In silico identification of novel protective VSG antigens expressed by Trypanosoma brucei and an effort for designing a highly immunogenic DNA vaccine using IL-12 as adjuvant Bashir Akhlaq Akhoon a, * ,1 , Parvez Singh Slathia a,1 , Preeti Sharma b , Shishir Kumar Gupta c , Vijeshwar Verma a a Centre of Bioinformatics, School of Biotechnology, Shri Mata Vaishno Devi (SMVD) University, Jammu, J&K 182320, India b Microbiology laboratory, School of Biotechnology, Shri Mata Vaishno Devi (SMVD) University, Jammu, J&K 182320, India c Society for Biological Research & Rural Development, Lucknow, Uttar Pradesh, India article info Article history: Received 17 August 2010 Received in revised form 24 January 2011 Accepted 24 January 2011 Available online 22 February 2011 Keywords: Trypanosoma brucei Antigenic variability Vaccine candidates African trypanosomiasis Immunoinformatics abstract African trypanosomiasis continues to be a major health problem, with more adults dying from this disease world-wide. As the sequence diversity of Trypanosoma brucei is extreme, with VSGs having 15 e25% identity with most other VSGs, hence it displays a huge diversity of adaptations and host speci- ficities. Therefore the need for an improved vaccine has become an international priority. The highly conserved and specific epitopes acting as both CD8 þ and CD4 þ T-cell epitopes (FLINKKPAL and FTALCTLAA) were predicted from large bunch of VSGs of T. brucei. Besides, some other potential epitopes with very high affinity for MHC I and II molecules were also determined while taking consideration on the most common HLA in the general population which accounts for major ethnicities. The vaccine candidates were found to be effective even for non-african populations as predicted by population coverage analysis. Hence the migrating travelers acting as a spread means of the infection can probably also be treated successfully after injection of such a multiepitopic vaccine. Exploiting the immu- noinformatics approaches, we designed a potential vaccine by using the consensus epitopic sequence of 388 VSG proteins of T. brucei and performed in silico cloning of multiepitopic antigenic DNA sequence in pBI-CMV1 vector. Moreover, various techniques like codon adaptation, CpG optimization, removal of self recognized epitopes, use of adjuvant and co-injection with plasmids expressing immune-stimulatory molecules were implemented to enhance the immunogenicity of the proposed in silico vaccine. Ó 2011 Elsevier Ltd. All rights reserved. 1. Introduction Parasitic protozoa continue to pose major threats to human health and animals of veterinary importance. Human African Trypanosomiasis (HAT) or African trypanosomiasis has been endemic in sub-Saharan Africa for thousands of years. Currently approximately 50 million people are at risk for this disease in an area of 10 million square kilometers [1]. The number of non- reported cases is probably much higher. Cases of HAT (about 50 cases annually) have also been reported outside Africa as a result of travelers or workers returning to North America or Europe [2]. Trypanosoma brucei is the causative agent of sleeping sickness in humans. The disease has a severe impact on humans [3] and is fatal if left untreated. African trypanosomiasis is represented in humans by a complex neuropsychiatric syndrome caused by T. brucei (subspecies rhodesiense and gambiense) [4]. This parasitic infection is hallmarked by disturbances of the sleep/wake (S/W) cycle and sleep pattern [4e6]. Due to sleep changes, the disease is also designated with the alternative name, sleeping sickness [7]. The extracellular African trypanosomes are directly exposed to the host immune system. T. brucei parasites subsist extracellularly in the blood and tissues of their mammalian hosts and are able to evade the humoral immune response by means of antigenic variation of the major surface antigen: the membrane form of the variant surface glycoproteins (VSGs). This creates a physical barrier that protects the parasite against the effector mechanisms of the * Corresponding author. Present address: BIF Centre, School of Biotechnology, Shri Mata Vaishno Devi University (SMVDU) Campus, Sub P.O. SMVD University, Katra 182320 (J&K), India. Tel.: þ91 9906751249. E-mail addresses: [email protected] (B.A. Akhoon), parvezsingh@gmail. com (P.S. Slathia), [email protected] (P. Sharma), [email protected] (S.K. Gupta), [email protected] (V. Verma). 1 Both the authors contribute equally. Contents lists available at ScienceDirect Microbial Pathogenesis journal homepage: www.elsevier.com/locate/micpath 0882-4010/$ e see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.micpath.2011.01.011 Microbial Pathogenesis 51 (2011) 77e87

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

lable at ScienceDirect
Microbial Pathogenesis 51 (2011) 77e87
Contents lists avai
Microbial Pathogenesis
journal homepage: www.elsevier .com/locate/micpath
In silico identification of novel protective VSG antigens expressed byTrypanosoma brucei and an effort for designing a highlyimmunogenic DNA vaccine using IL-12 as adjuvant
Bashir Akhlaq Akhoon a,*,1, Parvez Singh Slathia a,1, Preeti Sharma b,Shishir Kumar Gupta c, Vijeshwar Verma a
aCentre of Bioinformatics, School of Biotechnology, Shri Mata Vaishno Devi (SMVD) University, Jammu, J&K 182320, IndiabMicrobiology laboratory, School of Biotechnology, Shri Mata Vaishno Devi (SMVD) University, Jammu, J&K 182320, Indiac Society for Biological Research & Rural Development, Lucknow, Uttar Pradesh, India
a r t i c l e i n f o
Article history:Received 17 August 2010Received in revised form24 January 2011Accepted 24 January 2011Available online 22 February 2011
Keywords:Trypanosoma bruceiAntigenic variabilityVaccine candidatesAfrican trypanosomiasisImmunoinformatics
* Corresponding author. Present address: BIF CenShri Mata Vaishno Devi University (SMVDU) CampuKatra 182320 (J&K), India. Tel.: þ91 9906751249.
E-mail addresses: [email protected] (B.A. Acom (P.S. Slathia), [email protected] (P. Sharma(S.K. Gupta), [email protected] (V. Verma).
1 Both the authors contribute equally.
0882-4010/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.micpath.2011.01.011
a b s t r a c t
African trypanosomiasis continues to be a major health problem, with more adults dying from thisdisease world-wide. As the sequence diversity of Trypanosoma brucei is extreme, with VSGs having 15e25% identity with most other VSGs, hence it displays a huge diversity of adaptations and host speci-ficities. Therefore the need for an improved vaccine has become an international priority. The highlyconserved and specific epitopes acting as both CD8þ and CD4þ T-cell epitopes (FLINKKPAL andFTALCTLAA) were predicted from large bunch of VSGs of T. brucei. Besides, some other potential epitopeswith very high affinity for MHC I and II molecules were also determined while taking consideration onthe most common HLA in the general population which accounts for major ethnicities. The vaccinecandidates were found to be effective even for non-african populations as predicted by populationcoverage analysis. Hence the migrating travelers acting as a spread means of the infection can probablyalso be treated successfully after injection of such a multiepitopic vaccine. Exploiting the immu-noinformatics approaches, we designed a potential vaccine by using the consensus epitopic sequence of388 VSG proteins of T. brucei and performed in silico cloning of multiepitopic antigenic DNA sequence inpBI-CMV1 vector. Moreover, various techniques like codon adaptation, CpG optimization, removal of selfrecognized epitopes, use of adjuvant and co-injection with plasmids expressing immune-stimulatorymolecules were implemented to enhance the immunogenicity of the proposed in silico vaccine.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Parasitic protozoa continue to pose major threats to humanhealth and animals of veterinary importance. Human AfricanTrypanosomiasis (HAT) or African trypanosomiasis has beenendemic in sub-Saharan Africa for thousands of years. Currentlyapproximately 50 million people are at risk for this disease in anarea of 10 million square kilometers [1]. The number of non-reported cases is probably much higher. Cases of HAT (about 50
tre, School of Biotechnology,s, Sub P.O. SMVD University,
khoon), parvezsingh@gmail.), [email protected]
All rights reserved.
cases annually) have also been reported outside Africa as a result oftravelers or workers returning to North America or Europe [2].Trypanosoma brucei is the causative agent of sleeping sickness inhumans. The disease has a severe impact on humans [3] and is fatalif left untreated. African trypanosomiasis is represented in humansby a complex neuropsychiatric syndrome caused by T. brucei(subspecies rhodesiense and gambiense) [4]. This parasitic infectionis hallmarked by disturbances of the sleep/wake (S/W) cycle andsleep pattern [4e6]. Due to sleep changes, the disease is alsodesignated with the alternative name, sleeping sickness [7]. Theextracellular African trypanosomes are directly exposed to the hostimmune system. T. brucei parasites subsist extracellularly in theblood and tissues of their mammalian hosts and are able to evadethe humoral immune response by means of antigenic variation ofthe major surface antigen: the membrane form of the variantsurface glycoproteins (VSGs). This creates a physical barrier thatprotects the parasite against the effector mechanisms of the

Table 1Fragments selected for epitope prediction.
Fragment Length
AVLLLLLLLSAAAAAA 16GDNAAAFTALCTLAA 15AAAAAAAAAAYLAG 14LAALRALAKAAVAATAAAAA 20ATAACAALTEALYG 14ALAADLACLCAG 12AALAALKAAIKAATTD 16ADGACVKYSA 10ALKAKLKALLGTAKALYKAAF 21CKDSSFLINKKPALL 15
B.A. Akhoon et al. / Microbial Pathogenesis 51 (2011) 77e8778
immune system [8,9]. VSG molecules are immunodominant anti-gens that elicit B- and T-cell responses capable of providingtemporal protection for the host during infection [10e13] and suchVSGs undergoes antigenic variation, enabling the parasite to escapeimmune reactions. The overwhelming production of immuno-globulins, only partially directed against trypanosome antigens,added to the release of VSG fragments in the blood, contributes todisease immunopathogenicity [14]. Trypanosomes can activatemacrophages directly, through endotoxin-like substances of thefamily of coat lipopolysaccharides and the GPI anchor of VSG [15].This activation is enhanced by interferon-g (IFN-g). Interestingly, T-cells are triggered to produce IFN-g by a microtubule-bindingprotein trypanin [16,17], which in turn could be a growthpromoting signal for trypanosomes [18]. The immune responseelicited by infections can be predominantly of type Th1 or Th2,depending on the cytokines mainly produced. The treatment usedto combat HAT is not satisfactory because the drugs used for thetreatment are potentially toxic and often associated with the deathof patients in treatment [19]. Invariant surface glycoprotein (ISG)antigen candidate might be a promising antigen because it wasreported to be highly immunogenic, bloodstream-stage specificand expressed as a surface protein in the bloodstream forms ofT. brucei [20,21]. Indeed, it is generally accepted that DNA vaccineshave a significant Th1 bias [22,23]. Also, reorientation of theimmune response has been observed for DNA vaccine therapyagainst Leishmania and Mycobacterium infections [24,25]. Consid-ering the cellular complexity of T. brucei, it is believed that morethan one antigenic candidate may be necessary for an efficientvaccine preparation against HAT. Thus, to characterize antigenictargets in the context of DNA vaccination could be a rationalstrategy for vaccine development for this ailment [26].
2. Results and discussion
In the post-genomic era, strategies of vaccine development haveprogressed dramatically from traditional Pasteur’s principles ofisolating, inactivating and injecting the causative agent of aninfectious disease, to reverse vaccinology that starts from bio-informatics analysis of the genome information. Vaccination iswidely regarded as one of the most successful public health inter-vention measures in the fight against infectious diseases, allergies,neurodegenerative diseases, autoimmune disorders and somecancers [27]. An unguided experimental search for antigenic andimmunogenic regions is inherently laborious and resource inten-sive [28] and techniques used in this report have the potential tosimplify the process greatly. With an ever-increasing number ofpathogen genomes fully or partially sequenced there is a pressingneed for in silico methods that can identify potential vaccinecandidates. For a vaccine to be effective it must invoke a strongresponse from both T-cells and B cells; therefore, epitope mappingis a central issue in their design [29,30]. The traces of generatinghumoral responses by T-cell epitopes have also been found byimmunologists [31,32].
The feasible solution to prevent sleeping sickness in futureagainst T. brucei can be done by the development of new prophy-lactic DNA vaccine which can work against almost all different VSGvariants of T. brucei. In this research, we proceed to designcomputational DNA vaccine, taking VSG sequence variants ofdifferent strains of T. brucei. Antigenic variability is the majorobstacle for the development of urgently needed vaccines and toaddress this problem the VSG sequences were selected from almostall species of T. brucei available at NCBI and were subjected to MSA.Constructing a multiple alignment corresponds to developinga hypothesis of how a number of sequences have evolved throughthe processes of character substitution, insertion and deletion.
ClustalW was selected as an alignment method which uses a seriesof weight tables as the alignment progresses and here Gonnet 250was chosen as Protein Weight Matrix. The software can be run intwo modes: Fast-Approx. versus Slow-Accurate and to achieveoptimal MSA Slow-Accurate option was preferred. The resultedalignment reflected amino acid conservancy of VSG proteins andten conserved/consensus (Table 1) regions (�9-mer) were finallyselected for epitope prediction.
To view evolutionary relationships of VSG proteins predictedfrom the multiple sequence alignment a phylogenetic tree wasconstructed. A popular way of evaluating the reliability of aninferred phylogenetic tree is boot strap analysis and a high bootstrap score is a sign of greater reliability. This score indicates thenumber of times to compare the displayed tree with random treeconstructs that use the neighbor-joining methodology. The higherthe number, the better the result and the typical range is from 100to 1000. The maximum (1000) possible trails were selected forresampling and the phylogenetic tree of T. brucei VSGs is shown inFig. 1. To make the clear visualization of the tree, we grouped thesub-trees into 18 clusters on the basis of their evolutionarydistance. We observed that the 3 VSGs: CAA28882.1, AAA30274.1and AAB30175.1 don’t fall into any of the clusters. The cluster 18(red color) consisting of 2 VSG sequences with accession no.AAA63550.1 and CBH13985.1 was noticed to be enormouslydivergent from the rest 386 VSGs of T. brucei.
In silico work is the boon for experimental biologist for rapidscreening and identification of probable vaccine candidates [33].HLA class-I molecules (HLA-B, -C, -A) present foreign antigens toCD8þ CTLs and HLA class-II molecules (HLA-DP, DQ, DR) presentantigenic peptides to CD4þ helper T-lymphocytes. We tried toretrieve almost all available (388) VSGs of T. brucei at NCBI for theconservancy analysis of amino acid residues to minimize the anti-genic drift of the selected epitopes as vaccine candidates. All theVSGs selected for this analysis have been provided in the supple-mentary material. IEDB (ANN) was used to access the predictions ofpeptide binding to MHC class-I molecules. We defined predictedIC50 [nM] cutoff field as 50 to report only best epitopes. Besides,NetMHC 3.2 server which predicts binding of peptides to a numberof different HLA alleles using artificial neural networks (ANNs) wasused for the prediction of MHC class-I epitopes. ANNs have beentrained for 57 different Human MHC (HLA) alleles representing all12 HLA A and B Supertypes [34]. Only one strongest 9-mer bindingpeptide with lowest nM IC50 values was selected for each allele.Epitope Conservancy Analysis tool of IEDB was used to filter theidentical epitopes and to make the final selection of epitopes(Table 2a). The degree of conservation is defined as the fraction ofpeptide sequences containing the epitope at a given identity leveland we have chosen sequence identity threshold “�100” to displaycalculated conservancies. NetMHCII 2.2 and NetMHCIIpan 2.0servers were used to predict consensus 9-mers binding epitopes.NetMHCIIpan server predicts binding of peptides to more than 500

Fig. 1. (a) Evolutionary tree of 388 sequence variants of Trypanosoma brucei VSG proteins. The sequence divergence is shown by color coding. The numeric digits at terminal pointsdenote the number of sequences in that cluster (subtree). (b) Subgroups of each cluster showing the phylogenetic relationship of VSGs. The length of each pair of branchesrepresents the distance between sequence pairs, while the numbers at the end points indicate the accession numbers of the selected VSG sequences.
B.A. Akhoon et al. / Microbial Pathogenesis 51 (2011) 77e87 79
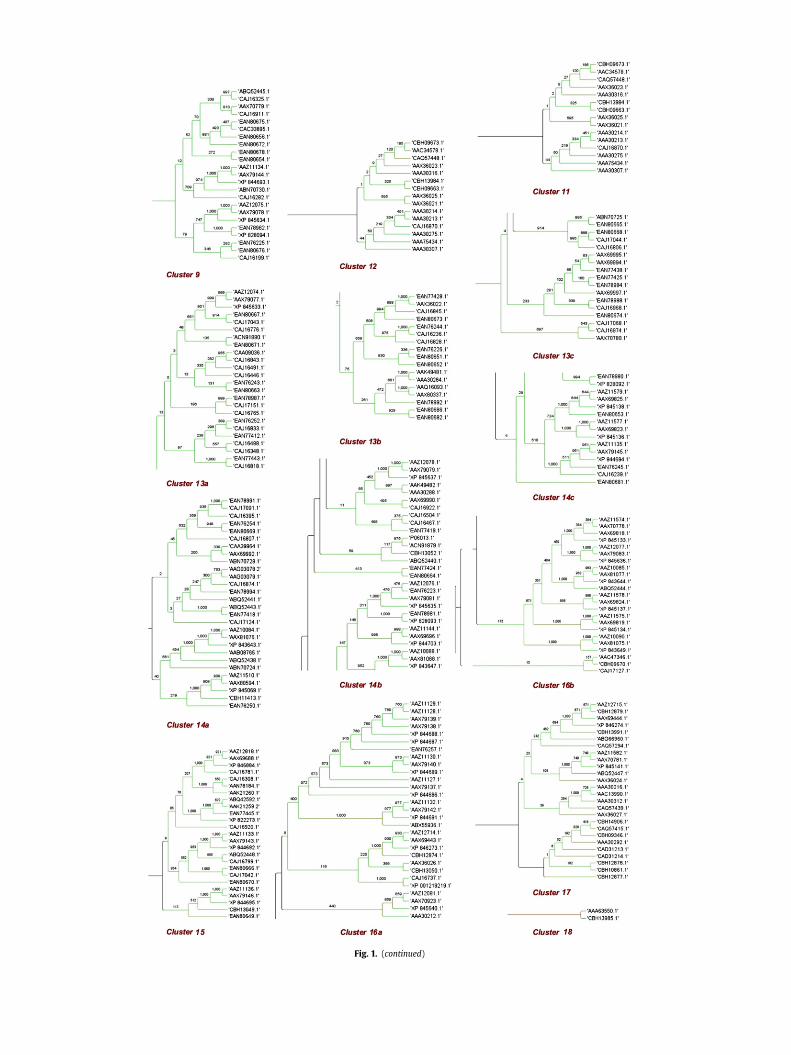
Fig. 1. (continued)
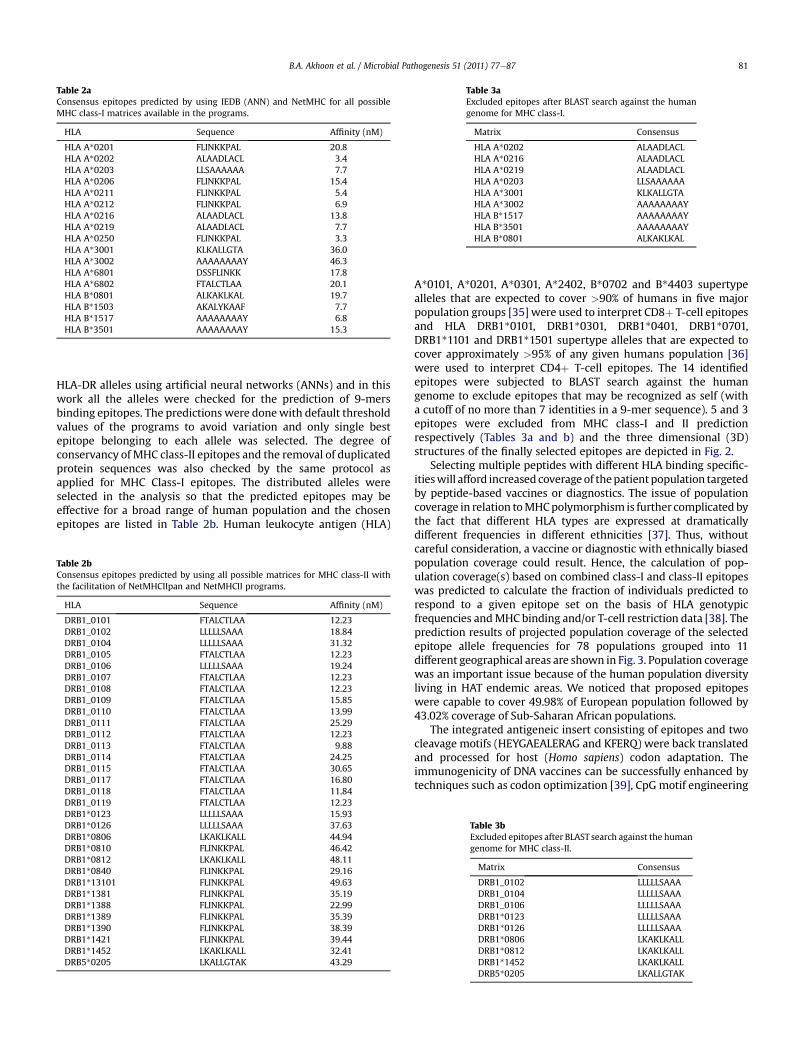
Table 2aConsensus epitopes predicted by using IEDB (ANN) and NetMHC for all possibleMHC class-I matrices available in the programs.
HLA Sequence Affinity (nM)
HLA A*0201 FLINKKPAL 20.8HLA A*0202 ALAADLACL 3.4HLA A*0203 LLSAAAAAA 7.7HLA A*0206 FLINKKPAL 15.4HLA A*0211 FLINKKPAL 5.4HLA A*0212 FLINKKPAL 6.9HLA A*0216 ALAADLACL 13.8HLA A*0219 ALAADLACL 7.7HLA A*0250 FLINKKPAL 3.3HLA A*3001 KLKALLGTA 36.0HLA A*3002 AAAAAAAAY 46.3HLA A*6801 DSSFLINKK 17.8HLA A*6802 FTALCTLAA 20.1HLA B*0801 ALKAKLKAL 19.7HLA B*1503 AKALYKAAF 7.7HLA B*1517 AAAAAAAAY 6.8HLA B*3501 AAAAAAAAY 15.3
Table 3aExcluded epitopes after BLAST search against the humangenome for MHC class-I.
Matrix Consensus
HLA A*0202 ALAADLACLHLA A*0216 ALAADLACLHLA A*0219 ALAADLACLHLA A*0203 LLSAAAAAAHLA A*3001 KLKALLGTAHLA A*3002 AAAAAAAAYHLA B*1517 AAAAAAAAYHLA B*3501 AAAAAAAAYHLA B*0801 ALKAKLKAL
B.A. Akhoon et al. / Microbial Pathogenesis 51 (2011) 77e87 81
HLA-DR alleles using artificial neural networks (ANNs) and in thiswork all the alleles were checked for the prediction of 9-mersbinding epitopes. The predictions were donewith default thresholdvalues of the programs to avoid variation and only single bestepitope belonging to each allele was selected. The degree ofconservancy of MHC class-II epitopes and the removal of duplicatedprotein sequences was also checked by the same protocol asapplied for MHC Class-I epitopes. The distributed alleles wereselected in the analysis so that the predicted epitopes may beeffective for a broad range of human population and the chosenepitopes are listed in Table 2b. Human leukocyte antigen (HLA)
Table 2bConsensus epitopes predicted by using all possible matrices for MHC class-II withthe facilitation of NetMHCIIpan and NetMHCII programs.
HLA Sequence Affinity (nM)
DRB1_0101 FTALCTLAA 12.23DRB1_0102 LLLLLSAAA 18.84DRB1_0104 LLLLLSAAA 31.32DRB1_0105 FTALCTLAA 12.23DRB1_0106 LLLLLSAAA 19.24DRB1_0107 FTALCTLAA 12.23DRB1_0108 FTALCTLAA 12.23DRB1_0109 FTALCTLAA 15.85DRB1_0110 FTALCTLAA 13.99DRB1_0111 FTALCTLAA 25.29DRB1_0112 FTALCTLAA 12.23DRB1_0113 FTALCTLAA 9.88DRB1_0114 FTALCTLAA 24.25DRB1_0115 FTALCTLAA 30.65DRB1_0117 FTALCTLAA 16.80DRB1_0118 FTALCTLAA 11.84DRB1_0119 FTALCTLAA 12.23DRB1*0123 LLLLLSAAA 15.93DRB1*0126 LLLLLSAAA 37.63DRB1*0806 LKAKLKALL 44.94DRB1*0810 FLINKKPAL 46.42DRB1*0812 LKAKLKALL 48.11DRB1*0840 FLINKKPAL 29.16DRB1*13101 FLINKKPAL 49.63DRB1*1381 FLINKKPAL 35.19DRB1*1388 FLINKKPAL 22.99DRB1*1389 FLINKKPAL 35.39DRB1*1390 FLINKKPAL 38.39DRB1*1421 FLINKKPAL 39.44DRB1*1452 LKAKLKALL 32.41DRB5*0205 LKALLGTAK 43.29
A*0101, A*0201, A*0301, A*2402, B*0702 and B*4403 supertypealleles that are expected to cover >90% of humans in five majorpopulation groups [35] were used to interpret CD8þ T-cell epitopesand HLA DRB1*0101, DRB1*0301, DRB1*0401, DRB1*0701,DRB1*1101 and DRB1*1501 supertype alleles that are expected tocover approximately >95% of any given humans population [36]were used to interpret CD4þ T-cell epitopes. The 14 identifiedepitopes were subjected to BLAST search against the humangenome to exclude epitopes that may be recognized as self (witha cutoff of no more than 7 identities in a 9-mer sequence). 5 and 3epitopes were excluded from MHC class-I and II predictionrespectively (Tables 3a and b) and the three dimensional (3D)structures of the finally selected epitopes are depicted in Fig. 2.
Selecting multiple peptides with different HLA binding specific-itieswill afford increased coverage of thepatientpopulation targetedby peptide-based vaccines or diagnostics. The issue of populationcoverage in relation toMHCpolymorphism is further complicated bythe fact that different HLA types are expressed at dramaticallydifferent frequencies in different ethnicities [37]. Thus, withoutcareful consideration, a vaccine or diagnostic with ethnically biasedpopulation coverage could result. Hence, the calculation of pop-ulation coverage(s) based on combined class-I and class-II epitopeswas predicted to calculate the fraction of individuals predicted torespond to a given epitope set on the basis of HLA genotypicfrequencies andMHC binding and/or T-cell restriction data [38]. Theprediction results of projected population coverage of the selectedepitope allele frequencies for 78 populations grouped into 11different geographical areas are shown in Fig. 3. Population coveragewas an important issue because of the human population diversityliving in HAT endemic areas. We noticed that proposed epitopeswere capable to cover 49.98% of European population followed by43.02% coverage of Sub-Saharan African populations.
The integrated antigeneic insert consisting of epitopes and twocleavage motifs (HEYGAEALERAG and KFERQ) were back translatedand processed for host (Homo sapiens) codon adaptation. Theimmunogenicity of DNA vaccines can be successfully enhanced bytechniques such as codon optimization [39], CpG motif engineering
Table 3bExcluded epitopes after BLAST search against the humangenome for MHC class-II.
Matrix Consensus
DRB1_0102 LLLLLSAAADRB1_0104 LLLLLSAAADRB1_0106 LLLLLSAAADRB1*0123 LLLLLSAAADRB1*0126 LLLLLSAAADRB1*0806 LKAKLKALLDRB1*0812 LKAKLKALLDRB1*1452 LKAKLKALLDRB5*0205 LKALLGTAK
Fig. 2. Optimal three dimensional structures of the selected epitopes.
B.A. Akhoon et al. / Microbial Pathogenesis 51 (2011) 77e8782
[40,41] and the introduction of promoter sequences [42,43]. Duringcodon optimization, a total of 10 codons were replaced in class-IMHC sequence and that of 3 in class-II MHC sequence. CpG opti-mization, can be used to optimize the codons with respect to CGdinucleotides and pattern recognition receptors that form part ofthe innate immune system can often distinguish prokaryotic DNA
Fig. 3. Graph shows the fraction of individuals predicted to respond to a given epitopeset on the basis of HLA genotypic frequencies.
from eukaryotic DNA by detecting unmethylated CpG dinucleotidesin particular base contexts, termed ‘CpG motifs’ [44]. The CPGmotifs are commonly associated with the induction of polarizedTh1-cell immune response, characterized by IFN-g production andTh1-associated antibody isotypes (IgG2a, IgG2b, and IgG3) anda reduction in IgG1 production [45,46]. An important study ofKlinman (2004) showed that the CpG oligodeoxynucleotide (ODN)treatment significantly enhances resistance to T. b. rhodesiense insusceptible mouse strains [47]. Therefore, CpG optimization wasattained that replaced 48 codons in class-I MHC sequence and thatof 14 in class-II MHC sequence.
Gene cloning facilitates the insertion of antigenic DNA into pBI-CMV1 vector to design the desired vaccine. The DNA plasmid vectorvaccine contains the genetic material to produce an antigen,allowing that antigen to be transcribed in the host cell. This leads togenerate humoral and cell-mediated immune responses against thevector as well as inserted gene products. Both MHC class-I and/orMHC class-II antigen processing pathway can be targeted when weuse pBI-CMV1. Annotations of specific regions of prepared DNAvaccine is depicted in Fig. 4. The predominance of IL-12 or IL-4during the early stages of the infections, as well as several otherfactors, skews the immune response toward a Th1 or Th2 type,respectively [48,49]. The Th1 cytokine milieu contributes to theresistance to many intracellular parasites [48]. Hence, we admin-istrated both p40 and p35 subunits of IL-12 in to two differentcloning sites of pVITRO2- hygro-mcs. pVITRO is a family of plasmidsdeveloped mainly for in vitro studies that allow the ubiquitous andconstitutive co-expression of two genes of interest. pVITRO plas-mids can be stably transfected in mammalian cells and the genes ofinterest are expressed at high levels. pVITRO2- hygro-mcs plasmidis selectable in Escherichia coli and mammalian cells withHygromycin B/HygroGold�. It contains two multiple cloning sites(MCS) for the convenient cloning of two cDNAs. SV40 enhancerwhich is comprised of a 72-base-pair repeat allows the enhance-ment of gene expression in a large host range. The enhancementvaries from 2-fold in non-permissive cells to 20-fold in permissivecells [50]. The cDNA of p40 and p35 subunits were insertedbetween the two restriction sites Eco RV-Bam HI and Bgl IIeEcoRIrespectively. The various features of pVITRO2- hygro-mcs vectoralong with the incorporated cDNA of IL-12, used as an adjuvant toinduce the protective CD8þ -T-cell responses against T. brucei isshown in Fig. 5. The overall methodology used in this work mighthave a tremendous potential to accelerate vaccine development. Inrecent years, there have been significant advances in vaccinedevelopment against these parasites, and the recent completion ofthe sequencing of their genomes has opened new opportunities tofurther our understanding of the mechanisms required forprotection and the identification of novel antigens.
3. Materials and methods
3.1. Sequence selection and evolutionary conservation studies
Almost all available (388) sequence variants of T. brucei VSGproteins were collected by using the key word “variant surfaceglycoprotein [T. brucei]” from the Entrez protein database availableat NCBI (http://www.ncbi.nlm.nih.gov). All the sequences weresubjected to multiple sequence alignment (MSA) and conservationor majority of VSG amino acid residues in 388 sequences werenoticed by using MegAlign module of DNASTAR Lasergene soft-ware. ClustalW was selected as an alignment method and theconserved fragments of �9-mers were chosen for T-cell epitopeprediction. Phylogenetic tree was constructed using MegAlign tovisualize the evolutionary relationships among VSG sequences and

Fig. 4. Annotation of DNA vaccine map.
B.A. Akhoon et al. / Microbial Pathogenesis 51 (2011) 77e87 83
to test the statistical reliability of the phylogenetic tree; the treewas subjected to boot strap analyses with 1000 replications.
3.2. Prediction of MHC class-I epitopes
MHC class-I molecules deliver peptides from the cytosol and arerecognized by CD8þ T-cells. The T-cell epitopes presented by MHCclass-I molecules are typically peptides between 8 and 11 aminoacids in length. Gowthaman and coworkers (2010) reviewed theprediction efficiency of eight different in silico programs/serverswith the experimentally verified data for MHC class-I epitopes andreported that IEDB (ANN) followed by NetMHC have the bestoverall performance measures as evidenced by high and balancedsensitivity, specificity and accuracy values [51]. The predictionswere done with default threshold values of the programs to avoidvariation. Hence, we predicted 9-mer MHC I epitopes by using theabove two programs with default threshold values.
3.3. Prediction of MHC class-II epitopes
To predict the immune response against given pathogens,a number of methods have been developed to predict peptide-MHCbinding. Dimitrov and coworkers (2010) evaluated prediction effi-ciency of eight different in silico programs/servers for MHC class-IIepitopes and it was confirmed that NetMHCIIpan and NetMHCII
performed well than the rest tools [52]. Therefore, in this work wehave chosen these two servers for the prediction of 9-mer epitopes.
3.4. Removal of self immune epitopes and population coverageanalysis
Epitopes were evaluated for homology with human sequencesusing the BLAST algorithm [53]. Clusters with no more than 7matches per 9-mer frame were selected for further study and therest were discarded. Population coverage analysis plays significantrole for the designing of epitope-based vaccine as the MHC mole-cule is highly polymorphic [54e56]. Hence, population coverage ofnovel epitopes was predicted by the population coverage tool ofIEDB. The tool provides allele frequencies for 78 populationsgrouped into 11 different geographical areas.
3.5. Addition of motifs for target specification
Specific motifs were inserted within optimized sequences fortarget specific cleavage in proteasomal and lysosomal degrad-ation machineries. HEYGAEALERAG was added at the both eNH2andeCOOH termini of eachMHC class-I epitope. Themotif containsall five cleavage sites Y3-G4, A5-E6, A7-L8, L8-E9 and R10-A11defined for eukaryotic proteasomes in which A5-E6 is majorcleavage site [57]. All the chaperone-mediated autophagy substrateproteins described to date are soluble cytosolic proteins containing

Fig. 5. IL-12-expressing vector (pVITRO2- hygro-mcs) used as an adjuvant. The Red and Cyan color shows the integrated p35 and p40 subunits of IL-12. (For interpretation of thereferences to colour in this figure legend, the reader is referred to the web version of this article.)
B.A. Akhoon et al. / Microbial Pathogenesis 51 (2011) 77e8784
a targeting motif biochemically related to the pentapeptide KFERQ[58,59]. This motif, present inw30% of the proteins in the cytosol, isrecognized by a cytosolic chaperone, the heat shock cognateprotein of 73 kDa (cyt-hsc70) [60,61] and the interaction with thischaperone, modulated by the hsc70 cochaperones targets thesubstrate to the lysosomal membrane, where it interacts with thelysosomal membrane protein type 2a [62]. Hence, KFERQmotif wasadded along the both termini of each epitope. The final array ofboth sequences was start codon, epitopes (peptide HEYGAEALERAGat eachMHC class-I epitope and peptide KFERQ at eachMHC class-IIepitope) and then stop codon respectively.
3.6. Reverse translation and optimization of DNA vaccinecandidates
Protein sequences were reverse translated into nucleotidesequences by using backtranseq program of mEMBOSS 6.0.1. Thecodon frequency table for human (Ehum.cut) was used to replaceeach amino acid of input sequence with the corresponding mostfrequently occurring codon [63,64] because the degeneracy of thegenetic code makes backtranslation potentially ambiguous sincemost amino acids are encoded by multiple codons.
There are many ways to optimize the efficiency of a DNAvaccinebeyond the choice of antigen. One of the most effective of thesetechniques has been codon optimization, which can enhance theefficiency of gene expression. It is a common technique whichenhances the efficiency of DNA expression vectors used in DNAvaccination to achieve optimum expression of a foreign gene in thehost’s cell system [65]. All codons inMHCclass-I and -II sequences ofT. brucei were adapted to H. sapiens codon usage by DyNAVacS [66]which computes the optimal codon for each amino acid encoded bythe stretch of DNA and replaces the codons with the most optimalcodon according to the codon usage tables derived from the KazusaCodonUsageDatabase (http://kazusa.or.jp/codon). CpGmotifswereadded to DNA or conventional protein vaccines to enhance the Th1immune response [67]. Antigen specific immunogenicity of a DNA
vaccine is significantly increased by coadministering exogenousCpG-containing DNA [40] and the process was done by DynaVacsserver in which a tool for mapping experimentally proven stimu-lating and suppressive CpG-motifs [68e70] has been incorporated.
3.7. Vector selection and in silico cloning
The pBI-CMV1 vector was chosen as an expression system todesign DNA vaccine. pBI-CMV1 is designed to simultaneously andconstitutively express two genes of interest. Gene expression isdriven by one of two constitutively active, minimal human cyto-megalovirus promoters, PminCMV1 and PminCMV2, located justupstream of two independent multiple cloning sites (MCS 1 andMCS 2, respectively). To allow propagation and selection in E. coli,the vector also contains a ColE1 origin of replication and an ampi-cillin resistance gene (Ampr). pBI-CMV1, which is a bidirectionalexpressionvector, requires each genemust be cloned into a differentMCS, and contain both an initiation codon and a stop codon. For thegeneration of first antigenic insert, we prepared a restrictionmap ofthe antigen sequence and selected two consecutive restriction sites(BamHI and SalI) from theMCS 1 of pBI-CMV1multiple cloning sitesthat were not present in the antigen. Start codon (ATG) and BamHIsitewere added at 50 primer and stop codon (TAA) and SalI sitewereadded at 30 primer of optimized antigenic sequence so that theantigen gene can be directionally cloned. Second antigenic insertwas also mapped but the restriction site XbaI was used at 50 primerand EcoRI at the 30 primer of optimized antigenic sequence. Startand stop codonswere same as added to the antigenic insert first andMCS 2was used for the second antigenic insert. The steps of in silicocloning [71] were performed using clc-DNA Workbench 5.2.0. Thefinal vaccine map was constructed by PlasMapper.
3.8. Adjuvant insertion for boosting the immune response
It has been examined that the vaccine efficacy of DNA encodinga specific antigen in combination with DNA encoding IL-12 could

Fig. 6. A pictorial flowchart representing the key steps of the complete methodology.
B.A. Akhoon et al. / Microbial Pathogenesis 51 (2011) 77e87 85
greatly improve the protective immunity against the protozoandisease [72]. Therefore, Murine IL-12 cDNAwas retrieved fromNCBIand it was successfully cloned in pVITRO2- hygro-mcs (www.invivogen.com) plasmid. The plasmid sequence was extractedfrom the vector database (http://genome-www.stanford.edu/vectordb/). The cDNA of IL-12, which contains p40 and p35subunits, was incorporated into two distinct multiple cloning sites(MCS) i.e., MCS 1 and MCS 2 already present in the vector. For theconvenient cloning of two cDNAs, the p40 and p35 subunits wereinserted between the two restriction sites Eco RV-Bam HI and BglII-EcoRI respectively. All these steps were done through Plasmiddrawing program by using BVTech Plasmid 5.0 software.
The different key steps performed in the complete methodologyare shown in the form of pictorial flowchart in Fig. 6.
4. Conclusion
Vaccines are among the most efficacious and cost-effective toolsfor reducing morbidity and mortality caused by infectious diseases.DNA vaccination represents a novel therapeutic strategy for T. bru-cei infection and will be more effective and long lasting comparedto normal virus vaccinations due to the ability of DNA vaccines toreadily elicit both CTLs and antibodies which distinguish them fromconventional vaccines and make them an attractive vaccineapproach to test with many infectious pathogens. Epitope-basedvaccines (EVs) have recently been attracting significant interest.They trigger an immune response by confronting the immunesystem with immunogenic peptides derived from parasite relatedproteins. Binding of these peptides to proteins from the major
histocompatibility complex (MHC) is crucial for immune systemactivation. However, since theMHC is highly polymorphic, differentpatients typically bind different repertoires of peptides. Hence, it iscrucial to identify the optimal set of peptides for a vaccine, givenconstraints such as MHC allele probabilities in the target pop-ulation and maximum number of selected peptides. Immuneresponses to the VSG (the most abundant surface antigen of thetrypanosomes) are not protective because of the vast repertoire ofimmunologically distinct variants. Therefore, we deriveda consensus sequence (conserved among the sequence variants ofVSGs) from the alignment of majority of the VSGs sequencesavailable at NCBI and identified novel consensus VSG epitopes thatcan serve as best vaccine candidates. It has been already demon-strated that Balb/C mice immunized intramuscularly with a singledose of a DNA plasmid encoding a bloodstream-stage specificinvariant surface glycoprotein (ISG) are partially protected froma lethal dose of T. brucei [73]. The results obtained are encouraging,strongly suggesting that this approach may serve as guideline todevelop new vaccine production strategies but clinical trials arerequired to validate the efficacy of designed vaccine.
Acknowledgment
The authors are highly thankful to Department of biotechnology,New Delhi for providing Bioinformatics Infra Structure Facility forcarrying out this work. SKG acknowledges Society for BiologicalResearch & Rural Development, India for the munificent support.We are also grateful toMr. Tinku Kumar for his technical assistance.

B.A. Akhoon et al. / Microbial Pathogenesis 51 (2011) 77e8786
References
[1] Fèver EM, Picozzi K, Fyfe J, Waiswa C, Odiit M, Coleman PG, et al. A burgeoningepidemic of sleeping sickness in Uganda. The Lancet 2005;366:745e7.
[2] Kennedy PG. The continuing problem of human African trypanosomiasis(sleeping sickness). Ann Neurol 2008;64:116e26.
[3] Taylor KA. Immune responses of cattle to African trypanosomes: protective orpathogenic. Int J Parasitol 1998;28:219e40.
[4] Dumas M, Girard PL. Sleeping sickness. Trends Neurosci 1979;2:24e6.[5] BuguestA,BertJ,TapieP,TabaraudF,DouaF,LonsdorferJ,etal.Sleep-wakecyclein
humanAfricantrypanosomiasis.JClinNeurophysiol1993;10:190e6.[6] Buguest A, Gati R, Severe JP, Develoux M, Bogui P, Lonsdorfer J. 24 hour pol-
ysomnographic evaluation in a patient with sleeping sickness. Electro-encephalogr Clin Neurophysiol 1989;72:471e8.
[7] Kennedy PG. Human African trypanosomiasis of the CNS: current issues andchallenges. J Clin Invest 2004;113:496e504.
[8] Cross GA. Cellular and genetic aspects on antigenic variation in trypanosomes.Annu Rev Immunol 1990;8:83e110.
[9] Blum ML, Down JA, Gurnett AM, Carrington M, Turner MJ, Wiley DC.A structural motif in the variant surface glycoproteins of Trypanosoma brucei.Nature 1993;362:603e9.
[10] Dempsey WL, Mansfield JM. Lymphocyte function in experimental Africantrypanosomiasis. V. Role of antibody and the mononuclear phagocyte systemin variant-specific immunity. J Immunol 1983;130:405e11.
[11] Hertz CJ, Filutowicz H, Mansfield JM. Resistance to the African trypanosomesis IFN-gamma dependent. J Immunol 1998;161:6775e83.
[12] Levine RF, Mansfield JM. Genetics of resistance to the African trypanosomes.III. Variant-specific antibody responses of H-2-compatible resistant andsusceptible mice. J Immunol 1984;133:1564e9.
[13] Schleifer KW, Filutowicz H, Schopf LR, Mansfield JM. Characterization of Thelper cell responses to the trypanosome variant surface glycoprotein.J Immunol 1993;150:2910e9.
[14] Bisser S, Ouwe-Missi-Oukem-Boyer ON, Toure FS, Taoufiq Z, Bouteille B,Buguet A, et al. Harbouring in the brain: a focus on immune evasion mech-anisms and their deleterious effects in malaria and human African trypano-somiasis. Int J Parasitol 2006;36:529e40.
[15] Rhind SG, Shek PN. Cytokines in the pathogenesis of human Africantrypanosomiasis: antagonistic roles of TNF-a and IL-10. In: Dumas M, Bou-teille B, Buguet A, editors. Progress in human African trypanosomiasis,sleeping sick-ness. Paris: Springer; 1999. p. 119e35.
[16] Hill KL, Hutchings NR, Grandgenett PM, Donelson JE. T lymphocyte-triggeringfactor of african trypanosomes is associated with the flagellar fraction of thecytoskeleton and represents a new family of proteins that are present inseveral divergent eukaryotes. J Biol Chem 2000;275:39369e78.
[17] Hutchings NR, Donelson JE, Hill KL. Trypanin is a cytoskeletal linker proteinand is required for cell motility in African trypanosomes. J Cell Biol2002;156:867e77.
[18] Bakhiet M, Olsson T, Edlund C, Hojeberg B, Holmberg K, Lorentzen J, et al.A trypanosoma brucei brucei-derived factor that triggers CD8C lymphocytesto interferon-gamma secretion: purification, characterization and protectiveeffects in vivo by treatment with a monoclonal antibody against the factor.Scand J Immunol 1993;37:165e78.
[19] Atouguia JM, Kennedy PGE. Neurological aspects of human African trypano-somiasis. In: Davis LE, Kennedy PGE, editors. Infectious diseases of thenervous system. Oxford (: Butterworth-Heinemann; 2000. p. 321e72.
[20] Ziegelbauer K, Overath P. Identification of invariant surface glycoprotein in thebloodstream stage of Trypanosoma brucei. J Biol Chem 1992;267:10791e6.
[21] Ziegelbauer K, Multhaup G, Overath P. Molecular characterization of twoinvariant surface glycoproteins specific for the bloodstream stage of Trypa-noma brucei. J Biol Chem 1992;267:10797e803.
[22] Dumonteil E, Escoberdo-Ortegon J, Reyes-Rodriguez N, Arjona-Torres A,Ramirez-Sierra MJ. Immunotherapy of Trypanosoma cruzi infection with DNAvaccines in mice. Infect Immun 2004;72:46e53.
[23] Seder RA, Hill AVS. Vaccine against intracellular infections requiring cellularimmunity. Nature 2000;406:793e8.
[24] Handman E, Noormohammadi AH, Curtis JM, Baldwin T, Sjolander A. Therapyof murine cutaneous leishmaniasis by DNA vaccination. Vaccine2000;18:3011e7.
[25] Lowrie DB, Tascon RE, Bonato VLD, Lima VMF, Faccioli LH, Stravropoulos E,et al. Therapy of tuberculosis in mice by DNA vaccination. Nature1999;400:269e71.
[26] Carvalho JA, Rodgers J, Atouguia J, Prazeres DM, Monteiro GA. DNA vaccines:a rational design against parasitic diseases. Expert Rev Vaccines2010;9:175e91.
[27] Tong JC, Ren EC. Immunoinformatics: current trends and future directions.Drug Discov Today 2009;14:684e9.
[28] Baloria U, Akhoon BA, Gupta SK, Sharma S, Verma V. In silico proteomiccharacterization of human epidermal growth factor receptor 2 (HER-2) for themapping of high affinity antigenic determinants against breast cancer. AminoAcids; 2011, doi:10.1007/s00726-010-0830-x.
[29] De Groot AS, Berzofsky JA. From genome to vaccine e new immu-noinformatics tools for vaccine design. Methods 2004;34:425e8.
[30] De Groot AS. Immunomics: discovering new targets for vaccines and thera-peutics. Drug Discov Today 2006;11:203e9.
[31] del Guercio MF, Alexander J, Kubo RT, Arrhenius T, Maewal A, Appella E, et al.Potent immunogenic short linear peptide constructs composed of B cellepitopes and Pan DR T helper epitopes (PADRE) for antibody responses invivo. Vaccine 1997;15:441e8.
[32] Franke ED, Hoffman SL, Sacci Jr JB, Wang R, Charoenvit Y, Appella E, et al. PanDR binding sequence provides T-cell help for induction of protective anti-bodies against Plasmodium yoeliisporozoites. Vaccine 1999;17:1201e5.
[33] Gupta SK, Smita S, Sarangi AN, Srivastava M, Akhoon BA, Rahman Q, et al. Insilico CD4þ T-cell epitope prediction and HLA distribution analysis for thepotential proteins of Neisseria meningitidis Serogroup B e a clue for vaccinedevelopment. Vaccine 2010;28:7092e7.
[34] Lund O, Nielsen M, Kesmir C, Petersen AG, Lundegaard C, Worning P, et al.Definition of supertypes for HLA molecules using clustering of specificitymatrices. Immunogenetics 2004;55:797e810.
[35] Sette A, Sidney J. Nine major HLA class-I supertypes account for the vastpreponderance of HLA-A and -B polymorphism. Immunogenetics 1999;50:201e12.
[36] Southwood S, Sidney J, Kondo A, del Guercio MF, Appella E, Hoffman S, et al.Several common HLA-DR types share largely overlapping peptide bindingrepertoires. J Immunol 1998;160:3363e73.
[37] Gupta SK, Srivastava M, Akhoon BA, Smita S, Schmitz U, Vera J, et al. Identi-fication of immunogenic consensus T-cell epitopes in globally distributedinfluenza-A H1N1 neuraminidase. Infect Genet Evol 2010b;11:308e19.
[38] Bui HH, Sidney J, Dinh K, Southwood S, Newman MJ, Sette A. Predictingpopulation coverage of T-cell epitope-based diagnostics and vaccines. BMCBioinform 2006;7:153.
[39] Uchijima M, Yoshida A, Nagata T, Koide Y. Optimization of codon usage ofplasmid DNA vaccine is required for the effective MHC class-I-restrictedT-cell responses against an intracellular bacterium. J Immunol 1998;161:5594e9.
[40] Klinman DM, Yamshchikov G, Ishigatsubo Y. Contribution of CpG motifs to theimmunogenicity of DNA vaccines. J Immunol 1997;158:3635e9.
[41] Booth JS, Nichania AK, Benjamina P, Dara A, Kriegb AM, Babiuka LA, et al.Innate immune responses induced by classes of CpG oligodeoxynucleotides inovine lymph node and blood mononuclear cells. Vet Immunol Immunopathol2007;115:24e34.
[42] Lee AH, Suh YS, Sung JH, Yang SH, Sung YC. Comparison of various expressionplasmids for the induction of immune response by DNA immunization. MolCells 1997;7:495e501.
[43] Xu ZL, Mizuguchi H, Ishii-Watabe A, Uchida E, Mayumi T, Hayakawa T.Optimization of transcriptional regulatory elements for constructing plasmidvectors. Gene 2001;272:149e56.
[44] Matthew ND, Darren RF. Harnessing bioinformatics to discover new vaccines.Drug Discov Today 2007;12:389e95.
[45] Harris TH, Mansfield JM, Paulnock DM. CpG oligodeoxynucleotide treatmentenhances innate resistance and acquired immunity to African trypanosomes.Infect Immun 2007;75:2366e73.
[46] Dittmer U, Olbrich AR. Treatment of infectious diseases with immunostimu-latory oligodeoxynucleotides containing CpG motifs. Curr Opin Microbiol2003;6:472e7.
[47] Klinman DM. Immunotherapy uses of CpG oligodeoxynucleotides. Nat RevImmunol 2004;4:249e58.
[48] Mosmann TR, Sad S. The expanding universe of T-cell subsets: Th1, Th2 andmore. Immunol Today 1996;17:138e46.
[49] Murray JS. How MHC selects Th1/Th2 immunity. Immunol Today 1998;19:157e63.
[50] Moreau P, Hen R, Wasylyk B, Everett R, Gaub MP, Chambon P. The SV40 72base repair repeat has a striking effect on gene expression both in SV40 andother chimeric recombinants. Nucleic Acids Res 1981;9:6047e68.
[51] Gowthaman U, Chodisetti SB, Parihar P, Agrewala JN. Evaluation ofdifferent generic in silico methods for predicting HLA class-I bindingpeptide vaccine candidates using a reverse approach. Amino Acids 2010;39:1333e42.
[52] Dimitrov I, Garnev P, Flower DR, Doytchinova I. MHC class-II bindingprediction-a Little help from a friend. J Biomed Biotechnol; 2010:705821.
[53] Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignmentsearch tool. J Mol Biol 1990;215:403e10.
[54] Stern LJ, Brown JH, Jardetzky TS, Gorga JC, Urban RG, Strominger JL, et al.Crystal structure of the human class-II MHC protein HLA-DR1 complexed withan influenza virus peptide. Nature 1994;368:215e21.
[55] Reche PA, Reinherz EL. Sequence variability analysis of human class-I andclass-II MHC molecules: functional and structural correlates of amino acidpolymorphisms. J Mol Biol 2003;331:623e41.
[56] Maenaka K, Jones EY. MHC superfamily structure and the immune system.Curr Opin Struct Biol 2009;9:745e53.
[57] Dolenc I, Seemuller E, Baumeister W. Decelerated degradation of shortpeptides 550 by the 20S proteasome. FEBS Lett 1998;434:357e61.
[58] Chiang HL, Dice JF. Peptide sequences that target proteins for enhanceddegradation during serum withdrawal. J Biol Chem 1988;263:6797e805.
[59] Dice J, Finn P, Majeski A, Mesieres N, Cuervo A. Chaperone-mediatedautophagy. In: Klionsky DJ, editor. Autophagy. Georgetown, TX: LandesBioscience; 2003. p. 158e77.
[60] Chiang HL, Terlecky S, Plant CP, Dice JF. A role for a 70-kilodalton heat shockprotein in lysosomal degradation of intracellular proteins. Science1989;246:382e5.

B.A. Akhoon et al. / Microbial Pathogenesis 51 (2011) 77e87 87
[61] Agarraberes FA, Dice JF. A molecular chaperone complex at the lysosomalmembrane is required for protein translocation. J Cell Sci 2001;114:2491e9.
[62] Cuervo AM, Dice JF. A receptor for the selective uptake and degradation ofproteins by lysosomes. Science 1996;27:501e3.
[63] Ferro A, Giugno R, Pigola G, Pulvirenti A, Pietro CD, Purrello M. Sequencesimilarity is more relevant than species specificity in probabilistic back-translation. BMC Bioinform 2007;8. WI. 58.
[64] Peter R, Ian L, Alan B. EMBOSS: the European molecular biology open suite.Trends Genet 2000;16:276e7.
[65] Kuljeet SS, Sunil P, Souvik M, Beena P. GASCO: genetic algorithm simulationforcodon optimization. Silico Biol 2008;8:187e92.
[66] Nagarajan H, Rekha G, Parul A, Vinod S, Beena P. DyNAVacS: an inte-grative tool for optimized DNA vaccine design. Nucleic Acids Res2006;34:264e6.
[67] Arthur MK, Ae-Kyung Y, Joachim S, Heathe LD. The role of CpG dinucleotidesin DNA vaccines. Trends Microb 1998;6:23e7.
[68] Bauer M, Redecke V, Ellwart JW, Scherer B, Kremer JP, Wagner H, et al.Bacterial CpG-DNA triggers activation and maturation of human CD11c-,CD123þ dendritic cells. J Immunol 2001;166:5000e7.
[69] Krieg AM, Yi AK, Matson S, Waldschmidt TJ, Bishop GA, Teasdale R, et al. CpGmotifs in bacterial DNA trigger direct B-cell activation.Nature 1995;374:546e9.
[70] Van-Uden J, Raz E. Introduction to immunostimulatory DNA sequences. SeminImmunopathol 2000;22:1e9.
[71] Gupta SK, Singh A, Srivastava M, Gupta SK, Akhoon BA. In silico DNA vaccinedesigning against human papillomavirus (HPV) causing cervical cancer.Vaccine 2009;28:120e31.
[72] Katae M, Miyahira Y, Takeda K, Matsuda H, Yagita H, Okumura K, et al.Coadministration of an Interleukin-12 gene and a Trypanosoma cruzi geneimproves vaccine efficacy. Infect Immun 2002;70:4833e40.
[73] Lança AS, de Sousa KP, Atouguia J, Prazeres DM, Monteiro GA, Silva MS.Trypanosoma brucei: immunisation with plasmid DNA encoding invariantsurface glycoprotein gene is able to induce partial protection in experimentalAfrican trypanosomiasis. Exp Parasitol 2011;127:18e24.
Related Documents











