624 Conservation Biology, Pages 624–633 Volume 15, No. 3, June 2001 Increased Herbivory in Forest Isolates: Implications for Plant Community Structure and Composition MADHU RAO,*§ JOHN TERBORGH,† AND PERCY NUÑEZ‡ *Nicholas School of the Environment, Box 90328, Duke University, Durham, NC 27708, U.S.A. †Center for Tropical Conservation, Duke University, P.O. Box 90381, Durham, NC 27708, U.S.A. ‡Herbario Vargas, Universidad Nacional San Antonio de Abad de Cusco, Cusco, Peru Abstract: Understanding processes driving population declines and, ultimately, species loss in forest isolates has significant implications for the long-term maintenance of species diversity. We investigated a potential mechanism driving loss of plant species in small, medium, and large land-bridge islands in Lago Guri, a 4300-km 2 hydroelectric impoundment in the State of Bolivar, Venezuela. Our hypothesis was that elevated Atta (leaf-cutter ants) herbivory on small Guri islands, attributable to release from predation, could result in reduced sapling densities, with potentially negative effects on preferred plant species. We experimentally es- tablished Atta preferences among 43 common tree species. Vegetation plots on small, medium, and large land masses were examined for variation in density, size-class distribution, and representation of preferred and less preferred species of small stems. Densities of adult trees of preferred and less preferred species were com- pared between Atta colony foraging zones and control plots without active Atta colonies, and between forag- ing zones on the small islands and the large island. Small stem densities were significantly reduced on the small islands, especially in the sapling category (1 m tall, 1 cm dbh). On medium and large land masses, preferred species were underrepresented in both sapling ( 1 m tall, 1 cm dbh) and juvenile (1 cm dbh, 10 cm dbh) size classes. On small islands, however, both preferred and less preferred species showed re- duced representation in the sapling size class. Adults of preferred species were significantly reduced within Atta foraging zones compared with control plots without Atta colonies. Small stem densities were independent of adult tree densities for preferred species on small islands and for both preferred and less preferred species on large land masses. High levels of Atta herbivory could be a potentially important cause of reduced plant re- cruitment on small Guri islands. Our findings contribute to understanding of the role of altered ecological in- teractions as deterministic mechanisms driving a process of change in the structure and composition of frag- mented communities. Incremento de la Herbivoría en Fragmentos Aislados de Bosque: Implicaciones en la Estructura y Composición de la Comunidad de Plantas Resumen: El entendimiento de los procesos que intervienen en la declinación de poblaciones y, finalmente, la pérdida de especies en fragmentos aislados de bosque tiene implicaciones significativas en el manten- imiento a lo largo de especies. Investigamos un mecanismo potencial de la pérdida de especies vegetales en is- las pequeñas, medianas y grandes en el Lago Guri, una presa hidroeléctrica de 4300 km 2 en el estado de Bolí- var, Venezuela. Nuestra hipótesis fue que alta herbivoría de Atta (hormigas arrieras) en islas pequeñas atribuible a la ausencia de depredación podría resultar en bajas densidades de renuevos con efectos poten- cialmente negativos en las especies preferidas. Experimentalmente establecimos las preferencias de Atta entre 43 especies de árboles comunes. Se examinaron parcelas de vegetación pequeñas, medianas y grandes para ver la variación en la densidad, distribución de clases de tamaño y la representación de especies preferidas y menos preferidas. Comparamos las densidades de árboles adultos de las especies preferidas y menos preferi- das en zonas con forrajeo de Atta y en parcelas control (sin colonias activas de Atta) y entre zonas de forrajeo en islas pequeñas y en la isla grande. Las densidades de plántulas fueron significativamente bajas en las islas pequeñas, especialmente en la categoría 1 m de altura y 1 cm dap. En parcelas medianas y grandes, las §Current address: Wildlife Conservation Society, 185th Street and Southern Boulevard, Bronx, NY 10460, U.S.A. Paper submitted October 1, 1999; revised manuscript accepted July 5, 2000.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

624
Conservation Biology, Pages 624–633Volume 15, No. 3, June 2001
Increased Herbivory in Forest Isolates: Implications for Plant Community Structure and Composition
MADHU RAO,*§ JOHN TERBORGH,† AND PERCY NUÑEZ‡
*Nicholas School of the Environment, Box 90328, Duke University, Durham, NC 27708, U.S.A.†Center for Tropical Conservation, Duke University, P.O. Box 90381, Durham, NC 27708, U.S.A.‡Herbario Vargas, Universidad Nacional San Antonio de Abad de Cusco, Cusco, Peru
Abstract:
Understanding processes driving population declines and, ultimately, species loss in forest isolateshas significant implications for the long-term maintenance of species diversity. We investigated a potentialmechanism driving loss of plant species in small, medium, and large land-bridge islands in Lago Guri, a4300-km
2
hydroelectric impoundment in the State of Bolivar, Venezuela. Our hypothesis was that elevated
Atta
(leaf-cutter ants) herbivory on small Guri islands, attributable to release from predation, could result inreduced sapling densities, with potentially negative effects on preferred plant species. We experimentally es-tablished
Atta
preferences among 43 common tree species. Vegetation plots on small, medium, and large landmasses were examined for variation in density, size-class distribution, and representation of preferred andless preferred species of small stems. Densities of adult trees of preferred and less preferred species were com-pared between
Atta
colony foraging zones and control plots without active
Atta
colonies, and between forag-ing zones on the small islands and the large island. Small stem densities were significantly reduced on thesmall islands, especially in the sapling category (
�
1 m tall,
�
1 cm dbh). On medium and large land masses,preferred species were underrepresented in both sapling (
�
1 m tall,
�
1 cm dbh) and juvenile (
�
1 cm dbh,
�
10 cm dbh) size classes. On small islands, however, both preferred and less preferred species showed re-duced representation in the sapling size class. Adults of preferred species were significantly reduced within
Atta
foraging zones compared with control plots without
Atta
colonies. Small stem densities were independentof adult tree densities for preferred species on small islands and for both preferred and less preferred specieson large land masses. High levels of
Atta
herbivory could be a potentially important cause of reduced plant re-cruitment on small Guri islands. Our findings contribute to understanding of the role of altered ecological in-teractions as deterministic mechanisms driving a process of change in the structure and composition of frag-mented communities.
Incremento de la Herbivoría en Fragmentos Aislados de Bosque: Implicaciones en la Estructura y Composición dela Comunidad de Plantas
Resumen:
El entendimiento de los procesos que intervienen en la declinación de poblaciones y, finalmente,la pérdida de especies en fragmentos aislados de bosque tiene implicaciones significativas en el manten-imiento a lo largo de especies. Investigamos un mecanismo potencial de la pérdida de especies vegetales en is-las pequeñas, medianas y grandes en el Lago Guri, una presa hidroeléctrica de 4300 km
2
en el estado de Bolí-var, Venezuela. Nuestra hipótesis fue que alta herbivoría de
Atta
(hormigas arrieras) en islas pequeñasatribuible a la ausencia de depredación podría resultar en bajas densidades de renuevos con efectos poten-cialmente negativos en las especies preferidas. Experimentalmente establecimos las preferencias de
Atta
entre43 especies de árboles comunes. Se examinaron parcelas de vegetación pequeñas, medianas y grandes paraver la variación en la densidad, distribución de clases de tamaño y la representación de especies preferidas ymenos preferidas. Comparamos las densidades de árboles adultos de las especies preferidas y menos preferi-das en zonas con forrajeo de
Atta
y en parcelas control (sin colonias activas de
Atta
) y entre zonas de forrajeoen islas pequeñas y en la isla grande. Las densidades de plántulas fueron significativamente bajas en las islaspequeñas, especialmente en la categoría
�
1 m de altura y
�
1 cm dap. En parcelas medianas y grandes, las
§
Current address: Wildlife Conservation Society, 185th Street and Southern Boulevard, Bronx, NY 10460, U.S.A.Paper submitted October 1, 1999; revised manuscript accepted July 5, 2000.

Conservation BiologyVolume 15, No. 3, June 2001
Rao et al. Herbivory in Forest Isolates
625
Introduction
Fragmentation of landscapes and the subsequent creationof isolated habitat patches are widespread phenomena withpotentially deleterious consequences for species diversity(Diamond 1984). Attempts to thwart species loss in rapidlyshrinking habitat requires an understanding of mechanismsdriving the extinction process. Some of the most instruc-tive lessons can be derived from the study of land-bridgeislands, which are islands once connected to the adjacentmainland. MacArthur and Wilson’s (1963, 1967) theoryof island biogeography provides a valuable conceptualfoundation for investigating the composition of insular bio-tas such as those found on land-bridge islands. The theorypredicts that land-bridge islands and habitat fragments thatare supersaturated with species at the time of isolation—that contain more species than can be maintained by colo-nization at equilibrium—will subsequently lose speciesthrough a process of species loss called “relaxation.” Dur-ing relaxation, extinction prevails over colonization, andthe system gradually collapses to a dynamic equilibriumcharacterized by simplified community structure and re-duced species diversity. Species loss is not random with re-spect to trophic role, with species at higher trophic levelsbeing most vulnerable to extinction and therefore amongthe first to become extinct locally. Thus, deterministic pat-terns of species loss on relaxing land-bridge islands result inthe complete absence of certain trophic levels, makingsuch islands ideally suited for examination of mechanistichypotheses related to the extinction process (Brown 1971;Heaney 1984; Lomolino et al. 1989).
Protected areas such as national parks and sanctuariesare becoming increasingly isolated within a human-dom-inated landscape matrix and thus are comparable to re-laxing land-bridge islands (Newmark 1987). Like recentlyisolated land-bridge islands, protected areas may be con-sidered supersaturated and could be expected to losespecies over time. Thus, understanding mechanisms driv-ing species loss on land-bridge islands has significant im-plications for the long-term maintenance of biodiversityin protected areas.
Our objective was to examine the role of altered eco-logical interactions as deterministic mechanisms under-
lying change in community composition and structurewithin a system of relaxing land-bridge islands in LagoGuri, Venezuela, a 4300-km
2
hydroelectric impoundmenton the Rio Caroní in the State of Bolivar. We specificallyinvestigated potential processes through which plantpopulations are driven to extinction in small forest iso-lates. Our primary hypothesis was that elevated herbi-vore density caused by reduced predation on small Guriislands results in declining sapling densities, with poten-tially negative effects on preferred plant species.
Leaf-cutter ants (genus
Atta
) are important selectiveherbivores throughout the Neotropics. Selective
Atta
herbivory reduces plant reproductive potential throughdecreased seed production and hinders seedling survi-vorship (Rockwood 1973; Blanton & Ewel 1985; Neps-tad et al. 1996; de Vasconcelos & Cherrett 1997).
Atta
colonies occur at much higher densities on smaller Guriislands than on larger land masses, primarily because ofthe reduced presence of
Atta
predators (Rao 1998). Weexamined patterns of stem density and composition atsites differing by an order of magnitude in
Atta
densities.Fragmentation-induced reduction in levels of predationcould potentially lead to elevated herbivory, whichcould influence the structure and composition of the un-derstory plant species community.
Methods
Study Areas
Lago Guri was formed by damming of the Caroní River inthe State of Bolivar, Venezuela (lat. 7
�
20
�
N, long. 62
�
48
�
W),and reached its final level in 1986 ( Morales & Gorzula1986). The formation of Lago Guri created several hun-dred forested islands (former hilltops) ranging in sizefrom
�
0.1 ha to
�
1000 ha which are ideally suited toexamine the consequences of fragmentation (Alvarez etal. 1986). Leaf-cutter ants (
Atta sexdens
and
Atta cephal-otes
) have attained high densities on small Lago Guri is-lands (Rao 1998, 2000). A survey of mature ant coloniesrevealed an order of magnitude increase in densities onsmall (0.3–3.0 ha) and medium (7–12 ha) islands (5.6
especies preferidas estaban poco representadas en las clases renuevo (
�
1 m de altura
�
1 cm dap) y juvenil(
�
1 cm DAP
�
10 cm dap). Sin embargo en islas pequeñas, tanto las especies preferidas como las no preferi-das mostraron baja representación en la clase renuevo. Adultos de las especies preferidas estaban significati-vamente reducidos dentro de las zonas de forrajeo en comparación con las parcelas control (sin colonias de
Atta
). Las densidades de renuevos fueron independientes de las densidades de árboles adultos de especiespreferidas en islas pequeñas y para especies preferidas y menos preferidas en islas grandes. Niveles altos deherbivoría de
Atta
pudieran ser un importante factor potencial de reducción del reclutamiento en islas pe-queñas de Guri. Nuestros hallazgos contribuyen al entendimiento del papel de interacciones ecológicas al-teradas como mecanismos determinísticos que dirigen un proceso de cambio en la estructura y la com-
posición de comunidades fragmentadas.

626
Herbivory in Forest Isolates Rao et al.
Conservation BiologyVolume 15, No. 3, June 2001
and 2.3 colonies per ha, respectively) in comparisonwith large islands (250–350 ha, 0.72 colonies/ha) andthe mainland (0 nests in 5.5 ha).
Reduced presence of
Atta
predators was the primarycause of observed variation in
Atta
densities (Rao 2000).Important predators of
Atta
include army ants (
Eciton
spp.), armadillos (
Dasypodidae
), and phorid parasitoids(
Neodohrniphora
and
Apocephalus
spp.) (Brian 1983;Feener & Brown 1993; Gotwald 1995; Swartz 1998).Army ants are suspected to preclude successful colony es-tablishment among
Atta
by frequently preying on thebrood and thereby undermining the reproductive poten-tial of the colonies. Army ant colonies require large, con-tiguous areas (up to 30 ha) and are among the first todisappear from smaller isolates following habitat fragmen-tation (Lovejoy et al. 1980; Franks 1983). Armadillos aresuspected to use their powerful foreclaws to dig up incip-ient and young
Atta
colonies (Weber 1972; Schade 1973).Home-range estimates for the common nine-banded arma-dillo (
Dasypus novemcinctus
) average 3.5 ha for adultsand explains their absence on small (
�
3.0 ha) Lago Guriislands (Breece & Dusi 1985). Phorid parasitoids associ-ated with leaf-cutter ants are known to deposit a singleegg in the head of
Atta
workers and to influence
Atta
for-aging rhythms (Orr 1992). Large land masses (mainlandand large islands) in our study area had all three predators(army ants, armadillos, and phorids) present at “natural”abundance levels; medium islands lacked army ants butretained armadillos and phorids, and small islands did notsupport army ants or armadillos, but some retained lowpopulations of phorids.
We examined 12 islands: 6 small (0.3–3.0 ha), 4 me-dium (7–12 ha), and 2 large (control; 250-ha Isla Grandeand 350-ha Danto Machado). A nearby mainland site, lo-cated on a peninsula, served as a third control site. Siteselection was based on land-mass size and not
Atta
den-sity, which was high on most small islands (Rao 2000).
The habitat of all study islands and the mainland issemideciduous tropical forest. The canopy is low, between15 and 20 m, with scattered emergents rising above 25 m.The climate is markedly seasonal, being dry from De-cember through April and wet from May through No-vember. Annual rainfall is 1100 mm at the Raul Leonidam 60 km to the north of the study area and increasestoward the south (Alvarez et al. 1986). Further informa-tion on the structure and composition of the forest atLago Guri is provided by Huber (1986) and Aymard et al.(1997).
Vegetation Plots
ADULT
TREE
AND
SMALL
-
STEM
PLOTS
Adult trees (
�
10 cm diameter at breast height [dbh]) andsmall stems (
�
1 m tall and
�
10 cm dbh) were mapped,measured, and tagged at all 13 sites. Every adult tree was
tagged on the six small islands. On other land masses,with the exception of one plot with 200 trees on thelarge island (Danto Machado), all other samples includedat least 300 trees, which is the approximate number oftrees in a hectare of dry forest at Lago Guri ( J.T., unpub-lished data). Plots on the small islands, unlike those atother sites, overlapped active ant-colony foraging zones.A single plot characterized all sites except for one of thelarge islands (Danto Machado, three plots) and the main-land (two plots). At most sites, the tree plots straddledthe highest point on the site so that one side faced intothe prevailing northeasterly wind (exposed) and theother side faced away (sheltered).
Two small stem subplots, each 15
�
15 m (225 m
2
),were located within the 1-ha mature tree plot at eachsite (only one small stem plot on Lomo, a medium is-land). Within the subplots, all small stems
�
1 m tall and
�
10 cm dbh were mapped, measured, and tagged.These were subsequently analyzed as two different sizeclasses, which we refer to as saplings (stems
�
1 m talland
�
1 cm dbh) and juveniles (stems
�
1 cm dbh and
�
10 cm dbh). All measurements were made 9 years af-ter isolation, making it likely that some to many of thesaplings were established after isolation of the islands,whereas many, if not all, juveniles probably were estab-lished prior to isolation. Vouchers of all plant specieswere collected and deposited in the Herbario Nacionalin Caracas and the Herbario Vargas in Cusco, Perú.
Data analyses were conducted with MathSoft Statisti-cal Software (1998). The three levels of the treatment“land-mass size” were: (1) small islands with high
Atta
densities (
n
�
6; Baya, Colón, Cola, Iguana, Palizada, andPerímetro); (2) medium islands with intermediate
Atta
densities (
n
�
4; Ambar, Chotacabra, Lomo, and Pan-orama), and (3) large land masses with low
Atta
densi-ties (
n
�
3, mainland, Isla Grande, and Danto Machado).We used a two-way analysis of variance (ANOVA) to ana-lyze data on saplings and juveniles. Land-mass category(small, medium, or large) and stem size (sapling or juve-nile) were the two fixed factors in the model.
ANT
-
COLONY
FORAGING
ZONES
Ant-colony foraging zones were demarcated by connect-ing the distal limits of all foraging trails used by focal
Atta
colonies during the study period (Rockwood & Hubbell1987). Every tree of
�
5 cm dbh within the foraging zoneof each colony was mapped, tagged, measured, and iden-tified. Nine such plots were surveyed: four plots on thelarge island (Danto Machado) and five on small islands(two each on Iguana and Búmeran and one on Colón).
Selectivity Experiment
The preferences by
Atta
colonies for particular plant spe-cies were measured during a 7-week period from late

Conservation BiologyVolume 15, No. 3, June 2001
Rao et al. Herbivory in Forest Isolates
627
June 1997 to mid-August 1997. The relative palatabilityof leaves to leaf-cutter ants is not expected to changesignificantly over so short a time period (Howard 1987).We used leaves of 43 common tree species found in thestudy area, which constitute approximately 44% of thetotal number of species (95) found across all the ant-col-ony foraging zones and 25% of the cumulative speciestotal (168) found in all the tree plots. We collected leavesfrom terminal branchlets by snipping off the petioles atthe point where they joined the stem.
During each trial, entire leaves of 21 plant species(one leaf per species) were placed in random successivesequence (20 cm apart) along an ant foraging trail at adistance of 2 m from the nest entrance, where they wereexposed to outbound foragers (Howard 1988). All trialslasted 1 hour and were completed within 3 hours of thetime of leaf collection to limit changes in leaf freshness.At the end of each 15-minute period, we recorded thenumber of leaf fragments cut from each plant speciesand removed to the nest entrance. We used frosted ce-real flakes as controls, and noted the number of flakesremoved during the same time period. This measure-ment was made to control for variation in activity levelsbetween sampling periods.
We counted the number of outbound ants passing afixed point on the trail during three 1-minute periods atthe end of each 15-minute period. Leaves that werecompletely cut and removed were replaced immedi-ately. At the end of every 15 minutes, the sequence ofleaves was altered to ensure uniform access and encoun-ter rates for all the leaves. The experiment was con-ducted during the peak activity period for two colonieson a large island (Danto Machado) and four colonies onsmall islands (Iguana, Baya, Colón, and Facil).
We conducted 34 trials of the experiment on small is-lands (Baya,
n
�
9; Colón,
n
�
17; Facil,
n
�
4; Iguana,
n
� 4) and 22 trials on one large island (Danto1, n � 12;Danto2, n � 10). The probability of the same ant en-countering the same species in two trials was low, whicheliminates the problem of pseudoreplication (Hurlbert1984). Overall, there were at least four replicate obser-vations per plant species for every colony and 56 inde-pendent runs of the experiment. The response variable(i.e., number of fragments of each species removed dur-ing each trial), was assumed to have a Poisson distribu-tion. The logarithm of the mean number of fragments re-moved per species per trial was modeled as a linearfunction of two factors: plant species (43 levels) andtrial (56 levels) (S-plus software program). The equationthat describes the model is log (nij) � �I
i � Jj,
where nij is number of counts (i.e., fragments cut and re-moved) at level i of factor I and at level j of factor J, I isthe plant species (i � 1. . . . . . .43 levels), and J is thetrial ( j � 1. . . . . . . . . .56 levels). The generalized linearmodel (GLM) specified in S-Plus does not force data intounnatural scales and incorporates separate functions to
allow for nonlinearity and heterogeneous variances (Cham-bers & Hastie 1993). The GLM function fits the coefficientsof the Poisson regression model based on maximumlikelihood in the binomial family. We used the partial re-gression coefficients in the fitted model to rank species.The most-preferred species had the largest positive devi-ation from the mean, and the least-preferred species hadthe largest negative deviation from the mean.
We performed an ANOVA to test the significance ofthe fit. The function ANOVA in S-Plus fits a sequence ofmodels by successively adding each of the factors. For aPoisson model, the changes in the residual sum ofsquares between nested models are treated as chi-squarevariables.
Analyses of Selectivity Data
The 43 plant species tested for preference by Atta work-ers were divided into two groups: preferred (top 10ranks) and less-preferred species (bottom 10 ranks) (Ta-ble 1). The null hypothesis of equal representation ofpreferred and less-preferred species in both preadultstem-size categories was tested independently for eachlandmass size. We performed an analysis of covariance(ANCOVA), with stem density (number per 225 m2) asthe response variable, preference category and stem-sizecategory as the main factors, and adult tree density (totalnumber of adult trees of preferred and less-preferredspecies of �10 cm dbh/ha) as a covariate. There weretwo levels in each of the two factors: preference cate-gory (preferred and less preferred) and stem size (sap-lings and juveniles). All data were square-root trans-formed (n 1) prior to analysis. We predicted that ifAtta hyperabundance had a selective effect on plant spe-cies recruitment, small stems of preferred plant speciesshould be underrepresented on the small islands. Sap-lings that were more likely to have become establishedafter isolation should be more negatively affected thanjuveniles that existed prior to isolation.
To test whether exposure to high levels of Atta her-bivory selectively depletes preferred species, we com-pared the relative adult tree abundance of preferred andless-preferred species between Atta foraging zones andcontrol plots (mature tree plots on the mainland and thelarge island, Danto Machado, that lacked Atta colonies).We calculated the relative abundance of adult trees(number of individuals of preferred or less-preferredspecies, divided by total number of individuals of all spe-cies per plot) for each plot and arcsine square-root trans-formed prior to analysis. There were nine ant foragingzones, four on Danto Machado (large land mass), two onBúmeran, two on Iguana, and one on Colón (all smallland masses) to compare with five control plots (two onthe mainland and three on the large island, DantoMachado). We tested the null hypothesis of no differ-

628 Herbivory in Forest Isolates Rao et al.
Conservation BiologyVolume 15, No. 3, June 2001
ence in the mean relative abundance of preferred orless-preferred species between foraging zones and con-trol plots using an ANOVA with plot type (foraging zonevs. controls) and preference (preferred vs. less preferred)as the two main factors.
We examined variation in the relative abundance ofpreferred species among ant-colony plots by groupingant colony plots on Danto Machado (n � 4) with the large-island category (treatment 1) and plots on Búmeran,Iguana, and Colón (n � 5) with the second, small-islandcategory (treatment 2). We tested the null hypothesis ofno difference in the mean relative abundance of pre-ferred species between the two treatments using a one-
way ANOVA. Variances were stabilized by arcsine square-root transformations of proportions prior to analyses.
Results
Density and Size-Class Distribution of Saplings and Juveniles
Analysis of square-root transformed data (small stemdensity) revealed significant main and interaction effectsfor both factors, land mass type and stem-size class (F �4.33; df � 5, 44; p � 0.05). Tukey’s test for multiplecomparisons of means revealed significantly lower sap-
Table 1. Plant species rankings for 43 species derived from the selectivity experiment conducted to establish preference categories.*
Species Family Fitted coefficient Ranking
Apeiba schomburgkii Tiliaceae 1.762 1Centrolobium paraense Leguminosae 1.355 2Randia armata Rubiaceae 1.316 3Alseis lobatioides Rubiaceae 1.296 4Bauhinia aculeata Leguminosae 1.233 5Pradosia caracasana Sapotaceae 1.076 6Peltogyne floribunda Leguminosae 1.047 7Tabebuia serratifolia Bignoniaceae 1.027 8Lepidocordia punctata Boraginaceae 0.963 9Casearia spinescens Flacourtiaceae 0.926 10Bunchosia mollis Malphigiaceae 0.902 11Actinostemon schomburgkiana Euphorbiaceae 0.749 12Casearia decandra Flacourtiaceae 0.619 13Melicoccus bijugatus Sapindaceae 0.492 14Acalypha cuspidata Euphorbiaceae 0.333 15Talisia heterodoxa Sapindaceae 0.287 16Lonchocarpus sericeus Leguminosae 0.276 17Coccoloba fallax Polygonaceae 0.191 18Machaerium acuminatum Leguminosae 0.185 19Perama hirsuta Rubiaceae 0.152 20Boureria cumanensis Apocynaceae �0.062 21Coccoloba striata Polygonaceae �0.069 22Casearia ulmifolia Flacourtiaceae �0.072 23Luehea speciosa Tiliaceae �0.134 24Pouteria leucentifolium Sapotaceae �0.257 25Trichilia lepidota Meliaceae �0.274 26Picramnia pentandra Simaroubiaceae �0.351 27Protium neglectum Burseraceae �0.397 28Brownea coccinea Leguminosae �0.459 29Fissycalyx fendleri Leguminosae �0.529 30Ocotea glomerata Lauraceae �0.568 31Citharexylum espinosum Verbenaceae �0.637 32Rhinorea riana Violaceae �0.713 33Erythroxylon habanense Erythroxylaceae �0.760 34Capparis amplisema Capparidaceae �0.794 35Calliandra laxa Leguminosae �0.809 36Myrcia lucida Myrtaceae �0.851 37Guatteria schomburgkiana Annonaceae �0.982 38Neea bracteosa Nyctaginaceae �1.161 39Hymenaea courbaril Leguminosae �1.187 40Copaifera pubiflora Leguminosae �1.317 41Ilex guianensis Aquifoliaceae �1.327 42Eugenia ligustrina Myrtaceae �1.753 43
*There were 34 trials of the experiment on small islands (Baya, n � 9; Colón, n � 17; Facil, n � 4; Iguana, n � 4) and 22 trials on one large is-land (Danto1, n � 12; Danto2, n � 10).
Conservation BiologyVolume 15, No. 3, June 2001
Rao et al. Herbivory in Forest Isolates 629
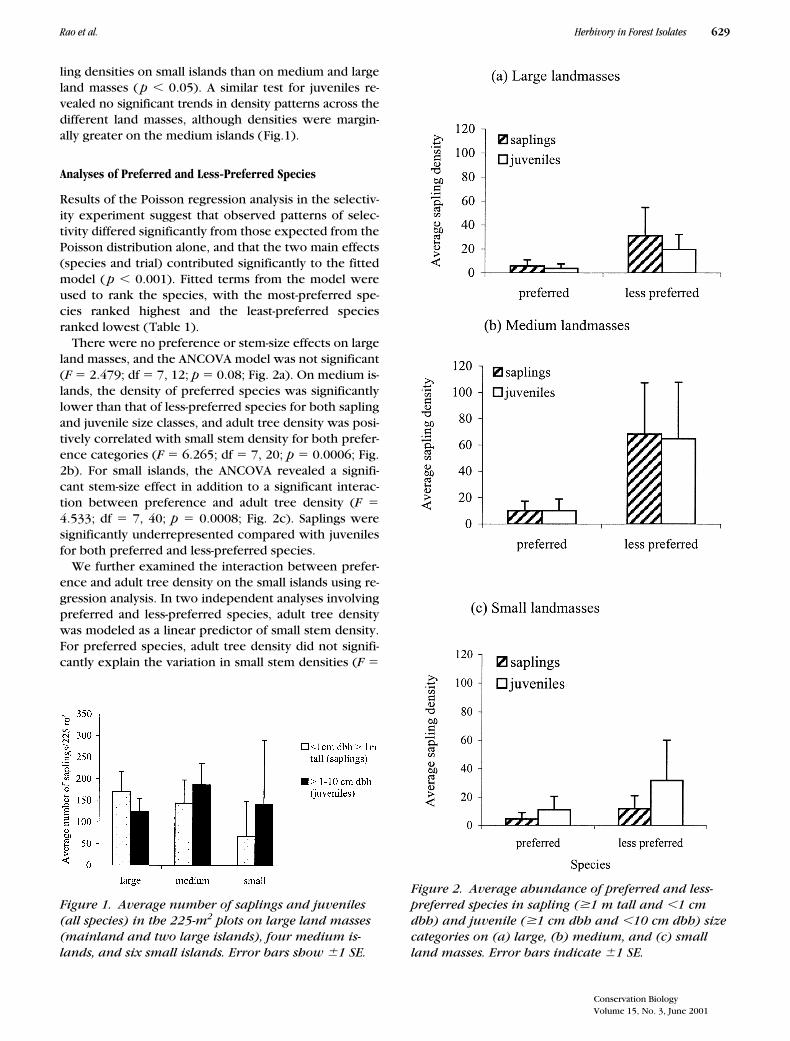
ling densities on small islands than on medium and largeland masses ( p � 0.05). A similar test for juveniles re-vealed no significant trends in density patterns across thedifferent land masses, although densities were margin-ally greater on the medium islands (Fig.1).
Analyses of Preferred and Less-Preferred Species
Results of the Poisson regression analysis in the selectiv-ity experiment suggest that observed patterns of selec-tivity differed significantly from those expected from thePoisson distribution alone, and that the two main effects(species and trial) contributed significantly to the fittedmodel ( p � 0.001). Fitted terms from the model wereused to rank the species, with the most-preferred spe-cies ranked highest and the least-preferred speciesranked lowest (Table 1).
There were no preference or stem-size effects on largeland masses, and the ANCOVA model was not significant(F � 2.479; df � 7, 12; p � 0.08; Fig. 2a). On medium is-lands, the density of preferred species was significantlylower than that of less-preferred species for both saplingand juvenile size classes, and adult tree density was posi-tively correlated with small stem density for both prefer-ence categories (F � 6.265; df � 7, 20; p � 0.0006; Fig.2b). For small islands, the ANCOVA revealed a signifi-cant stem-size effect in addition to a significant interac-tion between preference and adult tree density (F �4.533; df � 7, 40; p � 0.0008; Fig. 2c). Saplings weresignificantly underrepresented compared with juvenilesfor both preferred and less-preferred species.
We further examined the interaction between prefer-ence and adult tree density on the small islands using re-gression analysis. In two independent analyses involvingpreferred and less-preferred species, adult tree densitywas modeled as a linear predictor of small stem density.For preferred species, adult tree density did not signifi-cantly explain the variation in small stem densities (F �
Figure 1. Average number of saplings and juveniles (all species) in the 225-m2 plots on large land masses (mainland and two large islands), four medium is-lands, and six small islands. Error bars show 1 SE.
Figure 2. Average abundance of preferred and less-preferred species in sapling (�1 m tall and �1 cm dbh) and juvenile (�1 cm dbh and �10 cm dbh) size categories on (a) large, (b) medium, and (c) small land masses. Error bars indicate 1 SE.
630 Herbivory in Forest Isolates Rao et al.
Conservation BiologyVolume 15, No. 3, June 2001
0.2774; df � 1, 22; p � 0.6). There was, however, a sig-nificant negative relationship between density of adulttrees and small stems of less-preferred species (F � 6.363;df � 1, 22; p � 0.02).
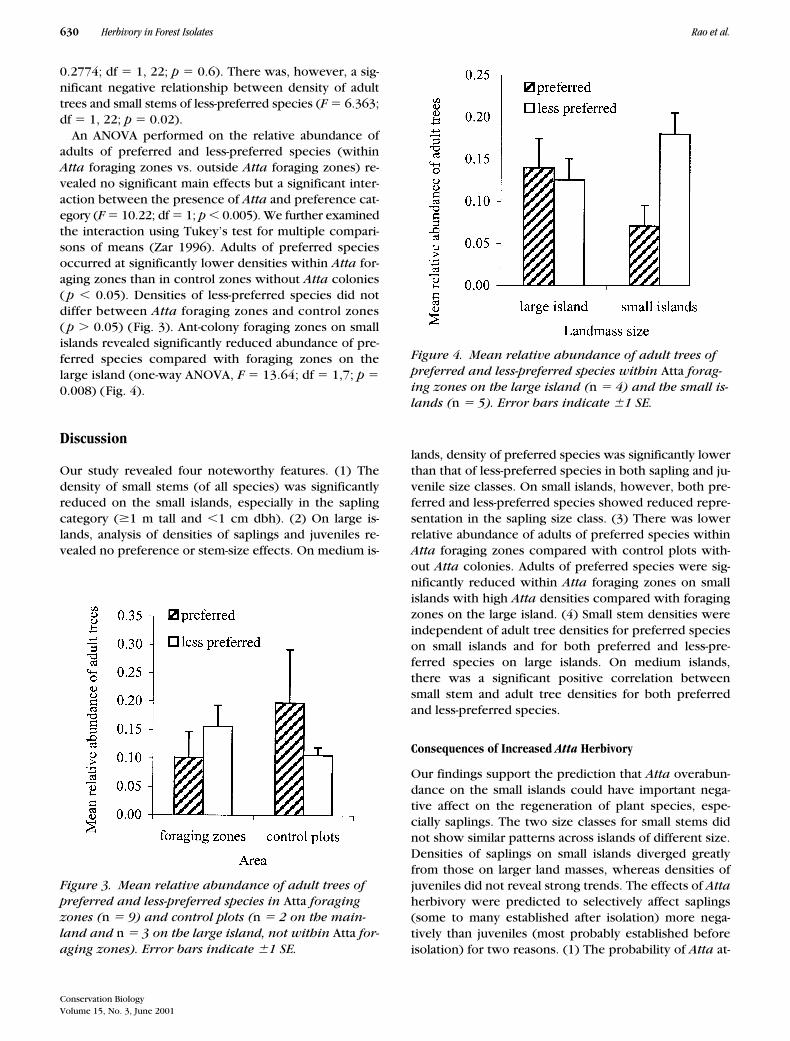
An ANOVA performed on the relative abundance ofadults of preferred and less-preferred species (withinAtta foraging zones vs. outside Atta foraging zones) re-vealed no significant main effects but a significant inter-action between the presence of Atta and preference cat-egory (F � 10.22; df � 1; p � 0.005). We further examinedthe interaction using Tukey’s test for multiple compari-sons of means (Zar 1996). Adults of preferred speciesoccurred at significantly lower densities within Atta for-aging zones than in control zones without Atta colonies( p � 0.05). Densities of less-preferred species did notdiffer between Atta foraging zones and control zones( p � 0.05) (Fig. 3). Ant-colony foraging zones on smallislands revealed significantly reduced abundance of pre-ferred species compared with foraging zones on thelarge island (one-way ANOVA, F � 13.64; df � 1,7; p �0.008) (Fig. 4).
Discussion
Our study revealed four noteworthy features. (1) Thedensity of small stems (of all species) was significantlyreduced on the small islands, especially in the saplingcategory (�1 m tall and �1 cm dbh). (2) On large is-lands, analysis of densities of saplings and juveniles re-vealed no preference or stem-size effects. On medium is-
lands, density of preferred species was significantly lowerthan that of less-preferred species in both sapling and ju-venile size classes. On small islands, however, both pre-ferred and less-preferred species showed reduced repre-sentation in the sapling size class. (3) There was lowerrelative abundance of adults of preferred species withinAtta foraging zones compared with control plots with-out Atta colonies. Adults of preferred species were sig-nificantly reduced within Atta foraging zones on smallislands with high Atta densities compared with foragingzones on the large island. (4) Small stem densities wereindependent of adult tree densities for preferred specieson small islands and for both preferred and less-pre-ferred species on large islands. On medium islands,there was a significant positive correlation betweensmall stem and adult tree densities for both preferredand less-preferred species.
Consequences of Increased Atta Herbivory
Our findings support the prediction that Atta overabun-dance on the small islands could have important nega-tive affect on the regeneration of plant species, espe-cially saplings. The two size classes for small stems didnot show similar patterns across islands of different size.Densities of saplings on small islands diverged greatlyfrom those on larger land masses, whereas densities ofjuveniles did not reveal strong trends. The effects of Attaherbivory were predicted to selectively affect saplings(some to many established after isolation) more nega-tively than juveniles (most probably established beforeisolation) for two reasons. (1) The probability of Atta at-
Figure 3. Mean relative abundance of adult trees of preferred and less-preferred species in Atta foraging zones (n � 9) and control plots (n � 2 on the main-land and n � 3 on the large island, not within Atta for-aging zones). Error bars indicate 1 SE.
Figure 4. Mean relative abundance of adult trees of preferred and less-preferred species within Atta forag-ing zones on the large island (n � 4) and the small is-lands (n � 5). Error bars indicate 1 SE.

Conservation BiologyVolume 15, No. 3, June 2001
Rao et al. Herbivory in Forest Isolates 631
tack on a small stem could decline with time, thus allow-ing the “escape” and continued persistence of individualstems in taller height categories (de Vasconcelos & Cher-rett 1997). (2) Taller (and older) stems may accumulateenergy reserves that help them survive defoliation, thusmaking them more tolerant to herbivory. Our results aregenerally compatible with other evidence of destructiveforaging by overabundant herbivores in terrestrial sys-tems (Merton et al. 1976; Oksanen 1983; Alverson et al.1988, 1994; McLaren & Peterson 1994).
Implications of Selective Herbivory for Plant Recruitment
The selectivity experiment provided a rigorous assess-ment of species preference and indicated that someplant species were consistently preferred over others.Other studies have used similar experiments to examinedeterminants underlying the selection process (Hubbellet al. 1984; Howard 1987, 1988). Results of these analy-ses need to be evaluated within limitations of the selec-tivity experiment. Observed trends in the representationof preferred and less-preferred species are valid only ifpreferences derived from the selectivity experiment re-flect actual preferences in natural settings. Experimen-tally derived species preferences in our study were com-parable with preferences recorded during natural foragingobservations. Rankings of certain preferred species (Bau-hinia aculeata, Pradosia caracasana, Machaerium acu-minatum, and Casearia spinescens) and some less-pre-ferred species (Hymenaea courbaril, Eugenia ligustrina,Calliandra laxa, Luehea speciosa, Guatteria schom-burkiana, Faessicalis fendleri, Neea bracteosa, Boureriacumanensis, and Picramnia pentandra) were fairly con-sistent with rankings obtained from natural foraging ob-servations (Rao 1998).
Hubbell et al. (1984) showed that seasonality influ-ences species selection through its effects on the con-centrations of secondary compounds. The majority ofspecies in their study showed a loss or substantial reduc-tion of repellency toward the end of the wet season, andHubbell et al. suggest that this loss is probably due to adecrease in the synthesis of antifungal compounds. Theselectivity experiment in our study was conducted dur-ing the middle of the wet season. Therefore, some of thespecies classified as less preferred could become morepalatable toward the end of the wet season, which per-haps explains some of the observed variability in therepresentation of less-preferred species.
Although small stems of preferred species were signif-icantly reduced in the understory of medium and largeland masses, there were no accompanying stem-size ef-fects as observed on the small islands. The lack of a sig-nificant interaction between stem size and preferencecategory on small Guri islands precludes firm conclu-sions regarding the effects of selective herbivory on theregeneration of preferred species. The reduction in sap-
ling density of both preferred and less-preferred specieson small islands could result from less-selective foragingat the early stages of sapling growth. Leaf-cutter ants pre-fer young leaves over mature leaves (Littledyke & Cher-rett 1978; Nichols-Orians & Schultz 1989, 1990). Conse-quently, selection based on leaf toughness does notdifferentiate between young leaves of preferred and less-preferred species and may explain the reduced repre-sentation of saplings of both preference categories. Fur-thermore, at densities of approximately 5/ha, colonieson the small islands experience strong intraspecific com-petition for limited food resources and, as predicted byoptimal foraging theory, may broaden their diets to in-clude less-preferred species (Emlen 1966; MacArthur &Pianka 1966; Schoener 1971). Natural foraging observa-tions also suggest that where they are overabundant,leaf-cutter ants are less selective than where they occurat the low densities typical of large land masses. Colo-nies on the small islands frequently forage on less-pre-ferred species, such as Erythroxylon habanense andBoureria cumanensis, that are rarely utilized by colo-nies on the large island (Rao 1998).
Our findings are nevertheless consistent with those ofan earlier study that showed reduced mortality of tallerseedlings compared with smaller seedlings and greatermortality of species preferred by leaf-cutter ants (de Vas-concelos & Cherrett 1997). Garrettson et al. (1998) alsofound reduced diversity and abundance of understoryplants in the vicinity of active leaf-cutter ant nests in aCosta Rican rainforest.
Atta foraging zones are defined by a trunk trail systemthat directs foraging effort to the most productive partsof the home range and facilitates rediscovery and contin-ued use of resources (Shepherd 1982). Shifts in foragingzones caused by variation in resource availability aretherefore represented in the trail system. Although Attaforaging zones in our study did not exhaustively includethe area utilized by the colony in the past, they never-theless represented the core foraging area of the colony.That adult densities of preferred tree species were lowerwithin Atta foraging zones than in tree plots lackingAtta colonies could be attributed to mortality caused byrepeated exposure to defoliation by Atta. Observed pat-terns of lower adult densities of preferred species inAtta foraging zones on small islands than on the large is-land are perhaps related to high levels of Atta herbivoryon small Guri islands. Herbivory-induced mortality ofadult trees could ultimately result in reduced regenera-tion of preferred species.
The relationship between the density of adult treesand small stems of preferred and less-preferred speciesvaried with island size. On large islands there was no sig-nificant correlation between adult tree density and smallstem densities for either preferred or less-preferred spe-cies. On medium islands there was a significant positivecorrelation between adult tree and small stem densities

632 Herbivory in Forest Isolates Rao et al.
Conservation BiologyVolume 15, No. 3, June 2001
for species at both ends of the preference spectrum. Onsmall islands, small stem densities of preferred specieswere independent of adult densities, whereas there wasa negative relationship between small stem and adultdensities among less-preferred species. These results areperhaps best explained by considering that fire occur-rence continued after isolation on larger land masses,while on medium and small islands there had been fewor no fires since the project began in 1993 (M.R., per-sonal observations). The relative effects of fire and her-bivory on understory species composition may be per-ceived as varying in opposing directions with island size.Although fire constitutes an important factor on thelarger land masses than on small and medium islands, in-creased levels of herbivory by overabundant Atta couldhave stronger effects on small islands than on larger landmasses. Mechanisms underlying the observed negativerelationship between adult tree and small stem densitiesof less-preferred species on small islands needs furtherexamination, however. There are some important cave-ats to consider in the interpretation of our results. Com-parisons between small islands with and without Attacolonies would have been ideal to control for highermortality of small stems and adult trees on wind-exposedsmall islands (Laurance 1997). Although the observedtrends are in the predicted direction, more detailed re-search on small stem and adult tree mortality will beneeded to remove possible confounding influences. Thesmall Guri islands support high populations of granivo-rous rodents, so selective seed predation could poten-tially interact with the effects of selective herbivory toultimately determine plant regeneration patterns (Ter-borgh et al. 1997).
Conservation Implications
Fragmentation unleashes complex, multitrophic-leveldistortions that portend long-term implications for plantspecies diversity and community composition (Ter-borgh 1992). Although reduction in species diversitywith habitat size is widely acknowledged, biologicalmechanisms driving population declines remain poorlyunderstood (Caughley 1994). Our study contributes toan understanding of how habitat loss alters ecological re-lationships, thereby driving a process of change in com-munity composition (Brown et al. 1986). Our findingsreaffirm the need for large, unfragmented protected ar-eas in which undistorted ecological processes can act toconserve biological diversity.
Acknowledgments
We are grateful to L. Balbas for generous logistical andadministrative support. We thank Electrificación del Ca-roní (EDELCA) and Consejo Nacional de Investigaciones
Científicas y Tecnológicas (CONICIT) for permits neces-sary to conduct research in the Guri impoundment. D.Higdon provided valuable statistical advice. Commentsby W. Morris, L. Mojonnier, and C.van Schaik helped im-prove the manuscript. We warmly thank C. Aponte, M.Riveros, O. Cornejo, N. Ceballos, L. Davenport, E. De-Mattia, P. Hankenson, G. Herzner, and G. Shahabuddinfor field assistance and two anonymous reviewers forsuggestions on the manuscript. We gratefully acknowl-edge financial support from Sigma Xi (M.R.) and the Na-tional Science Foundation (grant DEB 9707281 to J.T.).
Literature Cited
Alvarez, E., L. Balbas, I. Massa, and J. Pacheco. 1986. Aspectos ecologi-cos del embalse de Guri. Interciencia 11:325–333.
Alverson, W. S., D. M. Waller, and S. L. Solheim. 1988. Forests toodeer: edge effects in northern Wisconsin. Conservation Biology 2:348–358.
Alverson, W. S., W. Kuhlmann, and D. M. Waller. 1994. Wild forests: con-servation biology and public policy. Island Press, Washington, D.C.
Aymard, G., M. Norconk, and W. Kinzey. 1997. Composicion floristicade comunidades vegetales en islas en el embalse de Guri, Rio Ca-roní, Estado Bolivar, Venezuela. UNELLEZ-Guanare, ProgramaR.N.R. Herbario Universitario (PORT) Portuguesa, Venezuela.
Blanton, C. M., and J. J. Ewel. 1985. Leaf-cutter ant herbivory in succes-sional and agricultural tropical ecosystems. Ecology 66:861–869.
Breece, G. A., and J. L. Dusi. 1985. Food habits and home ranges of thecommon long-nosed armadillos Dasypus novemcinctus in Ala-bama. Pages 419–427 in G. G. Montgomery, editor. The evolutionand ecology of armadillos, sloths and vermilinguas. Smithsonian In-stitution Press, Washington, D.C.
Brian, M. V. 1983. Social insects: ecology and behavioural biology.Chapman and Hall, London.
Brown, J. H. 1971. Mammals on mountaintops: nonequilibrium insularbiogeography. The American Naturalist 105:467–478
Brown, J. H., D. W. Davidson, J. C. Munger, and R. S. Inouye. 1986. Ex-perimental community ecology: the desert granivore system. Pages41–61 in J. Diamond and T. J. Case, editors. Community ecology.Harper and Row, New York.
Caughley, G. 1994. Directions in conservation biology. Journal of Ani-mal Ecology 63:215–244.
Chambers, J. M., and T. J. Hastie. 1993. Statistical models in S. Chap-man & Hall, New York.
de Vasconcelos, H. L., and J. M. Cherrett. 1997. Leaf-cutting ants andearly forest regeneration in central Amazonia: effects of herbivoryon tree seedling establishment. Journal of Tropical Ecology 13:357–370.
Diamond, J. M. 1984. “Normal” extinction of isolated populations.Pages 191–246 in M. H. Nitecki, editor. Extinctions. Chicago Uni-versity Press, Chicago.
Emlen, J. M. 1966. The role of time and energy in food preference. TheAmerican Naturalist 100:611–617.
Feener, D. H., and B. V. Brown. 1993. Oviposition behaviour of an ant-parasitizing fly, Neodohrniphora curvinervis (Diptera: Phoridae),and defense behaviour by its leaf-cutting ant host Atta cephalotes(Hymenoptera: Formicidae). Journal of Insect Behaviour 6:675–687.
Franks, N. R. 1983. Ecology and population regulation in the army antEciton burchelli. Pages 389–394 in E. G. Leigh, A. S. Rand, andD. W. Windsor, editors. Ecology of a tropical rainforest: seasonalrhythms and long-term changes. Smithsonian Institution Press,Washington, D.C.
Garrettson, M., J. F. Stetzel, B. S. Halpern, D. J. Hearn, B. T. Lucey, andM. J. McKone. 1998. Diversity and abundance of understory plants

Conservation BiologyVolume 15, No. 3, June 2001
Rao et al. Herbivory in Forest Isolates 633
on active and abandoned nests of leaf-cutting ants (Atta cephalotes)in a Costa Rican rain forest. Journal of Tropical Ecology 14:17–26.
Gotwald, W. H., Jr. 1995. Army ants: the biology of social predation.Cornell University Press, New York.
Heaney, L. R. 1984. Mammalian species richness on islands on theSunda Shelf, Southeast Asia. Oecologia 61:11–17.
Howard, J. J. 1987. Leafcutting ant diet selection: the role of nutrients,water, and secondary chemistry. Ecology 68:503–515.
Howard, J. H. 1988. Leafcutting ant diet selection: relative influence ofleaf chemistry and physical chemistry. Ecology 69:250–260.
Hubbell, S. P., J. J. Howard, and D. F. Wiemer. 1984. Chemical leaf re-pellency to an Attine ant: seasonal distribution among potentialhost plant species. Ecology 65:1067–1076.
Huber, O. 1986. La vegetación de la cuenca del Rio Caroní. Intercien-cia 11:301–310.
Hurlbert, S. H. 1984. Pseudoreplication and the design of ecologicalfield experiments. Ecological Monographs 54:187–211.
Laurance, W. F. 1997. Hyper-disturbed parks: edge effects and theecology of isolated rainforest reserves in tropical Australia. Pages71–84 in W. F. Laurance and R. O. Bierregaard Jr., editors. Tropicalforest remnants: ecology, management and conservation of frag-mented communities. University of Chicago Press, Chicago.
Littledyke, M., and J. M. Cherrett. 1978. Defense mechanisms in youngand old leaves against cutting by the leaf-cutting ants (Atta cephal-otes (L.) and Acromyrmex octospinosus (Reich) (Hymenoptera:Formicidae). Bulletin for Entomological Research 68:263–271.
Lomolino, M. V., J. H. Brown, and R. Davis. 1989. Island biogeographyof montane forest mammals in the American Southwest. Ecology70:180–194.
Lovejoy, T. E., R. O. Bierregaard Jr., A. B. Rylands, J. R. Malcolm, C. E.Quintela, L. H. Harper, K. S. Brown Jr., A. H. Powell, G. V. N. Pow-ell, H. O. R. Schubart, and M. B. Hays, 1980. Edge and other effectsof isolation on Amazonian forest fragments. Pages 257–285 in M. E.Soule, editor. Conservation Biology: the science of scarcity and di-versity. Sinauer Associates, Sunderland, Massachusetts.
MacArthur, R. H., and E. R. Pianka. 1966. On optimal use of a patchyenvironment. The American Naturalist 100:603–609.
MacArthur, R. H., and E. O. Wilson. 1963. An equilibrium theory of in-sular zoogeography. Evolution 17:373–387.
MacArthur, R. H. and E. O. Wilson. 1967. The theory of island biogeog-raphy. Princeton University Press, Princeton, New Jersey.
MathSoft Statistical Software. 1998. S-Plus. Version 5.2. MathSoft, Seattle.McLaren, B. E., and R. O. Peterson. 1994. Wolves, moose and tree
rings on Isle Royale. Science 266:1555–1558.Merton, L. F. H., D. M. Bourn, and R. J. Hnatiuk. 1976. Giant tortoise
and vegetation interaction on Aldabra Atoll. 1. Inland. BiologicalConservation 9:293–304.
Morales, L. C., and S. Gorzula. 1986. The interrelations of the Caroníriver basin ecosystems and hydroelectric power projects. Inter-ciencia 11:272–277.
Nepstad, D. C., C. Uhl, C. A. Pereira, and J. M. Cardoso da Silva. 1996.A comparative study of tree establishment in abandoned pastureand mature forest of eastern Amazonia. Oikos 76:25–39.
Newmark, W. D. 1987. A land-bridge island perspective on mammalianextinctions in western North American parks. Nature 325:430–432.
Nichols-Orians, C. M., and J. C. Schultz. 1989. Leaf toughness affectsleaf harvesting by the leaf-cutter ant, Atta cephalotes (L.) (Hy-menoptera: Formicidae). Biotropica 21:80–83.
Nichols-Orians, C. M., and J. C. Schultz. 1990. Interactions among leaftoughness, chemistry, and harvesting by attine ants. Ecological En-tomology 15:311–320.
Oksanen, L. 1983. Trophic exploitation and Arctic phytomass pat-terns. The American Naturalist 122:45–52.
Orr, M. 1992. Parasitic flies (Diptera: Phoridae) influence foragingrhythms and caste division of labour in the leaf-cutter ant, Attacephalotes (Hymenoptera: Formicidae). Behavioural Ecology andSociobiology 30:395–402.
Rao, M. 1998. Relaxation in the Lago Guri islands, Venezuela: causesunderlying leaf-cutter ant (Atta sp.) hyperabundance and conse-quences for plant species diversity. Ph.D thesis. Duke University,Durham, North Carolina.
Rao, M. 2000. Variation in leaf-cutter ant (Atta sp.) densities in forestisolates: the potential role of predation. Journal of Tropical Ecology16:209–225.
Rockwood, L. L. 1973. The effect of defoliation on seed production ofsix Costa Rican tree species. Ecology 54:1363–1369.
Rockwood, L. L. and S. P. Hubbell. 1987. Host-plant selection, diet di-versity and optimal foraging in a tropical leaf-cutting ant. Oecologia74:55–61.
Schade, F. H. 1973. The ecology and control of the leaf-cutting ants ofParaguay. Pages 77–93 in J. R. Gorham, editor. Paraguay: ecologicalessays. Academy of the Arts and Sciences of the Americas, Miami.
Schoener, T. W. 1971. Theory of feeding strategies. Annual Review ofEcology and Systematics 2:369–404.
Shepherd, J. D. 1982. Trunk trails and the searching strategy of a leaf-cutter ant, Atta colombica. Behavioural Ecology & Sociobiology11:77–84.
Swartz, M. 1998. Predation on an Atta cephalotes colony by an armyant, Nomamyrmex esenbeckii. Biotropica 30:682–684.
Terborgh, J. 1992. Maintenance of diversity in tropical forests. Biotro-pica 24:283–292.
Terborgh, J., L. Lopez, J. Tello, D. Yu, and A. R. Bruni. 1997. Transitorystates in relaxing ecosystems of land-bridge islands. Pages 256–279in W. F. Laurance and R. O. Bierregaard Jr., editors. Tropical forestremnants: ecology, management and conservation of fragmentedcommunities. University of Chicago Press, Chicago.
Weber, N. A. 1972. Gardening ants. The attines. American Philosophi-cal Society, Philadelphia.
Zar, J. H. 1996. Biostatistical analysis. Prentice-Hall, New Jersey.
Related Documents











