RESEARCH ARTICLE Improvement of pesticide mineralization in on-farm biopuri¢cation systems by bioaugmentation with pesticide-primed soil Kristel Sniegowski 1 , Karolien Bers 1 , Kris Van Goetem 1 , Jaak Ryckeboer 1 , Peter Jaeken 2 , Pieter Spanoghe 3 & Dirk Springael 1 1 Division of Soil and Water management, Katholieke Universiteit Leuven, Heverlee, Belgium; 2 Ecology Department, Proefcentrum Fruitteelt vzw, Sint-Truiden, Belgium; and 3 Laboratory of Crop Protection Chemistry, Ghent University, Ghent, Belgium Correspondence: Kristel Sniegowski, Division of Soil and Water management, Katholieke Universiteit Leuven, Kasteelpark Arenberg 20, 3001 Heverlee, Belgium. Tel.: 132 16 32 16 04; fax: 132 16 19 97; e-mail: [email protected] Present address: Peter Jaeken, Phytofar, A. Reyerslaan 80, 1030 Brussels, Belgium. Received 28 June 2010; revised 3 December 2010; accepted 3 December 2010. Final version published online 13 January 2011. DOI:10.1111/j.1574-6941.2010.01031.x Editor: Kornelia Smalla Keywords biopurification system; pesticide-primed soil; bioaugmentation; linuron mineralization; Variovorax. Abstract Microcosms were used to examine whether pesticide-primed soils could be preferentially used over nonprimed soils for bioaugmentation of on-farm biopuri- fication systems (BPS) to improve pesticide mineralization. Microcosms contain- ing a mixture of peat, straw and either linuron-primed soil or nonprimed soil were irrigated with clean or linuron-contaminated water. The lag time of linuron mineralization, recorded for microcosm samples, was indicative of the dynamics of the linuron-mineralizing biomass in the system. Bioaugmentation with linuron- primed soil immediately resulted in the establishment of a linuron-mineralizing capacity, which increased in size when fed with the pesticide. Also, microcosms containing nonprimed soil developed a linuron-mineralizing population, but after extended linuron feeding. Additional experiments showed that linuron-miner- alization only developed with some nonprimed soils. Concomitant with the increase in linuron degradation capacity, targeted PCR-denaturing gradient gel electrophoresis showed the proliferation of a Variovorax phylotype related to the linuron-degrading Variovorax sp. SRS16 in microcosms containing linuron- primed soil, suggesting the involvement of Variovorax in linuron degradation. The correlation between the appearance of specific Variovorax phylotypes and linuron mineralization capacity was less clear in microcosms containing non- primed soil. The data indicate that supplementation of pesticide-primed soil results in the establishment of pesticide-mineralizing populations in a BPS matrix with more certainty and more rapidly than the addition of nonprimed soil. Introduction Since 1940, pesticides are intensively used worldwide. An important environmental issue of pesticide use is the pollu- tion of ground and surface water as a result of either diffuse (run-off, percolation and spray drift) or point contamina- tion (direct losses through spillage and leakages). Recent studies showed that direct losses account for 40–90% of the surface water pollution (De Wilde et al., 2007). To minimize direct pesticide losses, the installation of biopurification systems (BPS) to treat pesticide-contaminated wastewater on the farm yard has been proposed (Torstensson & del Pilar Castillo, 1997; Vischetti et al., 2004; De Wilde et al., 2007). In on-farm BPS, the contaminated water is conducted over a solid matrix, called a biomix, which is composed of a mixture of various materials, for example straw, peat and soil, in which biodegradation and sorption result in pesti- cide removal. BPS are considered a simple, low-cost, prac- tical and labor-extensive approach for farmers to treat pesticide-contaminated wastewater on the farm. Despite the high pesticide removal percentage observed in BPS (Fogg et al., 2003a, b, 2004; Pigeon et al., 2005), degradation remains poor for some pesticides (Fogg et al., 2003a, 2004). Moreover, a rapid complete degradation is advised to avoid possible toxicity effects of accumulated contaminants (Hen- riksen et al., 2003), aging (Johannesen et al., 2003; Zhao et al., 2003) and the occurrence of mobile and toxic metabolites (Coppola et al., 2007). In addition, degradation during start-up of the system is poor (Fogg et al., 2004) because the appropriate microorganisms need to proliferate FEMS Microbiol Ecol 76 (2011) 64–73 c 2011 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved MICROBIOLOGY ECOLOGY

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

R E S E A R C H A R T I C L E
Improvementof pesticidemineralization inon-farmbiopuri¢cationsystemsbybioaugmentationwith pesticide-primed soilKristel Sniegowski1, Karolien Bers1, Kris Van Goetem1, Jaak Ryckeboer1, Peter Jaeken2, PieterSpanoghe3 & Dirk Springael1
1Division of Soil and Water management, Katholieke Universiteit Leuven, Heverlee, Belgium; 2Ecology Department, Proefcentrum Fruitteelt vzw,
Sint-Truiden, Belgium; and 3Laboratory of Crop Protection Chemistry, Ghent University, Ghent, Belgium
Correspondence: Kristel Sniegowski,
Division of Soil and Water management,
Katholieke Universiteit Leuven, Kasteelpark
Arenberg 20, 3001 Heverlee, Belgium.
Tel.: 132 16 32 16 04; fax: 132 16 19 97;
e-mail: [email protected]
Present address: Peter Jaeken, Phytofar, A.
Reyerslaan 80, 1030 Brussels, Belgium.
Received 28 June 2010; revised 3 December
2010; accepted 3 December 2010.
Final version published online 13 January 2011.
DOI:10.1111/j.1574-6941.2010.01031.x
Editor: Kornelia Smalla
Keywords
biopurification system; pesticide-primed soil;
bioaugmentation; linuron mineralization;
Variovorax.
Abstract
Microcosms were used to examine whether pesticide-primed soils could be
preferentially used over nonprimed soils for bioaugmentation of on-farm biopuri-
fication systems (BPS) to improve pesticide mineralization. Microcosms contain-
ing a mixture of peat, straw and either linuron-primed soil or nonprimed soil were
irrigated with clean or linuron-contaminated water. The lag time of linuron
mineralization, recorded for microcosm samples, was indicative of the dynamics of
the linuron-mineralizing biomass in the system. Bioaugmentation with linuron-
primed soil immediately resulted in the establishment of a linuron-mineralizing
capacity, which increased in size when fed with the pesticide. Also, microcosms
containing nonprimed soil developed a linuron-mineralizing population, but after
extended linuron feeding. Additional experiments showed that linuron-miner-
alization only developed with some nonprimed soils. Concomitant with the
increase in linuron degradation capacity, targeted PCR-denaturing gradient gel
electrophoresis showed the proliferation of a Variovorax phylotype related to the
linuron-degrading Variovorax sp. SRS16 in microcosms containing linuron-
primed soil, suggesting the involvement of Variovorax in linuron degradation.
The correlation between the appearance of specific Variovorax phylotypes and
linuron mineralization capacity was less clear in microcosms containing non-
primed soil. The data indicate that supplementation of pesticide-primed soil
results in the establishment of pesticide-mineralizing populations in a BPS matrix
with more certainty and more rapidly than the addition of nonprimed soil.
Introduction
Since 1940, pesticides are intensively used worldwide. An
important environmental issue of pesticide use is the pollu-
tion of ground and surface water as a result of either diffuse
(run-off, percolation and spray drift) or point contamina-
tion (direct losses through spillage and leakages). Recent
studies showed that direct losses account for 40–90% of the
surface water pollution (De Wilde et al., 2007). To minimize
direct pesticide losses, the installation of biopurification
systems (BPS) to treat pesticide-contaminated wastewater
on the farm yard has been proposed (Torstensson & del Pilar
Castillo, 1997; Vischetti et al., 2004; De Wilde et al., 2007).
In on-farm BPS, the contaminated water is conducted over
a solid matrix, called a biomix, which is composed of a
mixture of various materials, for example straw, peat and
soil, in which biodegradation and sorption result in pesti-
cide removal. BPS are considered a simple, low-cost, prac-
tical and labor-extensive approach for farmers to treat
pesticide-contaminated wastewater on the farm. Despite
the high pesticide removal percentage observed in BPS
(Fogg et al., 2003a, b, 2004; Pigeon et al., 2005), degradation
remains poor for some pesticides (Fogg et al., 2003a, 2004).
Moreover, a rapid complete degradation is advised to avoid
possible toxicity effects of accumulated contaminants (Hen-
riksen et al., 2003), aging (Johannesen et al., 2003; Zhao
et al., 2003) and the occurrence of mobile and toxic
metabolites (Coppola et al., 2007). In addition, degradation
during start-up of the system is poor (Fogg et al., 2004)
because the appropriate microorganisms need to proliferate
FEMS Microbiol Ecol 76 (2011) 64–73c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
MIC
ROBI
OLO
GY
EC
OLO
GY

in the biomix before maximum degradation rates are
obtained. To ensure rapid and complete degradation of the
pesticides in BPS, bioaugmentation of the biomix with
microorganisms containing catabolic pathways enabling
complete mineralization of pesticides is suggested. Pesti-
cide-mineralizing microorganisms can often be found in
soils or other ecosystems with a long history of pesticide
contamination, designated as pesticide-primed materials.
The organisms can be inoculated either as (formulated)
cultured strains or along with the pesticide-primed material
(e.g. soil) in which they have developed. In comparison with
pure cultures, the latter is expected to contain (1) a larger
gene pool and higher diversity in microorganisms that
contribute to pesticide degradation and (2) populations
better adapted to in situ conditions. In addition, this
approach does not require the isolation of the appropriate
organisms. This is important since, to date, pure strains able
to mineralize a pesticide have been reported only for a few
pesticide compounds. Moreover, bioaugmentation using
lab-cultured pollutant-degrading isolates often had limited
success (Chatterjee et al., 1982; Grigg et al., 1997; Struthers
et al., 1998; Topp, 2001; Mertens et al., 2006; Moran et al.,
2006; Singh et al., 2006; Bazot & Lebeau, 2008). On the other
hand, the few reports on bioaugmentation of contaminated
matrices with pollutant-primed materials showed promising
results (Barbeau et al., 1997; Runes et al., 2001; Grundmann
et al., 2007). Despite the apparent high potential of applying
primed materials for bioaugmentation of contaminated
ecosystems, little research has been performed on this topic.
Moreover, no reports exist of using pesticide-primed mate-
rials for bioaugmenting BPS despite the low associated cost
of such an approach for farmers. However, BPS contain a
complex matrix prepared of components from different
ecosystems harboring different microbial communities and
provide a high nutrient content with multiple substrates for
microbial growth. It has to be examined whether pesticide-
mineralizing populations introduced by inoculating pesti-
cide-primed materials can compete and proliferate in such a
complex biotechnological matrix.
Therefore, in this study, the dynamics of the pesticide
mineralization capacity of lab-scale BPS microcosms (BM),
containing a mixture of soil, straw and peat, were compared
when inoculated with either a pesticide-primed soil posses-
sing a pesticide mineralization capacity or a nonprimed soil
without apparent pesticide mineralization capacity and
irrigated or not with a pesticide-containing solution. The
phenylurea herbicide linuron was used as model pesticide
and soil originating from an agricultural field that had been
treated annually with linuron and that contains linuron-
mineralizing organisms (Breugelmans et al., 2007) was used
as the model pesticide-primed material. The linuron miner-
alization capacity of the BMs was monitored by means of14C-linuron mineralization assays. Because linuron-degrad-
ing isolates originating from linuron-treated soils, including
the linuron-primed soil used in this study, almost exclu-
sively belong to the genus Variovorax (Dejonghe et al., 2003;
Sorensen et al., 2005; Breugelmans et al., 2007), the number
of Variovorax and composition of the Variovorax commu-
nity within the BMs was monitored by means of targeted
molecular techniques.
Materials and methods
Pesticides used
Linuron [3-(3,4-dichlorophenyl)-1-methoxy-1-methyl urea]
(purity, 99.5%) was purchased from Sigma Aldrich (Belgium).
[phenyl-U-14C] linuron (16.93 mCi mmol�1, radiochemical
purity 4 95%) was obtained from Izotop, Hungary.
BM set-ups
BM were set up in glass cylinders (height 10 cm; diameter
4 cm) containing a glass filter positioned at 8 cm depth and
filled with the appropriate mixture of soil, peat and straw
(Table 1). The linuron-primed soil (soil L) was sampled
from the A-horizon of a potato field in Halen, Belgium, in
April 2005. The field had been treated for several years with
linuron and was shown to contain a linuron-mineralizing
microbial community (Breugelmans et al., 2007). Non-linuron-
primed soils were obtained from seven different locations
representing four different ecosystems, i.e. garden (soils G1 and
Table 1. Overview of the different BM set-ups operated in this study
Set-up� Origin soil
Linuron-
primed soil
Linuron
treatment
Moisture
content
(w/w%) pH (� SD)
Experiment 1: mixture: straw (25 vol%); peat (25 vol%); soil (50 vol%)
L� Agriculture 1 � 56.12 6.20 (� 0.12)
L1 Agriculture 1 1 60.52 6.20 (� 0.12)
C� Construction
site
� � 48.00 6.05 (� 0.22)
C1 Construction
site
� 1 48.14 6.05 (� 0.22)
Experiment 2: mixture: straw (37.5 vol%); peat (37.5 vol%);
soil (25 vol%)
O1w
No soil � 1 217.78 5.11 (� 0.10)
L1 Agriculture 1 1 89.52 5.44 (� 0.26)
C1 Construction
site
� 1 71.03 5.49 (� 0.21)
A11 Agriculture � 1 42.67 5.56 (� 0.55)
A21 Agriculture � 1 34.64 5.09 (� 0.15)
F11 Forest � 1 82.33 5.61 (� 0.10)
F21 Forest � 1 57.54 4.87 (� 0.11)
G11 Garden � 1 74.19 5.43 (� 0.05)
G21 Garden � 1 42.25 5.38 (� 0.30)
�1 and � indicate treatment with and without linuron, respectively.wStraw: 50 vol%, peat: 50 vol%, soil: 0 vol%.
FEMS Microbiol Ecol 76 (2011) 64–73 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
65Pesticide-primed soil to improve on-farm biopurification

G2), agriculture field (soils A1 and A2), forest (soils F1 and F2)
and a construction site (soil C). To the best of our knowledge,
these soils were never treated with linuron and, in addition, did
not show any capacity to mineralize linuron for a period of 70
days in 14C-linuron mineralization assays (data not shown). All
soils were top soils sampled from the upper 20 cm. The soils
were stored at 4 1C in the dark and sieved before use. After
filling, the BMs were positioned on a dish to collect drainage
water and placed in a glass jar closed with an air-open lid to
avoid desiccation and contamination. No leaching of linuron
was observed.
Two BM experiments were set up as outlined in Table 1,
which provides an overview of the compositions, moisture
contents and pHs of the biomix in the various BM set-ups. The
moisture content, based on the weight of a sample (� 0.500 g)
taken from the upper layer of the BMs before and after 2 days
incubation at 60 1C, was determined at each sampling time.
The pH was measured at the start and the end of both
experiments in a 0.01 M CaCl2 extract (soil : liquid ratio 1 : 5).
The pHs determined at the start of the experiments did not
differ significantly within each set-up. In addition, neither pH
nor moisture content changed significantly during the incuba-
tion period (data not shown).
In the first experiment, BMs contained a mixture of
25 vol% cut straw (� 0.5 cm2), 25 vol% peat and 50 vol% soil.
The physico-chemical characteristics of the used substrata are
shown in Table 2. BMs of set-ups L� and L1 were inoculated
with the linuron-primed soil L and packed to a density of
0.94 g cm�3, while BMs of set-ups C� and C1 were inoculated
with nonprimed soil C packed to a density of 0.78 g cm�3.
BMs of set-ups C� and L�were irrigated with sterile tap water,
while BMs of set-ups C1 and L1 received sterile tap water
containing 60 mg L�1 linuron. Both solutions were manually
spread evenly over the surface of the matrix using a 1-mL
pipette during 12 weeks. Each week, the solutions were applied
on Monday (1 mL), Wednesday (1 mL) and Friday (1.5 mL),
resulting in an average added volume of 3.18 L m�3 day�1.
Each set-up included triplicate BMs. All BMs were incubated
in the dark at 25 1C. The upper 1 cm of the matrix in the BMs
was mixed with a sterile spatula, before taking samples to
examine the linuron mineralization capacity. The samples
were taken exactly 0, 2, 4, 8 and 12 weeks after starting the
treatments. Additional samples were taken at weeks 0, 2 and
12 for analysis of the Variovorax community.
In the second experiment, BMs contained eight different
mixtures consisting of 37.5 vol% cut straw, 37.5 vol% peat and
25 vol% of soil and were operated in triplicate. The straw and
peat used were the same as those used in the first experiment.
BMs of set-up L1 were inoculated with the linuron-primed
soil, while BMs of the other set-ups were inoculated with
either one of the seven nonprimed soils (G1, G2, A1, A2, F1, F2,
and C). As a control, a ninth set-up was included in which the
BMs contained a mixture of 50 vol% straw and 50 vol% peat
only. Each of these BM set-ups received a linuron-containing
solution for a period of 20 weeks as described above. Samples
were taken after 0, 15 and 20 weeks of incubation at 25 1C for
determination of the 14C-linuron mineralization capacity as
described for the first set-up. Additional samples were taken at
weeks 0 and 15 for analysis of the Variovorax community.
14C-linuron mineralization assays14C-linuron mineralization assays were performed as de-
scribed by Breugelmans et al. (2007). The matrix sample
(200 mg) was added to a volume of 5 mL MMN minimal
medium (Breugelmans et al., 2007), pH 5.8, containing both
unlabeled (20 mg L�1) and 14C-labeled (31mg L�1) linuron
as the only carbon and nitrogen source (a final radioactivity
of 213 Bq mL�1) in 20-mL pyrex tubes containing NaOH
traps. During incubation at 20 1C on a rotary shaker at
150 r.p.m., the amount of 14CO2 produced with reference to
the initial added amount of 14C-linuron was measured and
cumulative mineralization curves were established. As a
negative control, 14CO2 production of the medium without
inoculation of a biomix sample was monitored. The lag
phase, defined as the period between initiating the miner-
alization assay and start of mineralization, was calculated as
the intersection of the x-axis with the linear regression line
between two successive points of the mineralization curve,
where the amount of 14CO2 showed the largest increase. The
slope of the linear regression line of maximum mineraliza-
tion determined the maximum mineralization rate (Broos
et al., 2005).
Most probable number (MPN) mineralizationmethod
An estimation of the size of the active linuron-mineralizing
biomass in samples of the BM matrix was performed using
Table 2. Physicochemical characteristics of the substrata used in the biomix of the BMs
Substratum Moisture content % (w/w) pH Specific density (g cm�3) Total C (%) Total N (%) C/N (%)
Soil C 2.83 6.9 2.81 0.95 0.08 12.1
Soil L 11.53 4.9 ND 0.80 0.06 13.3
Straw 8.40 6.6 1.56 43.25 0.46 95.1
Peat 49.67 6.4 1.58 43.97 0.93 47.1
ND, not determined.
FEMS Microbiol Ecol 76 (2011) 64–73c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
66 K. Sniegowski et al.
an MPN approach. A sample (10 g, wet weight) taken from
the biomix of the BM was added to 25 mL MgSO4 (10�2 M)
and incubated overnight on a shaker to remove the cells
from the matrix. After settling of the matrix material for 2 h,
a decimal serial dilution was made with 2 mL of the aqueous
extract. Aliquots of 0.5 mL of these dilutions were used as
inoculum for 14C-linuron mineralization assays in triplicate,
operated as described above. Positive tubes for MPN calcu-
lation were those vials where 4 10% total 14CO2 was
produced within 60 days. MPN calculations were performed
using the computer-assisted method developed by Briones
& Reichardt (1999) and the MPN number was expressed as
active mineralizing units per gram dry weight (dw) of the
sample (AMU g�1 dw).
Molecular techniques
Total DNA was extracted from 400-mg samples of the
biomix as described by Uyttebroek et al. (2006). The copy
number of the Variovorax 16S rRNA gene was determined by
real-time PCR, performed in a Rotor Gene (RG 3000)
apparatus from Westburg using the Variovorax-specific 16S
rRNA gene primers VarF and VarR. Details of the method
are described in Bers et al. (2011). Real-time PCR deter-
mined the bacterial 16S rRNA gene copy number as
described by Haest et al. (2010) using primers Eub341F and
Eub534R (Muyzer et al., 1993). The detection limit of both
methods was 105 copies g�1 biomix dw.
Denaturing gradient gel electrophoresis (DGGE) finger-
prints of the Variovorax community were performed using
16S rRNA gene fragments amplified with primers VarF-GC
and VarR in either a single or a double PCR as described by
Bers et al. (2011) on a polyacrylamide gel (10%) with a
denaturating gradient from 45% to 75% as described by
Uyttebroek et al. (2006). The 16S rRNA gene fragments in
the gel were mobilized through an electric field set at 120 V
for 15 h. On each gel, a reference DGGE migration Variovorax
marker sample (ref-Var) was loaded, containing 16S rRNA
gene fragments PCR amplified with primers GC-VarF and
VarR from Variovorax sp. SRS16 (Sorensen et al., 2005),
Variovorax sp. DSM66 and Variovorax sp. WDL1 (Dejonghe
et al., 2003) representing three major phylotypes within the
Variovorax genus (Breugelmans et al., 2007). In addition,
reference DGGE migration marker samples (ref-nonVar)
containing 16S rRNA gene fragments with different GC
content from various bacteria (not belonging to the Variovor-
ax genus) amplified with bacterial primers 63F and 518R (El
Fantroussi et al., 1999) were loaded on each gel. The detection
limit of the Variovorax-specific PCR-DGGE method was
approximately 105 copies g�1 biomix dw. The significance of
differences between averages of the 16S rRNA gene copy
numbers of triplicate BM samples were analyzed by ANOVA
(Po 0.05).
Results
Linuron mineralization capacity of BMbioaugmented with linuron-primed soil andnonprimed soil
In the first experiment, bioaugmentation of BPS with linuron-
primed soil L and nonprimed soil C was compared. Initially,
only samples from BMs inoculated with linuron-primed soil
L, i.e. BMs of set-ups L� and L1, showed linuron mineraliza-
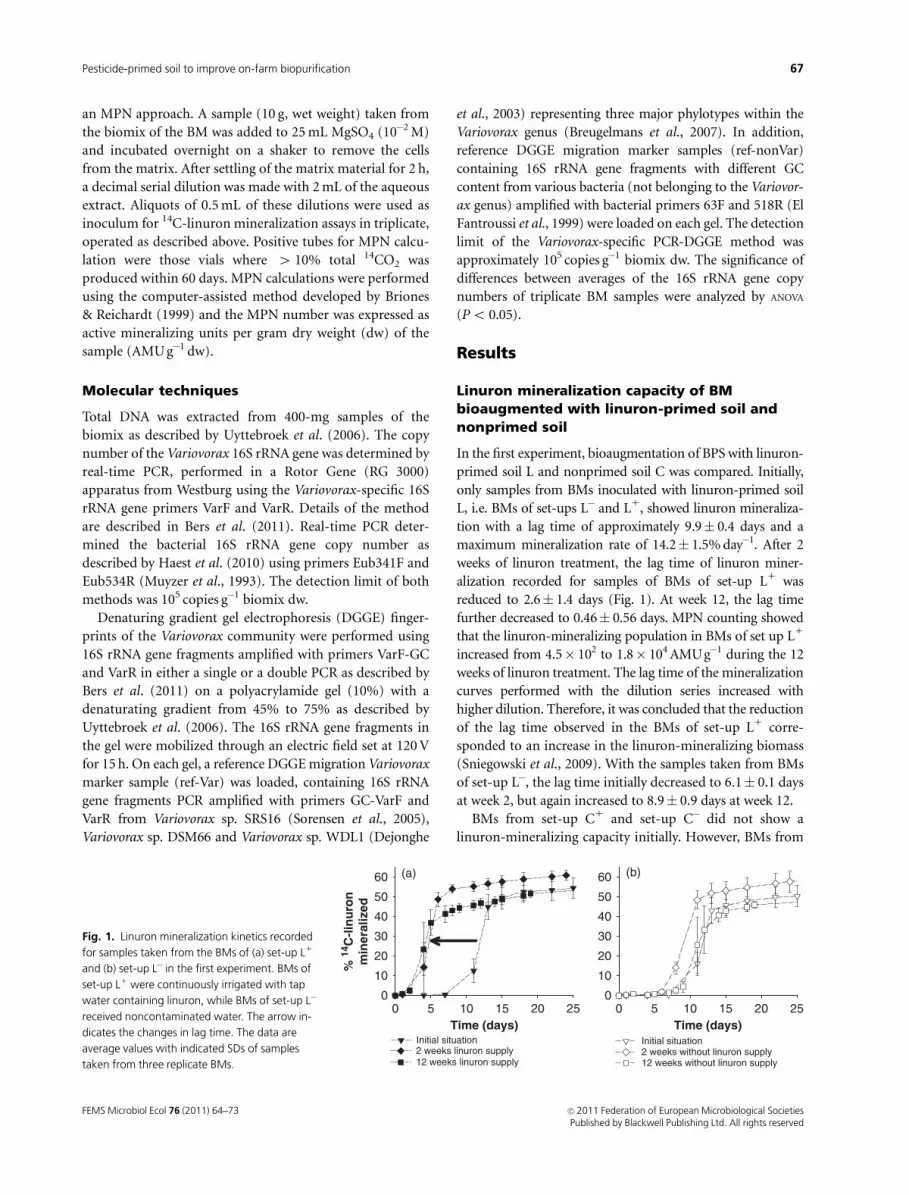
tion with a lag time of approximately 9.9� 0.4 days and a
maximum mineralization rate of 14.2� 1.5% day�1. After 2
weeks of linuron treatment, the lag time of linuron miner-
alization recorded for samples of BMs of set-up L1 was
reduced to 2.6� 1.4 days (Fig. 1). At week 12, the lag time
further decreased to 0.46� 0.56 days. MPN counting showed
that the linuron-mineralizing population in BMs of set up L1
increased from 4.5� 102 to 1.8� 104 AMUg�1 during the 12
weeks of linuron treatment. The lag time of the mineralization
curves performed with the dilution series increased with
higher dilution. Therefore, it was concluded that the reduction
of the lag time observed in the BMs of set-up L1 corre-
sponded to an increase in the linuron-mineralizing biomass
(Sniegowski et al., 2009). With the samples taken from BMs
of set-up L�, the lag time initially decreased to 6.1� 0.1 days
at week 2, but again increased to 8.9� 0.9 days at week 12.
BMs from set-up C1 and set-up C� did not show a
linuron-mineralizing capacity initially. However, BMs from
(b)
Time (days)
0
10
20
30
40
50
60
Initial situation2 weeks without linuron supply12 weeks without linuron supply
(a)
Time (days)0 5 10 15 20 250 5 10 15 20 25
% 14
C-l
inu
ron
min
eral
ized
0
10
20
30
40
50
60
Initial situation2 weeks linuron supply12 weeks linuron supply
Fig. 1. Linuron mineralization kinetics recorded
for samples taken from the BMs of (a) set-up L1
and (b) set-up L� in the first experiment. BMs of
set-up L1 were continuously irrigated with tap
water containing linuron, while BMs of set-up L�
received noncontaminated water. The arrow in-
dicates the changes in lag time. The data are
average values with indicated SDs of samples
taken from three replicate BMs.
FEMS Microbiol Ecol 76 (2011) 64–73 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
67Pesticide-primed soil to improve on-farm biopurification

set-up C1 clearly acquired the capacity to mineralize linuron
upon continuous irrigation with linuron. This capacity devel-
oped in the three replicate BMs, after different periods of
linuron feeding (Fig. 2). Moreover, for one replicate BM, a
significantly lower mineralization rate than that for the other
two was recorded. In addition, the mineralization curve
recorded with this replicate demonstrated a rather linear
mineralization curve, indicating that mineralization was not
linked to growth. At week 12, the average recorded lag time
was 4.5� 1.8 days. BMs from set-up C� did not develop an
observable linuron mineralization capacity during the 12-
week incubation period (data not shown).
Adaptability of BPS bioaugmented withnonprimed soils
Because BMs of set-up C1 developed a linuron mineraliza-
tion capacity upon the continuous supply of a linuron-
containing solution, it was tested whether inoculation with
other nonprimed soils led to a similar development. There-
fore, a second experiment was performed in which non-
primed soils of different origins were used to bioaugment
BMs. To ensure that the acquisition of a linuron mineraliza-
tion capacity was related to the inoculation of the soil, a set-
up containing control BMs without soil addition was
included. BMs containing linuron-primed soil L and non-
primed soil C were included in the experimental set-up. All
systems were irrigated with the linuron-containing solution.
Because in the first experiment, the development of a linuron-
mineralizing capacity in BMs containing nonprimed soil was
observed only after 12 weeks of linuron irrigation, matrix
samples in the current experiment were taken for linuron
mineralization assays after 15 and 20 weeks of incubation.
BMs of set-ups L1 and C1 behaved identical to those in the
first experiment (data not shown). However, the development
of a linuron mineralization capacity was observed with only
one of the other tested nonprimed soils, i.e. soil A1, and in
only one replicate BM (lag time 4.9 days). After 20 weeks of
linuron treatment, the recorded linuron mineralization curves
of the BMs were similar to those recorded after 15 weeks of
treatment (data not shown).
Size and composition of the Variovoraxcommunity in bioaugmented BPS
In the first experiment, the Variovorax 16S rRNA gene copy
number increased both in BMs of set-up L1 and in BMs of
set-up L� between the start of the experiment and week 12
while the number of bacterial 16S rRNA gene copies
remained stable (Table 3). The increases were, however,
insignificant. In BMs of set-ups C� and C1, the Variovorax
16S rRNA gene copy number decreased after 12 weeks of
incubation, but only the decrease observed in set-up C�was
significant (Table 3). Double PCR-DGGE analysis of the
Variovorax community in BMs of set-ups L�, L1, C� and C1
showed similar fingerprints at week 0 for all set-ups with
bands at positions V1, V2 and V4 (Fig. 3a). Position V1
corresponds to the migration position of the 16S rRNA gene
fragment amplified from the linuron-degrading Variovorax
sp. WDL-1, which belongs to Variovorax phylotype B (Bers
et al., 2011). 16S rRNA gene fragments at position V2
correspond to Variovorax members of phylotype C, which
does not contain Variovorax strains with a linuron degrada-
tion capacity (Bers et al., 2011). The fingerprints of week 2 and
week 12 showed the same bands at positions V1, V2 and V4.
However, in set-ups L� and L1 an additional band appeared at
position V3. This band especially dominated in set-up L1 at
week 12 and as such can be associated with the observed
increased capacity to mineralize linuron. Position V3 corre-
sponds to the migration position of the 16S rRNA gene
fragment amplified from the linuron-degrading Variovorax
sp. SRS16, which is a member of the Variovorax phylotype A
(Bers et al., 2011). A band at position V3 also appeared in the
profiles of set-ups C� and C1 at week 12, but no clear
differences in intensity were found between C� and C1.
Real-time PCR results performed with DNA extracted
from the samples of set-ups L1, C1, A11 and O1, taken at
weeks 0 and 15 in the second experiment, showed that the
Week 0
Time (days)
% 14
C-l
inu
ron
min
eral
ized
0102030405060
Week 2
Time (days)
Week 4
Time (days)
Week 12
Time (days)0 10 20 30 40 50 60 0 10 20 30 40 50 60 0 10 20 30 40 50 60 0 10 20 30 40 50 60
BM 1 BM 2 BM 3
Fig. 2. Evolution of the linuron mineralization kinetics recorded with samples taken from the three replicate BMs designated as BM1, BM2 and BM3 of
set-up C1 inoculated with the non-linuron-primed soil in the first experiment.
FEMS Microbiol Ecol 76 (2011) 64–73c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
68 K. Sniegowski et al.
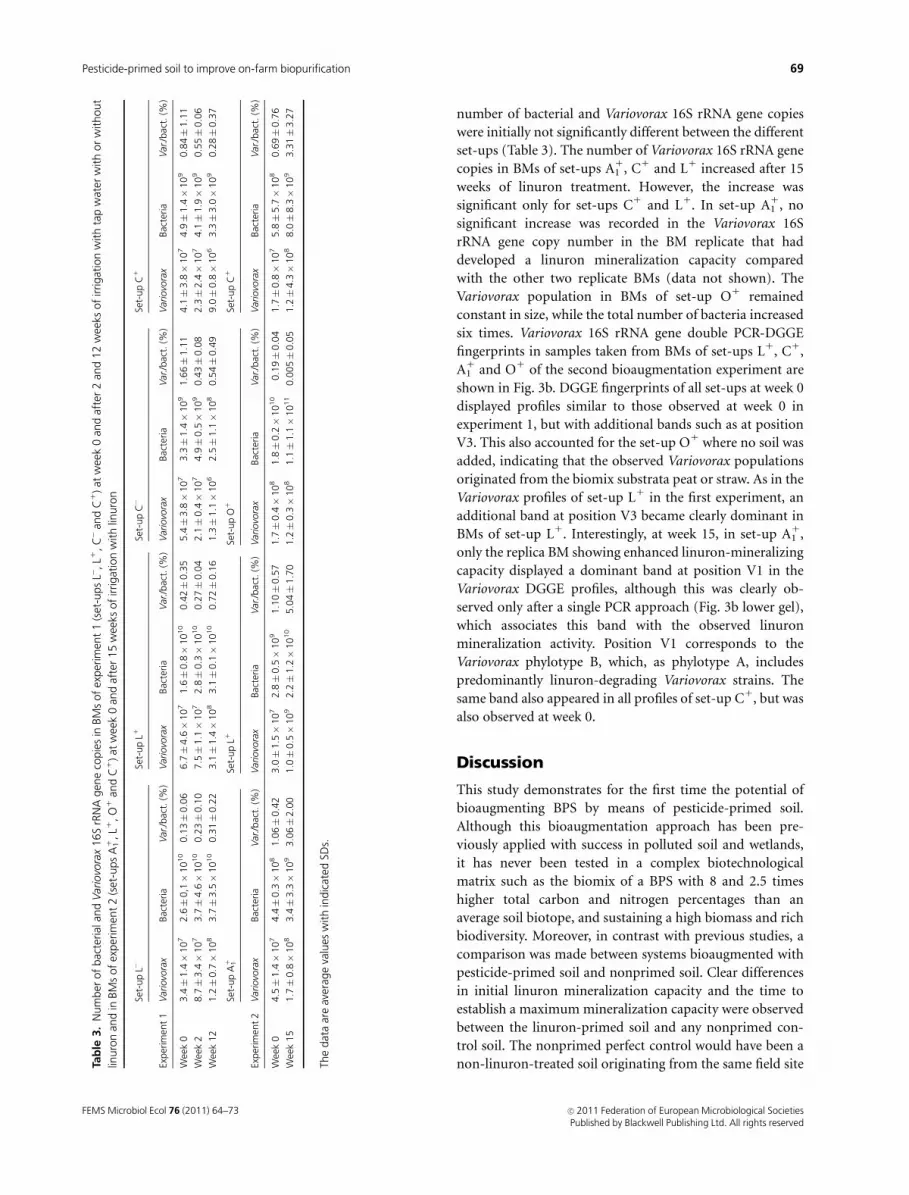
number of bacterial and Variovorax 16S rRNA gene copies
were initially not significantly different between the different
set-ups (Table 3). The number of Variovorax 16S rRNA gene
copies in BMs of set-ups A11, C1 and L1 increased after 15
weeks of linuron treatment. However, the increase was
significant only for set-ups C1 and L1. In set-up A11, no
significant increase was recorded in the Variovorax 16S
rRNA gene copy number in the BM replicate that had
developed a linuron mineralization capacity compared
with the other two replicate BMs (data not shown). The
Variovorax population in BMs of set-up O1 remained
constant in size, while the total number of bacteria increased
six times. Variovorax 16S rRNA gene double PCR-DGGE
fingerprints in samples taken from BMs of set-ups L1, C1,
A11 and O1 of the second bioaugmentation experiment are
shown in Fig. 3b. DGGE fingerprints of all set-ups at week 0
displayed profiles similar to those observed at week 0 in
experiment 1, but with additional bands such as at position
V3. This also accounted for the set-up O1 where no soil was
added, indicating that the observed Variovorax populations
originated from the biomix substrata peat or straw. As in the
Variovorax profiles of set-up L1 in the first experiment, an
additional band at position V3 became clearly dominant in
BMs of set-up L1. Interestingly, at week 15, in set-up A11,
only the replica BM showing enhanced linuron-mineralizing
capacity displayed a dominant band at position V1 in the
Variovorax DGGE profiles, although this was clearly ob-
served only after a single PCR approach (Fig. 3b lower gel),
which associates this band with the observed linuron
mineralization activity. Position V1 corresponds to the
Variovorax phylotype B, which, as phylotype A, includes
predominantly linuron-degrading Variovorax strains. The
same band also appeared in all profiles of set-up C1, but was
also observed at week 0.
Discussion
This study demonstrates for the first time the potential of
bioaugmenting BPS by means of pesticide-primed soil.
Although this bioaugmentation approach has been pre-
viously applied with success in polluted soil and wetlands,
it has never been tested in a complex biotechnological
matrix such as the biomix of a BPS with 8 and 2.5 times
higher total carbon and nitrogen percentages than an
average soil biotope, and sustaining a high biomass and rich
biodiversity. Moreover, in contrast with previous studies, a
comparison was made between systems bioaugmented with
pesticide-primed soil and nonprimed soil. Clear differences
in initial linuron mineralization capacity and the time to
establish a maximum mineralization capacity were observed
between the linuron-primed soil and any nonprimed con-
trol soil. The nonprimed perfect control would have been a
non-linuron-treated soil originating from the same field siteTab
le3.
Num
ber
of
bac
terial
and
Var
iovo
rax
16S
rRN
Agen
eco
pie
sin
BM
sof
exper
imen
t1
(set
-ups
L�,L1
,C�
and
C1
)at
wee
k0
and
afte
r2
and
12
wee
ksof
irrigat
ion
with
tap
wat
erw
ith
or
without
linuro
nan
din
BM
sof
exper
imen
t2
(set
-ups
A11
,L1
,O
1an
dC
1)at
wee
k0
and
afte
r15
wee
ksof
irrigat
ion
with
linuro
n
Exper
imen
t1
Set-
up
L�Se
t-up
L1Se
t-up
C�
Set-
up
C1
Var
iovo
rax
Bac
teria
Var
./bac
t.(%
)V
ario
vora
xBac
teria
Var
./bac
t.(%
)V
ario
vora
xBac
teria
Var
./bac
t.(%
)V
ario
vora
xBac
teria
Var
./bac
t.(%
)
Wee
k0
3.4�
1.4�
10
72.6�
0,1�
10
10
0.1
3�
0.0
66.7�
4.6�
10
71.6�
0.8�
10
10
0.4
2�
0.3
55.4�
3.8�
10
73.3�
1.4�
10
91.6
6�
1.1
14.1�
3.8�
10
74.9�
1.4�
10
90.8
4�
1.1
1
Wee
k2
8.7�
3.4�
10
73.7�
4.6�
10
10
0.2
3�
0.1
07.5�
1.1�
10
72.8�
0.3�
10
10
0.2
7�
0.0
42.1�
0.4�
10
74.9�
0.5�
10
90.4
3�
0.0
82.3�
2.4�
10
74.1�
1.9�
10
90.5
5�
0.0
6
Wee
k12
1.2�
0.7�
10
83.7�
3.5�
10
10
0.3
1�
0.2
23.1�
1.4�
10
83.1�
0.1�
10
10
0.7
2�
0.1
61.3�
1.1�
10
62.5�
1.1�
10
80.5
4�
0.4
99.0�
0.8�
10
63.3�
3.0�
10
90.2
8�
0.3
7
Set-
up
A11
Set-
up
L1Se
t-up
O1
Set-
up
C1
Exper
imen
t2
Var
iovo
rax
Bac
teria
Var
./bac
t.(%
)V
ario
vora
xBac
teria
Var
./bac
t.(%
)V
ario
vora
xBac
teria
Var
./bac
t.(%
)V
ario
vora
xBac
teria
Var
./bac
t.(%
)
Wee
k0
4.5�
1.4�
10
74.4�
0.3�
10
81.0
6�
0.4
23.0�
1.5�
10
72.8�
0.5�
10
91.1
0�
0.5
71.7�
0.4�
10
81.8�
0.2�
10
10
0.1
9�
0.0
41.7�
0.8�
10
75.8�
5.7�
10
80.6
9�
0.7
6
Wee
k15
1.7�
0.8�
10
83.4�
3.3�
10
93.0
6�
2.0
01.0�
0.5�
10
92.2�
1.2�
10
10
5.0
4�
1.7
01.2�
0.3�
10
81.1�
1.1�
10
11
0.0
05�
0.0
51.2�
4.3�
10
88.0�
8.3�
10
93.3
1�
3.2
7
The
dat
aar
eav
erag
eva
lues
with
indic
ated
SDs.
FEMS Microbiol Ecol 76 (2011) 64–73 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
69Pesticide-primed soil to improve on-farm biopurification

as the linuron-primed soil but such a soil was not available.
Nevertheless, the effect of linuron priming is accentuated in
the second experiment where only two of the tested non-
primed soils resulted into the development of a linuron
mineralization capacity.
In contrast to nonprimed soil, bioaugmentation with
linuron-primed soil immediately enhanced the linuron
mineralization capacity of the BMs. Moreover, feeding with
linuron further increased the linuron-mineralizing capacity
in the matrix. MPN counting demonstrated that the ob-
served enhancement in linuron mineralization capacity
could be related to an increase in size of the linuron-
mineralizing population. Therefore, it can be concluded
that despite the complex background, the high biomass and
high biodiversity, the community or at least the linuron-
degrading fraction in the added soil could proliferate in the
biomix in case fed with linuron. Various studies, mostly
performed with suspended cultures inoculated with pure
bacterial strains, reported on the effect of a highly degrad-
able carbon and nitrogen content on mineralization/degra-
dation of pesticides or other pollutants. Positive effects of
additional C and/or N sources on pesticide degradation
were reported by Cullington & Walker (1999), Aslan &
Turkman (2005) and Fogg et al. (2003b), while Breugelmans
et al. (2010) noticed negative effects on degradation of
linuron. On the other hand, the maintenance of the linuron-
mineralizing capacity in BMs of set-up L� throughout the
experiment indicate that the linuron-degrading population
present in the primed soil is sufficiently competitive in its
new biotope and can establish even without apparent
selective conditions. Similarly, Johnsen et al. (2007) reported
the long-term survival of a polyaromatic hydrocarbon
(PAH)-degrading community present in inoculum PAH-
primed soil, in soils without PAH pollution.
BMs of set-up C1, inoculated with nonprimed soil C,
started to mineralize linuron after an extended period of
linuron supply. A similar result was obtained with one soil of
the other tested non-linuron-primed soils. Development of
a linuron mineralization capacity in those BMs occured only
when the pesticide was supplied, showing the selective
nature of the process. Because the set-ups showing this
outcome had the maximum moisture contents and because
the pH did not significantly differ between set-ups, it is very
unlikely that such differences in environmental conditions
Week 15Week 0
V4V3
V1
V2
V4V3
V1
V2
A1+A1
+ C+ O+L+ O+L+C+RVR R– – – – – – + + + – – – + – – + + + + + + – – –
A1+
–––+++++–+––++–––– ––
Week 0
A1+ C+ O+
Week 15
L+ O+L+C+
V4V3
V1
V2
V4V3
V1
V2
RVR R
Double PCR
R
Week 12Week 2Week 0
C– C+ L– L+ C– C+ L– L+ C– C+ L– L+R RVR RV
V4V3V2
V3V2
V1
V4
V1
(a)
(b)
Single PCR
Double PCR
Fig. 3. 16S rRNA gene-based DGGE fingerprints of the Variovorax community obtained from (a) BM samples taken at weeks 0, 2 and 12 from BMs of
set-ups C�, C1, L� and L1 in the first experiment and (b) BM samples taken at weeks 0 and 15 from set-ups A11, C1, L1 and O1 in the second
experiment. (a) Only DGGE profiles obtained with 16S rRNA gene fragments produced by double PCR are shown. (b) DGGE profiles obtained with 16S
rRNA gene fragments produced by double PCR (upper gel) and single PCR (lower gel). DGGE fingerprints of reference DGGE migration markers ref-
nonVar and ref-Var are indicated with ‘R’ and ‘V’, respectively. Band V3 corresponds to phylotype A, band V2 with phylotype C and band V1 with
phylotype B.
FEMS Microbiol Ecol 76 (2011) 64–73c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
70 K. Sniegowski et al.

were responsible for the observed differences in linuron
mineralization development. Instead, it can be hypothesized
that either the microbial community in those BMs geneti-
cally adapted to degrade linuron or a linuron-mineralizing
bacterial population initially present in the soil at undetec-
tably low numbers proliferated when linuron was added.
Interestingly, different replicate BMs developed a linuron-
mineralizing capacity either after different treatment periods
(in case of bioaugmentation with soil C) or only in one BM
replicate (in case of bioaugmentation with soil A1). This can be
explained by the occurrence of genetic adaptation at different
time points or in one replicate, or by the fact that low initial
numbers of mineralizing biomass were unevenly distributed in
the soil sample used for inoculation. A similar observation was
reported by Cullington & Walker (1999) with diuron.
The number of Variovorax in BMs containing linuron-
primed soil L and receiving linuron (set-up L1) increased
concomitantly with the increase in linuron mineralization
capacity, but this was statistically significant only in BMs
operated in the second experiment. The increase in Vario-
vorax 16S rRNA gene copy number was less pronounced
compared with the 100-fold increase observed in soil L when
it was supplied linuron on a long-term base (Bers et al.,
2011). The reason for the observed difference in prolifera-
tion of Variovorax between set-ups L1 in the first and second
experiment might be due to the longer incubation period (3
weeks) and/or due to the higher amount of nutrients
because of the higher fraction of straw and peat (only
25 vol% soil inoculum) in the biomix used in the second
experiment. On the other hand, in both experiments,
Variovorax DGGE fingerprinting clearly demonstrated the
proliferation of a particular Variovorax population in BMs
bioaugmented with linuron-primed soil L and fed with
linuron. This Variovorax population belonged to phylotype
A, which includes only linuron-degrading Variovorax strains
such as Variovorax SRS16 (Breugelmans et al., 2007). The
same phylotype A band proliferated in soil L when con-
tinuously fed with a linuron-containing solution (Bers et al.,
2011). The Variovorax 16S rRNA gene PCR products
recovered from these BMs were cloned but only some
randomly chosen clones were sequenced because the asso-
ciation of bands with specific phylotypes was demonstrated
previously (Bers et al., 2011). Nevertheless, BLAST analysis of
these sequences identified them all as Variovorax 16S rRNA
genes (E-value o 10�18) with sequences 100% identical to
those reported by Bers et al. (2011). The enrichment of
phylotype A Variovorax in these BMs was probably not
detected by the Variovorax real-time PCR due to masking
by the large number of Variovorax initially present in the
biomix substrata. These data suggest that linuron-degrading
Variovorax populations endogenous to the primed soil were
successfully transferred to the BM biomix. In addition they
suggest that strains belonging to phylotype A proliferated in
the BM upon linuron supply, thereby enhancing the linuron
degradation capacity. Because those populations only pro-
liferated in case linuron was supplied and because they could
be associated with a phylotype containing only linuron-
degrading Variovorax, it can be suggested that these popula-
tions are involved in linuron mineralization in the L1 set-
up. Conclusive evidence for this link can be provided using
techniques such as DNA/RNA-stable isotope probing (Du-
mont & Murrell, 2005). It cannot be excluded that bacteria
other than Variovorax were also involved in linuron degra-
dation. DGGE analysis of bacterial 16S rRNA genes was
performed but no differences were observed between set-ups
L1 and L� (data not shown).
Linuron is degraded through 3,4-dichloroaniline (3,4-
DCA). Several 3,4-DCA-degrading strains have been iso-
lated previously from soil L in co-culture with linuron-
degrading Variovorax strains and it was suggested that the
former proliferated by growth on 3,4-DCA leaking from the
linuron-degrading Variovorax strains (Breugelmans et al.,
2007). Some of these 3,4-DCA-degrading strains belonged
to Variovorax phylotype C. The band associated with
Variovorax phylotype C in the Variovorax DGGE profiles
migrates at position V2, but its occurrence in BMs of set-up
L1 could not be correlated with the appearance of the
linuron mineralization capacity. An explanation is that the
release of 3,4-DCA was limited during linuron degradation
in the biomix, hindering the proliferation of 3,4-DCA-
degrading bacteria or that, alternatively, 3,4-DCA degrada-
tion in the biomix was performed by other genera such as
Comamonas as observed in other linuron-mineralizing
bacteria consortia (Dejonghe et al., 2003).
No relationship was found between the linuron-miner-
alization capacity dynamics and Variovorax community
dynamics (neither in size nor in structure) in BMs of set-
up C1 in the first experiment. On the other hand, in
experiment 2, a Variovorax population related to phylotype
B (at position V1) appeared in the DGGE profile concomi-
tantly with the appearance of a linuron mineralization
capacity. Moreover, the Variovorax number increased con-
comitantly with the linuron mineralization capacity. Phylo-
type B includes the linuron-degrading Variovorax sp.
WDL1. However, in experiment 2, no set-up was included
without linuron feed and therefore the appearance of this
phylotype B cannot be linked to the observed increased
linuron mineralization capacity. Moreover, the presence of a
band at position V1 was not correlated with an increase in
linuron mineralization capacity in the first experiment. On
the other hand, based on the single PCR-DGGE results, the
proliferation of a Variovorax phylotype B and hence a
WDL1-related Variovorax population could be clearly asso-
ciated with the increased linuron mineralization capacity in
BM1 of set-up A11 because neither of the other two BM
replicas in this set-up showed enhanced linuron
FEMS Microbiol Ecol 76 (2011) 64–73 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
71Pesticide-primed soil to improve on-farm biopurification

mineralization capacity nor the DGGE band associated with
Variovorax phylotype B. However, this association was less
clear in the DGGE profiles obtained with the product of the
double-PCR approach. Because a WDL1-related population
was also detected in BMs without inoculum and in BMs
with nonprimed soil A1 (as shown in the double PCR-
DGGE approach) that did not develop a linuron-degrading
capacity, it can be suggested that different WDL1-related
populations were initially present in the straw and peat mix
on the one hand and in the soil inoculum on the other.
In conclusion, the results suggest that it is preferable to
use pesticide-primed soils containing endogenous pesticide-
mineralizing microorganisms over nonprimed soil for
bioaugmentation of on-farm BPS for several reasons. First,
although microorganisms in soils without previous pesti-
cide treatment can apparently develop a capacity to miner-
alize the pesticide used, this is not always the case. Second,
the initiation of mineralization of the target pesticide, in
case nonprimed soil is used, takes much more time com-
pared with a system inoculated with pesticide-primed soil.
This is important because mineralization should start as fast
as possible in a BPS to minimize leaching of pesticides
during the start-up period of the BPS. Furthermore, the
results show that Variovorax populations originating from
the added soil contribute to linuron degradation in the BPS
at least in the systems containing linuron-primed soil.
Acknowledgements
This research was supported by IWT-Vlaanderen Strategic
Basic Research project 73352 and IWT-Vlaanderen Agricul-
tural Research project LBO 040272.
References
Aslan S & Turkman A (2005) Combined biological removal of
nitrate and pesticides using wheat straw as substrates. Process
Biochem 40: 934–943.
Barbeau C, Deschenes L, Karamanev D, Comeau Y & Samson R
(1997) Bioremediation of pentachlorophenol-contaminated
soil by bioaugmentation using activated soil. Appl Microbiol
Biot 48: 745–752.
Bazot S & Lebeau T (2008) Simultaneous mineralization of
glyphosate and diuron by a consortium of three bacteria as
free- and/or immobilized-cells formulations. Appl Microbiol
Biot 77: 1351–1358.
Bers K, Sniegowski K, Breugelmans P, Hendrickxs L, Mot RD &
Springael D (2011) A molecular toolbox to estimate the
number and diversity of Variovorax in the environment:
application in soils treated with the phenylurea herbicide
linuron. FEMS Microbiol Ecol DOI: 10.1111/j.1574-
6941.2010.01028.x.
Breugelmans P, D’huys PJ, De Mot R & Springael D (2007)
Characterization of novel linuron-mineralizing bacterial
consortia enriched from long-term linuron-treated
agricultural soils. FEMS Microbiol Ecol 62: 374–385.
Breugelmans P, Horemans B, Hofkens J & Springael D (2010)
Response of the structure and activity of a linuron-degrading
triple-species biofilm to mixed substrate feeds. Res Microbiol
161: 660–666.
Briones AM & Reichardt W (1999) Estimating microbial
population counts by ‘most probable number’ using Microsoft
Excels. J Microbiol Meth 35: 157–161.
Broos K, Mertens J & Smolders E (2005) Toxicity of heavy metals
in soil assessed with various soil microbial and plant growth
assays: a comparative study. Environ Toxicol Chem 24:
634–640.
Chatterjee DK, Kilbane JJ & Chakrabarty AM (1982)
Biodegradation of 2,4,5-trichlorophenoxyacetic acid in soil by
a pure culture of Pseudomonas cepacia. Appl Environ Microb
44: 514–516.
Coppola L, Castillo MD, Monaci E & Vischetti C (2007)
Adaptation of the biobed composition for chlorpyrifos
degradation to Southern Europe conditions. J Agr Food Chem
55: 396–401.
Cullington JE & Walker A (1999) Rapid biodegradation of diuron
and other phenylurea herbicides by a soil bacterium. Soil Biol
Biochem 31: 677–686.
Dejonghe W, Berteloot E, Goris J, Boon N, Crul K, Maertens S,
Hofte M, De Vos P, Verstraete W & Top EM (2003) Synergistic
degradation of linuron by a bacterial consortium and isolation
of a single linuron-degrading Variovorax strain. Appl Environ
Microb 69: 1532–1541.
De Wilde T, Spanoghe P, Debaer C, Ryckeboer J, Springael D &
Jaeken P (2007) Overview of on-farm bioremediation systems
to reduce the occurrence of point source contamination. Pest
Manag Sci 63: 111–128.
Dumont MG & Murrell JC (2005) Stable isotope probing –
linking microbial identity to function. Nat Rev Microbial 3:
499–504.
El Fantroussi S, Verschuere L, Verstraete W & Top EM (1999)
Effect of phenylurea herbicides on soil microbial communities
estimated by analysis of 16S rRNA gene fingerprints and
community-level physiological profiles. Appl Environ Microb
65: 982–988.
Fogg P, Boxall ABA & Walker A (2003a) Degradation of pesticides
in biobeds: the effect of concentration and pesticide mixtures.
J Agr Food Chem 51: 5344–5349.
Fogg P, Boxall ABA, Walker A & Jukes AA (2003b) Pesticide
degradation in a ‘biobed’ composting substrate. Pest Manag
Sci 59: 527–537.
Fogg P, Boxall ABA, Walker A & Jukes A (2004) Degradation and
leaching potential of pesticides in biobed systems. Pest Manag
Sci 60: 645–654.
Grigg BC, Assaf NA & Turco RF (1997) Removal of atrazine
contamination in soil and liquid systems using
bioaugmentation. Pestic Sci 50: 211–220.
Grundmann S, Fuss R, Schmid M, Laschinger M, Ruth B, Schulin
R, Munch JC & Schroll R (2007) Application of microbial hot
FEMS Microbiol Ecol 76 (2011) 64–73c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
72 K. Sniegowski et al.

spots enhances pesticide degradation in soils. Chemosphere 68:
511–517.
Haest PJ, Philips J, Springael D & Smolders E (2010) The reactive
transport of trichloroethene is influenced by residence time
and microbial numbers. J Contam Hydrol 119: 89–98.
Henriksen VV, Helweg A, Spliid NH, Felding G & Stenvang L
(2003) Capacity of model biobeds to retain and degrade
mecoprop and isoproturon. Pest Manag Sci 59: 1076–1082.
Johannesen H, Sorensen SR & Aamand J (2003) Mineralization of
soil-aged isoproturon and isoproturon metabolites by
Sphingomonas sp strain SRS2. J Environ Qual 32: 1250–1257.
Johnsen A, Schmidt S, Hybholt T, Henriksen S, Jacobsen C &
Andersen O (2007) Strong impact on the polycyclic aromatic
hydrocarbon (PAH)-degrading community of a PAH-polluted
soil but marginal effect on PAH degradation when priming
with bioremediated soil dominated by mycobacteria. Appl
Environ Microb 73: 1474–1480.
Mertens B, Boon N & Verstraete W (2006) Slow-release
inoculation allows sustained biodegradation of gamma-
hexachlorocyclohexane. Appl Environ Microb 72: 622–627.
Moran AC, Muller A, Manzano M & Gonzalez B (2006) Simazine
treatment history determines a significant herbicide
degradation potential in soils that is not improved by
bioaugmentation with Pseudomonas sp. ADP. J Appl Microbiol
101: 26–35.
Muyzer G, Dewaal EC & Uitterlinden AG (1993) Profiling of
complex microbial populations by denaturing gradient gel-
electrophoresis analysis of polymerase chain reaction-
amplified genes coding for 16S ribosomal-RNA. Appl Environ
Microb 59: 695–700.
Pigeon O, De Vleeschouwer C, Cors F, Weickmans B, De Ruyckel
B, Pussemier L, Debognie P & Culot M (2005) Development of
biofilters to treat the pesticide wastes from spraying
applications. Comm Agr Appl Biol Sci 70: 1003–1012.
Runes HB, Jenkins JJ & Bottomley PJ (2001) Atrazine
degradation by bioaugmented sediment from constructed
wetlands. Appl Microbiol Biot 57: 427–432.
Singh BK, Walker A & Wright DJ (2006) Bioremedial potential of
fenamiphos and chlorpyrifos degrading isolates: influence of
different environmental conditions. Soil Biol Biochem 38:
2682–2693.
Sniegowski K, Mertens J, Diels J, Smolders E & Springael D
(2009) Inverse modeling of pesticide degradation and
pesticide-degrading population size dynamics in a
bioremediation system: parameterizing the Monod model.
Chemosphere 75: 726–731.
Sorensen SR, Rasmussen J, Jacobsen CS, Jacobsen OS, Juhler RK
& Aamand J (2005) Elucidating the key member of a linuron-
mineralizing bacterial community by PCR and reverse
transcription-PCR denaturing gradient gel electrophoresis 16S
rRNA gene fingerprinting and cultivation. Appl Environ
Microb 71: 4144–4148.
Struthers JK, Jayachandran K & Moorman TB (1998)
Biodegradation of atrazine by Agrobacterium radiobacter J14a
and use of this strain in bioremediation of contaminated soil.
Appl Environ Microb 64: 3368–3375.
Topp E (2001) A comparison of three atrazine-degrading bacteria
for soil bioremediation. Biol Fert Soils 33: 529–534.
Torstensson L & del Pilar Castillo M (1997) Use of biobeds in
Sweden to minimize environmental spillages from agricultural
spraying equipment. Pesticide Outlook 8: 24–27.
Uyttebroek M, Breugelmans P, Janssen M et al. (2006) Distribution
of the Mycobacterium community and polycyclic aromatic
hydrocarbons (PAHs) among different size fractions of a
long-term PAH-contaminated soil. Environ Microbiol 8:
836–847.
Vischetti C, Capri E, Trevisan M, Casucci C & Perucci P (2004)
Biomassbed: a biological system to reduce pesticide point
contamination at farm level. Chemosphere 55: 823–828.
Zhao SH, Arthur EL & Coats JR (2003) Influence of microbial
inoculation (Pseudomonas sp strain ADP), the enzyme atrazine
chlorohydrolase, and vegetation on the degradation of
atrazine and metolachlor in soil. J Agr Food Chem 51:
3043–3048.
FEMS Microbiol Ecol 76 (2011) 64–73 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
73Pesticide-primed soil to improve on-farm biopurification
Related Documents











