Copyright 2010 by ESPGHAN and NASPGHAN. Unauthorized reproduction of this article is prohibited. Increased Heat Shock Protein 72 Expression in Celiac Disease z Erna Sziksz, Ga´borVeres, z A ´ da´mVannay, A ´ gnesPro´kai, KrisztinaGa´l, Anna O ´ nody, y Ilma Rita Korponay-Szabo´, Gyo¨rgyReusz, Andra´s Szabo´, Tivadar Tulassay, Andra´s Arato´, and z Bea´taSzebeni ABSTRACT Background and Objectives: Heat shock protein (HSP) 72, a known chaperone, has potential epithelial barrier protecting, antiapoptotic, and immune system regulatory effects; therefore, our aim was to study its involvement in the pathology of celiac disease (CD). Patients and Methods: Duodenal biopsy specimens were collected from children with untreated and treated CD and from controls. mRNA expression, protein level, and localization of HSP72 were determined. Results: Elevated HSP72 mRNA expression and higher protein levels were found in the duodenal mucosa of children with untreated CD as well as in children with treated CD compared with those in controls. In the duodenal mucosa of children with treated CD, HSP72 mRNA expression was decreased and HSP72 protein levels were lower than those in children with untreated CD. We detected intensive HSP72 staining in the villous enterocytes and immune cells of the lamina propria in the duodenal villi of children with untreated CD compared with that in controls. Conclusions: The increased expression and altered localization of HSP72 in CD indicate that HSP72 should have a role in protection against gliadin- induced cytotoxicity. HSP72 may exert antiapoptotic effect and contribute to preservation of intestinal epithelial barrier integrity. Moreover, HSP72 as a ligand of TLR2 and TLR4 may promote innate immune responses and warn the cells of the potential injury. Key Words: children, celiac disease, duodenal biopsies, heat shock protein 72 (JPGN 2010;51: 573–578) C eliac disease (CD) is an immunologically mediated entero- pathy of the small intestine, characterized by lifelong intol- erance to gliadin and related prolamines (1). The essential function of the intestinal epithelium is the maintenance of a selective barrier through which nutrients and electrolytes can permeate, whereas potentially harmful agents are excluded (2). In CD the structure of this barrier is damaged, which leads to the leakage of cereal proteins across the intestinal epithelium. Wheat gliadin then activates both the innate and the adaptive immune system (3 – 5) and increases the apoptosis (6) of enterocytes. Heat shock protein (HSP) 72 (7–8) was first described as a molecular chaperone induced by increased temperature. Later it was demonstrated that several other physiological or pathological stress factors can induce the synthesis of HSP72. HSP72 may alter diverse biological functions such as the inflammatory and the apoptotic processes. Increased intracellular HSP72 is expressed on the cell surface and is subsequently released into the circulation (9). Circulating HSP72 binds to antigen-presenting cells (APCs) and stimulates proinflammatory cytokine synthesis (10), chemokine (11), and reactive oxygen species release (12). Recent findings demonstrate that HSP72s are ligands for Toll-like receptors (TLRs), which play a crucial role in the defense mechanism of the innate immune system (13). HSP72 may exert immunoregulatory effects by binding especially to TLR2 and TLR4 on APCs (14). Moreover, using human colonic Caco-2/bbe (C2) cells the activation of caspase-9, a key early step leading to apoptosis, occurs earlier when cells express low levels of HSP72 (15). It suggests that HSP72 may also have a protective function through its antiapoptotic effect (16). On the basis of the interaction of increased mucosal expres- sion of TLR2 and TLR4 in CD (17) and potential immunostimu- latory and antiapoptotic features of HSP72 we investigated the modulatory role of this protein in CD. PATIENTS AND METHODS Patients Duodenal biopsy samples from 16 children with untreated (boys 6, girls 10; median age 6.7 years [3.7–13.9]) and 9 with treated (boys 4, girls 5; median age 6.7 years [4.9–12.7]) CD were collected. Biopsy samples of 7 children with untreated CD were taken at the time of diagnosis, before the introduction of a gluten- free diet. From the other 9 children duodenal biopsies were obtained before (untreated CD) and 1.5 years (range 1.1– 2.5) after exclusion of gluten from the diet (treated CD) (Table 1). The diagnosis of CD was based on the European Society for Pediatric Gastroenterology, Hepatology, and Nutrition criteria (18). All of the untreated patients with CD had anti-endomysium IgA positivity and subtotal villous atrophy of the intestinal mucosa. In cases of treated patients with CD full clinical remission was observed on the diet and no serum Received September 15, 2009; accepted May 27, 2010. From the First Department of Pediatrics, Semmelweis University, the y Department of Gastroenterology-Nephrology, Heim Pal Children’s Hospital, and the z Research Group for Pediatrics and Nephrology, Semmelweis University and Hungarian Academy of Sciences, Budapest, Hungary. Address correspondence and reprint requests to Erna Sziksz, Research Group for Pediatrics and Nephrology, Semmelweis University and Hungarian Academy of Sciences, H-1083 Budapest, Bo ´kay J. u. 53-54, Hungary (e-mail: [email protected]). Erna Sziksz and Ga ´bor Veres contributed equally to this work. This work was supported by funds from the Hungarian National Scientific Research Foundation grant (OTKA 71730, T046082, OTKA-K81117) and grant of the Ministry of Health (ETT 435/2006; ETT- 028-02), and TA ´ MOP-4.2.2-08/1/KMR-2008-0004. The authors report no conflicts of interest. Copyright # 2010 by European Society for Pediatric Gastroenterology, Hepatology, and Nutrition and North American Society for Pediatric Gastroenterology, Hepatology, and Nutrition DOI: 10.1097/MPG.0b013e3181ea0092 ORIGINAL ARTICLE:GASTROENTEROLOGY JPGN Volume 51, Number 5, November 2010 573

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Co
ORIGINAL ARTICLE: GASTROENTEROLOGY
Increased Heat Shock Protein 72 Expression in
Celiac DiseasezErna Sziksz, �Gabor Veres, zAdam Vannay, �Agnes Prokai, �Krisztina Gal, �Anna Onody,
y � � �
Ilma Rita Korponay-Szabo, Gyorgy Reusz, Andras Szabo, Tivadar Tulassay,nd zBeata Szebeni �Andras Arato, apyright 2010 by ESPGHAN and NASPGHAN. Unauthorized repro
C eliac disease (CD) is an immunologically mediated entero-pathy of the small intestine, characterized by lifelong intol-
atrophy of the intestinCD full clinical remiss
Received September 15, 2009; accepted May 27, 2010.From the �First Department of Pediatrics, Semmelweis University, theyDepartment of Gastroenterology-Nephrology, Heim Pal Children’sHospital, and the zResearch Group for Pediatrics and Nephrology,Semmelweis University and Hungarian Academy of Sciences, Budapest,Hungary.
Address correspondence and reprint requests to Erna Sziksz, ResearchGroup for Pediatrics and Nephrology, Semmelweis University andHungarian Academy of Sciences, H-1083 Budapest, Bokay J. u. 53-54,Hungary (e-mail: [email protected]).
Erna Sziksz and Gabor Veres contributed equally to this work.This work was supported by funds from the Hungarian National Scientific
Research Foundation grant (OTKA 71730, T046082, OTKA-K81117)and grant of the Ministry of Health (ETT 435/2006; ETT- 028-02), andTAMOP-4.2.2-08/1/KMR-2008-0004.
The authors report no conflicts of interest.Copyright # 2010 by European Society for Pediatric Gastroenterology,
Hepatology, and Nutrition and North American Society for PediatricGastroenterology, Hepatology, and Nutrition
DOI: 10.1097/MPG.0b013e3181ea0092
JPGN � Volume 51, Number 5, November 2010
ABSTRACT
Background and Objectives: Heat shock protein (HSP) 72, a known
chaperone, has potential epithelial barrier protecting, antiapoptotic, and
immune system regulatory effects; therefore, our aim was to study its
involvement in the pathology of celiac disease (CD).
Patients and Methods: Duodenal biopsy specimens were collected from
children with untreated and treated CD and from controls. mRNA
expression, protein level, and localization of HSP72 were determined.
Results: Elevated HSP72 mRNA expression and higher protein levels were
found in the duodenal mucosa of children with untreated CD as well as in
children with treated CD compared with those in controls. In the duodenal
mucosa of children with treated CD, HSP72 mRNA expression was
decreased and HSP72 protein levels were lower than those in children
with untreated CD. We detected intensive HSP72 staining in the villous
enterocytes and immune cells of the lamina propria in the duodenal villi of
children with untreated CD compared with that in controls.
Conclusions: The increased expression and altered localization of HSP72 in
CD indicate that HSP72 should have a role in protection against gliadin-
induced cytotoxicity. HSP72 may exert antiapoptotic effect and contribute to
preservation of intestinal epithelial barrier integrity. Moreover, HSP72 as a
ligand of TLR2 and TLR4 may promote innate immune responses and warn
the cells of the potential injury.
Key Words: children, celiac disease, duodenal biopsies, heat shock protein
72
(JPGN 2010;51: 573–578)
erance to gliadin and related prolamines (1). The essential functionof the intestinal epithelium is the maintenance of a selective barrierthrough which nutrients and electrolytes can permeate, whereaspotentially harmful agents are excluded (2). In CD the structure ofthis barrier is damaged, which leads to the leakage of cereal proteinsacross the intestinal epithelium. Wheat gliadin then activates boththe innate and the adaptive immune system (3–5) and increases theapoptosis (6) of enterocytes.
Heat shock protein (HSP) 72 (7–8) was first described as amolecular chaperone induced by increased temperature. Later it wasdemonstrated that several other physiological or pathological stressfactors can induce the synthesis of HSP72. HSP72 may alter diversebiological functions such as the inflammatory and the apoptoticprocesses.
Increased intracellular HSP72 is expressed on the cell surfaceand is subsequently released into the circulation (9). CirculatingHSP72 binds to antigen-presenting cells (APCs) and stimulatesproinflammatory cytokine synthesis (10), chemokine (11), andreactive oxygen species release (12). Recent findings demonstratethat HSP72s are ligands for Toll-like receptors (TLRs), which play acrucial role in the defense mechanism of the innate immune system(13). HSP72 may exert immunoregulatory effects by bindingespecially to TLR2 and TLR4 on APCs (14).
Moreover, using human colonic Caco-2/bbe (C2) cells theactivation of caspase-9, a key early step leading to apoptosis, occursearlier when cells express low levels of HSP72 (15). It suggests thatHSP72 may also have a protective function through its antiapoptoticeffect (16).
On the basis of the interaction of increased mucosal expres-sion of TLR2 and TLR4 in CD (17) and potential immunostimu-latory and antiapoptotic features of HSP72 we investigated themodulatory role of this protein in CD.
PATIENTS AND METHODS
PatientsDuodenal biopsy samples from 16 children with untreated
(boys 6, girls 10; median age 6.7 years [3.7–13.9]) and 9 withtreated (boys 4, girls 5; median age 6.7 years [4.9–12.7]) CD werecollected. Biopsy samples of 7 children with untreated CD weretaken at the time of diagnosis, before the introduction of a gluten-free diet. From the other 9 children duodenal biopsies were obtainedbefore (untreated CD) and 1.5 years (range 1.1–2.5) after exclusionof gluten from the diet (treated CD) (Table 1). The diagnosis of CDwas based on the European Society for Pediatric Gastroenterology,Hepatology, and Nutrition criteria (18). All of the untreated patientswith CD had anti-endomysium IgA positivity and subtotal villous
duction of this article is prohibited.
al mucosa. In cases of treated patients withion was observed on the diet and no serum
573
Co
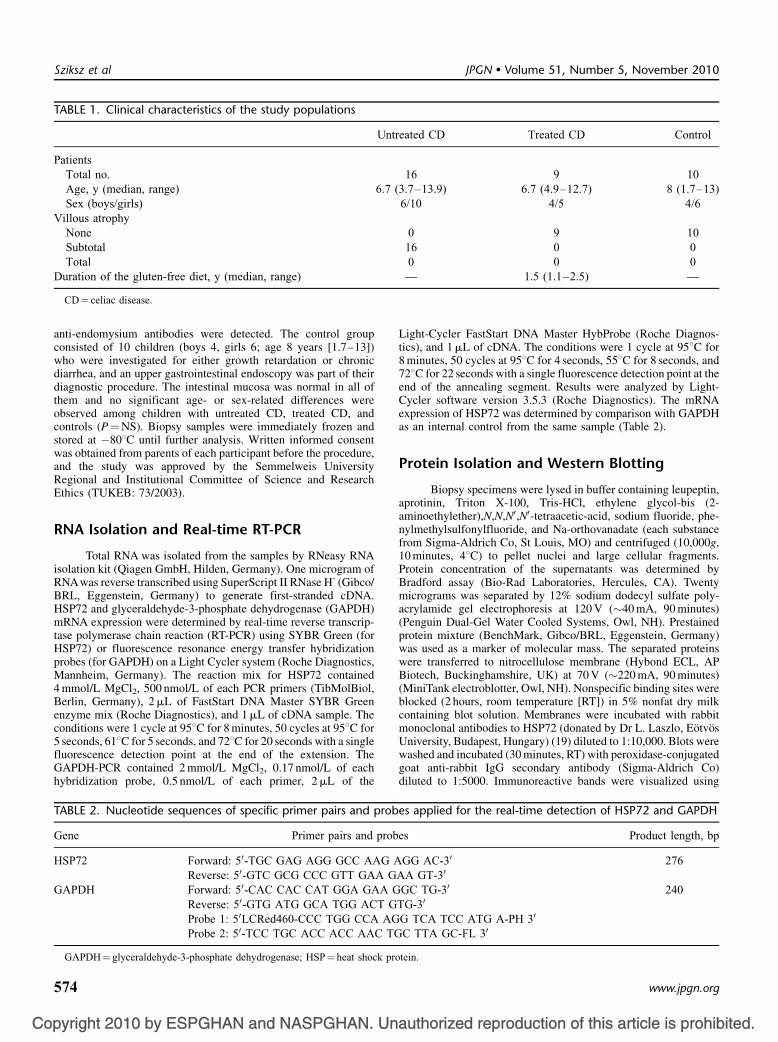
TABLE 1. Clinical characteristics of the study populations
Untreated CD Treated CD Control
PatientsTotal no. 16 9 10Age, y (median, range) 6.7 (3.7–13.9) 6.7 (4.9–12.7) 8 (1.7–13)Sex (boys/girls) 6/10 4/5 4/6
Villous atrophyNone 0 9 10Subtotal 16 0 0Total 0 0 0
Duration of the gluten-free diet, y (median, range) — 1.5 (1.1–2.5) —
Sziksz et al JPGN � Volume 51, Number 5, November 2010
anti-endomysium antibodies were detected. The control groupconsisted of 10 children (boys 4, girls 6; age 8 years [1.7–13])who were investigated for either growth retardation or chronicdiarrhea, and an upper gastrointestinal endoscopy was part of theirdiagnostic procedure. The intestinal mucosa was normal in all ofthem and no significant age- or sex-related differences wereobserved among children with untreated CD, treated CD, andcontrols (P¼NS). Biopsy samples were immediately frozen andstored at �808C until further analysis. Written informed consentwas obtained from parents of each participant before the procedure,and the study was approved by the Semmelweis UniversityRegional and Institutional Committee of Science and ResearchEthics (TUKEB: 73/2003).
RNA Isolation and Real-time RT-PCR
Total RNA was isolated from the samples by RNeasy RNAisolation kit (Qiagen GmbH, Hilden, Germany). One microgram ofRNAwas reverse transcribed using SuperScript II RNase H- (Gibco/BRL, Eggenstein, Germany) to generate first-stranded cDNA.HSP72 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH)mRNA expression were determined by real-time reverse transcrip-tase polymerase chain reaction (RT-PCR) using SYBR Green (forHSP72) or fluorescence resonance energy transfer hybridizationprobes (for GAPDH) on a Light Cycler system (Roche Diagnostics,Mannheim, Germany). The reaction mix for HSP72 contained4 mmol/L MgCl2, 500 nmol/L of each PCR primers (TibMolBiol,Berlin, Germany), 2 mL of FastStart DNA Master SYBR Greenenzyme mix (Roche Diagnostics), and 1 mL of cDNA sample. Theconditions were 1 cycle at 958C for 8 minutes, 50 cycles at 958C for5 seconds, 618C for 5 seconds, and 728C for 20 seconds with a singlefluorescence detection point at the end of the extension. The
CD¼ celiac disease.
pyright 2010 by ESPGHAN and NASPGHAN. Un
GAPDH-PCR contained 2 mmol/L MgCl2, 0.17 nmol/L of eachhybridization probe, 0.5 nmol/L of each primer, 2 mL of the
TABLE 2. Nucleotide sequences of specific primer pairs and prob
Gene Primer pairs and prob
HSP72 Forward: 50-TGC GAG AGG GCC AAG AReverse: 50-GTC GCG CCC GTT GAA G
GAPDH Forward: 50-CAC CAC CAT GGA GAA GReverse: 50-GTG ATG GCA TGG ACT GProbe 1: 50LCRed460-CCC TGG CCA AGProbe 2: 50-TCC TGC ACC ACC AAC TG
GAPDH¼ glyceraldehyde-3-phosphate dehydrogenase; HSP¼ heat shock pro
574
Light-Cycler FastStart DNA Master HybProbe (Roche Diagnos-tics), and 1 mL of cDNA. The conditions were 1 cycle at 958C for8 minutes, 50 cycles at 958C for 4 seconds, 558C for 8 seconds, and728C for 22 seconds with a single fluorescence detection point at theend of the annealing segment. Results were analyzed by Light-Cycler software version 3.5.3 (Roche Diagnostics). The mRNAexpression of HSP72 was determined by comparison with GAPDHas an internal control from the same sample (Table 2).
Protein Isolation and Western Blotting
Biopsy specimens were lysed in buffer containing leupeptin,aprotinin, Triton X-100, Tris-HCl, ethylene glycol-bis (2-aminoethylether),N,N,N0,N0-tetraacetic-acid, sodium fluoride, phe-nylmethylsulfonylfluoride, and Na-orthovanadate (each substancefrom Sigma-Aldrich Co, St Louis, MO) and centrifuged (10,000g,10 minutes, 48C) to pellet nuclei and large cellular fragments.Protein concentration of the supernatants was determined byBradford assay (Bio-Rad Laboratories, Hercules, CA). Twentymicrograms was separated by 12% sodium dodecyl sulfate poly-acrylamide gel electrophoresis at 120 V (�40 mA, 90 minutes)(Penguin Dual-Gel Water Cooled Systems, Owl, NH). Prestainedprotein mixture (BenchMark, Gibco/BRL, Eggenstein, Germany)was used as a marker of molecular mass. The separated proteinswere transferred to nitrocellulose membrane (Hybond ECL, APBiotech, Buckinghamshire, UK) at 70 V (�220 mA, 90 minutes)(MiniTank electroblotter, Owl, NH). Nonspecific binding sites wereblocked (2 hours, room temperature [RT]) in 5% nonfat dry milkcontaining blot solution. Membranes were incubated with rabbitmonoclonal antibodies to HSP72 (donated by Dr L. Laszlo, EotvosUniversity, Budapest, Hungary) (19) diluted to 1:10,000. Blots werewashed and incubated (30 minutes, RT) with peroxidase-conjugated
authorized reproduction of this article is prohibited.
goat anti-rabbit IgG secondary antibody (Sigma-Aldrich Co)diluted to 1:5000. Immunoreactive bands were visualized using
es applied for the real-time detection of HSP72 and GAPDH
es Product length, bp
GG AC-30 276AA GT-30
GC TG-30 240TG-30
G TCA TCC ATG A-PH 30
C TTA GC-FL 30
tein.
www.jpgn.org

Co
HSP72 immunoreactivity was observed. HSP72 was not present
FIGURE 1. HSP72 expression in the duodenal mucosa ofrepresentative samples from children with untreated CD,children with treated CD, and controls. Data for the mRNAexpression of HSP72 were obtained by computerized analysisof PCR products. Optical density of the investigated RT-PCRproducts was corrected for that of GAPDH. Data are shown asscatterplots. Analysis of significance was performed by Mann-Whitney U test. P¼0.003 untreated vs treated CD,P¼0.0002 untreated vs control, P¼0.023 treated vs control.CD¼ celiac disease; GAPDH¼glyceraldehyde-3-phosphatedehydrogenase; HSP¼heat shock protein; RT-PCR¼ reverse
HSP72 in Celiac Disease
enhanced chemiluminescence Western blotting detection protocol(AP Biotech, Buckinghamshire, UK). Negatives were analyzed bycomputerized densitometry (Gel-Pro Analyzer 3.1 software, MediaCybernetics, Bethesda, MD).
Immunofluorescent Staining
Duodenal biopsy samples were immediately snap frozen forimmunofluorescent analysis and embedded in Shandon cryomatrix(ThermoElectron Co, Waltham) and then cut to 5-mm slides. Slideswere incubated (1 hour, RT) with anti-HSP72 rabbit polyclonal IgGantibody (same antibody as in Western blot) diluted to 1:1000. Afterwashing, slides were incubated (30 minutes, RT) with Alexa Fluor488 F (ab0)2 fragment of goat anti-rabbit IgG (Invitrogen, Carlsbad,CA) diluted to 1:100. DNA was stained (10 minutes, RT) withHoechst 33342 (Sigma-Aldrich Co) diluted to 1:1000. Finally slideswere coverslipped with Vectashield fluorescent mounting medium(Vector Laboratories, Burlingame, CA). Appropriate controls wereperformed omitting the primary antibodies to ensure their specificityand avoid autofluorescence. To visualize the stained tissues on thesections, Zeiss LSM 510 Meta confocal laser scanning microscope(Carl Zeiss, Jena, Germany) was used equipped with an invertedAxiovert 200 M microscope, 20�Plan Apochromat (NA¼ 0.80), and63� Plan Apochromat oil immersion DIC objectives (NA¼ 1.4).
Statistical Analysis
Data were analyzed using Statistica 7.0 software (StatSoftInc, Tulsa, OK). After testing the normality with the Shapiro-Wilktest, the Mann-Whitney U test was used to determine the differencesamong all groups. Data are shown in a form of scatterplots. P values�0.05 were considered statistically significant.
RESULTS
HSP72 mRNA Expression in the DuodenalMucosa
Figure 1 shows the mRNA expression of HSP72 using real-time PCR as detected in the duodenal mucosa of samples fromchildren with untreated CD, children with treated CD, and controls.HSP72 mRNA expression was significantly increased in the duo-denal mucosa of children with untreated CD as well as children withtreated CD compared with that in controls (P¼ 0.0002 andP¼ 0.023, respectively). In the duodenal mucosa of children withtreated CD, the HSP72 mRNA level was decreased in comparison tochildren with untreated CD (P¼ 0.003).
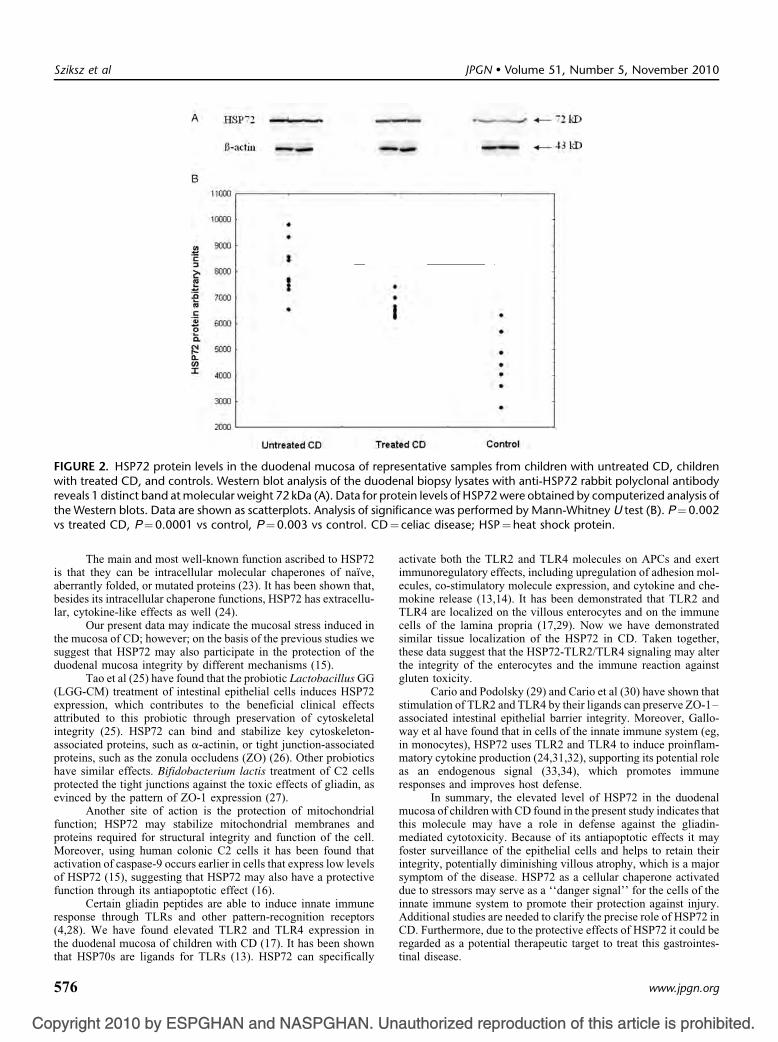
HSP72 Protein Levels in the Duodenal Mucosa
Western blot analysis of duodenal biopsy specimens fromchildren with untreated CD, children with treated CD, and controlsusing anti-HSP72 rabbit polyclonal antibody revealed distinctbands at 72 kDa (Fig. 2A). Significantly elevated HSP72 proteinlevels were detected in the duodenal mucosa of children withuntreated and also with treated CD compared with that in controls(P¼ 0.0001 and P¼ 0.003). In the duodenal mucosa of childrenwith treated CD, HSP72 protein levels were markedly higher than inuntreated CD (P¼ 0.002) (Fig. 2B).
Localization of HSP72 in the Duodenal Mucosa
JPGN � Volume 51, Number 5, November 2010
pyright 2010 by ESPGHAN and NASPGHAN. Un
The cellular distribution of HSP72 proteins was determinedby immunofluorescent staining. In the duodenal villi of children
www.jpgn.org
with untreated CD, strong HSP72 staining intensity was found in thevillous enterocytes and immune cells of the lamina propria com-pared with controls. In the duodenal villi of children with treatedCD, weaker HSP72 signal was found in villous enterocytes andimmune cells of the lamina propria compared with children withuntreated CD. In the normal duodenum of controls only weak
transcriptase polymerase chain reaction.
at detectable levels in the nuclear fractions (Fig. 3).
DISCUSSIONIn the present study we first demonstrated the increased
expression of HSP72 in the duodenal mucosa of children withCD. CD is a complex small intestinal disorder affecting 1% of thesociety and caused by intolerance against wheat gliadin and relatedproteins (20). Deregulated immune response and disturbance in theepithelial barrier function leads to the inflammation of the laminapropria (21) and increased enterocyte apoptosis (6).
In the intestinal mucosa the expression of HSP72 has beensparsely examined. There are only a few data about the importanceof HSP72 in inflammatory bowel diseases. Cuzzocrea et al (22)have revealed that the intraperitoneal administration of cyclopen-tenone prostaglandin (15-deoxy-D [12,14]-PGJ2)] stimulates theactivation of HSP72 in the inflamed colon of dinitrobenzenesulfonic acid–induced experimental rat colitis and causes substan-tial reduction of the degree of colonic injury (22).
In the present study we demonstrated significant upregula-tion of HSP72 in the villous enterocytes and immune cells of thelamina propria of children with untreated CD. In the duodenalmucosa of children maintained on gluten-free diet, the expression of
authorized reproduction of this article is prohibited.
HSP72 was markedly decreased compared with that of untreatedchildren; however; its level remained higher than that in controls.
575
Co
FIGURE 2. HSP72 protein levels in the duodenal mucosa of representative samples from children with untreated CD, childrenwith treated CD, and controls. Western blot analysis of the duodenal biopsy lysates with anti-HSP72 rabbit polyclonal antibodyreveals 1 distinct band at molecular weight 72 kDa (A). Data for protein levels of HSP72 were obtained by computerized analysis ofthe Western blots. Data are shown as scatterplots. Analysis of significance was performed by Mann-Whitney U test (B). P¼0.002
D¼
Sziksz et al JPGN � Volume 51, Number 5, November 2010
The main and most well-known function ascribed to HSP72is that they can be intracellular molecular chaperones of naıve,aberrantly folded, or mutated proteins (23). It has been shown that,besides its intracellular chaperone functions, HSP72 has extracellu-lar, cytokine-like effects as well (24).
Our present data may indicate the mucosal stress induced inthe mucosa of CD; however; on the basis of the previous studies wesuggest that HSP72 may also participate in the protection of theduodenal mucosa integrity by different mechanisms (15).
Tao et al (25) have found that the probiotic Lactobacillus GG(LGG-CM) treatment of intestinal epithelial cells induces HSP72expression, which contributes to the beneficial clinical effectsattributed to this probiotic through preservation of cytoskeletalintegrity (25). HSP72 can bind and stabilize key cytoskeleton-associated proteins, such as a-actinin, or tight junction-associatedproteins, such as the zonula occludens (ZO) (26). Other probioticshave similar effects. Bifidobacterium lactis treatment of C2 cellsprotected the tight junctions against the toxic effects of gliadin, asevinced by the pattern of ZO-1 expression (27).
Another site of action is the protection of mitochondrialfunction; HSP72 may stabilize mitochondrial membranes andproteins required for structural integrity and function of the cell.Moreover, using human colonic C2 cells it has been found thatactivation of caspase-9 occurs earlier in cells that express low levelsof HSP72 (15), suggesting that HSP72 may also have a protectivefunction through its antiapoptotic effect (16).
Certain gliadin peptides are able to induce innate immuneresponse through TLRs and other pattern-recognition receptors(4,28). We have found elevated TLR2 and TLR4 expression in
vs treated CD, P¼0.0001 vs control, P¼0.003 vs control. C
pyright 2010 by ESPGHAN and NASPGHAN. Un
the duodenal mucosa of children with CD (17). It has been shownthat HSP70s are ligands for TLRs (13). HSP72 can specifically
576
activate both the TLR2 and TLR4 molecules on APCs and exertimmunoregulatory effects, including upregulation of adhesion mol-ecules, co-stimulatory molecule expression, and cytokine and che-mokine release (13,14). It has been demonstrated that TLR2 andTLR4 are localized on the villous enterocytes and on the immunecells of the lamina propria (17,29). Now we have demonstratedsimilar tissue localization of the HSP72 in CD. Taken together,these data suggest that the HSP72-TLR2/TLR4 signaling may alterthe integrity of the enterocytes and the immune reaction againstgluten toxicity.
Cario and Podolsky (29) and Cario et al (30) have shown thatstimulation of TLR2 and TLR4 by their ligands can preserve ZO-1–associated intestinal epithelial barrier integrity. Moreover, Gallo-way et al have found that in cells of the innate immune system (eg,in monocytes), HSP72 uses TLR2 and TLR4 to induce proinflam-matory cytokine production (24,31,32), supporting its potential roleas an endogenous signal (33,34), which promotes immuneresponses and improves host defense.
In summary, the elevated level of HSP72 in the duodenalmucosa of children with CD found in the present study indicates thatthis molecule may have a role in defense against the gliadin-mediated cytotoxicity. Because of its antiapoptotic effects it mayfoster surveillance of the epithelial cells and helps to retain theirintegrity, potentially diminishing villous atrophy, which is a majorsymptom of the disease. HSP72 as a cellular chaperone activateddue to stressors may serve as a ‘‘danger signal’’ for the cells of theinnate immune system to promote their protection against injury.Additional studies are needed to clarify the precise role of HSP72 inCD. Furthermore, due to the protective effects of HSP72 it could be
celiac disease; HSP¼heat shock protein.
authorized reproduction of this article is prohibited.
regarded as a potential therapeutic target to treat this gastrointes-tinal disease.
www.jpgn.org

Co
FIGURE 3. Localization of HSP72 in the duodenal mucosa of representative samples from children with untreated CD, childrenwith treated CD, and controls. Frozen duodenal villous sections were fixed and immunofluorescent staining was performed usinganti-HSP72 antibody. Arrowheads indicate HSP72 staining of enterocytes (E) and lamina propria immune cells (LPC). Heavystaining of HSP72 was found in villous enterocytes and inflammatory cells of the lamina propria of untreated patients with CD.Immunofluorescent staining revealed decreased HSP72 positivity in the lamina propria immune cells of treated patients with CDcompared with untreated patients with CD, whereas no fluorescent signal was detected in villous enterocytes. Weak HSP72immunopositivity was observed in the immune cells of the lamina propria of controls. DIC pictures show longitudinal (untreatedCD and controls) or horizontal sections of duodenal villi (treated CD). Confocal images were taken on a Zeiss Axiovert LSM510
ase
JPGN � Volume 51, Number 5, November 2010 HSP72 in Celiac Disease
Acknowledgments: We are grateful to Maria Bernath forexcellent technical assistance.
REFERENCES1. Polanco I. Celiac disease. J Pediatr Gastroenterol Nutr 2008;47 (Suppl 1):
S3–6.2. Anderson JM, Van Itallie CM. Tight junctions and the molecular basis
for regulation of paracellular permeability. Am J Physiol GastrointestLiver Physiol 1995;269:G467–75.
3. Stepniak D, Koning F. Celiac disease–sandwiched between innate andadaptive immunity. Hum Immunol 2006;67:460–8.
4. Maiuri L, Ciacci C, Ricciardelli I, et al. Association between innateresponse to gliadin and activation of pathogenic T cells in coeliacdisease. Lancet 2003;362:30–7.
5. Koning F. Celiac disease: caught between a rock and a hard place.Gastroenterology 2005;129:1294–301.
6. Giovannini C, Matarrese P, Scazzocchio B, et al. Wheat gliadin inducesapoptosis of intestinal cells via an autocrine mechanism involving Fas-Fas ligand pathway. FEBS Lett 2003;540:117–24.
with the plan apochromat 63x/1.40 Oil DIC. CD¼ celiac dise
pyright 2010 by ESPGHAN and NASPGHAN. Un
7. Whitham M, Fortes MB. Heat shock protein 72: release and biologicalsignificance during exercise. Front Biosci 2008;13:1328–39.
www.jpgn.org
8. Odashima M, Otaka M, Jin M, et al. Induction of a 72-kDa heat-shockprotein in cultured rat gastric mucosal cells and rat gastric mucosa byzinc L-carnosine. Dig Dis Sci 2002;47:2799–804.
9. Asea A, Jean-Pierre C, Kaur P, et al. Heat shock protein-containing exo-somes in mid-trimester amniotic fluids. J Reprod Immunol 2008;79:12–7.
10. Asea A. Stress proteins and initiation of immune response: chaperokineactivity of hsp72. Exerc Immunol Rev 2005;11:34–45.
11. Lehner T, Bergmeier LA, Wang Y, et al. Heat shock proteins generatebeta-chemokines which function as innate adjuvants enhancing adaptiveimmunity. Eur J Immunol 2000;30:594–603.
12. Panjwani NN, Popova L, Srivastava PK. Heat shock proteins gp96 andhsp70 activate the release of nitric oxide by APCs. J Immunol 2002;168:2997–3003.
13. Asea A. Heat shock proteins and toll-like receptors. Handb ExpPharmacol 2008;183:111–27.
14. Wheeler DS, Chase MA, Senft AP, et al. Extracellular Hsp72, an endo-genous DAMP, is released by virally infected airway epithelial cells andactivates neutrophils via Toll-like receptor (TLR)-4. Respir Res 2009;10:31.
15. Liu TS, Musch MW, Sugi K, et al. Protective role of HSP72 against
; HSP¼heat shock protein.
authorized reproduction of this article is prohibited.
Clostridium difficile toxin A-induced intestinal epithelial cell dysfunc-tion. Am J Physiol Cell Physiol 2003;284:C1073–82.
577

Co
16. Lanneau D, de Thonel A, Maurel S, et al. Apoptosis versus celldifferentiation: role of heat shock proteins HSP90, HSP70 andHSP27. Prion 2007;1:53–60.
17. Szebeni B, Veres G, Dezsofi A, et al. Increased mucosal expression ofToll-like receptor (TLR)2 and TLR4 in coeliac disease. J PediatrGastroenterol Nutr 2007;45:187–93.
18. Walker-Smith JA, Guandalini S, Schmitz J, et al. Revised criteria fordiagnosis of coeliac disease. Report of Working Group of EuropeanSociety of Paediatric Gastroenterology and Nutrition. Arch Dis Child1990;65:909–11.
19. Kurucz I, Tombor B, Prechl J, et al. Ultrastructural localization of Hsp-72 examined with a new polyclonal antibody raised against the truncatedvariable domain of the heat shock protein. Cell Stress Chaperones1999;4:139–52.
20. Leeds JS, Hopper AD, Sanders DS. Coeliac disease. Br Med Bull2008;88:157–70.
21. Gianfrani C, Auricchio S, Troncone R. Adaptive and innate immuneresponses in celiac disease. Immunol Lett 2005;99:141–5.
22. Cuzzocrea S, Ianaro A, Wayman NS, et al. The cyclopentenoneprostaglandin 15-deoxy-delta(12,14)- PGJ2 attenuates the developmentof colon injury caused by dinitrobenzene sulphonic acid in the rat. Br JPharmacol 2003;138:678–88.
23. Lindquist S, Craig EA. The heat-shock proteins. Annu Rev Genet1988;22:631–77.
24. Asea A, Kraeft SK, Kurt-Jones EA, et al. HSP70 stimulates cytokineproduction through a CD14-dependant pathway, demonstrating its dualrole as a chaperone and cytokine. Nat Med 2000;6:435–42.
25. Tao Y, Drabik KA, Waypa TS, et al. Soluble factors from Lactobacillus
Sziksz et al
pyright 2010 by ESPGHAN and NASPGHAN. Un
intestinal epithelial cells. Am J Physiol Cell Physiol 2006;290:C1018–30.
578
26. Musch MW, Sugi K, Straus D, et al. Heat-shock protein 72protects against oxidant-induced injury of barrier function of humancolonic epithelial Caco2/bbe cells. Gastroenterology 1999;117:115–22.
27. Lindfors K, Blomqvist T, Juuti-Uusitalo K, et al. Live probioticBifidobacterium lactis bacteria inhibit the toxic effects induced bywheat gliadin in epithelial cell culture. Clin Exp Immunol 2008;152:552–8.
28. Schuppan D, Esslinger B, Dieterich W. Innate immunity and coeliacdisease. Lancet 2003;362:3–4.
29. Cario E, Podolsky DK. Differential alteration in intestinal epithelial cellexpression of toll-like receptor 3 (TLR3) and TLR4 in inflammatorybowel disease. Infect Immun 2000;68:7010–7.
30. Cario E, Gerken G, Podolsky DK. Toll-like receptor 2 controls mucosalinflammation by regulating epithelial barrier function. Gastroenterology2007;132:1359–74.
31. Asea A, Rehli M, Kabingu E, et al. Novel signal transduction pathwayutilized by extracellular HSP70: role of Toll-like receptor (TLR) 2 andTLR4. J Biol Chem 2002;277:15028–34.
32. Galloway E, Shin T, Huber N, et al. Activation of hepatocytes byextracellular heat shock protein 72. Am J Physiol Cell Physiol 2008;295:C514–20.
33. Chase MA, Wheeler DS, Lierl KM, et al. Hsp72 induces inflammationand regulates cytokine production in airway epithelium through aTLR4- and NF-kappaB-dependent mechanism. J Immunol 2007;179:6318–24.
34. Karoly E, Fekete A, Banki NF, et al. Heat shock protein 72 (HSPA1B)gene polymorphism and Toll-like receptor (TLR) 4 mutation are
JPGN � Volume 51, Number 5, November 2010
associated with increased risk of urinary tract infection in children.
GG activate MAPKs and induce cytoprotective heat shock proteins inauthorized reproduction of this article is prohibited.
Pediatr Res 2007;61:371–4.
www.jpgn.org
Related Documents











