Research Article Impairment of Electron Transfer Chain Induced by Acute Carnosine Administration in Skeletal Muscle of Young Rats José Roberto Macarini, 1 Soliany Grassi Maravai, 1 José Henrique Cararo, 1 Nádia Webber Dimer, 1 Cinara Ludvig Gonçalves, 2 Luiza Wilges Kist, 3,4,5 Mauricio Reis Bogo, 3,4,5 Patrícia Fernanda Schuck, 1 Emilio Luiz Streck, 2,5 and Gustavo Costa Ferreira 6 1 Laborat´ orio de Erros Inatos do Metabolismo, Programa de P´ os-Graduac ¸˜ ao em Ciˆ encias da Sa´ ude, Unidade Acadˆ emica de Ciˆ encias da Sa´ ude, Universidade do Extremo Sul Catarinense, 88801-600 Crici´ uma, SC, Brazil 2 Laborat´ orio de Bioenerg´ etica, Programa de P´ os-Graduac ¸˜ ao em Ciˆ encias da Sa´ ude, Unidade Acadˆ emica de Ciˆ encias da Sa´ ude, Universidade do Extremo Sul Catarinense, 88806-000 Crici´ uma, SC, Brazil 3 Laborat´ orio de Biologia Genˆ omica e Molecular, Faculdade de Biociˆ encias, Pontif´ ıcia Universidade Cat´ olica do Rio Grande do Sul, 90619-900 Porto Alegre, RS, Brazil 4 Programa de P´ os-Graduac ¸˜ ao em Medicina e Ciˆ encias da Sa´ ude, Pontif´ ıcia Universidade Cat´ olica do Rio Grande do Sul, 90619-900 Porto Alegre, RS, Brazil 5 Instituto Nacional de Ciˆ encia e Tecnologia Translacional em Medicina (INCT-TM), 90035-003 Porto Alegre, RS, Brazil 6 Laborat´ orio de Neuroqu´ ımica, Instituto de Biof´ ısica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, Avenida Carlos Chagas Filho 373, Cidade Universit´ aria, Ilha do Fund˜ ao, 21941-902 Rio de Janeiro, RJ, Brazil Correspondence should be addressed to Gustavo Costa Ferreira; [email protected] Received 10 February 2014; Revised 6 April 2014; Accepted 7 April 2014; Published 4 May 2014 Academic Editor: Marcos Roberto de Oliveira Copyright © 2014 Jos´ e Roberto Macarini et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Serum carnosinase deficiency is an inherited disorder that leads to an accumulation of carnosine in the brain tissue, cerebrospinal fluid, skeletal muscle, and other tissues of affected patients. Considering that high levels of carnosine are associated with neurological dysfunction and that the pathophysiological mechanisms involved in serum carnosinase deficiency remain poorly understood, we investigated the in vivo effects of carnosine on bioenergetics parameters, namely, respiratory chain complexes (I–III, II, and II- III), malate dehydrogenase, succinate dehydrogenase, and creatine kinase activities and the expression of mitochondrial-specific transcription factors (NRF-1, PGC-1, and TFAM) in skeletal muscle of young Wistar rats. We observed a significant decrease of complexes I–III and II activities in animals receiving carnosine acutely, as compared to control group. However, no significant alterations in respiratory chain complexes, citric acid cycle enzymes, and creatine kinase activities were found between rats receiving carnosine chronically and control group animals. As compared to control group, mRNA levels of NRF-1, PGC-1, and TFAM were unchanged. e present findings indicate that electron transfer through the respiratory chain is impaired in skeletal muscle of rats receiving carnosine acutely. In case these findings are confirmed by further studies and ATP depletion is also observed, impairment of bioenergetics could be considered a putative mechanism responsible for the muscle damage observed in serum carnosinase- deficient patients. 1. Introduction Carnosine (-alanyl-L-histidine) is an imidazole dipeptide abundant in skeletal muscle, as well as in central nervous system [1], cardiac muscle, kidney, stomach, and olfactory bulbs [2, 3] of most animals. Physiological levels of carnosine may reach up to 20 mM in muscle tissues [4], and its biological role remains unclear. Carnosine of human diet Hindawi Publishing Corporation BioMed Research International Volume 2014, Article ID 632986, 10 pages http://dx.doi.org/10.1155/2014/632986

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Research ArticleImpairment of Electron Transfer Chain Induced by AcuteCarnosine Administration in Skeletal Muscle of Young Rats
Joseacute Roberto Macarini1 Soliany Grassi Maravai1
Joseacute Henrique Cararo1 Naacutedia Webber Dimer1 Cinara Ludvig Gonccedilalves2
Luiza Wilges Kist345 Mauricio Reis Bogo345 Patriacutecia Fernanda Schuck1
Emilio Luiz Streck25 and Gustavo Costa Ferreira6
1 Laboratorio de Erros Inatos do Metabolismo Programa de Pos-Graduacao em Ciencias da Saude Unidade Academica deCiencias da Saude Universidade do Extremo Sul Catarinense 88801-600 Criciuma SC Brazil
2 Laboratorio de Bioenergetica Programa de Pos-Graduacao em Ciencias da Saude Unidade Academica deCiencias da Saude Universidade do Extremo Sul Catarinense 88806-000 Criciuma SC Brazil
3 Laboratorio de Biologia Genomica e Molecular Faculdade de Biociencias Pontifıcia Universidade Catolica do Rio Grande do Sul90619-900 Porto Alegre RS Brazil
4 Programa de Pos-Graduacao em Medicina e Ciencias da Saude Pontifıcia Universidade Catolica do Rio Grande do Sul90619-900 Porto Alegre RS Brazil
5 Instituto Nacional de Ciencia e Tecnologia Translacional em Medicina (INCT-TM) 90035-003 Porto Alegre RS Brazil6 Laboratorio de Neuroquımica Instituto de Biofısica Carlos Chagas Filho Universidade Federal do Rio de JaneiroAvenida Carlos Chagas Filho 373 Cidade Universitaria Ilha do Fundao 21941-902 Rio de Janeiro RJ Brazil
Correspondence should be addressed to Gustavo Costa Ferreira gustavolhegmailcom
Received 10 February 2014 Revised 6 April 2014 Accepted 7 April 2014 Published 4 May 2014
Academic Editor Marcos Roberto de Oliveira
Copyright copy 2014 Jose Roberto Macarini et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Serum carnosinase deficiency is an inherited disorder that leads to an accumulation of carnosine in the brain tissue cerebrospinalfluid skeletalmuscle and other tissues of affected patients Considering that high levels of carnosine are associatedwith neurologicaldysfunction and that the pathophysiological mechanisms involved in serum carnosinase deficiency remain poorly understood weinvestigated the in vivo effects of carnosine on bioenergetics parameters namely respiratory chain complexes (IndashIII II and II-III) malate dehydrogenase succinate dehydrogenase and creatine kinase activities and the expression of mitochondrial-specifictranscription factors (NRF-1 PGC-1120572 and TFAM) in skeletal muscle of young Wistar rats We observed a significant decreaseof complexes IndashIII and II activities in animals receiving carnosine acutely as compared to control group However no significantalterations in respiratory chain complexes citric acid cycle enzymes and creatine kinase activities were found between rats receivingcarnosine chronically and control group animals As compared to control group mRNA levels of NRF-1 PGC-1120572 and TFAM wereunchanged The present findings indicate that electron transfer through the respiratory chain is impaired in skeletal muscle of ratsreceiving carnosine acutely In case these findings are confirmed by further studies and ATP depletion is also observed impairmentof bioenergetics could be considered a putative mechanism responsible for the muscle damage observed in serum carnosinase-deficient patients
1 Introduction
Carnosine (120573-alanyl-L-histidine) is an imidazole dipeptideabundant in skeletal muscle as well as in central nervous
system [1] cardiac muscle kidney stomach and olfactorybulbs [2 3] of most animals Physiological levels of carnosinemay reach up to 20mM in muscle tissues [4] and itsbiological role remains unclear Carnosine of human diet
Hindawi Publishing CorporationBioMed Research InternationalVolume 2014 Article ID 632986 10 pageshttpdxdoiorg1011552014632986
2 BioMed Research International
is uptaken by intestinal proton-coupled peptide transporternamely human H+peptide cotransporter 1 (hPEPT1) [5]This dipeptide is synthesized starting from its componentamino acids histidine and 120573-alanine by enzyme carnosinesynthetase (EC 63211) On the other hand the clearanceof carnosine is essentially due to the hydrolytic action of ametalloprotease the carnosinase [6] This enzyme exists intwo isoforms The secreted form namely serum carnosinase(EC 341320) is found primarily in serum as well as inbrain and cerebrospinal fluid The cytosolic form tissuecarnosinase (EC 34133) is present in several tissues includ-ing liver kidney and spleen but not skeletal muscle [7]Both carnosinase isoforms differ not only in distribution andmolecular weight but also in substrate specificity
Serum carnosinase deficiency (OMIM number 212200)results in accumulation of carnosine and related compoundshomocarnosine and anserine in plasma (20ndash30 120583molmLserum) urine [8] and tissues of affected patientsThe clinicalpresentation includes tremor myoclonic seizures hypotoniapsychomotor retardation and inability to social relationship[9 10] The coexistence of this rare enzymatic deficiency anda deletion in the long arm of chromosome 18 in a child wasreported suggesting the localization of serum carnosinasegene [11] Currently there are no reports of effective treatmentfor the disease According to Willi and colleagues [11] dietrestricted in meat ameliorates the clinical symptoms but itdoes not fully eliminate them Due to rarity of the disease themechanisms of tissue damage observed in affected subjectsare still to be unraveled
Such patients accumulate carnosine in tissues and bodyfluids and present a wide range of muscle alterations Inthis scenario the present work aimed to investigate theinfluence of acute and chronic carnosine administration onsome parameters of energy homeostasis in skeletal muscle ofyoung rats namely respiratory chain complexes (IndashIII II andII-III) citric acid cycle enzymes (succinate dehydrogenaseand malate dehydrogenase) and creatine kinase activities aswell as the expression of transcription factors (NRF-1 PGC-1120572 and TFAM) related to mitochondrial biogenesis
2 Materials and Methods
21 Animals Twenty-four male Wistar rats (250ndash300 g age30 days) obtained from the Central Animal House ofUniversidade do Extremo Sul Catarinense Santa CatarinaBrazil were caged in groups of six provided with ad libitumcommercial rat chow and water and maintained on a 12 hlight-dark cycle at a temperature of 23plusmn 1∘CThe animalswererandomly divided into four groups (119899 = 6) sham (saline)and carnosine (100mgkg of body weight) acute and chronicgroups All studies were performed in accordance with theNational Institutes of Health guidelines and EU Directive201063EU for animal experiments and with the approvalof the Ethics Committee of Universidade do Extremo SulCatarinense (Protocol number 672012)
22 Carnosine Administration Male Wistar rats weredivided into acute and chronic treatment groups In
acute administration the animals received a single doseof carnosine (100mgkg of body weight) administeredintraperitoneally Twenty-four hours after administrationthe rats were euthanized In the chronic administration theanimals received a daily dose of the dipeptide (100mgkgof body weight) administered intraperitoneally for fivedays and one hour after the last injection the rats wereeuthanized by decapitation without anesthesia and theskeletal muscle was removed for subsequent biochemicalanalysis The animals of control group were subjected tosimilar experimental conditions as carnosine group butreceived vehicle (NaCl 09) instead of carnosine
23 Complexes I-CoQ-III Activities NADH oxidoreductasecytochrome c (complexes IndashIII) activity was determinedaccording to Schapira and colleagues [14] by the determi-nation of cytochrome 119888 reduction at 120582 = 550 nm Results areexpressed as nmolsdotminminus1sdotmg proteinminus1
24 Complex II Activity Succinate 26-dichloroindophenol-oxidoreductase (complex II) activity was evaluated using themethod described by Fischer et al [15] Complex II activitywas measured by following the decrease in absorbance dueto the reduction of 26-dichloroindophenol at 120582 = 600 nmResults are expressed as nmolsdotminminus1sdotmg proteinminus1
25 Complexes II-III Activities Succinate cytochrome 119888oxidoreductase (complexes II-III) activity was determinedaccording to Fischer et al [15] Complexes II-III activitieswere measured by cytochrome 119888 reduction using succi-nate as substrate at 120582 = 550 nm Results are expressed asnmolsdotminminus1sdotmg proteinminus1
26 Creatine Kinase Activity Creatine kinase (EC 2732)activity wasmeasured in total homogenates using themethoddescribed by Hughes [16] with slight modifications [17]Results are expressed as 120583mol creatinesdotminminus1sdotmg proteinminus1
27 Succinate Dehydrogenase Activity Succinate dehydroge-nase (EC 13991) activity was determined in homogenatesaccording to Fischer et al [15] Results are expressed asnmolsdotminminus1sdotmg proteinminus1
28 Malate Dehydrogenase Activity Malate dehydrogenase(EC 11137) activity was evaluated according to Kitto [18]by following the reduction of NADH at wavelengths ofexcitation and emission of 340 and 466 nm respectivelyResults were calculated as nmolsdotminminus1sdotmg proteinminus1
29 Expression of Mitochondrial-Specific Transcription Fac-tors Transcript abundance of key factors involved in mito-chondrial biogenesis was evaluated by quantitative real-timeRT-PCR (RT-qPCR) Total RNA was isolated with Trizolreagent (Invitrogen Carlsbad CA USA) in accordance withthe manufacturerrsquos instructions The total RNA was quanti-fied by spectrophotometry (A260280 nm) and after treatedwith deoxyribonuclease I (Invitrogen) to eliminate genomic
BioMed Research International 3
0
200
400
600
800
Control Carnosine100 mgkg
(nm
olmiddot
minus1
min
middotmg
prot
einminus
1 )
lowast
(a)
0
5
10
15
20
Control Carnosine100 mgkg
(nm
olmiddot
minus1
min
middotmg
prot
einminus
1 )
lowastlowast
(b)
00
25
50
75
100
Control Carnosine100 mgkg
(nm
olmiddot
minus1
min
middotmg
prot
einminus
1 )
(c)
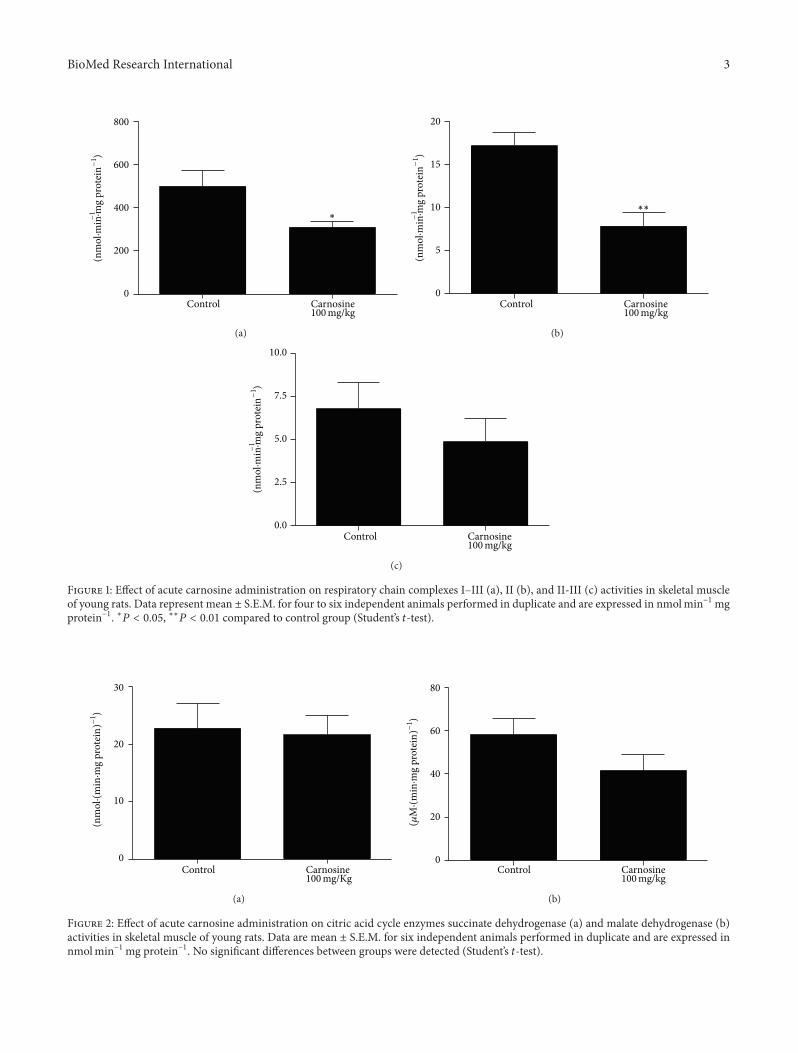
Figure 1 Effect of acute carnosine administration on respiratory chain complexes IndashIII (a) II (b) and II-III (c) activities in skeletal muscleof young rats Data represent mean plusmn SEM for four to six independent animals performed in duplicate and are expressed in nmolminminus1mgproteinminus1 lowast119875 lt 005 lowastlowast119875 lt 001 compared to control group (Studentrsquos 119905-test)
0
10
20
30
Control Carnosine100 mgKg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(a)
0
20
40
60
80
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
(b)
Figure 2 Effect of acute carnosine administration on citric acid cycle enzymes succinate dehydrogenase (a) and malate dehydrogenase (b)activities in skeletal muscle of young rats Data are mean plusmn SEM for six independent animals performed in duplicate and are expressed innmolminminus1mg proteinminus1 No significant differences between groups were detected (Studentrsquos 119905-test)
4 BioMed Research International
Table 1 Primer sequences for RT-qPCR experiments included in the study
Gene Forward primer Reverse primerGapda 51015840-GCTAAGCAGTTGGTGGTGCA-31015840 51015840-TCACCACCATGGAGAAGGC-31015840
Hprt1a 51015840-GCAGACTTTGCTTTCCTTGG-31015840 51015840-GCAGACTTTGCTTTCCTTGG-31015840
NRF-1b 51015840-TTACTCTGCTGTGGCTGATGG-31015840 51015840-CCTCTGATGCTTGCGTCGTCT-31015840
PGC-1120572c 51015840-CGTTACACCTGTGACGCTTTCGCTG-31015840 51015840-CATACTTGCTCTTGGTGGAAGCAGG-31015840
TFAMc 51015840-AATTGAAGCTTGTAAATCAGGCTTGG-31015840 51015840-CGGATGAGATCACTTCGCCCAAC-31015840
According to aBonefeld et al 2008 [12] bZhang et al 2012 [13] cdesigned by authors
0
1
2
3
4
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
Figure 3 Effect of acute carnosine administration on creatinekinase activity in skeletal muscle of young rats Data are mean plusmnSEM for six independent animals performed in duplicate andare expressed in 120583mol creatine minminus1mg proteinminus1 No significantdifference between groups was observed (Studentrsquos 119905-test)
DNA contamination in accordance with the manufacturerrsquosinstructions The cDNA was synthesized with ImProm-IIReverse Transcription System (Promega) from 1120583g totalRNA following the manufacturerrsquos instruction QuantitativePCR was performed using SYBR Green I (Invitrogen) todetect double-strand cDNA synthesis Reactions were donein a volume of 25120583L using 125 120583L of diluted cDNA contain-ing a final concentration of 02x SYBR Green I (Invitrogen)100 120583M dNTP 1x PCR Buffer 3mMMgCl
2 025U Platinum
Taq DNA Polymerase (Invitrogen) 05M of betaine (forGapd [12] Hprt1 [12] PGC-1120572 andTFAM) 2 of the reactionof DMSO (for NRF-1 [13]) and 200 nM of each of reverseand forward primers (Table 1) The PCR cycling conditionswere an initial polymerase activation step for 5min at 95∘C40 cycles of 15 s at 95∘C for denaturation 35 s at 60∘Cfor annealing and 15 s at 72∘C for elongation At the endof cycling protocol a melting-curve analysis was includedand fluorescence measured from 60 to 99∘C and showedin all cases one single peak Gapd and Hprt1 were used asreference genes for normalization Relative expression levelswere determined with 7500 and 7500 Fast Real-Time PCRSystems Software v206 (Applied Biosystems)The efficiencyper sample was calculated using LinRegPCR 20123 Software(httpLinRegPCRnl) Relative mRNA expression levelswere determined using the 2minusΔΔCT method
210 Protein Determination Protein was measured by Lowryand colleagues [19] method using bovine serum albumin asstandard
211 Statistical Analysis Results are presented as mean plusmnstandard error of mean (SEM) Assays were performed induplicate and the mean or median was used for statisticalanalysis Data was analyzed using Studentrsquos t-test Differencesbetween groups were rated significant at P le 005 Allanalyses were carried out in an IBM-compatible PC computerusing the Statistica version 7 software Molecular data wereexpressed as means plusmn SEM and analyzed by Studentrsquos t-test for unpaired samples considering 119875 lt 005 as statisticalsignificance
3 Results
31 Energy Metabolism Is Impaired by Acute CarnosineAdministration in Skeletal Muscle of Young Rats Initiallywe tested the influence of acute carnosine administrationIn such condition we observed a statistically significantdecrease of complexes IndashIII [119905
(10)= 241 119875 lt 005]
(Figure 1(a)) and II [t(8)
= 386 119875 lt 001] (Figure 1(b))activities in treated animals as compared to control grouprats Furthermore the complexes II-III [119905
(10)= 0918 119875 gt
005] (Figure 1(c)) succinate dehydrogenase [119905(10)
= 0211119875 gt005] (Figure 2(a)) malate dehydrogenase [119905
(10)= 156 119875 gt
005] (Figure 2(b)) and creatine kinase [119905(10)
= 105119875 gt 005](Figure 3) activities were not significantly different betweengroups However we identified a nonsignificant decreasein complexes II-III and malate dehydrogenase activities inanimals that received carnosine as compared to control rats(Figures 1(c) and 2(b) resp)
32 Chronic Carnosine Administration Does Not Affect EnergyMetabolism in Rat Skeletal Muscle The biochemical param-eters following chronic carnosine administration were alsoevaluated We detected no statistically significant differencein complexes IndashIII [t
(9)= minus0169 119875 gt 005] (Figure 4(a)) II
[t(9)
= minus0383 119875 gt 005] (Figure 4(b)) II-III [t(9)
= minus0289119875 gt 005] (Figure 4(c)) succinate dehydrogenase [119905
(10)=
0274 119875 gt 005] (Figure 5(a)) malate dehydrogenase [119905(10)
= 0374 119875 gt 005] (Figure 5(b)) and creatine kinase [119905(10)
=minus0113 119875 gt 005] (Figure 6) activities between groups in ratskeletal muscle However it was observed that there is a trend
BioMed Research International 5
0
100
200
300
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(a)
0
5
10
15
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(b)
0
5
10
15
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(c)
Figure 4 Effect of chronic carnosine administration on respiratory chain complexes IndashIII (a) II (b) and II-III (c) activities in skeletal muscleof young rats Data are mean plusmn SEM for five to six independent animals performed in duplicate and are expressed in nmolminminus1mgproteinminus1 No significant difference between groups was detected (Studentrsquos 119905-test)
0
5
10
15
20
25
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(a)
0
10
20
30
40
50
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
(b)
Figure 5 Effect of chronic carnosine administration on citric acid cycle enzymes succinate dehydrogenase (a) and malate dehydrogenase (b)activities in skeletal muscle of young rats Data are mean plusmn SEM for six independent animals performed in duplicate and are expressed innmolminminus1mg proteinminus1 No significant difference between groups was observed (Studentrsquos 119905-test)
6 BioMed Research International
00
05
10
15
20
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
Figure 6 Effect of chronic carnosine administration on creatinekinase activity in skeletal muscle of young rats Data are mean plusmnSEM for six independent animals performed in duplicate andare expressed in 120583mol creatine minminus1mg proteinminus1 No significantdifference between groups was detected (Studentrsquos 119905-test)
of decrease of complexes IndashIII activities in animals receivingcarnosine as compared to control rats (Figure 4(a))
33 Mitochondrial Biogenesis Transcript abundance of keyfactors involved in mitochondrial biogenesis was evaluatedbyRT-qPCR in skeletalmuscle of animals receiving carnosineacutely (Figure 7) or chronically (Figure 8) As compared tocontrol groups mRNA levels of NRF-1 PGC-1120572 and TFAMin the animals receiving carnosine under either regimen ofadministration were unchanged
4 Discussion
In skeletal muscle tissue of most vertebrates and some inver-tebrates the imidazole dipeptide carnosine is synthesizedfrom histidine and 120573-alanine by the ATP-dependent enzymecarnosine synthetase [20] The biological roles of carnosineand related dipeptides in skeletal muscle remain uncertainalthough it is possible that they act as proton buffering agents[4] Furthermore there are a growing number of evidencesrelated to carnosine effects on metabolism Aside from itsantioxidant activity [21 22] carbonyl-scavenger ability hasbeen assigned to carnosine [23] and other imidazole dipep-tides [24] Many of carbonyl compounds are metabolic toxicby-products and in the bulk of tissues they are detoxifiedby oxidoreductases These enzymes catalyze the oxidation orreduction of aldehydes or its conjugation with lowmolecularweight amines and thiols such as reduced glutathione andimidazole dipeptides [25] Carnosine was also shown toreact with methylglyoxal a toxic metabolic by-product ofglycolysis as well as to inhibit cancer and aging mechanisms[26] and the cellular respiration regulatory complex mTOR[27] and activate the gluconeogenic enzyme fructose-16-bisphosphatase [28]
It was also shown that carnosine intake significantlydiminishes the activity and mRNA expression of malic
enzyme fatty acid synthase 3-hydroxy-3-methylglutarylcoenzyme A (HMG-CoA) reductase sterol regulatoryelement-binding proteins (SREBP-1c) and SREBP-2 inmice that consumed high saturated fat diet [29] Carnosinewas able to inhibit Zn2+-induced death of GT1-7 neuronsthrough the inhibition of GADD34 p8 (endoplasmicreticulum stress-related genes) and Arc (calcium-relatedgene) expression [30] Finally Asperger and colleagues [31]through a proteomics study with glioblastoma cells receivingcarnosine detected 31 proteins expressed differentiallyunder the influence of dipeptide including BCL2-associatedathanogene 2 and von Hippel-Lindau binding protein 1
In the present study we observed a statistically sig-nificant decrease of complexes IndashIII and II activities inanimals receiving carnosine acutely compared to controlgroup rats Additionally we detected no significant differencein complexes II-III citric acid cycle enzymes and creatinekinase activities between groups Finally we were also notable to find any significant alteration of respiratory chaincomplexes citric acid cycle enzymes and creatine kinaseactivities Since several evidences suggest that many of thephysiological effects of carnosine administration initiate attranscriptional level as shown by changes in mRNA levels ofHIF-1120572 [32] runt-related transcription factor-2core bindingfactor alpha-1 (RUNX2Cbf1205721) Sox9 [33] Hsp70 and SOD2[34] mitochondrial biogenesis could explain these findingsNevertheless it was found that mRNA levels of NRF-1 PGC-1120572 and TFAM in skeletal muscle of rats receiving carnosineacutely or chronically were similar to those measured incontrol animals
Our finding showing that the malate dehydrogenaseactivity presented no statistically significant differencebetween groups in both treatments corroborates at least inpart a literature report It was found that a patient with serumcarnosinase deficiency exhibited malate dehydrogenase andlactate dehydrogenase activities within normal limits in liverkidney and spleen specimens despite the fact that patientswith this disorder present high levels of carnosine in theirplasma (carnosinemia) [8]
We also observed that succinate dehydrogenase activityin skeletal muscle of carnosine group was not significantlydifferent from those in control group in animals subjectedto either acute or chronic carnosine administration On theother hand a literature report showed that rats chronicallyintoxicated with ethanol presented reduced activity of somehepatic enzymes including succinate dehydrogenase [35] Itwas also showed in the same study that carnosine load for twoweeks previously or simultaneously with ethanol intoxicationprevents or reverses the toxicity of such alcohol on thisenzyme activity
It should be emphasized that in skeletal muscle of ratsreceiving carnosine acutely a statistically significant decreasein complexes IndashIII and II activities was identified as well asa trend of decrease in complexes II-III activities relatively tocontrol group However rats receiving carnosine chronicallyshowed only a trend of decrease in complexes IndashIII activitiesin skeletal muscle as compared to control group whereas theother complexes activities were not different between groups
BioMed Research International 7
00
05
10
15Re
lativ
e RN
A ex
pres
sion
Control Carnosine100 mgkg
(a)
Relat
ive R
NA
expr
essio
n
00
05
10
15
Control Carnosine100 mgkg
(b)
00
05
10
15
Relat
ive R
NA
expr
essio
n
Control Carnosine100 mgkg
(c)
Figure 7 Effect of acute carnosine administration on relative mRNA expression ofNRF-1 (a) PGC-1120572 (b) and TFAM (c) activities in skeletalmuscle of young rats Results are mean plusmn SEM for four to six independent animals performed in quadruplicate No significant differencebetween groups was detected
The same pattern of alteration was also observed inmalate dehydrogenase activity which was found increasedin rats receiving carnosine acutely and unaltered in animalsreceiving the dipeptide chronically Thus it may be proposedthat skeletal muscle is able to adapt to high carnosinelevels recovering the impaired enzyme activities back tocontrol conditions A putative mechanism involved in suchadaptation is increased mitochondrial synthesis since theenzymes affected by carnosine administration present mito-chondrial localization Considering that carnosine adminis-tration increases vimentin synthesis [36] and that vimentin isa cytoskeleton protein that may be related to mitochondrialmotility and localization [37] it is possible that this processis mediated by increased vimentin synthesis However dataon NRF-1 PGC-1120572 and TFAM mRNA levels suggest thatcarnosine interferes with these enzymatic activities throughmechanisms unrelated to mitochondrial biogenesis but theinfluence of carnosine on mitophagy cannot be ruled out
At present we cannot ascertain the pathophysiologicalrelevance of our findings In this communication we inducedcarnosinemia in the animals by administering carnosineintraperitoneally With respect to this fact it should be men-tioned that carnosine dietary administration elicits increase
of its content in human skeletal muscle [38] The carnosinelevels in this tissue are situated within 30ndash50mmolkg ofwet weight [39] and are prone to alterations according todiet gender and age of subject [40] Additionally in ourstudy carnosine is increased in skeletal muscle of rat suchas occurrence in patients with carnosinemia
5 Conclusion
It is here shown that carnosine administration impairs elec-tron transfer through the mitochondrial respiratory chainin skeletal muscle of young rats without changing mito-chondrial biogenesis-related transcription factors At leastin part this finding does not corroborate some reportswhich indicate that this dipeptide increases the efficiencyand intensity of oxidative phosphorylation in rats adaptedto hypobaric hypoxia [41] The human carnosinase levelsare almost undetectable in plasma of neonates increasingaccording to aging [6] rendering young individuals lessprone to metabolize carnosine In case these findings areconfirmed by further studies and ATP depletion is alsoobserved it is possible that an energy dysfunction secondaryto carnosine accumulationmay aid to explain someofmuscle
8 BioMed Research International
00
05
10
15Re
lativ
e RN
A ex
pres
sion
Control Carnosine100 mgkg
(a)
00
05
10
15
Relat
ive R
NA
expr
essio
n
Control Carnosine100 mgkg
(b)
Control Carnosine100 mgkg
00
05
10
15
Relat
ive R
NA
expr
essio
n
(c)
Figure 8 Effect of chronic carnosine administration on relative mRNA expression of NRF-1 (a) PGC-1120572 (b) and TFAM (c) activities inskeletal muscle of young rats Results are mean plusmn SEM for four to six independent animals performed in quadruplicate No significantdifference between groups was detected
symptoms commonly observed in young patients with serumcarnosinase deficiency
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This research was supported by resources from Universidadedo Extremo Sul Catarinense and Brazilian National Councilfor Scientific and Technological Development (CNPq) LuizaWilges Kist was recipient of fellowships from Coordenacaode Aperfeicoamento de Pessoal de Nıvel Superior (CAPES)
References
[1] L Bonfanti P Peretto S deMarchis andA Fasolo ldquoCarnosine-related dipeptides in the mammalian brainrdquo Progress in Neuro-biology vol 59 no 4 pp 333ndash353 1999
[2] W Gulewitsch and S Amiradzibi ldquoUeber das carnosineine neue organische base des fleischextraktesrdquo Berichte der
Deutschen ChemischenGesellschaft vol 33 no 2 pp 1902ndash19031900
[3] K Nagai and T Suda ldquoRealization of spontaneous healingfunction by carnosinerdquo Methods and Findings in Experimentaland Clinical Pharmacology vol 10 no 8 pp 497ndash507 1988
[4] H Abe ldquoRole of histidine-related compounds as intracellularproton buffering constituents in vertebrate musclerdquo Biochem-istry vol 65 no 7 pp 757ndash765 2000
[5] S Geissler M Zwarg I Knutter F Markwardt and M Brand-sch ldquoThe bioactive dipeptide anserine is transported by humanproton-coupled peptide transportersrdquo FEBS Journal vol 277no 3 pp 790ndash795 2010
[6] J F Lenney R P George and A M Weiss ldquoHuman serumcarnosinase characterization distinction from cellular carnosi-nase and activation by cadmiumrdquo Clinica Chimica Acta vol123 no 3 pp 221ndash231 1982
[7] C R Scriver and K M Gibson ldquoDisorders of 120573- and 120574-aminoacids in free and peptide-linked formsrdquo in The Metabolic andMolecular Bases of Inherit C R Scriver A L Beaudet S Slyand D Valle Eds vol 1 part 5 pp 1349ndash1368 McGraw-HillNew York NY USA 7th edition 1995
[8] W H Murphey D G Lindmark and L I Patchen ldquoSerumcarnosinase deficiency concomitant with mental retardationrdquoPediatric Research vol 7 no 7 pp 601ndash606 1973
BioMed Research International 9
[9] H Lunde O Sjaastad and L Gjessing ldquoHomocarnosinosishypercarnosinuriardquo Journal of Neurochemistry vol 38 no 1 pp242ndash245 1982
[10] T L Perry S Hansen B Tischler R Bunting and K BerryldquoCarnosinemia A new metabolic disorder associated withneurologic disease andmental defectrdquoTheNewEngland Journalof Medicine vol 277 no 23 pp 1219ndash1227 1967
[11] S M Willi Y Zhang J B Hill M C Phelan R C Michaelisand K R Holden ldquoA deletion in the long arm of chromosome18 in a child with serum carnosinase deficiencyrdquo PediatricResearch vol 41 no 2 pp 210ndash213 1997
[12] B E Bonefeld B Elfving and GWegener ldquoReference genes fornormalization a study of rat brain tissuerdquo Synapse vol 62 no4 pp 302ndash309 2008
[13] Q Zhang YWu H Sha et al ldquoEarly exercise affects mitochon-drial transcription factors expression after cerebral ischemia inratsrdquo International Journal of Molecular Sciences vol 13 no 2pp 1670ndash1679 2012
[14] A H V Schapira V M Mann J M Cooper et al ldquoAnatomicand disease specificity of NADH CoQ1 reductase (complex I)deficiency in Parkinsonrsquos diseaserdquo Journal of Neurochemistryvol 55 no 6 pp 2142ndash2145 1990
[15] J C Fischer W Ruitenbeek and J A Berden ldquoDifferentialinvestigation of the capacity of succinate oxidation in humanskeletal musclerdquo Clinica Chimica Acta vol 153 no 1 pp 23ndash261985
[16] B P Hughes ldquoA method for the estimation of serum creatinekinase and its use in comparing creatine kinase and aldolaseactivity in normal and pathological serardquo Clinica Chimica Actavol 7 no 5 pp 597ndash603 1962
[17] P F Schuck G Leipnitz C A J Ribeiro et al ldquoInhibition ofcreatine kinase activity in vitro by ethylmalonic acid in cerebralcortex of young ratsrdquoNeurochemical Research vol 27 no 12 pp1633ndash1639 2002
[18] G B Kitto ldquoIntra- and extramitochondrial malate dehydroge-nases from chicken and tuna heartrdquo Methods in Enzymologyvol 13 pp 106ndash116 1969
[19] O H Lowry N J Rosebrough A L Farr and R J RandallldquoProtein measurement with the Folin phenol reagentrdquo TheJournal of Biological Chemistry vol 193 no 1 pp 265ndash275 1951
[20] J Drozak M Veiga-da-Cunha D Vertommen V Stroobantand E van Schaftingen ldquoMolecular identification of carno-sine synthase as ATP-grasp domain-containing protein 1(ATPGD1)rdquoThe Journal of Biological Chemistry vol 285 no 13pp 9346ndash9356 2010
[21] P J Quinn A A Boldyrev and V E Formazuyk ldquoCarnosineits properties functions and potential therapeutic applicationsrdquoMolecular Aspects of Medicine vol 13 no 5 pp 379ndash444 1992
[22] P S Manhiani J K Northcutt I Han W C Bridges and PL Dawson ldquoAntioxidant activity of carnosine extracted fromvarious poultry tissuesrdquo Poultry Science vol 92 no 2 pp 444ndash453 2013
[23] G Aldini M Orioli G Rossoni et al ldquoThe carbonyl scavengercarnosine ameliorates dyslipidaemia and renal function inZucker obese ratsrdquo Journal of Cellular and Molecular Medicinevol 15 no 6 pp 1339ndash1354 2011
[24] G Vistoli D deMaddis V Straniero et al ldquoExploring the spaceof histidine containing dipeptides in search of novel efficientRCS sequestering agentsrdquo European Journal of Medicinal Chem-istry vol 66 pp 153ndash160 2013
[25] Z Xie S P Baba B R Sweeney and O A Barski ldquoDetoxi-fication of aldehydes by histidine-containing dipeptides fromchemistry to clinical implicationsrdquo Chemico-Biological Interac-tions vol 202 no 1ndash3 pp 288ndash297 2013
[26] A R Hipkiss ldquoAging proteotoxicity mitochondria glycationNAD+ and carnosine possible inter-relationships and resolu-tion of the oxygen paradoxrdquoFrontiers inAgingNeuroscience vol2 article 10 2010
[27] A R Hipkiss ldquoEnergy metabolism proteotoxic stress andage-related dysfunction - Protection by carnosinerdquo MolecularAspects of Medicine vol 32 no 4ndash6 pp 267ndash278 2011
[28] A R Hipkiss S P Cartwright C Bromley S R Gross and RM Bill ldquoCarnosine can understanding its actions on energymetabolism and protein homeostasis inform its therapeuticpotentialrdquo Chemistry Central Journal vol 7 no 1 article 382013
[29] M-C Mong C-Y Chao and M-C Yin ldquoHistidine andcarnosine alleviated hepatic steatosis in mice consumed highsaturated fat dietrdquo European Journal of Pharmacology vol 653no 1ndash3 pp 82ndash88 2011
[30] D Mizuno and M Kawahara ldquoThe molecular mechanisms ofzinc neurotoxicity and the pathogenesis of vascular type seniledementiardquo International Journal of Molecular Sciences vol 14no 11 pp 22067ndash22081 2013
[31] A Asperger C Renner M Menzel R Gebhardt J Meixens-berger and F Gaunitz ldquoIdentification of factors involved inthe anti-tumor activity of carnosine on glioblastomas using aproteomics approachrdquo Cancer Investigation vol 29 no 4 pp272ndash281 2011
[32] L A Bharadwaj G F Davies I J Xavier and N Ovsenek ldquoL-carnosine and verapamil inhibit hypoxia-induced expression ofhypoxia inducible factor (HIF-1120572) in H9c2 cardiomyoblastsrdquoPharmacological Research vol 45 no 3 pp 175ndash181 2002
[33] E Ito-Kato N Suzuki M Maeno et al ldquoEffect of carnosine onrunt-related transcription factor-2core binding factor 120572-1 andSox9 expressions of human periodontal ligament cellsrdquo Journalof Periodontal Research vol 39 no 3 pp 199ndash204 2004
[34] V Calabrese C Colombrita E Guagliano et al ldquoProtectiveeffect of carnosine during nitrosative stress in astroglial cellculturesrdquo Neurochemical Research vol 30 no 6-7 pp 797ndash8072005
[35] K M Soliman M A Hamed and S A Ali ldquoHepatoprotectiveeffect of carnosine on liver biochemical parameters in chronicethanol intoxicated ratrdquoTheMedical Journal of the IslamicWorldAcademy of Sciences vol 16 no 2 pp 77ndash86 2006
[36] D Ikeda S-I Wada C Yoneda H Abe and S WatabeldquoCarnosine stimulates vimentin expression in cultured ratfibroblastsrdquoCell Structure and Function vol 24 no 2 pp 79ndash871999
[37] O E Nekrasova M G Mendez I S Chernoivanenko et alldquoVimentin intermediate filaments modulate the motility ofmitochondriardquoMolecular Biology of the Cell vol 22 no 13 pp2282ndash2289 2011
[38] C Sale B Saunders and R C Harris ldquoEffect of beta-alaninesupplementation onmuscle carnosine concentrations and exer-cise performancerdquoAmino Acids vol 39 no 2 pp 321ndash333 2010
[39] A A Boldyrev and S E Severin ldquoThe histidine-containingdipeptides carnosine and anserine distribution properties andbiological significancerdquoAdvances in Enzyme Regulation vol 30pp 175ndash188 1990
[40] I Everaert A Mooyaart A Baguet et al ldquoVegetarianismfemale gender and increasing age but not CNDP1 genotype
10 BioMed Research International
are associatedwith reducedmuscle carnosine levels in humansrdquoAmino Acids vol 40 no 4 pp 1221ndash1229 2011
[41] V N Korobov N M Doliba and I V Telegus ldquoCarnosine inadaptation to hypobaric hypoxiardquo Biokhimiya vol 58 no 5 pp740ndash744 1993

2 BioMed Research International
is uptaken by intestinal proton-coupled peptide transporternamely human H+peptide cotransporter 1 (hPEPT1) [5]This dipeptide is synthesized starting from its componentamino acids histidine and 120573-alanine by enzyme carnosinesynthetase (EC 63211) On the other hand the clearanceof carnosine is essentially due to the hydrolytic action of ametalloprotease the carnosinase [6] This enzyme exists intwo isoforms The secreted form namely serum carnosinase(EC 341320) is found primarily in serum as well as inbrain and cerebrospinal fluid The cytosolic form tissuecarnosinase (EC 34133) is present in several tissues includ-ing liver kidney and spleen but not skeletal muscle [7]Both carnosinase isoforms differ not only in distribution andmolecular weight but also in substrate specificity
Serum carnosinase deficiency (OMIM number 212200)results in accumulation of carnosine and related compoundshomocarnosine and anserine in plasma (20ndash30 120583molmLserum) urine [8] and tissues of affected patientsThe clinicalpresentation includes tremor myoclonic seizures hypotoniapsychomotor retardation and inability to social relationship[9 10] The coexistence of this rare enzymatic deficiency anda deletion in the long arm of chromosome 18 in a child wasreported suggesting the localization of serum carnosinasegene [11] Currently there are no reports of effective treatmentfor the disease According to Willi and colleagues [11] dietrestricted in meat ameliorates the clinical symptoms but itdoes not fully eliminate them Due to rarity of the disease themechanisms of tissue damage observed in affected subjectsare still to be unraveled
Such patients accumulate carnosine in tissues and bodyfluids and present a wide range of muscle alterations Inthis scenario the present work aimed to investigate theinfluence of acute and chronic carnosine administration onsome parameters of energy homeostasis in skeletal muscle ofyoung rats namely respiratory chain complexes (IndashIII II andII-III) citric acid cycle enzymes (succinate dehydrogenaseand malate dehydrogenase) and creatine kinase activities aswell as the expression of transcription factors (NRF-1 PGC-1120572 and TFAM) related to mitochondrial biogenesis
2 Materials and Methods
21 Animals Twenty-four male Wistar rats (250ndash300 g age30 days) obtained from the Central Animal House ofUniversidade do Extremo Sul Catarinense Santa CatarinaBrazil were caged in groups of six provided with ad libitumcommercial rat chow and water and maintained on a 12 hlight-dark cycle at a temperature of 23plusmn 1∘CThe animalswererandomly divided into four groups (119899 = 6) sham (saline)and carnosine (100mgkg of body weight) acute and chronicgroups All studies were performed in accordance with theNational Institutes of Health guidelines and EU Directive201063EU for animal experiments and with the approvalof the Ethics Committee of Universidade do Extremo SulCatarinense (Protocol number 672012)
22 Carnosine Administration Male Wistar rats weredivided into acute and chronic treatment groups In
acute administration the animals received a single doseof carnosine (100mgkg of body weight) administeredintraperitoneally Twenty-four hours after administrationthe rats were euthanized In the chronic administration theanimals received a daily dose of the dipeptide (100mgkgof body weight) administered intraperitoneally for fivedays and one hour after the last injection the rats wereeuthanized by decapitation without anesthesia and theskeletal muscle was removed for subsequent biochemicalanalysis The animals of control group were subjected tosimilar experimental conditions as carnosine group butreceived vehicle (NaCl 09) instead of carnosine
23 Complexes I-CoQ-III Activities NADH oxidoreductasecytochrome c (complexes IndashIII) activity was determinedaccording to Schapira and colleagues [14] by the determi-nation of cytochrome 119888 reduction at 120582 = 550 nm Results areexpressed as nmolsdotminminus1sdotmg proteinminus1
24 Complex II Activity Succinate 26-dichloroindophenol-oxidoreductase (complex II) activity was evaluated using themethod described by Fischer et al [15] Complex II activitywas measured by following the decrease in absorbance dueto the reduction of 26-dichloroindophenol at 120582 = 600 nmResults are expressed as nmolsdotminminus1sdotmg proteinminus1
25 Complexes II-III Activities Succinate cytochrome 119888oxidoreductase (complexes II-III) activity was determinedaccording to Fischer et al [15] Complexes II-III activitieswere measured by cytochrome 119888 reduction using succi-nate as substrate at 120582 = 550 nm Results are expressed asnmolsdotminminus1sdotmg proteinminus1
26 Creatine Kinase Activity Creatine kinase (EC 2732)activity wasmeasured in total homogenates using themethoddescribed by Hughes [16] with slight modifications [17]Results are expressed as 120583mol creatinesdotminminus1sdotmg proteinminus1
27 Succinate Dehydrogenase Activity Succinate dehydroge-nase (EC 13991) activity was determined in homogenatesaccording to Fischer et al [15] Results are expressed asnmolsdotminminus1sdotmg proteinminus1
28 Malate Dehydrogenase Activity Malate dehydrogenase(EC 11137) activity was evaluated according to Kitto [18]by following the reduction of NADH at wavelengths ofexcitation and emission of 340 and 466 nm respectivelyResults were calculated as nmolsdotminminus1sdotmg proteinminus1
29 Expression of Mitochondrial-Specific Transcription Fac-tors Transcript abundance of key factors involved in mito-chondrial biogenesis was evaluated by quantitative real-timeRT-PCR (RT-qPCR) Total RNA was isolated with Trizolreagent (Invitrogen Carlsbad CA USA) in accordance withthe manufacturerrsquos instructions The total RNA was quanti-fied by spectrophotometry (A260280 nm) and after treatedwith deoxyribonuclease I (Invitrogen) to eliminate genomic
BioMed Research International 3
0
200
400
600
800
Control Carnosine100 mgkg
(nm
olmiddot
minus1
min
middotmg
prot
einminus
1 )
lowast
(a)
0
5
10
15
20
Control Carnosine100 mgkg
(nm
olmiddot
minus1
min
middotmg
prot
einminus
1 )
lowastlowast
(b)
00
25
50
75
100
Control Carnosine100 mgkg
(nm
olmiddot
minus1
min
middotmg
prot
einminus
1 )
(c)
Figure 1 Effect of acute carnosine administration on respiratory chain complexes IndashIII (a) II (b) and II-III (c) activities in skeletal muscleof young rats Data represent mean plusmn SEM for four to six independent animals performed in duplicate and are expressed in nmolminminus1mgproteinminus1 lowast119875 lt 005 lowastlowast119875 lt 001 compared to control group (Studentrsquos 119905-test)
0
10
20
30
Control Carnosine100 mgKg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(a)
0
20
40
60
80
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
(b)
Figure 2 Effect of acute carnosine administration on citric acid cycle enzymes succinate dehydrogenase (a) and malate dehydrogenase (b)activities in skeletal muscle of young rats Data are mean plusmn SEM for six independent animals performed in duplicate and are expressed innmolminminus1mg proteinminus1 No significant differences between groups were detected (Studentrsquos 119905-test)
4 BioMed Research International
Table 1 Primer sequences for RT-qPCR experiments included in the study
Gene Forward primer Reverse primerGapda 51015840-GCTAAGCAGTTGGTGGTGCA-31015840 51015840-TCACCACCATGGAGAAGGC-31015840
Hprt1a 51015840-GCAGACTTTGCTTTCCTTGG-31015840 51015840-GCAGACTTTGCTTTCCTTGG-31015840
NRF-1b 51015840-TTACTCTGCTGTGGCTGATGG-31015840 51015840-CCTCTGATGCTTGCGTCGTCT-31015840
PGC-1120572c 51015840-CGTTACACCTGTGACGCTTTCGCTG-31015840 51015840-CATACTTGCTCTTGGTGGAAGCAGG-31015840
TFAMc 51015840-AATTGAAGCTTGTAAATCAGGCTTGG-31015840 51015840-CGGATGAGATCACTTCGCCCAAC-31015840
According to aBonefeld et al 2008 [12] bZhang et al 2012 [13] cdesigned by authors
0
1
2
3
4
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
Figure 3 Effect of acute carnosine administration on creatinekinase activity in skeletal muscle of young rats Data are mean plusmnSEM for six independent animals performed in duplicate andare expressed in 120583mol creatine minminus1mg proteinminus1 No significantdifference between groups was observed (Studentrsquos 119905-test)
DNA contamination in accordance with the manufacturerrsquosinstructions The cDNA was synthesized with ImProm-IIReverse Transcription System (Promega) from 1120583g totalRNA following the manufacturerrsquos instruction QuantitativePCR was performed using SYBR Green I (Invitrogen) todetect double-strand cDNA synthesis Reactions were donein a volume of 25120583L using 125 120583L of diluted cDNA contain-ing a final concentration of 02x SYBR Green I (Invitrogen)100 120583M dNTP 1x PCR Buffer 3mMMgCl
2 025U Platinum
Taq DNA Polymerase (Invitrogen) 05M of betaine (forGapd [12] Hprt1 [12] PGC-1120572 andTFAM) 2 of the reactionof DMSO (for NRF-1 [13]) and 200 nM of each of reverseand forward primers (Table 1) The PCR cycling conditionswere an initial polymerase activation step for 5min at 95∘C40 cycles of 15 s at 95∘C for denaturation 35 s at 60∘Cfor annealing and 15 s at 72∘C for elongation At the endof cycling protocol a melting-curve analysis was includedand fluorescence measured from 60 to 99∘C and showedin all cases one single peak Gapd and Hprt1 were used asreference genes for normalization Relative expression levelswere determined with 7500 and 7500 Fast Real-Time PCRSystems Software v206 (Applied Biosystems)The efficiencyper sample was calculated using LinRegPCR 20123 Software(httpLinRegPCRnl) Relative mRNA expression levelswere determined using the 2minusΔΔCT method
210 Protein Determination Protein was measured by Lowryand colleagues [19] method using bovine serum albumin asstandard
211 Statistical Analysis Results are presented as mean plusmnstandard error of mean (SEM) Assays were performed induplicate and the mean or median was used for statisticalanalysis Data was analyzed using Studentrsquos t-test Differencesbetween groups were rated significant at P le 005 Allanalyses were carried out in an IBM-compatible PC computerusing the Statistica version 7 software Molecular data wereexpressed as means plusmn SEM and analyzed by Studentrsquos t-test for unpaired samples considering 119875 lt 005 as statisticalsignificance
3 Results
31 Energy Metabolism Is Impaired by Acute CarnosineAdministration in Skeletal Muscle of Young Rats Initiallywe tested the influence of acute carnosine administrationIn such condition we observed a statistically significantdecrease of complexes IndashIII [119905
(10)= 241 119875 lt 005]
(Figure 1(a)) and II [t(8)
= 386 119875 lt 001] (Figure 1(b))activities in treated animals as compared to control grouprats Furthermore the complexes II-III [119905
(10)= 0918 119875 gt
005] (Figure 1(c)) succinate dehydrogenase [119905(10)
= 0211119875 gt005] (Figure 2(a)) malate dehydrogenase [119905
(10)= 156 119875 gt
005] (Figure 2(b)) and creatine kinase [119905(10)
= 105119875 gt 005](Figure 3) activities were not significantly different betweengroups However we identified a nonsignificant decreasein complexes II-III and malate dehydrogenase activities inanimals that received carnosine as compared to control rats(Figures 1(c) and 2(b) resp)
32 Chronic Carnosine Administration Does Not Affect EnergyMetabolism in Rat Skeletal Muscle The biochemical param-eters following chronic carnosine administration were alsoevaluated We detected no statistically significant differencein complexes IndashIII [t
(9)= minus0169 119875 gt 005] (Figure 4(a)) II
[t(9)
= minus0383 119875 gt 005] (Figure 4(b)) II-III [t(9)
= minus0289119875 gt 005] (Figure 4(c)) succinate dehydrogenase [119905
(10)=
0274 119875 gt 005] (Figure 5(a)) malate dehydrogenase [119905(10)
= 0374 119875 gt 005] (Figure 5(b)) and creatine kinase [119905(10)
=minus0113 119875 gt 005] (Figure 6) activities between groups in ratskeletal muscle However it was observed that there is a trend
BioMed Research International 5
0
100
200
300
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(a)
0
5
10
15
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(b)
0
5
10
15
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(c)
Figure 4 Effect of chronic carnosine administration on respiratory chain complexes IndashIII (a) II (b) and II-III (c) activities in skeletal muscleof young rats Data are mean plusmn SEM for five to six independent animals performed in duplicate and are expressed in nmolminminus1mgproteinminus1 No significant difference between groups was detected (Studentrsquos 119905-test)
0
5
10
15
20
25
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(a)
0
10
20
30
40
50
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
(b)
Figure 5 Effect of chronic carnosine administration on citric acid cycle enzymes succinate dehydrogenase (a) and malate dehydrogenase (b)activities in skeletal muscle of young rats Data are mean plusmn SEM for six independent animals performed in duplicate and are expressed innmolminminus1mg proteinminus1 No significant difference between groups was observed (Studentrsquos 119905-test)
6 BioMed Research International
00
05
10
15
20
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
Figure 6 Effect of chronic carnosine administration on creatinekinase activity in skeletal muscle of young rats Data are mean plusmnSEM for six independent animals performed in duplicate andare expressed in 120583mol creatine minminus1mg proteinminus1 No significantdifference between groups was detected (Studentrsquos 119905-test)
of decrease of complexes IndashIII activities in animals receivingcarnosine as compared to control rats (Figure 4(a))
33 Mitochondrial Biogenesis Transcript abundance of keyfactors involved in mitochondrial biogenesis was evaluatedbyRT-qPCR in skeletalmuscle of animals receiving carnosineacutely (Figure 7) or chronically (Figure 8) As compared tocontrol groups mRNA levels of NRF-1 PGC-1120572 and TFAMin the animals receiving carnosine under either regimen ofadministration were unchanged
4 Discussion
In skeletal muscle tissue of most vertebrates and some inver-tebrates the imidazole dipeptide carnosine is synthesizedfrom histidine and 120573-alanine by the ATP-dependent enzymecarnosine synthetase [20] The biological roles of carnosineand related dipeptides in skeletal muscle remain uncertainalthough it is possible that they act as proton buffering agents[4] Furthermore there are a growing number of evidencesrelated to carnosine effects on metabolism Aside from itsantioxidant activity [21 22] carbonyl-scavenger ability hasbeen assigned to carnosine [23] and other imidazole dipep-tides [24] Many of carbonyl compounds are metabolic toxicby-products and in the bulk of tissues they are detoxifiedby oxidoreductases These enzymes catalyze the oxidation orreduction of aldehydes or its conjugation with lowmolecularweight amines and thiols such as reduced glutathione andimidazole dipeptides [25] Carnosine was also shown toreact with methylglyoxal a toxic metabolic by-product ofglycolysis as well as to inhibit cancer and aging mechanisms[26] and the cellular respiration regulatory complex mTOR[27] and activate the gluconeogenic enzyme fructose-16-bisphosphatase [28]
It was also shown that carnosine intake significantlydiminishes the activity and mRNA expression of malic
enzyme fatty acid synthase 3-hydroxy-3-methylglutarylcoenzyme A (HMG-CoA) reductase sterol regulatoryelement-binding proteins (SREBP-1c) and SREBP-2 inmice that consumed high saturated fat diet [29] Carnosinewas able to inhibit Zn2+-induced death of GT1-7 neuronsthrough the inhibition of GADD34 p8 (endoplasmicreticulum stress-related genes) and Arc (calcium-relatedgene) expression [30] Finally Asperger and colleagues [31]through a proteomics study with glioblastoma cells receivingcarnosine detected 31 proteins expressed differentiallyunder the influence of dipeptide including BCL2-associatedathanogene 2 and von Hippel-Lindau binding protein 1
In the present study we observed a statistically sig-nificant decrease of complexes IndashIII and II activities inanimals receiving carnosine acutely compared to controlgroup rats Additionally we detected no significant differencein complexes II-III citric acid cycle enzymes and creatinekinase activities between groups Finally we were also notable to find any significant alteration of respiratory chaincomplexes citric acid cycle enzymes and creatine kinaseactivities Since several evidences suggest that many of thephysiological effects of carnosine administration initiate attranscriptional level as shown by changes in mRNA levels ofHIF-1120572 [32] runt-related transcription factor-2core bindingfactor alpha-1 (RUNX2Cbf1205721) Sox9 [33] Hsp70 and SOD2[34] mitochondrial biogenesis could explain these findingsNevertheless it was found that mRNA levels of NRF-1 PGC-1120572 and TFAM in skeletal muscle of rats receiving carnosineacutely or chronically were similar to those measured incontrol animals
Our finding showing that the malate dehydrogenaseactivity presented no statistically significant differencebetween groups in both treatments corroborates at least inpart a literature report It was found that a patient with serumcarnosinase deficiency exhibited malate dehydrogenase andlactate dehydrogenase activities within normal limits in liverkidney and spleen specimens despite the fact that patientswith this disorder present high levels of carnosine in theirplasma (carnosinemia) [8]
We also observed that succinate dehydrogenase activityin skeletal muscle of carnosine group was not significantlydifferent from those in control group in animals subjectedto either acute or chronic carnosine administration On theother hand a literature report showed that rats chronicallyintoxicated with ethanol presented reduced activity of somehepatic enzymes including succinate dehydrogenase [35] Itwas also showed in the same study that carnosine load for twoweeks previously or simultaneously with ethanol intoxicationprevents or reverses the toxicity of such alcohol on thisenzyme activity
It should be emphasized that in skeletal muscle of ratsreceiving carnosine acutely a statistically significant decreasein complexes IndashIII and II activities was identified as well asa trend of decrease in complexes II-III activities relatively tocontrol group However rats receiving carnosine chronicallyshowed only a trend of decrease in complexes IndashIII activitiesin skeletal muscle as compared to control group whereas theother complexes activities were not different between groups
BioMed Research International 7
00
05
10
15Re
lativ
e RN
A ex
pres
sion
Control Carnosine100 mgkg
(a)
Relat
ive R
NA
expr
essio
n
00
05
10
15
Control Carnosine100 mgkg
(b)
00
05
10
15
Relat
ive R
NA
expr
essio
n
Control Carnosine100 mgkg
(c)
Figure 7 Effect of acute carnosine administration on relative mRNA expression ofNRF-1 (a) PGC-1120572 (b) and TFAM (c) activities in skeletalmuscle of young rats Results are mean plusmn SEM for four to six independent animals performed in quadruplicate No significant differencebetween groups was detected
The same pattern of alteration was also observed inmalate dehydrogenase activity which was found increasedin rats receiving carnosine acutely and unaltered in animalsreceiving the dipeptide chronically Thus it may be proposedthat skeletal muscle is able to adapt to high carnosinelevels recovering the impaired enzyme activities back tocontrol conditions A putative mechanism involved in suchadaptation is increased mitochondrial synthesis since theenzymes affected by carnosine administration present mito-chondrial localization Considering that carnosine adminis-tration increases vimentin synthesis [36] and that vimentin isa cytoskeleton protein that may be related to mitochondrialmotility and localization [37] it is possible that this processis mediated by increased vimentin synthesis However dataon NRF-1 PGC-1120572 and TFAM mRNA levels suggest thatcarnosine interferes with these enzymatic activities throughmechanisms unrelated to mitochondrial biogenesis but theinfluence of carnosine on mitophagy cannot be ruled out
At present we cannot ascertain the pathophysiologicalrelevance of our findings In this communication we inducedcarnosinemia in the animals by administering carnosineintraperitoneally With respect to this fact it should be men-tioned that carnosine dietary administration elicits increase
of its content in human skeletal muscle [38] The carnosinelevels in this tissue are situated within 30ndash50mmolkg ofwet weight [39] and are prone to alterations according todiet gender and age of subject [40] Additionally in ourstudy carnosine is increased in skeletal muscle of rat suchas occurrence in patients with carnosinemia
5 Conclusion
It is here shown that carnosine administration impairs elec-tron transfer through the mitochondrial respiratory chainin skeletal muscle of young rats without changing mito-chondrial biogenesis-related transcription factors At leastin part this finding does not corroborate some reportswhich indicate that this dipeptide increases the efficiencyand intensity of oxidative phosphorylation in rats adaptedto hypobaric hypoxia [41] The human carnosinase levelsare almost undetectable in plasma of neonates increasingaccording to aging [6] rendering young individuals lessprone to metabolize carnosine In case these findings areconfirmed by further studies and ATP depletion is alsoobserved it is possible that an energy dysfunction secondaryto carnosine accumulationmay aid to explain someofmuscle
8 BioMed Research International
00
05
10
15Re
lativ
e RN
A ex
pres
sion
Control Carnosine100 mgkg
(a)
00
05
10
15
Relat
ive R
NA
expr
essio
n
Control Carnosine100 mgkg
(b)
Control Carnosine100 mgkg
00
05
10
15
Relat
ive R
NA
expr
essio
n
(c)
Figure 8 Effect of chronic carnosine administration on relative mRNA expression of NRF-1 (a) PGC-1120572 (b) and TFAM (c) activities inskeletal muscle of young rats Results are mean plusmn SEM for four to six independent animals performed in quadruplicate No significantdifference between groups was detected
symptoms commonly observed in young patients with serumcarnosinase deficiency
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This research was supported by resources from Universidadedo Extremo Sul Catarinense and Brazilian National Councilfor Scientific and Technological Development (CNPq) LuizaWilges Kist was recipient of fellowships from Coordenacaode Aperfeicoamento de Pessoal de Nıvel Superior (CAPES)
References
[1] L Bonfanti P Peretto S deMarchis andA Fasolo ldquoCarnosine-related dipeptides in the mammalian brainrdquo Progress in Neuro-biology vol 59 no 4 pp 333ndash353 1999
[2] W Gulewitsch and S Amiradzibi ldquoUeber das carnosineine neue organische base des fleischextraktesrdquo Berichte der
Deutschen ChemischenGesellschaft vol 33 no 2 pp 1902ndash19031900
[3] K Nagai and T Suda ldquoRealization of spontaneous healingfunction by carnosinerdquo Methods and Findings in Experimentaland Clinical Pharmacology vol 10 no 8 pp 497ndash507 1988
[4] H Abe ldquoRole of histidine-related compounds as intracellularproton buffering constituents in vertebrate musclerdquo Biochem-istry vol 65 no 7 pp 757ndash765 2000
[5] S Geissler M Zwarg I Knutter F Markwardt and M Brand-sch ldquoThe bioactive dipeptide anserine is transported by humanproton-coupled peptide transportersrdquo FEBS Journal vol 277no 3 pp 790ndash795 2010
[6] J F Lenney R P George and A M Weiss ldquoHuman serumcarnosinase characterization distinction from cellular carnosi-nase and activation by cadmiumrdquo Clinica Chimica Acta vol123 no 3 pp 221ndash231 1982
[7] C R Scriver and K M Gibson ldquoDisorders of 120573- and 120574-aminoacids in free and peptide-linked formsrdquo in The Metabolic andMolecular Bases of Inherit C R Scriver A L Beaudet S Slyand D Valle Eds vol 1 part 5 pp 1349ndash1368 McGraw-HillNew York NY USA 7th edition 1995
[8] W H Murphey D G Lindmark and L I Patchen ldquoSerumcarnosinase deficiency concomitant with mental retardationrdquoPediatric Research vol 7 no 7 pp 601ndash606 1973
BioMed Research International 9
[9] H Lunde O Sjaastad and L Gjessing ldquoHomocarnosinosishypercarnosinuriardquo Journal of Neurochemistry vol 38 no 1 pp242ndash245 1982
[10] T L Perry S Hansen B Tischler R Bunting and K BerryldquoCarnosinemia A new metabolic disorder associated withneurologic disease andmental defectrdquoTheNewEngland Journalof Medicine vol 277 no 23 pp 1219ndash1227 1967
[11] S M Willi Y Zhang J B Hill M C Phelan R C Michaelisand K R Holden ldquoA deletion in the long arm of chromosome18 in a child with serum carnosinase deficiencyrdquo PediatricResearch vol 41 no 2 pp 210ndash213 1997
[12] B E Bonefeld B Elfving and GWegener ldquoReference genes fornormalization a study of rat brain tissuerdquo Synapse vol 62 no4 pp 302ndash309 2008
[13] Q Zhang YWu H Sha et al ldquoEarly exercise affects mitochon-drial transcription factors expression after cerebral ischemia inratsrdquo International Journal of Molecular Sciences vol 13 no 2pp 1670ndash1679 2012
[14] A H V Schapira V M Mann J M Cooper et al ldquoAnatomicand disease specificity of NADH CoQ1 reductase (complex I)deficiency in Parkinsonrsquos diseaserdquo Journal of Neurochemistryvol 55 no 6 pp 2142ndash2145 1990
[15] J C Fischer W Ruitenbeek and J A Berden ldquoDifferentialinvestigation of the capacity of succinate oxidation in humanskeletal musclerdquo Clinica Chimica Acta vol 153 no 1 pp 23ndash261985
[16] B P Hughes ldquoA method for the estimation of serum creatinekinase and its use in comparing creatine kinase and aldolaseactivity in normal and pathological serardquo Clinica Chimica Actavol 7 no 5 pp 597ndash603 1962
[17] P F Schuck G Leipnitz C A J Ribeiro et al ldquoInhibition ofcreatine kinase activity in vitro by ethylmalonic acid in cerebralcortex of young ratsrdquoNeurochemical Research vol 27 no 12 pp1633ndash1639 2002
[18] G B Kitto ldquoIntra- and extramitochondrial malate dehydroge-nases from chicken and tuna heartrdquo Methods in Enzymologyvol 13 pp 106ndash116 1969
[19] O H Lowry N J Rosebrough A L Farr and R J RandallldquoProtein measurement with the Folin phenol reagentrdquo TheJournal of Biological Chemistry vol 193 no 1 pp 265ndash275 1951
[20] J Drozak M Veiga-da-Cunha D Vertommen V Stroobantand E van Schaftingen ldquoMolecular identification of carno-sine synthase as ATP-grasp domain-containing protein 1(ATPGD1)rdquoThe Journal of Biological Chemistry vol 285 no 13pp 9346ndash9356 2010
[21] P J Quinn A A Boldyrev and V E Formazuyk ldquoCarnosineits properties functions and potential therapeutic applicationsrdquoMolecular Aspects of Medicine vol 13 no 5 pp 379ndash444 1992
[22] P S Manhiani J K Northcutt I Han W C Bridges and PL Dawson ldquoAntioxidant activity of carnosine extracted fromvarious poultry tissuesrdquo Poultry Science vol 92 no 2 pp 444ndash453 2013
[23] G Aldini M Orioli G Rossoni et al ldquoThe carbonyl scavengercarnosine ameliorates dyslipidaemia and renal function inZucker obese ratsrdquo Journal of Cellular and Molecular Medicinevol 15 no 6 pp 1339ndash1354 2011
[24] G Vistoli D deMaddis V Straniero et al ldquoExploring the spaceof histidine containing dipeptides in search of novel efficientRCS sequestering agentsrdquo European Journal of Medicinal Chem-istry vol 66 pp 153ndash160 2013
[25] Z Xie S P Baba B R Sweeney and O A Barski ldquoDetoxi-fication of aldehydes by histidine-containing dipeptides fromchemistry to clinical implicationsrdquo Chemico-Biological Interac-tions vol 202 no 1ndash3 pp 288ndash297 2013
[26] A R Hipkiss ldquoAging proteotoxicity mitochondria glycationNAD+ and carnosine possible inter-relationships and resolu-tion of the oxygen paradoxrdquoFrontiers inAgingNeuroscience vol2 article 10 2010
[27] A R Hipkiss ldquoEnergy metabolism proteotoxic stress andage-related dysfunction - Protection by carnosinerdquo MolecularAspects of Medicine vol 32 no 4ndash6 pp 267ndash278 2011
[28] A R Hipkiss S P Cartwright C Bromley S R Gross and RM Bill ldquoCarnosine can understanding its actions on energymetabolism and protein homeostasis inform its therapeuticpotentialrdquo Chemistry Central Journal vol 7 no 1 article 382013
[29] M-C Mong C-Y Chao and M-C Yin ldquoHistidine andcarnosine alleviated hepatic steatosis in mice consumed highsaturated fat dietrdquo European Journal of Pharmacology vol 653no 1ndash3 pp 82ndash88 2011
[30] D Mizuno and M Kawahara ldquoThe molecular mechanisms ofzinc neurotoxicity and the pathogenesis of vascular type seniledementiardquo International Journal of Molecular Sciences vol 14no 11 pp 22067ndash22081 2013
[31] A Asperger C Renner M Menzel R Gebhardt J Meixens-berger and F Gaunitz ldquoIdentification of factors involved inthe anti-tumor activity of carnosine on glioblastomas using aproteomics approachrdquo Cancer Investigation vol 29 no 4 pp272ndash281 2011
[32] L A Bharadwaj G F Davies I J Xavier and N Ovsenek ldquoL-carnosine and verapamil inhibit hypoxia-induced expression ofhypoxia inducible factor (HIF-1120572) in H9c2 cardiomyoblastsrdquoPharmacological Research vol 45 no 3 pp 175ndash181 2002
[33] E Ito-Kato N Suzuki M Maeno et al ldquoEffect of carnosine onrunt-related transcription factor-2core binding factor 120572-1 andSox9 expressions of human periodontal ligament cellsrdquo Journalof Periodontal Research vol 39 no 3 pp 199ndash204 2004
[34] V Calabrese C Colombrita E Guagliano et al ldquoProtectiveeffect of carnosine during nitrosative stress in astroglial cellculturesrdquo Neurochemical Research vol 30 no 6-7 pp 797ndash8072005
[35] K M Soliman M A Hamed and S A Ali ldquoHepatoprotectiveeffect of carnosine on liver biochemical parameters in chronicethanol intoxicated ratrdquoTheMedical Journal of the IslamicWorldAcademy of Sciences vol 16 no 2 pp 77ndash86 2006
[36] D Ikeda S-I Wada C Yoneda H Abe and S WatabeldquoCarnosine stimulates vimentin expression in cultured ratfibroblastsrdquoCell Structure and Function vol 24 no 2 pp 79ndash871999
[37] O E Nekrasova M G Mendez I S Chernoivanenko et alldquoVimentin intermediate filaments modulate the motility ofmitochondriardquoMolecular Biology of the Cell vol 22 no 13 pp2282ndash2289 2011
[38] C Sale B Saunders and R C Harris ldquoEffect of beta-alaninesupplementation onmuscle carnosine concentrations and exer-cise performancerdquoAmino Acids vol 39 no 2 pp 321ndash333 2010
[39] A A Boldyrev and S E Severin ldquoThe histidine-containingdipeptides carnosine and anserine distribution properties andbiological significancerdquoAdvances in Enzyme Regulation vol 30pp 175ndash188 1990
[40] I Everaert A Mooyaart A Baguet et al ldquoVegetarianismfemale gender and increasing age but not CNDP1 genotype
10 BioMed Research International
are associatedwith reducedmuscle carnosine levels in humansrdquoAmino Acids vol 40 no 4 pp 1221ndash1229 2011
[41] V N Korobov N M Doliba and I V Telegus ldquoCarnosine inadaptation to hypobaric hypoxiardquo Biokhimiya vol 58 no 5 pp740ndash744 1993
BioMed Research International 3
0
200
400
600
800
Control Carnosine100 mgkg
(nm
olmiddot
minus1
min
middotmg
prot
einminus
1 )
lowast
(a)
0
5
10
15
20
Control Carnosine100 mgkg
(nm
olmiddot
minus1
min
middotmg
prot
einminus
1 )
lowastlowast
(b)
00
25
50
75
100
Control Carnosine100 mgkg
(nm
olmiddot
minus1
min
middotmg
prot
einminus
1 )
(c)
Figure 1 Effect of acute carnosine administration on respiratory chain complexes IndashIII (a) II (b) and II-III (c) activities in skeletal muscleof young rats Data represent mean plusmn SEM for four to six independent animals performed in duplicate and are expressed in nmolminminus1mgproteinminus1 lowast119875 lt 005 lowastlowast119875 lt 001 compared to control group (Studentrsquos 119905-test)
0
10
20
30
Control Carnosine100 mgKg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(a)
0
20
40
60
80
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
(b)
Figure 2 Effect of acute carnosine administration on citric acid cycle enzymes succinate dehydrogenase (a) and malate dehydrogenase (b)activities in skeletal muscle of young rats Data are mean plusmn SEM for six independent animals performed in duplicate and are expressed innmolminminus1mg proteinminus1 No significant differences between groups were detected (Studentrsquos 119905-test)
4 BioMed Research International
Table 1 Primer sequences for RT-qPCR experiments included in the study
Gene Forward primer Reverse primerGapda 51015840-GCTAAGCAGTTGGTGGTGCA-31015840 51015840-TCACCACCATGGAGAAGGC-31015840
Hprt1a 51015840-GCAGACTTTGCTTTCCTTGG-31015840 51015840-GCAGACTTTGCTTTCCTTGG-31015840
NRF-1b 51015840-TTACTCTGCTGTGGCTGATGG-31015840 51015840-CCTCTGATGCTTGCGTCGTCT-31015840
PGC-1120572c 51015840-CGTTACACCTGTGACGCTTTCGCTG-31015840 51015840-CATACTTGCTCTTGGTGGAAGCAGG-31015840
TFAMc 51015840-AATTGAAGCTTGTAAATCAGGCTTGG-31015840 51015840-CGGATGAGATCACTTCGCCCAAC-31015840
According to aBonefeld et al 2008 [12] bZhang et al 2012 [13] cdesigned by authors
0
1
2
3
4
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
Figure 3 Effect of acute carnosine administration on creatinekinase activity in skeletal muscle of young rats Data are mean plusmnSEM for six independent animals performed in duplicate andare expressed in 120583mol creatine minminus1mg proteinminus1 No significantdifference between groups was observed (Studentrsquos 119905-test)
DNA contamination in accordance with the manufacturerrsquosinstructions The cDNA was synthesized with ImProm-IIReverse Transcription System (Promega) from 1120583g totalRNA following the manufacturerrsquos instruction QuantitativePCR was performed using SYBR Green I (Invitrogen) todetect double-strand cDNA synthesis Reactions were donein a volume of 25120583L using 125 120583L of diluted cDNA contain-ing a final concentration of 02x SYBR Green I (Invitrogen)100 120583M dNTP 1x PCR Buffer 3mMMgCl
2 025U Platinum
Taq DNA Polymerase (Invitrogen) 05M of betaine (forGapd [12] Hprt1 [12] PGC-1120572 andTFAM) 2 of the reactionof DMSO (for NRF-1 [13]) and 200 nM of each of reverseand forward primers (Table 1) The PCR cycling conditionswere an initial polymerase activation step for 5min at 95∘C40 cycles of 15 s at 95∘C for denaturation 35 s at 60∘Cfor annealing and 15 s at 72∘C for elongation At the endof cycling protocol a melting-curve analysis was includedand fluorescence measured from 60 to 99∘C and showedin all cases one single peak Gapd and Hprt1 were used asreference genes for normalization Relative expression levelswere determined with 7500 and 7500 Fast Real-Time PCRSystems Software v206 (Applied Biosystems)The efficiencyper sample was calculated using LinRegPCR 20123 Software(httpLinRegPCRnl) Relative mRNA expression levelswere determined using the 2minusΔΔCT method
210 Protein Determination Protein was measured by Lowryand colleagues [19] method using bovine serum albumin asstandard
211 Statistical Analysis Results are presented as mean plusmnstandard error of mean (SEM) Assays were performed induplicate and the mean or median was used for statisticalanalysis Data was analyzed using Studentrsquos t-test Differencesbetween groups were rated significant at P le 005 Allanalyses were carried out in an IBM-compatible PC computerusing the Statistica version 7 software Molecular data wereexpressed as means plusmn SEM and analyzed by Studentrsquos t-test for unpaired samples considering 119875 lt 005 as statisticalsignificance
3 Results
31 Energy Metabolism Is Impaired by Acute CarnosineAdministration in Skeletal Muscle of Young Rats Initiallywe tested the influence of acute carnosine administrationIn such condition we observed a statistically significantdecrease of complexes IndashIII [119905
(10)= 241 119875 lt 005]
(Figure 1(a)) and II [t(8)
= 386 119875 lt 001] (Figure 1(b))activities in treated animals as compared to control grouprats Furthermore the complexes II-III [119905
(10)= 0918 119875 gt
005] (Figure 1(c)) succinate dehydrogenase [119905(10)
= 0211119875 gt005] (Figure 2(a)) malate dehydrogenase [119905
(10)= 156 119875 gt
005] (Figure 2(b)) and creatine kinase [119905(10)
= 105119875 gt 005](Figure 3) activities were not significantly different betweengroups However we identified a nonsignificant decreasein complexes II-III and malate dehydrogenase activities inanimals that received carnosine as compared to control rats(Figures 1(c) and 2(b) resp)
32 Chronic Carnosine Administration Does Not Affect EnergyMetabolism in Rat Skeletal Muscle The biochemical param-eters following chronic carnosine administration were alsoevaluated We detected no statistically significant differencein complexes IndashIII [t
(9)= minus0169 119875 gt 005] (Figure 4(a)) II
[t(9)
= minus0383 119875 gt 005] (Figure 4(b)) II-III [t(9)
= minus0289119875 gt 005] (Figure 4(c)) succinate dehydrogenase [119905
(10)=
0274 119875 gt 005] (Figure 5(a)) malate dehydrogenase [119905(10)
= 0374 119875 gt 005] (Figure 5(b)) and creatine kinase [119905(10)
=minus0113 119875 gt 005] (Figure 6) activities between groups in ratskeletal muscle However it was observed that there is a trend
BioMed Research International 5
0
100
200
300
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(a)
0
5
10
15
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(b)
0
5
10
15
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(c)
Figure 4 Effect of chronic carnosine administration on respiratory chain complexes IndashIII (a) II (b) and II-III (c) activities in skeletal muscleof young rats Data are mean plusmn SEM for five to six independent animals performed in duplicate and are expressed in nmolminminus1mgproteinminus1 No significant difference between groups was detected (Studentrsquos 119905-test)
0
5
10
15
20
25
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(a)
0
10
20
30
40
50
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
(b)
Figure 5 Effect of chronic carnosine administration on citric acid cycle enzymes succinate dehydrogenase (a) and malate dehydrogenase (b)activities in skeletal muscle of young rats Data are mean plusmn SEM for six independent animals performed in duplicate and are expressed innmolminminus1mg proteinminus1 No significant difference between groups was observed (Studentrsquos 119905-test)
6 BioMed Research International
00
05
10
15
20
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
Figure 6 Effect of chronic carnosine administration on creatinekinase activity in skeletal muscle of young rats Data are mean plusmnSEM for six independent animals performed in duplicate andare expressed in 120583mol creatine minminus1mg proteinminus1 No significantdifference between groups was detected (Studentrsquos 119905-test)
of decrease of complexes IndashIII activities in animals receivingcarnosine as compared to control rats (Figure 4(a))
33 Mitochondrial Biogenesis Transcript abundance of keyfactors involved in mitochondrial biogenesis was evaluatedbyRT-qPCR in skeletalmuscle of animals receiving carnosineacutely (Figure 7) or chronically (Figure 8) As compared tocontrol groups mRNA levels of NRF-1 PGC-1120572 and TFAMin the animals receiving carnosine under either regimen ofadministration were unchanged
4 Discussion
In skeletal muscle tissue of most vertebrates and some inver-tebrates the imidazole dipeptide carnosine is synthesizedfrom histidine and 120573-alanine by the ATP-dependent enzymecarnosine synthetase [20] The biological roles of carnosineand related dipeptides in skeletal muscle remain uncertainalthough it is possible that they act as proton buffering agents[4] Furthermore there are a growing number of evidencesrelated to carnosine effects on metabolism Aside from itsantioxidant activity [21 22] carbonyl-scavenger ability hasbeen assigned to carnosine [23] and other imidazole dipep-tides [24] Many of carbonyl compounds are metabolic toxicby-products and in the bulk of tissues they are detoxifiedby oxidoreductases These enzymes catalyze the oxidation orreduction of aldehydes or its conjugation with lowmolecularweight amines and thiols such as reduced glutathione andimidazole dipeptides [25] Carnosine was also shown toreact with methylglyoxal a toxic metabolic by-product ofglycolysis as well as to inhibit cancer and aging mechanisms[26] and the cellular respiration regulatory complex mTOR[27] and activate the gluconeogenic enzyme fructose-16-bisphosphatase [28]
It was also shown that carnosine intake significantlydiminishes the activity and mRNA expression of malic
enzyme fatty acid synthase 3-hydroxy-3-methylglutarylcoenzyme A (HMG-CoA) reductase sterol regulatoryelement-binding proteins (SREBP-1c) and SREBP-2 inmice that consumed high saturated fat diet [29] Carnosinewas able to inhibit Zn2+-induced death of GT1-7 neuronsthrough the inhibition of GADD34 p8 (endoplasmicreticulum stress-related genes) and Arc (calcium-relatedgene) expression [30] Finally Asperger and colleagues [31]through a proteomics study with glioblastoma cells receivingcarnosine detected 31 proteins expressed differentiallyunder the influence of dipeptide including BCL2-associatedathanogene 2 and von Hippel-Lindau binding protein 1
In the present study we observed a statistically sig-nificant decrease of complexes IndashIII and II activities inanimals receiving carnosine acutely compared to controlgroup rats Additionally we detected no significant differencein complexes II-III citric acid cycle enzymes and creatinekinase activities between groups Finally we were also notable to find any significant alteration of respiratory chaincomplexes citric acid cycle enzymes and creatine kinaseactivities Since several evidences suggest that many of thephysiological effects of carnosine administration initiate attranscriptional level as shown by changes in mRNA levels ofHIF-1120572 [32] runt-related transcription factor-2core bindingfactor alpha-1 (RUNX2Cbf1205721) Sox9 [33] Hsp70 and SOD2[34] mitochondrial biogenesis could explain these findingsNevertheless it was found that mRNA levels of NRF-1 PGC-1120572 and TFAM in skeletal muscle of rats receiving carnosineacutely or chronically were similar to those measured incontrol animals
Our finding showing that the malate dehydrogenaseactivity presented no statistically significant differencebetween groups in both treatments corroborates at least inpart a literature report It was found that a patient with serumcarnosinase deficiency exhibited malate dehydrogenase andlactate dehydrogenase activities within normal limits in liverkidney and spleen specimens despite the fact that patientswith this disorder present high levels of carnosine in theirplasma (carnosinemia) [8]
We also observed that succinate dehydrogenase activityin skeletal muscle of carnosine group was not significantlydifferent from those in control group in animals subjectedto either acute or chronic carnosine administration On theother hand a literature report showed that rats chronicallyintoxicated with ethanol presented reduced activity of somehepatic enzymes including succinate dehydrogenase [35] Itwas also showed in the same study that carnosine load for twoweeks previously or simultaneously with ethanol intoxicationprevents or reverses the toxicity of such alcohol on thisenzyme activity
It should be emphasized that in skeletal muscle of ratsreceiving carnosine acutely a statistically significant decreasein complexes IndashIII and II activities was identified as well asa trend of decrease in complexes II-III activities relatively tocontrol group However rats receiving carnosine chronicallyshowed only a trend of decrease in complexes IndashIII activitiesin skeletal muscle as compared to control group whereas theother complexes activities were not different between groups
BioMed Research International 7
00
05
10
15Re
lativ
e RN
A ex
pres
sion
Control Carnosine100 mgkg
(a)
Relat
ive R
NA
expr
essio
n
00
05
10
15
Control Carnosine100 mgkg
(b)
00
05
10
15
Relat
ive R
NA
expr
essio
n
Control Carnosine100 mgkg
(c)
Figure 7 Effect of acute carnosine administration on relative mRNA expression ofNRF-1 (a) PGC-1120572 (b) and TFAM (c) activities in skeletalmuscle of young rats Results are mean plusmn SEM for four to six independent animals performed in quadruplicate No significant differencebetween groups was detected
The same pattern of alteration was also observed inmalate dehydrogenase activity which was found increasedin rats receiving carnosine acutely and unaltered in animalsreceiving the dipeptide chronically Thus it may be proposedthat skeletal muscle is able to adapt to high carnosinelevels recovering the impaired enzyme activities back tocontrol conditions A putative mechanism involved in suchadaptation is increased mitochondrial synthesis since theenzymes affected by carnosine administration present mito-chondrial localization Considering that carnosine adminis-tration increases vimentin synthesis [36] and that vimentin isa cytoskeleton protein that may be related to mitochondrialmotility and localization [37] it is possible that this processis mediated by increased vimentin synthesis However dataon NRF-1 PGC-1120572 and TFAM mRNA levels suggest thatcarnosine interferes with these enzymatic activities throughmechanisms unrelated to mitochondrial biogenesis but theinfluence of carnosine on mitophagy cannot be ruled out
At present we cannot ascertain the pathophysiologicalrelevance of our findings In this communication we inducedcarnosinemia in the animals by administering carnosineintraperitoneally With respect to this fact it should be men-tioned that carnosine dietary administration elicits increase
of its content in human skeletal muscle [38] The carnosinelevels in this tissue are situated within 30ndash50mmolkg ofwet weight [39] and are prone to alterations according todiet gender and age of subject [40] Additionally in ourstudy carnosine is increased in skeletal muscle of rat suchas occurrence in patients with carnosinemia
5 Conclusion
It is here shown that carnosine administration impairs elec-tron transfer through the mitochondrial respiratory chainin skeletal muscle of young rats without changing mito-chondrial biogenesis-related transcription factors At leastin part this finding does not corroborate some reportswhich indicate that this dipeptide increases the efficiencyand intensity of oxidative phosphorylation in rats adaptedto hypobaric hypoxia [41] The human carnosinase levelsare almost undetectable in plasma of neonates increasingaccording to aging [6] rendering young individuals lessprone to metabolize carnosine In case these findings areconfirmed by further studies and ATP depletion is alsoobserved it is possible that an energy dysfunction secondaryto carnosine accumulationmay aid to explain someofmuscle
8 BioMed Research International
00
05
10
15Re
lativ
e RN
A ex
pres
sion
Control Carnosine100 mgkg
(a)
00
05
10
15
Relat
ive R
NA
expr
essio
n
Control Carnosine100 mgkg
(b)
Control Carnosine100 mgkg
00
05
10
15
Relat
ive R
NA
expr
essio
n
(c)
Figure 8 Effect of chronic carnosine administration on relative mRNA expression of NRF-1 (a) PGC-1120572 (b) and TFAM (c) activities inskeletal muscle of young rats Results are mean plusmn SEM for four to six independent animals performed in quadruplicate No significantdifference between groups was detected
symptoms commonly observed in young patients with serumcarnosinase deficiency
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This research was supported by resources from Universidadedo Extremo Sul Catarinense and Brazilian National Councilfor Scientific and Technological Development (CNPq) LuizaWilges Kist was recipient of fellowships from Coordenacaode Aperfeicoamento de Pessoal de Nıvel Superior (CAPES)
References
[1] L Bonfanti P Peretto S deMarchis andA Fasolo ldquoCarnosine-related dipeptides in the mammalian brainrdquo Progress in Neuro-biology vol 59 no 4 pp 333ndash353 1999
[2] W Gulewitsch and S Amiradzibi ldquoUeber das carnosineine neue organische base des fleischextraktesrdquo Berichte der
Deutschen ChemischenGesellschaft vol 33 no 2 pp 1902ndash19031900
[3] K Nagai and T Suda ldquoRealization of spontaneous healingfunction by carnosinerdquo Methods and Findings in Experimentaland Clinical Pharmacology vol 10 no 8 pp 497ndash507 1988
[4] H Abe ldquoRole of histidine-related compounds as intracellularproton buffering constituents in vertebrate musclerdquo Biochem-istry vol 65 no 7 pp 757ndash765 2000
[5] S Geissler M Zwarg I Knutter F Markwardt and M Brand-sch ldquoThe bioactive dipeptide anserine is transported by humanproton-coupled peptide transportersrdquo FEBS Journal vol 277no 3 pp 790ndash795 2010
[6] J F Lenney R P George and A M Weiss ldquoHuman serumcarnosinase characterization distinction from cellular carnosi-nase and activation by cadmiumrdquo Clinica Chimica Acta vol123 no 3 pp 221ndash231 1982
[7] C R Scriver and K M Gibson ldquoDisorders of 120573- and 120574-aminoacids in free and peptide-linked formsrdquo in The Metabolic andMolecular Bases of Inherit C R Scriver A L Beaudet S Slyand D Valle Eds vol 1 part 5 pp 1349ndash1368 McGraw-HillNew York NY USA 7th edition 1995
[8] W H Murphey D G Lindmark and L I Patchen ldquoSerumcarnosinase deficiency concomitant with mental retardationrdquoPediatric Research vol 7 no 7 pp 601ndash606 1973
BioMed Research International 9
[9] H Lunde O Sjaastad and L Gjessing ldquoHomocarnosinosishypercarnosinuriardquo Journal of Neurochemistry vol 38 no 1 pp242ndash245 1982
[10] T L Perry S Hansen B Tischler R Bunting and K BerryldquoCarnosinemia A new metabolic disorder associated withneurologic disease andmental defectrdquoTheNewEngland Journalof Medicine vol 277 no 23 pp 1219ndash1227 1967
[11] S M Willi Y Zhang J B Hill M C Phelan R C Michaelisand K R Holden ldquoA deletion in the long arm of chromosome18 in a child with serum carnosinase deficiencyrdquo PediatricResearch vol 41 no 2 pp 210ndash213 1997
[12] B E Bonefeld B Elfving and GWegener ldquoReference genes fornormalization a study of rat brain tissuerdquo Synapse vol 62 no4 pp 302ndash309 2008
[13] Q Zhang YWu H Sha et al ldquoEarly exercise affects mitochon-drial transcription factors expression after cerebral ischemia inratsrdquo International Journal of Molecular Sciences vol 13 no 2pp 1670ndash1679 2012
[14] A H V Schapira V M Mann J M Cooper et al ldquoAnatomicand disease specificity of NADH CoQ1 reductase (complex I)deficiency in Parkinsonrsquos diseaserdquo Journal of Neurochemistryvol 55 no 6 pp 2142ndash2145 1990
[15] J C Fischer W Ruitenbeek and J A Berden ldquoDifferentialinvestigation of the capacity of succinate oxidation in humanskeletal musclerdquo Clinica Chimica Acta vol 153 no 1 pp 23ndash261985
[16] B P Hughes ldquoA method for the estimation of serum creatinekinase and its use in comparing creatine kinase and aldolaseactivity in normal and pathological serardquo Clinica Chimica Actavol 7 no 5 pp 597ndash603 1962
[17] P F Schuck G Leipnitz C A J Ribeiro et al ldquoInhibition ofcreatine kinase activity in vitro by ethylmalonic acid in cerebralcortex of young ratsrdquoNeurochemical Research vol 27 no 12 pp1633ndash1639 2002
[18] G B Kitto ldquoIntra- and extramitochondrial malate dehydroge-nases from chicken and tuna heartrdquo Methods in Enzymologyvol 13 pp 106ndash116 1969
[19] O H Lowry N J Rosebrough A L Farr and R J RandallldquoProtein measurement with the Folin phenol reagentrdquo TheJournal of Biological Chemistry vol 193 no 1 pp 265ndash275 1951
[20] J Drozak M Veiga-da-Cunha D Vertommen V Stroobantand E van Schaftingen ldquoMolecular identification of carno-sine synthase as ATP-grasp domain-containing protein 1(ATPGD1)rdquoThe Journal of Biological Chemistry vol 285 no 13pp 9346ndash9356 2010
[21] P J Quinn A A Boldyrev and V E Formazuyk ldquoCarnosineits properties functions and potential therapeutic applicationsrdquoMolecular Aspects of Medicine vol 13 no 5 pp 379ndash444 1992
[22] P S Manhiani J K Northcutt I Han W C Bridges and PL Dawson ldquoAntioxidant activity of carnosine extracted fromvarious poultry tissuesrdquo Poultry Science vol 92 no 2 pp 444ndash453 2013
[23] G Aldini M Orioli G Rossoni et al ldquoThe carbonyl scavengercarnosine ameliorates dyslipidaemia and renal function inZucker obese ratsrdquo Journal of Cellular and Molecular Medicinevol 15 no 6 pp 1339ndash1354 2011
[24] G Vistoli D deMaddis V Straniero et al ldquoExploring the spaceof histidine containing dipeptides in search of novel efficientRCS sequestering agentsrdquo European Journal of Medicinal Chem-istry vol 66 pp 153ndash160 2013
[25] Z Xie S P Baba B R Sweeney and O A Barski ldquoDetoxi-fication of aldehydes by histidine-containing dipeptides fromchemistry to clinical implicationsrdquo Chemico-Biological Interac-tions vol 202 no 1ndash3 pp 288ndash297 2013
[26] A R Hipkiss ldquoAging proteotoxicity mitochondria glycationNAD+ and carnosine possible inter-relationships and resolu-tion of the oxygen paradoxrdquoFrontiers inAgingNeuroscience vol2 article 10 2010
[27] A R Hipkiss ldquoEnergy metabolism proteotoxic stress andage-related dysfunction - Protection by carnosinerdquo MolecularAspects of Medicine vol 32 no 4ndash6 pp 267ndash278 2011
[28] A R Hipkiss S P Cartwright C Bromley S R Gross and RM Bill ldquoCarnosine can understanding its actions on energymetabolism and protein homeostasis inform its therapeuticpotentialrdquo Chemistry Central Journal vol 7 no 1 article 382013
[29] M-C Mong C-Y Chao and M-C Yin ldquoHistidine andcarnosine alleviated hepatic steatosis in mice consumed highsaturated fat dietrdquo European Journal of Pharmacology vol 653no 1ndash3 pp 82ndash88 2011
[30] D Mizuno and M Kawahara ldquoThe molecular mechanisms ofzinc neurotoxicity and the pathogenesis of vascular type seniledementiardquo International Journal of Molecular Sciences vol 14no 11 pp 22067ndash22081 2013
[31] A Asperger C Renner M Menzel R Gebhardt J Meixens-berger and F Gaunitz ldquoIdentification of factors involved inthe anti-tumor activity of carnosine on glioblastomas using aproteomics approachrdquo Cancer Investigation vol 29 no 4 pp272ndash281 2011
[32] L A Bharadwaj G F Davies I J Xavier and N Ovsenek ldquoL-carnosine and verapamil inhibit hypoxia-induced expression ofhypoxia inducible factor (HIF-1120572) in H9c2 cardiomyoblastsrdquoPharmacological Research vol 45 no 3 pp 175ndash181 2002
[33] E Ito-Kato N Suzuki M Maeno et al ldquoEffect of carnosine onrunt-related transcription factor-2core binding factor 120572-1 andSox9 expressions of human periodontal ligament cellsrdquo Journalof Periodontal Research vol 39 no 3 pp 199ndash204 2004
[34] V Calabrese C Colombrita E Guagliano et al ldquoProtectiveeffect of carnosine during nitrosative stress in astroglial cellculturesrdquo Neurochemical Research vol 30 no 6-7 pp 797ndash8072005
[35] K M Soliman M A Hamed and S A Ali ldquoHepatoprotectiveeffect of carnosine on liver biochemical parameters in chronicethanol intoxicated ratrdquoTheMedical Journal of the IslamicWorldAcademy of Sciences vol 16 no 2 pp 77ndash86 2006
[36] D Ikeda S-I Wada C Yoneda H Abe and S WatabeldquoCarnosine stimulates vimentin expression in cultured ratfibroblastsrdquoCell Structure and Function vol 24 no 2 pp 79ndash871999
[37] O E Nekrasova M G Mendez I S Chernoivanenko et alldquoVimentin intermediate filaments modulate the motility ofmitochondriardquoMolecular Biology of the Cell vol 22 no 13 pp2282ndash2289 2011
[38] C Sale B Saunders and R C Harris ldquoEffect of beta-alaninesupplementation onmuscle carnosine concentrations and exer-cise performancerdquoAmino Acids vol 39 no 2 pp 321ndash333 2010
[39] A A Boldyrev and S E Severin ldquoThe histidine-containingdipeptides carnosine and anserine distribution properties andbiological significancerdquoAdvances in Enzyme Regulation vol 30pp 175ndash188 1990
[40] I Everaert A Mooyaart A Baguet et al ldquoVegetarianismfemale gender and increasing age but not CNDP1 genotype
10 BioMed Research International
are associatedwith reducedmuscle carnosine levels in humansrdquoAmino Acids vol 40 no 4 pp 1221ndash1229 2011
[41] V N Korobov N M Doliba and I V Telegus ldquoCarnosine inadaptation to hypobaric hypoxiardquo Biokhimiya vol 58 no 5 pp740ndash744 1993
4 BioMed Research International
Table 1 Primer sequences for RT-qPCR experiments included in the study
Gene Forward primer Reverse primerGapda 51015840-GCTAAGCAGTTGGTGGTGCA-31015840 51015840-TCACCACCATGGAGAAGGC-31015840
Hprt1a 51015840-GCAGACTTTGCTTTCCTTGG-31015840 51015840-GCAGACTTTGCTTTCCTTGG-31015840
NRF-1b 51015840-TTACTCTGCTGTGGCTGATGG-31015840 51015840-CCTCTGATGCTTGCGTCGTCT-31015840
PGC-1120572c 51015840-CGTTACACCTGTGACGCTTTCGCTG-31015840 51015840-CATACTTGCTCTTGGTGGAAGCAGG-31015840
TFAMc 51015840-AATTGAAGCTTGTAAATCAGGCTTGG-31015840 51015840-CGGATGAGATCACTTCGCCCAAC-31015840
According to aBonefeld et al 2008 [12] bZhang et al 2012 [13] cdesigned by authors
0
1
2
3
4
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
Figure 3 Effect of acute carnosine administration on creatinekinase activity in skeletal muscle of young rats Data are mean plusmnSEM for six independent animals performed in duplicate andare expressed in 120583mol creatine minminus1mg proteinminus1 No significantdifference between groups was observed (Studentrsquos 119905-test)
DNA contamination in accordance with the manufacturerrsquosinstructions The cDNA was synthesized with ImProm-IIReverse Transcription System (Promega) from 1120583g totalRNA following the manufacturerrsquos instruction QuantitativePCR was performed using SYBR Green I (Invitrogen) todetect double-strand cDNA synthesis Reactions were donein a volume of 25120583L using 125 120583L of diluted cDNA contain-ing a final concentration of 02x SYBR Green I (Invitrogen)100 120583M dNTP 1x PCR Buffer 3mMMgCl
2 025U Platinum
Taq DNA Polymerase (Invitrogen) 05M of betaine (forGapd [12] Hprt1 [12] PGC-1120572 andTFAM) 2 of the reactionof DMSO (for NRF-1 [13]) and 200 nM of each of reverseand forward primers (Table 1) The PCR cycling conditionswere an initial polymerase activation step for 5min at 95∘C40 cycles of 15 s at 95∘C for denaturation 35 s at 60∘Cfor annealing and 15 s at 72∘C for elongation At the endof cycling protocol a melting-curve analysis was includedand fluorescence measured from 60 to 99∘C and showedin all cases one single peak Gapd and Hprt1 were used asreference genes for normalization Relative expression levelswere determined with 7500 and 7500 Fast Real-Time PCRSystems Software v206 (Applied Biosystems)The efficiencyper sample was calculated using LinRegPCR 20123 Software(httpLinRegPCRnl) Relative mRNA expression levelswere determined using the 2minusΔΔCT method
210 Protein Determination Protein was measured by Lowryand colleagues [19] method using bovine serum albumin asstandard
211 Statistical Analysis Results are presented as mean plusmnstandard error of mean (SEM) Assays were performed induplicate and the mean or median was used for statisticalanalysis Data was analyzed using Studentrsquos t-test Differencesbetween groups were rated significant at P le 005 Allanalyses were carried out in an IBM-compatible PC computerusing the Statistica version 7 software Molecular data wereexpressed as means plusmn SEM and analyzed by Studentrsquos t-test for unpaired samples considering 119875 lt 005 as statisticalsignificance
3 Results
31 Energy Metabolism Is Impaired by Acute CarnosineAdministration in Skeletal Muscle of Young Rats Initiallywe tested the influence of acute carnosine administrationIn such condition we observed a statistically significantdecrease of complexes IndashIII [119905
(10)= 241 119875 lt 005]
(Figure 1(a)) and II [t(8)
= 386 119875 lt 001] (Figure 1(b))activities in treated animals as compared to control grouprats Furthermore the complexes II-III [119905
(10)= 0918 119875 gt
005] (Figure 1(c)) succinate dehydrogenase [119905(10)
= 0211119875 gt005] (Figure 2(a)) malate dehydrogenase [119905
(10)= 156 119875 gt
005] (Figure 2(b)) and creatine kinase [119905(10)
= 105119875 gt 005](Figure 3) activities were not significantly different betweengroups However we identified a nonsignificant decreasein complexes II-III and malate dehydrogenase activities inanimals that received carnosine as compared to control rats(Figures 1(c) and 2(b) resp)
32 Chronic Carnosine Administration Does Not Affect EnergyMetabolism in Rat Skeletal Muscle The biochemical param-eters following chronic carnosine administration were alsoevaluated We detected no statistically significant differencein complexes IndashIII [t
(9)= minus0169 119875 gt 005] (Figure 4(a)) II
[t(9)
= minus0383 119875 gt 005] (Figure 4(b)) II-III [t(9)
= minus0289119875 gt 005] (Figure 4(c)) succinate dehydrogenase [119905
(10)=
0274 119875 gt 005] (Figure 5(a)) malate dehydrogenase [119905(10)
= 0374 119875 gt 005] (Figure 5(b)) and creatine kinase [119905(10)
=minus0113 119875 gt 005] (Figure 6) activities between groups in ratskeletal muscle However it was observed that there is a trend
BioMed Research International 5
0
100
200
300
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(a)
0
5
10
15
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(b)
0
5
10
15
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(c)
Figure 4 Effect of chronic carnosine administration on respiratory chain complexes IndashIII (a) II (b) and II-III (c) activities in skeletal muscleof young rats Data are mean plusmn SEM for five to six independent animals performed in duplicate and are expressed in nmolminminus1mgproteinminus1 No significant difference between groups was detected (Studentrsquos 119905-test)
0
5
10
15
20
25
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(a)
0
10
20
30
40
50
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
(b)
Figure 5 Effect of chronic carnosine administration on citric acid cycle enzymes succinate dehydrogenase (a) and malate dehydrogenase (b)activities in skeletal muscle of young rats Data are mean plusmn SEM for six independent animals performed in duplicate and are expressed innmolminminus1mg proteinminus1 No significant difference between groups was observed (Studentrsquos 119905-test)
6 BioMed Research International
00
05
10
15
20
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
Figure 6 Effect of chronic carnosine administration on creatinekinase activity in skeletal muscle of young rats Data are mean plusmnSEM for six independent animals performed in duplicate andare expressed in 120583mol creatine minminus1mg proteinminus1 No significantdifference between groups was detected (Studentrsquos 119905-test)
of decrease of complexes IndashIII activities in animals receivingcarnosine as compared to control rats (Figure 4(a))
33 Mitochondrial Biogenesis Transcript abundance of keyfactors involved in mitochondrial biogenesis was evaluatedbyRT-qPCR in skeletalmuscle of animals receiving carnosineacutely (Figure 7) or chronically (Figure 8) As compared tocontrol groups mRNA levels of NRF-1 PGC-1120572 and TFAMin the animals receiving carnosine under either regimen ofadministration were unchanged
4 Discussion
In skeletal muscle tissue of most vertebrates and some inver-tebrates the imidazole dipeptide carnosine is synthesizedfrom histidine and 120573-alanine by the ATP-dependent enzymecarnosine synthetase [20] The biological roles of carnosineand related dipeptides in skeletal muscle remain uncertainalthough it is possible that they act as proton buffering agents[4] Furthermore there are a growing number of evidencesrelated to carnosine effects on metabolism Aside from itsantioxidant activity [21 22] carbonyl-scavenger ability hasbeen assigned to carnosine [23] and other imidazole dipep-tides [24] Many of carbonyl compounds are metabolic toxicby-products and in the bulk of tissues they are detoxifiedby oxidoreductases These enzymes catalyze the oxidation orreduction of aldehydes or its conjugation with lowmolecularweight amines and thiols such as reduced glutathione andimidazole dipeptides [25] Carnosine was also shown toreact with methylglyoxal a toxic metabolic by-product ofglycolysis as well as to inhibit cancer and aging mechanisms[26] and the cellular respiration regulatory complex mTOR[27] and activate the gluconeogenic enzyme fructose-16-bisphosphatase [28]
It was also shown that carnosine intake significantlydiminishes the activity and mRNA expression of malic
enzyme fatty acid synthase 3-hydroxy-3-methylglutarylcoenzyme A (HMG-CoA) reductase sterol regulatoryelement-binding proteins (SREBP-1c) and SREBP-2 inmice that consumed high saturated fat diet [29] Carnosinewas able to inhibit Zn2+-induced death of GT1-7 neuronsthrough the inhibition of GADD34 p8 (endoplasmicreticulum stress-related genes) and Arc (calcium-relatedgene) expression [30] Finally Asperger and colleagues [31]through a proteomics study with glioblastoma cells receivingcarnosine detected 31 proteins expressed differentiallyunder the influence of dipeptide including BCL2-associatedathanogene 2 and von Hippel-Lindau binding protein 1
In the present study we observed a statistically sig-nificant decrease of complexes IndashIII and II activities inanimals receiving carnosine acutely compared to controlgroup rats Additionally we detected no significant differencein complexes II-III citric acid cycle enzymes and creatinekinase activities between groups Finally we were also notable to find any significant alteration of respiratory chaincomplexes citric acid cycle enzymes and creatine kinaseactivities Since several evidences suggest that many of thephysiological effects of carnosine administration initiate attranscriptional level as shown by changes in mRNA levels ofHIF-1120572 [32] runt-related transcription factor-2core bindingfactor alpha-1 (RUNX2Cbf1205721) Sox9 [33] Hsp70 and SOD2[34] mitochondrial biogenesis could explain these findingsNevertheless it was found that mRNA levels of NRF-1 PGC-1120572 and TFAM in skeletal muscle of rats receiving carnosineacutely or chronically were similar to those measured incontrol animals
Our finding showing that the malate dehydrogenaseactivity presented no statistically significant differencebetween groups in both treatments corroborates at least inpart a literature report It was found that a patient with serumcarnosinase deficiency exhibited malate dehydrogenase andlactate dehydrogenase activities within normal limits in liverkidney and spleen specimens despite the fact that patientswith this disorder present high levels of carnosine in theirplasma (carnosinemia) [8]
We also observed that succinate dehydrogenase activityin skeletal muscle of carnosine group was not significantlydifferent from those in control group in animals subjectedto either acute or chronic carnosine administration On theother hand a literature report showed that rats chronicallyintoxicated with ethanol presented reduced activity of somehepatic enzymes including succinate dehydrogenase [35] Itwas also showed in the same study that carnosine load for twoweeks previously or simultaneously with ethanol intoxicationprevents or reverses the toxicity of such alcohol on thisenzyme activity
It should be emphasized that in skeletal muscle of ratsreceiving carnosine acutely a statistically significant decreasein complexes IndashIII and II activities was identified as well asa trend of decrease in complexes II-III activities relatively tocontrol group However rats receiving carnosine chronicallyshowed only a trend of decrease in complexes IndashIII activitiesin skeletal muscle as compared to control group whereas theother complexes activities were not different between groups
BioMed Research International 7
00
05
10
15Re
lativ
e RN
A ex
pres
sion
Control Carnosine100 mgkg
(a)
Relat
ive R
NA
expr
essio
n
00
05
10
15
Control Carnosine100 mgkg
(b)
00
05
10
15
Relat
ive R
NA
expr
essio
n
Control Carnosine100 mgkg
(c)
Figure 7 Effect of acute carnosine administration on relative mRNA expression ofNRF-1 (a) PGC-1120572 (b) and TFAM (c) activities in skeletalmuscle of young rats Results are mean plusmn SEM for four to six independent animals performed in quadruplicate No significant differencebetween groups was detected
The same pattern of alteration was also observed inmalate dehydrogenase activity which was found increasedin rats receiving carnosine acutely and unaltered in animalsreceiving the dipeptide chronically Thus it may be proposedthat skeletal muscle is able to adapt to high carnosinelevels recovering the impaired enzyme activities back tocontrol conditions A putative mechanism involved in suchadaptation is increased mitochondrial synthesis since theenzymes affected by carnosine administration present mito-chondrial localization Considering that carnosine adminis-tration increases vimentin synthesis [36] and that vimentin isa cytoskeleton protein that may be related to mitochondrialmotility and localization [37] it is possible that this processis mediated by increased vimentin synthesis However dataon NRF-1 PGC-1120572 and TFAM mRNA levels suggest thatcarnosine interferes with these enzymatic activities throughmechanisms unrelated to mitochondrial biogenesis but theinfluence of carnosine on mitophagy cannot be ruled out
At present we cannot ascertain the pathophysiologicalrelevance of our findings In this communication we inducedcarnosinemia in the animals by administering carnosineintraperitoneally With respect to this fact it should be men-tioned that carnosine dietary administration elicits increase
of its content in human skeletal muscle [38] The carnosinelevels in this tissue are situated within 30ndash50mmolkg ofwet weight [39] and are prone to alterations according todiet gender and age of subject [40] Additionally in ourstudy carnosine is increased in skeletal muscle of rat suchas occurrence in patients with carnosinemia
5 Conclusion
It is here shown that carnosine administration impairs elec-tron transfer through the mitochondrial respiratory chainin skeletal muscle of young rats without changing mito-chondrial biogenesis-related transcription factors At leastin part this finding does not corroborate some reportswhich indicate that this dipeptide increases the efficiencyand intensity of oxidative phosphorylation in rats adaptedto hypobaric hypoxia [41] The human carnosinase levelsare almost undetectable in plasma of neonates increasingaccording to aging [6] rendering young individuals lessprone to metabolize carnosine In case these findings areconfirmed by further studies and ATP depletion is alsoobserved it is possible that an energy dysfunction secondaryto carnosine accumulationmay aid to explain someofmuscle
8 BioMed Research International
00
05
10
15Re
lativ
e RN
A ex
pres
sion
Control Carnosine100 mgkg
(a)
00
05
10
15
Relat
ive R
NA
expr
essio
n
Control Carnosine100 mgkg
(b)
Control Carnosine100 mgkg
00
05
10
15
Relat
ive R
NA
expr
essio
n
(c)
Figure 8 Effect of chronic carnosine administration on relative mRNA expression of NRF-1 (a) PGC-1120572 (b) and TFAM (c) activities inskeletal muscle of young rats Results are mean plusmn SEM for four to six independent animals performed in quadruplicate No significantdifference between groups was detected
symptoms commonly observed in young patients with serumcarnosinase deficiency
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This research was supported by resources from Universidadedo Extremo Sul Catarinense and Brazilian National Councilfor Scientific and Technological Development (CNPq) LuizaWilges Kist was recipient of fellowships from Coordenacaode Aperfeicoamento de Pessoal de Nıvel Superior (CAPES)
References
[1] L Bonfanti P Peretto S deMarchis andA Fasolo ldquoCarnosine-related dipeptides in the mammalian brainrdquo Progress in Neuro-biology vol 59 no 4 pp 333ndash353 1999
[2] W Gulewitsch and S Amiradzibi ldquoUeber das carnosineine neue organische base des fleischextraktesrdquo Berichte der
Deutschen ChemischenGesellschaft vol 33 no 2 pp 1902ndash19031900
[3] K Nagai and T Suda ldquoRealization of spontaneous healingfunction by carnosinerdquo Methods and Findings in Experimentaland Clinical Pharmacology vol 10 no 8 pp 497ndash507 1988
[4] H Abe ldquoRole of histidine-related compounds as intracellularproton buffering constituents in vertebrate musclerdquo Biochem-istry vol 65 no 7 pp 757ndash765 2000
[5] S Geissler M Zwarg I Knutter F Markwardt and M Brand-sch ldquoThe bioactive dipeptide anserine is transported by humanproton-coupled peptide transportersrdquo FEBS Journal vol 277no 3 pp 790ndash795 2010
[6] J F Lenney R P George and A M Weiss ldquoHuman serumcarnosinase characterization distinction from cellular carnosi-nase and activation by cadmiumrdquo Clinica Chimica Acta vol123 no 3 pp 221ndash231 1982
[7] C R Scriver and K M Gibson ldquoDisorders of 120573- and 120574-aminoacids in free and peptide-linked formsrdquo in The Metabolic andMolecular Bases of Inherit C R Scriver A L Beaudet S Slyand D Valle Eds vol 1 part 5 pp 1349ndash1368 McGraw-HillNew York NY USA 7th edition 1995
[8] W H Murphey D G Lindmark and L I Patchen ldquoSerumcarnosinase deficiency concomitant with mental retardationrdquoPediatric Research vol 7 no 7 pp 601ndash606 1973
BioMed Research International 9
[9] H Lunde O Sjaastad and L Gjessing ldquoHomocarnosinosishypercarnosinuriardquo Journal of Neurochemistry vol 38 no 1 pp242ndash245 1982
[10] T L Perry S Hansen B Tischler R Bunting and K BerryldquoCarnosinemia A new metabolic disorder associated withneurologic disease andmental defectrdquoTheNewEngland Journalof Medicine vol 277 no 23 pp 1219ndash1227 1967
[11] S M Willi Y Zhang J B Hill M C Phelan R C Michaelisand K R Holden ldquoA deletion in the long arm of chromosome18 in a child with serum carnosinase deficiencyrdquo PediatricResearch vol 41 no 2 pp 210ndash213 1997
[12] B E Bonefeld B Elfving and GWegener ldquoReference genes fornormalization a study of rat brain tissuerdquo Synapse vol 62 no4 pp 302ndash309 2008
[13] Q Zhang YWu H Sha et al ldquoEarly exercise affects mitochon-drial transcription factors expression after cerebral ischemia inratsrdquo International Journal of Molecular Sciences vol 13 no 2pp 1670ndash1679 2012
[14] A H V Schapira V M Mann J M Cooper et al ldquoAnatomicand disease specificity of NADH CoQ1 reductase (complex I)deficiency in Parkinsonrsquos diseaserdquo Journal of Neurochemistryvol 55 no 6 pp 2142ndash2145 1990
[15] J C Fischer W Ruitenbeek and J A Berden ldquoDifferentialinvestigation of the capacity of succinate oxidation in humanskeletal musclerdquo Clinica Chimica Acta vol 153 no 1 pp 23ndash261985
[16] B P Hughes ldquoA method for the estimation of serum creatinekinase and its use in comparing creatine kinase and aldolaseactivity in normal and pathological serardquo Clinica Chimica Actavol 7 no 5 pp 597ndash603 1962
[17] P F Schuck G Leipnitz C A J Ribeiro et al ldquoInhibition ofcreatine kinase activity in vitro by ethylmalonic acid in cerebralcortex of young ratsrdquoNeurochemical Research vol 27 no 12 pp1633ndash1639 2002
[18] G B Kitto ldquoIntra- and extramitochondrial malate dehydroge-nases from chicken and tuna heartrdquo Methods in Enzymologyvol 13 pp 106ndash116 1969
[19] O H Lowry N J Rosebrough A L Farr and R J RandallldquoProtein measurement with the Folin phenol reagentrdquo TheJournal of Biological Chemistry vol 193 no 1 pp 265ndash275 1951
[20] J Drozak M Veiga-da-Cunha D Vertommen V Stroobantand E van Schaftingen ldquoMolecular identification of carno-sine synthase as ATP-grasp domain-containing protein 1(ATPGD1)rdquoThe Journal of Biological Chemistry vol 285 no 13pp 9346ndash9356 2010
[21] P J Quinn A A Boldyrev and V E Formazuyk ldquoCarnosineits properties functions and potential therapeutic applicationsrdquoMolecular Aspects of Medicine vol 13 no 5 pp 379ndash444 1992
[22] P S Manhiani J K Northcutt I Han W C Bridges and PL Dawson ldquoAntioxidant activity of carnosine extracted fromvarious poultry tissuesrdquo Poultry Science vol 92 no 2 pp 444ndash453 2013
[23] G Aldini M Orioli G Rossoni et al ldquoThe carbonyl scavengercarnosine ameliorates dyslipidaemia and renal function inZucker obese ratsrdquo Journal of Cellular and Molecular Medicinevol 15 no 6 pp 1339ndash1354 2011
[24] G Vistoli D deMaddis V Straniero et al ldquoExploring the spaceof histidine containing dipeptides in search of novel efficientRCS sequestering agentsrdquo European Journal of Medicinal Chem-istry vol 66 pp 153ndash160 2013
[25] Z Xie S P Baba B R Sweeney and O A Barski ldquoDetoxi-fication of aldehydes by histidine-containing dipeptides fromchemistry to clinical implicationsrdquo Chemico-Biological Interac-tions vol 202 no 1ndash3 pp 288ndash297 2013
[26] A R Hipkiss ldquoAging proteotoxicity mitochondria glycationNAD+ and carnosine possible inter-relationships and resolu-tion of the oxygen paradoxrdquoFrontiers inAgingNeuroscience vol2 article 10 2010
[27] A R Hipkiss ldquoEnergy metabolism proteotoxic stress andage-related dysfunction - Protection by carnosinerdquo MolecularAspects of Medicine vol 32 no 4ndash6 pp 267ndash278 2011
[28] A R Hipkiss S P Cartwright C Bromley S R Gross and RM Bill ldquoCarnosine can understanding its actions on energymetabolism and protein homeostasis inform its therapeuticpotentialrdquo Chemistry Central Journal vol 7 no 1 article 382013
[29] M-C Mong C-Y Chao and M-C Yin ldquoHistidine andcarnosine alleviated hepatic steatosis in mice consumed highsaturated fat dietrdquo European Journal of Pharmacology vol 653no 1ndash3 pp 82ndash88 2011
[30] D Mizuno and M Kawahara ldquoThe molecular mechanisms ofzinc neurotoxicity and the pathogenesis of vascular type seniledementiardquo International Journal of Molecular Sciences vol 14no 11 pp 22067ndash22081 2013
[31] A Asperger C Renner M Menzel R Gebhardt J Meixens-berger and F Gaunitz ldquoIdentification of factors involved inthe anti-tumor activity of carnosine on glioblastomas using aproteomics approachrdquo Cancer Investigation vol 29 no 4 pp272ndash281 2011
[32] L A Bharadwaj G F Davies I J Xavier and N Ovsenek ldquoL-carnosine and verapamil inhibit hypoxia-induced expression ofhypoxia inducible factor (HIF-1120572) in H9c2 cardiomyoblastsrdquoPharmacological Research vol 45 no 3 pp 175ndash181 2002
[33] E Ito-Kato N Suzuki M Maeno et al ldquoEffect of carnosine onrunt-related transcription factor-2core binding factor 120572-1 andSox9 expressions of human periodontal ligament cellsrdquo Journalof Periodontal Research vol 39 no 3 pp 199ndash204 2004
[34] V Calabrese C Colombrita E Guagliano et al ldquoProtectiveeffect of carnosine during nitrosative stress in astroglial cellculturesrdquo Neurochemical Research vol 30 no 6-7 pp 797ndash8072005
[35] K M Soliman M A Hamed and S A Ali ldquoHepatoprotectiveeffect of carnosine on liver biochemical parameters in chronicethanol intoxicated ratrdquoTheMedical Journal of the IslamicWorldAcademy of Sciences vol 16 no 2 pp 77ndash86 2006
[36] D Ikeda S-I Wada C Yoneda H Abe and S WatabeldquoCarnosine stimulates vimentin expression in cultured ratfibroblastsrdquoCell Structure and Function vol 24 no 2 pp 79ndash871999
[37] O E Nekrasova M G Mendez I S Chernoivanenko et alldquoVimentin intermediate filaments modulate the motility ofmitochondriardquoMolecular Biology of the Cell vol 22 no 13 pp2282ndash2289 2011
[38] C Sale B Saunders and R C Harris ldquoEffect of beta-alaninesupplementation onmuscle carnosine concentrations and exer-cise performancerdquoAmino Acids vol 39 no 2 pp 321ndash333 2010
[39] A A Boldyrev and S E Severin ldquoThe histidine-containingdipeptides carnosine and anserine distribution properties andbiological significancerdquoAdvances in Enzyme Regulation vol 30pp 175ndash188 1990
[40] I Everaert A Mooyaart A Baguet et al ldquoVegetarianismfemale gender and increasing age but not CNDP1 genotype
10 BioMed Research International
are associatedwith reducedmuscle carnosine levels in humansrdquoAmino Acids vol 40 no 4 pp 1221ndash1229 2011
[41] V N Korobov N M Doliba and I V Telegus ldquoCarnosine inadaptation to hypobaric hypoxiardquo Biokhimiya vol 58 no 5 pp740ndash744 1993
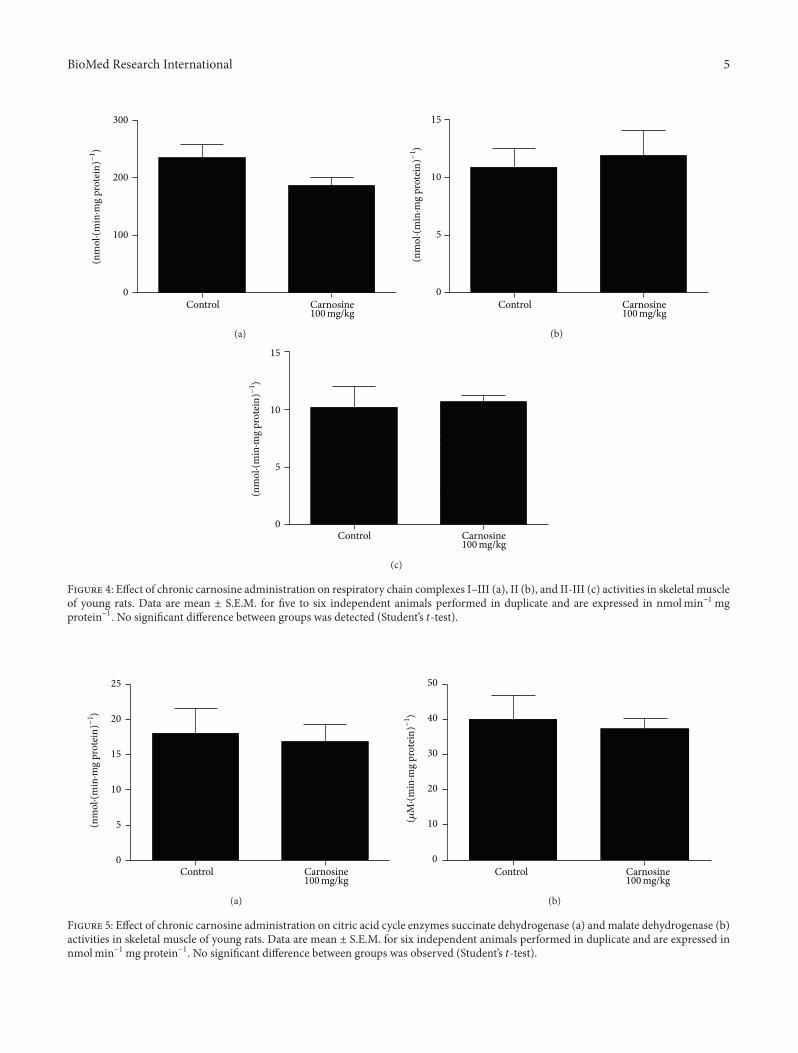
BioMed Research International 5
0
100
200
300
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(a)
0
5
10
15
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(b)
0
5
10
15
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(c)
Figure 4 Effect of chronic carnosine administration on respiratory chain complexes IndashIII (a) II (b) and II-III (c) activities in skeletal muscleof young rats Data are mean plusmn SEM for five to six independent animals performed in duplicate and are expressed in nmolminminus1mgproteinminus1 No significant difference between groups was detected (Studentrsquos 119905-test)
0
5
10
15
20
25
Control Carnosine100 mgkg
(nm
olmiddot(
min
middotmg
prot
ein)
minus1 )
(a)
0
10
20
30
40
50
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
(b)
Figure 5 Effect of chronic carnosine administration on citric acid cycle enzymes succinate dehydrogenase (a) and malate dehydrogenase (b)activities in skeletal muscle of young rats Data are mean plusmn SEM for six independent animals performed in duplicate and are expressed innmolminminus1mg proteinminus1 No significant difference between groups was observed (Studentrsquos 119905-test)
6 BioMed Research International
00
05
10
15
20
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
Figure 6 Effect of chronic carnosine administration on creatinekinase activity in skeletal muscle of young rats Data are mean plusmnSEM for six independent animals performed in duplicate andare expressed in 120583mol creatine minminus1mg proteinminus1 No significantdifference between groups was detected (Studentrsquos 119905-test)
of decrease of complexes IndashIII activities in animals receivingcarnosine as compared to control rats (Figure 4(a))
33 Mitochondrial Biogenesis Transcript abundance of keyfactors involved in mitochondrial biogenesis was evaluatedbyRT-qPCR in skeletalmuscle of animals receiving carnosineacutely (Figure 7) or chronically (Figure 8) As compared tocontrol groups mRNA levels of NRF-1 PGC-1120572 and TFAMin the animals receiving carnosine under either regimen ofadministration were unchanged
4 Discussion
In skeletal muscle tissue of most vertebrates and some inver-tebrates the imidazole dipeptide carnosine is synthesizedfrom histidine and 120573-alanine by the ATP-dependent enzymecarnosine synthetase [20] The biological roles of carnosineand related dipeptides in skeletal muscle remain uncertainalthough it is possible that they act as proton buffering agents[4] Furthermore there are a growing number of evidencesrelated to carnosine effects on metabolism Aside from itsantioxidant activity [21 22] carbonyl-scavenger ability hasbeen assigned to carnosine [23] and other imidazole dipep-tides [24] Many of carbonyl compounds are metabolic toxicby-products and in the bulk of tissues they are detoxifiedby oxidoreductases These enzymes catalyze the oxidation orreduction of aldehydes or its conjugation with lowmolecularweight amines and thiols such as reduced glutathione andimidazole dipeptides [25] Carnosine was also shown toreact with methylglyoxal a toxic metabolic by-product ofglycolysis as well as to inhibit cancer and aging mechanisms[26] and the cellular respiration regulatory complex mTOR[27] and activate the gluconeogenic enzyme fructose-16-bisphosphatase [28]
It was also shown that carnosine intake significantlydiminishes the activity and mRNA expression of malic
enzyme fatty acid synthase 3-hydroxy-3-methylglutarylcoenzyme A (HMG-CoA) reductase sterol regulatoryelement-binding proteins (SREBP-1c) and SREBP-2 inmice that consumed high saturated fat diet [29] Carnosinewas able to inhibit Zn2+-induced death of GT1-7 neuronsthrough the inhibition of GADD34 p8 (endoplasmicreticulum stress-related genes) and Arc (calcium-relatedgene) expression [30] Finally Asperger and colleagues [31]through a proteomics study with glioblastoma cells receivingcarnosine detected 31 proteins expressed differentiallyunder the influence of dipeptide including BCL2-associatedathanogene 2 and von Hippel-Lindau binding protein 1
In the present study we observed a statistically sig-nificant decrease of complexes IndashIII and II activities inanimals receiving carnosine acutely compared to controlgroup rats Additionally we detected no significant differencein complexes II-III citric acid cycle enzymes and creatinekinase activities between groups Finally we were also notable to find any significant alteration of respiratory chaincomplexes citric acid cycle enzymes and creatine kinaseactivities Since several evidences suggest that many of thephysiological effects of carnosine administration initiate attranscriptional level as shown by changes in mRNA levels ofHIF-1120572 [32] runt-related transcription factor-2core bindingfactor alpha-1 (RUNX2Cbf1205721) Sox9 [33] Hsp70 and SOD2[34] mitochondrial biogenesis could explain these findingsNevertheless it was found that mRNA levels of NRF-1 PGC-1120572 and TFAM in skeletal muscle of rats receiving carnosineacutely or chronically were similar to those measured incontrol animals
Our finding showing that the malate dehydrogenaseactivity presented no statistically significant differencebetween groups in both treatments corroborates at least inpart a literature report It was found that a patient with serumcarnosinase deficiency exhibited malate dehydrogenase andlactate dehydrogenase activities within normal limits in liverkidney and spleen specimens despite the fact that patientswith this disorder present high levels of carnosine in theirplasma (carnosinemia) [8]
We also observed that succinate dehydrogenase activityin skeletal muscle of carnosine group was not significantlydifferent from those in control group in animals subjectedto either acute or chronic carnosine administration On theother hand a literature report showed that rats chronicallyintoxicated with ethanol presented reduced activity of somehepatic enzymes including succinate dehydrogenase [35] Itwas also showed in the same study that carnosine load for twoweeks previously or simultaneously with ethanol intoxicationprevents or reverses the toxicity of such alcohol on thisenzyme activity
It should be emphasized that in skeletal muscle of ratsreceiving carnosine acutely a statistically significant decreasein complexes IndashIII and II activities was identified as well asa trend of decrease in complexes II-III activities relatively tocontrol group However rats receiving carnosine chronicallyshowed only a trend of decrease in complexes IndashIII activitiesin skeletal muscle as compared to control group whereas theother complexes activities were not different between groups
BioMed Research International 7
00
05
10
15Re
lativ
e RN
A ex
pres
sion
Control Carnosine100 mgkg
(a)
Relat
ive R
NA
expr
essio
n
00
05
10
15
Control Carnosine100 mgkg
(b)
00
05
10
15
Relat
ive R
NA
expr
essio
n
Control Carnosine100 mgkg
(c)
Figure 7 Effect of acute carnosine administration on relative mRNA expression ofNRF-1 (a) PGC-1120572 (b) and TFAM (c) activities in skeletalmuscle of young rats Results are mean plusmn SEM for four to six independent animals performed in quadruplicate No significant differencebetween groups was detected
The same pattern of alteration was also observed inmalate dehydrogenase activity which was found increasedin rats receiving carnosine acutely and unaltered in animalsreceiving the dipeptide chronically Thus it may be proposedthat skeletal muscle is able to adapt to high carnosinelevels recovering the impaired enzyme activities back tocontrol conditions A putative mechanism involved in suchadaptation is increased mitochondrial synthesis since theenzymes affected by carnosine administration present mito-chondrial localization Considering that carnosine adminis-tration increases vimentin synthesis [36] and that vimentin isa cytoskeleton protein that may be related to mitochondrialmotility and localization [37] it is possible that this processis mediated by increased vimentin synthesis However dataon NRF-1 PGC-1120572 and TFAM mRNA levels suggest thatcarnosine interferes with these enzymatic activities throughmechanisms unrelated to mitochondrial biogenesis but theinfluence of carnosine on mitophagy cannot be ruled out
At present we cannot ascertain the pathophysiologicalrelevance of our findings In this communication we inducedcarnosinemia in the animals by administering carnosineintraperitoneally With respect to this fact it should be men-tioned that carnosine dietary administration elicits increase
of its content in human skeletal muscle [38] The carnosinelevels in this tissue are situated within 30ndash50mmolkg ofwet weight [39] and are prone to alterations according todiet gender and age of subject [40] Additionally in ourstudy carnosine is increased in skeletal muscle of rat suchas occurrence in patients with carnosinemia
5 Conclusion
It is here shown that carnosine administration impairs elec-tron transfer through the mitochondrial respiratory chainin skeletal muscle of young rats without changing mito-chondrial biogenesis-related transcription factors At leastin part this finding does not corroborate some reportswhich indicate that this dipeptide increases the efficiencyand intensity of oxidative phosphorylation in rats adaptedto hypobaric hypoxia [41] The human carnosinase levelsare almost undetectable in plasma of neonates increasingaccording to aging [6] rendering young individuals lessprone to metabolize carnosine In case these findings areconfirmed by further studies and ATP depletion is alsoobserved it is possible that an energy dysfunction secondaryto carnosine accumulationmay aid to explain someofmuscle
8 BioMed Research International
00
05
10
15Re
lativ
e RN
A ex
pres
sion
Control Carnosine100 mgkg
(a)
00
05
10
15
Relat
ive R
NA
expr
essio
n
Control Carnosine100 mgkg
(b)
Control Carnosine100 mgkg
00
05
10
15
Relat
ive R
NA
expr
essio
n
(c)
Figure 8 Effect of chronic carnosine administration on relative mRNA expression of NRF-1 (a) PGC-1120572 (b) and TFAM (c) activities inskeletal muscle of young rats Results are mean plusmn SEM for four to six independent animals performed in quadruplicate No significantdifference between groups was detected
symptoms commonly observed in young patients with serumcarnosinase deficiency
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This research was supported by resources from Universidadedo Extremo Sul Catarinense and Brazilian National Councilfor Scientific and Technological Development (CNPq) LuizaWilges Kist was recipient of fellowships from Coordenacaode Aperfeicoamento de Pessoal de Nıvel Superior (CAPES)
References
[1] L Bonfanti P Peretto S deMarchis andA Fasolo ldquoCarnosine-related dipeptides in the mammalian brainrdquo Progress in Neuro-biology vol 59 no 4 pp 333ndash353 1999
[2] W Gulewitsch and S Amiradzibi ldquoUeber das carnosineine neue organische base des fleischextraktesrdquo Berichte der
Deutschen ChemischenGesellschaft vol 33 no 2 pp 1902ndash19031900
[3] K Nagai and T Suda ldquoRealization of spontaneous healingfunction by carnosinerdquo Methods and Findings in Experimentaland Clinical Pharmacology vol 10 no 8 pp 497ndash507 1988
[4] H Abe ldquoRole of histidine-related compounds as intracellularproton buffering constituents in vertebrate musclerdquo Biochem-istry vol 65 no 7 pp 757ndash765 2000
[5] S Geissler M Zwarg I Knutter F Markwardt and M Brand-sch ldquoThe bioactive dipeptide anserine is transported by humanproton-coupled peptide transportersrdquo FEBS Journal vol 277no 3 pp 790ndash795 2010
[6] J F Lenney R P George and A M Weiss ldquoHuman serumcarnosinase characterization distinction from cellular carnosi-nase and activation by cadmiumrdquo Clinica Chimica Acta vol123 no 3 pp 221ndash231 1982
[7] C R Scriver and K M Gibson ldquoDisorders of 120573- and 120574-aminoacids in free and peptide-linked formsrdquo in The Metabolic andMolecular Bases of Inherit C R Scriver A L Beaudet S Slyand D Valle Eds vol 1 part 5 pp 1349ndash1368 McGraw-HillNew York NY USA 7th edition 1995
[8] W H Murphey D G Lindmark and L I Patchen ldquoSerumcarnosinase deficiency concomitant with mental retardationrdquoPediatric Research vol 7 no 7 pp 601ndash606 1973
BioMed Research International 9
[9] H Lunde O Sjaastad and L Gjessing ldquoHomocarnosinosishypercarnosinuriardquo Journal of Neurochemistry vol 38 no 1 pp242ndash245 1982
[10] T L Perry S Hansen B Tischler R Bunting and K BerryldquoCarnosinemia A new metabolic disorder associated withneurologic disease andmental defectrdquoTheNewEngland Journalof Medicine vol 277 no 23 pp 1219ndash1227 1967
[11] S M Willi Y Zhang J B Hill M C Phelan R C Michaelisand K R Holden ldquoA deletion in the long arm of chromosome18 in a child with serum carnosinase deficiencyrdquo PediatricResearch vol 41 no 2 pp 210ndash213 1997
[12] B E Bonefeld B Elfving and GWegener ldquoReference genes fornormalization a study of rat brain tissuerdquo Synapse vol 62 no4 pp 302ndash309 2008
[13] Q Zhang YWu H Sha et al ldquoEarly exercise affects mitochon-drial transcription factors expression after cerebral ischemia inratsrdquo International Journal of Molecular Sciences vol 13 no 2pp 1670ndash1679 2012
[14] A H V Schapira V M Mann J M Cooper et al ldquoAnatomicand disease specificity of NADH CoQ1 reductase (complex I)deficiency in Parkinsonrsquos diseaserdquo Journal of Neurochemistryvol 55 no 6 pp 2142ndash2145 1990
[15] J C Fischer W Ruitenbeek and J A Berden ldquoDifferentialinvestigation of the capacity of succinate oxidation in humanskeletal musclerdquo Clinica Chimica Acta vol 153 no 1 pp 23ndash261985
[16] B P Hughes ldquoA method for the estimation of serum creatinekinase and its use in comparing creatine kinase and aldolaseactivity in normal and pathological serardquo Clinica Chimica Actavol 7 no 5 pp 597ndash603 1962
[17] P F Schuck G Leipnitz C A J Ribeiro et al ldquoInhibition ofcreatine kinase activity in vitro by ethylmalonic acid in cerebralcortex of young ratsrdquoNeurochemical Research vol 27 no 12 pp1633ndash1639 2002
[18] G B Kitto ldquoIntra- and extramitochondrial malate dehydroge-nases from chicken and tuna heartrdquo Methods in Enzymologyvol 13 pp 106ndash116 1969
[19] O H Lowry N J Rosebrough A L Farr and R J RandallldquoProtein measurement with the Folin phenol reagentrdquo TheJournal of Biological Chemistry vol 193 no 1 pp 265ndash275 1951
[20] J Drozak M Veiga-da-Cunha D Vertommen V Stroobantand E van Schaftingen ldquoMolecular identification of carno-sine synthase as ATP-grasp domain-containing protein 1(ATPGD1)rdquoThe Journal of Biological Chemistry vol 285 no 13pp 9346ndash9356 2010
[21] P J Quinn A A Boldyrev and V E Formazuyk ldquoCarnosineits properties functions and potential therapeutic applicationsrdquoMolecular Aspects of Medicine vol 13 no 5 pp 379ndash444 1992
[22] P S Manhiani J K Northcutt I Han W C Bridges and PL Dawson ldquoAntioxidant activity of carnosine extracted fromvarious poultry tissuesrdquo Poultry Science vol 92 no 2 pp 444ndash453 2013
[23] G Aldini M Orioli G Rossoni et al ldquoThe carbonyl scavengercarnosine ameliorates dyslipidaemia and renal function inZucker obese ratsrdquo Journal of Cellular and Molecular Medicinevol 15 no 6 pp 1339ndash1354 2011
[24] G Vistoli D deMaddis V Straniero et al ldquoExploring the spaceof histidine containing dipeptides in search of novel efficientRCS sequestering agentsrdquo European Journal of Medicinal Chem-istry vol 66 pp 153ndash160 2013
[25] Z Xie S P Baba B R Sweeney and O A Barski ldquoDetoxi-fication of aldehydes by histidine-containing dipeptides fromchemistry to clinical implicationsrdquo Chemico-Biological Interac-tions vol 202 no 1ndash3 pp 288ndash297 2013
[26] A R Hipkiss ldquoAging proteotoxicity mitochondria glycationNAD+ and carnosine possible inter-relationships and resolu-tion of the oxygen paradoxrdquoFrontiers inAgingNeuroscience vol2 article 10 2010
[27] A R Hipkiss ldquoEnergy metabolism proteotoxic stress andage-related dysfunction - Protection by carnosinerdquo MolecularAspects of Medicine vol 32 no 4ndash6 pp 267ndash278 2011
[28] A R Hipkiss S P Cartwright C Bromley S R Gross and RM Bill ldquoCarnosine can understanding its actions on energymetabolism and protein homeostasis inform its therapeuticpotentialrdquo Chemistry Central Journal vol 7 no 1 article 382013
[29] M-C Mong C-Y Chao and M-C Yin ldquoHistidine andcarnosine alleviated hepatic steatosis in mice consumed highsaturated fat dietrdquo European Journal of Pharmacology vol 653no 1ndash3 pp 82ndash88 2011
[30] D Mizuno and M Kawahara ldquoThe molecular mechanisms ofzinc neurotoxicity and the pathogenesis of vascular type seniledementiardquo International Journal of Molecular Sciences vol 14no 11 pp 22067ndash22081 2013
[31] A Asperger C Renner M Menzel R Gebhardt J Meixens-berger and F Gaunitz ldquoIdentification of factors involved inthe anti-tumor activity of carnosine on glioblastomas using aproteomics approachrdquo Cancer Investigation vol 29 no 4 pp272ndash281 2011
[32] L A Bharadwaj G F Davies I J Xavier and N Ovsenek ldquoL-carnosine and verapamil inhibit hypoxia-induced expression ofhypoxia inducible factor (HIF-1120572) in H9c2 cardiomyoblastsrdquoPharmacological Research vol 45 no 3 pp 175ndash181 2002
[33] E Ito-Kato N Suzuki M Maeno et al ldquoEffect of carnosine onrunt-related transcription factor-2core binding factor 120572-1 andSox9 expressions of human periodontal ligament cellsrdquo Journalof Periodontal Research vol 39 no 3 pp 199ndash204 2004
[34] V Calabrese C Colombrita E Guagliano et al ldquoProtectiveeffect of carnosine during nitrosative stress in astroglial cellculturesrdquo Neurochemical Research vol 30 no 6-7 pp 797ndash8072005
[35] K M Soliman M A Hamed and S A Ali ldquoHepatoprotectiveeffect of carnosine on liver biochemical parameters in chronicethanol intoxicated ratrdquoTheMedical Journal of the IslamicWorldAcademy of Sciences vol 16 no 2 pp 77ndash86 2006
[36] D Ikeda S-I Wada C Yoneda H Abe and S WatabeldquoCarnosine stimulates vimentin expression in cultured ratfibroblastsrdquoCell Structure and Function vol 24 no 2 pp 79ndash871999
[37] O E Nekrasova M G Mendez I S Chernoivanenko et alldquoVimentin intermediate filaments modulate the motility ofmitochondriardquoMolecular Biology of the Cell vol 22 no 13 pp2282ndash2289 2011
[38] C Sale B Saunders and R C Harris ldquoEffect of beta-alaninesupplementation onmuscle carnosine concentrations and exer-cise performancerdquoAmino Acids vol 39 no 2 pp 321ndash333 2010
[39] A A Boldyrev and S E Severin ldquoThe histidine-containingdipeptides carnosine and anserine distribution properties andbiological significancerdquoAdvances in Enzyme Regulation vol 30pp 175ndash188 1990
[40] I Everaert A Mooyaart A Baguet et al ldquoVegetarianismfemale gender and increasing age but not CNDP1 genotype
10 BioMed Research International
are associatedwith reducedmuscle carnosine levels in humansrdquoAmino Acids vol 40 no 4 pp 1221ndash1229 2011
[41] V N Korobov N M Doliba and I V Telegus ldquoCarnosine inadaptation to hypobaric hypoxiardquo Biokhimiya vol 58 no 5 pp740ndash744 1993
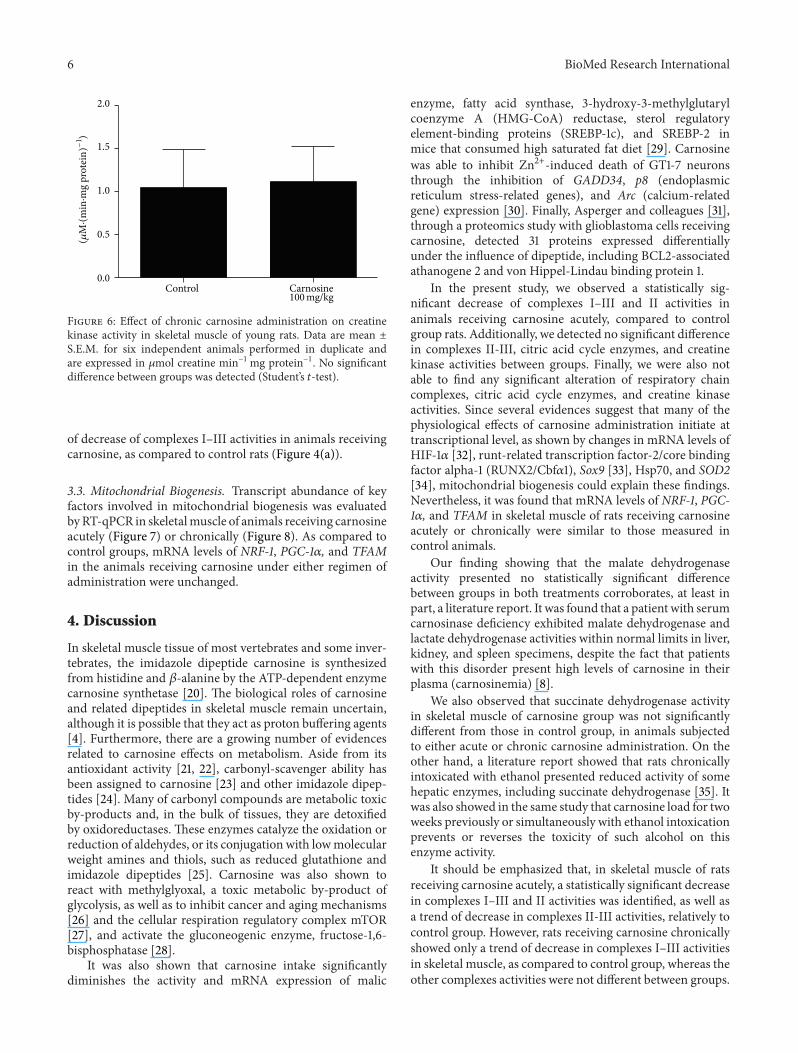
6 BioMed Research International
00
05
10
15
20
Control Carnosine100 mgkg
(120583M
middot(m
inmiddotm
gpr
otei
n)minus
1 )
Figure 6 Effect of chronic carnosine administration on creatinekinase activity in skeletal muscle of young rats Data are mean plusmnSEM for six independent animals performed in duplicate andare expressed in 120583mol creatine minminus1mg proteinminus1 No significantdifference between groups was detected (Studentrsquos 119905-test)
of decrease of complexes IndashIII activities in animals receivingcarnosine as compared to control rats (Figure 4(a))
33 Mitochondrial Biogenesis Transcript abundance of keyfactors involved in mitochondrial biogenesis was evaluatedbyRT-qPCR in skeletalmuscle of animals receiving carnosineacutely (Figure 7) or chronically (Figure 8) As compared tocontrol groups mRNA levels of NRF-1 PGC-1120572 and TFAMin the animals receiving carnosine under either regimen ofadministration were unchanged
4 Discussion
In skeletal muscle tissue of most vertebrates and some inver-tebrates the imidazole dipeptide carnosine is synthesizedfrom histidine and 120573-alanine by the ATP-dependent enzymecarnosine synthetase [20] The biological roles of carnosineand related dipeptides in skeletal muscle remain uncertainalthough it is possible that they act as proton buffering agents[4] Furthermore there are a growing number of evidencesrelated to carnosine effects on metabolism Aside from itsantioxidant activity [21 22] carbonyl-scavenger ability hasbeen assigned to carnosine [23] and other imidazole dipep-tides [24] Many of carbonyl compounds are metabolic toxicby-products and in the bulk of tissues they are detoxifiedby oxidoreductases These enzymes catalyze the oxidation orreduction of aldehydes or its conjugation with lowmolecularweight amines and thiols such as reduced glutathione andimidazole dipeptides [25] Carnosine was also shown toreact with methylglyoxal a toxic metabolic by-product ofglycolysis as well as to inhibit cancer and aging mechanisms[26] and the cellular respiration regulatory complex mTOR[27] and activate the gluconeogenic enzyme fructose-16-bisphosphatase [28]
It was also shown that carnosine intake significantlydiminishes the activity and mRNA expression of malic
enzyme fatty acid synthase 3-hydroxy-3-methylglutarylcoenzyme A (HMG-CoA) reductase sterol regulatoryelement-binding proteins (SREBP-1c) and SREBP-2 inmice that consumed high saturated fat diet [29] Carnosinewas able to inhibit Zn2+-induced death of GT1-7 neuronsthrough the inhibition of GADD34 p8 (endoplasmicreticulum stress-related genes) and Arc (calcium-relatedgene) expression [30] Finally Asperger and colleagues [31]through a proteomics study with glioblastoma cells receivingcarnosine detected 31 proteins expressed differentiallyunder the influence of dipeptide including BCL2-associatedathanogene 2 and von Hippel-Lindau binding protein 1
In the present study we observed a statistically sig-nificant decrease of complexes IndashIII and II activities inanimals receiving carnosine acutely compared to controlgroup rats Additionally we detected no significant differencein complexes II-III citric acid cycle enzymes and creatinekinase activities between groups Finally we were also notable to find any significant alteration of respiratory chaincomplexes citric acid cycle enzymes and creatine kinaseactivities Since several evidences suggest that many of thephysiological effects of carnosine administration initiate attranscriptional level as shown by changes in mRNA levels ofHIF-1120572 [32] runt-related transcription factor-2core bindingfactor alpha-1 (RUNX2Cbf1205721) Sox9 [33] Hsp70 and SOD2[34] mitochondrial biogenesis could explain these findingsNevertheless it was found that mRNA levels of NRF-1 PGC-1120572 and TFAM in skeletal muscle of rats receiving carnosineacutely or chronically were similar to those measured incontrol animals
Our finding showing that the malate dehydrogenaseactivity presented no statistically significant differencebetween groups in both treatments corroborates at least inpart a literature report It was found that a patient with serumcarnosinase deficiency exhibited malate dehydrogenase andlactate dehydrogenase activities within normal limits in liverkidney and spleen specimens despite the fact that patientswith this disorder present high levels of carnosine in theirplasma (carnosinemia) [8]
We also observed that succinate dehydrogenase activityin skeletal muscle of carnosine group was not significantlydifferent from those in control group in animals subjectedto either acute or chronic carnosine administration On theother hand a literature report showed that rats chronicallyintoxicated with ethanol presented reduced activity of somehepatic enzymes including succinate dehydrogenase [35] Itwas also showed in the same study that carnosine load for twoweeks previously or simultaneously with ethanol intoxicationprevents or reverses the toxicity of such alcohol on thisenzyme activity
It should be emphasized that in skeletal muscle of ratsreceiving carnosine acutely a statistically significant decreasein complexes IndashIII and II activities was identified as well asa trend of decrease in complexes II-III activities relatively tocontrol group However rats receiving carnosine chronicallyshowed only a trend of decrease in complexes IndashIII activitiesin skeletal muscle as compared to control group whereas theother complexes activities were not different between groups
BioMed Research International 7
00
05
10
15Re
lativ
e RN
A ex
pres
sion
Control Carnosine100 mgkg
(a)
Relat
ive R
NA
expr
essio
n
00
05
10
15
Control Carnosine100 mgkg
(b)
00
05
10
15
Relat
ive R
NA
expr
essio
n
Control Carnosine100 mgkg
(c)
Figure 7 Effect of acute carnosine administration on relative mRNA expression ofNRF-1 (a) PGC-1120572 (b) and TFAM (c) activities in skeletalmuscle of young rats Results are mean plusmn SEM for four to six independent animals performed in quadruplicate No significant differencebetween groups was detected
The same pattern of alteration was also observed inmalate dehydrogenase activity which was found increasedin rats receiving carnosine acutely and unaltered in animalsreceiving the dipeptide chronically Thus it may be proposedthat skeletal muscle is able to adapt to high carnosinelevels recovering the impaired enzyme activities back tocontrol conditions A putative mechanism involved in suchadaptation is increased mitochondrial synthesis since theenzymes affected by carnosine administration present mito-chondrial localization Considering that carnosine adminis-tration increases vimentin synthesis [36] and that vimentin isa cytoskeleton protein that may be related to mitochondrialmotility and localization [37] it is possible that this processis mediated by increased vimentin synthesis However dataon NRF-1 PGC-1120572 and TFAM mRNA levels suggest thatcarnosine interferes with these enzymatic activities throughmechanisms unrelated to mitochondrial biogenesis but theinfluence of carnosine on mitophagy cannot be ruled out
At present we cannot ascertain the pathophysiologicalrelevance of our findings In this communication we inducedcarnosinemia in the animals by administering carnosineintraperitoneally With respect to this fact it should be men-tioned that carnosine dietary administration elicits increase
of its content in human skeletal muscle [38] The carnosinelevels in this tissue are situated within 30ndash50mmolkg ofwet weight [39] and are prone to alterations according todiet gender and age of subject [40] Additionally in ourstudy carnosine is increased in skeletal muscle of rat suchas occurrence in patients with carnosinemia
5 Conclusion
It is here shown that carnosine administration impairs elec-tron transfer through the mitochondrial respiratory chainin skeletal muscle of young rats without changing mito-chondrial biogenesis-related transcription factors At leastin part this finding does not corroborate some reportswhich indicate that this dipeptide increases the efficiencyand intensity of oxidative phosphorylation in rats adaptedto hypobaric hypoxia [41] The human carnosinase levelsare almost undetectable in plasma of neonates increasingaccording to aging [6] rendering young individuals lessprone to metabolize carnosine In case these findings areconfirmed by further studies and ATP depletion is alsoobserved it is possible that an energy dysfunction secondaryto carnosine accumulationmay aid to explain someofmuscle
8 BioMed Research International
00
05
10
15Re
lativ
e RN
A ex
pres
sion
Control Carnosine100 mgkg
(a)
00
05
10
15
Relat
ive R
NA
expr
essio
n
Control Carnosine100 mgkg
(b)
Control Carnosine100 mgkg
00
05
10
15
Relat
ive R
NA
expr
essio
n
(c)
Figure 8 Effect of chronic carnosine administration on relative mRNA expression of NRF-1 (a) PGC-1120572 (b) and TFAM (c) activities inskeletal muscle of young rats Results are mean plusmn SEM for four to six independent animals performed in quadruplicate No significantdifference between groups was detected
symptoms commonly observed in young patients with serumcarnosinase deficiency
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This research was supported by resources from Universidadedo Extremo Sul Catarinense and Brazilian National Councilfor Scientific and Technological Development (CNPq) LuizaWilges Kist was recipient of fellowships from Coordenacaode Aperfeicoamento de Pessoal de Nıvel Superior (CAPES)
References
[1] L Bonfanti P Peretto S deMarchis andA Fasolo ldquoCarnosine-related dipeptides in the mammalian brainrdquo Progress in Neuro-biology vol 59 no 4 pp 333ndash353 1999
[2] W Gulewitsch and S Amiradzibi ldquoUeber das carnosineine neue organische base des fleischextraktesrdquo Berichte der
Deutschen ChemischenGesellschaft vol 33 no 2 pp 1902ndash19031900
[3] K Nagai and T Suda ldquoRealization of spontaneous healingfunction by carnosinerdquo Methods and Findings in Experimentaland Clinical Pharmacology vol 10 no 8 pp 497ndash507 1988
[4] H Abe ldquoRole of histidine-related compounds as intracellularproton buffering constituents in vertebrate musclerdquo Biochem-istry vol 65 no 7 pp 757ndash765 2000
[5] S Geissler M Zwarg I Knutter F Markwardt and M Brand-sch ldquoThe bioactive dipeptide anserine is transported by humanproton-coupled peptide transportersrdquo FEBS Journal vol 277no 3 pp 790ndash795 2010
[6] J F Lenney R P George and A M Weiss ldquoHuman serumcarnosinase characterization distinction from cellular carnosi-nase and activation by cadmiumrdquo Clinica Chimica Acta vol123 no 3 pp 221ndash231 1982
[7] C R Scriver and K M Gibson ldquoDisorders of 120573- and 120574-aminoacids in free and peptide-linked formsrdquo in The Metabolic andMolecular Bases of Inherit C R Scriver A L Beaudet S Slyand D Valle Eds vol 1 part 5 pp 1349ndash1368 McGraw-HillNew York NY USA 7th edition 1995
[8] W H Murphey D G Lindmark and L I Patchen ldquoSerumcarnosinase deficiency concomitant with mental retardationrdquoPediatric Research vol 7 no 7 pp 601ndash606 1973
BioMed Research International 9
[9] H Lunde O Sjaastad and L Gjessing ldquoHomocarnosinosishypercarnosinuriardquo Journal of Neurochemistry vol 38 no 1 pp242ndash245 1982
[10] T L Perry S Hansen B Tischler R Bunting and K BerryldquoCarnosinemia A new metabolic disorder associated withneurologic disease andmental defectrdquoTheNewEngland Journalof Medicine vol 277 no 23 pp 1219ndash1227 1967
[11] S M Willi Y Zhang J B Hill M C Phelan R C Michaelisand K R Holden ldquoA deletion in the long arm of chromosome18 in a child with serum carnosinase deficiencyrdquo PediatricResearch vol 41 no 2 pp 210ndash213 1997
[12] B E Bonefeld B Elfving and GWegener ldquoReference genes fornormalization a study of rat brain tissuerdquo Synapse vol 62 no4 pp 302ndash309 2008
[13] Q Zhang YWu H Sha et al ldquoEarly exercise affects mitochon-drial transcription factors expression after cerebral ischemia inratsrdquo International Journal of Molecular Sciences vol 13 no 2pp 1670ndash1679 2012
[14] A H V Schapira V M Mann J M Cooper et al ldquoAnatomicand disease specificity of NADH CoQ1 reductase (complex I)deficiency in Parkinsonrsquos diseaserdquo Journal of Neurochemistryvol 55 no 6 pp 2142ndash2145 1990
[15] J C Fischer W Ruitenbeek and J A Berden ldquoDifferentialinvestigation of the capacity of succinate oxidation in humanskeletal musclerdquo Clinica Chimica Acta vol 153 no 1 pp 23ndash261985
[16] B P Hughes ldquoA method for the estimation of serum creatinekinase and its use in comparing creatine kinase and aldolaseactivity in normal and pathological serardquo Clinica Chimica Actavol 7 no 5 pp 597ndash603 1962
[17] P F Schuck G Leipnitz C A J Ribeiro et al ldquoInhibition ofcreatine kinase activity in vitro by ethylmalonic acid in cerebralcortex of young ratsrdquoNeurochemical Research vol 27 no 12 pp1633ndash1639 2002
[18] G B Kitto ldquoIntra- and extramitochondrial malate dehydroge-nases from chicken and tuna heartrdquo Methods in Enzymologyvol 13 pp 106ndash116 1969
[19] O H Lowry N J Rosebrough A L Farr and R J RandallldquoProtein measurement with the Folin phenol reagentrdquo TheJournal of Biological Chemistry vol 193 no 1 pp 265ndash275 1951
[20] J Drozak M Veiga-da-Cunha D Vertommen V Stroobantand E van Schaftingen ldquoMolecular identification of carno-sine synthase as ATP-grasp domain-containing protein 1(ATPGD1)rdquoThe Journal of Biological Chemistry vol 285 no 13pp 9346ndash9356 2010
[21] P J Quinn A A Boldyrev and V E Formazuyk ldquoCarnosineits properties functions and potential therapeutic applicationsrdquoMolecular Aspects of Medicine vol 13 no 5 pp 379ndash444 1992
[22] P S Manhiani J K Northcutt I Han W C Bridges and PL Dawson ldquoAntioxidant activity of carnosine extracted fromvarious poultry tissuesrdquo Poultry Science vol 92 no 2 pp 444ndash453 2013
[23] G Aldini M Orioli G Rossoni et al ldquoThe carbonyl scavengercarnosine ameliorates dyslipidaemia and renal function inZucker obese ratsrdquo Journal of Cellular and Molecular Medicinevol 15 no 6 pp 1339ndash1354 2011
[24] G Vistoli D deMaddis V Straniero et al ldquoExploring the spaceof histidine containing dipeptides in search of novel efficientRCS sequestering agentsrdquo European Journal of Medicinal Chem-istry vol 66 pp 153ndash160 2013
[25] Z Xie S P Baba B R Sweeney and O A Barski ldquoDetoxi-fication of aldehydes by histidine-containing dipeptides fromchemistry to clinical implicationsrdquo Chemico-Biological Interac-tions vol 202 no 1ndash3 pp 288ndash297 2013
[26] A R Hipkiss ldquoAging proteotoxicity mitochondria glycationNAD+ and carnosine possible inter-relationships and resolu-tion of the oxygen paradoxrdquoFrontiers inAgingNeuroscience vol2 article 10 2010
[27] A R Hipkiss ldquoEnergy metabolism proteotoxic stress andage-related dysfunction - Protection by carnosinerdquo MolecularAspects of Medicine vol 32 no 4ndash6 pp 267ndash278 2011
[28] A R Hipkiss S P Cartwright C Bromley S R Gross and RM Bill ldquoCarnosine can understanding its actions on energymetabolism and protein homeostasis inform its therapeuticpotentialrdquo Chemistry Central Journal vol 7 no 1 article 382013
[29] M-C Mong C-Y Chao and M-C Yin ldquoHistidine andcarnosine alleviated hepatic steatosis in mice consumed highsaturated fat dietrdquo European Journal of Pharmacology vol 653no 1ndash3 pp 82ndash88 2011
[30] D Mizuno and M Kawahara ldquoThe molecular mechanisms ofzinc neurotoxicity and the pathogenesis of vascular type seniledementiardquo International Journal of Molecular Sciences vol 14no 11 pp 22067ndash22081 2013
[31] A Asperger C Renner M Menzel R Gebhardt J Meixens-berger and F Gaunitz ldquoIdentification of factors involved inthe anti-tumor activity of carnosine on glioblastomas using aproteomics approachrdquo Cancer Investigation vol 29 no 4 pp272ndash281 2011
[32] L A Bharadwaj G F Davies I J Xavier and N Ovsenek ldquoL-carnosine and verapamil inhibit hypoxia-induced expression ofhypoxia inducible factor (HIF-1120572) in H9c2 cardiomyoblastsrdquoPharmacological Research vol 45 no 3 pp 175ndash181 2002
[33] E Ito-Kato N Suzuki M Maeno et al ldquoEffect of carnosine onrunt-related transcription factor-2core binding factor 120572-1 andSox9 expressions of human periodontal ligament cellsrdquo Journalof Periodontal Research vol 39 no 3 pp 199ndash204 2004
[34] V Calabrese C Colombrita E Guagliano et al ldquoProtectiveeffect of carnosine during nitrosative stress in astroglial cellculturesrdquo Neurochemical Research vol 30 no 6-7 pp 797ndash8072005
[35] K M Soliman M A Hamed and S A Ali ldquoHepatoprotectiveeffect of carnosine on liver biochemical parameters in chronicethanol intoxicated ratrdquoTheMedical Journal of the IslamicWorldAcademy of Sciences vol 16 no 2 pp 77ndash86 2006
[36] D Ikeda S-I Wada C Yoneda H Abe and S WatabeldquoCarnosine stimulates vimentin expression in cultured ratfibroblastsrdquoCell Structure and Function vol 24 no 2 pp 79ndash871999
[37] O E Nekrasova M G Mendez I S Chernoivanenko et alldquoVimentin intermediate filaments modulate the motility ofmitochondriardquoMolecular Biology of the Cell vol 22 no 13 pp2282ndash2289 2011
[38] C Sale B Saunders and R C Harris ldquoEffect of beta-alaninesupplementation onmuscle carnosine concentrations and exer-cise performancerdquoAmino Acids vol 39 no 2 pp 321ndash333 2010
[39] A A Boldyrev and S E Severin ldquoThe histidine-containingdipeptides carnosine and anserine distribution properties andbiological significancerdquoAdvances in Enzyme Regulation vol 30pp 175ndash188 1990
[40] I Everaert A Mooyaart A Baguet et al ldquoVegetarianismfemale gender and increasing age but not CNDP1 genotype
10 BioMed Research International
are associatedwith reducedmuscle carnosine levels in humansrdquoAmino Acids vol 40 no 4 pp 1221ndash1229 2011
[41] V N Korobov N M Doliba and I V Telegus ldquoCarnosine inadaptation to hypobaric hypoxiardquo Biokhimiya vol 58 no 5 pp740ndash744 1993
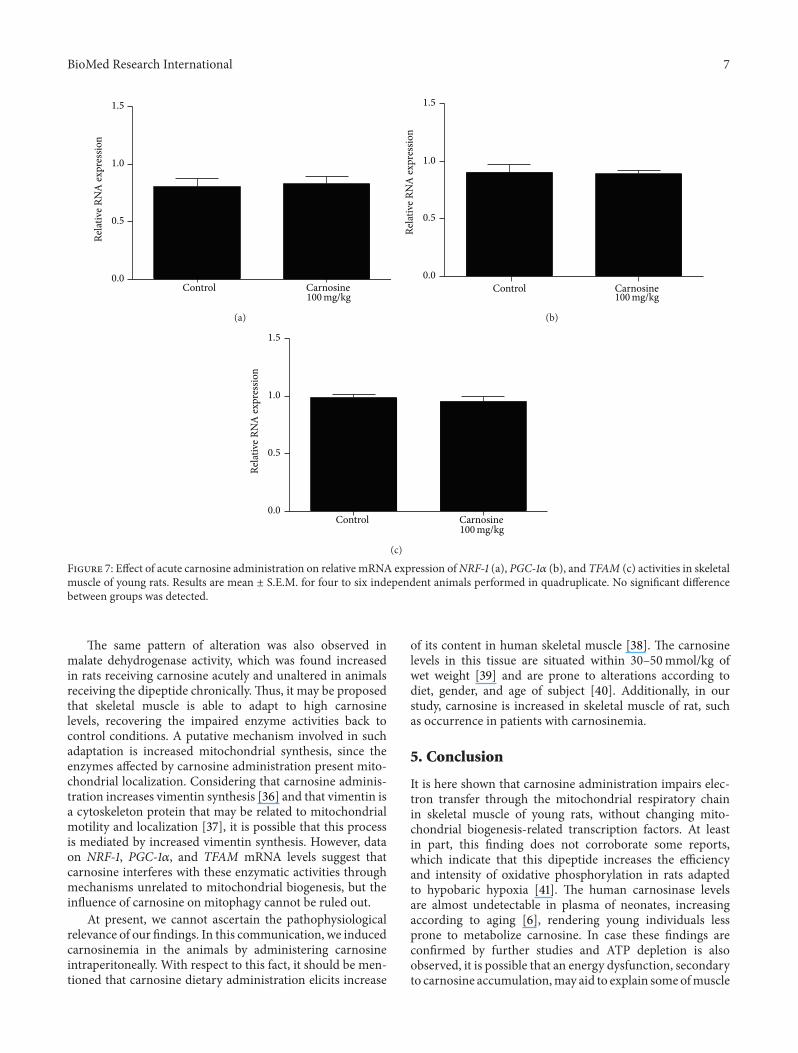
BioMed Research International 7
00
05
10
15Re
lativ
e RN
A ex
pres
sion
Control Carnosine100 mgkg
(a)
Relat
ive R
NA
expr
essio
n
00
05
10
15
Control Carnosine100 mgkg
(b)
00
05
10
15
Relat
ive R
NA
expr
essio
n
Control Carnosine100 mgkg
(c)
Figure 7 Effect of acute carnosine administration on relative mRNA expression ofNRF-1 (a) PGC-1120572 (b) and TFAM (c) activities in skeletalmuscle of young rats Results are mean plusmn SEM for four to six independent animals performed in quadruplicate No significant differencebetween groups was detected
The same pattern of alteration was also observed inmalate dehydrogenase activity which was found increasedin rats receiving carnosine acutely and unaltered in animalsreceiving the dipeptide chronically Thus it may be proposedthat skeletal muscle is able to adapt to high carnosinelevels recovering the impaired enzyme activities back tocontrol conditions A putative mechanism involved in suchadaptation is increased mitochondrial synthesis since theenzymes affected by carnosine administration present mito-chondrial localization Considering that carnosine adminis-tration increases vimentin synthesis [36] and that vimentin isa cytoskeleton protein that may be related to mitochondrialmotility and localization [37] it is possible that this processis mediated by increased vimentin synthesis However dataon NRF-1 PGC-1120572 and TFAM mRNA levels suggest thatcarnosine interferes with these enzymatic activities throughmechanisms unrelated to mitochondrial biogenesis but theinfluence of carnosine on mitophagy cannot be ruled out
At present we cannot ascertain the pathophysiologicalrelevance of our findings In this communication we inducedcarnosinemia in the animals by administering carnosineintraperitoneally With respect to this fact it should be men-tioned that carnosine dietary administration elicits increase
of its content in human skeletal muscle [38] The carnosinelevels in this tissue are situated within 30ndash50mmolkg ofwet weight [39] and are prone to alterations according todiet gender and age of subject [40] Additionally in ourstudy carnosine is increased in skeletal muscle of rat suchas occurrence in patients with carnosinemia
5 Conclusion
It is here shown that carnosine administration impairs elec-tron transfer through the mitochondrial respiratory chainin skeletal muscle of young rats without changing mito-chondrial biogenesis-related transcription factors At leastin part this finding does not corroborate some reportswhich indicate that this dipeptide increases the efficiencyand intensity of oxidative phosphorylation in rats adaptedto hypobaric hypoxia [41] The human carnosinase levelsare almost undetectable in plasma of neonates increasingaccording to aging [6] rendering young individuals lessprone to metabolize carnosine In case these findings areconfirmed by further studies and ATP depletion is alsoobserved it is possible that an energy dysfunction secondaryto carnosine accumulationmay aid to explain someofmuscle
8 BioMed Research International
00
05
10
15Re
lativ
e RN
A ex
pres
sion
Control Carnosine100 mgkg
(a)
00
05
10
15
Relat
ive R
NA
expr
essio
n
Control Carnosine100 mgkg
(b)
Control Carnosine100 mgkg
00
05
10
15
Relat
ive R
NA
expr
essio
n
(c)
Figure 8 Effect of chronic carnosine administration on relative mRNA expression of NRF-1 (a) PGC-1120572 (b) and TFAM (c) activities inskeletal muscle of young rats Results are mean plusmn SEM for four to six independent animals performed in quadruplicate No significantdifference between groups was detected
symptoms commonly observed in young patients with serumcarnosinase deficiency
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This research was supported by resources from Universidadedo Extremo Sul Catarinense and Brazilian National Councilfor Scientific and Technological Development (CNPq) LuizaWilges Kist was recipient of fellowships from Coordenacaode Aperfeicoamento de Pessoal de Nıvel Superior (CAPES)
References
[1] L Bonfanti P Peretto S deMarchis andA Fasolo ldquoCarnosine-related dipeptides in the mammalian brainrdquo Progress in Neuro-biology vol 59 no 4 pp 333ndash353 1999
[2] W Gulewitsch and S Amiradzibi ldquoUeber das carnosineine neue organische base des fleischextraktesrdquo Berichte der
Deutschen ChemischenGesellschaft vol 33 no 2 pp 1902ndash19031900
[3] K Nagai and T Suda ldquoRealization of spontaneous healingfunction by carnosinerdquo Methods and Findings in Experimentaland Clinical Pharmacology vol 10 no 8 pp 497ndash507 1988
[4] H Abe ldquoRole of histidine-related compounds as intracellularproton buffering constituents in vertebrate musclerdquo Biochem-istry vol 65 no 7 pp 757ndash765 2000
[5] S Geissler M Zwarg I Knutter F Markwardt and M Brand-sch ldquoThe bioactive dipeptide anserine is transported by humanproton-coupled peptide transportersrdquo FEBS Journal vol 277no 3 pp 790ndash795 2010
[6] J F Lenney R P George and A M Weiss ldquoHuman serumcarnosinase characterization distinction from cellular carnosi-nase and activation by cadmiumrdquo Clinica Chimica Acta vol123 no 3 pp 221ndash231 1982
[7] C R Scriver and K M Gibson ldquoDisorders of 120573- and 120574-aminoacids in free and peptide-linked formsrdquo in The Metabolic andMolecular Bases of Inherit C R Scriver A L Beaudet S Slyand D Valle Eds vol 1 part 5 pp 1349ndash1368 McGraw-HillNew York NY USA 7th edition 1995
[8] W H Murphey D G Lindmark and L I Patchen ldquoSerumcarnosinase deficiency concomitant with mental retardationrdquoPediatric Research vol 7 no 7 pp 601ndash606 1973
BioMed Research International 9
[9] H Lunde O Sjaastad and L Gjessing ldquoHomocarnosinosishypercarnosinuriardquo Journal of Neurochemistry vol 38 no 1 pp242ndash245 1982
[10] T L Perry S Hansen B Tischler R Bunting and K BerryldquoCarnosinemia A new metabolic disorder associated withneurologic disease andmental defectrdquoTheNewEngland Journalof Medicine vol 277 no 23 pp 1219ndash1227 1967
[11] S M Willi Y Zhang J B Hill M C Phelan R C Michaelisand K R Holden ldquoA deletion in the long arm of chromosome18 in a child with serum carnosinase deficiencyrdquo PediatricResearch vol 41 no 2 pp 210ndash213 1997
[12] B E Bonefeld B Elfving and GWegener ldquoReference genes fornormalization a study of rat brain tissuerdquo Synapse vol 62 no4 pp 302ndash309 2008
[13] Q Zhang YWu H Sha et al ldquoEarly exercise affects mitochon-drial transcription factors expression after cerebral ischemia inratsrdquo International Journal of Molecular Sciences vol 13 no 2pp 1670ndash1679 2012
[14] A H V Schapira V M Mann J M Cooper et al ldquoAnatomicand disease specificity of NADH CoQ1 reductase (complex I)deficiency in Parkinsonrsquos diseaserdquo Journal of Neurochemistryvol 55 no 6 pp 2142ndash2145 1990
[15] J C Fischer W Ruitenbeek and J A Berden ldquoDifferentialinvestigation of the capacity of succinate oxidation in humanskeletal musclerdquo Clinica Chimica Acta vol 153 no 1 pp 23ndash261985
[16] B P Hughes ldquoA method for the estimation of serum creatinekinase and its use in comparing creatine kinase and aldolaseactivity in normal and pathological serardquo Clinica Chimica Actavol 7 no 5 pp 597ndash603 1962
[17] P F Schuck G Leipnitz C A J Ribeiro et al ldquoInhibition ofcreatine kinase activity in vitro by ethylmalonic acid in cerebralcortex of young ratsrdquoNeurochemical Research vol 27 no 12 pp1633ndash1639 2002
[18] G B Kitto ldquoIntra- and extramitochondrial malate dehydroge-nases from chicken and tuna heartrdquo Methods in Enzymologyvol 13 pp 106ndash116 1969
[19] O H Lowry N J Rosebrough A L Farr and R J RandallldquoProtein measurement with the Folin phenol reagentrdquo TheJournal of Biological Chemistry vol 193 no 1 pp 265ndash275 1951
[20] J Drozak M Veiga-da-Cunha D Vertommen V Stroobantand E van Schaftingen ldquoMolecular identification of carno-sine synthase as ATP-grasp domain-containing protein 1(ATPGD1)rdquoThe Journal of Biological Chemistry vol 285 no 13pp 9346ndash9356 2010
[21] P J Quinn A A Boldyrev and V E Formazuyk ldquoCarnosineits properties functions and potential therapeutic applicationsrdquoMolecular Aspects of Medicine vol 13 no 5 pp 379ndash444 1992
[22] P S Manhiani J K Northcutt I Han W C Bridges and PL Dawson ldquoAntioxidant activity of carnosine extracted fromvarious poultry tissuesrdquo Poultry Science vol 92 no 2 pp 444ndash453 2013
[23] G Aldini M Orioli G Rossoni et al ldquoThe carbonyl scavengercarnosine ameliorates dyslipidaemia and renal function inZucker obese ratsrdquo Journal of Cellular and Molecular Medicinevol 15 no 6 pp 1339ndash1354 2011
[24] G Vistoli D deMaddis V Straniero et al ldquoExploring the spaceof histidine containing dipeptides in search of novel efficientRCS sequestering agentsrdquo European Journal of Medicinal Chem-istry vol 66 pp 153ndash160 2013
[25] Z Xie S P Baba B R Sweeney and O A Barski ldquoDetoxi-fication of aldehydes by histidine-containing dipeptides fromchemistry to clinical implicationsrdquo Chemico-Biological Interac-tions vol 202 no 1ndash3 pp 288ndash297 2013
[26] A R Hipkiss ldquoAging proteotoxicity mitochondria glycationNAD+ and carnosine possible inter-relationships and resolu-tion of the oxygen paradoxrdquoFrontiers inAgingNeuroscience vol2 article 10 2010
[27] A R Hipkiss ldquoEnergy metabolism proteotoxic stress andage-related dysfunction - Protection by carnosinerdquo MolecularAspects of Medicine vol 32 no 4ndash6 pp 267ndash278 2011
[28] A R Hipkiss S P Cartwright C Bromley S R Gross and RM Bill ldquoCarnosine can understanding its actions on energymetabolism and protein homeostasis inform its therapeuticpotentialrdquo Chemistry Central Journal vol 7 no 1 article 382013
[29] M-C Mong C-Y Chao and M-C Yin ldquoHistidine andcarnosine alleviated hepatic steatosis in mice consumed highsaturated fat dietrdquo European Journal of Pharmacology vol 653no 1ndash3 pp 82ndash88 2011
[30] D Mizuno and M Kawahara ldquoThe molecular mechanisms ofzinc neurotoxicity and the pathogenesis of vascular type seniledementiardquo International Journal of Molecular Sciences vol 14no 11 pp 22067ndash22081 2013
[31] A Asperger C Renner M Menzel R Gebhardt J Meixens-berger and F Gaunitz ldquoIdentification of factors involved inthe anti-tumor activity of carnosine on glioblastomas using aproteomics approachrdquo Cancer Investigation vol 29 no 4 pp272ndash281 2011
[32] L A Bharadwaj G F Davies I J Xavier and N Ovsenek ldquoL-carnosine and verapamil inhibit hypoxia-induced expression ofhypoxia inducible factor (HIF-1120572) in H9c2 cardiomyoblastsrdquoPharmacological Research vol 45 no 3 pp 175ndash181 2002
[33] E Ito-Kato N Suzuki M Maeno et al ldquoEffect of carnosine onrunt-related transcription factor-2core binding factor 120572-1 andSox9 expressions of human periodontal ligament cellsrdquo Journalof Periodontal Research vol 39 no 3 pp 199ndash204 2004
[34] V Calabrese C Colombrita E Guagliano et al ldquoProtectiveeffect of carnosine during nitrosative stress in astroglial cellculturesrdquo Neurochemical Research vol 30 no 6-7 pp 797ndash8072005
[35] K M Soliman M A Hamed and S A Ali ldquoHepatoprotectiveeffect of carnosine on liver biochemical parameters in chronicethanol intoxicated ratrdquoTheMedical Journal of the IslamicWorldAcademy of Sciences vol 16 no 2 pp 77ndash86 2006
[36] D Ikeda S-I Wada C Yoneda H Abe and S WatabeldquoCarnosine stimulates vimentin expression in cultured ratfibroblastsrdquoCell Structure and Function vol 24 no 2 pp 79ndash871999
[37] O E Nekrasova M G Mendez I S Chernoivanenko et alldquoVimentin intermediate filaments modulate the motility ofmitochondriardquoMolecular Biology of the Cell vol 22 no 13 pp2282ndash2289 2011
[38] C Sale B Saunders and R C Harris ldquoEffect of beta-alaninesupplementation onmuscle carnosine concentrations and exer-cise performancerdquoAmino Acids vol 39 no 2 pp 321ndash333 2010
[39] A A Boldyrev and S E Severin ldquoThe histidine-containingdipeptides carnosine and anserine distribution properties andbiological significancerdquoAdvances in Enzyme Regulation vol 30pp 175ndash188 1990
[40] I Everaert A Mooyaart A Baguet et al ldquoVegetarianismfemale gender and increasing age but not CNDP1 genotype
10 BioMed Research International
are associatedwith reducedmuscle carnosine levels in humansrdquoAmino Acids vol 40 no 4 pp 1221ndash1229 2011
[41] V N Korobov N M Doliba and I V Telegus ldquoCarnosine inadaptation to hypobaric hypoxiardquo Biokhimiya vol 58 no 5 pp740ndash744 1993
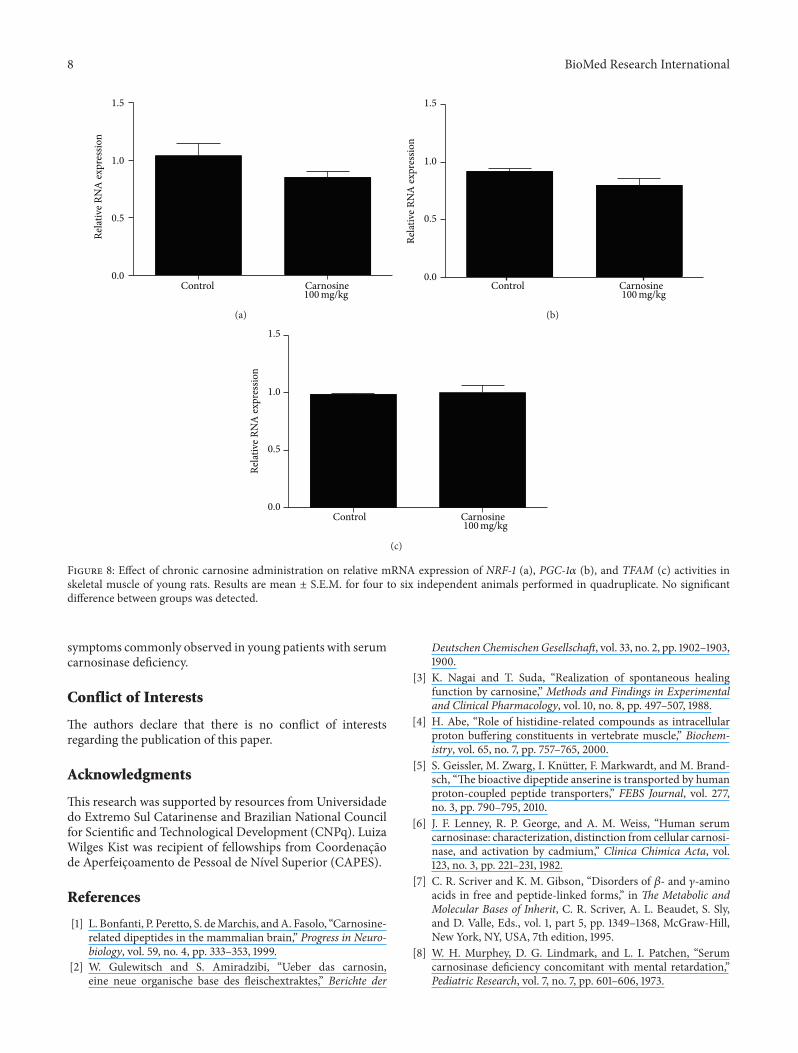
8 BioMed Research International
00
05
10
15Re
lativ
e RN
A ex
pres
sion
Control Carnosine100 mgkg
(a)
00
05
10
15
Relat
ive R
NA
expr
essio
n
Control Carnosine100 mgkg
(b)
Control Carnosine100 mgkg
00
05
10
15
Relat
ive R
NA
expr
essio
n
(c)
Figure 8 Effect of chronic carnosine administration on relative mRNA expression of NRF-1 (a) PGC-1120572 (b) and TFAM (c) activities inskeletal muscle of young rats Results are mean plusmn SEM for four to six independent animals performed in quadruplicate No significantdifference between groups was detected
symptoms commonly observed in young patients with serumcarnosinase deficiency
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This research was supported by resources from Universidadedo Extremo Sul Catarinense and Brazilian National Councilfor Scientific and Technological Development (CNPq) LuizaWilges Kist was recipient of fellowships from Coordenacaode Aperfeicoamento de Pessoal de Nıvel Superior (CAPES)
References
[1] L Bonfanti P Peretto S deMarchis andA Fasolo ldquoCarnosine-related dipeptides in the mammalian brainrdquo Progress in Neuro-biology vol 59 no 4 pp 333ndash353 1999
[2] W Gulewitsch and S Amiradzibi ldquoUeber das carnosineine neue organische base des fleischextraktesrdquo Berichte der
Deutschen ChemischenGesellschaft vol 33 no 2 pp 1902ndash19031900
[3] K Nagai and T Suda ldquoRealization of spontaneous healingfunction by carnosinerdquo Methods and Findings in Experimentaland Clinical Pharmacology vol 10 no 8 pp 497ndash507 1988
[4] H Abe ldquoRole of histidine-related compounds as intracellularproton buffering constituents in vertebrate musclerdquo Biochem-istry vol 65 no 7 pp 757ndash765 2000
[5] S Geissler M Zwarg I Knutter F Markwardt and M Brand-sch ldquoThe bioactive dipeptide anserine is transported by humanproton-coupled peptide transportersrdquo FEBS Journal vol 277no 3 pp 790ndash795 2010
[6] J F Lenney R P George and A M Weiss ldquoHuman serumcarnosinase characterization distinction from cellular carnosi-nase and activation by cadmiumrdquo Clinica Chimica Acta vol123 no 3 pp 221ndash231 1982
[7] C R Scriver and K M Gibson ldquoDisorders of 120573- and 120574-aminoacids in free and peptide-linked formsrdquo in The Metabolic andMolecular Bases of Inherit C R Scriver A L Beaudet S Slyand D Valle Eds vol 1 part 5 pp 1349ndash1368 McGraw-HillNew York NY USA 7th edition 1995
[8] W H Murphey D G Lindmark and L I Patchen ldquoSerumcarnosinase deficiency concomitant with mental retardationrdquoPediatric Research vol 7 no 7 pp 601ndash606 1973
BioMed Research International 9
[9] H Lunde O Sjaastad and L Gjessing ldquoHomocarnosinosishypercarnosinuriardquo Journal of Neurochemistry vol 38 no 1 pp242ndash245 1982
[10] T L Perry S Hansen B Tischler R Bunting and K BerryldquoCarnosinemia A new metabolic disorder associated withneurologic disease andmental defectrdquoTheNewEngland Journalof Medicine vol 277 no 23 pp 1219ndash1227 1967
[11] S M Willi Y Zhang J B Hill M C Phelan R C Michaelisand K R Holden ldquoA deletion in the long arm of chromosome18 in a child with serum carnosinase deficiencyrdquo PediatricResearch vol 41 no 2 pp 210ndash213 1997
[12] B E Bonefeld B Elfving and GWegener ldquoReference genes fornormalization a study of rat brain tissuerdquo Synapse vol 62 no4 pp 302ndash309 2008
[13] Q Zhang YWu H Sha et al ldquoEarly exercise affects mitochon-drial transcription factors expression after cerebral ischemia inratsrdquo International Journal of Molecular Sciences vol 13 no 2pp 1670ndash1679 2012
[14] A H V Schapira V M Mann J M Cooper et al ldquoAnatomicand disease specificity of NADH CoQ1 reductase (complex I)deficiency in Parkinsonrsquos diseaserdquo Journal of Neurochemistryvol 55 no 6 pp 2142ndash2145 1990
[15] J C Fischer W Ruitenbeek and J A Berden ldquoDifferentialinvestigation of the capacity of succinate oxidation in humanskeletal musclerdquo Clinica Chimica Acta vol 153 no 1 pp 23ndash261985
[16] B P Hughes ldquoA method for the estimation of serum creatinekinase and its use in comparing creatine kinase and aldolaseactivity in normal and pathological serardquo Clinica Chimica Actavol 7 no 5 pp 597ndash603 1962
[17] P F Schuck G Leipnitz C A J Ribeiro et al ldquoInhibition ofcreatine kinase activity in vitro by ethylmalonic acid in cerebralcortex of young ratsrdquoNeurochemical Research vol 27 no 12 pp1633ndash1639 2002
[18] G B Kitto ldquoIntra- and extramitochondrial malate dehydroge-nases from chicken and tuna heartrdquo Methods in Enzymologyvol 13 pp 106ndash116 1969
[19] O H Lowry N J Rosebrough A L Farr and R J RandallldquoProtein measurement with the Folin phenol reagentrdquo TheJournal of Biological Chemistry vol 193 no 1 pp 265ndash275 1951
[20] J Drozak M Veiga-da-Cunha D Vertommen V Stroobantand E van Schaftingen ldquoMolecular identification of carno-sine synthase as ATP-grasp domain-containing protein 1(ATPGD1)rdquoThe Journal of Biological Chemistry vol 285 no 13pp 9346ndash9356 2010
[21] P J Quinn A A Boldyrev and V E Formazuyk ldquoCarnosineits properties functions and potential therapeutic applicationsrdquoMolecular Aspects of Medicine vol 13 no 5 pp 379ndash444 1992
[22] P S Manhiani J K Northcutt I Han W C Bridges and PL Dawson ldquoAntioxidant activity of carnosine extracted fromvarious poultry tissuesrdquo Poultry Science vol 92 no 2 pp 444ndash453 2013
[23] G Aldini M Orioli G Rossoni et al ldquoThe carbonyl scavengercarnosine ameliorates dyslipidaemia and renal function inZucker obese ratsrdquo Journal of Cellular and Molecular Medicinevol 15 no 6 pp 1339ndash1354 2011
[24] G Vistoli D deMaddis V Straniero et al ldquoExploring the spaceof histidine containing dipeptides in search of novel efficientRCS sequestering agentsrdquo European Journal of Medicinal Chem-istry vol 66 pp 153ndash160 2013
[25] Z Xie S P Baba B R Sweeney and O A Barski ldquoDetoxi-fication of aldehydes by histidine-containing dipeptides fromchemistry to clinical implicationsrdquo Chemico-Biological Interac-tions vol 202 no 1ndash3 pp 288ndash297 2013
[26] A R Hipkiss ldquoAging proteotoxicity mitochondria glycationNAD+ and carnosine possible inter-relationships and resolu-tion of the oxygen paradoxrdquoFrontiers inAgingNeuroscience vol2 article 10 2010
[27] A R Hipkiss ldquoEnergy metabolism proteotoxic stress andage-related dysfunction - Protection by carnosinerdquo MolecularAspects of Medicine vol 32 no 4ndash6 pp 267ndash278 2011
[28] A R Hipkiss S P Cartwright C Bromley S R Gross and RM Bill ldquoCarnosine can understanding its actions on energymetabolism and protein homeostasis inform its therapeuticpotentialrdquo Chemistry Central Journal vol 7 no 1 article 382013
[29] M-C Mong C-Y Chao and M-C Yin ldquoHistidine andcarnosine alleviated hepatic steatosis in mice consumed highsaturated fat dietrdquo European Journal of Pharmacology vol 653no 1ndash3 pp 82ndash88 2011
[30] D Mizuno and M Kawahara ldquoThe molecular mechanisms ofzinc neurotoxicity and the pathogenesis of vascular type seniledementiardquo International Journal of Molecular Sciences vol 14no 11 pp 22067ndash22081 2013
[31] A Asperger C Renner M Menzel R Gebhardt J Meixens-berger and F Gaunitz ldquoIdentification of factors involved inthe anti-tumor activity of carnosine on glioblastomas using aproteomics approachrdquo Cancer Investigation vol 29 no 4 pp272ndash281 2011
[32] L A Bharadwaj G F Davies I J Xavier and N Ovsenek ldquoL-carnosine and verapamil inhibit hypoxia-induced expression ofhypoxia inducible factor (HIF-1120572) in H9c2 cardiomyoblastsrdquoPharmacological Research vol 45 no 3 pp 175ndash181 2002
[33] E Ito-Kato N Suzuki M Maeno et al ldquoEffect of carnosine onrunt-related transcription factor-2core binding factor 120572-1 andSox9 expressions of human periodontal ligament cellsrdquo Journalof Periodontal Research vol 39 no 3 pp 199ndash204 2004
[34] V Calabrese C Colombrita E Guagliano et al ldquoProtectiveeffect of carnosine during nitrosative stress in astroglial cellculturesrdquo Neurochemical Research vol 30 no 6-7 pp 797ndash8072005
[35] K M Soliman M A Hamed and S A Ali ldquoHepatoprotectiveeffect of carnosine on liver biochemical parameters in chronicethanol intoxicated ratrdquoTheMedical Journal of the IslamicWorldAcademy of Sciences vol 16 no 2 pp 77ndash86 2006
[36] D Ikeda S-I Wada C Yoneda H Abe and S WatabeldquoCarnosine stimulates vimentin expression in cultured ratfibroblastsrdquoCell Structure and Function vol 24 no 2 pp 79ndash871999
[37] O E Nekrasova M G Mendez I S Chernoivanenko et alldquoVimentin intermediate filaments modulate the motility ofmitochondriardquoMolecular Biology of the Cell vol 22 no 13 pp2282ndash2289 2011
[38] C Sale B Saunders and R C Harris ldquoEffect of beta-alaninesupplementation onmuscle carnosine concentrations and exer-cise performancerdquoAmino Acids vol 39 no 2 pp 321ndash333 2010
[39] A A Boldyrev and S E Severin ldquoThe histidine-containingdipeptides carnosine and anserine distribution properties andbiological significancerdquoAdvances in Enzyme Regulation vol 30pp 175ndash188 1990
[40] I Everaert A Mooyaart A Baguet et al ldquoVegetarianismfemale gender and increasing age but not CNDP1 genotype
10 BioMed Research International
are associatedwith reducedmuscle carnosine levels in humansrdquoAmino Acids vol 40 no 4 pp 1221ndash1229 2011
[41] V N Korobov N M Doliba and I V Telegus ldquoCarnosine inadaptation to hypobaric hypoxiardquo Biokhimiya vol 58 no 5 pp740ndash744 1993

BioMed Research International 9
[9] H Lunde O Sjaastad and L Gjessing ldquoHomocarnosinosishypercarnosinuriardquo Journal of Neurochemistry vol 38 no 1 pp242ndash245 1982
[10] T L Perry S Hansen B Tischler R Bunting and K BerryldquoCarnosinemia A new metabolic disorder associated withneurologic disease andmental defectrdquoTheNewEngland Journalof Medicine vol 277 no 23 pp 1219ndash1227 1967
[11] S M Willi Y Zhang J B Hill M C Phelan R C Michaelisand K R Holden ldquoA deletion in the long arm of chromosome18 in a child with serum carnosinase deficiencyrdquo PediatricResearch vol 41 no 2 pp 210ndash213 1997
[12] B E Bonefeld B Elfving and GWegener ldquoReference genes fornormalization a study of rat brain tissuerdquo Synapse vol 62 no4 pp 302ndash309 2008
[13] Q Zhang YWu H Sha et al ldquoEarly exercise affects mitochon-drial transcription factors expression after cerebral ischemia inratsrdquo International Journal of Molecular Sciences vol 13 no 2pp 1670ndash1679 2012
[14] A H V Schapira V M Mann J M Cooper et al ldquoAnatomicand disease specificity of NADH CoQ1 reductase (complex I)deficiency in Parkinsonrsquos diseaserdquo Journal of Neurochemistryvol 55 no 6 pp 2142ndash2145 1990
[15] J C Fischer W Ruitenbeek and J A Berden ldquoDifferentialinvestigation of the capacity of succinate oxidation in humanskeletal musclerdquo Clinica Chimica Acta vol 153 no 1 pp 23ndash261985
[16] B P Hughes ldquoA method for the estimation of serum creatinekinase and its use in comparing creatine kinase and aldolaseactivity in normal and pathological serardquo Clinica Chimica Actavol 7 no 5 pp 597ndash603 1962
[17] P F Schuck G Leipnitz C A J Ribeiro et al ldquoInhibition ofcreatine kinase activity in vitro by ethylmalonic acid in cerebralcortex of young ratsrdquoNeurochemical Research vol 27 no 12 pp1633ndash1639 2002
[18] G B Kitto ldquoIntra- and extramitochondrial malate dehydroge-nases from chicken and tuna heartrdquo Methods in Enzymologyvol 13 pp 106ndash116 1969
[19] O H Lowry N J Rosebrough A L Farr and R J RandallldquoProtein measurement with the Folin phenol reagentrdquo TheJournal of Biological Chemistry vol 193 no 1 pp 265ndash275 1951
[20] J Drozak M Veiga-da-Cunha D Vertommen V Stroobantand E van Schaftingen ldquoMolecular identification of carno-sine synthase as ATP-grasp domain-containing protein 1(ATPGD1)rdquoThe Journal of Biological Chemistry vol 285 no 13pp 9346ndash9356 2010
[21] P J Quinn A A Boldyrev and V E Formazuyk ldquoCarnosineits properties functions and potential therapeutic applicationsrdquoMolecular Aspects of Medicine vol 13 no 5 pp 379ndash444 1992
[22] P S Manhiani J K Northcutt I Han W C Bridges and PL Dawson ldquoAntioxidant activity of carnosine extracted fromvarious poultry tissuesrdquo Poultry Science vol 92 no 2 pp 444ndash453 2013
[23] G Aldini M Orioli G Rossoni et al ldquoThe carbonyl scavengercarnosine ameliorates dyslipidaemia and renal function inZucker obese ratsrdquo Journal of Cellular and Molecular Medicinevol 15 no 6 pp 1339ndash1354 2011
[24] G Vistoli D deMaddis V Straniero et al ldquoExploring the spaceof histidine containing dipeptides in search of novel efficientRCS sequestering agentsrdquo European Journal of Medicinal Chem-istry vol 66 pp 153ndash160 2013
[25] Z Xie S P Baba B R Sweeney and O A Barski ldquoDetoxi-fication of aldehydes by histidine-containing dipeptides fromchemistry to clinical implicationsrdquo Chemico-Biological Interac-tions vol 202 no 1ndash3 pp 288ndash297 2013
[26] A R Hipkiss ldquoAging proteotoxicity mitochondria glycationNAD+ and carnosine possible inter-relationships and resolu-tion of the oxygen paradoxrdquoFrontiers inAgingNeuroscience vol2 article 10 2010
[27] A R Hipkiss ldquoEnergy metabolism proteotoxic stress andage-related dysfunction - Protection by carnosinerdquo MolecularAspects of Medicine vol 32 no 4ndash6 pp 267ndash278 2011
[28] A R Hipkiss S P Cartwright C Bromley S R Gross and RM Bill ldquoCarnosine can understanding its actions on energymetabolism and protein homeostasis inform its therapeuticpotentialrdquo Chemistry Central Journal vol 7 no 1 article 382013
[29] M-C Mong C-Y Chao and M-C Yin ldquoHistidine andcarnosine alleviated hepatic steatosis in mice consumed highsaturated fat dietrdquo European Journal of Pharmacology vol 653no 1ndash3 pp 82ndash88 2011
[30] D Mizuno and M Kawahara ldquoThe molecular mechanisms ofzinc neurotoxicity and the pathogenesis of vascular type seniledementiardquo International Journal of Molecular Sciences vol 14no 11 pp 22067ndash22081 2013
[31] A Asperger C Renner M Menzel R Gebhardt J Meixens-berger and F Gaunitz ldquoIdentification of factors involved inthe anti-tumor activity of carnosine on glioblastomas using aproteomics approachrdquo Cancer Investigation vol 29 no 4 pp272ndash281 2011
[32] L A Bharadwaj G F Davies I J Xavier and N Ovsenek ldquoL-carnosine and verapamil inhibit hypoxia-induced expression ofhypoxia inducible factor (HIF-1120572) in H9c2 cardiomyoblastsrdquoPharmacological Research vol 45 no 3 pp 175ndash181 2002
[33] E Ito-Kato N Suzuki M Maeno et al ldquoEffect of carnosine onrunt-related transcription factor-2core binding factor 120572-1 andSox9 expressions of human periodontal ligament cellsrdquo Journalof Periodontal Research vol 39 no 3 pp 199ndash204 2004
[34] V Calabrese C Colombrita E Guagliano et al ldquoProtectiveeffect of carnosine during nitrosative stress in astroglial cellculturesrdquo Neurochemical Research vol 30 no 6-7 pp 797ndash8072005
[35] K M Soliman M A Hamed and S A Ali ldquoHepatoprotectiveeffect of carnosine on liver biochemical parameters in chronicethanol intoxicated ratrdquoTheMedical Journal of the IslamicWorldAcademy of Sciences vol 16 no 2 pp 77ndash86 2006
[36] D Ikeda S-I Wada C Yoneda H Abe and S WatabeldquoCarnosine stimulates vimentin expression in cultured ratfibroblastsrdquoCell Structure and Function vol 24 no 2 pp 79ndash871999
[37] O E Nekrasova M G Mendez I S Chernoivanenko et alldquoVimentin intermediate filaments modulate the motility ofmitochondriardquoMolecular Biology of the Cell vol 22 no 13 pp2282ndash2289 2011
[38] C Sale B Saunders and R C Harris ldquoEffect of beta-alaninesupplementation onmuscle carnosine concentrations and exer-cise performancerdquoAmino Acids vol 39 no 2 pp 321ndash333 2010
[39] A A Boldyrev and S E Severin ldquoThe histidine-containingdipeptides carnosine and anserine distribution properties andbiological significancerdquoAdvances in Enzyme Regulation vol 30pp 175ndash188 1990
[40] I Everaert A Mooyaart A Baguet et al ldquoVegetarianismfemale gender and increasing age but not CNDP1 genotype
10 BioMed Research International
are associatedwith reducedmuscle carnosine levels in humansrdquoAmino Acids vol 40 no 4 pp 1221ndash1229 2011
[41] V N Korobov N M Doliba and I V Telegus ldquoCarnosine inadaptation to hypobaric hypoxiardquo Biokhimiya vol 58 no 5 pp740ndash744 1993

10 BioMed Research International
are associatedwith reducedmuscle carnosine levels in humansrdquoAmino Acids vol 40 no 4 pp 1221ndash1229 2011
[41] V N Korobov N M Doliba and I V Telegus ldquoCarnosine inadaptation to hypobaric hypoxiardquo Biokhimiya vol 58 no 5 pp740ndash744 1993
Related Documents











