO 2 ]) significantly increased nitrogen-use efficiency (NUE; net primary produc- tivity per unit of annual N uptake), decreased N concentration in most plant tissues, but did not significantly change cumula- tive N uptake by trees over the rotation. Total soil N was de- pleted more in elevated [CO 2 ] than in ambient [CO 2 ], although not significantly for all soil layers. The effect of elevated [CO 2 ] was usually similar for all species, although differences among species were sometimes significant. During the first 3-year ro- tation, productivity of the plantation remained high in the ele- vated [CO 2 ] treatment. However, we observed a potential re- duction in N availability in response to elevated [CO 2 ]. Keywords: nitrogen, FACE, Populus, SRF, NUE, N uptake. Introduction Growth of trees is often largely regulated by nitrogen (N) availability. When plants are exposed to elevated atmospheric CO 2 concentration ([CO 2 ]), they often exhibit enhanced growth with increased biomass or net primary productivity (NPP), or both ( King et al. 2005, Norby et al. 2005), which, in the long-term, can lead to increased N uptake from the soil. Ni- trogen and carbon (C) cycles are closely coordinated in plants, and N metabolism is regulated by signals that are derived from C metabolism (Stitt and Krapp 1999). Elevated [CO 2 ] usually decreases N concentrations in all plant tissues (Wong 1979, Cotrufo et al. 1998, Johnson 2006), although this decrease can be the result of an increase in nonstructural carbohydrates, mainly starch, which usually in- creases in response to elevated [CO 2 ] (Wong 1979, Stitt and Krapp 1999). However, in some cases, reduced N concentra- tion may be observed in response to elevated [CO 2 ] even after correcting for increased carbohydrates (Poorter et al. 1997, Tissue et al. 1997). Lowered plant tissue N concentration may be caused by depletion of available N in the soil as a conse- quence of enhanced growth in elevated [CO 2 ] ( Bazzaz 1990). Short-term experiments indicate that soil N availability can decrease, increase or exhibit no significant change beneath temperate trees grown in an atmosphere with an elevated [CO 2 ] ( Zak et al. 2000). Changes in plant litter production and quality in response to elevated [CO 2 ] could alter microbial N demand and the amounts of N available for plant uptake (Zak et al. 2003, Chapman et al. 2005). Microbial biomass has an Tree Physiology 27, 1153–1163 © 2007 Heron Publishing—Victoria, Canada Increased nitrogen-use efficiency of a short-rotation poplar plantation in elevated CO 2 concentration CARLO CALFAPIETRA, 1,2 PAOLO DE ANGELIS, 1 BIRGIT GIELEN, 3 MARTIN LUKAC, 4 MARIA CRISTINA MOSCATELLI, 5 GIUSEPPINA AVINO, 1 ALESSANDRA LAGOMARSINO, 1 ANDREA POLLE, 6 REINHART CEULEMANS, 3 GIUSEPPE SCARASCIA MUGNOZZA, 1,7 MARCEL R. HOOSBEEK 8 and MARIA FRANCESCA COTRUFO 9 1 University of Tuscia, Department of Forest Environment and Resources (DISAFRI), Via S. Camillo De Lellis, I-01100 Viterbo, Italy 2 Corresponding author ([email protected]) 3 University of Antwerpen, Campus Drie Eiken, Department of Biology, Research Group of Plant and Vegetation Ecology, Universiteitsplein 1, B-2610 Wilrijk, Belgium 4 University of Wales, School of Agricultural and Forest Sciences, Bangor, Gwynedd LL57 2UW, United Kingdom 5 University of Tuscia, Soil Biochemistry Laboratory, Department of Agrobiology and Agrochemistry, Via S. Camillo De Lellis, I-01100 Viterbo, Italy 6 Institut für Forstbotanik, Georg-August Universität, Büsgenweg 2, 37077 Göttingen, Germany 7 Institute of Agro-Environmental and Forest Biology – (IBAF-CNR), Via G. Marconi, 2 “Villa Paolina,” 05010 Porano (TR), Italy 8 Wageningen University, Department of Environmental Sciences, Earth System Science Group, P.O. Box 37, 6700 AA Wageningen, The Netherlands 9 Second University of Naples, Department of Environmental Sciences, Via Vivaldi 43, I-81100 Caserta, Italy Received July 31, 2006; accepted October 23, 2006; published online May 1, 2007 by guest on May 15, 2011 treephys.oxfordjournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Summary We estimated nitrogen (N) use by trees of threepoplar species exposed for 3 years to free air CO2 enrichment(FACE) and determined whether the CO2 treatment affectedthe future N availability of the plantation. Trees were harvestedat the end of the first 3-year rotation and N concentration andcontent of woody tissues determined. Nitrogen uptake of fineroots and litter was measured throughout the first crop rotation.The results were related to previously published variations insoil N content during the same period. We estimated retrans-location from green leaves and processes determining N mobi-lization and immobilization, such as mineralization and nitrifi-cation, and N immobilization in litter and microbial biomass.In all species, elevated CO2 concentration ([CO2]) significantlyincreased nitrogen-use efficiency (NUE; net primary produc-tivity per unit of annual N uptake), decreased N concentrationin most plant tissues, but did not significantly change cumula-tive N uptake by trees over the rotation. Total soil N was de-pleted more in elevated [CO2] than in ambient [CO2], althoughnot significantly for all soil layers. The effect of elevated [CO2]was usually similar for all species, although differences amongspecies were sometimes significant. During the first 3-year ro-tation, productivity of the plantation remained high in the ele-vated [CO2] treatment. However, we observed a potential re-duction in N availability in response to elevated [CO2].
Keywords: nitrogen, FACE, Populus, SRF, NUE, N uptake.
Introduction
Growth of trees is often largely regulated by nitrogen (N)availability. When plants are exposed to elevated atmosphericCO2 concentration ([CO2]), they often exhibit enhancedgrowth with increased biomass or net primary productivity(NPP), or both (King et al. 2005, Norby et al. 2005), which, inthe long-term, can lead to increased N uptake from the soil. Ni-trogen and carbon (C) cycles are closely coordinated in plants,and N metabolism is regulated by signals that are derived fromC metabolism (Stitt and Krapp 1999).
Elevated [CO2] usually decreases N concentrations in allplant tissues (Wong 1979, Cotrufo et al. 1998, Johnson 2006),although this decrease can be the result of an increase innonstructural carbohydrates, mainly starch, which usually in-creases in response to elevated [CO2] (Wong 1979, Stitt andKrapp 1999). However, in some cases, reduced N concentra-tion may be observed in response to elevated [CO2] even aftercorrecting for increased carbohydrates (Poorter et al. 1997,Tissue et al. 1997). Lowered plant tissue N concentration maybe caused by depletion of available N in the soil as a conse-quence of enhanced growth in elevated [CO2] (Bazzaz 1990).
Short-term experiments indicate that soil N availability candecrease, increase or exhibit no significant change beneathtemperate trees grown in an atmosphere with an elevated[CO2] (Zak et al. 2000). Changes in plant litter production andquality in response to elevated [CO2] could alter microbial Ndemand and the amounts of N available for plant uptake (Zaket al. 2003, Chapman et al. 2005). Microbial biomass has an
Tree Physiology 27, 1153–1163© 2007 Heron Publishing—Victoria, Canada
Increased nitrogen-use efficiency of a short-rotation poplar plantationin elevated CO2 concentration
CARLO CALFAPIETRA,1,2 PAOLO DE ANGELIS,1 BIRGIT GIELEN,3 MARTIN LUKAC,4
MARIA CRISTINA MOSCATELLI,5 GIUSEPPINA AVINO,1 ALESSANDRA LAGOMARSINO,1
ANDREA POLLE,6 REINHART CEULEMANS,3 GIUSEPPE SCARASCIA MUGNOZZA,1,7
MARCEL R. HOOSBEEK8 and MARIA FRANCESCA COTRUFO9
1 University of Tuscia, Department of Forest Environment and Resources (DISAFRI), Via S. Camillo De Lellis, I-01100 Viterbo, Italy2 Corresponding author ([email protected])3 University of Antwerpen, Campus Drie Eiken, Department of Biology, Research Group of Plant and Vegetation Ecology, Universiteitsplein 1, B-2610
Wilrijk, Belgium4 University of Wales, School of Agricultural and Forest Sciences, Bangor, Gwynedd LL57 2UW, United Kingdom5 University of Tuscia, Soil Biochemistry Laboratory, Department of Agrobiology and Agrochemistry, Via S. CamilloDe Lellis, I-01100 Viterbo, Italy6 Institut für Forstbotanik, Georg-August Universität, Büsgenweg 2, 37077 Göttingen, Germany7 Institute of Agro-Environmental and Forest Biology – (IBAF-CNR), Via G. Marconi, 2 “Villa Paolina,” 05010 Porano (TR), Italy8 Wageningen University, Department of Environmental Sciences, Earth System Science Group, P.O. Box37, 6700 AA Wageningen, The Netherlands9 Second University of Naples, Department of Environmental Sciences, Via Vivaldi 43, I-81100 Caserta, Italy
Received July 31, 2006; accepted October 23, 2006; published online May 1, 2007
by guest on May 15, 2011
treephys.oxfordjournals.orgD
ownloaded from

important role in determining the availability of N to plants: anincrease in easily degradable C may stimulate microbial activ-ity and hence nutrient cycling. Elevated [CO2] caused a 10.7%reduction in N in leaf litter at the AspenFACE site (Liu et al.2005), and a 7.1% reduction in leaf litter N was found in ameta-analysis across different plants (Norby et al. 2001).Moreover, elevated [CO2] may cause a negative feedback onsoil N availability through reduced litter decomposition and Nmineralization (Gill et al. 2006). Soil N depletion, which is of-ten observed in long-term CO2 experiments (Luo et al. 2004),is often advocated as the reason why plants change their re-sponse to elevated [CO2] over time.
An understanding of N cycling is essential to the effectivemanagement of short-rotation forestry (SRF) plantationswhich have been introduced with the aim of producing bio-mass for energy and mitigating greenhouse gases. Most fre-quently, nutrient cycling in an SRF system is managed to pro-vide sustainable biomass production. Applying the rightamount of fertilizer is important to minimizing productioncosts and limiting off-site fertilizer pollution (Heilman andNorby 1998). Although there has been little research on long-term ecosystem-level sustainability of nutrient cycles and stor-age in agro-forestry systems (Isaac et al. 2005), there are indi-cations that the fertilizer requirements of well-established andhigh-yielding (> 12 Mg dry mass ha–1 year –1) plantations aresmall, about 30 kg N ha–1 year –1, because of efficient recy-cling of N from litter and the relatively low nutrient content inthe harvested biomass (stem) (Ericsson 1994). But, as sug-gested in the cited studies, increased atmospheric [CO2] canalter both N uptake by the trees and the mechanisms influenc-ing the availability of N in the soil, adding uncertainty to esti-mates of N fertilization requirements.
We investigated N use by three poplar species in a short-ro-tation plantation (POP-EUROFACE) exposed to a [CO2] of550 ppm for three years, from planting to full canopy closure.Free air CO2 enrichment (FACE) experiments offer the oppor-tunity to study the effects of elevated [CO2] on nutrient cyclingat the ecosystem level, including long-term feedbacks (John-son et al. 2001). Where available, we present data about pro-cesses that may help explain previous findings and possiblechanges in N cycling in response to elevated [CO2]. Three hy-potheses were tested: (1) elevated [CO2] stimulates plant pro-ductivity, leading to increased nitrogen-use efficiency (NUE)and to decreased N concentration in plant tissues; (2) soil Nstocks are reduced in elevated [CO2] as a result of reduced Ninput from litter fall; and (3) N immobilization is increased byelevated [CO2], leading to decreases in available N.
Materials and methods
Experimental site
The POP-EUROFACE experimental site is located in centralItaly close to Tuscania (42°37′ N, 11°51′ E, 150 m a.s.l). The9-ha poplar plantation was established in 1999 with hardwoodcuttings of Populus × euramericana at a planting density of2 × 1 m. Six experimental plots (30 × 30 m) were selected andplanted at a density of 1 × 1 m with three poplar genotypes(Scarascia Mugnozza et al. 2006): P. × euramericana Dode(Guinier) ( P. deltoides Bart. ex Marsh. × P. nigra L.) genotypeI-214, P. nigra L. (genotype Jean Pourtet) and P. alba L. (geno-type 2AS11) (for further information on genotype propertiessee Calfapietra et al. 2001). Three of the six experimental plotswere maintained at ambient [CO2] (~ 370 ppm) and three weremaintained at elevated [CO2] (~ 550 ppm) with the FACE tech-nique (Miglietta et al. 2001). Within the elevated [CO2] plots,mean [CO2] was 544 ± 48, 532 ± 83 and 554 ± 95 ppm duringthe three consecutive years of the study. Because the plantationwas established on a former agricultural field, the soil nutrientavailability at the beginning of the experiment was high (Ta-ble 1). The upper 20 cm of the topsoil had a silt loam textureand a pH (KCl) of 5.04 (mean for all the plots); the bulk den-sity was 1.34 g cm– 3. Further information on soil propertieshas been described by Hoosbeek et al. (2004).
Plant nitrogen assessments
The plantation was harvested at the end of the first 3-year rota-tion (i.e., at the end of the third growing season) in Novemberand December 2001 (Calfapietra et al. 2003). Four trees perspecies per plot were selected, representing the size variabilityof the population. Half of the harvested trees, two per speciesper plot, were selected for belowground assessment by exca-vating a 1 × 1 m area of soil up to 70–80 cm in depth. Allwoody components were oven dried for 5 days at 70 °C andweighed. The woody components were classified into: (1)three stem portions corresponding to the three annual heightgrowth increments (HGI); (2) sylleptic branches (developedfrom axillary buds not undergoing a winter rest period); (3)proleptic branches (developed from buds that have undergonea winter rest period); (4) stump (the portion of the stem stand-ing below ground from which the main roots originated); and(5) coarse roots (all roots with diameter > 2 mm). A 5-cm-longstem section was collected from each harvested sample and,after bark removal, ground to a fine powder for C and N assess-ments with a CHN elemental analyzer (NA1500, Carlo ErbaInstruments, Rodano, Italy).
1154 CALFAPIETRA ET AL.
TREE PHYSIOLOGY VOLUME 27, 2007
Table 1. Soil conditions in ambient (~370 ppm) and elevated (~ 550 ppm) CO2 concentration ([CO2]) experimental plots at the POP-EUROFACEsite in November 1998, before plantation establishment. Values are means (± SE) of n = 3 plots. Abbreviations: Ctotal, total carbon concentration;Ntotal, total nitrogen concentration; NO3-N, nitrate nitrogen; and NH4-N, ammonium nitrogen.
[CO2] treatment Ctotal Ntotal NO3-N NH4-N pH(%) (%) (µg N g–1 soil) (µg N g–1 soil)
Ambient 1.10 (0.02) 0.13 (0.00) 9.22 (0.76) 0.67 (0.05) 5.13 (0.09)Elevated 0.99 (0.07) 0.13 (0.01) 7.41 (0.65) 0.60 (0.08) 4.93 (0.03)
by guest on May 15, 2011
treephys.oxfordjournals.orgD
ownloaded from

Canopy N content was determined in August of the thirdyear, at the time of peak leaf area index (LAI). Calculationswere made for all species based on total LAI, LAI in each 1-mheight interval of the canopy and vertical profiles of leaf Ncontent and specific leaf area (SLA) as reported by Gielen etal. (2003).
During the second and the third growing seasons, litter pro-duction was monitored with leaf litter traps (Cotrufo et al.2005). Litter production of the first year was estimated foreach canopy sector by multiplying the peak values of LAI bythe mean SLA of the canopy. On a monthly basis and for eachyear, N concentration was determined on a subsample of leaflitter analyzed independently with a CHN elemental analyzer.Annual N input to soil via litter fall was calculated for the threegrowing seasons as the sum of each periodic collection of littermultiplied by the corresponding N concentration.
Fine root biomass was sampled throughout the first rotationcycle and in February 2002, shortly after harvesting of theplantation. Auger coring was carried out with an 8-cm-diame-ter corer at depths of 0–20 and 20–40 cm. Fine roots werewashed free of soil, dried at 60 °C for 5 days and weighed. Fineroot N concentration was determined as for the woodycomponents.
Nitrogen uptake, as defined by Cole and Rapp (1981), wascalculated as described by Johnson et al. (2004). By assumingthat fine root N is not resorbed into perennial tissue before rootdeath (Nambiar and Fife 1991), annual N uptake was calcu-lated for the whole plant as the sum of N content of annualwoody increment (stem, branches, stump and coarse roots),annual fine root production and annual litter fall. Nitrogenpools for the different tissues were calculated at the end of thefirst rotation by summing the N contents of the annualincrements.
The foliar leaching component (throughfall minus deposi-tion) was negligible relative to the quantity of N cycling be-tween the plant and the soil. It is usually considered to be lessthan 1% of the total annual N uptake in deciduous trees (Aertsand Chapin 2000). As reported in a previous paper, the pres-ence of insects and herbivores was insignificant (Gielen et al.2005).
Nitrogen-use efficiency
Nitrogen-use efficiency (NUE) was calculated for both thesecond and the third growing seasons of the first crop rotationby dividing annual net primary productivity (NPP) (Gielen etal. 2005) by annual N uptake of the whole plant (Berendse andAerts 1987). As reported by Berendse and Arts (1987), NUEcomprised two components: N productivity (AN) and meanresidence time (MRT):
AN
NPP
peak N content=
MRTpeak N content
annual N uptake=
Peak N content of the whole tree was calculated as the sum ofN in standing wood, N in leaves at peak canopy mass and N infine roots at peak fine root standing crop.
Soil nitrogen
Estimates of total soil N stocks are derived from Hoosbeek etal. (2004) and are reported here to provide a comprehensivepicture of the N pools at the ecosystem level.
Nitrogen dynamics
Mean N content for the canopy was estimated in August of thethird year as explained previously. The leaf biomass of thetrees in August represented about 75% of the mean total leafbiomass produced across treatments and species during thethird year. Total N contained in the green canopy in Augustwas estimated per square meter of ground. The mean value ofN concentration of leaf litter samples collected from Septem-ber onwards was used to estimate N retranslocation, takinginto account that green leaves standing in August started tosenesce and fall in September.
Retranslocation efficiency (RE) was calculated as (Wang etal. 2003):
REN N
Ncanopy litter
canopy
=[ ] – [ ]
[ ]
where [Ncanopy] is the nitrogen concentration in green canopyand [Nlitter] is the nitrogen concentration in leaf litter. This esti-mate may be biased because of mass loss during senescence(van Heerwaarden and Aerts 2003). The pool of N retrans-located from the green canopy was estimated from the totalstanding biomass of green leaves in August of the third yearand RE for each species and treatment.
Net N mineralization and nitrification were measured ac-cording to Eno (1960) during the second cycle. Two adjacentsoil cores (10 cm in diameter and 20 cm in depth) were col-lected in June and October of the third year (data of the cam-paigns were averaged) for each species in each plot. The firstcore (initial content) was transported to the laboratory within6 h of collection, sieved and nitrate and ammonium contentsmeasured as described by Cataldo et al. (1975) and Andersonand Ingram (1993), respectively. The second core (final con-tent) was placed in a polyethylene plastic bag to avoid waterentry and incubated in situ in the field for 1 month. After1 month it was removed from the field and treated in the sameway as the first core. Net N mineralization and net nitrificationwere calculated as the difference between final and initial con-tents of ammonium plus nitrate and only nitrate, respectively.The values in June and October were averaged to representfield conditions, thus including the two seasons that most in-fluence soil microbial activity in the plantation. In the field,soil water content ranged from 20 to 29% in June and from 32to 40% in October, with the highest values recorded at the endof the month (Probe TDR-TRIME; IMKO Micromodultech-nik GmbH, Ettlingen, Germany; 15-cm depth). Soil tempera-ture ranged from 18 to 23 °C in June and from 14 to 16 °C in
TREE PHYSIOLOGY ONLINE at http://heronpublishing.com
EFFECT OF ELEVATED [CO2] ON N CYCLING IN POPLAR 1155
by guest on May 15, 2011
treephys.oxfordjournals.orgD
ownloaded from

October (EGM-4; PP-Systems, Hertfordshire, U.K.).Litter decomposition was studied by the litterbag technique
(Cotrufo et al. 2005). Two litter-bag types were considered: (1)litter produced in ambient [CO2] plots and incubated in ambi-ent [CO2] plots; and (2) litter produced in elevated [CO2] plotsand incubated in elevated [CO2] plots.
Litter N immobilization was calculated as the difference be-tween the N content in litterbags at the beginning and at theend of 250 days of incubation. Extrapolation of N dynamics tothe plot scale was obtained by normalizing the net change in Ncontent per bag to grams of litter and multiplying it by total lit-ter production. Thus, positive values indicate a net microbialuptake of N from the soil to the litter layer, whereas negativevalues show net mineralization from the decomposing leaflitter.
To provide information on the possible mechanisms of im-mobilization of N in ambient and elevated [CO2], nitrogen inmicrobial biomass was estimated during the second crop rota-tion by averaging the campaigns in June and October. Quanti-fication of microbial biomass was provided by Moscatelli et al.(2005) following the fumigation extraction (FE) method(Vance et al. 1987). Microbial biomass N (MBN) was deter-mined as ninhydrin-reactive N in 0.5 M K2SO4 soil extracts(fumigated and non-fumigated) according to Joergensen andBrookes (1990).
Statistical analysis
Analysis of variance (ANOVA) was carried out to evaluate themain effects of elevated [CO2], species and their interaction onN pools and dynamics with a randomized complete-block de-sign with treatment, species and treatment × species interac-tion as fixed factors and block as a random factor. Data werefirst tested for normality by the Shapiro-Wilk statistic. Plot(3 ambient CO2 plots and 3 elevated CO2 plots) was the unit ofreplication. Where the ANOVA F test indicated an interactionbetween treatment and species, an a posteriori comparison ofmeans was performed. Bonferroni’s method was used for mul-tiple comparisons. Differences between means were consid-
ered significant at P < 0.05. The elevated [CO2] effect was pre-sented as an elevated [CO2]:ambient [CO2] (E:A) ratio.
Results
Plant N assessments
For the aboveground components, elevated [CO2] caused a de-crease in N concentration in the lower part of the stem (HGI 1and 2), proleptic branches, leaves and leaf litter (Table 2). Ni-trogen concentration was higher in branches than in stems andincreased acropetally. For the belowground components, theE:A ratio ranged from 0.40 to 0.91, depending on species andcomponent. However, the CO2 effect was statistically signifi-cant only for the stump (Table 2). For coarse roots, the a poste-riori comparison revealed a significant CO2 effect for P. nigra.Nitrogen concentration decreased from fine roots to coarseroots to stumps.
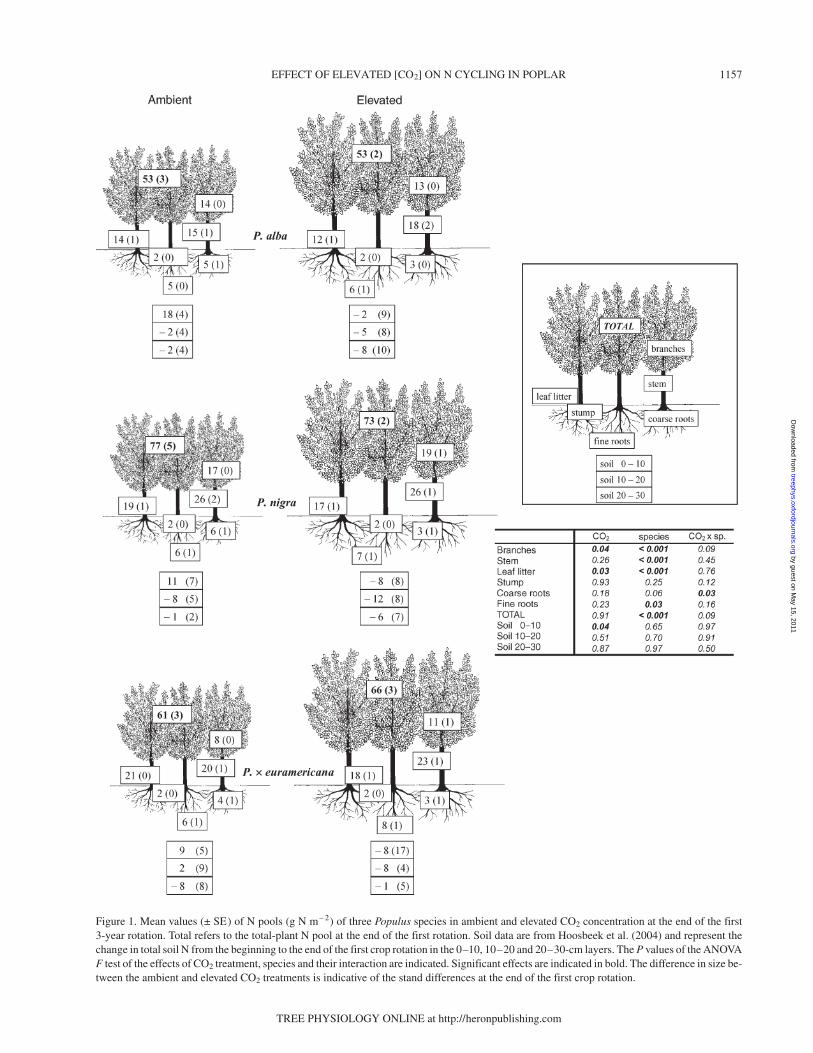
Total-plant N pool at the end of the first crop rotation rangedfrom about 53 to about 77 g m– 2, depending on species andtreatment (Figure 1). Total-plant N pool differed significantlyamong species, mainly because of differences in overall bio-mass, although no significant effect of elevated [CO2] was ob-served. Populus nigra had the highest (77 g m– 2 in elevated[CO2]) and P. alba the lowest (53 g m– 2 in both CO2 treat-ments) total-plant N pool values. Nitrogen was stored mainlyin the stems, but leaf litter and branches represented importantN pools for all three species. Elevated [CO2] had no effect onmost plant N pools. However, elevated [CO2] significantly in-creased the branch N pool and decreased the leaf litter N pool.The relationship between N content of leaf litter in ambientand elevated [CO2] (Figure 2) revealed that the slope was sig-nificantly different from 1, indicating that the differences be-tween treatments increased with the increase in N content inlitter fall between the second and third year.
The coarse root N pool decreased considerably in responseto elevated [CO2] (E:A = 0.45) for P. nigra (P = 0.03 in thea posteriori comparison). Fine roots N pool did not differ sig-nificantly between CO2 treatments, whereas the species effect
1156 CALFAPIETRA ET AL.
TREE PHYSIOLOGY VOLUME 27, 2007
Table 2. Nitrogen concentration (% dry mass) in plant tissues of three Populus species after 3 years in ambient (~370 ppm) or elevated (~550 ppm)CO2 concentration. Values represent means (± SE) with n = 3 plots. The P values of the ANOVA F test of the effects of CO2 treatment (C), species(S) and their interaction (C × S) are indicated. Stem HGI 1, 2 and 3 denote the stem height growth increment in years 1, 2 and 3, respectively.
Tissue P. alba P. nigra P. × euramericana C S C × S
Ambient Elevated Ambient Elevated Ambient Elevated
Stem HGI 1 0.39 (0.01) 0.34 (0.01) 0.50 (0.05) 0.42 (0.01) 0.51 (0.03) 0.46 (0.03) 0.04 < 0.01 0.67Stem HGI 2 0.50 (0.05) 0.40 (0.03) 0.55 (0.08) 0.46 (0.02) 0.60 (0.05) 0.51 (0.04) 0.02 0.21 0.99Stem HGI 3 0.70 (0.08) 0.66 (0.00) 0.81 (0.05) 0.76 (0.06) 0.77 (0.05) 0.77 (0.03) 0.46 0.10 0.86Sylleptic branches 1.25 (0.05) 1.04 (0.06) 1.51 (0.06) 1.40 (0.03) 1.46 (0.05) 1.37 (0.11) 0.19 < 0.001 0.35Proleptic branches 1.18 (0.02) 0.97 (0.03) 1.54 (0.04) 1.28 (0.10) 1.24 (0.05) 1.16 (0.09) 0.01 < 0.01 0.37Stump 0.61 (0.10) 0.34 (0.05) 0.70 (0.03) 0.39 (0.02) 0.56 (0.06) 0.49 (0.11) 0.04 0.51 0.20Coarse roots 0.96 (0.29) 0.46 (0.08) 1.14 (0.23) 0.46 (0.09) 0.78 (0.12) 0.58 (0.18) 0.12 0.44 0.07Fine roots 0.99 (0.10) 0.90 (0.12) 1.20 (0.28) 0.72 (0.14) 0.88 (0.18) 0.80 (0.12) 0.33 0.19 0.02Leaves 2.69 (0.10) 2.26 (0.12) 2.55 (0.11) 2.31 (0.05) 2.77 (0.01) 2.25 (0.15) 0.02 0.63 0.27Leaf litter 1.46 (0.02) 1.26 (0.01) 1.75 (0.02) 1.46 (0.03) 1.86 (0.04) 1.58 (0.06) < 0.01 < 0.001 0.20
by guest on May 15, 2011
treephys.oxfordjournals.orgD
ownloaded from
TREE PHYSIOLOGY ONLINE at http://heronpublishing.com
EFFECT OF ELEVATED [CO2] ON N CYCLING IN POPLAR 1157
Figure 1. Mean values (± SE) of N pools (g N m– 2) of three Populus species in ambient and elevated CO2 concentration at the end of the first3-year rotation. Total refers to the total-plant N pool at the end of the first rotation. Soil data are from Hoosbeek et al. (2004) and represent thechange in total soil N from the beginning to the end of the first crop rotation in the 0–10, 10–20 and 20–30-cm layers. The P values of the ANOVAF test of the effects of CO2 treatment, species and their interaction are indicated. Significant effects are indicated in bold. The difference in size be-tween the ambient and elevated CO2 treatments is indicative of the stand differences at the end of the first crop rotation.
by guest on May 15, 2011
treephys.oxfordjournals.orgD
ownloaded from
was significant, with P. × euramericana having the highestvalues (mean of 7 g m– 2 for the two treatments) and P. alba thelowest value of 5 g m– 2.
Nitrogen-use efficiency
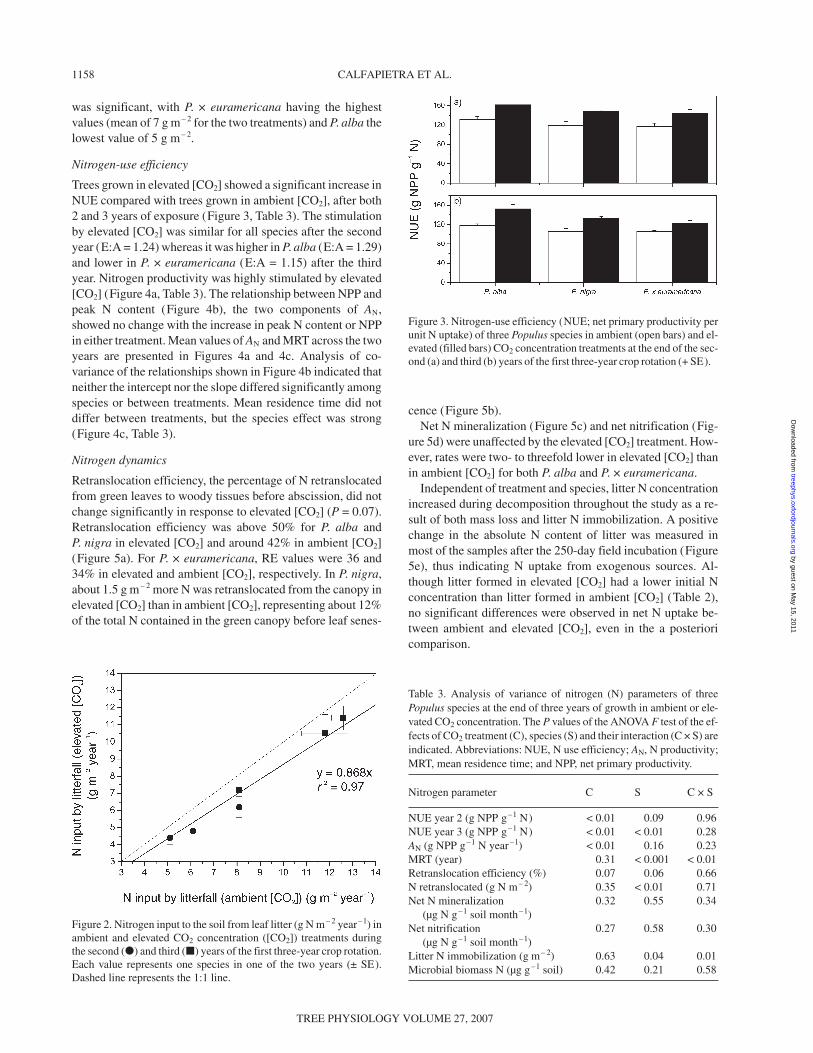
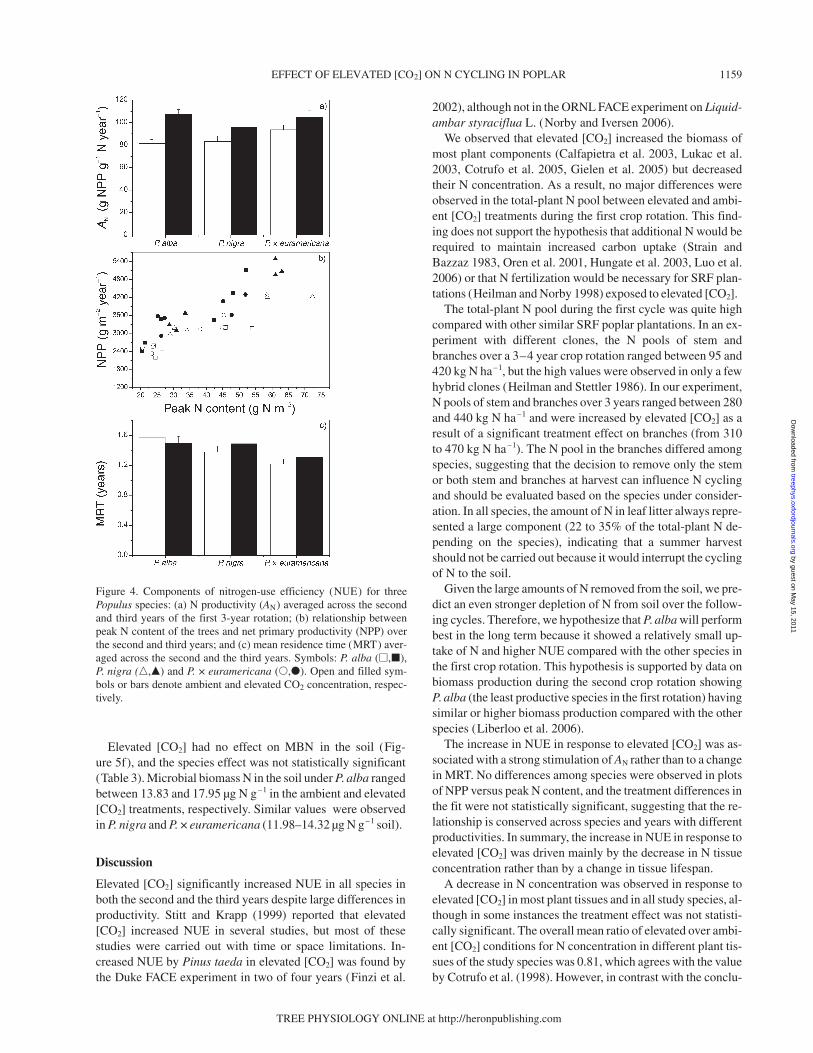
Trees grown in elevated [CO2] showed a significant increase inNUE compared with trees grown in ambient [CO2], after both2 and 3 years of exposure (Figure 3, Table 3). The stimulationby elevated [CO2] was similar for all species after the secondyear (E:A = 1.24) whereas it was higher in P. alba (E:A = 1.29)and lower in P. × euramericana (E:A = 1.15) after the thirdyear. Nitrogen productivity was highly stimulated by elevated[CO2] (Figure 4a, Table 3). The relationship between NPP andpeak N content (Figure 4b), the two components of AN,showed no change with the increase in peak N content or NPPin either treatment. Mean values of AN and MRT across the twoyears are presented in Figures 4a and 4c. Analysis of co-variance of the relationships shown in Figure 4b indicated thatneither the intercept nor the slope differed significantly amongspecies or between treatments. Mean residence time did notdiffer between treatments, but the species effect was strong(Figure 4c, Table 3).
Nitrogen dynamics
Retranslocation efficiency, the percentage of N retranslocatedfrom green leaves to woody tissues before abscission, did notchange significantly in response to elevated [CO2] (P = 0.07).Retranslocation efficiency was above 50% for P. alba andP. nigra in elevated [CO2] and around 42% in ambient [CO2](Figure 5a). For P. × euramericana, RE values were 36 and34% in elevated and ambient [CO2], respectively. In P. nigra,about 1.5 g m– 2 more N was retranslocated from the canopy inelevated [CO2] than in ambient [CO2], representing about 12%of the total N contained in the green canopy before leaf senes-
cence (Figure 5b).Net N mineralization (Figure 5c) and net nitrification (Fig-
ure 5d) were unaffected by the elevated [CO2] treatment. How-ever, rates were two- to threefold lower in elevated [CO2] thanin ambient [CO2] for both P. alba and P. × euramericana.
Independent of treatment and species, litter N concentrationincreased during decomposition throughout the study as a re-sult of both mass loss and litter N immobilization. A positivechange in the absolute N content of litter was measured inmost of the samples after the 250-day field incubation (Figure5e), thus indicating N uptake from exogenous sources. Al-though litter formed in elevated [CO2] had a lower initial Nconcentration than litter formed in ambient [CO2] (Table 2),no significant differences were observed in net N uptake be-tween ambient and elevated [CO2], even in the a posterioricomparison.
1158 CALFAPIETRA ET AL.
TREE PHYSIOLOGY VOLUME 27, 2007
Figure 2. Nitrogen input to the soil from leaf litter (g N m– 2 year–1) inambient and elevated CO2 concentration ([CO2]) treatments duringthe second (�) and third (�) years of the first three-year crop rotation.Each value represents one species in one of the two years (± SE).Dashed line represents the 1:1 line.
Figure 3. Nitrogen-use efficiency (NUE; net primary productivity perunit N uptake) of three Populus species in ambient (open bars) and el-evated (filled bars) CO2 concentration treatments at the end of the sec-ond (a) and third (b) years of the first three-year crop rotation (+ SE).
Table 3. Analysis of variance of nitrogen (N) parameters of threePopulus species at the end of three years of growth in ambient or ele-vated CO2 concentration. The P values of the ANOVA F test of the ef-fects of CO2 treatment (C), species (S) and their interaction (C × S) areindicated. Abbreviations: NUE, N use efficiency; AN, N productivity;MRT, mean residence time; and NPP, net primary productivity.
Nitrogen parameter C S C × S
NUE year 2 (g NPP g–1 N) < 0.01 0.09 0.96NUE year 3 (g NPP g–1 N) < 0.01 < 0.01 0.28AN (g NPP g–1 N year–1) < 0.01 0.16 0.23MRT (year) 0.31 < 0.001 < 0.01Retranslocation efficiency (%) 0.07 0.06 0.66N retranslocated (g N m– 2) 0.35 < 0.01 0.71Net N mineralization 0.32 0.55 0.34
(µg N g–1 soil month–1)Net nitrification 0.27 0.58 0.30
(µg N g–1 soil month–1)Litter N immobilization (g m– 2) 0.63 0.04 0.01Microbial biomass N (µg g–1 soil) 0.42 0.21 0.58
by guest on May 15, 2011
treephys.oxfordjournals.orgD
ownloaded from
Elevated [CO2] had no effect on MBN in the soil (Fig-ure 5f), and the species effect was not statistically significant(Table 3). Microbial biomass N in the soil under P. alba rangedbetween 13.83 and 17.95 µg N g–1 in the ambient and elevated[CO2] treatments, respectively. Similar values were observedin P. nigra and P. × euramericana (11.98–14.32 µg N g–1 soil).
Discussion
Elevated [CO2] significantly increased NUE in all species inboth the second and the third years despite large differences inproductivity. Stitt and Krapp (1999) reported that elevated[CO2] increased NUE in several studies, but most of thesestudies were carried out with time or space limitations. In-creased NUE by Pinus taeda in elevated [CO2] was found bythe Duke FACE experiment in two of four years (Finzi et al.
2002), although not in the ORNL FACE experiment on Liquid-ambar styraciflua L. (Norby and Iversen 2006).
We observed that elevated [CO2] increased the biomass ofmost plant components (Calfapietra et al. 2003, Lukac et al.2003, Cotrufo et al. 2005, Gielen et al. 2005) but decreasedtheir N concentration. As a result, no major differences wereobserved in the total-plant N pool between elevated and ambi-ent [CO2] treatments during the first crop rotation. This find-ing does not support the hypothesis that additional N would berequired to maintain increased carbon uptake (Strain andBazzaz 1983, Oren et al. 2001, Hungate et al. 2003, Luo et al.2006) or that N fertilization would be necessary for SRF plan-tations (Heilman and Norby 1998) exposed to elevated [CO2].
The total-plant N pool during the first cycle was quite highcompared with other similar SRF poplar plantations. In an ex-periment with different clones, the N pools of stem andbranches over a 3–4 year crop rotation ranged between 95 and420 kg N ha–1, but the high values were observed in only a fewhybrid clones (Heilman and Stettler 1986). In our experiment,N pools of stem and branches over 3 years ranged between 280and 440 kg N ha–1 and were increased by elevated [CO2] as aresult of a significant treatment effect on branches (from 310to 470 kg N ha–1). The N pool in the branches differed amongspecies, suggesting that the decision to remove only the stemor both stem and branches at harvest can influence N cyclingand should be evaluated based on the species under consider-ation. In all species, the amount of N in leaf litter always repre-sented a large component (22 to 35% of the total-plant N de-pending on the species), indicating that a summer harvestshould not be carried out because it would interrupt the cyclingof N to the soil.
Given the large amounts of N removed from the soil, we pre-dict an even stronger depletion of N from soil over the follow-ing cycles. Therefore, we hypothesize that P. alba will performbest in the long term because it showed a relatively small up-take of N and higher NUE compared with the other species inthe first crop rotation. This hypothesis is supported by data onbiomass production during the second crop rotation showingP. alba (the least productive species in the first rotation) havingsimilar or higher biomass production compared with the otherspecies (Liberloo et al. 2006).
The increase in NUE in response to elevated [CO2] was as-sociated with a strong stimulation of AN rather than to a changein MRT. No differences among species were observed in plotsof NPP versus peak N content, and the treatment differences inthe fit were not statistically significant, suggesting that the re-lationship is conserved across species and years with differentproductivities. In summary, the increase in NUE in response toelevated [CO2] was driven mainly by the decrease in N tissueconcentration rather than by a change in tissue lifespan.
A decrease in N concentration was observed in response toelevated [CO2] in most plant tissues and in all study species, al-though in some instances the treatment effect was not statisti-cally significant. The overall mean ratio of elevated over ambi-ent [CO2] conditions for N concentration in different plant tis-sues of the study species was 0.81, which agrees with the valueby Cotrufo et al. (1998). However, in contrast with the conclu-
TREE PHYSIOLOGY ONLINE at http://heronpublishing.com
EFFECT OF ELEVATED [CO2] ON N CYCLING IN POPLAR 1159
Figure 4. Components of nitrogen-use efficiency (NUE) for threePopulus species: (a) N productivity (AN) averaged across the secondand third years of the first 3-year rotation; (b) relationship betweenpeak N content of the trees and net primary productivity (NPP) overthe second and third years; and (c) mean residence time (MRT) aver-aged across the second and the third years. Symbols: P. alba (�,�),P. nigra (�,�) and P. × euramericana (�,�). Open and filled sym-bols or bars denote ambient and elevated CO2 concentration, respec-tively.
by guest on May 15, 2011
treephys.oxfordjournals.orgD
ownloaded from

sions of Cotrufo et al. (1998), we observed no greater reduc-tions in N concentration under elevated [CO2] in the above-ground woody structures compared with the belowground sys-tem. Although a decrease in N concentration in response to el-evated [CO2] is often caused by an increase in nonstructuralcarbohydrates (Stitt and Krapp 1999), this decrease in N con-centration could induce a decrease of photosynthetic capacity(cf. Stitt and Krapp 1999) because much of the leaf N is accu-mulated in Rubisco (Cooke and Weih 2005). However, weshould take into account that elevated [CO2] frequently in-duces a change in specific leaf area (Gielen and Ceulemans2001), thus reducing or cancelling the differences in leaf-area-based N concentration, the most common unit for photo-synthetic assessments. No differences between treatmentswere observed in our experiment in leaf-area-based N concen-tration or in the maximum carboxylation rate of Rubisco(Vcmax) (Calfapietra et al. 2005), which is in agreement with theconclusion reached by Ellsworth et al. (2004). This suggeststhat the increase in AN by elevated [CO2] observed at our sitecan be explained by an effect of elevated [CO2] on photo-synthetic NUE, which is primarily a function of an increasedsubstrate (CO2) supply for photosynthesis rather than a changein N metabolism (Sholtis et al. 2004).
Nitrogen cycling includes a complex series of processes thatremain largely unknown, thus increasing the uncertainties inestimating the effects of elevated [CO2] on N use (Johnson andBall 1996). Nitrogen uptake by trees is separated into a long-lived component (woody tissues) and a fast-turnover compo-nent (leaves and fine roots). Because of the rapid decomposi-tion of leaves and fine roots, N in the fast-turnover componentbecomes available for plant uptake relatively soon after de-composition and subsequent mineralization of organic N. Inour experiment, the amount of N from leaf litter was consider-ably higher in ambient [CO2] than in elevated [CO2]. Thismight explain the observed increases in the total N pool in thetop soil layer in ambient [CO2], whereas in all other soil layers,in both CO2 treatments, total N decreased throughout the ex-periment, with the greatest declines observed in elevated[CO2]. Preliminary data on N2O emissions from soil indicatethat these emissions are too small to influence the variations insoil N (Castaldi, Second University of Naples, pers. comm.).Also, N deposition in this area was considerably below themean values for European regions (Mosello et al. 2002) andwas not considered in our analysis.
Our data suggest that elevated [CO2] had a negative effect onnet nitrification and net N mineralization rates for P. alba and
1160 CALFAPIETRA ET AL.
TREE PHYSIOLOGY VOLUME 27, 2007
Figure 5. Mobilization (a–d) and immobilization (e, f) mechanisms of N occurring for three Populus species in ambient (white bars) and elevated(black bars) CO2 concentration treatments. (a) Retranslocation efficiency (RE), (b) N retranslocated from the standing green canopy in August ofthe third year before leaf fall, (c) soil net N mineralization, (d) soil net nitrification, (e) litter N immobilization after 250 days of incubation, and (f)N content in soil microbial biomass (MBN). Bars represent means of n = 3 (+ SE).
by guest on May 15, 2011
treephys.oxfordjournals.orgD
ownloaded from

P. × euramericana, but the overall treatment effect was not sig-nificant. However, real differences may have been undetect-able because of the great variability among plots and the lim-ited number of replicates (three plots). A significant decline innitrification rate in elevated [CO2] was found by Ambus andRobertson (1999) in Populus. This could partly explain thelarge decrease in soil nitrate in elevated [CO2] during the thirdyear, the most productive year of the first crop rotation(Moscatelli et al. 2005).
The conversion of inorganic N (NO3– and NH4
+) into organicforms (amino acids and proteins), mediated by microrganismsthat decompose carbonaceous organic residues in the soil, usu-ally requires more N than is contained in the residues. Nitro-gen immobilized in microbial cells in the short-term can latereither be converted into forms that make up the humus com-plex, thus leading to long-term immobilization, or released asNO3
– or NH4+ (Brady and Weil 2002). The immobilization of N
in the soil microbial biomass was unaffected by the CO2 treat-ments, which is in agreement with the findings of Holmes et al.(2003) at the Aspen FACE site and of Finzi et al. (2006) at theDuke FACE site. However, this finding could be an indicationthat there is still a strong influence of the native pool of soil or-ganic matter on microbial activity, because Van Groenigen etal. (2006) showed that microbial N immobilization is the mainmechanism involved in N retention in soils in elevated [CO2],especially when nutrients become limiting.
Mineralization and immobilization processes occur simul-taneously in soil. The balance between the two processes canbe shifted by several factors, including the quality of the sub-strates. The observed increase in the C:N ratio of litter (Cotru-fo et al. 2005) and labile soil organic matter (Lagomarsino etal. 2006) in response to elevated [CO2] was predicted to inducea negative trend in N mineralization.
In mature forests, > 80% of annual tree N uptake is recycled(Cole and Rapp 1981). It has been calculated that, in the firstseason of growth, 50% or more of the total N in a Populus treeis in the green canopy; a significant amount of N is subse-quently retranslocated from the canopy to the perennial tissuesbefore leaf fall (Cooke et al. 2003). We observed a trend of in-creased retranslocation efficiency in elevated [CO2] (P = 0.07)for all species, thus leading to an increase of the long-termstorage of N (wood) versus its fast turnover (leaf litter). Thisfinding is in agreement with other studies, where elevated[CO2] slightly increased N resorption efficiency from thegreen canopy (Norby et al. 2000, Finzi et al. 2001). This effectbecame more significant when it ameliorated the negative ef-fects of O3 exposure (Lindroth et al. 2001).
It appeared that the elevated [CO2] treatment affected thedecay rate of leaf litter in two opposing ways, at least duringthe first stage of litter decay. Litter produced in elevated [CO2]decomposed more slowly than litter produced in ambient[CO2], regardless of where the litter was incubated, and leaflitter incubated in elevated [CO2] decomposed faster than leaflitter incubated in ambient [CO2], regardless of the origin ofthe litter (Cotrufo et al. 2005). Despite its lower N concentra-tion, litter grown in elevated [CO2] did not immobilize N dif-ferently from litter grown in ambient [CO2].
In conclusion, although elevated [CO2] increased productiv-ity of poplar trees, it had no effect on N uptake. As a conse-quence, NUE increased in trees in the elevated [CO2] treat-ment, thus negating the idea that additional N is required tomaintain increased carbon uptake in an elevated [CO2] atmo-sphere in short-rotation poplar plantations. However, a greaterdecrease in soil N concentration was observed at the end of thefirst crop rotation in the elevated [CO2] treatment than in theambient [CO2] treatment, probably because of decreased N in-put by leaf litter. Moreover, we observed a trend, although notsignificant, of decreased N mobilization in elevated [CO2]. Inthe following rotations, we will be able to verify if these pro-cesses are exacerbated over longer time periods under elevated[CO2] and determine if the future productivity of such planta-tions could be compromised by increasing atmospheric [CO2].
Acknowledgments
This research was conducted under the following programs: theEU-POPFACE (ENV4-CT97-0657), EU-EUROFACE (EVR1-CT-2002-40027), Center of Excellence “Forest and Climate” (MIUR Ital-ian Ministry of University and Research), the Bilateral Project It-aly-USA on Climate Change (Italian Ministry of Environment) andthe Marie-Curie fellowship GLOBALVOC (MOIF-CT-2005-7692).The authors acknowledge S. Linder for useful comments on nutrientcycling, T. Sharkey for language revision, T. Oro and M. Tamantinifor technical assistance, and B. Bartoli and R. Pigliacelli for help dur-ing field and laboratory activities. B. Gielen is a post-doctoral re-search fellow of the Fund for Scientific Research-Flanders (F.W.O.-Vlaanderen).
References
Aerts, R. and F.S. Chapin. 2000. The mineral nutrition of wild plantsrevisited: a re-evaluation of processes and patterns. Adv. Ecol. Res.30:1–67.
Ambus, P. and G.P. Robertson. 1999. Fluxes of CH4 and N2O in aspenstands grown under ambient and twice-ambient CO2. Plant Soil209:1–8.
Anderson, J.M. and J.S.I. Ingram. 1993. Colorimetric determinationof ammonium. In Tropical Soil Biology and Fertility. A Handbookof Methods. 2nd Edn. Eds. J.M. Anderson and J.S.I. Ingram. CABInternational, Wallingford, pp 73–74.
Bazzaz, F.A. 1990. Response of annual ecosystems to the risingglobal CO2 levels. Annu. Rev. Ecol. Syst. 21:167–196.
Berendse, F. and R. Aerts. 1987. Nitrogen use efficiency: a biologi-cally meaningful definition? Funct. Ecol. 1:293–296.
Brady, N.C. and R.R. Weil. 2002. The nature and properties of soils.13th Edn. Prentice Hall, Upper Saddle River, NJ, 960 p.
Calfapietra, C., B. Gielen, M. Sabatti, P. De Angelis, G. Scara-scia-Mugnozza and R. Ceulemans. 2001. Growth performance ofPopulus exposed to “Free Air Carbon dioxide Enrichment” duringthe first growing season in the POPFACE experiment. Ann. For.Sci. 58:819–828.
Calfapietra, C., B. Gielen, A.N.J. Galema, M. Lukac, P. De Angelis,M.C. Moscatelli, R. Ceulemans and G. Scarascia-Mugnozza.2003. Free-air CO2 enrichment (FACE) enhances biomass produc-tion in a short-rotation poplar plantation (POPFACE). TreePhysiol. 23:805–814.
TREE PHYSIOLOGY ONLINE at http://heronpublishing.com
EFFECT OF ELEVATED [CO2] ON N CYCLING IN POPLAR 1161
by guest on May 15, 2011
treephys.oxfordjournals.orgD
ownloaded from

Calfapietra, C., I. Tulva, E. Eensalu, M. Perez, P. De Angelis, G. Sca-rascia-Mugnozza and O. Kull. 2005. Canopy profiles of photo-synthetic parameters under elevated CO2 and N fertilization in apoplar plantation. Environ. Pollut. 137:525–535.
Cataldo, D., M. Haroon, L. Schrader and M. Young. 1975. Rapidcolorimetric determination of nitrate in plant tissue by nitration ofsalycilic acid. Commun. Soil Sci. Plan. Anal. 6:71–80.
Chapman, J.A., J.S. King, K.S. Pregitzer and D.R. Zak. 2005. Effectsof elevated concentrations of atmospheric CO2 and tropospheric O3
on decomposition of tree fine roots. Tree Physiol. 25:1501–1510.Cole, D. and M. Rapp. 1981. Elemental cycling in forest ecosystems.
In Dynamic Properties of Forest Ecosystems. Ed. D. Reichle. Cam-bridge University Press, London, pp 341–409.
Cooke, J., K. Brown, R. Wu and J. Davis. 2003. Gene expression as-sociated with nitrogen-induced shifts in allocation in poplar. PlantCell Environ. 26:757–770.
Cooke, J. and M. Weih. 2005. Nitrogen storage and seasonal nitrogencycling in Populus: bridging molecular physiology and ecophys-iology. New Phytol. 167:19–30.
Cotrufo, M.F., P. Ineson and A. Scott. 1998. Elevated CO2 reduces thenitrogen concentration of plant tissues. Global Change Biol. 4:43–54.
Cotrufo, M.F., P. De Angelis and A. Polle. 2005. Leaf litter produc-tion and decomposition in a poplar short-rotation coppice exposedto free air CO2 enrichment (POPFACE). Global Change Biol.11:971–982.
Ellsworth, D.S., P.B. Reich, E.S. Naumburg, G.W. Koch,M.E. Kubiske and S.D. Smith. 2004. Photosynthesis, carboxyl-ation and leaf nitrogen responses of 16 species to elevated pCO2
across four free-air CO2 enrichment experiments in forest, grass-land and desert. Global Change Biol. 10:2121–2138.
Eno, C.F. 1960. Nitrate production in the field by incubating the soilin polyethylene bags. Soil Sci. Soc. Am. Proc. 24:277–279.
Ericsson, T. 1994. Nutrient cycling in energy forest plantations. Bio-mass Bioenergy 6:115–121.
Finzi, A.C., A.S. Allen, E.H. DeLucia, D.S. Ellsworth andW.H. Schlesinger. 2001. Forest litter production, chemistry and de-composition following two years of free-air CO2 enrichment. Ecol-ogy 82:470–484.
Finzi, A.C., E.H. DeLucia, J.G. Hamilton, D.D. Richter andW.H. Schlesinger. 2002. The nitrogen budget of a pine forest underfree air CO2 enrichment. Oecologia 132:567–578.
Finzi, A.C., D.J.P. Moore, E.H. DeLucia et al. 2006. Progressive ni-trogen limitation of ecosystem processes under elevated CO2 in awarm-temperate forest. Ecology 87:15–25.
Gielen, B. and R. Ceulemans. 2001. The likely impact of rising atmo-spheric CO2 on natural and managed Populus: a literature review.Environ. Pollut. 115:335–358.
Gielen, B., M. Liberloo, J. Bogaert, C. Calfapietra, P. De Angelis,F. Miglietta, G. Scarascia-Mugnozza and R. Ceulemans. 2003.Three years of free-air CO2 enrichment (POPFACE) only slightlyaffect profiles of light and leaf characteristics in closed canopies ofPopulus. Global Change Biol. 9:1022–1037.
Gielen, B., C. Calfapietra, M. Lukac et al. 2005. Net carbon storage ina poplar plantation (POPFACE) after three years of free-air CO2
enrichment. Tree Physiol. 25:1399–1408.Gill, R.A., L.J. Anderson, H.W. Polley, H.B. Johnson and R.B. Jack-
son. 2006. Potential nitrogen constraints on soil carbon sequestra-tion under low and elevated atmospheric CO2. Ecology 87:41–52.
Heilman, P.E. and R.F. Stettler. 1986. Nutritional concerns in selec-tion of black cottonwood and hybrid clones for short rotation. Can.J. For. Res. 16:860–863.
Heilman, P.E. and R.J. Norby. 1998. Nutrient cycling and fertilitymanagement in temperate short rotation forest. Biomass Bioenergy14:361–370.
Holmes, W.E., D.R. Zak, K.S. Pregitzer and J.S. King. 2003. Soil ni-trogen transformations under Populus tremuloides, Betula papyri-fera and Acer saccharum following 3 years exposure to elevatedCO2 and O3. Global Change Biol. 9:1743–1750.
Hoosbeek, M.R., M. Lukac, D. van Dam et al. 2004. More new carbonin the mineral soil of a poplar plantation under Free Air Carbon En-richment (POPFACE): cause of increased priming effect? GlobalBiogeochem. Cycles 18:GB1040.
Hungate, B., J. Dukes, M. Shaw, Y. Luo and C. Field. 2003. Nitrogenand climate change. Science 302:1512–1513.
Isaac, M.E., A.M. Gordon, N. Thevathasan, S.K. Oppong andJ. Quashie-Sam. 2005. Temporal changes in soil carbon and nitro-gen in west African multistrata agroforestry systems: a chrono-sequence of pools and fluxes. Agrofor. Syst. 65:23–31.
Joergensen, R.G. and P. Brookes. 1990. Ninhydrin-reactive nitrogenmeasurements of microbial biomass in 0.5 M K2SO4 soil extracts.Soil Biol. Biochem. 22:1023–1027.
Johnson, D.W. and J.T. Ball. 1996. Interactions between CO2 and ni-trogen in forests: can we extrapolate from the seedlings to the standlevel? In Carbon Dioxide and Terrestrial Ecosystems. Eds. G. Kochand H. Mooney. Academic Press, San Diego, pp 283–297.
Johnson, D.W., R.J. Norby and B.A. Hungate. 2001. Effects of ele-vated CO2 on nutrient cycling in forests. In The Impact of CarbonDioxide and Other Greenhouse Gases on Forest Ecosystems. Eds.D.F. Karnosky, R. Ceulemans, G. Scarascia-Mugnozza andJ.L. Innes. CAB International, Wallingford, pp 237–252.
Johnson, D.W., W. Cheng, J.D. Joslin, R.J. Norby, N.T. Edwards andD.E. Todd, Jr. 2004. Effects ofelevated CO2 on nutrient cycling in asweetgum plantation. Biogeochemistry 69:379–403.
Johnson, D.W. 2006. Progressive N limitation in forests: review andimplications for long-term responses to elevated CO2. Ecology87:64–75.
King, J.S., M.E. Kubiske, K.S. Pregitzer, G.R. Hendrey, C. Giardina,E. McDonald, V.S. Quinn and D.F. Karnosky. 2005. Ozone de-creases the capacity of CO2 fertilization of forests to offset fossilfuel emissions. New Phytol. 168:623–636.
Lagomarsino, A., M.C. Moscatelli, P. De Angelis and S. Grego. 2006.Labile substrates quality as the main driving force of microbialmineralization activity in a poplar plantation soil under elevatedCO2 and nitrogen fertilization. Sci. Total Environ. 372:256–265.
Liberloo, M., C. Calfapietra, M. Lukac et al. 2006. Woody biomassproduction of a bioenergy forest increases in a future high CO2
world. Global Change Biol. 12:1094–1106.Lindroth, R.L., B.J. Kopper, W.F.J. Parsons, J.G. Bockheim,
D.F. Karnosky, G. Hendrey, K.S. Pregitzer, J.G. Isebrands andJ. Sober. 2001. Consequences of elevated carbon dioxide andozone for foliar chemical composition and dynamics in tremblingaspen (Populus tremuloides) and paper birch (Betula papyrifera).Environ. Pollut. 115:395–404.
Liu, L., J. King and C. Giardina. 2005. Effects of elevated concentra-tions of atmospheric CO2 and tropospheric O3 on leaf litter produc-tion and chemistry in trembling aspen and paper birchcommunities. Tree Physiol. 25:1511–1522.
Lukac, M., C. Calfapietra and D.L. Godbold. 2003. Production, turn-over and mycorrhizal colonisation of root systems of three Populusspecies grown under elevated CO2 (POPFACE). Global ChangeBiol. 9:838–848.
Luo, Y., B. Su, W. Currie et al. 2004. Progressive nitrogen limitationof ecosystem responses to rising atmospheric carbon dioxide. Bio-science 54:731–739.
1162 CALFAPIETRA ET AL.
TREE PHYSIOLOGY VOLUME 27, 2007
by guest on May 15, 2011
treephys.oxfordjournals.orgD
ownloaded from

Luo, Y., D. Hui and D. Zhang. 2006. Elevated CO2 stimulated net ac-cumulations of carbon and nitrogen in land ecosystems: a meta-analysis. Ecology 87:53–63.
Miglietta, F., A. Peressotti, F.P. Vaccari, A. Zaldei, P. De Angelis andG. Scarascia-Mugnozza. 2001. Free-air CO2 enrichment (FACE)of a poplar plantation: the POPFACE fumigation system. NewPhytol. 150:465–476.
Moscatelli, M.C., A. Lagomarsino, P. De Angelis and S. Grego. 2005.Seasonality of soil biological properties in a poplar plantationgrowing under elevated atmospheric CO2. Appl. Soil Ecol. 30:162–173.
Nambiar, E.K.S. and D.N. Fife. 1991. Nutrient retranslocation in tem-perate conifers. Tree Physiol. 9:185–207.
Norby, R.J., T. Long, J. Hartz and E.G. O’Neill. 2000. Nitrogen re-sorption in senescing tree leaves in a warmer, CO2-enriched atmo-sphere. Plant Soil 224:15–29.
Norby, R.J., M.F. Cotrufo, P. Ineson, E.G. O’Neill and J.G. Canadell.2001. Elevated CO2, litter chemistry, and decomposition: a synthe-sis. Oecologia 127:153–165.
Norby, R., E. DeLucia, B. Gielen et al. 2005. Forest response to ele-vated CO2 is conserved across a broad range of productivity. Proc.Natl. Acad. Sci. USA 102:18,052–18,056.
Norby, R.J. and C.M. Iversen. 2006. Nitrogen uptake, distribution,turnover and efficiency of use in a CO2-enriched sweetgum forest.Ecology 87:5–14.
Oren, R., D.S. Ellsworth, K.H. Johnsen et al. 2001. Soil fertility limitscarbon sequestration by forest ecosystems in a CO2-enriched atmo-sphere. Nature 411:469–472.
Poorter, H., Y. Van Berkel, R. Baxter et al. 1997. The effect of ele-vated CO2 on the chemical composition and construction costs ofleaves of 27 C3 species. Plant Cell Environ. 20:472–482.
Scarascia-Mugnozza, G., C. Calfapietra, R. Ceulemans et al. 2006.Responses to elevated CO2 of a short rotation, multispecies poplarplantation. In Managed Ecosystems and Elevated CO2: Case Stud-ies, Processes and Perspectives. Eds. J. Nosberger, S.P. Long,R.J. Norby, M. Stitt, G.R. Hendrey and H. Blum. Springer-Verlag,Berlin, pp 173–195.
Sholtis, J.D., C.A. Gunderson, R.J. Norby and D.T. Tissue. 2004. Per-sistent stimulation of photosynthesis by elevated CO2 in a sweet-gum (Liquidambar styraciflua L.) forest stand. New Phytol. 162:343–354.
Stitt, M. and A. Krapp. 1999. The interaction between elevated carbondioxide and nitrogen nutrition: the physiological and molecularbackground. Plant Cell Environ. 22:583–621.
Strain, B.R. and F.A. Bazzaz. 1983. Terrestrial plant communities. InCO2 and Plants: The Response of Plants to Rising Levels of Atmo-spheric CO2. Ed. E.R. Lemon. Westview Press, Boulder,pp 177–222.
Tissue, D.T., R.B. Thomas and B.R. Strain. 1997. Atmospheric CO2
enrichment increases growth and photosynthesis of Pinus taeda: a4 year experiment in the field. Plant Cell Environ. 20:1123–1134.
van Heerwaarden, T.S. and R. Aerts. 2003. Current measures of nutri-ent resorption efficiency lead to a substantial underestimation ofreal resorption efficiency: facts and solutions. Oikos 101:664–669.
Vance, E., P. Brookes and D. Jenkinson. 1987. An extraction methodfor measuring soil microbial biomass C. Soil Biol. Biochem. 19:703–707.
van Groenigen, K.J., M.A. de Graaff, J. Six, D. Harris, P. Kuikmanand C. van Kessel. 2006. The impact of elevated atmospheric [CO2]on soil C and N dynamics: a meta-analysis. In Managed Ecosys-tems and Elevated CO2: Case Studies, Processes and Perspectives.Eds. J. Nosberger, S.P. Long, R.J. Norby, M. Stitt, G.R. Hendreyand H. Blum. Springer-Verlag, Berlin, pp 373–391.
Wang, W., M. Wang and P. Lin. 2003. Seasonal changes in elementcontents in mangrove element retranslocation during leaf senes-cence. Plant Soil 252:187–193.
Wong, S.C. 1979. Elevated atmospheric partial pressure of carbon di-oxide and plant growth. I. Interaction of nitrogen nutrition andphotosynthetic capacity in C3 and C4 plants. Oecologia 22:68–74.
Zak, D.R., K.S. Pregitzer, P.S. Curtis, C.S. Vogel, W.E. Holmes andJ. Lussenhop. 2000. Atmospheric CO2, soil-N availability, and al-location of biomass and nitrogen by Populus tremuloides. Ecol.Appl. 10:34–46.
Zak, D.R., W.E. Holmes, A.C. Finzi, R.J. Norby and W.H. Schle-singer. 2003. Soil nitrogen cycling under elevated CO2: a synthesisof forest FACE experiments. Ecol. Appl. 13:1508–1514.
TREE PHYSIOLOGY ONLINE at http://heronpublishing.com
EFFECT OF ELEVATED [CO2] ON N CYCLING IN POPLAR 1163
by guest on May 15, 2011
treephys.oxfordjournals.orgD
ownloaded from
Related Documents











