Annals of Anatomy 193 (2011) 479–485 Contents lists available at SciVerse ScienceDirect Annals of Anatomy jo ur n al ho mepage: www.elsevier.de/aanat Research article In vitro evidence of involvement of the epithelial y + transporter in -defensin production on the ocular surface Kristin Jäger a,∗ , Andrea Nielitz a , Fabian Garreis a,b , Saadettin Sel c , Heike Nave a , Friedrich P. Paulsen a,b a Department of Anatomy and Cell Biology, Martin Luther University of Halle-Wittenberg, D-06097 Halle, Saale, Germany b Department of Anatomy II, Friedrich Alexander University of Erlangen-Nuremberg, Erlangen, Germany c Department of Ophthalmology, Martin-Luther-University Halle-Wittenberg, Halle, Saale, Germany a r t i c l e i n f o Article history: Received 19 September 2011 Received in revised form 6 October 2011 Accepted 8 October 2011 Keywords: -Defensins Cationic amino acid transporter Innate immune system Corneal epithelium s u m m a r y To analyse the hypothesis as to whether there is a functional relationship between human cationic amino acid transporters (hCATs, y + transporter, the main transporter of l-arginine and l-lysine) and human -defensin (important components of immune function) production on the ocular surface, arginase and nitrate monoxide synthase (NOS), enzymes that compete for l-arginine, were inhibited by norNOHA (N(omega)-hydroxy-nor-l-arginine) and/or L-NAME (NG-nitro-l-arginine methyl ester) in cultured human corneal epithelial cells. In addition, the transport activity of hCAT proteins was inhib- ited or activated through -tocopherol or PMA (phorbol myristate acetate), respectively. Concentrations of the human inducible -defensins (hBD) 2 and 3 were determined by ELISA experiments. The basic expression of hBD3 in non-stimulated HCE cells significantly exceeded that of hBD2. Both -defensins also differed as to how readily their excretion could be stimulated. HBD2 excretion rate was 3.5 time more by L-NAME, whereas norNOHA had no effect. In contrast, hBD3 excretion was increased by norNOHA by a factor of 1.5 but L-NAME alone had no effect. The excretion of both -defensins was increased 3- and 6-fold by combined administration of L-NAME, norNOHA and interleukin (IL)-1. Administration of - tocopherol increased hBD2 excretion twofold. No effect was observed for hBD3. With PMA, on the other hand, a reduction in secretion for both -defensins was observed. These in vitro findings provide evi- dence of a functional association between CAT proteins and -defensins 2 and 3 opening up a new field of research with pharmacological perspectives for treatment of inflammatory diseases such as keratitis or dry eye disease. © 2011 Elsevier GmbH. All rights reserved. 1. Introduction Bacterial keratitis is one of the most frequent diagnoses in opthalmological practice. The causal pathogens of keratitis are fre- quently Staphylococcus aureus and Pseudomonas aeruginosa (Fleisig et al., 1992; O’Callaghan et al., 1997). The bacterial metabolites and toxins, as well as the inflammatory reactions of the host, often result in severe tissue damage with permanent scarring and even loss of eyesight (Baum, 1995). Administration of antibiotics is the only fast and effective method available to date of treat- ing such bacterial infections. This approach is becoming more and more problematic in view of increasing resistance to “conven- tional” antibiotics and the search for effective alternatives. Potential alternatives include so called host-defence peptides (HDPs) which have been shown to prevent infection and are likely involved in ∗ Corresponding author. Tel.: +49 3455571740; fax: +49 34505571700. E-mail address: [email protected] (K. Jäger). wound-healing responses and tissue remodelling (Schroder, 1999; Harder et al., 2001) also on the ocular surface (for review see Kolar and McDermott, 2011; McDermott, 2009). These endogenous HDPs may be involved in ocular immune and inflammatory responses although much remains to be discovered about such roles (Kolar and McDermott, 2011). Devel- opment of an HDP or derivative/mimic thereof for therapeutic purposes is an attractive proposition, and because of ease of application to the ocular surface (topical or local injection), reducing some of the confounding factors associated with sys- temic delivery, is indeed a viable option (Kolar and McDermott, 2011). Although HDPs occur as natural antibiotics in various organisms, pathogenic germs have not developed any signifi- cant resistances in the course of evolution. The HDPs on the ocular surface that have been most thoroughly described and investigated are the -defensins (Kolar and McDermott, 2011; McDermott, 2009; Gottsch et al., 1998; Haynes et al., 1998). They are produced by the outmost epithelial cells of the cornea and con- junctiva in keeping with their role as a chemical shield to invading 0940-9602/$ – see front matter © 2011 Elsevier GmbH. All rights reserved. doi:10.1016/j.aanat.2011.10.001

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

R
Ip
Ka
b
c
a
ARRA
K�CIC
1
oqeaoeiimtah
0d
Annals of Anatomy 193 (2011) 479– 485
Contents lists available at SciVerse ScienceDirect
Annals of Anatomy
jo ur n al ho mepage: www.elsev ier .de /aanat
esearch article
n vitro evidence of involvement of the epithelial y+ transporter in �-defensinroduction on the ocular surface
ristin Jägera,∗, Andrea Nielitza, Fabian Garreisa,b, Saadettin Sel c, Heike Navea, Friedrich P. Paulsena,b
Department of Anatomy and Cell Biology, Martin Luther University of Halle-Wittenberg, D-06097 Halle, Saale, GermanyDepartment of Anatomy II, Friedrich Alexander University of Erlangen-Nuremberg, Erlangen, GermanyDepartment of Ophthalmology, Martin-Luther-University Halle-Wittenberg, Halle, Saale, Germany
r t i c l e i n f o
rticle history:eceived 19 September 2011eceived in revised form 6 October 2011ccepted 8 October 2011
eywords:-Defensinsationic amino acid transporter
nnate immune systemorneal epithelium
s u m m a r y
To analyse the hypothesis as to whether there is a functional relationship between human cationicamino acid transporters (hCATs, y+ transporter, the main transporter of l-arginine and l-lysine) andhuman �-defensin (important components of immune function) production on the ocular surface,arginase and nitrate monoxide synthase (NOS), enzymes that compete for l-arginine, were inhibitedby norNOHA (N(omega)-hydroxy-nor-l-arginine) and/or L-NAME (NG-nitro-l-arginine methyl ester) incultured human corneal epithelial cells. In addition, the transport activity of hCAT proteins was inhib-ited or activated through �-tocopherol or PMA (phorbol myristate acetate), respectively. Concentrationsof the human inducible �-defensins (hBD) 2 and 3 were determined by ELISA experiments. The basicexpression of hBD3 in non-stimulated HCE cells significantly exceeded that of hBD2. Both �-defensinsalso differed as to how readily their excretion could be stimulated. HBD2 excretion rate was 3.5 time moreby L-NAME, whereas norNOHA had no effect. In contrast, hBD3 excretion was increased by norNOHA bya factor of 1.5 but L-NAME alone had no effect. The excretion of both �-defensins was increased 3- and
6-fold by combined administration of L-NAME, norNOHA and interleukin (IL)-1�. Administration of �-tocopherol increased hBD2 excretion twofold. No effect was observed for hBD3. With PMA, on the otherhand, a reduction in secretion for both �-defensins was observed. These in vitro findings provide evi-dence of a functional association between CAT proteins and �-defensins 2 and 3 opening up a new fieldof research with pharmacological perspectives for treatment of inflammatory diseases such as keratitisor dry eye disease.© 2011 Elsevier GmbH. All rights reserved.
. Introduction
Bacterial keratitis is one of the most frequent diagnoses inpthalmological practice. The causal pathogens of keratitis are fre-uently Staphylococcus aureus and Pseudomonas aeruginosa (Fleisigt al., 1992; O’Callaghan et al., 1997). The bacterial metabolitesnd toxins, as well as the inflammatory reactions of the host,ften result in severe tissue damage with permanent scarring andven loss of eyesight (Baum, 1995). Administration of antibioticss the only fast and effective method available to date of treat-ng such bacterial infections. This approach is becoming more and
ore problematic in view of increasing resistance to “conven-
ional” antibiotics and the search for effective alternatives. Potentiallternatives include so called host-defence peptides (HDPs) whichave been shown to prevent infection and are likely involved in∗ Corresponding author. Tel.: +49 3455571740; fax: +49 34505571700.E-mail address: [email protected] (K. Jäger).
940-9602/$ – see front matter © 2011 Elsevier GmbH. All rights reserved.oi:10.1016/j.aanat.2011.10.001
wound-healing responses and tissue remodelling (Schroder, 1999;Harder et al., 2001) also on the ocular surface (for review see Kolarand McDermott, 2011; McDermott, 2009).
These endogenous HDPs may be involved in ocular immuneand inflammatory responses although much remains to bediscovered about such roles (Kolar and McDermott, 2011). Devel-opment of an HDP or derivative/mimic thereof for therapeuticpurposes is an attractive proposition, and because of ease ofapplication to the ocular surface (topical or local injection),reducing some of the confounding factors associated with sys-temic delivery, is indeed a viable option (Kolar and McDermott,2011). Although HDPs occur as natural antibiotics in variousorganisms, pathogenic germs have not developed any signifi-cant resistances in the course of evolution. The HDPs on theocular surface that have been most thoroughly described and
investigated are the �-defensins (Kolar and McDermott, 2011;McDermott, 2009; Gottsch et al., 1998; Haynes et al., 1998). Theyare produced by the outmost epithelial cells of the cornea and con-junctiva in keeping with their role as a chemical shield to invading
4 Anato
oMeo1eiacblMSwa(ctb2boH2ien2
twaste
amefcst(
h(antilctisieti
uttm(
80 K. Jäger et al. / Annals of
rganisms. As recently thoroughly reviewed by Kolar andcDermott (2011), corneal and conjunctival cells constitutively
xpress hBD-1. In some studies, expression of hBD2 is found onlyccasionally in normal tissue (Hattenbach et al., 1998; Haynes et al.,999; McNamara et al., 1999; Lehmann et al., 2000; McDermottt al., 2003; Narayanan et al., 2003) whereas a recent study revealedts constitutive expression in superficial epithelial cells of corneand conjunctiva (Garreis et al., 2010). In vitro studies with culturedorneal and conjunctival epithelial cells have demonstrated thatacterial products and proinflammatory cytokines such as inter-
eukin (IL)-1� upregulate hBD2 expression (McNamara et al., 1999;cDermott et al., 2003; Narayanan et al., 2003; Garreis et al., 2010;
hin et al., 2004; Kumar et al., 2006, 2007; Li et al., 2008) and studiesith supernatants of S. aureus in mice have demonstrated that this
lso occurs in vivo by upregulating the mouse homologue of hBD2Garreis et al., 2010). Corneal and conjunctival epithelial cells alsoonstitutively express hBD3 which is upregulated in vitro in cul-ured ocular surface epithelial cells in response to cytokines andacterial supernatants (McNamara et al., 1999; McDermott et al.,003; Garreis et al., 2010; McIntosh et al., 2005). Further mem-ers detected on the ocular surface are hBD4 which was foundnly infrequently in actual tissue samples (McIntosh et al., 2005;uang et al., 2007) and hBD9 (Abedin et al., 2008; Mohammed et al.,010). Of interest, hBD9 reveal decreased expression in response to
nflammation and infection (Abedin et al., 2008). Expression of sev-ral other members of the �-defensin family has been sought, butone appear to be present on the ocular surface (McIntosh et al.,005; Huang et al., 2007).
�-Defensins are low molecular weight cationic oligopeptideshat bring about lysis of a broad spectrum of microorganisms byay of their interaction with biomembranes (Schroder, 1999). They
re able to interact with microbial membranes only due to theirtrong net positive charge, a result of their extremely high propor-ion of cationic amino acids, especially l-arginine and l-lysine (Zout al., 2007).
For the synthesis of �-defensins, high concentrations of themino acids l-arginine and l-lysine are necessary. During inflam-atory processes these amino acids must be supplied by the
pithelial cells that produce the �-defensins. The limiting factoror this supply is the transport of l-arginine and l-lysine into theell. The transport is enabled to nearly 80% by only one transporterystem, human cationic amino acid transporter proteins (hCAT pro-eins, y+ transporters) a subfamily of the solute carrier family 7SLC7) (Deves and Boyd, 1998).
CAT proteins are encoded by at least 4 transcripts in humans:CAT1 (SLC7A1), 2a (SLC7A2A), 2b (SLC7A2B) and 3 (SLC7A3)Hoshide et al., 1996; Vekony et al., 2001). Other transporters suchs B0,+, b0,+ and y+-L, which can also transport l-arginine and l-lysineon-specifically, tend to play a lesser role in humans in transport ofhese two cationic amino acids (Deves and Boyd, 1998). Althought has just recently been demonstrated that an increased supply of-arginine correlates with increased expression of hBD-1 in colonarcinoma cells (HCT-116) (Sherman et al., 2006), the limiting fac-or to transport of the cationic amino acids into the cell has not beennvestigated to date. In previous investigations we could demon-trate that hCAT1 and hCAT2 are expressed on the ocular surfacen healthy cornea and conjunctiva whereas hCAT3 is absent (Jägert al., 2009, 2010a,b). Cell culture experiments have demonstratedhat proinflammatory cytokines and bacterial products are able toncrease hCAT1 and hCAT2 expression (Jäger et al., 2009).
The detection of the presence of CATs on the ocular surface leds to speculate about a possible functional relationship between
hem and �-defensins in a regulatory circuit via which the balanceoward �-defensin production could be shifted to treat inflam-atory diseases such as bacterial keratitis or dry eye syndromeJäger et al., 2010a,b). A changed supply of the cationic amino acids
my 193 (2011) 479– 485
l-lysine and l-arginine in the cell due to activity blockade of theCATs or a balance shift in the metabolic pathways competing forthe amino acid l-arginine should result in a change in �-defensinproduction. To test this hypothesis the present in vitro study wasundertaken in which l-arginine metabolism was experimentallyimpaired in several ways described in detail in the following.l-Arginine is a semi-essential amino acid that plays a central
role in metabolism. In the human body, l-arginine is metabo-lized via two main pathways. One leads from arginase to ureaand l-ornithine (Jenkinson and Grigor, 1994) and the other fromnitrate monoxide synthase (NOS) to nitrogen monoxide and l-citrulline (Feelisch and Stammler, 1996) (Fig. 1). Both enzymes arecan be inhibited and production in each case can be shifted to theother respective metabolic pathway (Balkan et al., 1997). NorNOHA(Nω-hydroxy-nor-l-arginine) is used as an effective competitiveinhibitor of arginase (Tate et al., 2008; Tenu et al., 1999), but isneither a substrate nor an inhibitor of iNOS (inducible NOS). L-NAME (N(G)-nitro-l-arginine methyl ester) is an arginine derivatethat acts as a competitive inhibitor of nitrate monoxide synthase(Balkan et al., 1997). For blocking experiments, norNOHA and L-NAME were tested as potential inhibitors of cationic amino acidtransporter proteins hCAT1, 2a and b.
2. Materials and methods
2.1. Cell lines
SV40-transformed human corneal epithelial cells in the 36thpassage (HCE cells, a kind gift from Kaoru Araki-Sasaki, Tane Memo-rial Eye Hospital, Osaka, Japan; Araki-Sasaki et al., 1995) werecultured as monolayers and used for stimulation experiments.
2.2. Cell culture and stimulation
HCE cells were cultured in Dulbecco modified Eagle medium(DMEM/HAMs F12 1:1; PAA Laboratories GmbH, Pasching, Austria)containing 10% foetal calf serum (FCS; Biochrom AG, Berlin,Germany), in a humidified 5% CO2 incubator at 37 ◦C. When conflu-ence was reached, cells were washed in phosphate-buffered saline(PBS) and incubated in serum-free media overnight. Cells were thentreated with different dilutions of stimulants and for different incu-bation times, 2 mM norNOHA for 48 h (Tenu et al., 1999), 2 mML-NAME for 24 h (Balkan et al., 1997), 50 �M �-tocopherol for 24 h(Ricciarelli et al., 1998), 100 nM phorbol myristate acetate (PMAfor 4 h; Krotova et al., 2003) and 10 ng/ml interleukin-1� (IL-1� for24 h; Jäger et al., 2009).
To ensure that norNOHA and L-NAME inhibit arginase and NOsynthetase respectively and do not completely block the cationicamino acid transporter proteins hCAT1, 2a and 2b themselves,uptake investigations were undertaken previously to the stimu-lation experiments revealing that norNOHA and L-NAME have onlyan inhibitory effect of 15 up to 20% on hCATs and therefore areuseful for the present study (data not shown).
2.2.1. ELISACells and culture supernatants obtained during the experiments
were processed for enzyme-linked immunosorbent assay (ELISA)of hBD2 and hBD3. Sandwich ELISA 96-well immunoplates (Max-iSorp, Nunc, Roskilde, Denmark) were coated at 4 ◦C overnight with0.5 �g/ml of capture antibody (goat anti-hBD2, 500-P161G; rabbitanti-hBD3, 500-P241; PeproTech Inc., Rocky Hill, USA). One hun-dred microlitres of supernatants of each sample were analyzed as
previously described (Garreis et al., 2010; Varoga et al., 2004). Wellswere incubated for one hour at room temperature with 0.2 �g/ml ofbiotinylated detection antibody (biotinylated goat anti-hBD2, 500-P161GBt; biotinylated rabbit anti hBD3, 500-P241Bt; PeproTech
K. Jäger et al. / Annals of Anatomy 193 (2011) 479– 485 481
Fig. 1. Scheme of l-arginine metabolism. The incorporation of the individual proteins (human �-defensins; hCAT = y+-transporter = human cationic amino acid transporter)a = nitrc y of inc argini
IaPPlrrceho
2
stDt
3
3n
c(s
3ps
h2
11700.0 ± 2151 pg/ml, over three times the hBD3 control value. Theblockade of NOS with L-NAME or the inflammatory stimulation byIL-1�, on the other hand, had no significant influence (Fig. 3B).
Fig. 2. Quantification of hBD2 and hBD3 concentrations in SV40-transformedhuman corneal epithelial cells. The supernatants of non-stimulated HCE cells were
nd their metabolites (L-arg = l-arginine; L-orn = l-ornithine; L-citr = l-citruline; NOan be restored or pushed in a desired direction by means of alterations in the supplircuit. L-lys = l-lysine; NOS = nitric oxide synthetase; L-NAME = Nω-hydroxy-nor-l-
nc., Rocky Hill, USA). Absorbance was measured at 405 nm with multichannel photometer. Human recombinant hBD2 (300-49;eproTech Inc., Rocky Hill, USA,) and recombinant hBD3 (300-52;eproTech Inc., Rocky Hill, USA) served as the standard at the fol-owing concentrations: 0, 0.078, 0.156, 0.31, 0.625, 1.25, 2.5 ng/mlecombinant hBD2 and 0, 0.156, 0.31, 0.625, 1.25, 2.5, 5, 10 ng/mlecombinant hBD3. Standard, sample supernatants and negativeontrols were performed in duplicate. Additionally, for all ELISAxperiments, cell proteins were measured, with only weak hBD2 orBD3 signals measurable, in this paper all values are shown basedn measurements of the cell supernatants.
.3. Statistical analysis
Data are expressed as the mean ± standard error (SEM) of testedamples. Statistical significance was evaluated with the Student’s-test using InStat Statistical software (Graphpad Software, Saniego, CA, USA). P values of less than 0.05 were considered sta-
istically significant.
. Results
.1. Comparison of hBD2 and hBD3 concentrations inon-stimulated HCE cells
Comparison of the constitutive expression of hBD2 and hBD3 inultured HCE cells revealed a protein concentration of 9.2 pg/ml±4.9 pg/ml) hBD2, and 3701 pg/ml (±874 pg/ml) hBD3 in non-timulated human corneal epithelial cells (Fig. 2, n = 9).
.2. Inhibition of NOS by L-NAME induces an increase of the hBD2rotein concentration whereas inhibition of arginase by norNOHAtimulates the hBD3 protein concentration
The NOS inhibitor L-NAME stimulated the expression ofBD2 in HCE cells significantly, 3.5-fold, from 8.5 ± 2.17 pg/ml to9.0 ± 3 pg/ml (Fig. 3A). The blockade of the arginase metabolic
ic oxide; urea) in a regulatory circuit is the basis of the present study. The balancedividual metabolites and stimulation or inhibition of the proteins in this regulatoryne; norNOHA = N(G)-nitro-l-arginine methyl ester.
pathway with norNOHA revealed no effect, or no additional effect,on hBD2 secretion (Fig. 3A). The inflammatory stimulation with10 ng/ml IL1� 18.0 ± 2.2 pg/ml had no significant effect of thehBD2 secretion. Addition of the pro-inflammatory cytokine IL-1�in combination with norNOHA and L-NAME resulted in a sig-nificant, six-fold, increase in concentration (to 60.0 ± 7.3 pg/ml).Blocking the arginase with norNOHA, on the other hand, resultedin a significant increase in concentration of hBD3 from approxi-mately 3610.0 ± 375 pg/ml to 5050 ± 473 pg/ml (Fig. 3B). Combinedapplication of norNOHA + L-NAME + IL-1� even raised the level to
collected and determined by enzyme-linked immunosorbent assay (ELISA, n = 9).Values were compared between hBD2 and hBD3 concentrations in non-stimulatedHCE cells. Amounts of hBD2 and hBD3 concentrations were normalized with totalprotein concentration from cell extracts. Columns represent mean ± SEM of fourindependent measurements. ***p < 0.001.
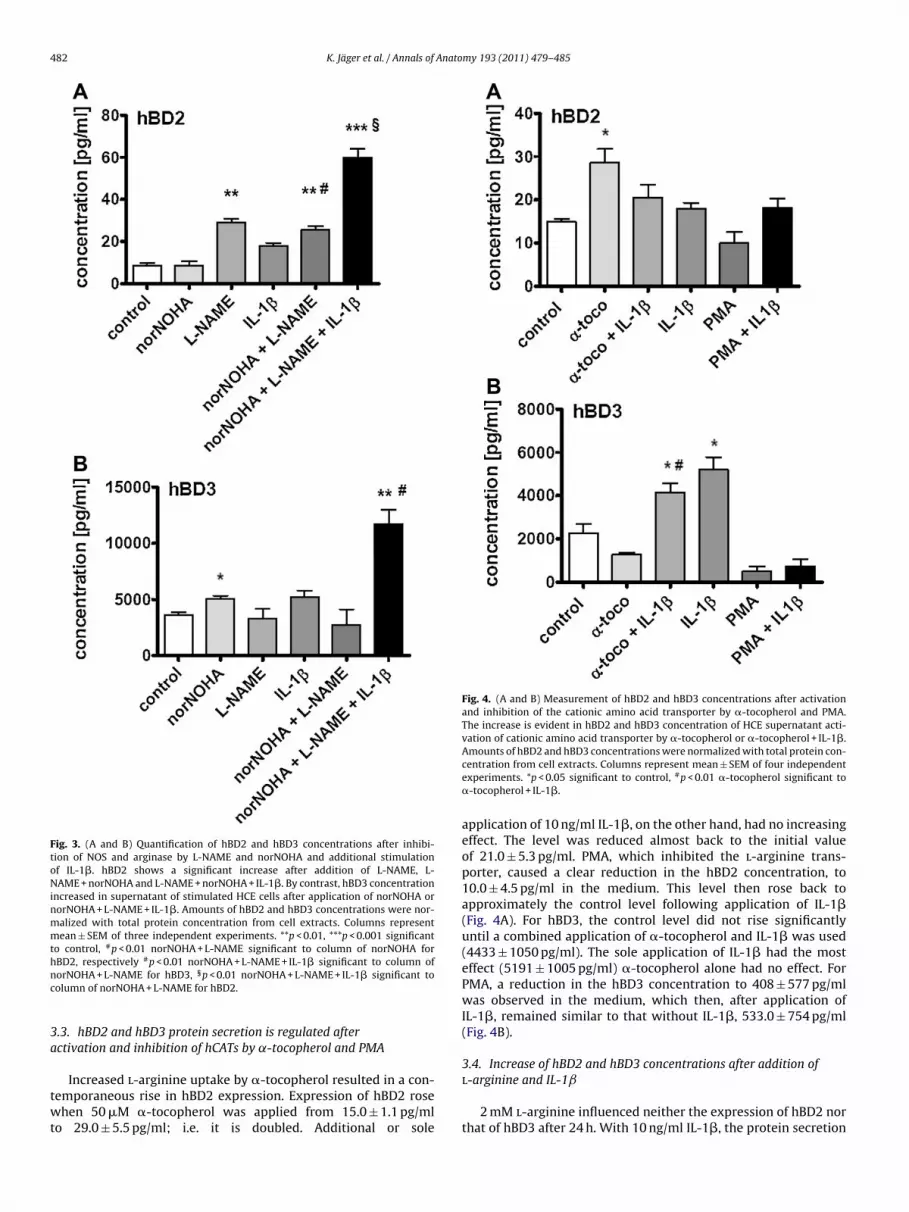
482 K. Jäger et al. / Annals of Anatomy 193 (2011) 479– 485
Fig. 3. (A and B) Quantification of hBD2 and hBD3 concentrations after inhibi-tion of NOS and arginase by L-NAME and norNOHA and additional stimulationof IL-1�. hBD2 shows a significant increase after addition of L-NAME, L-NAME + norNOHA and L-NAME + norNOHA + IL-1�. By contrast, hBD3 concentrationincreased in supernatant of stimulated HCE cells after application of norNOHA ornorNOHA + L-NAME + IL-1�. Amounts of hBD2 and hBD3 concentrations were nor-malized with total protein concentration from cell extracts. Columns representmean ± SEM of three independent experiments. **p < 0.01, ***p < 0.001 significantto control, #p < 0.01 norNOHA + L-NAME significant to column of norNOHA forhBD2, respectively #p < 0.01 norNOHA + L-NAME + IL-1� significant to column ofnc
3a
twt
Fig. 4. (A and B) Measurement of hBD2 and hBD3 concentrations after activationand inhibition of the cationic amino acid transporter by �-tocopherol and PMA.The increase is evident in hBD2 and hBD3 concentration of HCE supernatant acti-vation of cationic amino acid transporter by �-tocopherol or �-tocopherol + IL-1�.Amounts of hBD2 and hBD3 concentrations were normalized with total protein con-
l-arginine and IL-1ˇ
orNOHA + L-NAME for hBD3, §p < 0.01 norNOHA + L-NAME + IL-1� significant toolumn of norNOHA + L-NAME for hBD2.
.3. hBD2 and hBD3 protein secretion is regulated afterctivation and inhibition of hCATs by ˛-tocopherol and PMA
Increased l-arginine uptake by �-tocopherol resulted in a con-
emporaneous rise in hBD2 expression. Expression of hBD2 rosehen 50 �M �-tocopherol was applied from 15.0 ± 1.1 pg/mlo 29.0 ± 5.5 pg/ml; i.e. it is doubled. Additional or sole
centration from cell extracts. Columns represent mean ± SEM of four independentexperiments. *p < 0.05 significant to control, #p < 0.01 �-tocopherol significant to�-tocopherol + IL-1�.
application of 10 ng/ml IL-1�, on the other hand, had no increasingeffect. The level was reduced almost back to the initial valueof 21.0 ± 5.3 pg/ml. PMA, which inhibited the l-arginine trans-porter, caused a clear reduction in the hBD2 concentration, to10.0 ± 4.5 pg/ml in the medium. This level then rose back toapproximately the control level following application of IL-1�(Fig. 4A). For hBD3, the control level did not rise significantlyuntil a combined application of �-tocopherol and IL-1� was used(4433 ± 1050 pg/ml). The sole application of IL-1� had the mosteffect (5191 ± 1005 pg/ml) �-tocopherol alone had no effect. ForPMA, a reduction in the hBD3 concentration to 408 ± 577 pg/mlwas observed in the medium, which then, after application ofIL-1�, remained similar to that without IL-1�, 533.0 ± 754 pg/ml(Fig. 4B).
3.4. Increase of hBD2 and hBD3 concentrations after addition of
2 mM l-arginine influenced neither the expression of hBD2 northat of hBD3 after 24 h. With 10 ng/ml IL-1�, the protein secretion

K. Jäger et al. / Annals of Anato
Fig. 5. (A and B) Determination of hBD2 and hBD3 secretions after addition of l-arginine and IL-1�. An increase is shown for hBD2 after 24 h of stimulation with50 ng/ml IL-1� which is not boosted by additional l-arginine. Not until l-arginineand IL-1� are both added to HCE medium does the hBD3 concentration increase.Amounts of hBD2 and hBD3 concentrations were normalized with total protein con-ceL
otharfi9
4
cihtdrctlms
entration from cell extracts. Columns represent mean ± SEM of four independentxperiments. **p < 0.01, significant to control, #p < 0.05, L-Arg + IL-1� significant to-Arg and/or IL-1�.
f hBD2 could be raised from 4.2 ± 1.7 pg/ml to 18.0 ± 2.2 pg/ml,hus by a factor of 4.5 (Fig. 5A), but no effect was observed onBD3 (Fig. 5B). Following combined application of IL-1� and l-rginine, the hBD2 concentration increased, with protein secretionemaining at about 4 times the control level, as was observedor IL-1� alone (Fig. 5A). The hBD3 concentration rose follow-ng additional application of l-arginine from 4492.0 ± 288 pg/ml to155.0 ± 1065 pg/ml; i.e. to double the control value (Fig. 5B).
. Discussion
In the present study we addressed the question of whether ahanged supply of the cationic amino acids l-lysine and l-argininen cultured corneal epithelial cells, due to activity blockade of theCATs or a balance shift in the metabolic pathways competing forhe amino acid l-arginine, has an effect on hBD2 and/or hBD3 pro-uction. A recent study of the Kinoshita group showed that theelative composition of l-arginine (Arg) in the tear fluid was signifi-antly higher than in plasma and aqueous humor. They propose that
he steady-state tear-fluid amino acid profiles might reflect ocu-ar surface homeostasis and the observed changes of amino acidsight have a close relation to the disease conditions on the ocularurface (Nakatsukasa et al., 2011). Thus, this group also provides
my 193 (2011) 479– 485 483
evidence of the correlation of l-arginine and immune defence inocular surface.
As hBD1 has been shown to be only constitutively expressedon the ocular surface and some studies on ocular surface epithe-lia and cell lines have demonstrated that it is not upregulated byproinflammatory cytokines or bacterial products in these tissues(for review see Kolar and McDermott, 2011) we limited our studyto the two inducible �-defensins hBD2 and hBD3. During prepa-ration of this manuscript a study came out demonstrating thatapplication of l-arginine results in increased expression of hBD-1 in colon carcinoma cells (HCT-116; Sherman et al., 2006), whichincreases the availability of l-arginine; i.e. has a direct influence onthe expression of hBD1 and thus supports our hypothesis.
Previous results as well as our present data reveal with regardto the non-stimulated expression of hBD2 and hBD3 in the super-natant of cultured HCE cells that hBD3 originally shows a muchhigher level of baseline expression, varying between 125 and 400times that found for hBD2 (Fig. 2). Interestingly, the measured base-line concentration of hBD2 and hBD3 in healthy cornea is nearly thesame in relation to mg total protein (Garreis et al., 2010). Earlierinvestigations found that mRNA expression of hBD2 is variable;only occasionally being present in normal cornea (Haynes et al.,1999; McNamara et al., 1999; Lehmann et al., 2000; McDermottet al., 2003) and more frequently in inflamed ocular tissue, whereashBD3 is constitutively expressed in healthy cornea and culturedcorneal epithelial cells (McDermott et al., 2003; McIntosh et al.,2005). The cell proteins of the non stimulated cells were also inves-tigated in each case in our present study, revealing only barelymeasurable results for hBD2 and hBD3 (data not shown). For skinHDPs, studies have shown that defensins are localized to structurescalled lamellar bodies, which are lipid-containing vesicles secretedinto the extracellular space having the effect of raising the localconcentration of these antimicrobial peptides (Oren et al., 2003).As in vitro antimicrobial activity is rapid (15–90 min; McDermott,2009) and we could measure in a previous study significant hBD3peptide concentrations as early as 5 min after exposure to a bacte-rial supernatant (Garreis et al., 2010) we speculate that hBD2 andhBD3 are probably stored in HCE cells in an intracellular form (forexample a precursor) that is not detected by our ELISA experiments.
We were able to demonstrate, based on inhibition of the main l-arginine metabolic pathways with L-NAME and norNOHA, that notonly the enzymes arginase and nitrate monoxide synthase competefor the amino acid l-arginine in the HCE cells, but the enzymesthat mediate the constitutive expression of �-defensins as well.In addition, hBD2 and hBD3 are seemingly regulated in differentways in the HCE cells. The hBD2 protein secretion is clearly stim-ulated by the blockade of NOS by L-NAME. Additional applicationof norNOHA has no additive effect (Fig. 3A). These results contrastwith those obtained for hBD3 selection, which is enhanced aboveall when the arginase blocker norNOHA is blocked. On the otherhand, the blockade of NOS appears to have no influence (Fig. 3B).However, the cells react with a clearly increased secretion of both�-defensins to blockade of both metabolic pathways that com-pete for l-arginine and additional stimulation by IL-1�. In this case,secretion increases to 6 fold (hBD2) respectively 3 fold (hBD3) theinitial value (Fig. 3). The proinflammatory cytokine IL-1� alone hadno effect. First the combined inhibition of arginase and the NOSpathway reveals a significant effect on the secretion rate of hBD2and -3 (Fig. 3). This experiment demonstrates that all three ways(NO production by NOS; urea and ornithine production by arginase,and the �-defensin production, Fig. 1) compete for l-arginine undersimulated inflammatory conditions through IL-1�.
Differential regulation of �-defensin expression has also beenobserved in other organs such as human skin or gingiva (Yin et al.,2010; Sørensen et al., 2005). It is notable that increased hBD2and arginase expression has been described repeatedly in various

4 Anato
aetwimewAis
atpepdipimristoadap1sdbcAde(
otsttldlobmmpllfdvsT4re
t
84 K. Jäger et al. / Annals of
rticles in correlation with wound healing processes (McDermottt al., 2001; Dressel et al., 2010; Bruch-Gerharz et al., 2003). In con-rast, the expression of hBD3 shows no change in expression inound healing processes (Dressel et al., 2010). This suggests that an
ncreased arginase expression, for instance after an L-NAME treat-ent (Balkan et al., 1997), could function as a trigger for the hBD2
xpression. HBD3 and NOS, on the other hand, are increased mainlyhile an inflammatory reaction is maintained (Harder et al., 2001;lderton et al., 2001; Kleinert et al., 2004; Korhonen et al., 2005). It
s thus conceivable that the NOS stimulation also stimulates hBD3ecretion by way of the arginase blockade (Balkan et al., 1997).
Another way to influence the availability of l-lysine and l-rginine for �-defensin production is to intervene directly in theransport of the two amino acids into the cell (Fig. 1). The hCATroteins have phosphorylation sites for proteinkinase C (Varogat al., 2004; Albritton et al., 1989; Closs, 2002). When these phos-horylation sites are activated, the proteinkinase C (PKC) is able toecrease the uptake of l-arginine. The PKC itself can be activated or
nhibited. An important inhibitor is �-tocopherol, which preventshosphorylation of the CAT proteins by inhibiting PKC and thus
ncreases l-arginine uptake (Ricciarelli et al., 1998). PMA (phorbolyristate acetate), on the other hand, activates PKC, resulting in
educed l-arginine transport, but begins to degrade the proteink-nase C after 4 h (Krotova et al., 2003). The different effect of theubstance that increases l-arginine uptake on �-defensin secre-ion can certainly be explained by the differing baseline expressionf the two �-defensins (Fig. 2). �-Tocopherol only increases l-rginine uptake within a specific scope. l-Arginine is then initiallyistributed in the cell between arginase and NOS, which also ensurenabolic processes such as growth (Anderson and Dobson, 1959),rotein synthesis, and wound healing (Barbul et al., 1990; Kirk et al.,993) during a non-inflammatory phase. Since hBD2 shows a con-titutively weak expression, a slight excess of l-arginine suffices toouble expression (Fig. 4A). On the other hand, this is contradictedy sole application of 2 mM l-arginine (Fig. 5A), whereupon the HCEells show increased secretion of neither hBD2 nor hBD3 (Fig. 5).pparently, �-tocopherol influences hBD2 expression/secretionirectly. Some data reveal the influence of �-tocopherol on thexpression of inflammation-associated genes or their modulationParola et al., 1992; Azzi et al., 2004).
For hBD3, this influence is too weak to raise the expressionf the �-defensin by way of increased arginine uptake due to �-ocopherol. A cytokine-induced phlogiston process must also betimulated in this case but the effect is not as intense as comparedo IL-1� alone (Fig. 4). PMA, on the other hand, has no effect onhe constitutive expression of hBD2 since the baseline expressionevel is too low, so that the loss of a few l-arginine molecules is notecisive for hBD2. For hBD3, however, a clear effect is observed fol-
owing application of PMA. A direct influence of the hCAT proteinsn �-defensin production would also explain the results obtainedy Harder et al., 2004. After 16 h of incubation of cultured pri-ary skin keratinocytes with 50 ng/ml PMA, there was an obviousassive rise in hBD2 and hBD3 expression. After 16 h, PMA com-
letely degrades the PKC of the y+ transporter, so that it could noonger exert an inhibitory effect on l-arginine transport and more-arginine could enter the cell (Krotova et al., 2003) and be availableor �-defensin expression. These experiments demonstrate that �-efensin secretion and expression can be controlled and influencedia the y+ transporter. This was also demonstrated in the presenttudy by contemporaneous stimulation with l-arginine and IL-1�.he cells react to this with both increased secretion of hBD2 (raised-fold; Fig. 5A) and of hBD3 (raised 2-fold, Fig. 5B). Comparable
esults in colon carcinoma cells were also obtained by Shermant al. (2006) for hBD1 following application of l-arginine.In conclusion, these results demonstrate that �-defensin secre-ion and production are integrated in the regulatory cascade of
my 193 (2011) 479– 485
arginase and NOS. Synthesis of �-defensins also competes for theamino acid l-arginine, as do the enzymes arginase and nitro-gen monooxidase. l-Arginine and �-tocopherol exert a significantenhancing effect on hBD2 and hBD3 expression in the presence ofthe proinflammatory protein IL-1�. Also, L-NAME and norNOHA,especially when combined, greatly increase hBD production underexperimental inflammatory conditions in cultured HCE cells.
Although these results potentially open up new perspectives forthe treatment of inflammatory and immune responses not only onthe ocular surface they also raise several new questions and muchremains to be discovered about the connection.
Conflict of interest
The authors have no financial conflict of interest.
Acknowledgments
We thank Ute Beyer, Stephanie Beilecke and Susann Möschterfor excellent technical assistance. We also thank the groupof Prof. Renke Maas, Institute of Experimental and Clini-cal Pharmacology and Toxicology, Friedrich-Alexander-UniversityErlangen-Nuremberg, Germany for helpful discussions. This workwas supported by the Deutsche Forschungsgemeinschaft (DFG) –program grants PA 738/9-1 and PA 738/9-2, BMBF – Wilhelm RouxProgram, Halle, Germany – program grants FKZ 09/16, FKZ 14/25,and FKZ 16/35, as well as Sicca Forschungsförderung of the profes-sional Association of German Ophthalmologists.
References
Abedin, A., Mohammed, I., Hopkinson, A., Dua, H.S., 2008. A novel antimicrobialpeptide on the ocular surface shows decreased expression in inflammation andinfection. Invest. Ophthalmol. Vis. Sci. 49, 28–33.
Albritton, L.M., Tseng, L., Scadden, D., Cunningham, J.M., 1989. A putative murineecotropic retrovirus receptor gene encodes a multiple membrane-spanning pro-tein and confers susceptibility to virus infection. Cell 57, 659–666.
Alderton, W.K., Cooper, C.E., Knowles, R.G., 2001. Nitric oxide synthases: structure,function and inhibition. Biochem. J. 357, 593–615.
Anderson, J.O., Dobson, D.C., 1959. Amino acids requirements of the chick: effect oftotal essential amino acid level in the diet on the arginine and lysine require-ments. Poult. Sci. 38, 1140–1147.
Araki-Sasaki, K., Ohashi, Y., Sasabe, T., et al., 1995. An SV40-immortalized humancorneal epithelial cell line and its characterization. Invest. Ophthalmol. Vis. Sci.36, 614–621.
Azzi, A., Gysin, R., Kempná, P., Munteanu, A., Negis, Y., Villacorta, L., Visarius, T., Zingg,J.M., 2004. Vitamin E mediates cell signaling and regulation of gene expression.Ann. N.Y. Acad. Sci. 1031, 86–95.
Balkan, E., Balkan, S., Ozben, T., Serteser, M., Gümüslü, S., Oguz, N., 1997. The effects ofnitric oxide synthase inhibitor, L-NAME on NO production during focal cerebralischemia in rats: could L-NAME be the future treatment of sudden deafness? Int.J. Neurosci. 89, 61–77.
Barbul, A., Lazarou, S.A., Efron, D.T., Wasserkrug, H.L., Efron, G., 1990. Arginineenhances wound healing and lymphocyte immune responses in humans.Surgery 108, 331–337.
Baum, J., 1995. Infection of the eye. Clin. Infect. Dis. 21, 479–488.Bruch-Gerharz, D., Schnorr, O., Suschek, C., Beck, K.F., Pfeilschifter, J., Ruzicka, T.,
Kolb-Bachofen, V., 2003. Arginase 1 overexpression in psoriasis: limitation ofinducible nitric oxide synthase activity as a molecular mechanism for ker-atinocyte hyperproliferation. Am. J. Pathol. 162, 203–211.
Closs, E.I., 2002. Expression, regulation and function of carrier proteins for cationicamino acids. Curr. Opin. Nephrol. Hypertens. 11, 99–107.
Deves, R., Boyd, C.A.R., 1998. Transporters for cationic amino acids in animal cells:discovery, structure, and function. Physiol. Rev. 78, 478–545.
Dressel, S., Harder, J., Cordes, J., Wittersheim, M., Meyer-Hoffert, U., Sunderkötter,C., Gläser, R., 2010. Differential expression of antimicrobial peptides in marginsof chronic wounds. Exp. Dermatol. 19, 628–632.
Feelisch, M., Stammler, S.J., 1996. Methods in Nitric Oxide Research. Wiley Inc.,Chichester.
Fleisig, S.M.J., Efron, N., Pier, G.B., 1992. Extended contact lens wear enhancesPseudomonas aeruginosa adherence to human corneal epithelium. Invest. Oph-
thalmol. Vis. Sci. 33, 2908–2916.Garreis, F., Schlorf, T., Worlitzsch, D., Steven, P., Bräuer, L., Jäger, K., Paulsen, F.P., 2010.Roles of human beta-defensins in innate immune defence at the ocular surface:arming and alarming corneal and conjunctival epithelial cells. Histochem. CellBiol. 134, 59–73.

Anato
G
H
H
H
H
H
H
H
J
J
J
J
K
K
K
K
K
K
K
L
L
M
K. Jäger et al. / Annals of
ottsch, J.D., Li, Q., Ashraf, M.F., O’Brien, T.P., Stark, W.J., Liu, S.H., 1998. Defensingene expression in the cornea. Curr. Eye Res. 17, 1082–1086.
arder, J., Bartels, J., Christophers, E., Schroder, J.M., 2001. Isolation and characteri-zation of human beta -defensin-3, a novel human inducible peptide antibiotic.J. Biol. Chem. 276, 5707–5713.
arder, J., Meyer-Hoffert, U., Wehkamp, K., Schwichtenberg, L., Schröder, J.M., 2004.Differential gene induction of human beta-defensins (hBD-1, -2, -3, and -4) inkeratinocytes is inhibited by retinoic acid. J. Invest. Dermatol. 123, 522–529.
attenbach, L.O., Gumbel, H., Kippenberger, S., 1998. Identification of beta-defensinsin human conjunctiva. Antimicrob. Agents Chemother. 42, 3332.
aynes, R.J., Tighe, P.J., Dua, H.S., 1999. Antimicrobial defensin peptides of the humanocular surface. Br. J. Ophthalmol. 83, 737–741.
aynes, R.J., Tighe, P.J., Dua, H.S., 1998. Innate defence of the eye by antimicrobialdefensin peptides. Lancet 352, 451–452.
oshide, R., Ikeda, Y., Karamshima, S., et al., 1996. Molecular cloning, tissue distribu-tion; and chromosomal localization of human cationic amino acid transporter2(HCAT2). Genomics 38, 174–178.
uang, L.C., Jean, D., Proske, R.J., Reins, R.Y., McDermott, A.M., 2007. Ocular sur-face expression and in vitro activity of antimicrobial peptides. Curr. Eye Res. 32,595–609.
äger, K., Boenisch, U., Risch, M., Worlitzsch, D., Paulsen, F.P., 2009. Detection ofcationic amino acid transporters in the human lacrimal system and their involve-ment in ocular surface disease. Invest. Ophthalmol. Vis. Sci. 50, 1112–1121.
äger, K., Garreis, F., Dunse, M., Paulsen, F.P., 2010a. Cationic amino acid transportersand beta-defensins in dry eye syndrome. Dev. Ophthalmol. 45, 12–15.
äger, K., Garreis, F., Posa, A., Dunse, M., Paulsen, F.P., 2010b. Functional relationshipbetween cationic amino acid transporters and beta-defensins: implications fordry skin diseases and the dry eye. Ann. Anat. 192, 65–69.
enkinson, C.P., Grigor, M.R., 1994. Rat mammary arginase: isolation and character-ization. Biochem. Med. Metab. Biol. 51, 156–165.
irk, S.J., Hurson, M., Regan, M.C., Holt, D.R., Wasserkrug, H.L., Barbul, A., 1993. Argi-nine stimulates wound healing and immune function in elderly human beings.Surgery 114, 155–160.
leinert, H., Pautz, A., Linker, K., Schwarz, P.M., 2004. Regulation of the expressionof inducible nitric oxide synthase. Eur. J. Pharmacol. 500, 255–266.
olar, S.S., McDermott, A.M., 2011. Role of host-defence peptides in eye diseases.Cell. Mol. Life Sci. 68, 2201–2213.
orhonen, R., Lahti, A., Kankaanranta, H., Moilanen, E., 2005. Nitric oxide productionand signaling in inflammation. Curr. Drug Targets Inflamm. Allergy 4, 471–474.
rotova, K.Y., Zharikov, S.I., Block, E.R., 2003. Classical isoforms of PKC as regulators ofCAT-1 transporter activity in pulmonary artery endothelial cells. Am. J. Physiol.Lung Cell Mol. Physiol. 284, 1037–1044.
umar, A., Yin, J., Zhang, J., Yu, F-S.X., 2007. Modulation of corneal epithelial innateimmune response to Pseudomonas infection by flagellin pretreatment. Invest.Ophthalmol. Vis. Sci. 48, 4664–4670.
umar, A., Zhang, J., Yu, F.S., 2006. Toll-like receptor 2-mediated expression of beta-defensin 2 in human corneal epithelial cells. Microbes Infect. 8, 380–389.
ehmann, O.J., Hussain, I.R., Watt, P.J., 2000. Investigation of �-defensin geneexpression in the ocular anterior segment by semiquantitative RT-PCR. Br. J.Ophthalmol. 84, 523–526.
i, Q., Kumar, A., Gui, J.F., Yu, F.S., 2008. Staphylococcus aureus lipoproteins triggerhuman corneal epithelial innate response through toll-like receptor-2. Microb.
Pathog. 44, 426–434.cDermott, A.M., Redfern, R.L., Zhang, B., Pei, Y., Huang, L., Proske, R.J., 2003.Defensin expression by the cornea: multiple signaling pathways mediate IL-1beta stimulation of hBD-2 expression by corneal epithelial cells. Invest. Oph-thalmol. Vis. Sci. 44, 1856–1859.
my 193 (2011) 479– 485 485
McDermott, A.M., Redfern, R.L., Zhang, B., 2001. Human beta-defensin 2 is up-regulated during re-epithelialization of the cornea. Curr. Eye Res. 22, 64–67.
McDermott, A.M., 2009. The role of antimicrobial peptides at the ocular surface.Ophthalmic Res. 41, 60–75.
McIntosh, R.S., Cade, J.E., Al-Abed, M., et al., 2005. The spectrum of antimicrobial pep-tide expression at the ocular surface. Invest. Ophthalmol. Vis. Sci. 46, 1379–1385.
McNamara, N.A., Van, R., Tuchin, O.S., Fleiszig, S.M.J., 1999. Ocular surface epitheliaexpress mRNA for human beta defensin-2. Exp. Eye Res. 69, 483–490.
Mohammed, I., Suleman, H., Otri, A., Kulkarni, B.B., Chen, P., Hopkinson, A., Dua, H.S.,2010. Localization and gene expression of human �-defensin 9 at the humanocular surface epithelium. Invest. Ophthalmol. Vis. Sci. 51, 4677–4682.
Nakatsukasa, M., Sotozono, C., Shimbo, K., Ono, N., Miyano, H., Okano, A., Hamuro, J.,Kinoshita, S., 2011. Amino acid profiles in human tear fluids analyzed by high-performance liquid chromatography and electrospray ionization tandem massspectrometry. Am. J. Ophthalmol. 151 (5), 799–808.
Narayanan, S., Miller, W.L., McDermott, A.M., 2003. Expression of human beta-defensins in conjunctival epithelium: relevance to dry eye disease. Invest.Ophthalmol. Vis. Sci. 44, 3795–3801.
O’Callaghan, R.J., Cellegan, M.C., Moreau, J.M., et al., 1997. Specific roles of alpha-toxin and beta-toxin during Staphylococcus aureus corneal infection. Infect.Immun. 65, 1571–1578.
Oren, A., Ganz, T., Liu, L., Meerloo, T., 2003. In human epidermis, beta-defensin 2 ispackaged in lamellar bodies. Exp. Mol. Pathol. 74, 180–182.
Parola, M., Muraca, R., Dianzani, I., et al., 1992. Vitamin E dietary supplementationinhibits transforming growth factor beta 1 gene expression in the rat liver. FEBSLett. 308, 267–270.
Ricciarelli, R., Tasinato, A., Clément, S., Ozer, N.K., Boscoboinik, D., Azzi, A., 1998.alpha-Tocopherol specifically inactivates cellular protein kinase C alpha bychanging its phosphorylation state. Biochem. J. 334, 243–249.
Schroder, J.M., 1999. Clinical significance of epithelial peptide antibiotics. BioDrugs11, 293–300.
Sherman, H., Chapnik, N., Froy, O., 2006. Albumin and amino acids upregulate theexpression of human beta-defensin 1. Mol. Immunol. 43, 1617–1623.
Shin, J.S., Kim, C.W., Kwon, Y.S., Kim, J.C., 2004. Human beta-defensin 2 is inducedby interleukin-1 beta in the corneal epithelial cells. Exp. Mol. Med. 36, 204–210.
Sørensen, O.E., Thapa, D.R., Rosenthal, A., Liu, L., Roberts, A.A., Ganz, T., 2005. Differ-ential regulation of beta-defensin expression in human skin by microbial stimuli.J. Immunol. 174, 4870–4879.
Tate Jr., D.J., Vonderhaar, D.J., Caldas, Y.A., et al., 2008. Effect of arginase II on l-arginine depletion and cell growth in murine cell lines of renal cell carcinoma.J. Hematol. Oncol. 25, 1–14.
Tenu, J.P., Lepoivre, M., Moali, C., Brollo, M., Mansuy, D., Boucher, J.L., 1999. Effects ofthe new arginase inhibitor N(omega)-hydroxy-nor-l-arginine on NO synthaseactivity in murine macrophages. Nitric Oxide 3, 427–438.
Varoga, D., Pufe, T., Harder, J., Meyer-Hoffert, U., Mentlein, R., Schröder, J.M., Petersen,W.J., Tillmann, B.N., Proksch, E., Goldring, M.B., Paulsen, F.P., 2004. Production ofendogenous antibiotics in articular cartilage. Arthritis Rheum. 50, 3526–3534.
Vekony, N., Wolf, S., Boissel, J.P., Gnauert, K., Closs, E.I., 2001. Human cationic aminoacid transporter hCAT-3 is preferentially expressed in peripheral tissue. Bio-chemistry 40, 12387–12394.
Yin, L., Chino, T., Horst, O.V., Hacker, B.M., Clark, E.A., Dale, B.A., Chung, W.O., 2010.Differential and coordinated expression of defensins and cytokines by gingival
epithelial cells and dendritic cells in response to oral bacteria. BMC Immunol.11, 37.Zou, G., de Leeuw, E., Li, C., et al., 2007. Toward understanding the cationicity ofdefensins. (2007) Arg and Lys versus their noncoded analogs. J. Biol. Chem. 282,19653–19665.
Related Documents











