Brief communication In vitro assessment of the neurotoxic and neuroprotective effects of N-acetyl-L-cysteine (NAC) on the rat sciatic nerve fibers Magdalini Moschou a , Efstratios K. Kosmidis a , Martha Kaloyianni a , A. Geronikaki b , N. Dabarakis c , George Theophilidis a, * a Laboratory of Animal Physiology, Department of Zoology, School of Biology, Aristotle University, Thessaloniki 54124, Hellas, Greece b School of Pharmacy, Department of Pharmaceutical Chemistry, Aristotle University, Thessaloniki 54124, Hellas, Greece c Department of Oral Surgery, Implantology and Roentgenology, Aristotle University, Thessaloniki 54124, Hellas, Greece Received 4 May 2007; accepted 7 September 2007 Available online 15 September 2007 Abstract N-acetyl-L-cysteine (NAC), at a concentration of 1–60 mM, has been previously used extensively for protection in a variety of cell cultures against the deleterious effects of various compounds. The results of this in vitro study show that NAC has certain unusual effects on the evoked compound action potential (CAP) of the rat sciatic nerve fibers. Firstly, at concentrations of 5.0, 3.5 and 2.5 mM, con- centrations used by others as a protectant for cell cultures, NAC inhibits the action potentials of the sciatic nerve fibers completely in a concentration-dependent manner within a few minutes or hours (2.5 mM). Secondly, the acute inhibitory action of NAC on the CAP of the nerve fibers was not spontaneously reversible, but as soon as NAC was replaced with saline there was a partial (75%) recovery in the function of the nerve fibers. Thirdly, the no observed effect concentration for NAC was estimated to be 1 mM. The paradox is that NAC at 1 mM not only had no effect on the nerve fibers, but it became an excellent neuroprotective compound, giving almost 100% neuro- protection against cadmium-induced neurotoxicity. The results show a possible effect of NAC on voltage-gated sodium and potassium channels. The observed neuroprotective-neurotoxic properties of NAC require careful reconsideration of its use in either in vitro studies or in vivo pharmaceutical applications. Ó 2007 Elsevier Ltd. All rights reserved. Keywords: N-acetyl-L-cysteine (NAC); Neurotoxicity; Neuroprotection; Rat sciatic nerve; In vitro; Compound action potential (CAP) 1. Introduction The antioxidant and free-radical scavenger N-acetyl-L- cysteine (NAC) is used extensively as a conditional nutri- ent. NAC acts as a cysteine donor and maintains or even increases the intracellular levels of glutathione, a tripeptide which protects cells from toxins such as free-radicals. It is currently the dietary supplement of choice for building up or conserving the body’s stores of glutathione, cysteine, and other sulfhydryl antioxidant resources. It has also been shown to have antiviral effects in patients with HIV due to inhibition of viral stimulation by reactive oxygen interme- diates. NAC is used in a broad range of concentrations, varying from 1 to 60 mM in in vitro studies. At these con- centrations it has been found to inhibit or delay certain types of cell death induced by, for example, TNF-a, (Mayer and Noble, 1994; Cossarizza et al., 1995; Schulze- Osthoff et al., 1994), ethanol (Higuchi et al., 2001), antican- cer drugs (Wang et al., 2000), environmental pollutants (Munday, 1989; Oh and Lima, 2006) active substances (Mohamadin and Abdel-Naim, 1999; Atkins et al., 2000) and heavy metals (Seaton et al., 1997; Fern et al., 1996; Carageorgiou et al., 2004; Oh and Lima, 2006). However, there are some reservations about the protec- tive properties of NAC since several researchers have reported unusual toxicity. For example, inhibitory effects 0887-2333/$ - see front matter Ó 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.tiv.2007.09.005 * Corresponding author. Tel.: +30 2310 998261, 998275; fax: +30 2310 998269. E-mail address: [email protected] (G. Theophilidis). www.elsevier.com/locate/toxinvit Available online at www.sciencedirect.com Toxicology in Vitro 22 (2008) 267–274

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Available online at www.sciencedirect.com
www.elsevier.com/locate/toxinvit
Toxicology in Vitro 22 (2008) 267–274
Brief communication
In vitro assessment of the neurotoxic and neuroprotective effectsof N-acetyl-L-cysteine (NAC) on the rat sciatic nerve fibers
Magdalini Moschou a, Efstratios K. Kosmidis a, Martha Kaloyianni a, A. Geronikaki b,N. Dabarakis c, George Theophilidis a,*
a Laboratory of Animal Physiology, Department of Zoology, School of Biology, Aristotle University, Thessaloniki 54124, Hellas, Greeceb School of Pharmacy, Department of Pharmaceutical Chemistry, Aristotle University, Thessaloniki 54124, Hellas, Greecec Department of Oral Surgery, Implantology and Roentgenology, Aristotle University, Thessaloniki 54124, Hellas, Greece
Received 4 May 2007; accepted 7 September 2007Available online 15 September 2007
Abstract
N-acetyl-L-cysteine (NAC), at a concentration of 1–60 mM, has been previously used extensively for protection in a variety of cellcultures against the deleterious effects of various compounds. The results of this in vitro study show that NAC has certain unusual effectson the evoked compound action potential (CAP) of the rat sciatic nerve fibers. Firstly, at concentrations of 5.0, 3.5 and 2.5 mM, con-centrations used by others as a protectant for cell cultures, NAC inhibits the action potentials of the sciatic nerve fibers completely in aconcentration-dependent manner within a few minutes or hours (2.5 mM). Secondly, the acute inhibitory action of NAC on the CAP ofthe nerve fibers was not spontaneously reversible, but as soon as NAC was replaced with saline there was a partial (�75%) recovery in thefunction of the nerve fibers. Thirdly, the no observed effect concentration for NAC was estimated to be 1 mM. The paradox is that NACat 1 mM not only had no effect on the nerve fibers, but it became an excellent neuroprotective compound, giving almost 100% neuro-protection against cadmium-induced neurotoxicity. The results show a possible effect of NAC on voltage-gated sodium and potassiumchannels. The observed neuroprotective-neurotoxic properties of NAC require careful reconsideration of its use in either in vitro studiesor in vivo pharmaceutical applications.� 2007 Elsevier Ltd. All rights reserved.
Keywords: N-acetyl-L-cysteine (NAC); Neurotoxicity; Neuroprotection; Rat sciatic nerve; In vitro; Compound action potential (CAP)
1. Introduction
The antioxidant and free-radical scavenger N-acetyl-L-cysteine (NAC) is used extensively as a conditional nutri-ent. NAC acts as a cysteine donor and maintains or evenincreases the intracellular levels of glutathione, a tripeptidewhich protects cells from toxins such as free-radicals. It iscurrently the dietary supplement of choice for buildingup or conserving the body’s stores of glutathione, cysteine,and other sulfhydryl antioxidant resources. It has also beenshown to have antiviral effects in patients with HIV due to
0887-2333/$ - see front matter � 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.tiv.2007.09.005
* Corresponding author. Tel.: +30 2310 998261, 998275; fax: +30 2310998269.
E-mail address: [email protected] (G. Theophilidis).
inhibition of viral stimulation by reactive oxygen interme-diates. NAC is used in a broad range of concentrations,varying from 1 to 60 mM in in vitro studies. At these con-centrations it has been found to inhibit or delay certaintypes of cell death induced by, for example, TNF-a,(Mayer and Noble, 1994; Cossarizza et al., 1995; Schulze-Osthoff et al., 1994), ethanol (Higuchi et al., 2001), antican-cer drugs (Wang et al., 2000), environmental pollutants(Munday, 1989; Oh and Lima, 2006) active substances(Mohamadin and Abdel-Naim, 1999; Atkins et al., 2000)and heavy metals (Seaton et al., 1997; Fern et al., 1996;Carageorgiou et al., 2004; Oh and Lima, 2006).
However, there are some reservations about the protec-tive properties of NAC since several researchers havereported unusual toxicity. For example, inhibitory effects

Fig. 1. (A) Overlapping records of the evoked compound actionpotentials (CAPs) from the isolated sciatic nerve of the rat in the three-chamber recording bath. The amplitude of the CAP increased inincrements (see thin lines) according to stimulus intensity. When thenerve was stimulated with supramaximal stimuli, all the axons wereactivated, the recorded CAP (see thick line) had the maximum value,indicated by the vertical scale bar. The dotted lines indicate the duration ofthe CAP. a–d: different phases of the CAP as explained in the text. Verticalscale bar: 5 mV. (B) Samples of the evoked CAP of the sciatic nerve fibers,defined between the sets of parallel lines, obtained every 1 h. The nervefibers were incubated in normal saline. The vertical line indicates theamplitude of the CAP (5 mV) while the horizontal line shows the durationof the CAP. (C) as in B, but the nerve was incubated in saline with 3.5 mMof NAC. (D) as in C but the NAC was replaced with normal saline at thetime indicated by the arrow (after 5 h).
268 M. Moschou et al. / Toxicology in Vitro 22 (2008) 267–274
on Na+ absorption in human nasal epithelia were observedat a concentration near 10 mM NAC (Rochat et al., 2004).Also, NAC, at a concentration greater than or equal to30 mM, was toxic, causing a prompt irreversible decreasein maximal tetanic stress on diaphragm muscles in vitro
(Khawli and Reid, 1994). Finally, 5–10 mM NAC causeddose-dependent reductions in viability of rat and humanaortic smooth muscle cells (Tsai et al., 1996). Due to theseunusual protective-toxic properties of NAC, further studiesare required to establish in detail the possible side effectswhich NAC may have on other tissues, like the nervoussystem, before further use of this compound in anyin vitro or in vivo study or in pharmaceutical applications.
The purpose of this study was first to assess in vitro theeffects of different concentrations of NAC on the actionpotentials of sciatic nerve fibers of the rat, and then to testthe possible neuroprotective properties of this compound.The sciatic nerve preparation used in this study is a reliablein vitro method which has been used successfully for theassessment of the neurotoxic effects of a number of pyre-throid insecticides (Theophilidis et al., 1997), various herbi-cides (Papaefthimiou et al., 2004), plasma ultrafiltrate fromuraemic patients (Cantaro et al., 1998), dental impressionpastes (Andreou et al., 2007) and the action of local anaes-thetics (Geronikaki et al., 2003).
2. Materials and methods
Male Wistar rats weighing between 220 and 280 g weresacrificed by intraperitoneal injection of sodium pentabar-bitone (100 mg/kg bodyweight, i.p.). All experimental pro-cedures were conducted in accordance with the protocolsoutlined by Aristotle University of Thessaloniki, Greece,regarding the recommended standard practices for Biolog-ical Investigations. The sciatic nerve of the rat, which con-sists of about 7500 nerve fibers (Schmalbruch, 1986), wasrapidly dissected from the spinal cord to the knee andimmersed in a standard oxygenated (O2 100%) saline solu-tion of the following composition (in mM): 140 NaCl, 11glucose, 2.7 KCl, 2.4 CaCl2, 1.1 MgCl2, 1 NaHCO3, 10HEPES, pH 7.2. All experiments were performed at a con-stant temperature of 26� ± 1 �C. The nerve was placed in aPetri dish and the perineurium was removed under a dis-secting stereoscope. Then, the nerve was mounted acrossa three-chambered recording bath, made of Plexiglas. Therecording system has been described elsewhere and it hasbeen used in a variety of in vitro neurotoxicological studies(e.g. Papaefthimiou et al., 2004; Andreou et al., 2007), buta short description will be given below. The recording bathconsists of three-chambers. The first is the recording cham-ber, where the active electrode of an A.C. differentialamplifier (Neurolog, NL822, Digitimer Ltd., UK) wereplaced. The second is the perfusion chamber where 70%of the 10 cm long nerve was incubated and the groundsof the recording and stimulating electrodes were immersed.The third is the stimulating chamber where the proximalpart of the nerve and the active stimulating electrode con-
nected to a constant voltage stimulator (Digitimer, DS9AEngland, UK) were immersed. The three-chambers werefilled with saline to cover the nerve. All electrodes forrecording stimulating and grounding, which were incontact with the saline, were made of 24 carat gold. Thedimensions of each chamber were 24 mm length, 36 mmwidth and 10 mm depth, with a total capacity of10–12 ml.
As an indication of the proper physiological functioningof the sciatic nerve fibers, their evoked compound actionpotentials (CAP) were recorded (Fig. 1A). The CAPs wereevoked by the stimulating unit delivering to the stimulatingelectrodes constant voltage stimuli of 2–3 V with a dura-tion of 0.01 ms at a frequency of 1 Hz. The propagatedCAPs were measured as the potential difference betweenthe recording and the perfusion chamber (in mV). The sig-nal of the CAPs was displayed using a digital oscilloscope(Hameg 205) and were digitized (200 kHz) continuously,while every 15 min data were stored in a computer, using
M. Moschou et al. / Toxicology in Vitro 22 (2008) 267–274 269
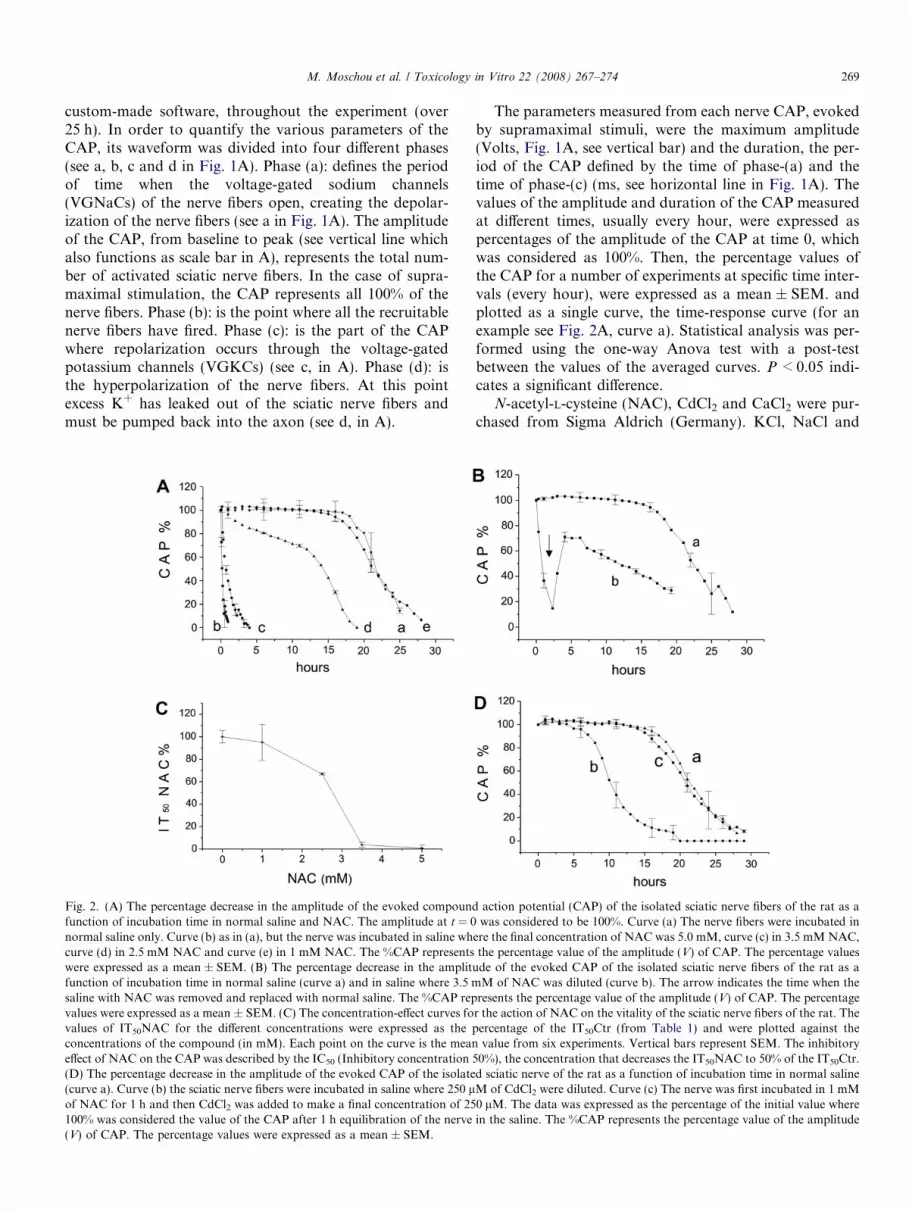
custom-made software, throughout the experiment (over25 h). In order to quantify the various parameters of theCAP, its waveform was divided into four different phases(see a, b, c and d in Fig. 1A). Phase (a): defines the periodof time when the voltage-gated sodium channels(VGNaCs) of the nerve fibers open, creating the depolar-ization of the nerve fibers (see a in Fig. 1A). The amplitudeof the CAP, from baseline to peak (see vertical line whichalso functions as scale bar in A), represents the total num-ber of activated sciatic nerve fibers. In the case of supra-maximal stimulation, the CAP represents all 100% of thenerve fibers. Phase (b): is the point where all the recruitablenerve fibers have fired. Phase (c): is the part of the CAPwhere repolarization occurs through the voltage-gatedpotassium channels (VGKCs) (see c, in A). Phase (d): isthe hyperpolarization of the nerve fibers. At this pointexcess K+ has leaked out of the sciatic nerve fibers andmust be pumped back into the axon (see d, in A).
Fig. 2. (A) The percentage decrease in the amplitude of the evoked compoundfunction of incubation time in normal saline and NAC. The amplitude at t = 0normal saline only. Curve (b) as in (a), but the nerve was incubated in saline whcurve (d) in 2.5 mM NAC and curve (e) in 1 mM NAC. The %CAP representswere expressed as a mean ± SEM. (B) The percentage decrease in the amplitfunction of incubation time in normal saline (curve a) and in saline where 3.5 msaline with NAC was removed and replaced with normal saline. The %CAP repvalues were expressed as a mean ± SEM. (C) The concentration-effect curves fovalues of IT50NAC for the different concentrations were expressed as the pconcentrations of the compound (in mM). Each point on the curve is the meaneffect of NAC on the CAP was described by the IC50 (Inhibitory concentration(D) The percentage decrease in the amplitude of the evoked CAP of the isolate(curve a). Curve (b) the sciatic nerve fibers were incubated in saline where 250 lof NAC for 1 h and then CdCl2 was added to make a final concentration of 25100% was considered the value of the CAP after 1 h equilibration of the nerve(V) of CAP. The percentage values were expressed as a mean ± SEM.
The parameters measured from each nerve CAP, evokedby supramaximal stimuli, were the maximum amplitude(Volts, Fig. 1A, see vertical bar) and the duration, the per-iod of the CAP defined by the time of phase-(a) and thetime of phase-(c) (ms, see horizontal line in Fig. 1A). Thevalues of the amplitude and duration of the CAP measuredat different times, usually every hour, were expressed aspercentages of the amplitude of the CAP at time 0, whichwas considered as 100%. Then, the percentage values ofthe CAP for a number of experiments at specific time inter-vals (every hour), were expressed as a mean ± SEM. andplotted as a single curve, the time-response curve (for anexample see Fig. 2A, curve a). Statistical analysis was per-formed using the one-way Anova test with a post-testbetween the values of the averaged curves. P < 0.05 indi-cates a significant difference.
N-acetyl-L-cysteine (NAC), CdCl2 and CaCl2 were pur-chased from Sigma Aldrich (Germany). KCl, NaCl and
action potential (CAP) of the isolated sciatic nerve fibers of the rat as awas considered to be 100%. Curve (a) The nerve fibers were incubated in
ere the final concentration of NAC was 5.0 mM, curve (c) in 3.5 mM NAC,the percentage value of the amplitude (V) of CAP. The percentage values
ude of the evoked CAP of the isolated sciatic nerve fibers of the rat as aM of NAC was diluted (curve b). The arrow indicates the time when the
resents the percentage value of the amplitude (V) of CAP. The percentager the action of NAC on the vitality of the sciatic nerve fibers of the rat. Theercentage of the IT50Ctr (from Table 1) and were plotted against thevalue from six experiments. Vertical bars represent SEM. The inhibitory
50%), the concentration that decreases the IT50NAC to 50% of the IT50Ctr.d sciatic nerve of the rat as a function of incubation time in normal salineM of CdCl2 were diluted. Curve (c) The nerve was first incubated in 1 mM0 lM. The data was expressed as the percentage of the initial value wherein the saline. The %CAP represents the percentage value of the amplitude

Table 1The effects of different concentrations of NAC tested on the evoked CAPof the isolated sciatic nerve fibers of the rat
Concentration NAC (mM) IT50 NAC (h) IT50 NAC% of IT50Ctr
0.0 21.94 ± 0.78 100.00.05 22.61 ± 0.84 103.05 ± 3.831.0 21.61 ± 0.86 98.47 ± 3.922.5 15.28 ± 0.04 69.77 ± 0.18**
3.5 0.785 ± 0.06 3.57 ± 0.27***
5.0 0.1815 ± 0.017 0.82 ± 0.08***
IT50 NAC represents the time (h) required for different concentrations ofNAC to decrease the amplitude of the CAP to 50% of its initial value(before the application of the compound). At 0.0 mM NAC, IT50NACequals IT50Ctr. Each value is the mean and the SEM of six experiments,eight experiments for the control. Also, the IT50 NAC was expressed as thepercentage of the IT50Ctr which was considered as 100%. The percentagevalues for each concentration of NAC were compared with the controlusing the Student’s t-test. *P < 0.05; **P < 0.01; ***P < 0.001.
270 M. Moschou et al. / Toxicology in Vitro 22 (2008) 267–274
MgCl2 from Panreac (Spain), HEPES from BiochemicaFluka (Switzerland), glucose from Riedel-de Haen (Ger-many) and NaHCO3 from Merck (Germany).
3. Results
3.1. The effects of NAC on the amplitude of the CAP
In this study, the effects of different concentrations ofNAC on the electrophysiological properties of the isolatedrat sciatic nerve fibers in saline were investigated. The CAPrecorded from the distal end of the sciatic nerve is a gradedresponse whose amplitude is proportional to the intensityof stimulation applied at its proximal end (see thin linesin Fig. 1A). The CAP was measured as the summed electri-cal response of all the excited nerve fibers of the sciaticnerve, since the stimulus intensity was supramaximal (seeCAP with the thick line in Fig. 1A). The amplitude ofthe CAP reflects the number of synchronously stimulatednerve fibers; thus, it can be used as an index of neuronalviability.
The main advantage of the three-chambered bath is thatunder normal recording conditions the shape of the evokedmaximum CAP remains constant for over 10 h (Fig. 1B).After that, there is a gradual decrease in the amplitudedue to gradual nerve fiber inactivation (data not shown).However, when the nerve was incubated in NAC, forexample, at a concentration of 3.5 mM, the CAP decreasedmuch faster (Fig. 1C, n = 5), although the nerve was stim-ulated with supramaximal stimuli, an indication that anumber of nerve fibers were inactivated. When the sciaticnerve was incubated continuously in 3.5 mM NAC therewas an acute inhibitory effects on the nerve fibers and theCAP was completely eliminated within 2–3 h. This effectwas not spontaneously reversible since the CAP neverrecovered (Fig. 1C). However, in similar experiments whenthe CAP was inhibited and NAC was removed immediately(the nerve was washed with normal saline and left in sal-ine), there was a gradual, but not complete, recovery toalmost 75% of the initial CAP (see arrow in Fig. 1D).
The effects of NAC on the vitality of sciatic nerve fibers,expressed as a decrease in the amplitude of the CAP, weredemonstrated more accurately using the time-responsecurves for the nerves exposed to different concentrationsof 5.0 mM, 3.5 mM, 2.5 mM, and 1.0 mM (Fig. 2 A, curvesb, c, d and e), compared to nerves exposed to normal saline(see curve a). A lower concentration of NAC of 0.05 mMwas also tested, but the time-response curve was identicalto the control curves in nerves exposed to saline (datanot shown). The unpaired Student’s t-test was used to com-pare the values between the time-response curves from thenerves treated with different concentrations of NAC andthe nerves treated with normal saline only. The values ofthe time-response curves of the nerves treated with5.0 mM, 3.5 mM and 2.5 mM were found to be signifi-cantly different when compared to controls (P < 0.01).Only the values of the curve (e) (NAC 1 mM) were found
to have no significant difference when compared to the con-trol (P > 0.05).
In order to simplify the presentation of the results con-cerned with the amplitude of the CAP, another parameter,the time required for the CAP to reach 50% of the value att = 0 h, was introduced. This parameter called here Inhibi-tion Time 50%, is represented in the text as IT50 and is mea-sured in hours. For the nerves kept in control conditions(normal saline) the IT50 was called IT50Ctr. From curvea, shown in Fig. 2A, the IT50Ctr was estimated to be21.94 ± 0.78 h (n = 8). The IT50 for the nerves incubatedin NAC was called IT50NAC. The IT50NAC for the con-centrations of 5.0, 3.5, 2.5, 1.0 and 0.05 mM tested, werealso estimated and the values are shown in Table 1.
Finally, the values of the IT50 NAC were expressed as apercentage of the IT50Ctrl (see Table 1). From the percent-age values shown in Table 1, it was possible to plot the con-centration-effect curve of NAC for the sciatic nerve fibers(Fig. 2C). Finally, using the concentration-effect curve,the IC50 (Inhibitory Concentration 50%), for the NACwas estimated to be 3.02 ± 0.21 mM (n = 5).
At this point, it is worth mentioning that despite the factthat the NAC completely inhibited the CAP of the nervefibers (Fig. 2A, curves b,c and d), it did not cause the acutedeath of nerve fibers. The results shown in Fig. 2B clearlyindicate that after the removal of 3.5 mM NAC from theperfusion chamber and its replacement with normal saline(see arrow in Fig. 2B, curve b), there was almost 75%recovery of the CAP. This pattern of action, where theCAP was complete eliminated in the presence of NACwhile there was over 75% recovery after the removal ofNAC from the nerve preparation, was here called a localanesthetic-like effect. The data in Fig. 2B show a fast recov-ery in the function of the nerve fibers, an indication that nopermanent injury was sustained by the sciatic nerve fibers;however, long-term recording, immediately after the recov-ery of the CAP, shows that the NAC has a delayed delete-rious effect. This is because although NAC was removedfrom the saline and there was a partial recovery of the

M. Moschou et al. / Toxicology in Vitro 22 (2008) 267–274 271
CAP, the vitality of the sciatic nerve fibers was drasticallyreduced, with an IT50NAC of 11.31 ± 1.2 h (from Fig. 2Bcurve b) while the IT50Ctr was 21.94 ± 0.78 h (curve a).From this kind of experiment, it is very clear indeed thatNAC at concentrations of 5.0 mM, 3.5 mM and 2.5 mMcannot be classified as a non-toxic compound for the nervefibers of mammalians.
3.2. The effects of NAC on the duration of the CAP
So far, the time-response curves used to describe theeffects of NAC were limited to the changes in the amplitudeof the CAP, a parameter related to the vitality of the sciaticnerve fibers; however, it seems that NAC has a strong effecton the functioning of the nerve fibers since it caused a dras-tic dose-dependent increase in the duration of the CAP (seehorizontal dotted lines in the CAP of Fig. 3 records A, Band C). The duration of the CAP is a property which isrelated to the functioning of the VGNaCs (phase-a,Fig. 1A) and VGKCs (phase-c) of the sciatic nerve fibers.
Fig. 3. A, B and C are recordings of the evoked CAP of the sciatic nervefibers of the rat incubated in normal saline obtained at time t = 0 h and attime t = IT50. (A) the sciatic nerve fibers were exposed to normal saline.The vertical line indicates the amplitude of the CAP which in this case is5 mV, while the horizontal dotted line shows the duration of the CAP.(B) The nerve fibers were exposed to 2.5 mM NAC and (C) to 1 mMNAC. (D). The percentage increase in the duration of the evoked CAP ofthe isolated sciatic nerve fibers of the rat as a function of incubation timein normal saline and NAC, where 100% was considered the duration att = 0. Curve (a) The nerve fibers were incubated in normal saline only.Curve (b) as in (a) but the nerve was incubated in saline where the finalconcentration of NAC was 3.5 mM, curve (c) in 1.0 mM NAC and curve(d) in 1.0 mM NAC + 250 lM of CdCl2. The dashed vertical line indicatesthe IT50NAC for 2.5 mM. The solid vertical line indicates the IT50 for thenormal saline which is similar to the IT50NAC for 1 mM and IT50NAC for1 mM + 250 lM CdCl2, an is applicable for curves a, c, and d.
In this study, the dispersion of the evoked CAP was alsoused to assess the effects of NAC on the function of the sci-atic nerve fibers and was measured by estimating the dura-tion of the CAP, the time interval between the onset of thepositive peak and return to the baseline. From samplerecords like those shown in Fig. 3, records A, B and C, itwas possible to measure the duration of the CAP accu-rately, during the course of the experiment for nervesexposed to normal saline (A), to NAC 2.5 mM (B) andNAC 1.0 mM (C). From the plots of duration vs. timeshown in Fig. 3D it was possible to estimate the durationof the CAP at time t = 0, which was in all cases0.92 ± 0.1 ms, and time t = IT50. Thus, for the nerves incu-bated in normal saline there was an increase in the durationof the CAP to 123.6 ± 6.96% (estimated from curve a inFig. 3D) at the point of IT50Ctr. The values of the IT50
are obtained from Table 1 and are marked on curve (a)with a solid vertical line (see Fig. 3D). For the nerves incu-bated in 2.5 mM NAC, there was an increase in the dura-tion of the CAP to 192.3 ± 10.13% (n = 5) at the point ofIT50NAC, estimated from curve (b). The value of theIT50NAC (taken from Table 1) on curve (b) is marked bythe dashed vertical line (Fig. 3D). Finally, for the nervesincubated in 1.0 mM NAC there was an increase to161.35 ± 13.87% (n = 5) as was estimated from curve (c).The values of the IT50 on this curve taken also from Table1, are marked by the solid vertical line (Fig. 3D).
The values of the duration of the CAP at 2.5 mM NAC(curve b) were found to be extremely different from those ofthe control (curve a) (P < 0.001). When 1 mM of NAC wasapplied, there was a significant (P < 0.01) increase in theduration of the CAP although there was no significantchange in the amplitude of the CAP, since IT50NAC wasidentical to the value of the IT50Ctr (see Table 1).
3.3. Neuroprotection against Cadmium-induced
neurotoxicity
The results so far have shown that ‘‘high’’ concentra-tions of NAC (2.5 to 5 mM) caused strong inhibition ofthe CAP, while at concentrations equal to less than1 mM (the no observed effect concentration, NOEC) theyhad almost no effect (Fig. 2A). The next question iswhether NAC has a neuroprotective effect on the sciaticnerve fiber preparation or not.
Cadmium-induced neurotoxicity is confirmed in thiscase. Incubation of the sciatic nerve in 250 lM of CdCl2resulted in a drastic decrease in the CAPs amplitude(Fig. 2D, curve b) (n = 6). However, incubation with1 mM NAC for 1 h prior to co-exposure to NAC andCdCl2 proved to be neuroprotective, as the amplitude ofthe CAP remained unaffected (Fig. 2D, curve c) (n = 6),compared to the nerve fibers incubated in the control saline(Fig. 2D curve a). Neuroprotection was not limited to theamplitude of the CAP, since minor effects were observedin the duration of the CAP of the nerves exposed to CdCl2and NAC as is shown by the comparison of curve (d) and

272 M. Moschou et al. / Toxicology in Vitro 22 (2008) 267–274
curve (a) in Fig. 3D. Finally, when the same experimentwas repeated incubating the nerve in CdCl2 in 50 lMNAC, there was no neuroprotection (data not shown)(n = 6).
4. Discussion
The results of this in vitro study, show that NAC hascertain unusual effects on the sciatic nerve fibers of therat. Firstly, at concentrations of 5.0, 3.5 and 2.5 mM, itcompletely inhibits the CAP of the nerve fibers. This effectis observed after a concentration-dependent delay and isnot spontaneously reversible. Such an unusual effect isreported for the first time, indicating that the sciatic nervefibers of the rat are very sensitive in comparison with otherexcitable tissues. NAC, at concentrations greater than orequal to 30 mM, had a toxic effect on diaphragm musclesin vitro (Khawli and Reid, 1994) and 5–10 mM causedreductions in viability in rat and human aortic smoothmuscle cells (Tsai et al., 1996). It is worth noticing that con-centrations of NAC which are already extremely toxic forthe sciatic nerve fibers, namely those of 5.0, 3.5 and 2.5,are far below the concentrations of 10–60 mM, used forthe protection of various cells in cultures with a varietyof chemical compounds (Schulze-Osthoff et al., 1994; Linet al., 1995; Tsai et al., 1996; Seaton et al., 1997; Moham-adin and Abdel-Naim, 1999, 2000; Stredrick et al., 2004;Oh and Lima, 2006). It is obvious that the sciatic nervefibers are far more sensitive to NAC than the cells in cul-tures and this is significant if someone considers the factthat most of the in vitro studies are performed on cell cul-tures and only very few are performed on nerve tissue suchas the optic nerve (Fern et al., 1996).
Secondly, the acute inhibitory action of NAC on thenerve fibers, their CAP was completely eliminated, provedto be reversible only when the NAC was replaced with nor-mal saline, a response similar to the inhibitory action oflocal anesthetics, compounds binding with the VGNaCson the nerve fibers (example, Ritchie, 1979; Balser et al.,1996). This local anesthetic-like effect, combined with theprolongation of the CAP, caused by the increase in theduration of phase-a (VGNaCs) and phase-c (VGKCs) dur-ing prolonged exposure to NAC, is an indication of a pos-sible involvement of voltage-gated channels in theinhibitory action of NAC on the nerve fibers. In this study,we assume that the decrease in amplitude and the increasein duration of the CAP is due to loss of individual fiberaction potential and their parallel prolongation. However,there is a remote possibility that these effects could be dueto the increase of variability in individual fiber actionpotential latency (due to partial depolarization). Thus,the decrease in amplitude and increase in duration that isseen in this study could in fact represent purely a loss insynchronicity of individually perfectly normal actionpotentials. Whatever the mechanism is which changes theshape of the CAP so drastically, the point is that theseeffects appeared only when the nerve was incubated in cer-
tain concentrations of NAC. Although the effects on thesciatic nerve fibers CAP are very clear, there is no informa-tion in the literature about any effect of NAC on the volt-age-gated sodium or potassium channels, except for onecase where it was shown that 10 mM NAC inhibits Na+
absorption across human nasal epithelial cells, affectingthe amiloride-sensitive sodium current (Rochat et al.,2004).
Thirdly, the effects of NAC on the sciatic nerve fiberswere dose-dependent and as was expected, 1 mM had noeffect on the vitality and the amplitude of the CAP in thenerve fibers, while a significant increase in the duration ofthe CAP was still observable. What was not expected wasthe fact that NAC, at a concentration of 1 mM, becamean excellent neuroprotective compound, causing almost100% neuroprotection against cadmium-induced neurotox-icity. Such neurotoxicity is caused mainly by the increase ofreactive oxygen species (ROS) (Fern et al., 1996; Lopezet al., 2006). It is an interesting coincidence that the sameconcentration of NAC, 1 mM, was found to be neuropro-tective against: (a) cadmium-induced neurotoxicityexpressed as inhibition of the brain AChE (Carageorgiouet al., 2004), (b) 4-hydroxynonenal induced neurotoxicityin cerebellar granule neurons (Arakawa et al., 2007) and(c) cadmium-induced neurotoxicity expressed as inhibitionof the evoked CAP of the optic nerve (Fern et al., 1996).
The results of the optic nerve exposed to CdCl2,obtained using suction electrodes (Fern et al., 1996), arethe only experiments which can be compared with thoseof the sciatic nerve exposed to CdCl2, obtained using thethree-chamber recording bath. Although the large sciaticnerve allows recordings of the CAP of better quality, therewere some interesting similarities in the results obtainedwith completely different methodologies. The IT50Cd forthe optic nerve exposed to 200 lM CdCl2 was estimatedto be 35% of the IT50Ctr (calculated from Fern et al.,1996), while for the sciatic nerve exposed to 250 lM itwas the 51% of the IT50Ctr. There is a relatively small dif-ference which could be due either to the difference in thetemperature of the experimental set-ups it was 37 �C forthe optic and 26 �C for the sciatic nerve, or the differencein the thickness of the two nerves. Also, it is worth noticingthat both methods show that 50–60 lM of CdCl2 has noeffect on the nerve fibers (sciatic or optic). This coincidencein the results obtained from two completely differentrecording methods, using similar concentrations of CdCl2(200 and 250 lM) on the same tissue of mammalian nervefibers (from optic and sciatic nerve), indicates that bothmethods are equally reliable for the assessment of theaction of CdCl2 and obviously for other neurotoxicologicalstudies, like assessing the action of NAC.
The neuroprotection at 1 mM NAC, demonstrated inthe sciatic and optic nerves, is due to the reaction againstROS (Fern et al., 1996; Lopez et al., 2006), but it is ratherdifficult to explain how neuroprotection became neurotox-icity as soon as the concentration of NAC increased �2-fold (2.5 mM) causing the complete elimination of the

M. Moschou et al. / Toxicology in Vitro 22 (2008) 267–274 273
CAP. It would be expected that higher concentrations ofNAC, of over 2.5 mM, would completely eliminate allavailable ROS in the sciatic nerve fibers, improving theirfunctioning. However, it seems that certain quantities ofROS are significant for the functioning of the nerve fibers,since there was an almost 75% recovery of the CAP as soonas NAC, 2.5 mM, was removed from the nerve. There is noinformation about the possible role of very low concentra-tion of ROS in the proper functioning of the sciatic nervefibers, but in another case (Xi et al., 2005) ROS were foundto play a significant role in the function of the cells. In thedilation of cerebral arteries, mitochondrial depolarizationincreased the generation of mitochondria-derived ROS thatstimulated Ca2+ sparks and large-conductance Ca2+ -acti-vated potassium (KCa) channels, leading to transient KCa
current activation (Xi et al., 2005). There is a possibilitythat low concentration of ROS interfere also with theproper functioning of sciatic nerve fibers of the rat. Thus,complete deprivation of ROS may led to the malfunctionof nerve fibers which developed by either increasing theduration of the CAP only, phase-(a) (VGNaCs) andphase-(c) (VGKCs), as observed for the concentration of1 mM, or by increasing the duration of the CAP and even-tually eliminating the CAP at higher concentrations over2.5 mM. The interesting point is that these effects werereversed as soon as NAC was removed from the prepara-tion, an indication that the reappearance of ROS initiatealmost immediately the function of the ceased nerve fibers.Finally, it is worth noticing that the vitality of the recov-ered nerve fibers from their exposure to high concentra-tions of NAC was reduced drastically, compared to thecontrols, indicating a delayed neurotoxic effect of this com-pound. Further studies are required to identify the target ofthe action of NAC in the nerve fibers and to establish amodel of action for ROS, like the model described for cere-bral arteries mentioned above (Xi et al., 2005).
NAC is a compound used extensively against oxidativestress and so there is some possibility of its being used asa drug against certain diseases. For example, in vivo mod-ulator action of NAC on glutathione (GSH) status inlead-exposed brain regions of the rat in vivo has beenreported (Nehru and Kanwar, 2007). Oxidative stress hasbeen shown to be a key cause of neuronal dysfunction,and such dysfunction is involved in various neurodegener-ative disorders, including spinocerebellar degeneration,Alzheimer’s disease and Parkinson’s disease (Jesbergerand Richardson, 1991; Simonian and Coyle, 1996;Yamashita et al., 2000). However, other studies haveshown that NAC, also has a toxic effect (Harada et al.,2003; Khawli and Reid, 1994; Brandao et al., 2006). There-fore, due to the unusual neurotoxic-neuroprotective prop-erties of this compound on the sciatic nerve fibersreported here, for the further use of NAC in any in vitro
study or pharmaceutical application of NAC in vivo, fur-ther studies are required to establish in detail the possibleside effects which NAC may have on other parts of the ner-vous system and other excitable tissues.
Acknowledgements
This research was financed by the Greek Ministry of Na-tional Education and Religious Affairs and the EuropeanUnion, within the framework of EPEAEK (PythagorasII). The authors would like to thank Ms. Carol Everhard,Foreign Instructor in the School of English, Aristotle Uni-versity, for proof-reading the manuscript and the anony-mous referee for their valuable corrections.
References
Arakawa, M., Ishimura, A., Arai, Y., Kawabe, K., Suzuki, S., Ishige, K.,Ito, Y., 2007. N-Acetylcysteine and ebselen but not nifedipineprotected cerebellar granule neurons against 4-hydroxynonenal-induced neuronal death. Neuroscience Research 57, 220–229.
Andreou, A., Dabarakis, N., Kagiava, A., Kosmidis, E.K., Geronikaki,A., Theophilidis, G., 2007. Assessing the effects of three dentalimpression materials on the isolated sciatic nerve of rat and frog.Toxicology in Vitro 21, 103–108.
Atkins, K.B., Lodhi, I.J., Hurley, L.L., Hinshaw, D.B., 2000. N-acetylcysteine and endothelial cell injury by sulfur mustard. Journalof Applied Toxicology 20, S125–S128.
Balser, J.R., Nuss, H.B., Romashko, D.N., Marban, E., Tomasselli, G.F.,1996. Functional consequences of lidocaine binding to slow-inacti-vated sodium channels. Journal of General Physiology 107, 643–658.
Brandao, R., Santos, F.W., Zeni, G., Rocha, J.B.T., Nogueira, C.W.,2006. DMPS and N-acetylcysteine induced renal toxicity in miceexposed to mercury. Biometals 19, 389–398.
Cantaro, S., Zara, G., Battaggia, C., di Loreto, P.L., Calo, L., D’Angelo,A., Theophilidis, G., 1998. In vivo and in vitro neurotoxic action ofplasma ultrafiltrate from uraemic patients. Nephrology DialysisTransplantation 13, 2288–2293.
Carageorgiou, H., Tzotzes, V., Pantos, C., Mourouzis, C., Zarros, A.,Tsakiris, S., 2004. In vivo and in vitro effects of cadmium on adult ratbrain total antioxidant status, acetylcholinesterase, (Na+,K+)-ATPaseand Mg2+ -ATPase activities: protection by L-Cysteine. Basic ClinicalPharmacology and Toxicology 94, 112–118.
Cossarizza, A., Franceschi, C., Monti, D., Salvioli, S., Bellesia, E.,Rivabene, R., Biondo, L., Rainaldi, G., Tinari, A., Malorni, W., 1995.Protective effect of N-acetylcysteine in tumor necrosis factor-a-inducedapoptosis in U937 cells: the role of mitochondria. Experimental CellResearch 220, 232–240.
Fern, R., Black, J.A., Ransom, B.R., Waxman, S.G., 1996. Cd2+-inducedinjury in CNS white matter. Journal of Neurophysiology 76, 3264–3273.
Geronikaki, A., Vicini, P., Theophilidis, G., Lagunin, A., Poroikov, V.,Dearden, J.C., 2003. Study of local anesthetic activity of somederivatives of 3-amino-benzo-[d]-isothiazole. SAR QSAR Environ-mental Research 14, 485–495.
Harada, M., Kishimoto, K., Furuhashi, T., Naito, K., Nakashima, Y.,Kawaguchi, Y., Hiraoka, I., 2003. Infertility observed in reproductivetoxicity study of N-acetyl-L-cysteine in rats. Biological Reproduction69, 242–247.
Higuchi, H., Adachi, M., Miura, S., Gores, G.J., Ishii, H., 2001. Themitochondrial permeability transition contributes to acute ethanol-induced apoptosis in rat hepatocytes. Hepatology 34, 320–328.
Jesberger, J.A., Richardson, J.S., 1991. Oxygen free radicals and braindysfunction. International Journal of Neuroscience 57, 1–17.
Khawli, F.A., Reid, M.B., 1994. N-acetylcysteine depresses contractilefunction and inhibits fatigue of diaphragm in vitro. Journal AppliedPhysiology 77, 317–324.
Lin, K.I., Lee, S.H., Narayanan, R., Baraban, J.M., Hardwick, J.M.,Ratan, R.R., 1995. Thiol agents and Bcl-2 identify an alphavirus-induced apoptotic pathway that requires activation of the transcriptionfactor NF-kappa B. The Journal of Cell Biology 131, 1149–1161.

274 M. Moschou et al. / Toxicology in Vitro 22 (2008) 267–274
Lopez, E., Arce, C., Oset-Gasque, M.J., Canadas, S., Gonzalez, M.P.,2006. Cadmium induces reactive oxygen species generation and lipidperoxidation in cortical neurons in culture. Free Radical BiologyMedicine 15, 940–951.
Mayer, M., Noble, M., 1994. N-acetyl-L-cysteine is a pluripotent protectoragainst cell death and enhancer of trophic factor-mediated cell survivalin vitro. Proceedings National Academy of Science 91, 7496–7500.
Mohamadin, A.M., Abdel-Naim, A.B., 1999. Chloroacetonitrile-inducedtoxicity and oxidative stress in rat gastric epithelial cells. Pharmacol-ogy Research 40, 377–383.
Munday, R., 1989. Toxicity of thiols and disulphides: involvement of free-radical species. Free Radical Biology Medicine 7, 659–673.
Nehru, B., Kanwar, S.S., 2007. Modulation by N-Acetylcysteine of lead-induced alterations in rat Brain: reduced glutathione levels andmorphology. Toxicology Mechanisms and Methods 17, 289–293.
Oh, S.H., Lima, S.C., 2006. A rapid and transient ROS generation bycadmium triggers apoptosis via caspase-dependent pathway in HepG2cells and this is inhibited through N-acetylcysteine-mediated catalaseupregulation. Toxicology and Applied Pharmacology 212, 212–223.
Papaefthimiou, C., Cabral, M.D., Mixailidou, C., Viegas, C.A., Sa-Correia, I., Theophilidis, G., 2004. Comparison of two screeningbioassays, based on the frog sciatic nerve and yeast cells, for theassessment of herbicide toxicity. Environmental Toxicology andChemistry 23, 1211–1218.
Ritchie, J.M., 1979. A pharmacological approach to the structure ofsodium channels in myelinated axons. Annual Review Neuroscience 2,341–362.
Rochat, T., Lacroix, J.S., Jornot, I., 2004. N-Acetylcysteine inhibits Na+
absorption across human nasal epithelial cells. Journal of CellPhysiology 201, 106–116.
Schmalbruch, H., 1986. Fiber composition of the rat sciatic nerve.Anatomical Records 215, 2157–2181.
Schulze-Osthoff, K., Krammer, P.H., Droge, W., 1994. Divergent signal-ling via APO-1/Fas and the TNF receptor, two homologous moleculesinvolved in physiological cell death. EMBO Journal 13, 4587–4596.
Seaton, T.A., Cooper, J.M., Schapira, A.H., 1997. Free radical scavengersprotect dopaminergic cell lines from apoptosis induced by complex Iinhibitors. Brain Research 777, 110–118.
Simonian, N.A., Coyle, J.T., 1996. Oxidative stress in neurodegenerativediseases. Annual Review of Pharmacology and Toxicology 36, 83–106.
Stredrick, D.L., Stokes, A.H., Worst, T.J., Freeman, W.M., Johnson,E.A., Lash, L.H., Aschner, M., Vrana, K.E., 2004. Manganese-induced cytotoxicity in dopamine-producing cells. Neurotoxicology25, 543–553.
Theophilidis, G., Benaki, M., Papadopoulou-Mourkidou, E., 1997.Neurotoxic action of six pyrethroid insecticides on the isolated sciaticnerve of a frog (Rana ridibunda). Comparative Biochemistry andPhysiology C Pharmacology Toxicology and Endocrinology 118, 97–103.
Tsai, J.C., Jain, M., Hsieh, C.M., Lee, W.S., Yoshizumi, M., Patterson,C., Perrella, M.A., Cooke, C., Wang, H., Haber, E., Schlegel, R., Lee,M.E., 1996. Induction of apoptosis by pyrrolidinedithiocarbamate andN-acetylcysteine in vascular smooth muscle cells. Journal of BiologicalChemistry 271, 3667–3670.
Wang, X., Martindale, J.L., Holbrook, N.J., 2000. Requirement for ERKactivation in cisplatin-induced apoptosis. Journal of Biological Chem-istry 275 (50), 39435–39443.
Xi, Q., Cheranov, S.Y., Jaggar, J.H., 2005. Mitochondria-derived reactiveoxygen species dilate cerebral arteries by activating Ca2+ sparks.Circulation Research 19, 302–304.
Yamashita, T., Ando, Y., Obayashi, K., Terazaki, H., Sakashita, N.,Uchida, K., Ohama, E., Ando, M., Uchino, M., 2000. Oxidative injuryis present in Purkinje cells in patients with olivopontocerebellaratrophy. Journal of Neurological Science 175, 107–110.
Related Documents











