Behavioral Neuroscience 2000, Vol. 114, No. 3, 514-525 Copyright 2000 by the American Psychological Association Inc 0735-7044/00/S5.00 DOI: 10.1037/AJ735-7044.114.3.514 Impairment of Olfactory Discrimination by Blockade of GABA and Nitric Oxide Activity in the Honey Bee Antennal Lobes Jay S. Hosier, Kristi L. Buxton, and Brian H. Smith The Ohio State University Honey bees readily associate an odor with sucrose reinforcement, and the response generalizes to other odors as a function of structural similarity to the conditioned odor. Recent studies have shown that a portion of odor memory is consolidated in the antennal lobes (AL), where first-order synaptic processing of sensory information takes place. The AL and/or the sensory afferents that project into them show staining patterns for the enzyme nitric oxide synthase, which catalyzes the release of the gaseous transmitter nitric oxide (NO). The results show that pharmacological blockade of NO release impairs olfactory discrimination only when release is blocked before conditioning. Blockade of GABAergic transmission disrupts discrimination of similar but not dissimilar odorants, and does so when the block occurs before condition or before testing. These results show that GABA and NO regulate the specificity of associative olfactory memory in the AL. Several neuromodulators and neurotransmitters play important roles in information processing in both the vertebrate olfactory bulb (OB) and in the insect antennal lobe (AL; Hildebrand & Shepherd, 1997; Shipley & Ennis, 1996). Of particular interest is the gaseous neurotransmitter nitric oxide (NO), which is present in vertebrate and invertebrate olfactory systems and has a variety of signaling functions within neuronal (Snyder, Jaffrey, & Zakhary, 1998) and nonneuronal systems (Ignarro, Buga, Wood, Byrnes, & Chaudhuri, 1987; Palmer, Ferrige, & Moncada, 1987). It most likely serves as a transmitter or as a modulator in a variety of neural systems (Schuman & Madison, 1994), where it may signal in an anterograde or a retrograde manner (Park, Straub, & O'Shea, 1998). Maintenance of long-term potentiation and long-term de- pression in the vertebrate brain requires the release of NO from postsynaptic cells (Lev-Ram, Makings, Keitz, Kao, & Tsien, 1995; Schuman & Madison, 1991, 1994). The sensory and first-order synaptic processing areas (AL and OB) show prominent staining patterns for NO itself or for NO synthase, an enzyme that catalyzes the release of NO by conver- sion of arginine to citrulline (Brennan & Keverne, 1997; Kendrick, Guevara-Guzman, Zorrilla, Hinton, Broad, Mimmack, & Ohkura, 1997; Miiller, 1997; Nighom, Gibson, Rivers, Hildebrand, & Mor- ton, 1998). Release of NO in these areas is activity dependent and triggers several biochemical pathways, both in the cells in which it is produced and in the target cells to which it diffuses (Miiller, 1997). The distribution and biochemical properties of NO could Jay S. Hosier, Kristi L. Buxton, and Brian H. Smith, Department of Entomology, The Ohio State University. This work was supported by Grant 9 R01-RR14166-06 from the Na- tional Institutes of Health/National Center for Research Resources. Correspondence concerning this article should be addressed to Brian H. Smith, Department of Entomology, 103 Botany and Zoology Building, 1735 Neil Avenue, The Ohio State University, Columbus, Ohio 43210- 1220. Electronic mail may be sent to [email protected]. predispose it to mediate processes of adaptation, in sensory cells (Breer & Shepherd, 1993), as well as habituation (Muller & Hildebrandt, 1995), and associative memory (Muller, 1996). The broad phylogenetic distribution of NO in the OB and AL suggests that it subserves fundamental properties of olfactory signal pro- cessing. Yet, particularly for insects, those properties still remain to be revealed in behavioral investigations. In addition, GABAergic transmission is involved in generating the spatial and/or temporal patterns of activity in the AL that are critical for representing olfactory information in the central ner- vous system (CNS). In the locust (Laurent, 1996; Laurent & Davidowitz, 1994; Laurent & Naraghi, 1994; Wehr & Laurent, 1996) and the honey bee (Stopfer, Bhagavan, Smith, & Laurent, 1997), odor stimulation induces an evolving set of spatial arrays in the AL. Stimulation gives rise to regular 20-30 Hz oscillations in the local field potential measured in the projection fields of the AL projection neurons. Each cycle in the local field potential oscilla- tion is caused by an array of synchronously activated projection neurons, and each succeeding cycle is caused by a partially or completely different set of activated neurons. Each array thus consists of a set of neurons that show phase-locked oscillations in their membrane potentials. Application of picrotoxin (PCT) to the AL, which blocks GABA-gated Cl— currents mediated by a sub- class of AL interaeurons (Christensen, Waldrop, & Hildebrand, 1998; Hildebrand, 1995; Homberg, Christensen, & Hildebrand, 1989; Homberg, Kingan, & Hildebrand, 1987; Leitch & Laurent, 1996), eliminates this synchronization but not the characteristic projection neuron response patterns to odor (MacLeod & Laurent, 1996). Treatment with PCT also impairs olfactory discrimination in the honey bee (Stopfer et al., 1997). Honey bees can be easily condi- tioned to respond to an odor conditioned stimulus (CS, Odor C), and their response to that odor remains for several days after multiple-trial conditioning (Menzel, 1990; Menzel & Muller, 1996). Subjects generalize the response to an odorant that is structurally similar (Odor S) to Odor C, but the generalization response is typically lower than the response to the conditioned 514

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Behavioral Neuroscience2000, Vol. 114, No. 3, 514-525
Copyright 2000 by the American Psychological Association Inc0735-7044/00/S5.00 DOI: 10.1037/AJ735-7044.114.3.514
Impairment of Olfactory Discrimination by Blockade of GABA and
Nitric Oxide Activity in the Honey Bee Antennal Lobes
Jay S. Hosier, Kristi L. Buxton, and Brian H. SmithThe Ohio State University
Honey bees readily associate an odor with sucrose reinforcement, and the response generalizes to other
odors as a function of structural similarity to the conditioned odor. Recent studies have shown that a
portion of odor memory is consolidated in the antennal lobes (AL), where first-order synaptic processing
of sensory information takes place. The AL and/or the sensory afferents that project into them showstaining patterns for the enzyme nitric oxide synthase, which catalyzes the release of the gaseous
transmitter nitric oxide (NO). The results show that pharmacological blockade of NO release impairsolfactory discrimination only when release is blocked before conditioning. Blockade of GABAergictransmission disrupts discrimination of similar but not dissimilar odorants, and does so when the block
occurs before condition or before testing. These results show that GABA and NO regulate the specificityof associative olfactory memory in the AL.
Several neuromodulators and neurotransmitters play important
roles in information processing in both the vertebrate olfactory
bulb (OB) and in the insect antennal lobe (AL; Hildebrand &
Shepherd, 1997; Shipley & Ennis, 1996). Of particular interest is
the gaseous neurotransmitter nitric oxide (NO), which is present in
vertebrate and invertebrate olfactory systems and has a variety of
signaling functions within neuronal (Snyder, Jaffrey, & Zakhary,
1998) and nonneuronal systems (Ignarro, Buga, Wood, Byrnes, &
Chaudhuri, 1987; Palmer, Ferrige, & Moncada, 1987). It most
likely serves as a transmitter or as a modulator in a variety of
neural systems (Schuman & Madison, 1994), where it may signal
in an anterograde or a retrograde manner (Park, Straub, & O'Shea,
1998). Maintenance of long-term potentiation and long-term de-
pression in the vertebrate brain requires the release of NO from
postsynaptic cells (Lev-Ram, Makings, Keitz, Kao, & Tsien, 1995;
Schuman & Madison, 1991, 1994).
The sensory and first-order synaptic processing areas (AL and
OB) show prominent staining patterns for NO itself or for NO
synthase, an enzyme that catalyzes the release of NO by conver-
sion of arginine to citrulline (Brennan & Keverne, 1997; Kendrick,
Guevara-Guzman, Zorrilla, Hinton, Broad, Mimmack, & Ohkura,
1997; Miiller, 1997; Nighom, Gibson, Rivers, Hildebrand, & Mor-
ton, 1998). Release of NO in these areas is activity dependent and
triggers several biochemical pathways, both in the cells in which it
is produced and in the target cells to which it diffuses (Miiller,
1997). The distribution and biochemical properties of NO could
Jay S. Hosier, Kristi L. Buxton, and Brian H. Smith, Department of
Entomology, The Ohio State University.This work was supported by Grant 9 R01-RR14166-06 from the Na-
tional Institutes of Health/National Center for Research Resources.Correspondence concerning this article should be addressed to Brian H.
Smith, Department of Entomology, 103 Botany and Zoology Building,1735 Neil Avenue, The Ohio State University, Columbus, Ohio 43210-1220. Electronic mail may be sent to [email protected].
predispose it to mediate processes of adaptation, in sensory cells
(Breer & Shepherd, 1993), as well as habituation (Muller &
Hildebrandt, 1995), and associative memory (Muller, 1996). The
broad phylogenetic distribution of NO in the OB and AL suggests
that it subserves fundamental properties of olfactory signal pro-
cessing. Yet, particularly for insects, those properties still remain
to be revealed in behavioral investigations.
In addition, GABAergic transmission is involved in generating
the spatial and/or temporal patterns of activity in the AL that are
critical for representing olfactory information in the central ner-
vous system (CNS). In the locust (Laurent, 1996; Laurent &
Davidowitz, 1994; Laurent & Naraghi, 1994; Wehr & Laurent,
1996) and the honey bee (Stopfer, Bhagavan, Smith, & Laurent,
1997), odor stimulation induces an evolving set of spatial arrays in
the AL. Stimulation gives rise to regular 20-30 Hz oscillations in
the local field potential measured in the projection fields of the AL
projection neurons. Each cycle in the local field potential oscilla-
tion is caused by an array of synchronously activated projection
neurons, and each succeeding cycle is caused by a partially or
completely different set of activated neurons. Each array thus
consists of a set of neurons that show phase-locked oscillations in
their membrane potentials. Application of picrotoxin (PCT) to the
AL, which blocks GABA-gated Cl— currents mediated by a sub-
class of AL interaeurons (Christensen, Waldrop, & Hildebrand,
1998; Hildebrand, 1995; Homberg, Christensen, & Hildebrand,
1989; Homberg, Kingan, & Hildebrand, 1987; Leitch & Laurent,
1996), eliminates this synchronization but not the characteristic
projection neuron response patterns to odor (MacLeod & Laurent,
1996).
Treatment with PCT also impairs olfactory discrimination in the
honey bee (Stopfer et al., 1997). Honey bees can be easily condi-
tioned to respond to an odor conditioned stimulus (CS, Odor C),
and their response to that odor remains for several days after
multiple-trial conditioning (Menzel, 1990; Menzel & Muller,
1996). Subjects generalize the response to an odorant that is
structurally similar (Odor S) to Odor C, but the generalization
response is typically lower than the response to the conditioned
514

NITRIC OXIDE MODULATION OF OLFACTORY PROCESSING 515
odorant (Smith & Menzel, 1989). Thus, subjects can normally
discriminate Odor C from Odor S, and they can also easily dis-
criminate Odor C from an odor that is from a different, dissimilar
molecular class (Odor D). PCT treatment eliminates subjects'
ability to discriminate Odor C from Odor S but leaves intact their
ability to discriminate Odor C from Odor D (Stopfer et al., 1997).
Thus, the temporal information that is provided by PN synchroni-
zation within the AL appears to be necessary for fine odor dis-
criminations. Discrimination of a dissimilar odor, which probably
activates different spatial arrays due to the activation of nonover-
lapping arrays of sensory cells (Joerges, Kilttner, Galizia, & Men-
zel, 1997), may not depend as much on that temporal information.
NO is also involved in the maintenance of oscillations in the
mollusk olfactory system. In the mollusk Limax, the circuitry of
the procerebral lobes gives rise to slow (0.5-2 Hz) propagating
waves of activity that travel along the lobe's longitudinal axis
(Gelperin & Tank, 1990). Natural odor stimulation causes electri-
cal activity to transiently switch from a propagating to a standing
waveform (Delaney et al., 1994). Drugs that block the activity of
nitric oxide synthase (NOS) eliminate the oscillations (Gelperin,
1994), just as PCT does in honey bees (Stopfer et al., 1997) and
locusts (MacLeod & Laurent, 1996). Although the spatial and
temporal characteristics of oscillations in the mollusk may be
different from those in insects, the data nevertheless provide a
testable prediction that GABA and NO mediate different parts of
the same pathway in the olfactory system.
The honey bee AL, like those of other insects, shows strong
staining patterns for NOS (Miiller, 1994; Miiller & Bicker, 1994),
although the cellular localization of NOS has yet to be revealed in
detail. To date, all identified sequences for NOS isoforms in
insects show strong similarity to mammalian neuronal NOS (Nig-
horn et al., 1998; Regulski & Tully, 1995). Because of the poten-
tial for involvement of NOS in synchronization among AL neu-
rons, we predicted that blockade of NO release in the honey bee
would disrupt olfactory discrimination ability. Indeed, the results
support this hypothesis. However, they also reveal that the disrup-
tion is either quantitatively or qualitatively different from that
produced by PCT treatment (Stopfer et al., 1997). In particular,
NO release is apparently involved in the formation of components
of associative memory that are localized to the AL (Griinbaum &
Miiller, 1998).
Method
Subjects
We conditioned a total of 724 worker honey bees for this study. Subjects
were from queens that were bred from a controlled breeding population
maintained at The Ohio State University.
Proboscis Extension Response (PER) Conditioning
We used a PER conditioning procedure to assay subjects' responses to
odors (Menzel & Bitterman, 1983). We collected worker honey bees as
they departed from colonies located either out-of-doors or in a flight room
maintained at room temperature and on a 16:8-hr light-dark cycle. Sub-
jects were collected individually in glass vials the morning of the day they
were to be used in a conditioning procedure. The vials were then placed
immediately into an ice-water bath until the bees ceased moving. They
were then placed into restraining harnesses that allowed movement of
antennae and mouthparts.
To facilitate pharmacological treatment of the AL, we immobilized each
subject's head by allowing molten dental wax to flow into and harden in
the space between the head and the strip of tape that restrained subjects in
the harnesses. We then cut a small window in the head capsule just above
the antennae to expose the ALs. After this surgery, each subject was fed
a 0.4-u.l droplet of 1.25 M sucrose-water solution and allowed to remain
undisturbed on a holding tray for 1 hr.
The specific protocol we used to condition individual subjects has been
extensively described elsewhere (Smith, 1997; Stopfer et al., 1997; http://
iris.biosci.ohio-state.edu/honeybee). Briefly, a subject is moved from the
holding tray into a conditioning arena through which air is continuously
drawn into an exhaust system. Approximately 30 s after placement, odor-
laden air is injected for 4 s into the exhaust stream that is drawn over the
subject's antennae. Three seconds after odor onset, the subject is stimulated
by antennal contact with a 0.4-/iil droplet of 1.25 M sucrose solution, which
the subject then consumes once its mouth parts are extended. All subjects
receive six such acquisition trials with Odor C separated by a 2-min
intertrial interval. For these acquisition trials, we scored a subject as having
responded if it extended its proboscis after the onset of odor but prior to
presentation of the sucrose unconditioned stimulus (US). If it failed to
extend its proboscis during this time, it was scored as having failed to
respond to the Odor CS.
Critical tests of hypotheses were based on each subject's proboscis
extension performance during test trials with Odors C, S, and D. Test trials
were performed identically to the reinforced (acquisition) trials just de-
scribed, except that sucrose stimulation and feeding was omitted. Subjects
therefore had the entire 4-s period of odor stimulation to respond to the test
odorant. These three trials, spaced 2 min apart and performed beginning 90
or 100 min after the start of conditioning (Figure 1, A and B), were
performed in a randomized sequence across subjects within treatment
groups.
During these test trials, we scored a positive response when a subject
extended its proboscis after the onset and before the offset of odor. In
addition, for some of the experiments, we videotaped the test trials to
analyze response duration to the 4-s odor burst (Smith, 1997, 1998).
Response duration refers to the total elapsed time that the proboscis
remains extended beyond the line that connects the tips of the open
mandibles. The taping began with the onset of odor and continued for a
total of 20 s, which allowed for 16 s to elapse between odor offset and the
end of the videotape recording.
Tests for Nonassociative Effects of N-Nitro-L-Arginine
Methyl Ester (LNAME) and PCT
It is possible that PCT and NO sensitize an animal's response to odor
and/or to the mechanosensory stimulation that accompanies it. We tested
this possibility with a sensitization assay, in which subjects were fed on a
sucrose-water droplet just before being tested with odor (Benatar, Cobey,
& Smith, 1995). In this procedure, subjects are not conditioned to odor
prior to testing (Figure ID). Within 15 s of feeding, subjects' responses to
odor are elevated by the sensitizing properties of the sucrose US. Sixty
seconds after sucrose stimulation, the sensitization responses decline to
near baseline response levels. If PCT and LNAME increase sensitivity in
a nonspecific manner, we would expect a drug-treated group to show
higher responsiveness to odor stimulation.
Pharmacological Treatment
Muller (1996) showed that LNAME rapidly (within 20 min) blocks
honey bee neuronal NOS. Therefore, we used the same pharmacological
treatment to block NOS in the AL. We used two general protocols for
pharmacological treatment: First (Figure 1 A), we applied a 3-nl droplet of

516 HOSLER, BUXTON, AND SMITH
B
inject withsaline/drug
• 10
inject withsaline/drug
Jl 10
v*
min
nun
conditioningwith 'C'
»l^JLUllllconditioning
with 'C'
-kl aUlillLl
90 min
90 min
Test with:C/S/D
*nnnillili]inject withsaline/drug
n 10Test with:
C/S/D
min^ nnnI I U U
15 or 60 sec
Test with:geraniol or 1-hexanol
Figure 1. Summary of experimental designs. Subjects received either a single (A) or dual (B) application ofsaline or saline + drui treatment to the brain regions described in the text. Ten minutes elapsed between theinitial treatment and six conditioning trials with the conditioned odor (C), which were separated by a 2-minintertrial interval. Ninety rain later subjects were either (A) tested with C, similar (S), and dissimilar (D) odors,in randomized order, or (B) given a second application of saline or saline + drug 10 min before testing withOdors C, S. and D. (C) Schematic diagram of the honey bee head with a cutaway view of the brain. CE =compound eye; Oc = ocelli; An = antenna; Md = mandibles; OL = optic lobes of the brain; AL = antennallobes; MB = mushroom body. Diagram of experimental protocol (D) used for tests of effects of nitric oxidesynthase inhibitors on nonassociative components of proboscis extension response conditioning.
either saline (amounts/L: 0.2 g KC1, 0.2 g CaCl, 9 g NaCl, 4 gsuciose, 1.19 g 4-(2-hydroxyethyl)-l-piperazine thanesculfonic acid[HEPESD or the indicated saline-drug treatment (PCT 100 fjM; LNAMEor N-nitro-D-arginine methyl ester (DNAME) at either 20 jtM or 200 ju-M;S-nitrosoacetylpenicillamine [SNAP] at 200 ^M) to each AL 10 min priorto conditioning. The droplet was applied to the dorsal anterior surface ofeach AL, Second (Figure IB), the same type of droplet was applied to eachAL 10 min before conditioning and 10 rain before testing. This latterprotocol generated four treatment groups, which corresponded to all pos-sible combinations of saline and drug treatment at the two treatment points.In both protocols, 90 min elapsed between conditioning and testing.
Odor Generalization Assay
We used a generalization assay to examine the effect of blockade ofGAB A and NO modulation on odor responses. We tested each subject withOdor C, with the structurally similar Odor S, and with the dissimilar OdorD, presented in a randomized order 90 min after conditioning was com-pleted. Generalization occurs when the response to a CS can be elicited byother stimuli that differ from the CS along defined stimulus dimensions(Mackintosh, 1983), Typically, a gradient of response levels can be estab-lished, and the response decreases as the test stimulus becomes less similarto the CS. In our work, hexanol and octanol are counterbalanced as the CS(Odor C) and similar (Odor S) odorants, respectively. There is empirical
support from the rabbit OB that shows that these odorants define a"dimension" in neural coding in that neuropil (Mori, 1995). Geraniol, aterpene, is always our dissimilar odor (Odor D). The response levelsdecrease from Odor C to Odor S to Odor D in a reliable fashion acrossexperiments (Smith & Menzel, 1989). We expect untreated subjects torespond less to Odor S and Odor D than they do to Odor C, in a mannerthat is graded by the molecular similarity between the odorants (Smith &.Menzel, 1989; Stopfer et al., 1997).
We also make the assumption that increased generalization due to drugtreatment reflects reduced discriminability of odors. However, this shouldnot be taken to imply that there is a complete lack of discrimination.
Generalization from Odor C to Odor S or to Odor D varies with thephysiological status of worker honey bees. Several experiments reportedbelow and in Stopfer et al. (1997, Figure 4) were carried out with workerscaught as they exited from colonies maintained in the field during thesummer months. In such workers, the percentage of subjects in a giventreatment group that respond to Odors C, S, or D is sufficient to demon-strate significant differences in the generalization gradient. When thosedifferences are evident in terms of the percentage of subjects that showproboscis extension, they are always evident according to the durationmeasure from videotape analyses (Smith, 1998). When the percentagemeasure fails to reveal significant differences, then the duration measurewill often reveal differences because it is more sensitive (Smith, 1997,1998).

NITRIC OXIDE MODULATION OF OLFACTORY PROCESSING 517
Statistical Analyses
In cases in which the proportion of responders and nonresponders was
analyzed, a chi-squared analysis was used. Before statistical analysis,
duration measures were log transformed; 1.0 was added to each subject's
duration score to avoid transformation of 0.0, which was the score attrib-
uted to subjects that failed to respond to the test odor. The log-transformed
duration scores to Odors C, S, and D were analyzed with two-way analysis
of variance (ANOVA) without replication (see Sokal & Rohlf, 1995).
Subjects were included as a random treatment factor, whereas the response
to Odors C, S, and D was a fixed treatment effect. Because subjects were
not tested multiple times with each of the three odorants, an interaction
term could not be calculated. However, plots of subjects' response to the
three odorants revealed that the lines were parallel, which provides an
indication that little interaction was present. Most of the experiments
reported below yielded a significant F ratio for differences across subjects.
The fact that individuals differ in overall response level reflects other
factors that vary among them (e.g., age genotype, motivational state), but
the random sampling of subjects from large colonies ensures that these
factors are randomly distributed across treatments. We thus do not report
these F ratios below, because they were not critical for interpretation of
differences in the fixed treatment effect.
When the ANOVA revealed differences in response levels to Odors C,
S, and D, we performed a series of one-tailed post hoc comparisons of C
to S and C to D, to determine whether either of the latter two odorants
elicited a response level lower than that to Odor C. These comparisons
were performed with paired-comparison t tests (Sokal & Rohlf, 1995).
In a few cases, we performed cross-group comparisons with standard
one-way ANOVA of responses to Odors C, S, or D. In those cases, post
hoc comparisons were performed with standard least significant difference
tests (Sokal & Rohlf, 1995).
Results
LNAME Decreases Discriminability of Odors
We first examined whether blockade of NO has an effect on
odor discrimination by using the same conditioning protocol that
was used to demonstrate the effect of GABA blockade (Stopfer et
al., 1997, Figure 1). Figure 2 shows the acquisition curves for all
treatment groups in each set of experiments reported below. All
treatment groups showed the same relative levels of acquisition
performance (80-100% of subjects respond by the sixth acquisi-
tion trial). The curves within each graph never differed signifi-
cantly from one another, Panel A: F(6, 283) = 1.6, ns; Panel B:
F(4, 235) = 0.3, ns; Panel C: F(3, 185) = 1.8, ns; Panel D: F(6,
313) = 1.3, ns. This provides an indication that drug treatment did
not affect acquisition performance to Odor C or motor tasks
involved in appetitive feeding movements.
We performed two initial investigations of the effect of LNAME
and the optically less active isomer DNAME on olfactory discrim-
ination. In these studies, we recorded the percentage of subjects
that responded to odor presentation during the test trials. In both
studies 89 to 100% of subjects responded to the conditioned Odor
C during the test (Tables 1 and 2), and the response levels of drug
treatment groups to Odor C did not differ from the saline control
groups. Furthermore, subjects within both saline treatment groups
showed significantly lower responses to Odor S and to Odor D
compared with their responses to Odor C, which replicated the
generalization gradient between these odors (Stopfer et al., 1997).
In the two groups treated with 200 pM LNAME, subjects showed
only a slight, nonsignificant response decrement from Odor C to
Odor S or to Odor D. A 10-fold lower concentration of LNAME
(Table 2) produced a significant generalization decrement that did
not qualitatively differ from the relevant control group. Finally,
200 jxM DNAME produced somewhat elevated response levels to
Odor S, but the response levels to Odor D were not different from
the control.
Reconstitution of C From D Discrimination
With an NO Donor
Thus, LNAME has the capacity to disrupt discrimination of
Odor C from Odor S, as does PCT treatment (Stopfer et al., 1997).
Therefore, we performed an additional investigation that was de-
signed to further delimit the nature of this effect. In this experi-
ment, we also videotaped subjects' responses to Odors C, S, and D,
to obtain more precise parametric measures of the response topol-
ogy (Smith, 1997, 1998).
Subjects that received saline application to each AL showed the
characteristic generalization gradient from Odor C to Odors S and
D when they were tested 90 min after conditioning (Figure 3A).
There was a significant effect of test odor, F(2, 125) = 10.9, p <
.001. Subjects responded significantly more to C than to S,
/(42) = 1.8, p < .05, and significantly more to C than to D,
7(42) = 5.6, p < .001.
The pattern of generalization in the saline treatment group
contrasted markedly with those of some of the drug treatment
groups. Subjects that received saline + LNAME application to
each AL responded equally often to all three odors, both in regard
to response probability and duration (Figure 3B). In this group the
effect of test condition was not significant; for C, S, and D: F(2,
104) = 0.7, ns. In contrast, treatment with DNAME (Figure 3C)
produced a significant effect of test odor, F(2, 107) = 5.6,p < .01.
Subjects in this group discriminated C from D, /(36) = 4.5, p <
.001, although the discrimination of C from S was impaired,r(36) = 1.1, ns.
We also ran several control procedures to more closely tie the
effect of LNAME to NO release in the AL. SNAP is an NO donor.
Therefore, treatment with SNAP would be expected to attenuate
the impact of LNAME (Kendrick et al., 1997). Subjects that
received coapplication of SNAP and LNAME responded differen-
tially to the test odors, F(2, 101) = 6.2, p < .01 (see Figure 3D).
They retained the ability to discriminate Odor C from Odor D,
J(34) = 0.9, ns, although the discrimination of C from S was still
impaired, 1(34) = 3.7, p < .001.
It is also possible that LNAME spreads rapidly to other brain
areas on application to the AL. If that were true, then LNAME
could exert its effect by suppressing NO release in other areas that
show NOS activity, such as the mushroom bodies (Muller, 1994).
We tested for this possibility in a group of subjects that received
application of LNAME to the optic lobes, which do not show
strong staining patterns for NOS (Muller, 1996). These subjects
responded differentially to the test odors, F(l, 113) = 12.9, p <
.001 (see Figure 3E). They displayed robust C from S, t(38) = 2.8,
p < .01, and C from D discrimination, r(38) = 5.5,p < .001. Thus
it is unlikely that the spread of LNAME to other neuropils in thebrain could account for the impairment in discrimination that
arises in conjunction with AL treatment.
We also performed across-groups comparisons of response lev-
els to the three test odorants. The five groups did not differ in

518 HOSLER, BUXTON, AND SMITH
100
80
salineLNAME (200)DNAME (200)
saline-*- LNAME (200)-e- LNAME (20)-*- DNAME (200)-e- DNAME (20)
UGO
§100o
80
« 6 0
20 •
0
saline-salinesaline-LNAME
LNAME-saline
LNAME-LNAME
1 3 4TRIAL
salineLNAME
DNAMELNAME+SNAPLNAME(OL)
saline-salinesaline-LNAMELNAME-salineLNAME-LNAME
saline-salinesaline-PCTPCT-salinePCT-PCT
3 4
TRIAL
Figure 2. Acquisition to conditioned odor (Odor C). A: Acquisition across eight independent groups of 26-40
subjects for test data presented in Tables 1 and 2. The first three groups and the last five groups were ran in
parallel to one another. Each of the two sets contained a saline control group. N-nitro-L-arginine methyl ester
(LNAME) and N-nitro-D-arginine methyl ester (DNAME) were applied at the juM concentrations indicated in
parentheses. B: Acquisition across five independent treatment groups of 35-43 subjects for test data presented
in Figure 3. Treatments of LNAME and DNAME were at 200 iM. LNAME + SNAP refers to the addition of
200 pM of the nitric oxide donor S-nitrosacetylpenicillamine to the LNAME treatment. LNAME(OL) refers to
application of LNAME to the optic lobes (Figure 1, Panel C). C-D: Acquisition for dual treatment experiments
reported in Table 3 and Figure 5, respectively. Sample sizes per group ranged from 37 to 40 subjects for each
experiment. LNAME was applied at 200-/iM concentration. PCT = picrotoxin.
response levels to Odor C, f(4, 185) = 1.3, ns (see Figure 3F), or
to Odor S, F(4, 185) = 0.5, ns (see Figure 3G). In regard to levelsof response to Odor D, only the LNAME group displayed a
response level that was significantly different from that of the
saline control group, F(4, 185) = 2.0, p < .05; LSD for saline-
LNAME comparison, p < .05 (see Figure 3H).
LNAME treatment does not increase the responsiveness to
background stimuli. We conditioned two groups of subjects, one
treated before conditioning with saline and the other treated withLNAME. These groups were tested for their responsiveness to air
alone, which is the mechanosensory background in which odor is
presented. The two groups did not differ in the mean response
during the test with the air stimulus (saline: M — 0.253;
SEM = 0.124 s; LNAME: M = 0.226; SEM = 0.130 s). Thus, the
increase in response to odor in LNAME-treated groups is specific
to the presence of odor.
SNAP Recovers C-From-S Discrimination After
DNAME Treatment
We performed a replicate of the design shown in Figure 1A to
investigate whether the partial effect of DNAME on impairment of
C-from-S discrimination might arise from a partial effect of
DNAME on NOS activity. We hypothesized that, if this were the

NITRIC OXIDE MODULATION OF OLFACTORY PROCESSING 519
Table 1
Effect of LNAME and DNAME on the Percentage of Subjects
That Responded to a Conditioned Odor (C), a Structurally
Similar Odor (S), and a Dissimilar Odor (D)
Treatment n C S D x*
SalineLNAME (200 jiM)DNAME (200 |iM)
/
292626
89.7
100.096.2
1.4ns
72.4100.096.2
9.9*
51.796.2
53.815.0**
16.7**2.2ns
20.6**
Note. Bees were treated 10 min before conditioning with either saline ora saline-containing drug treatment at the indicated concentration (seeMethod section, Figure 1). Ninety minutes later each bee was tested withOdors C, S, and D, presented in a randomized sequence. For each group,values indicate the percentage of n that responded to the odors. LNAME —
N-nitro-L-arginine methyl ester; DNAME = N-nitro-D-arginine methylester. Chi-square values to the right indicate comparison within treatments
across the three odors. Chi-square values below the columns indicatecomparison of responses to odor across treatment groups. Significance
levels are adjusted to maintain an experimentwise error rate of .05 (Sokal&Rohlf, 1995; *p< .01; ** p < .005).
case, then SNAP should recover C-from-S discrimination in that
group. In this replicate, the saline-treated group responded differ-
entially to the test odors, F(2,58) = 8.2, p < .001 (see Figure 4A).
Further analysis revealed low responses to Odor S, r(30) = 2.3,
p < .05, and Odor D, f(30) = 3.9, p < .001, when compared with
the response level to Odor C. The DNAME, F(2, 34) = 5.0, p <
.05 (Figure 4B), and the DNAME + SNAP, F(2, 34) = 8.2, p <
.01. treatment groups also responded differentially to the three test
odors. As expected, the DNAME group generalized from Odor C
to Odor S such that the response levels to those odors were
equivalent, /(18) — 0.9, ns. That group also responded differen-
tially to Odors C and D, ((18) = 2.8, p < .05. In the DNAME +
SNAP group, the pattern of C-from-S, ((18) = 2.8, p < .05, and
C-from-D, r(18) = 3.7, p < .01, discrimination was qualitatively
the same as in the saline group.
PCT and LNAME Have Different Effects in Storage and
Recall Tests
To investigate the physiological mechanisms in the AL that
underlie disruption of odor discrimination, it is critical to establish
whether the effects of LNAME and PCT are limited to treatment
of the AL before conditioning. To test this hypothesis, we modi-
fied our drug treatment protocol to include a second application 10
min prior to testing (Figure IB). This protocol gave rise to four
treatment groups that differed in regard to when they received drug
application (saline vs. drug before conditioning and/or before
recall testing).
In an initial experiment with LNAME application at 200 /jM
(Table 3) responses were not recorded on videotape. Subjects in
the control (saline-saline) group responded most often to Odor C
and significantly less often to Odors S and D. Response levels to
Odor S were relatively elevated in the saline-LNAME group, but
subjects in that group discriminated Odor C from Odor D. A
qualitatively different pattern of generalization was observed in the
LNAME-saline and LNAME-LNAME groups. In those groups,
the slight decrement in response frequency from Odor C to Odor
D was not significant. Comparison of response frequencies to Odor
D across groups (compare down the "D" column in Table 3)
showed that the elevated responses to that test odorant were
significantly higher whenever LNAME was active before
conditioning.
When we replicated this experiment using videotaped duration
scores, the same qualitative patterns emerged (see Figure 5).
Subjects' responses to Odors C, S, and D differed in most cases,
saline-saline: F(2, 110) = 11.7, p < .001; saline-LNAME: F(2,
116) = 3.5, p < .05; LNAME-saline: F(2, 110) = 6.3, p < .01;
LNAME-LNAME: F(2, 107) = 3.6, p < .05. Response duration
scores are more sensitive for revealing response decrements than
are frequency data (Smith, 1997, 1998). Thus it is not surprising
that the decrements are significant with this response metric.
Nevertheless, the generalization gradients were flatter when
LNAME was active before conditioning. The responses to Odor D
in those groups were significantly higher than in the two groups in
which saline was applied before conditioning, F(3, 149) = 4.5,
p < .01. Response levels to Odor C, F(3, 149) = 0.7, ns, and to
Odor S, F(3, 149) = 1.2, ns, did not differ across groups, although
there was a trend toward higher response to Odor S in the LNAME
pretreatment groups.
Dual application of PCT revealed a slightly different effect (see
Figure 6). Comparison of response duration scores within groups
revealed a significant decrement in all cases, saline-saline: F(2,
56) = 7.9, p < .01; saline-PCT: F(2, 56) = 5.6, p < .01;
PCT-saline: F(2, 50) = 9.2, p < .001; PCT-PCT: F(2,54) = 5.6,
p < .01. Post hoc testing revealed that the response to Odor C was
on average longer than that to Odors S and D in the saline control
group, ((27) = 4.7 and 3.4, for C versus S and C versus D,
respectively, p < .01 in both cases (see Figure 6A). In all three
remaining dual-injection treatment groups the response to Odor C
was greater than that to Odor D, saline-PCT: t(21) = 2.8,p< .01;
PCT-saline: t(26) = 3.9, p < .001; PCT-PCT: r(26) = 3.4, p <
Table 2
Effect of Different Concentrations of LNAME and DNAME
on the Percentage of Subjects That Responded to a
Conditioned Odor (C), a Structurally Similar Odor (S),and a Dissimilar Odor (D)
Treatment X2
SalineLNAME
200 ^M20 /AM
DNAME200 /iM
20 ^M
X*
37
3839
4039
91.9
97.497.4
92.589.73.2 ns
67.6
81.674.4
85.076.9
3.9 ns
45.9
78.948.7
45.051.313.0*
18.1**
6.3ns23.8**
27.2**15.1**
Note. Bees were treated 10 min before conditioning with either saline ora saline-containing drug treatment at the indicated concentration (seeMethod section; Figure 1). Ninety minutes later, each bee was tested with
Odors C, S, and D, presented in a randomized sequence. For each group,the numbers indicate the percentage of n that responded to the odors.LNAME = N-nitro-L-arginine methyl ester; DNAME = N-nitro-D-arginine methyl ester. Chi-square values to the right indicate comparisonwithin individuals across the three odors. Chi-square values below thecolumns indicate comparison of responses to odor across treatment groups.Significance levels are adjusted to maintain an experimentwise error rate of.05 (Sokal & Rohlf, 1995; *p < .01; **/? < .005).

520 HOSLER, BUXTON, AND SMITH
O
A. saline
B.
c
D
LNAME
D
DNAME
D
LNAME+SNAP
C s DLNAME(OL)
S DC
I I saline
^H LNAME
I I DNAME
E%%3 LNAME+SNAPI : : : : • : :| LNAME on optic lobes
* different from C (in A-E) orsaline (in F-H), p < 0.05
Figure 3. The effect of nitric oxide synthase inhibition by N-nitro-L-arginine methyl ester (LNAME) appliedto the antennal lobe and its recovery by coapplication of the nitric oxide donor S-nitrosacetylpenicillamine(SNAP). In this experiment, subjects received a single application of saline or saline + drug treatment to eachantennal lobe 10 min before conditioning with the conditioned odor C (Figure 1A). Ninety minutes later, theywere tested with C, a similar odor (S), and a dissimilar odor (D), in randomized order. Subjects' responses werevideotaped for calculation of response duration to a standard 4-s odor test pulse (see Method). The mean (± SEM)response duration to each test stimulus is presented in each graph, A-E: Comparison of response levels to OdorsC, S, and D within each treatment group. Asterisks indicate significant differences of respective points from C,which was assayed by I test (Sokal & Rohlf, 1995). Sample sizes: saline (A: n = 43), LNAME (B: n ~ 36),N-nitro-D-arginine methyl ester (DNAME) (C: n - 37), and LNAME + SNAP {D: n = 35) applications to eachantennal lobe. E: Same information for the group (n = 38) that received LNAME application to each opticlobe 10 min before conditioning. F-H: Comparison of response levels to Odors C, S, and D across groups.
.01. But the response levels to Odors C and S were statisticallyequivalent, saline-PCT: f(27) = 0.5, ns; PCT-saline: f(26) =1.1,ns; PCT-PCT: r(26) = 0.9, ns.
Comparison of response levels to Odors C, S, and D acrossgroups-(Figures 6E-F) confirm stronger responses to Odor S in alldrug treatment groups, F(3, 108) - 3.3, p < .05. But responselevels to odor never differed across groups, F(3, 108) = 1.0, ns, orto D, F(3, 108) = 0,5, ns.
Tests for Nonassociative Effects of LNAME and PCT
Treatment with PCT or LNAME failed to increase sensitizationresponses to odor (Figure 7). In the groups that were tested 15 safter feeding, the mean response duration was slightly lower in theLNAME-treated group, and it was substantially lower in the PCT-treated group, relative to the group treated with saline. But thesetrends are opposite the prediction for sensitization. Thus, sensiti-zation cannot explain the elevation in responses after associativeconditioning. Response durations differed across groups at this testpoint, F(2, 85) = 5.0, p < .01. Response levels in the PCT groupare significantly shorter than those of the saline control (LSD, p <
.05). The trend between LNAME and saline groups fell short ofstatistical significance. The groups tested 60 s after feeding did notdiffer in regard to response probability or mean duration, F(2,86) = 0.5, ns.
Discussion
Our results imply that NO release is involved in syaaptic plas-ticity that is important for setting up odor representations in thehoney bee AL. Blockade of NO release impairs C-from-D discrim-ination when the drug is active at the time of conditioning. It isreasonable to conclude that the locus for this effect lies in the ALcircuitry, because application to the optic neuropil failed to affectolfactory discrimination. If the drug had spread from the AL to thecorpora pedunculata, which are also involved in olfactory learningand show NOS staining, then it should also have spread there fromthe optic neuropils. Furthermore, the activity in the AL mayinvolve both nonassociative and associative mechanisms in syn-aptic transmission (Miiller & Hildebrandt, 1995). Inhibition ofNOS tended to reduce sensitization responses. The associativeeffect was revealed by an elevated generalization response when

NITRIC OXIDE MODULATION OF OLFACTORY PROCESSING 521
A. saline C. I DNAME+SNAP
B. DNAME
* different from C, p < 0.05
Figure 4. The effect of nitric oxide synthase inhibition by N-nitro-D-arginine methyl ester (DNAME) applied
to the antennal lobe and its recovery by coapplication of the nitric oxide donor S-nitrosacetylpenicillamine
(SNAP). In this experiment, subjects received a single application of saline or saline -f drug treatment to each
antennal lobe 10 min before conditioning with the conditioned odor (Odor C; Figure 1A). Ninety minutes later,
they were tested with Odor C, a similar odor (S), and a dissimilar odor (D), in randomized order. Subjects'
responses were videotaped for calculation of response duration to a standard 4-s odor test pulse (see Method).
The mean (± SEM) response duration to each test stimulus is presented in each graph. A-C: Comparison of
response levels to Odors C, S, and D within each treatment group. Asterisks indicate significant differences of
respective points from C. which was assayed by t test (Sokal & Rohlf, 1995). Sample sizes: saline (A: n = 21),
DNAME (B: n = 20), DNAME+SNAP (C: n = 19).
NOS was blocked before conditioning but not when it was blocked
before testing.
We feel that the effect of LNAME can in large part be attributed
to a blockade of NO release. Two lines of argument support this
conclusion: First, the effect of LNAME on C-from-D discrimina-
tion is recovered by coapplication of SNAP, which is an NO
donor. If the effect of LNAME were not caused by blockade of NO
Table 3
Effect of Dual Application of Saline and/or LNAME to the
Honey Bee Antennal Lobes, Before Conditioning and Testing
Treatment
Saline-salineSaline-LNAME
LNAME-salineLNAME-LNAME
if
n
37
37
40
37
C
86.582.5
86.591.9
1.5 ns
S
64.980.0
86.586.57.1*
D
59.547.5
75.781.112.0**
X2
7.25*14.5**
2.0 ns1.9 ns
Note, flees were treated 10 min before conditioning with either saline ora saline-containing drug treatment at the indicated concentration (seeMethod section: Figure 1). Ninety minutes later, each bee was treated again
with either the same application (saline-saline and N-nitro-L-argininemethyl ester [LNAMEJ-LNAME) or with a different one (saline-LNAMEand LNAME-saline). Ten minutes after the second application, each beewas tested with the conditioned odor (C), a structurally similar odor (S),and a dissimilar odor (D), presented in a randomized sequence. For eachgroup, the numbers indicate the percentage of n that responded to theodors. Chi-square values to the right indicate comparison within individ-uals across the three odors. Chi-square values below the columns indicatecomparison of responses to odor across treatment groups. Significancelevels are adjusted to maintain an experimentwise error rale of .05 (Sokal& Rohlf, 1995; * p < .01; ** p < .005).
but rather by a different mechanism or artifact of the injection, then
artificial elevation of NO with SNAP would not be expected to
recover the behavior. Second, DNAME failed to have the same
magnitude of impact as LNAME, although it did impair discrim-
ination of Odor C from Odor S. Physiological analyses should now
be used to reveal the nature of the DNAME effect. It could arise
if DNAME were not completely inactive, but instead just has a
dramatically reduced impact on NOS. Therefore, when the dis-
crimination became difficult, as with Odors C and S, and the
behavioral test became very sensitive, the effect of DNAME might
be more easily revealed. This interpretation is given weight by our
finding that SNAP can recover the C-from-S discrimination in
DNAME-treated groups. That C-from-S discrimination was recov-
ered by SNAP in the DNAME group but not in the LNAME group
could be due to the relatively stronger effect of LNAME in
blocking NOS. It is also possible that the DNAME was contami-
nated by a small amount of LNAME, although this seems unlikely
because we observed the effect of DNAME across two investiga-
tors and different sources of the drug.
At present we do not have detailed electrophysiological analyses
of NO effects in the honey bee AL, as has been done for GABA
(Stopfer et al., 1997). However, drugs that block NO or GABA
have somewhat different effects in our behavioral assays. PCT
blockade of GABAergic transmission impairs C-from-S discrimi-
nation, but it never impaired discrimination of Odor C from the
dissimilar Odor D. Furthermore, PCT had this effect regardless of
whether the drug was active before conditioning or before testing.
PCT treatment also blocks synchronization of membrane poten-
tials among AL interneurons, but it leaves their response patterns
to odor intact. Thus, when two odors elicit activity in spatially

522 HOSLER, BUJCTON, AND SMITH
OH
§
I
I
A. saline-saline C. N\LNAME-saline
D
D. T LNAME-LNAME
D C S D
I I saline-saline
^^ saline-LNAME
I I LNAME-saline
^B LNAME-LNAME
* different from C (A-D) orsaline-saline (E-G), p < 0.05 or 0.01
Figure 5. Effect of inhibition of nitric oxide release during conditioning and/or recall testing. Treatment groupsin this experiment received two applications of saline (with or without drug treatment) to each antenna! lobe(Figure IB). A-D: Within-group comparisons of responses to the conditioned odor (C), a similar odor (S), anda dissimilar odor (D) after (conditioning-testing) treatment with saline-saline (A: n — 38), saline-N-nitro-L-arginine methyl ester (LNAME, B: n = 40), LNAME-saline (C: n = 38), or LNAME-LNAME (D: n = 37).Mean (± SEM) response durations and percentage data presented as in Figures 3 and 4. E-G: Between-groupscomparisons of responses to Odors C, S, and D.
segregated, distinct sets of interneurons in the AL (Joerges et al,,1997), as one might expect of Odors C and D, then the two odorscan still be discriminated even when temporal information is lostunder PCT treatment (Laurent, 1996; MacLeod & Laurent, 1996;Stopfer et al., 1997; Wehr & Laurent, 1996). When there issignificant overlap in neural representation, such as when activityin the same cells code for two odors, as has been proposed forOdors C and S (Stopfer et al., 1997), then the information that ispresent in the sequence of evolving assemblies (Laurent, 1996) isnecessary for effective discrimination. PCT treatment is sufficientto eliminate all detectable traces of this temporal coding, and itdisrupts C-from-S, but not C-from-D, discrimination.
If the effect of LNAME treatment was simply to eliminatetemporal synchronization, then it should have had a behavioraleffect that is identical to PCT treatment. But blockade of NOS byLNAME treatment impairs C-from-D discrimination, which wehave never observed for PCT treatment {Stopfer et al., 1997). Itcould be that NO and GAB A affect the different physiological andbiochemical processes in the same pathway. LNAME might havea qualitatively stronger effect on NOS than PCT has on blockadeof Cl— channels associated with GABA receptors. Thus, thedifferences in our study could be attributable to quantitative ratherthan qualitative differences in the drug effects. Indeed, a lowerconcentration of LNAME, and higher concentrations of DNAME,produced generalization patterns similar to PCT, which would beconsistent with this interpretation.
However, we must also consider whether NO and GABA affectdifferent processes in the AL. That LNAME has its effect when itis active before conditioning would support this argument. It mightbe that die role of NO release in olfactory pathways may bedifferent in the honey bee AL than it is in the mollusk procerebralLobe (Gelperin, 1994). In the locust, oscillations develop as a resultof multiple stimulations over short time intervals (Stopfer & Lau-rent, 1998). After three to five stimulations, the oscillation ampli-tude increases and the spectral peak sharpens considerably. PCTtreatment eliminates oscillation in the local field potential evenafter it has formed, which would seem to implicate GABA in themaintenance of synchronization. But it is conceivable that NOrelease is involved in the formation, but not the maintenance, ofsynchronization over multiple trials, which might account for ourdata.
In the moilusk, the circuitry of the lobes gives rise to sponta-neous synchronized activity under an unstimulated condition(Gelperin & Tank, 1990). Natural odor stimulation causes theoscillations to switch from a propagating to a standing waveform(Delaney et al,, 1994). At 0.5-2.0 Hz, the oscillations are slowerthen they are hi the locust (20 Hz; Laurent & Davidowitz, 1994) orin the honey bee (30 Hz; Stopfer et al., 1997). Moreover, there isat present no indication that a propagating wave exists in the insectAL under unstimulated conditions. In this study, we provide evi-dence from our behavioral analyses that the physiological role of
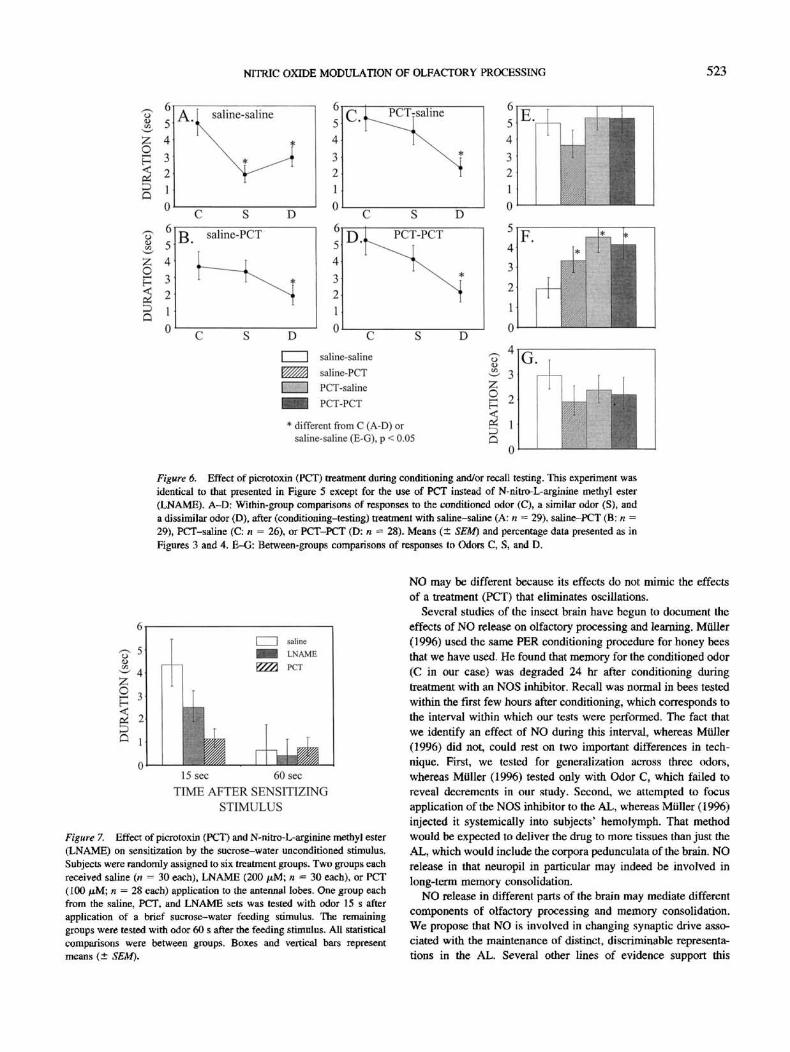
NITRIC OXIDE MODULATION OF OLFACTORY PROCESSING 523
zo
iDQ
O
saline-saline
D
g saline-PCT
D0
saline-saline
saline-PCT
PCT-saline
PCT-PCT
1 different from C (A-D) orsaline-saline (E-G), p < 0.05
Figure 6. Effect of picrotoxin (PCT) treatment during conditioning and/or recall testing. This experiment wasidentical to that presented in Figure 5 except for the use of PCT instead of N-nitro-L-arginine methyl ester(LNAME). A—D: Within-group comparisons of responses to the conditioned odor (C), a similar odor (S), anda dissimilar odor (D), after (conditioning-testing) treatment with saline-saline (A; n = 29), saline-PCT (B; n =29), PCT-saline (C: n = 26), or PCT-PCT (D: n = 28). Means (± SEM) and percentage data presented as inFigures 3 and 4. E-G: Between-groups comparisons of responses to Odors C, S, and D.
6
t? 5
u- 4
|3H
I I salineHi LNAME
15 sec 60 sec
TIME AFTER SENSITIZINGSTIMULUS
Figure 7. Effect of picrotoxin (PCT) and N-nitro-L-arginine methyl ester(LNAME) on sensitization by the sucrose-water unconditioned stimulus.Subjects were randomly assigned to six treatment groups. Two groups eachreceived saline (n = 30 each), LNAME (200 jttM; n = 30 each), or PCT(100 fiM; n - 28 each) application to the antennal lobes. One group eachfrom the saline, PCT, and LNAME sets was tested with odor 15 s afterapplication of a brief sucrose-water feeding stimulus. The remaininggroups were tested with odor 60 s after the feeding stimulus. All statisticalcomparisons were between groups. Boxes and vertical bars representmeans (± SEM).
NO may be different because its effects do not mimic the effectsof a treatment (PCT) that eliminates oscillations.
Several studies of the insect brain have begun to document theeffects of NO release on olfactory processing and learning. Milller(1996) used the same PER conditioning procedure for honey beesthat we have used. He found that memory for the conditioned odor(C in our case) was degraded 24 hr after conditioning duringtreatment with an NOS inhibitor. Recall was normal in bees testedwithin the first few hours after conditioning, which corresponds tothe interval within which our tests were performed. The fact thatwe identify an effect of NO during this interval, whereas Mtiller(1996) did not, could rest on two important differences in tech-nique. First, we tested for generalization across three odors,whereas Miiller (1996) tested only with Odor C, which failed toreveal decrements in our study. Second, we attempted to focusapplication of the NOS inhibitor to the AL, whereas Miiller (1996)injected it systemically into subjects' hemolymph. That methodwould be expected to deliver the drug to more tissues than just theAL, which would include the corpora pedunculata of the brain. NOrelease in that neuropil in particular may indeed be involved inlong-term memory consolidation,
NO release in different parts of the brain may mediate differentcomponents of olfactory processing and memory consolidation.We propose that NO if, involved in changing synaptic drive asso-ciated with the maintenance of distinct, discriminable representa-tions in the AL. Several other lines of evidence support this

524 HOSLER, BUXTON, AND SMITH
position. NO can affect several physiological processes (Mullet,
1997), one of which is the activation of a specific cyclic guanosine
monophosphate (cGMP). Nighotn, Gibson, Rivers, Hildebrand,
and Morton (1998) showed that NO elevates cGMP in projection
neurons once it is released from sensory afferents in the moth,
Manduca xexta. Furthermore, Griinbaum and Mu'ller (1998)
showed that a protein kinase (PKC) is activated in the AL after
associative pairing of an odor with sucrose reinforcement. The
pattern of activation is consistent with an effect of PKC on
intermediate-term components of protein-synthesis-dependent
memory in the AL.
At present it is unclear how the physiological effects of NO in
the AL are involved in setting up the representations we propose.
But a recent computational model that incorporated known neural
elements of AL processing in the honey bee demonstrated how
associative modulation of lateral inhibitory processes can serve to
enhance (or degrade if blocked) discriminability of odors (Linster
& Smith, 1997). The model incorporated a representation of an
identified neuron in the honey bee brain that represents the sucrose
US (Hammer, 1993; Hammer & Menzel, 1995). That cell, called
VUMmxl (Hammer, 1993), shows little or no response to an odor
before pairing of the odor with sucrose reinforcement. But, on
forward pairing of odor and sucrose reinforcement, the VUM
begins to respond to odor. Furthermore, depolarization of this cell
in the context of an odor is sufficient to replace the US (Hammer,
1993). VUM arborizes within many if not all of the glomeruli in
the AL. Linster and Smith (1997) proposed that coactivation of
afferent sensory input and the VUM modifies and thereby poten-
tiates lateral inhibitory processes. This enhances the ability of an
odor representation in the AL to suppress activation of neighboring
glomeruli, which would enhance discrimination of that odor from
others that activate similar sets of glomeruli. Tf NO is involved in
associative modification of synapses onto lateral inhibitory pro-
cesses, then blockade of NO release may have the effect we
observed.
Interestingly, the disruption of discrimination that we report is
analogous to the pattern established for NO-mediated discrimina-
tion of maternal odors in sheep (Kendrick et al., 1997). In the
sheep, synaptic plasticity is involved in maternal recognition of
lamb odor, which is learned around the time of birth (Kendrick,
Levy, & Keverne, 1992). Disruption of NO release at this time
impairs a ewe's ability to discriminate her own offspring's odor
from that of a foreign offspring. However, if NO release is dis-
rupted at the time of testing, this discrimination is left intact.
The similarity of these patterns provides a basis for arguing that
there is a functional analogy between NO release in the vertebrate
OB and the insect AL, which has also been argued on the basis of
anatomical similarities (Hildebrand & Shepherd, 1997). NO re-
lease from pre- and postsynaptic sites in the vertebrate OB poten-
tiates the sensitivity of mitral-to-granule cell synapses to glutamate
(Kendrick et al., 1992, 1997). The increase in sensitivity potenti-
ates the NMDA-driven plasticity that occurs at these synapses
(Kendrick et al., 1997). This synaptic modification is analogous to
the changes in synaptic efficacy that take place at dendrodendritic
synapses in the mouse accessory olfactory bulb as a result of
experience with odor (Brennan, Kaba, & Keverne, 1990). Further
physiological investigation of the insect AL should now be under-
taken to reveal in more detail these similarities and differences.
References
Benatar, S. T., Cobey, S., & Smith, B. H. (1995). Selection on a haploid
genotype for discrimination learning performance: Correlation between
drone honey bees (Apis tnellifera) and their worker progeny. Journal of
Insect Behavior, 8, 637-652.
Breer, H., & Shepherd, G. M. (1993). Implications of the NO/cGMP
system for olfaction. Trends in Neuroscience, 16, 5-9.
Brennan, P., Kaba, H., & Keverae, E. B. (1990, November 30). Olfactory
recognition: A simple memory system. Science, 250, 1223-1226.
Brennan, P. A., & Keverne, E. B. (1997). Neural mechanisms of mamma-
lian olfactory learning. Progress in Neurobiology, 51, 457-481.
Christensen, T. A., Waldrop, B. R., & Hildebrand, J. G. (1998). Multitask-
ing in the olfactory system: Context-dependent responses to odors reveal
dual GABA-regulated coding mechanisms in single olfactory projection
neurons. Journal of Neuroscience, IS, 5999-6008.
Delaney, K. R., Gelperin, A., Fee, M. S., Flores, J. A., Gervais, R., Tank,
D. W., & Kleinfeld, D. (1994). Waves and stimulus-modulated dynam-
ics in an oscillating olfactory network. Proceedings of the National
Academy of Sciences, USA, 91, 669-673.
Gelperin, A. (1994, May 5). Nitric oxide mediates network oscillations of
olfactory interneurons in a terrestrial mollusc. Nature, 369, 61-63.
Gelperin, A., & Tank, D. W. (1990, May 31). Odour-modulated collective
network oscillations of olfactory interneurons in a terrestrial mollusc.
Nature, 345, 437-440.
Griinbaum, L., & Miiller, U. (1998). Induction of specific olfactory mem-
ory leads to a long-lasting activation of protein kinase C in the antennal
lobe of the honey bee. Journal of Neuroscience, 11, 4384-4392.
Hammer, M. (1993, November 4). An identified neuron mediates the
unconditioned stimulus in associative olfactory learning in honeybees.
Nature, 366, 59-63.
Hammer, M., & Menzel, R. (1995). Learning and memory in the honeybee.
Journal of Neuroscience, 15, 1617-1630.
Hildebrand, J. G. (1995). Analysis of chemical signals by nervous systems.
Proceedings of the National Academy of Sciences, USA, 92, 67-74.
Hildebrand, J. G., & Shepherd, G. M. (1997). Mechanisms of olfactory
discrimination: Converging evidence for common principles across
phyla. Annual Review of Neuroscience, 20, 595-631.
Homberg, U., Christensen, T. A., & Hildebrand, J. G, (1989). Structure and
function of the deuterocerebrum in insects. Annual Review of Entomol-
ogy, 43, 477-501.
Homberg, U., Kingan, T. G., & Hildebrand, J. G. (1987). Immunocyto-
chemistry of GABA in the brain and suboesophageal ganglion of
Manduca sexta. Cell and Tissue Research, 248, 1-24.
Ignarro, L. J., Buga, G. M., Wood, K. S., Byrnes, R. E., & Chaudhuri, G.
(1987). Endothelium-derived relaxing factor produced and released from
artery and vein is nitric oxide. Proceedings of the National Academy of
Sciences, USA, 84, 9265-9269.
Joerges, J., KUttner, A., Galizia, C. G., & Menzel, R. (1997, May 15).
Representations of odours and odour mixtures visualized in the honey-
bee brain. Nature, 387, 285-288.
Kendrick, K. M., Guevara-Guzman, R., Zorrilla, J.. Hinton, M. R., Broad,
K. D., Mimmack, M., & Ohkura, S. (1997. August 14). Formation of
olfactory memories mediated by nitric oxide. Nature, 388, 670-674.
Kendrick, K. M., Levy, F., & Keverne, E. B. (1992, May 8). Changes in the
sensory processing of olfactory signals induced by birth in sheep. Sci-
ence, 256. 833-836.
Laurent, G. (1996). Dynamical representation of odors by oscillating and
evolving neural assemblies. Trends in Neuroscience, 19, 489-496.
Laurent, G., & Davidowitz, H. (1994, September 23). Encoding of olfac-
tory information with oscillating neural assemblies. Science, 265, 1872-
1875.
Laurent, G., & Naraghi, M. (1994). Odorant-induced oscillations in the
mushroom bodies of the locust. Journal of Neuroscience, 14, 2993-
3004.

NITRIC OXIDE MODULATION OF OLFACTORY PROCESSING 525
Leitch, B., & Laurent, G. (1996). GAB Aergic synapses in the antennal lobe
and mushroom body of the locust olfactory system. Journal of Compar-
ative Neurology, 372, 487-514.
Lev-Ram, V., Makings, L. R., Keitz, P. F., Kao, J. P. Y., & Tsien, R. Y.
(1995). Long-term depression in cerebellar purkinje neurons results from
coincidence of nitric oxide and depolarization-induced Ca2+ transients.
Neuron, 15, 407.
Linster, C, & Smith, B. H. (1997). A computational model of the response
of honey bee antennal lobe circuitry to odor blends: Overshadowing,
blocking and unblocking can arise from lateral inhibition. Behavioral
Brain Research, 87, 1-14.
Mackintosh, N. J. (1983). Conditioning and associative learning. Oxford,
England: Oxford University Press.
MacLeod, K., & Laurent, G. (1996, November 8). Distinct mechanisms for
synchronization and temporal patterning of odor-encoding neural assem-
blies. Science, 274, 976-979.
Menzel, R. (1990). Learning, memory and "cognition" in honey bees. In
R. P. Kesner & D. S. Olton (Fxls.), Neurobiology of comparative cog-
nition (pp. 237-292). Hillsdale, NJ: Erlbaum.
Menzel, R., & Bitterman, M. E. (1983). Learning by honeybees in an
unnatural situation. In F. Huber & H. Markl (Ms.), Neuroethology and
behavioral physiology (pp. 206-215). New York: Springer-Verlag.
Menzel, R., & Mdller, U. (1996). Learning and memory in honeybees:
From behavior to neural substrates. Annual Review of Neuroscience, 19,
379-404.
Mori, K. (1995). Relation of chemical structure to specificity of response
in olfactory glomeruli. Current Opinion in Neurobiology, 5, 467-474.
MUIler, U. (1994). Ca2+/calmodulin-dependent nitric oxide synthase in
Apis mellifera and Drosophila melanogaster. European Journal of Neu-
roscience, 6, 1362-1370.
Miiller, U. (1996). Inhibition of nitric oxide synthase impairs a distinct
form of long-term memory in the honeybee, Apis mellifera. Neuron, 16,
541-549.
MUIler, U. (1997). The nitric oxide system in insects. Progress in Neuro-
biology, 51, 363-381.
MUIler, U., & Bicker, G. (1994). Calcium-activated release of nitric oxide
and cellular distribution of nitric-oxide-synthesizing neurons in the
nervous system of the locust. Journal of Neuroscience, 14, 7521-7528.
Mltller, U., & Hildebrandt, H. (1995). The nitric oxide/cGMP system in the
antennal lobe of Apis mellifera is implicated in integrative processing of
chemosensory stimuli. European Journal of Neuroscience, 7, 2240-
2248.
Nighom, A., Gibson, N. J., Rivers, D. M., Hildebrand, J. G., & Morton,
D. B. (1998). The nitric oxide-cGMP pathway may mediate communi-
cation between sensory afferents and projection neurons in the antennal
lobe of Manduca sexta. Journal of Neuroscience, 18, 7244-7255.
Palmer, R. J., Ferrige, A. G., & Moncada, S. (1987, June 11). Nitric oxide
release accounts for the biological activity of endothehum-derived re-
laxing factor. Nature, 327, 524-526.
Park, J.-H., Straub, V. A., & O'Shea, M. (1998). Anterograde signaling by
nitric oxide: Characterization and in vitro reconstitution of an identified
nitrergic synapse. Journal of Neuroscience, 14, 5463-5476.
Regulski, M., & Tully, T. (1995). Molecular and biochemical character-
ization of dNOS: A Drosophila Ca+ +/calmodulin-dependent nitric ox-
ide synthase. Proceedings of the National Academy of Sciences,
USA, 92, 9072-9076.
Shipley, M. T., & Ennis, M. (1996). Functional organization of olfactory
system. Journal of Neurobiology, 30, 123-176.
Schuman, E. M., & Madison, D. V. (1991, December 6). A requirement for
the intercellular messenger nitric oxide in long-term potentiation. Sci-
ence, 254, 1503-1506.
Schuman, E. M., & Madison, D. V. (1994). Nitric oxide and synaptic
function. Annual Review of Neuroscience, 17, 153-183.
Smith, B. H. (1997). An analysis of blocking in binary odorant mixtures:
An increase but not a decrease in intensity of reinforcement produces
unblocking. Behavioral Neuroscience, 11, 57-69.
Smith, B. H. (1998). An analysis interaction in binary odorant mixtures.
Physiology and Behavior, 65, 397-407.
Smith, B. H., & Menzel, R. (1989). The use of electromyogram recordings
to quantify odorant discrimination in the honey bee, Apis mellifera.
Journal of Insect Physiology, 35, 369-375.
Snyder, S. H., Jaffrey, S. R., & Zakhary, R. (1998). Nitric oxide and carbon
monoxide: Parallel roles as neural messengers. Brain Research Re-
views, 26, 167-175.
Sokal, R. R., & Rohlf, F. J. (1995). Biometry. New York: W. H. Freeman.
Stopfer, M., Bhagavan, S., Smith, B. H., & Laurent, G. (1997, November
6). Impaired odour discrimination on desynchronization of odour-
encoding neural assemblies. Nature, 390, 70-74,
Stopfer, M., & Laurent, G. (1998). Short term plasticity in locust olfaction.
Society for Neuroscience Abstracts, 24 (Pan 1), 910.
Wehr, M., & Laurent, G. (1996, November 14). Odour encoding by
temporal sequences of firing in oscillating neural assemblies. Nature,
384, 162-166.
Received January 29, 1999
Revision received October 31, 1999
Accepted December 8, 1999
Related Documents











