PLEASE SCROLL DOWN FOR ARTICLE This article was downloaded by: [Khanizadeh, Shahrokh] On: 15 November 2010 Access details: Access Details: [subscription number 929334870] Publisher Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37- 41 Mortimer Street, London W1T 3JH, UK Archives Of Phytopathology And Plant Protection Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713454295 In vitro antifungal activity and mode of action of selected polyphenolic antioxidants on Botrytis cinerea Shutian Tao ab ; Shaoling Zhang a ; Rong Tsao c ; Marie Thérèse Charles b ; Raymond Yang c ; Shahrokh Khanizadeh b a College of Horticulture, Nanjing Agricultural University, Nanjing, Jiangsu Province, P.R. China b Horticultural Research and Development Centre, Agriculture and Agri-Food Canada, St-Jean-sur- Richelieu, Quebec, Canada c Food Research Program, Agriculture and Agri-Food Canada, Guelph, Ontario, Canada Online publication date: 09 November 2010 To cite this Article Tao, Shutian , Zhang, Shaoling , Tsao, Rong , Charles, Marie Thérèse , Yang, Raymond and Khanizadeh, Shahrokh(2010) 'In vitro antifungal activity and mode of action of selected polyphenolic antioxidants on Botrytis cinerea', Archives Of Phytopathology And Plant Protection, 43: 16, 1564 — 1578 To link to this Article: DOI: 10.1080/03235400802583834 URL: http://dx.doi.org/10.1080/03235400802583834 Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Khanizadeh, Shahrokh]On: 15 November 2010Access details: Access Details: [subscription number 929334870]Publisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Archives Of Phytopathology And Plant ProtectionPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713454295
In vitro antifungal activity and mode of action of selected polyphenolicantioxidants on Botrytis cinereaShutian Taoab; Shaoling Zhanga; Rong Tsaoc; Marie Thérèse Charlesb; Raymond Yangc; ShahrokhKhanizadehb
a College of Horticulture, Nanjing Agricultural University, Nanjing, Jiangsu Province, P.R. China b
Horticultural Research and Development Centre, Agriculture and Agri-Food Canada, St-Jean-sur-Richelieu, Quebec, Canada c Food Research Program, Agriculture and Agri-Food Canada, Guelph,Ontario, Canada
Online publication date: 09 November 2010
To cite this Article Tao, Shutian , Zhang, Shaoling , Tsao, Rong , Charles, Marie Thérèse , Yang, Raymond andKhanizadeh, Shahrokh(2010) 'In vitro antifungal activity and mode of action of selected polyphenolic antioxidants onBotrytis cinerea', Archives Of Phytopathology And Plant Protection, 43: 16, 1564 — 1578To link to this Article: DOI: 10.1080/03235400802583834URL: http://dx.doi.org/10.1080/03235400802583834
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

In vitro antifungal activity and mode of action of selected polyphenolic
antioxidants on Botrytis cinerea
Shutian Taoa,b, Shaoling Zhanga, Rong Tsaoc, Marie Therese Charlesb,Raymond Yangc and Shahrokh Khanizadehb*
aCollege of Horticulture, Nanjing Agricultural University, 1 Weigang, Nanjing 210095, JiangsuProvince, P.R. China; bHorticultural Research and Development Centre, Agriculture and Agri-FoodCanada, 430 Gouin Blvd., St-Jean-sur-Richelieu, J3B 3E6, Quebec, Canada; cFood Research Program,Agriculture and Agri-Food Canada, 93 Stone Road West, Guelph N1G 5C9, Ontario, Canada
(Received 22 September 2008; final version received 22 October 2008)
Six selected antioxidants (catechin, quercetin-3-galactoside, cyanidin-3-glucoside,pelargonidin-3-glucoside, ellagic and gallic acids) were evaluated in vitro for theirantifungal activities and mode of action on Botrytis cinerea Pers., one of the mostimportant pathogens of strawberries. Inhibitory effects were found for all the testedantioxidants, but varied at different fungal developmental stages. Catechin andquercetin-3-galactoside showed linear inhibitory effects on germ tube elongation, withthe highest suppression ratios of 54.8% and 58.8% respectively. No significant effectwas found on spore germination between treatments and control. Gallic acid showedvery strong and linear inhibition on spore germination (r ¼ 70.95), but the effectdiminished after spore germination. Cyanidin-3-glucoside and pelargonidin-3-glucosideprovided effective control on the fungi as concentrations increased. The arresting effectof ellagic acid on development of B. cinerea was quadratic. Ellagic acid inhibited germtube elongation and mycelial growth at its highest and lowest concentrations, while noeffects were observed at its medium concentration used in this study.
Keywords: Botrytis cinerea; ellagic acid; gallic acid; anthocyanin; catechin; flavonol
Introduction
Strawberry (Fragaria x ananassa Duch) as an important cash crop in Canada is verynutritional but also delicate and perishable. Gray mold is the most destructive, commonand widely distributed fungal disease caused by Botrytis cinerea Pers., causing seriouslosses to strawberry during the growing season and storage. Disease control is difficult,because the pathogen can infect all plant parts at any stages of growth and also it is notpossible to use fungicide at harvesting. The effect of gray mold is more pronounced duringthe post-harvest storage due to environmental conditions and the declining resistance offruit with its ripening process which causes short shelf life and decay by as much as 55%(Martınez-Romero et al. 2007).
The sensitivity of Botrytis cinerea to plant phytochemicals such as phenolic compoundshave been studied by several research groups (Hebert et al. 2002; Khanizadeh et al.2002b; Daferera et al. 2003; Soylu 2006; Lee et al. 2007; Martınez-Romero et al. 2007;
*Corresponding author. Email: [email protected]
Archives of Phytopathology and Plant Protection
Vol. 43, No. 16, 1 November 2010, 1564–1578
ISSN 0323-5408 print/ISSN 1477-2906 online
� 2010 Taylor & Francis
DOI: 10.1080/03235400802583834
http://www.informaworld.com
Downloaded By: [Khanizadeh, Shahrokh] At: 16:29 15 November 2010

Ehsani-Moghaddam et al. 2008; Khanizadeh et al. 2008a, 2008b); Chinese medicinal plantextracts, plant essential oil, rich in phenolics and carvacrol (a kind of phenolic compound)were found to be effective at inhibiting Botrytis cinerea growth in vitro (Daferera et al.2003; Lee et al. 2007; Martınez-Romero et al. 2007). Feucht et al. (1992) detected aboundary zone surrounding Botrytis-infected strawberry tissues enriched in catechin, themain constitutive unit of proanthocyanidin, which are oligomers of flavan-3-ols. In ourlab, we also established a positive correlation between the antioxidant capacity, shelf lifeand disease resistance (Khanizadeh et al. 2002b, 2007, 2008a, 2008b; Ehsani-Moghaddamet al. 2008).
Hebert et al. (2002) reported a higher degree of resistant to gray mold growth of sixstrawberry cultivars and concluded that ‘Seascape’ proanthocyanidins (PA) extract gavethe highest inhibition of mold radial growth (76.2%) and that its fruit was also the mostresistant to the appearance of mold during storage. However, the result did not indicatewhich phenolic composition or combinations are responsible for inhibiting growth of thegray mold in vitro.
Strawberries have been shown to be a rich source of phenolic compounds withantioxidant activity (Wang et al. 1996; Guo et al. 2003; Scalzo et al. 2005; Khanizadehet al. 2006, 2008a), but several factors influence the antioxidant capacity and thecomposition of bioactivity compounds including cultural practices, pre-harvest conditions,maturity, post-harvest handling and processing (Gil et al. 1997; Wang and Lin 2000, 2003;Wang and Zhang 2001; Ayala-Zavala et al. 2004; Kosar et al. 2004; Cordenunsi et al.2005; Vicente et al. 2006; Ferreyra et al. 2007; Hernanz et al. 2007; Wang et al. 2007) andgenetic variability is one of the most important factors.
In spite of many studies on the antioxidant and bioactive composition of strawberryfruits (Heinonen et al. 1998; Hebert et al. 2002; Wang et al. 2002; Terry et al. 2004; Heoand Lee 2005; Klopotek et al. 2005; Rekika et al. 2005; Khanizadeh et al. 2006, 2008a;Hernanz et al. 2007; Pinto et al. 2007), no study has been done on individual antioxidantsand their effect on gray mold growth and development.
The objective of this study is to simulate the effect of selected antioxidants on graymold growth and development including spore germination, germ tube elongation andradial growth of Botrytis cinerea, and if they can be of any use as biochemical markers toselect for B. cinerea resistance in a strawberry breeding program.
Materials and method
Chemicals
Catechin, cyaniding-3-glucoside, pelargonidin-3-glucoside and quercetin-3-galactosidewere purchased from Indofine Chemical Co. (Somerville, NJ, USA); ellagic acid, gallicacid, glycerol, NaOH and glucose were from Sigma-Aldrich (St. Louis, MO, USA); potatodextrose agar (PDA) was obtained from Difco Laboratories, Detroit (Sparks, MD, USA);KH2PO4 was from Fisher Scientific Company (Fair Lawn, NJ, USA); ethanol was fromCommercial alcohol Inc. (Brampton, ON, Canada).
Media preparation
Ellagic acid was dissolved in 0.2% NaOH aqueous solution; gallic acid, catechin andquercetin-3-galactoside were dissolved in 25% ethanol; cyaniding-3-glucoside andpelargonidin-3-glucoside were dissolved in double distilled water directly. All solutions
Archives of Phytopathology and Plant Protection 1565
Downloaded By: [Khanizadeh, Shahrokh] At: 16:29 15 November 2010

were then sterilised with a 0.2 mm syringe filter (Nalgene, Rochester, NY, USA) and usedas stock solutions for making culture media.
The media were prepared by appropriate dilution of the stock solutions with PDA toevaluate mycelial growth. The following concentrations were made based on a preliminaryrange-finding study: ellagic acid, 18, 36, 54, 72, and 90 ppm; gallic acid, 50, 100, 150, 200and 250 ppm; catechin, 120, 240, 360, 480 and 600 ppm; cyaniding-3-glucoside, 10, 20,30,40 and 50 ppm; pelargonidin-3-glucoside, 10, 20, 30,40 and 50 ppm and quercetin-3-galactoside, 12, 24, 36, 48 and 60 ppm. PDA alone and PDA with NaOH or ethanol wereused together as controls when NaOH or ethanol was used to dissolve the antioxidants.
Mixtures of 0.3% KH2PO4, 1% glucose and 2.5% glycerol with a selectedconcentration of antioxidants were prepared for spore germination and germ tube growth(Li et al. 2003).
Gray mold isolate preparation
The isolation was performed based on Koch’s postulate and according to the method ofAgrios (1997) and Raposo et al. (1995) with some modification.
Strawberry fruits with typical symptoms of gray mold were collected and brought tothe lab from the green house of Horticultural Research and Development Centre(CRDH), Agriculture and Agri-Food Canada (AAFC-AAC) in St-Jean-Sur-Richelieu.Spores on the furthermost top surface of the fruit were picked out with inoculating needleunder a microscope, and transferred to PDA plates. The plates with spores were thenincubated in the dark at 18.58C for 7 d, and fungal culture was purified through sub-culturing. Finally, the pure isolates of Botrytis cinerea Pers. were maintained on PDAplates until required for the experiment.
The conidia of Botrytis cinerea Pers. were obtained by flooding dishes with sterile 5%glycerol aqueous solution. The suspensions were filtered through three lays of sterilecheesecloth in order to remove the adhering mycelia, and the concentration of conidia wasadjusted to 20,000 conidia/ml using a hemocytometer.
Inoculation and measurement
The PDAs enriched with different levels of antioxidants were inoculated with 4 mmdimeter mycelial plugs cut from the edge of seven-day-old actively growing cultures ofBotrytis cinerea Pers. (Tremblay et al. 2003). The cultures were then incubated in the darkat 18.58C for 72 h and the mycelia were measured at 24 h, 36 h, 48 h and 72 h intervalsafter incubation using a calliper. Spore germination and germ tubes were measured on asuspension of conidia adjusted to a final concentration of 105 conidia/ml after 24 hincubated in the dark at 18.58C (Li et al. 2003). An inverted microscope (Leica DMIL)connected with a cooled CCD camera (Lumenera, Infinty 3–1) was used to measure sporegermination and germ tube at 12 h, 18 h and 24 h intervals.
Experimental design and data analysis
A completely randomised design with three replicates was used to test the selected levels ofantioxidant at 24, 36, 48 and 72 h intervals for mycelial growth and 12, 18 and 24 hintervals for germ tube and spore germination. Three plates were used for each replicateand the experiment was repeated twice under the same conditions. Each experiment wasanalysed separately and the two set of data were pooled together after testing the
1566 S. Tao et al.
Downloaded By: [Khanizadeh, Shahrokh] At: 16:29 15 November 2010

homogeneity of the two experimental errors. Two controls (PDA and PDA plus solvent)were used to test the efficacy of the selected levels and a least significant test (LSD) wasused to separate the means. The effect of different levels of antioxidant for each timeinterval was tested using the orthogonal polynomial (Steel and Torrie 1980) contrast ofSAS using the GLM procedure (SAS Institute 1989).
Results
No significant difference was observed between the two controls used for testing ellagicand gallic acids, catechin and quercertin-3-galactoside, indicating that the addition ofeither 2% NaOH or 25% ethanol as a solvent during the media preparation had noeffect on spore germination rate, germ tube length and mycelial growth (Tables 1, 2, 5,and 6).
Ellagic acid
Ellagic acid had no effect on spore germination rate after inoculation (Table 1) while thegerm tube length and mycelial diameter were reduced significantly at all intervals afterinoculation. The effect of concentration of ellagic acid on germ tube and mycelial growthwas quadratic, which means the lower levels of ellagic acid had a negative effect on thegerm tube length and mycelial growth diameter, whereas higher concentrations elongatedthe germ tube and promoted mycelial growth. The optimum level to reduce the germ tubelength and mycelial growth diameter was estimated at about 36 ppm.
Gallic acid
Similar to ellagic acid, gallic acid did not have any effect on spore germination rate except24 h after inoculation when a significant negative linear relationship was observed betweengallic acid concentration and the spore germination rate (Table 2). Gallic acid had asignificant linear negative effect on germ tube length and it was very pronounced at 18 hafter inoculation (F ¼ 12.19) but its effect diminished after 24 h (Table 2). Mycelial growthdiameter was not affected by any level of gallic acid at any interval applied in this studyafter inoculation.
Cyanidin-3-glucoside
Cyanidin-3-glucoside showed significantly linear negative effects on spore germination,but its effect diminished after 18 h. Cyanidin-3-glucoside had a distinct effect on reducingthe germ tube length as well as mycelial growth diameters and this effect was carried overall examined periods. The most extreme effect was observed at 24 h (9.76***) and 72 h(14.90***) respectively for germ tube and mycelial growth (Table 3).
Pelargonidin-3-glucoside
Pelargonidin-3-glucoside, another anthocyanin, showed a significantly linear effect onspore germination over the examined period, however in germ tube growth, theeffect was quadratic until 18 h after inoculation and then linear at 24 h. Ana-logously, the linear effect did not appear in initial periods but was significant from 48 hon (Table 4).
Archives of Phytopathology and Plant Protection 1567
Downloaded By: [Khanizadeh, Shahrokh] At: 16:29 15 November 2010
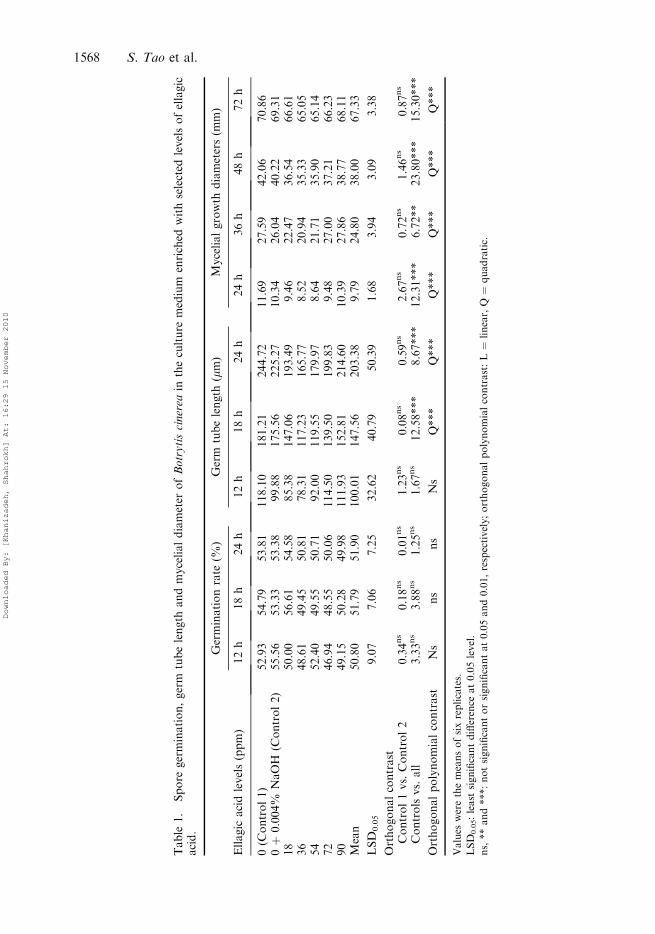
Table
1.
Spore
germination,germ
tubelength
andmycelialdiameter
ofBotrytiscinerea
intheculture
medium
enriched
withselected
levelsofellagic
acid.
Germinationrate
(%)
Germ
tubelength
(mm)
Mycelialgrowth
diameters(m
m)
Ellagic
acidlevels(ppm)
12h
18h
24h
12h
18h
24h
24h
36h
48h
72h
0(C
ontrol1)
52.93
54.79
53.81
118.10
181.21
244.72
11.69
27.59
42.06
70.86
0þ
0.004%
NaOH
(Control2)
55.56
53.33
53.38
99.88
175.56
225.27
10.34
26.04
40.22
69.31
18
50.00
56.61
54.58
85.38
147.06
193.49
9.46
22.47
36.54
66.61
36
48.61
49.45
50.81
78.31
117.23
165.77
8.52
20.94
35.33
65.05
54
52.40
49.55
50.71
92.00
119.55
179.97
8.64
21.71
35.90
65.14
72
46.94
48.55
50.06
114.50
139.50
199.83
9.48
27.00
37.21
66.23
90
49.15
50.28
49.98
111.93
152.81
214.60
10.39
27.86
38.77
68.11
Mean
50.80
51.79
51.90
100.01
147.56
203.38
9.79
24.80
38.00
67.33
LSD
0.05
9.07
7.06
7.25
32.62
40.79
50.39
1.68
3.94
3.09
3.38
Orthogonalcontrast
Control1vs.Control2
0.34ns
0.18ns
0.01ns
1.23ns
0.08ns
0.59ns
2.67ns
0.72ns
1.46ns
0.87ns
Controlsvs.all
3.33ns
3.88ns
1.25ns
1.67ns
12.58***
8.67***
12.31***
6.72**
23.80***
15.30***
Orthogonalpolynomialcontrast
Ns
ns
ns
Ns
Q***
Q***
Q***
Q***
Q***
Q***
Values
werethemeansofsixreplicates.
LSD
0.05:least
significantdifference
at0.05level.
ns,**and***:notsignificantorsignificantat0.05and0.01,respectively;orthogonalpolynomialcontrast:L¼
linear,Q¼
quadratic.
1568 S. Tao et al.
Downloaded By: [Khanizadeh, Shahrokh] At: 16:29 15 November 2010
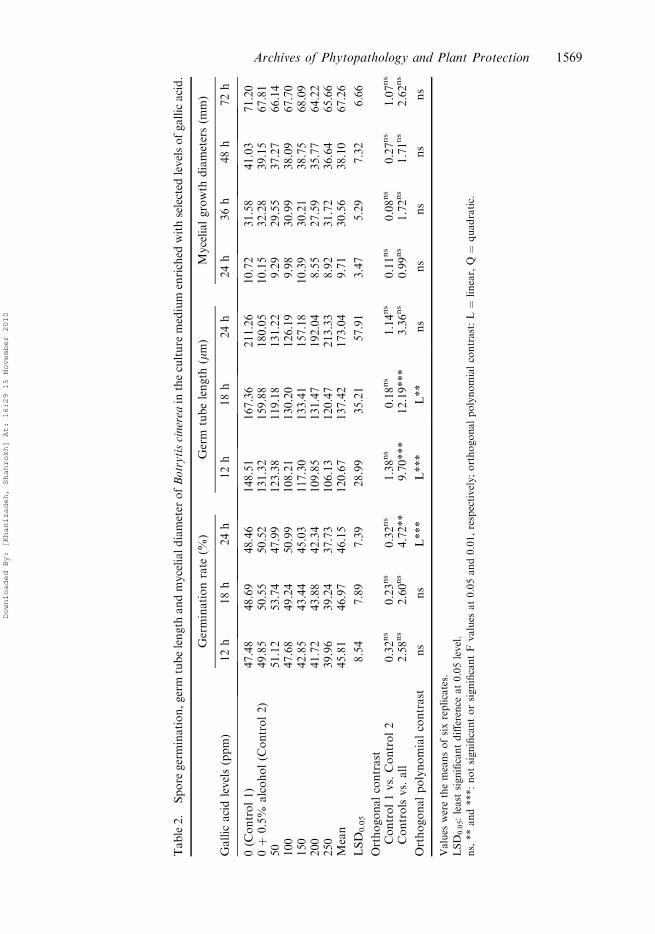
Table2.
Spore
germination,germ
tubelength
andmycelialdiameter
ofBotrytiscinerea
intheculture
medium
enriched
withselected
levelsofgallicacid.
Germinationrate
(%)
Germ
tubelength
(mm)
Mycelialgrowth
diameters(m
m)
Gallic
acidlevels(ppm)
12h
18h
24h
12h
18h
24h
24h
36h
48h
72h
0(C
ontrol1)
47.48
48.69
48.46
148.51
167.36
211.26
10.72
31.58
41.03
71.20
0þ
0.5%
alcohol(C
ontrol2)
49.85
50.55
50.52
131.32
159.88
180.05
10.15
32.28
39.15
67.81
50
51.12
53.74
47.99
123.38
119.18
131.22
9.29
29.55
37.27
66.14
100
47.68
49.24
50.99
108.21
130.20
126.19
9.98
30.99
38.09
67.70
150
42.85
43.44
45.03
117.30
133.41
157.18
10.39
30.21
38.75
68.09
200
41.72
43.88
42.34
109.85
131.47
192.04
8.55
27.59
35.77
64.22
250
39.96
39.24
37.73
106.13
120.47
213.33
8.92
31.72
36.64
65.66
Mean
45.81
46.97
46.15
120.67
137.42
173.04
9.71
30.56
38.10
67.26
LSD
0.05
8.54
7.89
7.39
28.99
35.21
57.91
3.47
5.29
7.32
6.66
Orthogonalcontrast
Control1vs.Control2
0.32ns
0.23ns
0.32ns
1.38ns
0.18ns
1.14ns
0.11ns
0.08ns
0.27ns
1.07ns
Controlsvs.all
2.58ns
2.60ns
4.72**
9.70***
12.19***
3.36ns
0.99ns
1.72ns
1.71ns
2.62ns
Orthogonalpolynomialcontrast
ns
ns
L***
L***
L**
ns
ns
ns
ns
ns
Values
werethemeansofsixreplicates.
LSD
0.05:least
significantdifference
at0.05level.
ns,**and***:notsignificantorsignificantF
values
at0.05and0.01,respectively;orthogonalpolynomialcontrast:L¼
linear,Q¼
quadratic.
Archives of Phytopathology and Plant Protection 1569
Downloaded By: [Khanizadeh, Shahrokh] At: 16:29 15 November 2010
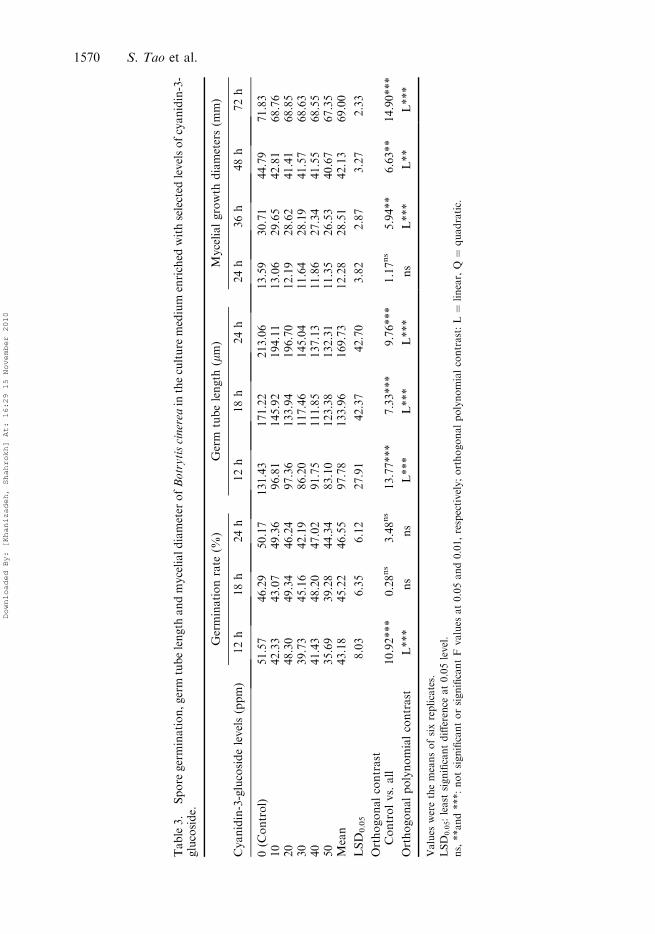
Table3.
Spore
germination,germ
tubelength
andmycelialdiameter
ofBotrytiscinerea
intheculture
medium
enriched
withselected
levelsofcyanidin-3-
glucoside.
Germinationrate
(%)
Germ
tubelength
(mm)
Mycelialgrowth
diameters(m
m)
Cyanidin-3-glucosidelevels(ppm)
12h
18h
24h
12h
18h
24h
24h
36h
48h
72h
0(C
ontrol)
51.57
46.29
50.17
131.43
171.22
213.06
13.59
30.71
44.79
71.83
10
42.33
43.07
49.36
96.81
145.92
194.11
13.06
29.65
42.81
68.76
20
48.30
49.34
46.24
97.36
133.94
196.70
12.19
28.62
41.41
68.85
30
39.73
45.16
42.19
86.20
117.46
145.04
11.64
28.19
41.57
68.63
40
41.43
48.20
47.02
91.75
111.85
137.13
11.86
27.34
41.55
68.55
50
35.69
39.28
44.34
83.10
123.38
132.31
11.35
26.53
40.67
67.35
Mean
43.18
45.22
46.55
97.78
133.96
169.73
12.28
28.51
42.13
69.00
LSD
0.05
8.03
6.35
6.12
27.91
42.37
42.70
3.82
2.87
3.27
2.33
Orthogonalcontrast
Controlvs.all
10.92***
0.28ns
3.48ns
13.77***
7.33***
9.76***
1.17ns
5.94**
6.63**
14.90***
Orthogonalpolynomialcontrast
L***
ns
ns
L***
L***
L***
ns
L***
L**
L***
Values
werethemeansofsixreplicates.
LSD
0.05:least
significantdifference
at0.05level.
ns,**and***:notsignificantorsignificantF
values
at0.05and0.01,respectively;orthogonalpolynomialcontrast:L¼
linear,Q¼
quadratic.
1570 S. Tao et al.
Downloaded By: [Khanizadeh, Shahrokh] At: 16:29 15 November 2010
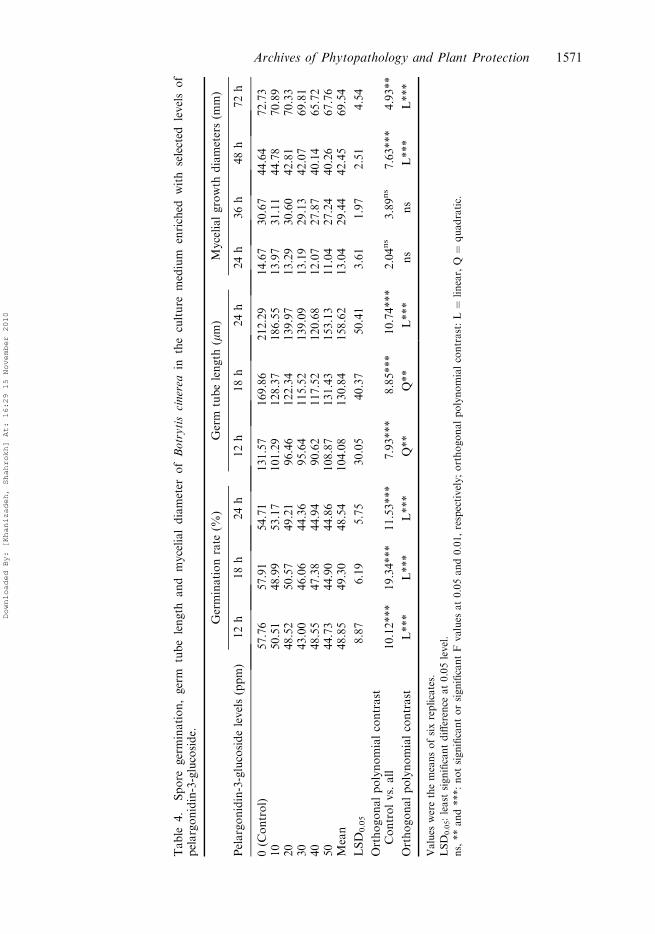
Table
4.
Spore
germination,germ
tubelength
and
mycelialdiameter
ofBotrytiscinerea
intheculture
medium
enriched
with
selected
levelsof
pelargonidin-3-glucoside.
Germinationrate
(%)
Germ
tubelength
(mm)
Mycelialgrowth
diameters(m
m)
Pelargonidin-3-glucosidelevels(ppm)
12h
18h
24h
12h
18h
24h
24h
36h
48h
72h
0(C
ontrol)
57.76
57.91
54.71
131.57
169.86
212.29
14.67
30.67
44.64
72.73
10
50.51
48.99
53.17
101.29
128.37
186.55
13.97
31.11
44.78
70.89
20
48.52
50.57
49.21
96.46
122.34
139.97
13.29
30.60
42.81
70.33
30
43.00
46.06
44.36
95.64
115.52
139.09
13.19
29.13
42.07
69.81
40
48.55
47.38
44.94
90.62
117.52
120.68
12.07
27.87
40.14
65.72
50
44.73
44.90
44.86
108.87
131.43
153.13
11.04
27.24
40.26
67.76
Mean
48.85
49.30
48.54
104.08
130.84
158.62
13.04
29.44
42.45
69.54
LSD
0.05
8.87
6.19
5.75
30.05
40.37
50.41
3.61
1.97
2.51
4.54
Orthogonalpolynomialcontrast
Controlvs.all
10.12***
19.34***
11.53***
7.93***
8.85***
10.74***
2.04ns
3.89ns
7.63***
4.93**
Orthogonalpolynomialcontrast
L***
L***
L***
Q**
Q**
L***
ns
ns
L***
L***
Values
werethemeansofsixreplicates.
LSD
0.05:least
significantdifference
at0.05level.
ns,**and***:notsignificantorsignificantFvalues
at0.05and0.01,respectively;orthogonalpolynomialcontrast:L¼
linear,Q¼
quadratic.
Archives of Phytopathology and Plant Protection 1571
Downloaded By: [Khanizadeh, Shahrokh] At: 16:29 15 November 2010
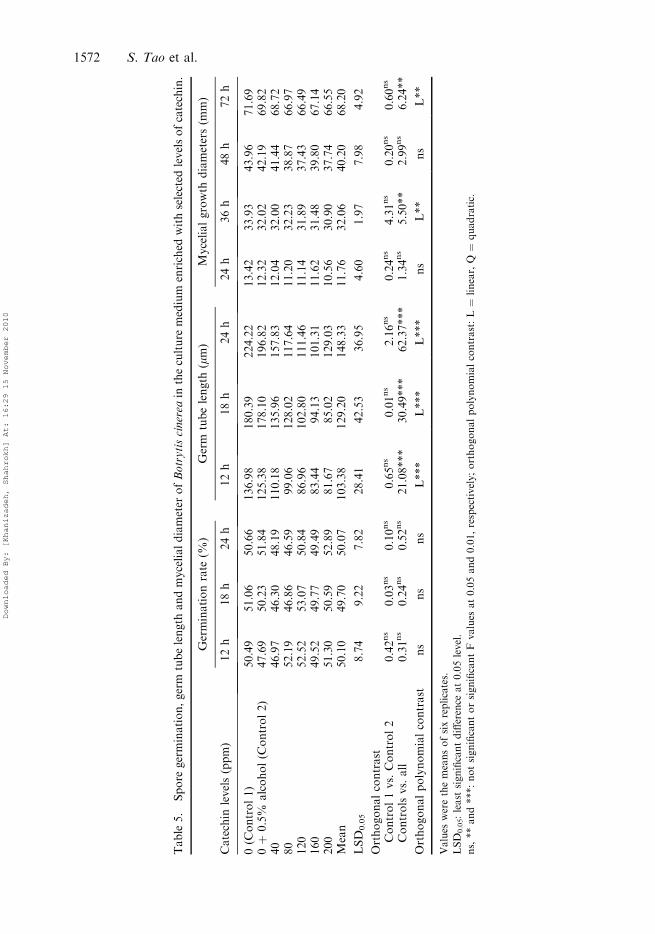
Table5.
Spore
germination,germ
tubelength
andmycelialdiameter
ofBotrytiscinerea
intheculture
medium
enriched
withselected
levelsofcatechin.
Germinationrate
(%)
Germ
tubelength
(mm)
Mycelialgrowth
diameters(m
m)
Catechin
levels(ppm)
12h
18h
24h
12h
18h
24h
24h
36h
48h
72h
0(C
ontrol1)
50.49
51.06
50.66
136.98
180.39
224.22
13.42
33.93
43.96
71.69
0þ
0.5%
alcohol(C
ontrol2)
47.69
50.23
51.84
125.38
178.10
196.82
12.32
32.02
42.19
69.82
40
46.97
46.30
48.19
110.18
135.96
157.83
12.04
32.00
41.44
68.72
80
52.19
46.86
46.59
99.06
128.02
117.64
11.20
32.23
38.87
66.97
120
52.52
53.07
50.84
86.96
102.80
111.46
11.14
31.89
37.43
66.49
160
49.52
49.77
49.49
83.44
94.13
101.31
11.62
31.48
39.80
67.14
200
51.30
50.59
52.89
81.67
85.02
129.03
10.56
30.90
37.74
66.55
Mean
50.10
49.70
50.07
103.38
129.20
148.33
11.76
32.06
40.20
68.20
LSD
0.05
8.74
9.22
7.82
28.41
42.53
36.95
4.60
1.97
7.98
4.92
Orthogonalcontrast
Control1vs.Control2
0.42ns
0.03ns
0.10ns
0.65ns
0.01ns
2.16ns
0.24ns
4.31ns
0.20ns
0.60ns
Controlsvs.all
0.31ns
0.24ns
0.52ns
21.08***
30.49***
62.37***
1.34ns
5.50**
2.99ns
6.24**
Orthogonalpolynomialcontrast
ns
ns
ns
L***
L***
L***
ns
L**
ns
L**
Values
werethemeansofsixreplicates.
LSD
0.05:least
significantdifference
at0.05level.
ns,**and***:notsignificantorsignificantFvalues
at0.05and0.01,respectively;orthogonalpolynomialcontrast:L¼
linear,Q¼
quadratic.
1572 S. Tao et al.
Downloaded By: [Khanizadeh, Shahrokh] At: 16:29 15 November 2010
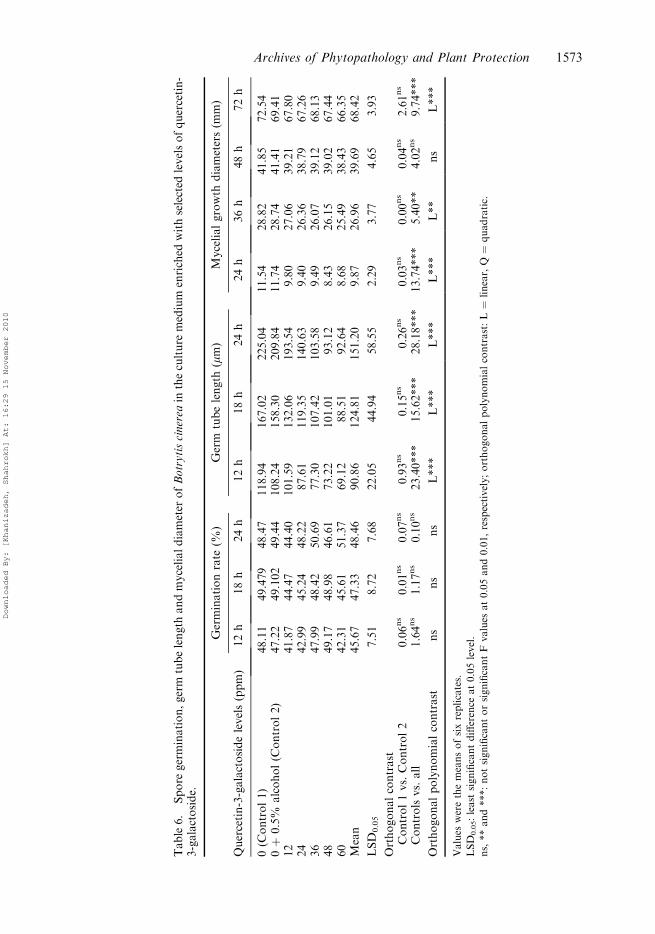
Table6.
Spore
germination,germ
tubelength
andmycelialdiameter
ofBotrytiscinerea
intheculture
medium
enriched
withselected
levelsofquercetin-
3-galactoside.
Germinationrate
(%)
Germ
tubelength
(mm)
Mycelialgrowth
diameters(m
m)
Quercetin-3-galactosidelevels(ppm)
12h
18h
24h
12h
18h
24h
24h
36h
48h
72h
0(C
ontrol1)
48.11
49.479
48.47
118.94
167.02
225.04
11.54
28.82
41.85
72.54
0þ
0.5%
alcohol(C
ontrol2)
47.22
49.102
49.44
108.24
158.30
209.84
11.74
28.74
41.41
69.41
12
41.87
44.47
44.40
101.59
132.06
193.54
9.80
27.06
39.21
67.80
24
42.99
45.24
48.22
87.61
119.35
140.63
9.40
26.36
38.79
67.26
36
47.99
48.42
50.69
77.30
107.42
103.58
9.49
26.07
39.12
68.13
48
49.17
48.98
46.61
73.22
101.01
93.12
8.43
26.15
39.02
67.44
60
42.31
45.61
51.37
69.12
88.51
92.64
8.68
25.49
38.43
66.35
Mean
45.67
47.33
48.46
90.86
124.81
151.20
9.87
26.96
39.69
68.42
LSD
0.05
7.51
8.72
7.68
22.05
44.94
58.55
2.29
3.77
4.65
3.93
Orthogonalcontrast
Control1vs.Control2
0.06ns
0.01ns
0.07ns
0.93ns
0.15ns
0.26ns
0.03ns
0.00ns
0.04ns
2.61ns
Controlsvs.all
1.64ns
1.17ns
0.10ns
23.40***
15.62***
28.18***
13.74***
5.40**
4.02ns
9.74***
Orthogonalpolynomialcontrast
ns
ns
ns
L***
L***
L***
L***
L**
ns
L***
Values
werethemeansofsixreplicates.
LSD
0.05:least
significantdifference
at0.05level.
ns,**and***:notsignificantorsignificantF
values
at0.05and0.01,respectively;orthogonalpolynomialcontrast:L¼
linear,Q¼
quadratic.
Archives of Phytopathology and Plant Protection 1573
Downloaded By: [Khanizadeh, Shahrokh] At: 16:29 15 November 2010

Catechin
Catechin had no effect on spore germination rate but significantly reduced the germ tubelength and mycelial growth diameter and its effect was very pronounced especially after24 h (62.37***) and 72 h (6.24**), respectively.
Quercetin-3-galactoside
Quercetin-3-galactoside showed a similar effect to catechin. It had no effect on sporegermination rate, but significantly reduced the germ tube length and mycelial growthdiameter and its effect was very pronounced especially after 24 h (628.187***) and 72 h(9.74**), respectively.
Discussion
Gray mold development is one of the principal causes of premature termination of post-harvest shelf life of strawberry. Numerous studies have conclusively established a key rolefor constitutive and induced phenolic compounds with antioxidant properties in thedefence of plant tissues against pathogen attacks (Nicholson and Hammerschmidt 1992;Prusky 1996). Strawberry fruit, ellagic acid, gallic acid along with anthocyanins(cyaniding-3-glucoside, pelargonidin-3-glucoside), proanthocyanidins (catechin) andflavonols (quercetin-3-galactoside) are the main phenolic compounds with antioxidantproperties. These compounds are known to influence the disease resistance and shelf life ofthe fruit. A threefold increase in ellagic acid content was observed in raspberry leavesinfected by orange rust (Wang et al. 1994), and high level of catechins, the mainconstitutive unit of proanthocyanidins (PA), found in immature strawberry fruits wasassociated with resistance to gray mold (Feucht et al. 1992). In vitro studies revealed thatproanthocyanidins (PA) of some strawberry cultivars reduced hyphal growth of graymold, and fruit of such cultivars high in PA content were more resistant to storage rotdevelopment (Hebert et al. 2002). In the present study, antifungal activity was observed forall antioxidants, and dose dependent antifungal activity of the antioxidants was also foundat every developmental stage of B. cinerea, and almost every antioxidant showed asignificantly linear inhibitory effect on pathogen progress. For example, spore germinationwas strongly inhibited by increased concentrations of gallic acid (r ¼ 70.95); catechin andquercetin-3-galactoside show a negative relationship with germ tube elongation(r ¼ 70.96 and r ¼ 70.98, respectively), and cyanidin-3-glucoside and pelargonidin-3-glucoside also provided effective control of the fungi as concentrations increased. Theseindicate that strawberry cultivars with varied phenolic contents (Meyers et al. 2003; Aabyet al. 2005; Rekika et al. 2005; Khanizadeh et al. 2006, 2008a) should have different shelflives and susceptibility to pathogen infection. It has been reported that high antioxidantcapacity was found in Harmonie, Saint-Jean d’Orleans and Saint-Laurent d’Orleans,which exhibited better shelf life stability than Kent (Khanizadeh et al. 2005a, 2005b,2005c).
However, the rule is not always straightforward. For example, Saint Pierre which ischaracterised by very good shelf life (Khanizadeh et al. 2002a) displayed a similar level ofantioxidants (gallic acid, epicatechin, catechin and ellagic acid) to other cultivars with ashort shelf life. Rosemary oil had half the amount of eucalyptol compared with the sageoil, but it was shown to be more effective at fungal inhibition (Daferera et al. 2003). It isinteresting to note that the effects of almost all tested antioxidants were linear except for
1574 S. Tao et al.
Downloaded By: [Khanizadeh, Shahrokh] At: 16:29 15 November 2010

ellagic acid which was quadratic (Table 1). This indicates that ellagic acid has an effect ongerm tube and mycelial growth at low concentration (36 ppm), whereas the higher level ofellagic acid promotes germ tube elongation and mycelial growth similar to the effect ofgrowth hormone.
Several papers in the literature demonstrated that plants accumulate phenolic compoundsunder infection or elicited resistance to pathogen (Wang et al. 1994; Soylu 2006; Liu et al.2007), however, it is important to note that there might be an interaction between individualphenolic compounds which may cause a synergistic effect, and the presence of other enzymessuch as PPO, POD and H2O2 burst in the plant might also affect the susceptibility of acultivar (Khanam et al. 2005). It has been reported that grape pranthocyanidinscompetitively inhibit the activity of laccase produced by B. cinerea, but only in the presenceof tannins (Elad and Evensen 1995). Lee et al. (2007) reported that combinations of herbextracts had a higher inhibitory effect towards tested fungi than that of individual extract.Although the catabolic phloridzin derivatives did not inhibit fungal mycerial growth ofPhlyctaeea vagabunda, Lattanzio et al. (2001) reported that phloridzin and chlorogenic acidin combination with polyphenol oxidase activity could function to arrest P. vagabunda inquiescent infections associated with immature and ripening apple fruit.
In the present study, the effects of the six antioxidants were different at differentdevelopmental stages of the fungi progress, for example, catechin and quercetin-3-galactoside exhibited very significant linear inhibitory effects on germ tube elongation,with the highest suppression ratio of 54.8% and 58.8%, respectively, but no significanteffects were found on spore germination. Meanwhile, gallic acid inhibited sporegermination strongly but weakly on germ tube elongation and mycelial growth, andhigher concentrations may be needed (Hur et al. 2003). Similar results were found betweenchitosan and Penicillium expansum and B. cinerea (Liu et al. 2007). Furthermore, whenlettuce leaves were inoculated with B. cinerea, lettucenin A accumulation was associatedwith limited lesions restricting the spread of disease, but when leaves were inoculated withmycelial suspensions, lettucenin A accumulation appeared to be overcome by degradationprocesses, and the disease spread (Bennett et al. 1994). These indicate that differentcompounds have different antifungal mechanisms and/or the sensitivity of the fungi maychange with different stages of development. Some small phenolic compounds disruptfungal membranes and attach and polymerise within the wall of the fungal hyphae, thusreducing the plasticity of the fungus for growth (Wang et al. 1994). Gallic acid and methylgallate can act on a cAMP-related signalling pathway regulating appressorium formationin Magnaporthe grisea (Ahn et al. 2005).
In conclusion, almost every antioxidant used in the present study exerted significantinhibitory effects on B. cinerea progress, and their effects were significantly linear, exceptfor ellagic acid which was quadratic, however, different antioxidants have differentantifungal mechanisms and the sensitivity of the B. cinerea to the antioxidants may changeat different stages of development. The objective of future work is to test the combinationeffect of selected antioxidants and determine if there is an interaction between the selectedcompounds on gray mold growth and development.
The observed results will help to select lines in vitro in order to select lines resistant togray mold.
Acknowledgements
The authors would like to acknowledge the support received from ABIP-100 project entitled:Nutraceuticals Emerging from Agricultural Technologies Network (NEATNet) and ChinaScholarship Council (CSC) and International Scientific Cooperation Bureau (ISCB) of Agriculture
Archives of Phytopathology and Plant Protection 1575
Downloaded By: [Khanizadeh, Shahrokh] At: 16:29 15 November 2010

and AgriFood Canada (AAFC). The authors also wish to thank Audrey Levasseur and MathieuTremblay from the Horticultural Research and Development Centre, Agriculture and Agri-FoodCanada for their technical assistance with this work.
References
Aaby K, Skrede G, Wrolstad RE. 2005. Phenolic composition and antioxidant activities in flesh andachenes of strawberries (Fragaria ananassa). J Agr Food Chem. 53:4032–4040.
Agrios GN. 1997. Plant pathology, 4th ed. San Diego (CA): Academic Press.Ahn Y, Lee H, Oh H, Kim H, Lee Y. 2005. Antifungal activity and mode of action of Galla rhois-
derived phenolics against phytopathogenic fungi. Pestic Biochem Phys. 81:105–112.Ayala-Zavala JF, Wang SY, Wang CY, Gonzalez-Aguilar GA. 2004. Effect of storage temperatures
on antioxidant capacity and aroma compounds in strawberry fruit. LWT-Food Sci Technol.37:687–695.
Bennett MH, Gallagher MDS, Bestwick CS, Rossiter JT, Mansfield JW. 1994. The phytoalexinresponse of lettuce to challenge by Botrytis cinerea, Bremia lactucae and Pseudomonas syringaepv. phaseolicola. Physiological and Molecular Plant Pathology. 44(5):321–333.
Cordenunsi BR, Genovese MI, Nascimento JRO, Hassimotto NMA, Santos RJ, Lajolo FM. 2005.Effects of temperature on the chemical composition and antioxidant activity of three strawberrycultivars. Food Chem. 91:113–121.
Daferera DJ, Ziogas BN, Polissiou MG. 2003. The effectiveness of plant essential oils on the growthof Botrytis cinerea, Fusarium sp. and Lavibacter michiganensis subsp. michiganensis. Crop Prot.22:39–44.
Ehsani-Moghaddam B, Khanizadeh S, Sullivan JA. 2008. Biochemical components of advancedselections and raspberry cultivars. Can. J. Plant Sci. 88(1):175–177.
Elad Y, Evensen K. 1995. Physiological aspects of resistance to Botrytis cinerea. Phytopathology.85(6):637–643.
Ferreyra RM, Vina SZ, Mugridge A, Chaves AR. 2007. Growth and ripening season effects onantioxidant capacity of strawberry cultivar Selva. Sci Hortic-Amsterdam. 112(1):27–32.
Feucht W, Treutter D, Christ E. 1992. The precise localization of catechins and proanthocyanidinsin protective layers around fungal infections. J Plant Dis Prot. 99(4):404–413.
Gil MI, Holcroft DM, Kader AA. 1997. Changes in strawberry anthocyanins and other polyphenolsin response to carbon dioxide treatments. J Agric Food Chem. 45:1662–1667.
Guo CJ, Yang JJ, Wei JY, Li YF, Xu J, Jiang YG. 2003. Antioxidant activities of peel, pulp and seedfractions of common fruits as determined by FRAP assay. Nutr Res. 23:1719–1726.
Hebert C, Charles MT, Gauthier L, Willemot C, Khanizadeh S, Cousineau J. 2002. Strawberryproanthocyanidins: biochemical markers for Botrytis cinerea resistance and shelf-life predict-ability. Acta Hort. 567:659–662.
Heinonen IM, Meyer AS, Frankel EN. 1998. Antioxidant activity of berry phenolics on human low-density lipoprotein and liposome oxidation. J Agric Food Chem. 46:4107–4112.
Heo HJ, Lee CY. 2005. Strawberry and its anthocyanins reduce oxidative stress-induced apoptosis inPC12 cells. J Agric Food Chem. 53:1984–1989.
Hernanz D, Recamales AF, Melendez-Martinze AJ, Gonzalez-Miret ML, Heredia FJ. 2007.Assessment of the difference in the phenolic composition of five strawberry cultivars (Fragaria xananassa Duch) grown in two different soil less systems. J Agric Food Chem. 55:1846–1852.
Hur JS, Oh SO, Jung JS, Koh YJ, Park JG, Park JC. 2003. Antifungal properties of Eucalyptusdarlympleana against postharvest pathogens of kiwifruits. ISHS Acta Hort 610: V InternationalSymposium on Kiwifruit. p. 425–431.
Khanam NN, Ueno M, Kihara J, Honda Y, Arase S. 2005. Suppression of red light-inducedresistance in broad beans to Botrytis cinerea by salicylic acid. Physiol Mol Plant Pathol. 66:20–29.
Khanizadeh S, Behrouz EM, Levasseur A. 2006. Antioxidant capacity in June-bearing and day-neutral strawberry. Can J Plant Sci. 86:1387–1390.
Khanizadeh S, Cousineau J, Deschenes M, Levasseur A, Carisse O, DeEll J, Gauthier L, Sullivan A.2002a. ‘Saint-Pierre’ Strawberry. HortScience. 37(7):1133–1134.
Khanizadeh S, Cousineau J, Gauthier L, Buszard D, Hebert C. 2002b. ‘‘L’ Authentique Orleans’’: Anew strawberry cultivar with high level of antioxidant. Acta Hort. 567:175–176.
1576 S. Tao et al.
Downloaded By: [Khanizadeh, Shahrokh] At: 16:29 15 November 2010

Khanizadeh S, Deschenes M, Levasseur A, Carisse O, Charles MT, Rekika D, Tsao R, Yang R,DeEll J, Thibeault P, Prive JP, Davidson C, Bors B. 2005a. ‘Harmonie’ strawberry. HortScience.40:480–481.
Khanizadeh S, Deschenes M, Levasseur A, Carisse O, Charles MT, Rekika D, Tsao R, Yang R,DeEll J, Gauthier L, Gosselin A, Sullivan JA, Davidson C. 2005b. ‘Saint-Laurent d’Orleans’strawberry. HortScience. 40(7):2195–2196.
Khanizadeh S, Deschenes M, Levasseur A, Carisse O, Charles MT, Rekika D, Tsao R, Yang R,DeEll J, Gauthier L. 2005c. Saint-Laurent d’Orleans strawberry. HortScience. 40:2195–2196.
Khanizadeh S, Deschenes M, Levasseur A, Carisse O, Charles MT, Rekika D, Gauthier L, GosselinA, Tsao R, Yang R, DeEll J, Sullivan JA. 2007. ‘St-Jean d’Orleans’ Strawberry. HortScience.42(1):168–169.
Khanizadeh S, Tao S, Zhang S, Tsao R, Rekika D, Yang R, Charles MT. 2008a. Antioxidantactivities of newly developed day-neutral and June-bearing strawberry lines. J Food AgrEnviron. 6(2):306–311.
Khanizadeh S, Tsao R, Rekika D, Yang R, Charles MT, Rupasinghe V. 2008b. Polyphenolcomposition and total antioxidant capacity of selected apple genotypes for processing. J FoodComposition Analysis. 21(5):396–401.
Klopotek Y, Otto K, Bohm V. 2005. Processing strawberries to different products alters contents ofvitamin C, total phenolics, total anthocyanins, and antioxidant capacity. J Agric Food Chem.53:5640–5646.
Kosar M, Kafkas E, Paydas S, Baser KHS. 2004. Phenolic composition of strawberry genotypes atdifferent maturation stages. J Agric Food Chem. 52:1586–1589.
Lattanzio V, Venere DD, Linsalata V, Bertolini P, Ippolito A, Salerno M. 2001. Low temperaturemetabolism of apple phenolics and quiescence of Phlyctaena vagabunda. J Agric Food Chem.49:5817–5821.
Lee SH, Chang KS, Su MS, Huang YS, Jang HD. 2007. Effects of some Chinese medicinal plantextract on five different fungi. Food Control. 18(12):1547–1554.
Li BJ, Chen LQ, Meng WJ, Wang FJ. 2003. Effects of humidity regulation on the infection ofBotrytis cinerea to tomato. Acta Phytopathol Sin. 33(2):167–169.
Liu J, Tian S, Meng X, Xu Y. 2007. Effects of chitosan on control of postharvest diseases andphysiological responses of tomato fruit. Postharvest Biol Technol. 44(3):300–306.
Martınez-Romero D, Guillen F, Valverde JM, Bailen G, Zapata G, Serrano M, Castillo S, Valero D.2007. Influence of carvacrol on survival of Botrytis cinerea inoculated in table grapes. Int J FoodMicrobiol. 115(2):144–148.
Meyers KJ, Watkins CB, Pritts MP, Liu RH. 2003. Antioxidant and antiproliferative activities ofstrawberries. Am J Enol Viticult. 51:6887–6892.
Nicholson RL, Hammerschmidt R. 1992. Phenolic compounds and their role in disease resistance.Annu Rev Phytopathol. 30:369–389.
Pinto MDS, Lajolo FM, Genovese MI. 2007. Bioactive compounds and antioxidant capacity ofstrawberry jams. Plant Food Hum Nutr. 62(3):127–131.
Prusky D. 1996. Pathogen quiescence in postharvest disease. Annu Rev Phytopathol. 34:413–434.Raposo R, Colgan R, Delcan J, Melgarejo P. 1995. Application of an automated quantitative
method to determine fungicide resistance in Botrytis cinerea. Plant Dis. 79(3):294–296.Rekika D, Khanizadeh S, Deschenes M, Levasseur A, Charles MT. 2005. Atioxidant capacity and
phenolic content of selected strawberry genotypes. HortScience. 40(6):1777–1781.Scalzo J, Politi A, Pellegrini N, Mezzetti B, Battino M. 2005. Plant genotype affects total antioxidant
capacity and phenolic contents in fruit. Nutrition. 21:207–213.Soylu S. 2006. Accumulation of cell-wall bound phenolic compounds and phytoalexin in
Arabidopsis thaliana leaves following inoculation with pathovars of Pseudomonas syringae.Plant Sci. 170:942–952.
SAS Institute. 1989. SAS user’s guide: statistics. Version 6. Cary (NC): SAS Institute.Steel RGD, Torrie JH. 1980. Principles and procedures of statistics, 2nd ed. New York: McGraw-
Hill.Terry LA, Joyce DC, Adikaram NKB, Khambay BPS. 2004. Preformed antifungal compounds in
strawberry fruit and flower tissues. Postharvest Biol Technol. 31(2):201–212.Tremblay DM, Talbot BG, Carisse O. 2003. Sensitivity of Botrytis squamosa to different classes of
fungicides. Plant Dis. 87:573–578.
Archives of Phytopathology and Plant Protection 1577
Downloaded By: [Khanizadeh, Shahrokh] At: 16:29 15 November 2010

Vicente AR, Martınez GA, Chaves AR, Civello PM. 2006. Effect of heat treatment on strawberryfruit damage and oxidative metabolism during storage. Postharvest Biol Technol. 40:116–122.
Wang H, Cao G, Prior RL. 1996. Total antioxidant capacity of fruits. J Agric Food Chem. 44:701–705.
Wang QL, Khanizadeh S, Vigneault C. 2007. Preharvest ways of enhancing the phytochemicalcontent of fruits and vegetables. Stewart Postharvest Rev. 3(3):3.1–3.8.
Wang SY, Lin HS. 2000. Antioxidant activity in fruits and leaves of blackberry, raspberry, andstrawberry varies with cultivar and developmental stage. J Agric Food Chem. 48:140–146.
Wang SY, Lin HS. 2003. Compost as a soil supplement increases the level of antioxidant compoundsand oxygen radical absorbance capacity in strawberries. J Agric Food Chem. 51:6844–6850.
Wang SY, Mass JL, Payne JA, Galletta GJ. 1994. Ellagic acid content in small fruits, mayhaws, andother plants. J Small Fruit Viticult. 2(4):39–49.
Wang SY, Zhang W. 2001. Effect of plant growth temperature on antioxidant capacity instrawberry. J Agric Food Chem. 49:4977–4982.
Wang SY, Zhang W, Galletta GJ. 2002. Culture system affects fruit quality and antioxidant capacityin strawberries. J Agric Food Chem. 50(22):6534–6542.
1578 S. Tao et al.
Downloaded By: [Khanizadeh, Shahrokh] At: 16:29 15 November 2010
Related Documents











