Implications of seasonal priming and reproductive activity on the interpretation of Comet assay data derived from the clam, Tapes semidecussatus Reeves 1864, exposed to contaminated sediments M.G.J. Hartl a,b , B.M. Coughlan a,b , D. Sheehan a,c , C. Mothersill a,d , F.N.A.M. van Pelt a,e , S.J. O’Reilly a,f , J.J.A. Heffron a,f , J. O’Halloran a,b , N.M. O’Brien a,g, * a Environmental Research Institute, University College Cork, Ireland b Department of Zoology, Ecology & Plant Science, University College Cork, Ireland c Department of Biochemistry, University College Cork, Ireland d Radiation and Environmental Science Centre, Dublin Institute of Technology, Dublin, Ireland e Department of Pharmacology & Therapeutics, University College Cork, Ireland f Biochemical Toxicology Laboratory, Department of Biochemistry, University College Cork, Ireland g Department of Food and Nutritional Sciences, University College Cork, Ireland Received 9 January 2003; received in revised form 12 October 2003; accepted 19 November 2003 Abstract We explore the use of the clam Tapes semidecussatus Reeves 1864 as an indicator for the presence of potentially genotoxic substances in estuarine sediments. The limitations associated with the interpretation of Comet assay data (expressed as % DNA in tail) in terms of clam reproductive state, size (age) and thermal exposure history following laboratory acclimation are discussed. Hatchery-reared clams, subjected to ambient temperature fluctuations during growth, were exposed in vivo under laboratory conditions for three weeks to sediment sam- ples collected from a polluted site and a ‘‘clean’’ reference site. The DNA damage observed in haemocytes, gill and digestive gland cells was significantly higher in animals exposed to contaminated sediment compared to those exposed to sediment from the reference site. The extent of DNA damage recorded was not correlated with size (age). Spawning was not Marine Environmental Research 57 (2004) 295–310 www.elsevier.com/locate/marenvrev 0141-1136/$ - see front matter # 2003 Elsevier Ltd. All rights reserved. doi:10.1016/j.marenvres.2003.11.004 * Corresponding author. Tel.: +353-21-490-2884; fax: +353-21-427-0244. E-mail address: [email protected] (N.M. O’Brien).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Implications of seasonal priming andreproductive activity on the interpretation ofComet assay data derived from the clam,
Tapes semidecussatus Reeves 1864, exposed tocontaminated sediments
M.G.J. Hartla,b, B.M. Coughlana,b, D. Sheehana,c,C. Mothersilla,d, F.N.A.M. van Pelta,e, S.J. O’Reillya,f,J.J.A. Heffrona,f, J. O’Hallorana,b, N.M. O’Briena,g,*
aEnvironmental Research Institute, University College Cork, IrelandbDepartment of Zoology, Ecology & Plant Science, University College Cork, Ireland
cDepartment of Biochemistry, University College Cork, IrelanddRadiation and Environmental Science Centre, Dublin Institute of Technology, Dublin, Ireland
eDepartment of Pharmacology & Therapeutics, University College Cork, IrelandfBiochemical Toxicology Laboratory, Department of Biochemistry, University College Cork, Ireland
gDepartment of Food and Nutritional Sciences, University College Cork, Ireland
Received 9 January 2003; received in revised form 12 October 2003; accepted 19 November 2003
Abstract
We explore the use of the clam Tapes semidecussatus Reeves 1864 as an indicator for thepresence of potentially genotoxic substances in estuarine sediments. The limitations associatedwith the interpretation of Comet assay data (expressed as % DNA in tail) in terms of clam
reproductive state, size (age) and thermal exposure history following laboratory acclimationare discussed. Hatchery-reared clams, subjected to ambient temperature fluctuations duringgrowth, were exposed in vivo under laboratory conditions for three weeks to sediment sam-
ples collected from a polluted site and a ‘‘clean’’ reference site. The DNA damage observedin haemocytes, gill and digestive gland cells was significantly higher in animals exposed tocontaminated sediment compared to those exposed to sediment from the reference site. The
extent of DNA damage recorded was not correlated with size (age). Spawning was not
Marine Environmental Research 57 (2004) 295–310
www.elsevier.com/locate/marenvrev
0141-1136/$ - see front matter # 2003 Elsevier Ltd. All rights reserved.
doi:10.1016/j.marenvres.2003.11.004
* Corresponding author. Tel.: +353-21-490-2884; fax: +353-21-427-0244.
E-mail address: [email protected] (N.M. O’Brien).

observed during the experiment. Nevertheless, clams with well-developed gonads showed a
statistically higher degree of DNA damage in gill and digestive gland cells- but not haemocytes,demonstrating an increased sensitivity to potential genotoxic compounds, possibly caused byimpaired DNA repair capacity due to reproductive activity. Furthermore, the degree of DNA
damage in clams exposed to contaminated sediments was higher in autumn and wintercompared to spring and summer, suggesting an effect of seasonal priming.# 2003 Elsevier Ltd. All rights reserved.
Keywords: Comet assay; Tapes semidecussatus; Sediment contamination; Reproductive and thermal
effects; DNA damage
1. Introduction
The single cell gel electrophoresis test (Comet assay) has been successfully adaptedto various aquatic organisms, including fish (Belpaeme, Cooreman, & Kirsch-Volders,1998; Devaux, Pesonen, & Monod, 1997; Mitchelmore & Chipman, 1998), mussels(Pavlica, Klobucar, Mojas, Erben, & Papes, 2001; Rank, 1999; Steinert, Streib-Montee,Leather, & Chadwick, 1998) and clams (Coughlan et al., 2002; Hamoutene, Payne,Rahimtula, & Lee, 2002). Most applications of the Comet assay to aquatic organismshave focussed on metals and organic compounds either in solution or aqueoussuspension. Recently, however, estuarine sediments have become an area of growingconcern, due to their potential for accumulating toxic compounds and acting as asecondary source of contaminant exposure to benthic fauna (Baumann, 1998;Edwards, Edyvane, Boxall, Hamann, & Soole, 2001; French et al., 1996; Stein, Hom,Collier, Brown, & Varanasi, 1995). Accordingly, both in vitro (Kammann, Riggers,Theobald, & Steinhart, 2000) and in vivo (Coughlan et al., 2002) adaptations of theComet assay for the assessment of sediment genotoxicity have been reported. Thelatter was a laboratory study designed to assess the suitability of the estuarine-dwelling Manila clam, Tapes semidecussatus Reeves 1864, as a biomarker organismfor the presence of potentially genotoxic agents in estuarine sediments. In anyecotoxicology assessment, knowledge of the contaminant pre-exposure history of thetest organism is paramount (Boisson, Hartl, Fowler, & Amiard-Triquet, 1998).Therefore, as in the present study, farmed rather than wild clams were obtainedfrom a commercial hatchery. However, these were exposed to ambient fluctuations intemperature and salinity during growth and development. Coughlan et al. (2002) didnot address the potential influence of seasonality and reproduction of clams on thelevel of genotoxic insult. The reproductive cycle of clams, which is linked to ambienttemperature, has been shown to modulate biomarker expression and influence theirsusceptibility to some contaminants (Blaise, Gagne, Pellerin, Hansen, & Trottier, 2002).
In the present laboratory study, using the Comet assay, we investigate if and towhat extent the reproductive state and size (age) of T. semidecussatus can influencethe occurrence of single-strand breaks in DNA. Furthermore, we explore thepotential effects of age, reproductive state and thermal exposure history on DNAdamage sustained by clams during exposure to field-collected estuarine sedimentsunder controlled laboratory conditions.
296 M.G.J. Hartl et al. /Marine Environmental Research 57 (2004) 295–310
2. Materials and methods
2.1. Site characteristics
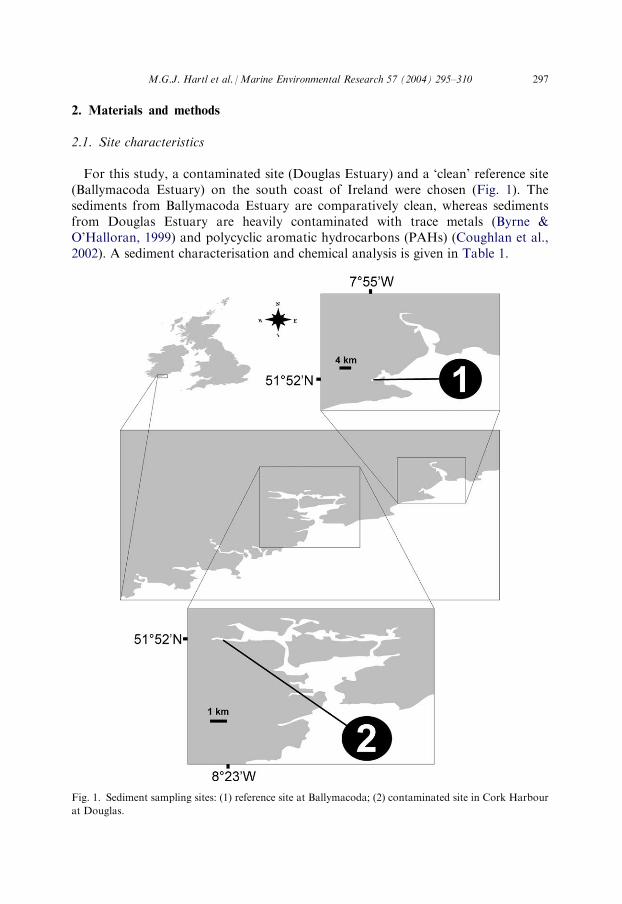
For this study, a contaminated site (Douglas Estuary) and a ‘clean’ reference site(Ballymacoda Estuary) on the south coast of Ireland were chosen (Fig. 1). Thesediments from Ballymacoda Estuary are comparatively clean, whereas sedimentsfrom Douglas Estuary are heavily contaminated with trace metals (Byrne &O’Halloran, 1999) and polycyclic aromatic hydrocarbons (PAHs) (Coughlan et al.,2002). A sediment characterisation and chemical analysis is given in Table 1.
Fig. 1. Sediment sampling sites: (1) reference site at Ballymacoda; (2) contaminated site in Cork Harbour
at Douglas.
M.G.J. Hartl et al. /Marine Environmental Research 57 (2004) 295–310 297

2.2. Collection and preparation of sediment
Prior to each experiment, approximately 40 kg of surface sediment (the topoxygenated 0.5–1 cm) were collected at low tide in summer, autumn and winter,2001 and spring, 2002 from both sites using a plastic trowel. Sediment chemistry andthe bioavailability of sediment-associated contaminants have been shown to dependon sampling and treatment conditions (Luoma & Ho, 1998) and can also vary withseason (Chong & Wang, 2000, 2001). Accordingly, following each sampling eventthe collected sediment was thoroughly mixed until it was of a consistent colour, thusinsuring a homogenous chemical contaminant distribution for each separate seasonalexperiment (Werner, Kline, & Hollibaugh, 1998). The samples were transported tothe laboratory, where 100 g sub-samples were immediately frozen for chemicalanalysis and the rest stored at 4 �C for up to 96 h before starting the exposureexperiments.
2.3. Clams
In each season, Manila clams, T. semidecussatus, of known history (1+ age class)were obtained from Cartron Point Shellfish Ltd, a commercial hatchery in a remotearea of County Clare on the west coast of Ireland. The animals were transported tothe laboratory and acclimated in aerated seawater (30�2%) for up to 2 weeks prior
Table 1
Sediment characterisation and analysis for selected chemical classes in sediment samples collected from
Ballymacoda Estuary (‘clean’ site) and Douglas Estuary (contaminated site)
Site characteristics
Sediment samplesBallymacoda Estuary
Douglas EstuaryOrg. carbon (%)
1.8 0.98<63 mm fraction (%)
36 54PAHs (ng g�1)
Fluorene
46.04 1120Phenanthrene
64.6 116.5Pyrene
23.65 493.9Benzo(a)anthracene
41.3 177.3Chyrsene
29.8 117.9Benzo(b)fluoranthene
7.6 1.2Benzo(k)fluoranthene
30.2 62.3Benzo(a)pyrene
14.58 244.1Metals (mg kg�1)
Meana �S.D. Meana �S.D.Copper
8.1 1.1 54.3 2.2Lead
15.6 1.8 51.8 5.1Cadmium
0.3 0.03 0.64 0.07Zinc
78.6 9.4 214.1 10.5a Mean value obtained from analyses of six sediment samples.
298 M.G.J. Hartl et al. /Marine Environmental Research 57 (2004) 295–310

to exposure to sediments. The average weight of individuals used varied with season:spring 1.53 g�0.6, summer 1.26 g�0.3, autumn 3.54 g�0.3 and winter 0.66 g�0.2.
2.4. Exposure of clams
The in vivo experiments were conducted in a controlled environment(15.0�0.5 �C; 12 h light/dark cycle). One kg aliquots (two replicates for eachsampling site) of sediment was added to 2 l of seawater (collected from Ballycotton,County Cork, an area with no industrial activity; S=30�2%) in a 5 l acid-washedpolyethylene aquarium, and allowed to settle over-night. Once the sediment sampleshad settled, 15 clams were randomly selected from a stock population and gentlyplaced onto the sediment surface in each aquarium. An additional 10 clams wererandomly selected from the stock population, sacrificed and single-cell suspensionsisolated from haemolymph, gill and digestive gland tissue to establish backgroundlevels of DNA damage at time 0 days (t0) of exposure. At 7-day intervals (t7, t14, t21)over the following 3 weeks, 10 clams were taken from each sediment type (tworeplicates of five clams) and the single-cell suspensions prepared for the Comet assayas for t0. The data from each treatment group were pooled for each time period andcell type and subjected to statistical analysis (see below).
The seawater in the tanks was continuously aerated and changed once a weekwhile carefully avoiding re-suspension of the sediments.
2.5. Isolation of single cell suspensions
The Comet Assay was used to analyse the haemocytes (to estimate the overallcondition of the clam), in conjunction with the gill and digestive gland tissues fromthe same animal enabling the study of potential effects on DNA stability covering allroutes of exposure.
Haemolymph was collected from both the posterior and anterior adductormuscles using a hypodermic syringe (Pavlica, Klobucar, Vetma, Erben, & Papes,2000) in an equal volume of osmotically corrected Hank’s balanced salt solution(HBSS; pH 7.4; 1000 mOsmol kg�1). Single-cell suspensions of gill and digestivegland tissue were obtained from the same animals using a method adapted fromWilson, Pascoe, Parry, and Dixon (1998) and described in detail in Coughlan et al.(2002). Briefly, each organ was excised and chopped separately 10 times in HBSSand incubated in trypsin (final conc. 0.05%; 1000 mOsm l�1) for 10 min at roomtemperature. The homogenates were passed through a 40-mm sieve, centrifuged at800g for 5 min, the supernatant discarded and the pellet carefully resuspended in 0.5ml of HBSS. Finally, cell viability was tested by means of the trypan-blue exclusionmethod (Absolom, 1986).
Well-developed gonads were clearly visible in clams collected in autumn, but nospawning occurred during the acclimation period or the experiment. In order to identifypotential effects of reproductive activity on the Comet assay data, the values fromclams with well-developed gonads were compared with those from clams at an earlierstage in their reproductive cycle, when gonads were not or less well-developed.
M.G.J. Hartl et al. /Marine Environmental Research 57 (2004) 295–310 299

Care was taken not to disrupt the gonads during dissection, and as a precaution,excised gill and digestive gland tissues were rinsed thoroughly in HBSSto remove any gametes from the surface prior to the preparation of single-cellsuspensions.
2.6. Comet assay
The Comet procedure was performed according to a protocol developed in ourlaboratory by Woods, O’Leary, McCarthy, and O’Brien (1999). Briefly, cells weresandwiched in 1% low-melting agarose onto frosted microscope slides, lysed in ahigh salt buffer [2.5M NaCl, 10 mM Tris, 100 mM EDTA, 1% (v/v) Triton X-100and 10% (v/v) DMSO, pH 10.0, in the dark at 4 �C] for 90 min, immersed in analkaline solution (0.3 M NaOH, 1 mM EDTA; pH>12) for 30 min at 4 �C in orderfor the DNA to unwind and a current (25 V, 300 mA) applied for 25 min in ahorizontal electrophoresis tank. The pH was neutralized with Tris buffer (0.4 MTris–HCl, pH 7.4), the DNA stained with ethidium bromide and the nucleoidsanalysed using an epifluorescence microscope (Nikon EFD-3). DNA damage wasexpressed as % tail DNA and determined using the imaging analysis softwarepackage Komet 4.0. (Kinetic Imaging Ltd).
2.7. Data analyses
The data from clams exposed to sediments from the two sites were analysed usinga one-way ANOVA followed by a Post-hoc Tukey pair-wise multiple comparisontest. The interaction of season and reproductive development stage with site wasassessed using a two-way ANOVA followed by a Post-hoc Tukey pair-wise multiplecomparison test. The influence of animal size was analysed by linear regression. AP<0.05 was considered significant.
3. Results
3.1. Cell viability
The cell viability for each tissue type was recorded for every single cell suspension,resulting in cell viabilities of over 85% for all cell types tested (Table 2) andremained stable for at least 90 min.
3.2. Comet assay results expressed as % tail DNA
3.2.1. Effect of sizeThe size of the clams depended on age and ranged from 0.34 to 6.10 g. There
was no correlation between size and % tail DNA in blood, gill or digestive glandcells in clams exposed to sediment from Ballymacoda Estuary or Douglas Estuary,respectively (Fig. 2).
300 M.G.J. Hartl et al. /Marine Environmental Research 57 (2004) 295–310

3.2.2. Effect of season and site on haemocytesThe background % tail DNA values in haemocytes, measured before exposure to
sediments (t0), in spring, summer, autumn and winter (Fig. 3a–d) did not differsignificantly.
Following 7 days of exposure (t7) in the laboratory to collected sediment from thereference and contaminated sites in spring and summer,% tail DNA values did notchange and there was also no difference between the two sites (Fig. 3a and b). Inautumn and winter the % tail DNA values at t7 for the contaminated site hadincreased significantly above both the reference site and the initial values at t0(Fig. 3c and d).
Table 2
Cell viability using the Trypan-Blue exclusion method for each cell type excised from clams, following the
preparation of single-cell suspensions
Season
Time(days)Cell type (% viable cells)
Blood
Gills Digestive glandB
D B D B DSpring
0 94 96 95 94 96 897
98 95 94 95 95 93 14 98 97 92 92 91 95 21 94 96 91 92 94 95Average
96 96 93 93 94 93 S.D. 2.31 0.82 1.83 1.50 2.16 2.83Summer
0 99 96 86 87 88 94 7 100 98 91 88 90 8814
100 95 86 89 87 89 21 95 97 87 89 90 95 Average 98 96 87 88 88 91S.D.
2.38 1.29 2.38 0.96 1.50 3.51Autumn
0 97 94 94 87 87 94 7 98 98 86 86 87 9014
96 N.D. 86 N.D. 96 N.D. 21 95 94 85 85 85 90 Average 96 95 87 86 88 91S.D.
1.29 2.31 4.19 1.00 4.92 2.31Winter
0 88 95 88 85 87 907
87 95 90 89 89 94 14 94 97 87 85 91 95 21 96 95 86 90 92 88Average
91 95 87 87 89 91 S.D. 4.43 1.00 1.71 2.63 2.22 3.30B: Reference site (Ballymacoda); D: polluted site (Douglas estuary); N.D.: no data
M.G.J. Hartl et al. /Marine Environmental Research 57 (2004) 295–310 301

After 14 days of exposure (t14), % tail DNA values for the contaminated sedimentin spring, summer, autumn and winter increased significantly above the initialbackground values and were also significantly higher than the respective values forthe reference sediment (Fig. 3a–d). In spring, summer and autumn % tail DNAfor both the contaminated and reference sediments on day 21 of exposure (t21)remained unchanged from those at t14 (Fig. 3a–c). However, in winter,% tail DNAin clams exposed in the laboratory to both contaminated and reference sedimentincreased significantly (Fig. 3d).
Fig. 2. Relationship between% DNA in tail and total clam wet weight.
302 M.G.J. Hartl et al. /Marine Environmental Research 57 (2004) 295–310

3.2.3. Effect of season and site on gill cellsBackground values for % tail DNA from gill single-cell preparations measured at
t0 for spring, summer, autumn and winter (Fig. 3e–h) did not differ significantly.Following 7 days of exposure (t7) to contaminated sediments, the % tail DNA
values in spring, summer, autumn and winter were significantly elevated above thebackground and the respective reference site values.
The % tail DNA values at t14 and t21 for the sediments from the contaminated siteremained above t0 and the reference site. During the entire experiment, % tail DNAvalues for gills from clams exposed to sediments from the reference site did notincrease above background levels.
Fig. 3. Degree of seasonal DNA damage expressed as % DNA in tail recorded in cells isolated from
haemolymph (a–d), gills (e–h) and digestive gland (i–l) following chronic exposure of the Manila clam
(Tapes semidecussatus) to sediment collected from a contaminated site, Douglas, and a reference site,
Ballymacoda, County Cork, Ireland; mean�S.E.; n=10; (*): significant difference (P<0.05) between
Douglas and Ballymacoda at time tn; (y): significant difference (P<0.05) between Douglas or reference
site at tn and initial values at t0.
M.G.J. Hartl et al. /Marine Environmental Research 57 (2004) 295–310 303

3.2.4. Effect of season and site on digestive gland cellsBackground values for % tail DNA from gill single-cell preparations measured at
t0 for spring, summer, autumn and winter (Fig. 3i–l) did not differ significantly.The % tail DNA values at t7 in spring, summer, autumn and winter for the
contaminated site increased above those of t0 and where also significantlyhigher than the respective reference site values. No change was observed att14 for the % tail DNA values from the polluted site in spring and summer.The % tail DNA values at t21 for autumn decreased and those for summer andwinter increased; however, they all remained significantly higher than the t0 and thereference site values.
Fig. 4. Effect of developmental stage on the size of % DNA in tail, in haemocytes, gill and digestive gland
cells. (*) significant difference (P<0.05) between clams with and without well-developed gonads.
304 M.G.J. Hartl et al. /Marine Environmental Research 57 (2004) 295–310

3.2.5. Effect of developmental stage and site on haemocytesAt t0 there was no significant difference in % tail DNA in haemocytes between
clams with and without well-defined gonads, indicating that the background valueswere independent of gonad development (Fig. 4a). Further examination of thehaemocyte data at t7, t14 and t21 revealed that the presence of gonads showed noeffect on the % tail DNA values of haemocytes in clams exposed to contaminatedand reference site sediment (Fig. 4a and b).
3.2.6. Effect of developmental stage and site on gill cellsAt t0 % tail DNA in gill cells showed no significant differences between indivi-
duals with and without gonads (Fig. 4c). At t7, t14 and t21 the presence of well-developed gonads had no effect on % tail DNA in animals exposed to sedimentsfrom the reference site. However, respective % tail DNA values from animalsexposed to contaminated sediments were significantly higher in animals with visiblegonads (Fig. 4c and d).
3.2.7. Effect of developmental stage and site on digestive gland cellsAt t0, there was no significant difference in % tail DNA values of digestive gland
cells between clams with or without gonad development (Fig. 4e). At t7, t14 and t21the % tail DNA values for clams with gonads exposed to contaminated sedimentwas significantly higher than those without (Fig. 4f), whereas the clams from thereference site remained unchanged (Fig. 4e).
4. Discussion
In vivo ecotoxicological testing generally requires the use of a well-defined popu-lation of test organisms of similar size, age, condition and contaminant exposurehistory (Boisson et al., 1998; Steinert, 1999). In this study, the weight of the clams,supplied by a hatchery, varied with season. Work carried out by Giovannelli,Decorosi, Dolara, and Pulvirenti (2003) on rats and Pruski and Dixon (2003) on thevent mussels, Bathymodiolus azoricus suggested that the DNA repair capacity maybereduced with size (age). Nonetheless, the lack of correlation between animal weightand % tail DNA in blood, gill and digestive gland cells in clams suggests that, withinthe groups studied, the degree of both background and DNA damage observedfollowing exposure to sediment-associated contaminants were independent of size(age) (Fig. 2). This is supported by Wilson et al. (1998), who found no relationshipbetween size (age) and DNA damage in gill cell preparations from the musselMytilus edulis L. exposed to H2O2 in vivo.
In order to evaluate all routes of exposure, single-cell suspensions derived fromthe haemolymph were used, as well as gill and digestive gland tissue from the sameanimal. The respective physical and biochemical extraction procedures were adaptedfrom Wilson et al. (1998) and Pavlica et al. (2000) and refined by Coughlan et al.(2002). The consistently low background values at t0 (Figs. 3a–l) and high cellviabilities (Table 2) within the different cell suspensions indicates that the level of
M.G.J. Hartl et al. /Marine Environmental Research 57 (2004) 295–310 305

artefactual DNA damage due to handling recorded in clams exposed to referenceand contaminated sediment was kept at a reproducible minimum. These values aresimilar to those by Pruski and Dixon (2002) for M. edulis, who reported controlvalues of 12% and 20% DNA in the tail for blood and gill cells, respectively.Mitchelmore, Birmelin, Chipman, and Livingstone (1998) reported values of 10–16% DNA in the tail for digestive gland cells, also from M. edulis, which were lowerthan the background values in the present study (Fig. 3i–l). Although the use oftrypsin for the preparation of gill single-cell suspensions did not seem to have aneffect on background levels, this may not be the case for digestive gland cells andcould explain the higher background DNA damage observed in this study (Birmelin,Mitchelmore, Goldfarb, & Livingstone, 1998).
Unlike in vitro work with cell lines, where the cells analysed are clones, single cellsuspensions derived from complex tissues are composed of a variety of cell types.Molluscan gills are composed of columnar, epithelial, squamous endothelial, andmucous cells (Carpence & George, 1981; Domouhtsidou & Dimitriadis, 2000). Thedigestive gland of bivalves is a paired gland consisting of numerous blind endingtubules, the epithelium of which is composed of basophilic and digestive cells(Domouhtsidou & Dimitriadis, 2000). Both organs contain essentially the samedetoxification enzyme systems, whereby the level of expression and activity isgenerally much higher in the digestive gland due to its role as the primary siteof xenobiotic metabolism (Doyotte, Cossu, Jacquin, Babut, & Vasseur, 1997;Mitchelmore et al., 1998). However, the Comet assay, in the present approach, wasunable to distinguish between differential effects of endogenous enzyme activity inindividual cell types due to the complex nature of the cell suspensions.
The sediments from the contaminated site at Douglas contained elevated levels ofboth metals and polycyclic aromatic hydrocarbons (PAHs) (Table 1). Heavy metalshave a tendency to bind to phosphates and a wide variety of organic molecules,including base residues of DNA, which can lead to mutations by altering primaryand secondary structures of the DNA (Wong, 1988). PAHs can cause DNA damagein marine invertebrates either by direct DNA strand breakage or are metabolizedinto reactive intermediates that form unstable DNA adducts (Nacci, Nelson,Nelson, & Jackim, 1992; Nacci, Stephanie, & Jackim, 1996; Mitchelmore, et al.,1998). Both processes are likely to trigger DNA excision repair mechanisms incurringDNA fragments detectable with the Comet assay (Collins, Dobson, Dusinska,Kennedy, & Stetina, 1997; Speit & Hartmann, 1995) and thus indicate clam interactionwith potential genotoxic compounds.
Significantly higher levels of DNA strand breaks were found in the present studyin clams with well-developed gonads (Fig. 4), notably in gill and digestive gland tis-sue preparations. In the rare cases during the preparation of single-cell suspensionswhere the gonads were damaged, the majority of oocytes were retained by thefiltration step (40 mm) (Birmelin, Pipe, Goldfarb, & Livingstone, 1999). This issupported by Xie and Burnell (1994) who demonstrated that 85% of the oocytes inripe females of T. semidecussatus were >35 mm in diameter with an average of 42.5mm. The occasional presence of oocytes was unlikely to have biased the results, asoocytes were easily distinguishable (much larger) from somatic cells and were
306 M.G.J. Hartl et al. /Marine Environmental Research 57 (2004) 295–310

therefore not measured. Spermatozoa require special treatment prior to analysis andtheir presence in Comet preparations is therefore likely to under estimate potentialgenotoxic damage (Frenzilli, Nigro, Scarcelli, Gorbi, & Regoli, 2001). The precau-tionary rinse of excised tissue during preparation was sufficient to remove mostspermatozoa. However, fewer than 1% slides were found to contain obviousspermatozoon nuclei (much smaller) and were therefore discarded.
Blaise et al. (2002), studying the soft-shell clam Mya arenaria exposed to multiplesources of contamination, reported vitellogenesis and spawning to be positivelycorrelated with DNA damage. In southern Irish waters T. semidecussatus ischaracterised by a unimodal gametogenesis cycle, with the onset of gametogenesis inspring and major spawning occurring in late autumn (Xie & Burnell, 1994). Sastry(1966) and Sastry and Blake (1971) showed that, during gametogenesis in the scallopAequipecten irradians, material stored in the digestive gland tissue is transferred tothe gonad. The diversion of stored energy to gametogenesis is likely to leave lessenergy for somatic growth and baseline metabolic maintenance processes such asDNA repair. Although DNA repair itself was not determined in the present study,this may explain in part the elevated susceptibility of ripe clams to DNA strandbreaks when exposed to contaminated sediment.
During the hatchery rearing process, clams were exposed to natural local fluctua-tions in temperature and salinity and therefore required acclimation to laboratoryconditions (15 �C, S=32) before the commencement of experiments. The averageseawater temperature on the west coast of Ireland varies with season: 7 �C (Spring),9 �C (Summer), 14 �C (Autumn) and 12 �C (Winter) (Briton, 1991). All three celltypes from animals exposed to contaminated sediments showed a clear trendtowards an increase in single-strand breaks in autumn and winter (Fig. 3). Similarly,using the Comet assay, Frenzilli et al. (2001) described a seasonal effect on the DNAintegrity of Mytilus galloprovincialis collected from a highly eutrophic coastallagoon. The DNA integrity and the antioxidant scavenging efficiency in their studywere both inversely correlated with seawater temperature. The clams in the presentstudy were acclimated to 15 �C for 2 weeks prior to and not fed during the experi-ments—clams were assumed to be feeding on detritus contained in the top layer ofsediment (Byrne & O’Halloran, 2000). The observed ‘‘seasonal’’ differences in DNAstrand breaks between the cold (7–9 �C) and the warm (12–14 �C) pre-conditionedclams may be due to paradoxical or reverse acclimation; a phenomenon described byBerg (1953) as a response of aquatic invertebrates to low food availability in winter,when their metabolism is depressed in an attempt to conserve energy. Wilson andElkaim (1997) identified a considerable degree of reverse acclimation as the maincause of observed lower oxygen consumption rates of summer-conditioned individualsof Cerastoderma glaucum, compared to those of winter-conditioned specimens whenheld at the same experimental temperature.
Seasonality has a profound effect on sediment chemistry and bioavailability ofcontaminants (Del Valls, Forja, & Gomez-Parra, 2002; Rice, Seltenrich, Spies, &Keller, 1993). The sediments were collected and thoroughly homogenised prior touse in laboratory experiments in spring, summer, autumn and winter. Althoughthese experiments are not a representation of the natural environment as the process
M.G.J. Hartl et al. /Marine Environmental Research 57 (2004) 295–310 307

of mixing broke down redox zones within the sediment, it produces a substratecontaining a homogenous contaminant distribution. This controlled laboratoryexperiment demonstrated the effects of seasonal priming and reproductive activityon DNA damage in T. semidecussatus and highlighted potential limitations for thefuture in situ use of clams as indicators of sediments contaminated with potentiallygenotoxic agents.
5. Conclusion
The present study highlights the importance of test organism exposure history, notonly in terms of pollutant exposure but also thermal history, condition and repro-ductive state. We conclude that (1) the degree of DNA damage recorded by meansof the Comet assay in the Manila clam T. semidecussatus was independent of size(age); (2) although spawning did not take place, the DNA damage observed ingill and digestive gland cells—but not haemocytes—was affected by reproductiveactivity; (3) a 2-week acclimation period to laboratory conditions may be too shortto eliminate the effects of seasonal priming; (4) the lack of correlation between theDNA damage to haemocytes and that of gill or digestive gland cells demonstratesthat haemocytes should not be used to predict the level of DNA damage in cells ofother organs in this species.
Acknowledgements
This work was funded by The Higher Education Authority under the Programmeof Research in Third Level Institutions (Cycle 2) as part of the EnvironmentalResearch Institute, University College Cork. We are grateful to Prof. Maire F.Mulcahy for critically reading an earlier draft of this manuscript.
References
Absolom, D. R. (1986). Basic methods for the study of phagocytosis.Methods in Enzymology, 132, 95–182.
Baumann, P. C. (1998). Epizootics of cancer in fish associated with genotoxins in sediment and water.
Mutation Research—Reviews in Mutation Research, 411, 227–233.
Belpaeme, K., Cooreman, K., & Kirsch-Volders, M. (1998). Development and validation of the in vivo
alkaline comet assay for detecting genomic damage in marine flatfish. Mutation Research—Genetic
Toxicology and Environmental Mutagenesis, 415, 167–184.
Berg, K. (1953). The problem of respiratory acclimatization. Hydrobiologia, 5, 331–350.
Birmelin, C., Mitchelmore, C. L., Goldfarb, P. S., & Livingstone, D. R. (1998). Characterisation of
biotransformation enzyme activities and DNA integrity in isolated cells of the digestive gland of the
common mussel, Mytilus edulis L. Comparative Biochemistry and Physiology a—Molecular and
Integrative Physiology, 120, 51–56.
Birmelin, C., Pipe, R. K., Goldfarb, P. S., & Livingstone, D. R. (1999). Primary cell-culture of the
digestive gland of the marine mussel Mytilus edulis: a time-course study of antioxidant- and bio-
transformation-enzyme activity and ultrastructural changes. Marine Biology, 135, 65–75.
308 M.G.J. Hartl et al. /Marine Environmental Research 57 (2004) 295–310

Blaise, C., Gagne, F., Pellerin, J., Hansen, P. D., & Trottier, S. (2002). Molluscan shellfish biomarker
study of the Quebec, Canada, Saguenay Fjord with the soft-shell clam, Mya arenaria. Environmental
Toxicology, 17, 170–186.
Boisson, F., Hartl, M. G. J., Fowler, S. W., & Amiard-Triquet, C. (1998). Influence of chronic exposure
to silver and mercury in the field on the bioaccumulation potential of the bivalve Macoma balthica.
Marine Environmental Research, 45, 325–340.
Briton, W. (1991). Clam cultivation manual. Dublin: Bord Iascaigh Mhara & Taighde Mara Teo (pp. 1–60).
Byrne, P. A., & O’Halloran, J. (1999). Aspects of assaying sediment toxicity in Irish estuarine ecosystems.
Marine Pollution Bulletin, 39, 97–105.
Byrne, P. A., & O’Halloran, J. (2000). Acute and sublethal toxicity of estuarine sediments to the manila
clam, Tapes semidecussatus. Environmental Toxicology, 15, 456–468.
Carpene, E., & George, S. G. (1981). Absorption of cadmium by gills of Mytilus edulis (L.). Molecular
Physiology, 1, 23–34.
Chong, K., & Wang, W. X. (2000). Bioavailability of sediment-bound Cd, Cr and Zn to the green mussel
Perna viridis and the Manila clam Ruditapes philippinarum. Journal of Experimental Marine Biology and
Ecology, 255, 75–92.
Chong, K., & Wang, W. X. (2001). Comparative studies on the biokinetics of Cd, Cr, and Zn in the green
mussel Perna viridis and theManila clamRuditapes philippinarum. Environmental Pollution, 115, 107–121.
Collins, A. R., Dobson, V. L., Dusinska, M., Kennedy, G., & Stetina, R. (1997). The comet assay: what
can it really tell us? Mutation Research—Fundamental and Molecular Mechanisms of Mutagenesis, 375,
183–193.
Coughlan, B.M., Hartl, M. G. J., O’Reilly, S. J., Sheehan, D., Morthersill, C., van Pelt, F., O’Halloran, J., &
O’Brien, N.M. (2002). Detecting genotoxicity using the Comet assay following chronic exposure ofManila
clam Tapes semidecussatus to polluted estuarine sediments. Marine Pollution Bulletin, 44, 1359–1365.
Del Valls, T. A., Forja, J. M., & Gomez-Parra, A. (2002). Seasonality of contamination, toxicity, and
quality values in sediments from littoral ecosystems in the Gulf of Cadiz (SW Spain). Chemosphere, 46,
1033–1043.
Devaux, A., Pesonen, M., & Monod, G. (1997). Alkaline comet assay in rainbow trout hepatocytes.
Toxicology in Vitro, 11, 71–79.
Domouhtsidou, G. P., & Dimitriadis, V. K. (2000). Ultrastructural localization of heavy metals (Hg, Ag,
Pb, and Cu) in gills and digestive gland of mussels, Mytilus galloprovincialis (L.). Archives of Environ-
mental Contamination and Toxicology, 38, 472–478.
Doyotte, A., Cossu, C., Jacquin, M. C., Babut, M., & Vasseur, P. (1997). Antioxidant enzymes,
glutathione and lipid peroxidation as relevant biomarkers of experimental or field exposure in the gills
and the digestive gland of the freshwater bivalve Unio tumidus. Aquatic Toxicology, 39, 93–110.
Edwards, J. W., Edyvane, K. S., Boxall, V. A., Hamann, M., & Soole, K. L. (2001). Metal levels in seston
and marine fish flesh near industrial and metropolitan centres in South Australia. Marine Pollution
Bulletin, 42, 389–396.
French, B. L., Reichert, W. L., Hom, T., Nishimoto, M., Sanborn, H. R., & Stein, J. E. (1996).
Accumulation and dose-response of hepatic DNA adducts in English sole. AquaticToxicology, 36, 1–16.
Frenzilli, G., Nigro, M., Scarcelli, V., Gorbi, S., & Regoli, F. (2001). DNA integrity and total oxyradical
scavenging capacity in the Mediterranean mussel, Mytilus galloprovincialis: a field study in a highly
eutrophicated coastal lagoon. Aquatic Toxicology, 53, 19–32.
Giovannelli, L., Decorosi, F., Dolara, P., & Pulvirenti, L. (2003). Vulnerability to DNA damage in the
aging rat substantia nigra: a study with the comet assay. Brain Research, 969, 244–247.
Hamoutene, D., Payne, J. F., Rahimtula, A. D., & Lee, K. (2002). Use of the Comet assay to assess DNA
damage in hemocytes and digestive gland cells of mussels and clams exposed to water contaminated
with petroleum hydrocarbons. Marine Environmental Research, 54, 471–474.
Kammann, U., Riggers, J. C., Theobald, N., & Steinhart, H. (2000). Genotoxic potential of marine
sediments from the North Sea. Mutation Research—Genetic Toxicology and Environmental Mutagenesis,
467, 161–168.
Luoma, S. N., & Ho, K. T. (1998). Appropriate uses of marine and estuarine sediment bioassays. In
P. Calow (Ed.), Handbook of ecotoxicology (pp. 193–226). Berlin: Blackwell Science.
M.G.J. Hartl et al. /Marine Environmental Research 57 (2004) 295–310 309

Mitchelmore, C. L., Birmelin, C., Chipman, J. K., & Livingstone, D. R. (1998). Evidence for cytochrome
P-450 catalysis and free radical involvement in the production of DNA strand breaks by benzo[a]pyrene
and nitroaromatics in mussel (Mytilus edulis L.) digestive gland cells. Aquatic Toxicology, 41, 193–212.
Mitchelmore, C. L., & Chipman, J. K. (1998). Detection of DNA strand breaks in brown trout (Salmo
trutta) hepatocytes and blood cells using the single cell gel electrophoresis (comet) assay. Aquatic
Toxicology, 41, 161–182.
Nacci, D., Nelson, S., Nelson, W., & Jackim, E. (1992). Application of the DNA alkaline unwinding assay
to detect DNA strand breaks in marine bivalves. Marine Environmental Research, 33, 83–100.
Nacci, D. E., Stephanie, C., & Jackim, E. (1996). Detection of DNA damage in individual cells from
marine organisms using the single cell gel assay. Aquatic Toxicology, 35, 197–210.
Pavlica, M., Klobucar, G. I. V., Mojas, N., Erben, R., & Papes, D. (2001). Detection of DNA damage
in haemocytes of zebra mussel using the comet assay. Mutation Research—Genetic Toxicology and
Environmental Mutagenesis, 490, 209–214.
Pavlica, M., Klobucar, G. I. V., Vetma, N., Erben, R., & Papes, D. (2000). Detection of micronuclei in
haemocytes of zebra mussel and great ramshorn snail exposed to pentachlorphenol. Mutation Research,
465, 145–150.
Pruski, A. M., & Dixon, D. R. (2002). Effects of cadmium on nuclear integrity and DNA repair efficiency
in the gill cells of Mytilus edulis L. Aquatic Toxicology, 57, 127–137.
Pruski, A. M., & Dixon, D. R. (2003). Toxic vents and DNA damage: first evidence from a naturally
contaminated deep-sea environment. Aquatic Toxicology, 64, 1–13.
Rank, J. (1999). Use of the comet assay on the blue mussel, Mytilus edulis, from coastal waters in
Denmark. Neoplasma, 46, 9–10.
Rice, D. W., Seltenrich, C. P., Spies, R. B., & Keller, M. L. (1993). Seasonal and annual distribution of
organic contaminants in marine sediments from Elkhorn Slough, Moss Landing Harbor and nearshore
Monterey Bay, California. Environmental Pollution, 82, 79–91.
Sastry, A. N. (1966). Temperature effects in reproduction of the bay scallop, Aequipecten irradians
Lamarck. Biological Bulletin 130. Woods Hole, MA: Marine Biological Laboratory (pp. 118–134).
Sastry, A. N., & Blake, N. J. (1971). Regulation of gonad development in the bay scallop, Aequipecten
irradians Lamarck. Biological Bulletin 140. Woods Hole, MA: Marine Biological Laboratory (pp. 274–283).
Speit, G., & Hartmann, A. (1995). The contribution of excision repair to the DNA effects seen in the
alkaline single-cell gel test (Comet assay). Mutagenesis, 10, 555–559.
Stein, J. E., Hom, T., Collier, T. K., Brown, D. W., & Varanasi, U. (1995). Contaminant exposure and
biochemical effects in outmigrant juvenile Chinook Salmon from urban and nonurban estuaries of
Puget Sound, Washington. Environmental Toxicology and Chemistry, 14, 1019–1029.
Steinert, S. A. (1999). DNA damage as a bivalve biomarker. Biomarkers, 4, 492–496.
Steinert, S. A., Streib-Montee, R., Leather, J.M., & Chadwick, D. B. (1998). DNAdamage inmussels at sites
in SanDiego Bay.Mutation Research—Fundamental andMolecularMechanisms ofMutagenesis, 399, 65–85.
Werner, I., Kline, K. F., &Hollibaugh, J. T. (1998). Stress protein expression inAmpelisca abdita (Amphipoda)
exposed to sediments from San Francisco Bay. Marine Environmental Research, 45, 417–430.
Wilson, J. G., & Elkaim, B. (1997). Seasonal and geographical differences in oxygen consumption with
temperature of Cerastoderma glaucum (Poiret) and a comparison with C. edule (L.). Estuarine Coastal
and Shelf Science, 45, 571–577.
Wilson, J. T., Pascoe, P. L., Parry, J. M., & Dixon, D. R. (1998). Evaluation of the comet assay as a method
for the detection of DNA damage in the cells of a marine invertebrate, Mytilus edulis L. (Mollusca:
Pelecypoda).Mutation Research—Fundamental and Molecular Mechanisms of Mutagenesis, 399, 87–95.
Wong, P. K. (1988). Mutagenicity of heavy metals. Bulletin of Environmental Contamination and
Toxicology, 40, 597–603.
Woods, J. A., O’Leary, K. A., McCarthy, R. P., & O’Brien, N. M. (1999). Preservation of comet assay
slides: comparison with fresh slides. Mutation Research-Fundamental and Molecular Mechanisms of
Mutagenesis, 429, 181–187.
Xie, Q. S., & Burnell, G. M. (1994). A comparative study of the gametogenic cycles of the clams Tapes
philippinarum (Adams,a. and Reeve 1850) and Tapes decussatus (Linnaeus) on the south coast of
Ireland. Journal of Shellfish Research, 13, 467–472.
310 M.G.J. Hartl et al. /Marine Environmental Research 57 (2004) 295–310
Related Documents











