In vivo infection of IgG-containing cells by Jembrana disease virus during acute infection Moira Desport a, ⁎, I.W. Masa Tenaya a,b , Alexander McLachlan a,1 , Tegan J. McNab a , Judhi Rachmat a,2 , Nining Hartaningsih b , Graham E. Wilcox a a School of Veterinary and Biomedical Science, Murdoch University, Murdoch WA 6150, Australia b Disease Investigation Centre, PO Box 3322, Denpasar, Bali, Indonesia abstract article info Article history: Received 17 March 2009 Returned to author for revision 14 July 2009 Accepted 23 July 2009 Available online 5 September 2009 Keywords: Bovine Lentivirus Plasma cells Tropism Jembrana disease virus Lymphocytes IgG-containing cells Jembrana disease virus (JDV) is an unusual bovine lentivirus which causes a non-follicular proliferation of lymphocytes, a transient immunosuppression and a delayed humoral response in infected Bali cattle in Indonesia. A double-immunofluorescent labeling method was developed to identify the subset of mononuclear cells in which the viral capsid protein could be detected. Viral antigen was present in pleomorphic centroblast-like cells which were identified as IgG-containing cells, including plasma cells, in lymphoid tissues. There was no evidence of infection of CD3 + T-cells or MAC387 + monocytes in tissues but large vacuolated cells with a macrophage-like morphology in the lung were found to contain viral antigen although they could not be shown conclusively to be infected. The tropism of JDV for mature IgG-containing cells may be relevant to understanding the pathogenesis of Jembrana disease, the delayed antibody responses and the genetic composition of this atypical lentivirus. © 2009 Elsevier Inc. All rights reserved. Introduction Jembrana disease virus (JDV) causes an acute and severe infection of the immune system of Bali cattle in Indonesia. Experimental infection of Bali cattle with JDV typically results in elevated rectal temperatures beginning 5–10 days post-infection which resolve within 14–17 days. This clinical response is accompanied by a leucopenia and high viral titers which peak on average 11.5 days post-infection at approximately 10 10 RNA genome copies/ml plasma (Desport et al., 2009). Most of the infectious agent is found in the plasma fraction of whole blood during this stage of the disease and is reduced to 10 5 ID 50 /ml as the temperature resolves and persists at levels as low as 10 ID 50 /ml for at least 25 months after recovery. Cattle that have been infected with JDV were found to be resistant to re- infection with a heterologous virus strain and there is no evidence of recurrence of clinical disease in recovered animals (Soeharsono et al., 1990; Soeharsono et al., 1995). Experimental infection with JDV initially causes a non-follicular lymphoproliferative response in lymphoid organs with a loss of IgG- containing cells and a decrease in CD4:CD8 lymphocyte numbers during the first 2 weeks of infection (Dharma et al., 1991; Dharma et al., 1994). The distribution of infected cells in post-mortem tissues taken at this stage of the disease was found to be predominantly in the parafollicular areas of the spleen and lymph node with little or no signal in the follicles (Chadwick et al., 1998). Follicular architecture was found to be obliterated by proliferating cells by 8 days after infection and marked follicular lymphoid reactions and plasma cell formation were only observed from the fifth week after infection (Dharma et al., 1991). The phenotype of the proliferating lymphocytes during the acute stage of the disease was originally suggested to be T- cells. We have recently identified B-lineage cells (CD79α positive) in the cortico-medullary region which were found to coalesce to efface the paracortex of lymph nodes and tonsils, and replace the marginal zone of splenic follicles during the early stages of the acute infection (second day of fever). A CD3 + T-cell proliferative response was identified in lymphoid tissues taken at a later stage of infection after the resolution of the febrile response (unpublished data). The cell tropism of JDV is unknown but has been suggested to be lymphocytes and/or monocyte/macrophage lineage cells (Chadwick et al., 1998). The genetically related bovine immunodeficiency virus (BIV) exhibits a broad cell tropism in vivo which includes T-cells, B- cells and monocyte/macrophage cells although it is not clear whether all of these cell types are productively infected since only BIV proviral DNA was detected using PCR in these studies (Heaton et al., 1998; Whetstone et al., 1997). Other experimental infection studies in cattle Virology 393 (2009) 221–227 ⁎ Corresponding author. Fax: +61 8 93104144. E-mail address: [email protected] (M. Desport). 1 Current address: New Zealand Veterinary Pathology, Cnr Anglesea and Thackeray Streets, PO Box 944, Hamilton, New Zealand. 2 Current address: R&D Centre for Biotechnology, LIPI, Jln Raya Bogor, Cibinong 16911, Indonesia. 0042-6822/$ – see front matter © 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.virol.2009.07.027 Contents lists available at ScienceDirect Virology journal homepage: www.elsevier.com/locate/yviro

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Virology 393 (2009) 221–227
Contents lists available at ScienceDirect
Virology
j ourna l homepage: www.e lsev ie r.com/ locate /yv i ro
In vivo infection of IgG-containing cells by Jembrana disease virus duringacute infection
Moira Desport a,⁎, I.W. Masa Tenaya a,b, Alexander McLachlan a,1, Tegan J. McNab a, Judhi Rachmat a,2,Nining Hartaningsih b, Graham E. Wilcox a
a School of Veterinary and Biomedical Science, Murdoch University, Murdoch WA 6150, Australiab Disease Investigation Centre, PO Box 3322, Denpasar, Bali, Indonesia
⁎ Corresponding author. Fax: +61 8 93104144.E-mail address: [email protected] (M. Des
1 Current address: New Zealand Veterinary PathologyStreets, PO Box 944, Hamilton, New Zealand.
2 Current address: R&D Centre for Biotechnology, L16911, Indonesia.
0042-6822/$ – see front matter © 2009 Elsevier Inc. Adoi:10.1016/j.virol.2009.07.027
a b s t r a c t
a r t i c l e i n f oArticle history:Received 17 March 2009Returned to author for revision 14 July 2009Accepted 23 July 2009Available online 5 September 2009
Keywords:BovineLentivirusPlasma cellsTropismJembrana disease virusLymphocytesIgG-containing cells
Jembrana disease virus (JDV) is an unusual bovine lentivirus which causes a non-follicular proliferation oflymphocytes, a transient immunosuppression and a delayed humoral response in infected Bali cattle inIndonesia. A double-immunofluorescent labeling method was developed to identify the subset ofmononuclear cells in which the viral capsid protein could be detected. Viral antigen was present inpleomorphic centroblast-like cells which were identified as IgG-containing cells, including plasma cells, inlymphoid tissues. There was no evidence of infection of CD3+ T-cells or MAC387+ monocytes in tissues butlarge vacuolated cells with a macrophage-like morphology in the lung were found to contain viral antigenalthough they could not be shown conclusively to be infected. The tropism of JDV for mature IgG-containingcells may be relevant to understanding the pathogenesis of Jembrana disease, the delayed antibodyresponses and the genetic composition of this atypical lentivirus.
© 2009 Elsevier Inc. All rights reserved.
Introduction
Jembrana disease virus (JDV) causes an acute and severe infectionof the immune system of Bali cattle in Indonesia. Experimentalinfection of Bali cattle with JDV typically results in elevated rectaltemperatures beginning 5–10 days post-infection which resolvewithin 14–17 days. This clinical response is accompanied by aleucopenia and high viral titers which peak on average 11.5 dayspost-infection at approximately 1010 RNA genome copies/ml plasma(Desport et al., 2009). Most of the infectious agent is found in theplasma fraction of whole blood during this stage of the disease and isreduced to 105 ID50/ml as the temperature resolves and persists atlevels as low as 10 ID50/ml for at least 25months after recovery. Cattlethat have been infected with JDV were found to be resistant to re-infection with a heterologous virus strain and there is no evidence ofrecurrence of clinical disease in recovered animals (Soeharsono et al.,1990; Soeharsono et al., 1995).
Experimental infection with JDV initially causes a non-follicularlymphoproliferative response in lymphoid organs with a loss of IgG-
port)., Cnr Anglesea and Thackeray
IPI, Jln Raya Bogor, Cibinong
ll rights reserved.
containing cells and a decrease in CD4:CD8 lymphocyte numbersduring the first 2 weeks of infection (Dharma et al., 1991; Dharmaet al., 1994). The distribution of infected cells in post-mortem tissuestaken at this stage of the diseasewas found to be predominantly in theparafollicular areas of the spleen and lymph node with little or nosignal in the follicles (Chadwick et al., 1998). Follicular architecturewas found to be obliterated by proliferating cells by 8 days afterinfection and marked follicular lymphoid reactions and plasma cellformation were only observed from the fifth week after infection(Dharma et al., 1991). The phenotype of the proliferating lymphocytesduring the acute stage of the disease was originally suggested to be T-cells. We have recently identified B-lineage cells (CD79α positive) inthe cortico-medullary region which were found to coalesce to effacethe paracortex of lymph nodes and tonsils, and replace the marginalzone of splenic follicles during the early stages of the acute infection(second day of fever). A CD3+ T-cell proliferative response wasidentified in lymphoid tissues taken at a later stage of infection afterthe resolution of the febrile response (unpublished data).
The cell tropism of JDV is unknown but has been suggested to belymphocytes and/or monocyte/macrophage lineage cells (Chadwicket al., 1998). The genetically related bovine immunodeficiency virus(BIV) exhibits a broad cell tropism in vivo which includes T-cells, B-cells and monocyte/macrophage cells although it is not clear whetherall of these cell types are productively infected since only BIV proviralDNA was detected using PCR in these studies (Heaton et al., 1998;Whetstone et al., 1997). Other experimental infection studies in cattle

Fig. 1. Mapping the reactivity of mAb LD1 raised against recombinant JDV capsidprotein. Schematic representation of JDVTAB/87 genome location (Chadwick et al., 1995)of biotinylated JDV gag constructs expressed using Pinpoint X-A-1 system in E. coli (A).Western immunoblotting with mAb LD1 revealed that its reactivity maps to the aminoterminus of capsid in the region encompassed by Jgag11 (expressed by nucleotides604–810). Lane 1, Marker; lane2, Jgag1; lane 3, Jgag2; lane 4, Jgag3; lane 5, Jgag4; lane 6,Jgag5; lane 7, Jgag6; lane 8, Jgag7; lane 9, Jgag11; lane 10, Jgag13 (B).
222 M. Desport et al. / Virology 393 (2009) 221–227
have reported that BIV could only be isolated frommonocytes but notfrom T-cells (Onuma et al., 1992) and when inoculated into rabbits,BIV antigen was detected in atypical blastic mononuclear cells whichwere presumed to be of the macrophage lineage in the red pulp of thespleen (Pifat et al., 1992). We have recently reported that JDV strainsin Bali are genetically stable with little variation even in env sequencesin isolates from over a period of 20 years (Desport et al., 2007). Thiscould indicate that JDV has a narrow host cell range, potentiallytargeting long-lived cells with a slow turnover rate. Attempts toculture JDV in cell lines that support BIV infection have beenunsuccessful and only in vitro cultivation of bovine mononuclearcells derived from peripheral blood could be shown to support JDVreplication when inoculated with plasma from infected cattle (Wilcoxet al., 1992).
Perturbations of the immune system are a consequence of manylentiviral infections and a hallmark of Jembrana disease is thetransient immunosuppression and delay in the development ofvirus specific antibodies of between 5–15 weeks post-infection(Desport et al., 2009; Hartaningsih et al., 1994; Wareing et al.,1999). A lack of virus specific humoral immune responses has beenreported in rapid progressors in both HIV and SIV infections(Dykhuizen et al., 1998; Michael et al., 1997) and has been attributedto a progressive depletion of proliferating B-cells as early as 20 dayspost-infection (Zhang et al., 2007). To better understand the delayedimmunological responses and the genetic stability of JDV, we soughtto determine the cell tropism of the virus using double-immunoflu-orescent staining to detect JDV capsid in subsets of leukocytesidentified using specific antibodies in formalin-fixed tissues.
Results
Characterization of JDV capsid-specific mAb
In order to undertake further investigations into the cell tropism ofJDV during acute infection a virus specific monoclonal antibody of adifferent isotype (IgG2b) to the cell surface markers that were used inthis study (IgG1) was selected. MAb LD1 was produced using arecombinant capsid protein as the immunizing antigen in mice andwas screened against a range of JDV gag truncated proteins todetermine the specificity of the antibody binding. The JDV gagsequence encoded by each of the DNA constructs used in this study isshown in Fig. 1 together with the Western immunoblot showing thatthe mAb was reactive against Jgag2, Jgag3, Jgag5, Jgag6 and Jgag11.This indicated that mAb LD1 bound to an epitope located between JDVnucleotides 604–810 (Chadwick et al., 1995) at the N′-terminus of thecapsid protein.
Analysis of infected cell phenotype
Productive infection of bovine lymphocytes during acute infectionwith JDV was determined using immunofluorescent staining techni-ques on formalin-fixed paraffin-embedded tissues. JDV-infected cellswere identified using mAb LD1 and a secondary goat anti-mouseantibody labeled with Alexa Fluor 568, without any further amplifi-cation of the fluorescent signal (Figs. 2 and 3). CD3-positive T-cellswere simultaneously labeled andwere found to be highly abundant inthe lymphoid tissues examined (Figs. 2A and D). There was no co-localization of signal when the fluorescent images for JDV capsid andCD3 staining were merged (Figs. 2C and F).
B-lymphocytes were identified initially using mAb anti-humanCD79α and were predominantly located in the germinal centers ofthe lymphoid follicles as expected. No co-localization of stainingwith LD1 was observed with CD79α positive cells in this locationbut occasionally CD79α positive cells outside the germinal centerswere also positive with mAb LD1 indicating that a small proportionof B-lymphocytes are productively infected with JDV (Fig. 3C
arrow). Plasma cells were identified in lymph node sections bydetecting the presence of bovine IgG in the cytoplasm (Figs. 3D andG) and many plasma cells were also found to contain JDV capsidprotein (Figs. 3F and I). Some background signal was observed withthe staining for IgG in lymph node sections (Figs. 3G and I),presumably as it is present both intracellularly and extracellularly inthese locations.
Distribution of cells of monocyte/macrophage lineage
Recently blood-derived monocytes and neutrophils were identi-fied initially in tissues using mAbMAC387 (Figs. 2G and I). Antibodiesdirected against different regions of human CD68 have been used tosuccessfully identify macrophages including mAb EBM11 which hasalso been applied to bovine tissues (Bielefeldt-Ohmann et al., 1988;Greywoode et al., 1990). Unfortunately the antigen retrieval requiredfor LD1 was not compatible with that required for EBM11 and it wasnot possible to perform double immunostaining with this combina-tion of antibodies. However, when single IHC was performed on serialsections of lung, the distribution of EBM11 positive cells (Fig. 4A)compared to MAC387 positive cells (Fig. 4C) was markedly differentin the lung confirming that these antibodies identified differentsubsets of cells of the mononuclear phagocyte system. There wereEBM11 positive cells in the alveoli, whereas JDV capsid-immunopo-sitive cells were scattered throughout the alveolar septae (Figs. 4B andD). It is possible that at least some of these alveolar macrophages wereinfected (Figss. 4A and B, D and E). Similarly, the distribution ofmature tissue macrophages identified using EBM11 in the medullarysinuses of the lymph node (Fig. 4F) was different to that of the JDV-positive cells in the medullary cords (Fig. 4G). Despite similarities inthe morphology and distribution of MAC387 positive cells and JDV-infected cells, there was no co-localization of staining of monocytes orneutrophils and JDV-infected cells in the lymph node (Fig. 2I) or lung(data not shown).
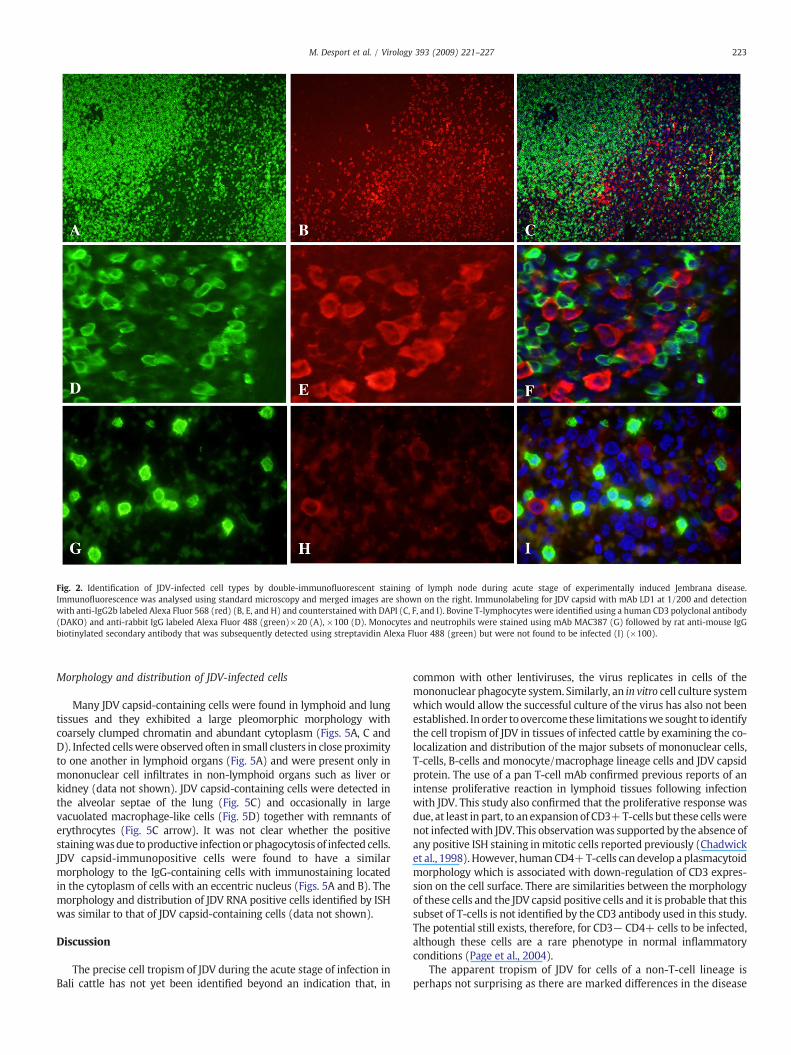
Fig. 2. Identification of JDV-infected cell types by double-immunofluorescent staining of lymph node during acute stage of experimentally induced Jembrana disease.Immunofluorescence was analysed using standard microscopy and merged images are shown on the right. Immunolabeling for JDV capsid with mAb LD1 at 1/200 and detectionwith anti-IgG2b labeled Alexa Fluor 568 (red) (B, E, and H) and counterstained with DAPI (C, F, and I). Bovine T-lymphocytes were identified using a human CD3 polyclonal antibody(DAKO) and anti-rabbit IgG labeled Alexa Fluor 488 (green)×20 (A), ×100 (D). Monocytes and neutrophils were stained using mAb MAC387 (G) followed by rat anti-mouse IgGbiotinylated secondary antibody that was subsequently detected using streptavidin Alexa Fluor 488 (green) but were not found to be infected (I) (×100).
223M. Desport et al. / Virology 393 (2009) 221–227
Morphology and distribution of JDV-infected cells
Many JDV capsid-containing cells were found in lymphoid and lungtissues and they exhibited a large pleomorphic morphology withcoarsely clumped chromatin and abundant cytoplasm (Figs. 5A, C andD). Infected cellswere observed often in small clusters in close proximityto one another in lymphoid organs (Fig. 5A) and were present only inmononuclear cell infiltrates in non-lymphoid organs such as liver orkidney (data not shown). JDV capsid-containing cells were detected inthe alveolar septae of the lung (Fig. 5C) and occasionally in largevacuolated macrophage-like cells (Fig. 5D) together with remnants oferythrocytes (Fig. 5C arrow). It was not clear whether the positivestainingwasdue toproductive infection or phagocytosis of infected cells.JDV capsid-immunopositive cells were found to have a similarmorphology to the IgG-containing cells with immunostaining locatedin the cytoplasm of cells with an eccentric nucleus (Figs. 5A and B). Themorphology and distribution of JDV RNA positive cells identified by ISHwas similar to that of JDV capsid-containing cells (data not shown).
Discussion
The precise cell tropism of JDV during the acute stage of infection inBali cattle has not yet been identified beyond an indication that, in
common with other lentiviruses, the virus replicates in cells of themononuclear phagocyte system. Similarly, an in vitro cell culture systemwhich would allow the successful culture of the virus has also not beenestablished. Inorder toovercome these limitationswe sought to identifythe cell tropism of JDV in tissues of infected cattle by examining the co-localization and distribution of the major subsets of mononuclear cells,T-cells, B-cells and monocyte/macrophage lineage cells and JDV capsidprotein. The use of a pan T-cell mAb confirmed previous reports of anintense proliferative reaction in lymphoid tissues following infectionwith JDV. This study also confirmed that the proliferative response wasdue, at least in part, to anexpansionof CD3+T-cells but these cellswerenot infectedwith JDV. This observationwas supported by the absence ofany positive ISH staining inmitotic cells reported previously (Chadwicket al., 1998). However, humanCD4+T-cells can develop a plasmacytoidmorphology which is associated with down-regulation of CD3 expres-sion on the cell surface. There are similarities between the morphologyof these cells and the JDV capsid positive cells and it is probable that thissubset of T-cells is not identified by the CD3 antibody used in this study.The potential still exists, therefore, for CD3− CD4+ cells to be infected,although these cells are a rare phenotype in normal inflammatoryconditions (Page et al., 2004).
The apparent tropism of JDV for cells of a non-T-cell lineage isperhaps not surprising as there are marked differences in the disease
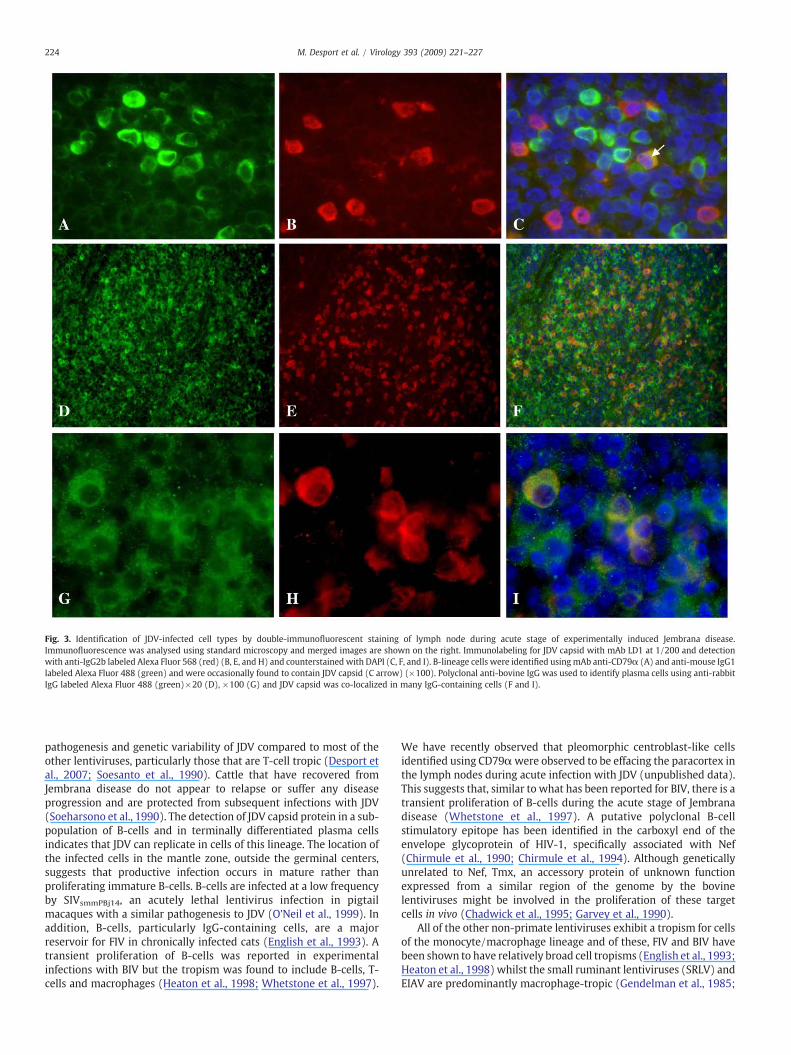
Fig. 3. Identification of JDV-infected cell types by double-immunofluorescent staining of lymph node during acute stage of experimentally induced Jembrana disease.Immunofluorescence was analysed using standard microscopy and merged images are shown on the right. Immunolabeling for JDV capsid with mAb LD1 at 1/200 and detectionwith anti-IgG2b labeled Alexa Fluor 568 (red) (B, E, and H) and counterstained with DAPI (C, F, and I). B-lineage cells were identified usingmAb anti-CD79α (A) and anti-mouse IgG1labeled Alexa Fluor 488 (green) and were occasionally found to contain JDV capsid (C arrow) (×100). Polyclonal anti-bovine IgG was used to identify plasma cells using anti-rabbitIgG labeled Alexa Fluor 488 (green)×20 (D), ×100 (G) and JDV capsid was co-localized in many IgG-containing cells (F and I).
224 M. Desport et al. / Virology 393 (2009) 221–227
pathogenesis and genetic variability of JDV compared to most of theother lentiviruses, particularly those that are T-cell tropic (Desport etal., 2007; Soesanto et al., 1990). Cattle that have recovered fromJembrana disease do not appear to relapse or suffer any diseaseprogression and are protected from subsequent infections with JDV(Soeharsono et al., 1990). The detection of JDV capsid protein in a sub-population of B-cells and in terminally differentiated plasma cellsindicates that JDV can replicate in cells of this lineage. The location ofthe infected cells in the mantle zone, outside the germinal centers,suggests that productive infection occurs in mature rather thanproliferating immature B-cells. B-cells are infected at a low frequencyby SIVsmmPBj14, an acutely lethal lentivirus infection in pigtailmacaques with a similar pathogenesis to JDV (O'Neil et al., 1999). Inaddition, B-cells, particularly IgG-containing cells, are a majorreservoir for FIV in chronically infected cats (English et al., 1993). Atransient proliferation of B-cells was reported in experimentalinfections with BIV but the tropism was found to include B-cells, T-cells and macrophages (Heaton et al., 1998; Whetstone et al., 1997).
We have recently observed that pleomorphic centroblast-like cellsidentified using CD79αwere observed to be effacing the paracortex inthe lymph nodes during acute infection with JDV (unpublished data).This suggests that, similar to what has been reported for BIV, there is atransient proliferation of B-cells during the acute stage of Jembranadisease (Whetstone et al., 1997). A putative polyclonal B-cellstimulatory epitope has been identified in the carboxyl end of theenvelope glycoprotein of HIV-1, specifically associated with Nef(Chirmule et al., 1990; Chirmule et al., 1994). Although geneticallyunrelated to Nef, Tmx, an accessory protein of unknown functionexpressed from a similar region of the genome by the bovinelentiviruses might be involved in the proliferation of these targetcells in vivo (Chadwick et al., 1995; Garvey et al., 1990).
All of the other non-primate lentiviruses exhibit a tropism for cellsof the monocyte/macrophage lineage and of these, FIV and BIV havebeen shown to have relatively broad cell tropisms (English et al., 1993;Heaton et al., 1998) whilst the small ruminant lentiviruses (SRLV) andEIAV are predominantly macrophage-tropic (Gendelman et al., 1985;
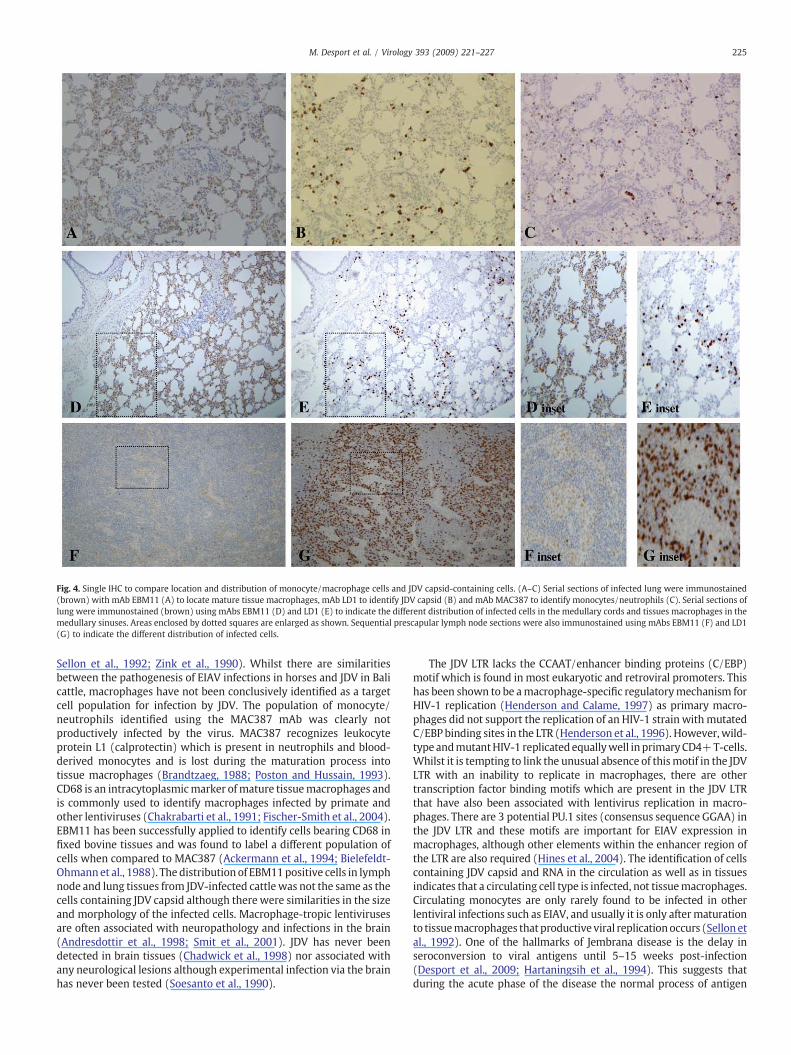
Fig. 4. Single IHC to compare location and distribution of monocyte/macrophage cells and JDV capsid-containing cells. (A–C) Serial sections of infected lung were immunostained(brown) with mAb EBM11 (A) to locate mature tissue macrophages, mAb LD1 to identify JDV capsid (B) and mAb MAC387 to identify monocytes/neutrophils (C). Serial sections oflung were immunostained (brown) using mAbs EBM11 (D) and LD1 (E) to indicate the different distribution of infected cells in the medullary cords and tissues macrophages in themedullary sinuses. Areas enclosed by dotted squares are enlarged as shown. Sequential prescapular lymph node sections were also immunostained using mAbs EBM11 (F) and LD1(G) to indicate the different distribution of infected cells.
225M. Desport et al. / Virology 393 (2009) 221–227
Sellon et al., 1992; Zink et al., 1990). Whilst there are similaritiesbetween the pathogenesis of EIAV infections in horses and JDV in Balicattle, macrophages have not been conclusively identified as a targetcell population for infection by JDV. The population of monocyte/neutrophils identified using the MAC387 mAb was clearly notproductively infected by the virus. MAC387 recognizes leukocyteprotein L1 (calprotectin) which is present in neutrophils and blood-derived monocytes and is lost during the maturation process intotissue macrophages (Brandtzaeg, 1988; Poston and Hussain, 1993).CD68 is an intracytoplasmicmarker ofmature tissuemacrophages andis commonly used to identify macrophages infected by primate andother lentiviruses (Chakrabarti et al., 1991; Fischer-Smith et al., 2004).EBM11 has been successfully applied to identify cells bearing CD68 infixed bovine tissues and was found to label a different population ofcells when compared to MAC387 (Ackermann et al., 1994; Bielefeldt-Ohmannet al., 1988). The distribution of EBM11positive cells in lymphnode and lung tissues from JDV-infected cattle was not the same as thecells containing JDV capsid although there were similarities in the sizeand morphology of the infected cells. Macrophage-tropic lentivirusesare often associated with neuropathology and infections in the brain(Andresdottir et al., 1998; Smit et al., 2001). JDV has never beendetected in brain tissues (Chadwick et al., 1998) nor associated withany neurological lesions although experimental infection via the brainhas never been tested (Soesanto et al., 1990).
The JDV LTR lacks the CCAAT/enhancer binding proteins (C/EBP)motif which is found in most eukaryotic and retroviral promoters. Thishas been shown to be amacrophage-specific regulatorymechanism forHIV-1 replication (Henderson and Calame, 1997) as primary macro-phages did not support the replication of an HIV-1 strain withmutatedC/EBP binding sites in the LTR (Henderson et al., 1996). However, wild-type andmutantHIV-1 replicated equallywell inprimaryCD4+T-cells.Whilst it is tempting to link the unusual absence of thismotif in the JDVLTR with an inability to replicate in macrophages, there are othertranscription factor binding motifs which are present in the JDV LTRthat have also been associated with lentivirus replication in macro-phages. There are 3 potential PU.1 sites (consensus sequence GGAA) inthe JDV LTR and these motifs are important for EIAV expression inmacrophages, although other elements within the enhancer region ofthe LTR are also required (Hines et al., 2004). The identification of cellscontaining JDV capsid and RNA in the circulation as well as in tissuesindicates that a circulating cell type is infected, not tissuemacrophages.Circulating monocytes are only rarely found to be infected in otherlentiviral infections such as EIAV, and usually it is only aftermaturationto tissuemacrophages that productive viral replicationoccurs (Sellonetal., 1992). One of the hallmarks of Jembrana disease is the delay inseroconversion to viral antigens until 5–15 weeks post-infection(Desport et al., 2009; Hartaningsih et al., 1994). This suggests thatduring the acute phase of the disease the normal process of antigen
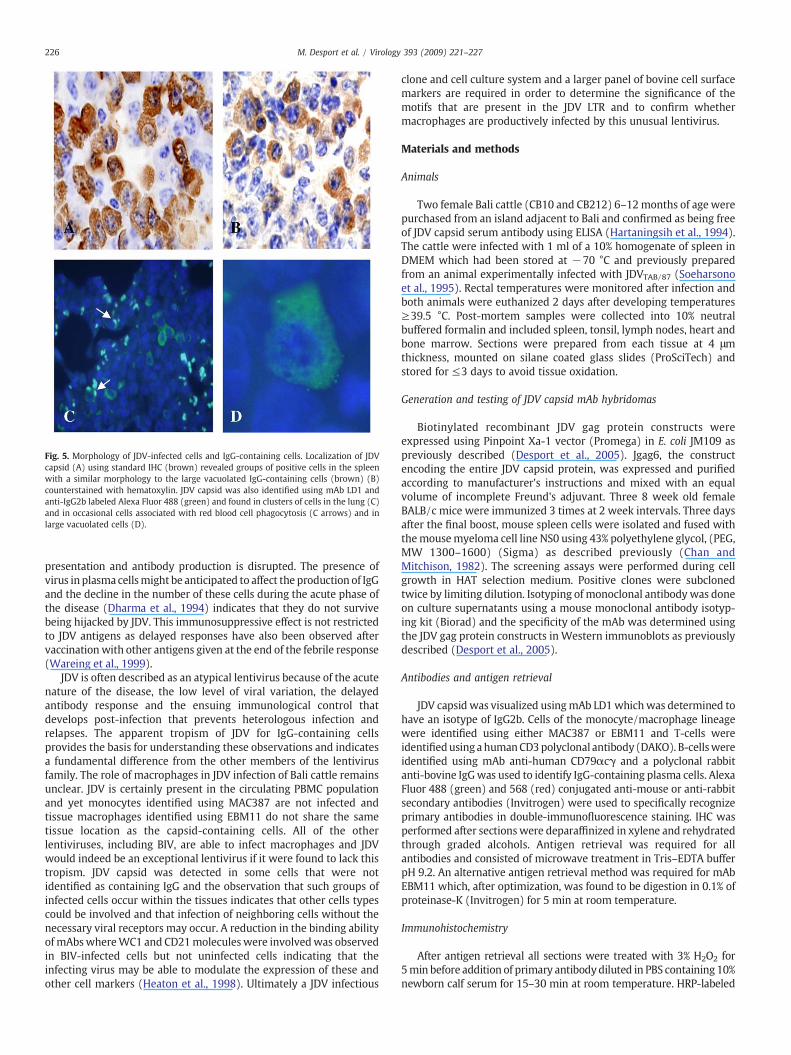
Fig. 5. Morphology of JDV-infected cells and IgG-containing cells. Localization of JDVcapsid (A) using standard IHC (brown) revealed groups of positive cells in the spleenwith a similar morphology to the large vacuolated IgG-containing cells (brown) (B)counterstained with hematoxylin. JDV capsid was also identified using mAb LD1 andanti-IgG2b labeled Alexa Fluor 488 (green) and found in clusters of cells in the lung (C)and in occasional cells associated with red blood cell phagocytosis (C arrows) and inlarge vacuolated cells (D).
226 M. Desport et al. / Virology 393 (2009) 221–227
presentation and antibody production is disrupted. The presence ofvirus in plasma cellsmight be anticipated to affect the production of IgGand the decline in the number of these cells during the acute phase ofthe disease (Dharma et al., 1994) indicates that they do not survivebeing hijacked by JDV. This immunosuppressive effect is not restrictedto JDV antigens as delayed responses have also been observed aftervaccinationwith other antigens given at the end of the febrile response(Wareing et al., 1999).
JDV is often described as an atypical lentivirus because of the acutenature of the disease, the low level of viral variation, the delayedantibody response and the ensuing immunological control thatdevelops post-infection that prevents heterologous infection andrelapses. The apparent tropism of JDV for IgG-containing cellsprovides the basis for understanding these observations and indicatesa fundamental difference from the other members of the lentivirusfamily. The role of macrophages in JDV infection of Bali cattle remainsunclear. JDV is certainly present in the circulating PBMC populationand yet monocytes identified using MAC387 are not infected andtissue macrophages identified using EBM11 do not share the sametissue location as the capsid-containing cells. All of the otherlentiviruses, including BIV, are able to infect macrophages and JDVwould indeed be an exceptional lentivirus if it were found to lack thistropism. JDV capsid was detected in some cells that were notidentified as containing IgG and the observation that such groups ofinfected cells occur within the tissues indicates that other cells typescould be involved and that infection of neighboring cells without thenecessary viral receptors may occur. A reduction in the binding abilityof mAbswhereWC1 and CD21molecules were involvedwas observedin BIV-infected cells but not uninfected cells indicating that theinfecting virus may be able to modulate the expression of these andother cell markers (Heaton et al., 1998). Ultimately a JDV infectious
clone and cell culture system and a larger panel of bovine cell surfacemarkers are required in order to determine the significance of themotifs that are present in the JDV LTR and to confirm whethermacrophages are productively infected by this unusual lentivirus.
Materials and methods
Animals
Two female Bali cattle (CB10 and CB212) 6–12months of age werepurchased from an island adjacent to Bali and confirmed as being freeof JDV capsid serum antibody using ELISA (Hartaningsih et al., 1994).The cattle were infected with 1 ml of a 10% homogenate of spleen inDMEM which had been stored at −70 °C and previously preparedfrom an animal experimentally infected with JDVTAB/87 (Soeharsonoet al., 1995). Rectal temperatures were monitored after infection andboth animals were euthanized 2 days after developing temperatures≥39.5 °C. Post-mortem samples were collected into 10% neutralbuffered formalin and included spleen, tonsil, lymph nodes, heart andbone marrow. Sections were prepared from each tissue at 4 μmthickness, mounted on silane coated glass slides (ProSciTech) andstored for ≤3 days to avoid tissue oxidation.
Generation and testing of JDV capsid mAb hybridomas
Biotinylated recombinant JDV gag protein constructs wereexpressed using Pinpoint Xa-1 vector (Promega) in E. coli JM109 aspreviously described (Desport et al., 2005). Jgag6, the constructencoding the entire JDV capsid protein, was expressed and purifiedaccording to manufacturer's instructions and mixed with an equalvolume of incomplete Freund's adjuvant. Three 8 week old femaleBALB/c mice were immunized 3 times at 2 week intervals. Three daysafter the final boost, mouse spleen cells were isolated and fused withthemousemyeloma cell line NS0 using 43% polyethylene glycol, (PEG,MW 1300–1600) (Sigma) as described previously (Chan andMitchison, 1982). The screening assays were performed during cellgrowth in HAT selection medium. Positive clones were subclonedtwice by limiting dilution. Isotyping of monoclonal antibody was doneon culture supernatants using a mouse monoclonal antibody isotyp-ing kit (Biorad) and the specificity of the mAb was determined usingthe JDV gag protein constructs in Western immunoblots as previouslydescribed (Desport et al., 2005).
Antibodies and antigen retrieval
JDV capsid was visualized usingmAb LD1whichwas determined tohave an isotype of IgG2b. Cells of the monocyte/macrophage lineagewere identified using either MAC387 or EBM11 and T-cells wereidentifiedusing ahumanCD3polyclonal antibody (DAKO). B-cellswereidentified using mAb anti-human CD79αcγ and a polyclonal rabbitanti-bovine IgGwas used to identify IgG-containing plasma cells. AlexaFluor 488 (green) and 568 (red) conjugated anti-mouse or anti-rabbitsecondary antibodies (Invitrogen) were used to specifically recognizeprimary antibodies in double-immunofluorescence staining. IHC wasperformed after sectionswere deparaffinized in xylene and rehydratedthrough graded alcohols. Antigen retrieval was required for allantibodies and consisted of microwave treatment in Tris–EDTA bufferpH 9.2. An alternative antigen retrieval method was required for mAbEBM11 which, after optimization, was found to be digestion in 0.1% ofproteinase-K (Invitrogen) for 5 min at room temperature.
Immunohistochemistry
After antigen retrieval all sections were treated with 3% H2O2 for5min before addition of primary antibodydiluted in PBS containing 10%newborn calf serum for 15–30 min at room temperature. HRP-labeled

227M. Desport et al. / Virology 393 (2009) 221–227
streptavidin–biotin reagents (LSAB2, DAKO) with specificity for mouseIgGwere used to directly detect primarymouse antibodies using DAB asthe substrate chromogen. Specificity of antibodies was confirmed byomission of primary antibody and testing on uninfected tissues.
Double-immunofluorescent staining
The slides were incubated with mixtures of 2 primary antibodies,using dilutions which had been established after preliminary titra-tions, for 15–30 min at room temperature. After washing 3 times withTBS, the appropriate secondary antibodies labeled with Alexa Fluor488 or 568, either fromdifferent species orwith differing Ig subclasses,were applied in TBS and incubated in the dark for 15–30 min.Additional sensitivity was obtained, where necessary, by using a ratanti-mouse IgG biotinylated secondary antibody thatwas subsequent-ly detected using streptavidin Alexa Fluor 488 (Invitrogen). Finally,slides were washed with TBS and dried in the dark for 15 min, beforebeing mounted in DAPI mounting medium (Vector Laboratories).
Acknowledgments
Weare extremely grateful to G. Thomson,M. Slaven andG. Spoelstrafor their expert technical assistance and helpful discussions. We aregrateful to the staff at the Disease Investigation Centre in Bali for theircareful collection and processing of samples. All animal researchcomplied with the ethics guidelines at the Disease Investigation Centre,Denpasar. This work was funded by the Australian Centre forInternational Agricultural Research grant No. AS1/2004/074.
References
Ackermann, M.R., DeBey, B.M., Stabel, T.J., Gold, J.H., Register, K.B., Meehan, J.T., 1994.Distribution of anti-CD68 (EBM11) immunoreactivity in formalin-fixed, paraffin-embedded bovine tissues. Vet. Pathol. 31 (3), 340–348.
Andresdottir, V., Tang, X., Agnarsdottir, G., Andresson, O.S., Georgsson, G., Skraban, R.,Torsteinsdottir, S., Rafnar, B., Benediktsdottir, E., Matthiasdottir, S., Arnadottir, S.,Hognadottir, S., Palsson, P.A., Petursson, G., 1998. Biological and genetic differencesbetween lung- and brain-derived isolates of maedi-visna virus. Virus Genes 16 (3),281–293.
Bielefeldt-Ohmann, H., Sabara, M., Lawman, M.J., Griebel, P., Babiuk, L.A., 1988. Amonoclonal antibody detects macrophage maturation antigen which appearsindependently of class II antigen expression. Reactivity of monoclonal EBM11 withbovine macrophages. J. Immunol. 140 (7), 2201–2209.
Brandtzaeg, P., 1988. The new monoclonal antibody (Mac 387) that reacts withmacrophages on paraffin sections detects the well-known leukocyte L1 antigen.J. Histochem. Cytochem. 36 (9), 1203–1206.
Chadwick, B.J., Coelen, R.J., Wilcox, G.E., Sammels, L.M., Kertayadnya, G., 1995.Nucleotide sequence analysis of Jembrana disease virus: a bovine lentivirusassociated with an acute disease syndrome. J. Gen. Virol. 76 (Pt 7), 1637–1650.
Chadwick, B.J., Desport, M., Brownlie, J., Wilcox, G.E., Dharma, D.M., 1998. Detection ofJembrana disease virus in spleen, lymph nodes, bonemarrow and other tissues by insitu hybridization of paraffin-embedded sections. J. Gen. Virol. 79 (Pt. 1), 101–106.
Chakrabarti, L., Hurtrel, M., Maire, M.A., Vazeux, R., Dormont, D., Montagnier, L., Hurtrel,B., 1991. Early viral replication in the brain of SIV-infected rhesus monkeys. Am. J.Pathol. 139 (6), 1273–1280.
Chan, W.L., Mitchison, N.A., 1982. The use of somatic cell hybrids for the production ofmonospecific viral antibodies. Lab. Res. Methods Biol. Med. 5, 125–141.
Chirmule, N., Kalyanaraman, V.S., Saxinger, C., Wong-Staal, F., Ghrayeb, J., Pahwa, S.,1990. Localization of B-cell stimulatory activity of HIV-1 to the carboxyl terminus ofgp41. AIDS Res. Hum. Retroviruses 6 (3), 299–305.
Chirmule, N., Oyaizu, N., Saxinger, C., Pahwa, S., 1994. Nef protein of HIV-1 has B-cellstimulatory activity. Aids 8 (6), 733–734.
Desport,M., Ditcham,W.G., Lewis, J.R.,McNab, T.J., Stewart,M.E., Hartaningsih,N.,Wilcox,G.E., 2009. Analysis of Jembrana disease virus replication dynamics in vivo revealsstrain variation and atypical responses to infection. Virology 386 (2), 310–316.
Desport, M., Stewart, M.E., Sheridan, C.A., Ditcham,W.G., Setiyaningsih, S., Tenaya,W.M.,Hartaningsih, N., Wilcox, G.E., 2005. Recombinant Jembrana disease virus gagproteins identify several different antigenic domains but do not facilitate serologicaldifferentiation of JDV and nonpathogenic bovine lentiviruses. J. Virol. Methods 124(1–2), 135–142.
Desport, M., Stewart, M.E., Mikosza, A.S., Sheridan, C.A., Peterson, S.E., Chavand, O.,Hartaningsih, N., Wilcox, G.E., 2007. Sequence analysis of Jembrana disease virusstrains reveals a genetically stable lentivirus. Virus Res. 126 (1–2), 233–244.
Dharma, D.M., Budiantono, A., Campbell, R.S., Ladds, P.W., 1991. Studies onexperimental Jembrana disease in Bali cattle. III. Pathology. J. Comp. Pathol. 105(4), 397–414.
Dharma, D.M., Ladds, P.W., Wilcox, G.E., Campbell, R.S., 1994. Immunopathology ofexperimental Jembrana disease in Bali cattle. Vet. Immunol. Immunopathol. 44 (1),31–44.
Dykhuizen, M., Mitchen, J.L., Montefiori, D.C., Thomson, J., Acker, L., Lardy, H., Pauza,C.D., 1998. Determinants of disease in the simian immunodeficiency virus-infected rhesus macaque: characterizing animals with low antibody responsesand rapid progression. J. Gen. Virol. 79 (Pt. 10), 2461–2467.
English, R.V., Johnson, C.M., Gebhard, D.H., Tompkins, M.B., 1993. In vivo lymphocytetropism of feline immunodeficiency virus. J. Virol. 67 (9), 5175–5186.
Fischer-Smith, T., Croul, S., Adeniyi, A., Rybicka, K., Morgello, S., Khalili, K., Rappaport, J.,2004. Macrophage/microglial accumulation and proliferating cell nuclear antigenexpression in the central nervous system in human immunodeficiency virusencephalopathy. Am. J. Pathol. 164 (6), 2089–2099.
Garvey, K.J., Oberste, M.S., Elser, J.E., Braun, M.J., Gonda, M.A., 1990. Nucleotidesequence and genome organization of biologically active proviruses of the bovineimmunodeficiency-like virus. Virology 175 (2), 391–409.
Gendelman, H.E., Narayan, O., Molineaux, S., Clements, J.E., Ghotbi, Z., 1985. Slow,persistent replication of lentiviruses: role of tissue macrophages and macrophageprecursors in bone marrow. Proc. Natl. Acad. Sci. U. S. A. 82 (20), 7086–7090.
Greywoode, G.I., McCarthy, S.P., McGee, J.O., 1990. Labelling of cells of the mononuclearphagocyte system in routinely processed archival biopsy specimens withmonoclonal antibody EBM/11. J. Clin. Pathol. 43 (12), 992–996.
Hartaningsih, N., Wilcox, G.E., Kertayadnya, G., Astawa, M., 1994. Antibody response toJembrana disease virus in Bali cattle. Vet. Microbiol. 39 (1–2), 15–23.
Heaton, P.R., Johnstone, P., Brownlie, J., 1998. Investigation of the cellular tropism ofbovine immunodeficiency-like virus. Res. Vet. Sci. 65 (1), 33–40.
Henderson, A.J., Calame, K.L., 1997. CCAAT/enhancer binding protein (C/EBP) sites arerequired for HIV-1 replication in primary macrophages but not CD4(+) T cells.Proc. Natl. Acad. Sci. U. S. A. 94 (16), 8714–8719.
Henderson, A.J., Connor, R.I., Calame, K.L., 1996. C/EBP activators are required for HIV-1replication and proviral induction in monocytic cell lines. Immunity 5 (1), 91–101.
Hines, R., Sorensen, B.R., Shea, M.A., Maury, W., 2004. PU.1 binding to ETS motifs withinthe equine infectious anemia virus long terminal repeat (LTR) enhancer: regulationof LTR activity and virus replication in macrophages. J. Virol. 78 (7), 3407–3418.
Michael, N.L., Brown, A.E., Voigt, R.F., Frankel, S.S., Mascola, J.R., Brothers, K.S., Louder,M., Birx, D.L., Cassol, S.A., 1997. Rapid disease progression without seroconversionfollowing primary human immunodeficiency virus type 1 infection—evidence forhighly susceptible human hosts. J. Infect. Dis. 175 (6), 1352–1359.
O'Neil, S.P., Mossman, S.P., Maul, D.H., Hoover, E.A., 1999. In vivo cell and tissue tropismof SIVsmmPBj14-bcl.3. AIDS Res. Hum. Retroviruses 15 (2), 203–215.
Onuma, M., Koomoto, E., Furuyama, H., Yasutomi, Y., Taniyama, H., Iwai, H., Kawakami,Y., 1992. Infection and dysfunction of monocytes induced by experimentalinoculation of calves with bovine immunodeficiency-like virus. J. Acquir. Immune.Defic. Syndr. 5 (10), 1009–1015.
Page, G., Sattler, A., Kersten, S., Thiel, A., Radbruch, A., Miossec, P., 2004. Plasma cell-likemorphology of Th1-cytokine-producing cells associated with the loss of CD3expression. Am. J. Pathol. 164 (2), 409–417.
Pifat, D.Y., Ennis, W.H., Ward, J.M., Oberste, M.S., Gonda, M.A., 1992. Persistent infectionof rabbits with bovine immunodeficiency-like virus. J. Virol. 66 (7), 4518–4524.
Poston, R.N., Hussain, I.F., 1993. The immunohistochemical heterogeneity of atheromamacrophages: comparison with lymphoid tissues suggests that recently blood-derivedmacrophages can be distinguished from longer-resident cells. J. Histochem.Cytochem. 41 (10), 1503–1512.
Sellon, D.C., Perry, S.T., Coggins, L., Fuller, F.J., 1992. Wild-type equine infectious anemiavirus replicates in vivo predominantly in tissue macrophages, not in peripheralblood monocytes. J. Virol. 66 (10), 5906–5913.
Smit, T.K., Wang, B., Ng, T., Osborne, R., Brew, B., Saksena, N.K., 2001. Varied tropism ofHIV-1 isolates derived from different regions of adult brain cortex discriminatebetween patients with and without AIDS dementia complex (ADC): evidence forneurotropic HIV variants. Virology 279 (2), 509–526.
Soeharsono, S., Hartaningsih, N., Soetrisno, M., Kertayadnya, G., Wilcox, G.E., 1990.Studies of experimental Jembrana disease in Bali cattle. I. Transmission andpersistence of the infectious agent in ruminants and pigs, and resistance ofrecovered cattle to re-infection. J. Comp. Pathol. 103 (1), 49–59.
Soeharsono, S., Wilcox, G.E., Putra, A.A., Hartaningsih, N., Sulistyana, K., Tenaya, M.,1995. The transmission of Jembrana disease, a lentivirus disease of Bos javanicuscattle. Epidemiol. Infect. 115 (2), 367–374.
Soesanto, M., Soeharsono, S., Budiantono, A., Sulistyana, K., Tenaya, M., Wilcox, G.E.,1990. Studies on experimental Jembrana disease in Bali cattle. II. Clinical signs andhaematological changes. J. Comp. Pathol. 103 (1), 61–71.
Wareing, S., Hartaningsih, N., Wilcox, G.E., Penhale, W.J., 1999. Evidence forimmunosuppression associated with Jembrana disease virus infection of cattle.Vet. Microbiol. 68 (1–2), 179–185.
Whetstone, C.A., Suarez, D.L., Miller, J.M., Pesch, B.A., Harp, J.A., 1997. Bovine lentivirusinduces early transient B-cell proliferation in experimentally inoculated cattle andappears to be pantropic. J. Virol. 71 (1), 640–644.
Wilcox, G.E., Kertayadnya, G., Hartaningsih, N., Dharma, D.M., Soeharsono, S.,Robertson, T., 1992. Evidence for a viral aetiology of Jembrana disease in Balicattle. Vet. Microbiol. 33 (1–4), 367–374.
Zhang, Z.Q., Casimiro, D.R., Schleif, W.A., Chen, M., Citron, M., Davies, M.E., Burns, J.,Liang, X., Fu, T.M., Handt, L., Emini, E.A., Shiver, J.W., 2007. Early depletion ofproliferating B cells of germinal center in rapidly progressive simian immunode-ficiency virus infection. Virology 361 (2), 455–464.
Zink, M.C., Yager, J.A., Myers, J.D., 1990. Pathogenesis of caprine arthritis encephalitisvirus. Cellular localization of viral transcripts in tissues of infected goats. Am. J.Pathol. 136 (4), 843–854.
Related Documents











