doi:10.1182/blood-2004-11-4272 Prepublished online February 10, 2005; and Magnar Bjoras Pal Aukrust, Luisa Luna, Thor Ueland, Rune F Johansen, Fredrik Muller, Stig S Froland, Erling C Seeberg in CD4+ T cells of HIV-infected patients Impaired base excision repair and accumulation of oxidative base lesions (5019 articles) Immunobiology Articles on similar topics can be found in the following Blood collections http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requests Information about reproducing this article in parts or in its entirety may be found online at: http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprints Information about ordering reprints may be found online at: http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtml Information about subscriptions and ASH membership may be found online at: digital object identifier (DOIs) and date of initial publication. the indexed by PubMed from initial publication. Citations to Advance online articles must include final publication). Advance online articles are citable and establish publication priority; they are appeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet Copyright 2011 by The American Society of Hematology; all rights reserved. 20036. the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.org From

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

doi:10.1182/blood-2004-11-4272Prepublished online February 10, 2005;
and Magnar BjorasPal Aukrust, Luisa Luna, Thor Ueland, Rune F Johansen, Fredrik Muller, Stig S Froland, Erling C Seeberg in CD4+ T cells of HIV-infected patientsImpaired base excision repair and accumulation of oxidative base lesions
(5019 articles)Immunobiology �Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
digital object identifier (DOIs) and date of initial publication. theindexed by PubMed from initial publication. Citations to Advance online articles must include
final publication). Advance online articles are citable and establish publication priority; they areappeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet
Copyright 2011 by The American Society of Hematology; all rights reserved.20036.the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

1
Regular manuscript, Immunobiology
Impaired base excision repair and accumulation of oxidative base
lesions in CD4+ T cells of HIV-infected patients
Running head: Aukrust et al. Repair of oxidative DNA damage in HIV infection
Pål Aukrust1,2, Luisa Luna4,5, Thor Ueland2,3, Rune F. Johansen4,5, Fredrik Müller2, Stig
S. Frøland1,2, Erling C. Seeberg4,5, Magnar Bjørås4,5
1Section of Clinical Immunology and Infectious Diseases, 2Research Institute for Internal
Medicine, 3Section of Endocrinology, Medical Department, 4Department of Molecular
Biology, Institute of Medical Microbiology, 5Centre for Molecular Biology and
Neuroscience, University of Oslo, The National Hospital, N-0027, Oslo, Norway.
Total word counts: 3700
Abstract word counts: 199
Corresponding authors:
Pål Aukrust, Section of Clinical Immunology and Infectious Diseases, Medical
Department, Rikshospitalet, N-0027 Oslo, Norway.
Phone: 47-23070000, Fax: 47-23073630, e-mail: [email protected].
Magnar Bjørås, Department of Molecular Biology, Institute of Medical Microbiology,
University of Oslo, The National Hospital, N-0027, Oslo, Norway.
Phone: 47-23074060, Fax: 47-23074061, e-mail: [email protected]
This work was supported by the Norwegian Cancer Society and the Research Council of
Norway, Anders Jahre´s Foundation, Medinnova Foundation and Odd Kåre Rabben´s
Memorial Fund for AIDS Research.
Blood First Edition Paper, prepublished online February 10, 2005; DOI 10.1182/blood-2004-11-4272
Copyright © 2005 American Society of Hematology
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

2
Abstract
Several studies have reported enhanced oxidative stress in HIV infection. An important
pathophysiological consequence of increased oxidative stress is endogenous DNA
damage, and the base excision repair pathway is the most important mechanism to
withstand such deleterious effects. To investigate the role of base excision repair in HIV
infection we examined 8-oxoguanine (8-oxoG) levels, as a marker of oxidative DNA
damage, and DNA glycosylase activities in CD4+ and CD8+ T cells from HIV-infected
patients and controls. These results showed that HIV-infected patients, particularly those
with advanced disease, were characterized by increased accumulation of 8oxoG in CD4+
T cells accompanied by a marked decline in DNA glycosylase activity for repair of
oxidative base lesions in these cells. In contrast, CD8+ T cells from HIV-infected
patients, with 8-oxoG levels similar to those in healthy controls, showed enhanced
capacity to repair oxidative DNA damage. Finally, highly active antiretroviral therapy
induced increased glycosylase activity in CD4+ T cells accompanied by normalization of
8-oxoG levels. This imbalance between accumulation of oxidative DNA damage and the
capacity to repair such lesions in CD4+ T cells may represent a previous unrecognized
mechanism involved in the numerical and functional impairment of CD4+ T cells in HIV
infection.
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

3
Introduction
Highly reactive oxygen species (ROS) are formed as by-products during a variety of
biochemical reactions and this steady-state formation of pro-oxidants are normally
balanced by a similar rate of consumption by antioxidants. Oxidative stress results from
imbalance between formation and neutralization of pro-oxidants resulting in
accumulation of pro-oxidants with potential harmful consequences.1 Enhanced oxidative
stress has been implicated in the pathogenesis of several clinical disorders such as heart
and brain ischemic diseases, neurodegenerative disorders, various autoimmune diseases
and seems to be involved in carcinogenesis and aging.1-5 Several lines of evidence
suggest that enhanced oxidative stress also plays a pathogenic role in human
immunodeficiency virus (HIV) infection. Thus, a number of reports have found impaired
antioxidant defence, particularly manifested by disturbed glutathione metabolism in HIV-
infected patients.5-9 Moreover, enhanced oxidative stress may be involved in the
pathogenesis of impaired T cell responsiveness and enhanced T cell apoptosis during
HIV infection, and may also play a role in the development of certain HIV-related
clinical disorders such as malignancies and HIV-related encephalopathy.5,10,11
An important pathophysiological consequence of increased intracellular oxidative
stress is endogenous DNA damage.12 Several types of oxidative DNA lesions have been
reported, including strand breaks, baseless sugars (AP-sites) and oxidized base residues,
with 7,8-dihydro-8-oxoguanine (8-oxoG) and 5-hydroxycytosine (5-ohC) representing
the most frequent mutagenic base lesions.13-15 DNA repair mechanisms have evolved
specifically to counteract the biological effects of DNA damage. Of particular importance
for removal of oxidative damage is the base excision repair pathway, which is initiated by
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

4
the action of DNA glycosylases removing different types of modified bases by cleavage
of the N-glycosylic bond.16,17 Five DNA glycosylases for removal of oxidative base
residues, hNth1, hOgg1, Neil1, Neil2 and Neil3 (hFpg2) have been cloned and
characterized in human cells.18-28 hNth1, Neil1 and Neil2 remove oxidized pyrimidines
such as 5-ohC, whereas hOgg1 is removing oxidized purines such as 8-oxoG. However,
Neil1 appear to be a backup function for hOgg1 for removal of 8-oxoG. All five enzymes
have been shown to remove cytotoxic imidazole-ring fragmented formamidopyrimidine
(faPy) residues which represent blocks to replication.
Based on the potential pathogenic role of enhanced oxidative stress in HIV
infection we examined 8-oxoG levels, as a marker of oxidative DNA damage, in relation
to DNA glycosylase activity for repair of oxidative damage in T cell subsets from HIV-
infected patients and healthy controls. We found increased level of 8-oxoG in DNA in
CD4+ T cells from HIV-infected patients, along with a reduced capacity for repair of
oxidized base residues in these cells.
Materials and methods
Patients and controls
Fifteen HIV-infected patients were consecutively recruited for the study (Table 1). The
patients were clinically classified according to the revised criteria from Center for
Disease Control and Prevention (CDC) in AIDS patients (CDC group C, n=7) and non-
AIDS patients (CDC group A+B, n=8). Patients with ongoing acute or exacerbation of
chronic infection at the time of blood collection were not included. Eleven of the patients
received antiretroviral therapy with nucleoside analog(s), but except for one sub-study
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

5
(see below), none received highly active antiretroviral therapy (HAART). None had
initiated or changed therapy during the last 5 months. None of the patients had abnormal
liver or kidney function, were abusing drugs or alcohol or were taking any regular
medication except for nucleoside analog(s) as described above. Controls in the study
were 13 healthy HIV-seronegative sex- and age-matched healthy blood donors (Table 1).
Approval was obtained from the National Hospital in Oslo institutional review board for
these studies. Informed consent was provided according to the Declaration of Helsinki.
Isolation of cells
Peripheral blood mononuclear cells (PBMC) were obtained from heparinized blood by
Isopaque-Ficoll (Lymphoprep; Nycomed, Oslo, Norway) gradient centrifugation within
45 minutes, and further positive selection of CD4+ and CD8+ T cells was done at 4oC as
previously described.7 Briefly, PBMC were mixed with Dynabeads (Dynal, Oslo,
Norway) coated with appropriate antibodies (anti-CD4, Dynabeads M-450 CD4; anti-
CD8, Dynabeads M-450 CD8) in a cell-to-bead ratio of 1:10. The mixture was incubated
in a test tube on a rocking platform for 30 minutes and rosetting cells isolated by
application of a magnet. After five consecutive washes in cold phosphate-buffered
saline/bovine serum albumin (Calbiochem, La Jolla, CA), the positively selected cells
(cell pellet) were immediately stored in liquid nitrogen. Storage of cells with
immunomagnetic beads did not influence glycosylase or 8-oxoG levels. The purity of cell
populations was >98% as assessed by staining of cytospin preparations of positively
selected cells by the alkaline phosphatase-antialkaline phosphatase procedure.7
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

6
Preparation of nuclear DNA and analyses of 8-oxoG by high-performance liquid
chromatography-electrochemical detection (HPLC-ECD)
Extraction of DNA and hydrolyses to nucleosides by nucleaseP1 and alkaline phosphatase
were performed as described elsewhere.29 To reduce oxidation during the preparation of
DNA, TEMPO (2,2,6,6-tetramethylpiperidine-N-oxyl) was added to all solutions at 100
µM immediately before use. 8-Hydroxy-2'-deoxyguanosine and 2'-deoxyguanosine were
separated by HPLC and analyzed by ECD (+300 mV) and UV light (290 nm). Results
were expressed as the ratio of 8-oxoG/106 bp in each DNA sample.
Preparation of cellular extracts
Cell pellets were resuspended in 25 µl 84 % sucrose/10 mM ethyleneglycol tetraacetic
acid. After 10 minutes incubation on ice the cells were frozen and thawed three times
with subsequent centrifugation (15000g for 15 minutes) to remove cell debris.
Supernatant was stored at –20°C.
Assays for DNA glycosylase activity
All enzyme activities were assayed in a reaction buffer containing 70 mM
morpholinopropanesulfonic acid (MOPS), pH 7.5, 1 mM ethylenediamide tetra-acetic
acid (EDTA), 5% glycerol and 1 mM dithiothreitol, and the mixtures were incubated at
37oC for 30 minutes. For analyzing faPy removal N-[3H]methyl-N´-nitrosourea
(18Ci/mmol) was used to prepare poly(dG-dC) DNA containing faPy residues (5000
dpm/µg DNA).30 FaPy DNA glycosylase activity was measured in a total volume of 50
µl containing 0.4 µg faPy-DNA substrate. For analyzing DNA glycosylase activity for
repair of alkylating damage the cell extracts were mixed with 0.3 µg of [methyl 3H]
methylated DNA (containing 5-10 pmol of methylated bases) in a total volume of 50 µl.
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

7
Assays for cleavage of 8-oxoguanine and 5-hydroxycytosine containing DNA.
Duplex DNA containing a single 8-oxoG residue at position 10 (5’-ATCACCGGC[8-
oxoG]CCACACGAGCTG) opposite C and duplex DNA containing a single 5-OHC
residue at position 22 (AATTGCGATCTAGCTCGCCAG [5OHC]AGCGACCTTA
TCTGATGA) were purified by 20% non denaturing polyacrylamide gel electrophoresis.
For all substrates, the 5’ends were 32P-labeled by T4 polynucleotide kinase and [γ-
32P]adenosine triphosphate (3000Ci/mmol; Amersham, Aylesbury, UK). The reaction
mixtures contained 50 fmol substrates and protein extracts as indicated in a total volume
of 10 µl. After incubation at 37°C for 30 minutes, the reaction products were separated on
20 % polyacrylamide/7 M urea denaturing gels (Hydrolink, FMC, AT Biochem, Malvern,
PA) with 1xTris-Borate-EDTA and radiolabeled fragments finally visualized by
PhosphorImaging [Molecular Dynamics (model 445 SI), Sunnyvale, CA].
Miscellaneous
HIV RNA levels were measured in EDTA plasma by quantitative reverse PCR (Amplicor
HIV Monitor, Roche Diagnostic Systems, Brancburg, NY; detection limit 50 copies/mL).
Plasma levels of tumor necrosis factor (TNF)α were measured by enzyme immunoassay
(BioSource, Camarillo, CA).
Statistical analysis
When comparing three groups of individuals, the Kruskal-Wallis test was used a priori.
If a significant difference was found, Mann-Whitney U test (two-tailed) was used to
determine the differences between each pair of groups. For comparisons within the same
individuals, the Wilcoxon matched pairs test was used. Coefficients of correlation were
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

8
calculated by the Spearman´s rank test. If not otherwise quoted, data are given as medians
and 25th-75th percentiles. P-values (two-sided) are considered significant when <0.05.
Results
Accumulation of 8-oxoG in the DNA of CD4+ and CD8+ T cells from HIV-infected
patients
We have previously reported markedly disturbed intracellular redox balance in CD4+ T
cells from HIV-infected patients.7 To examine if such disturbances were associated with
enhanced oxidative DNA damage, we analysed 8-oxoG levels in genomic DNA of CD4+
and CD8+ T cells from 5 AIDS patients, 4 non-AIDS HIV-infected patients and 7 healthy
controls. As shown in Figure 1, CD4+ T cells from HIV-infected patients were
characterized by significantly raised 8-oxoG contents, with particularly high levels in
AIDS patients (~6-fold increase). In contrast, 8-oxoG levels were similar in CD8+ T cells
from HIV-infected patients and controls (Figure 1). In fact, rather than increased, the 8-
oxoG level in the CD8 subset tended to decrease in the non-AIDS patients compared with
healthy controls, although the difference did not reach statistical significance (p=0.07,
Figure 1). In HIV-infected patients, 8-oxoG levels in CD4+ T cells were inversely
correlated with numbers of HIV RNA copies (r=-0.77, p<0.03) and positively correlated
with TNFα (r= 0.70, p<0.05) levels in plasma.
DNA glycosylase activity in CD4+ and CD8+ T cells during HIV infection
We next analysed if accumulation of 8-oxoG in CD4+ T cells was accompanied by any
alterations in DNA glycosylase activity for repair of oxidative damage. FaPy is a
common substrate for DNA glycosylases involved in repair of oxidative DNA lesions and
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

9
removal of faPy was measured in CD4+ and CD8+ T cells from 15 HIV-infected patients
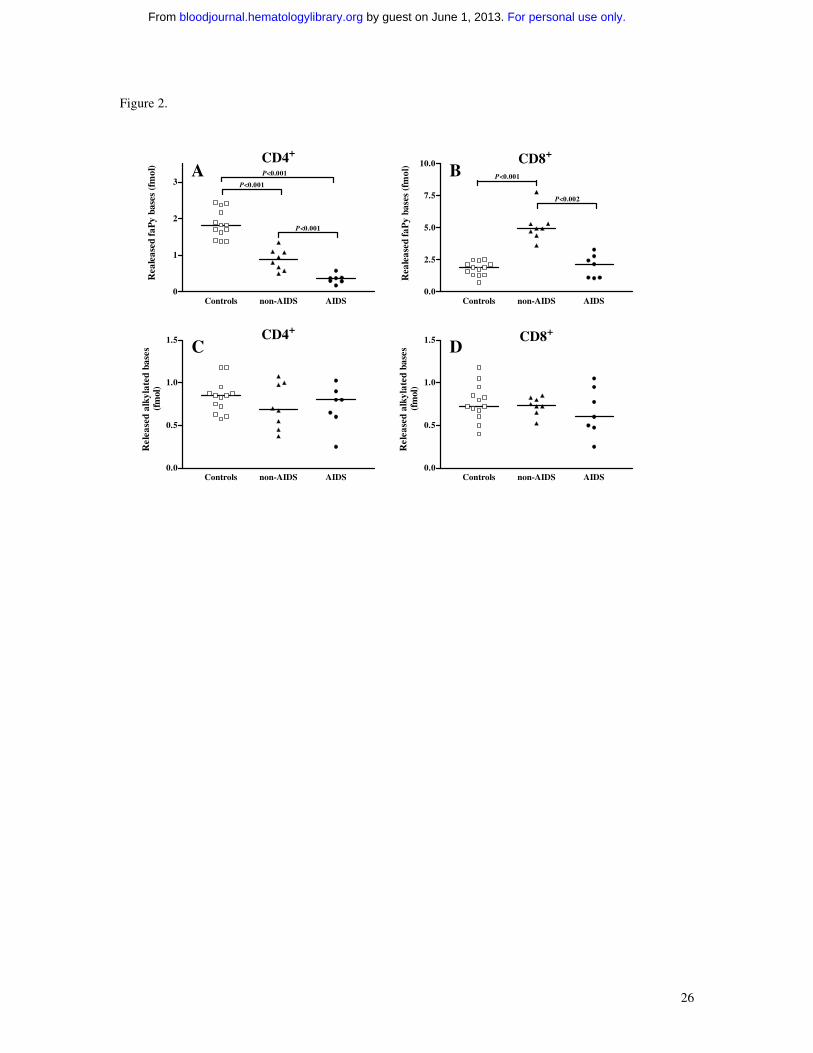
and 13 healthy controls. Notably, while a slight but significant decrease in removal of
faPy was found in CD4+ T cells from non-AIDS patients, CD4+ T cells in AIDS patients
had profoundly decreased faPy activity reaching only ~20% of activity in healthy
controls (Figure 2). In contrast, faPy excision in CD8+ T cells were either markedly
raised (~3.5-fold increase, non-AIDS patients) or similar (AIDS patients) to levels in
healthy controls (Figure 2). In HIV-infected patients, the decrease in faPy activity in
CD4+ T cells was inversely correlated with numbers of HIV RNA copies (r=-0.53,
p<0.05) and positively correlated with TNFα (r=0.61, p<0.01) levels in plasma. In
contrast to faPy excision, DNA glycosylase activity for repair of alkylating damage
showed no significant differences in these T cell subsets between HIV-infected patients
and controls (Figure 2), suggesting a specific down-regulation of DNA glycosylases
repairing oxidative DNA damage in HIV-infected patients.
Levels of DNA glycosylase activity for repair of 8-oxoG and 5-ohC in T cell subsets
during HIV infection
While the assay for measuring DNA glycosylase activity for repair of alkylating damage
is specific for the human Aag DNA glycosylase, the faPy assay may detect activity of
five different DNA glycosylases for repair of oxidative damage; hOgg1, hNth1, Neil1,
Neil2 and Neil3. We therefore examined removal of 8-oxoG and 5-ohC to further
distinguish between these DNA glycosylases. Notably, the removal of 8-oxoG in CD4+ T
cells of AIDS patients showed only ~15% of the activity in CD4+ T cells from healthy
controls suggesting a decrease in the major 8-oxoG DNA glycosylase activity (i.e.,
hOgg1)(Figure 3). Excision of 5-ohC in CD4+ T cells was also decreased in AIDS
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

10
patients (~70% of activity in healthy controls) suggesting a moderate reduction also in
one or more of the three glycosylases reported to remove 5-ohC (hNth1, Neil1 and
Neil2). Furthermore and in accordance with our findings for faPy activity, CD8+ T cells
from non-AIDS patients showed a 3-fold increase in 8-oxoG removal and a 4-fold
increase in 5ohC removal comparing CD8+ T cells from healthy controls, indicating an
enhancement of hOgg1 activity and an up-regulation of one or several of the DNA
glycosylases excising oxidized pyrimidines (hNth1, Neil1 and Neil2). In contrast,
although CD8+ T cells from AIDS patients showed no changes in faPy removal, we
found a 3-fold increase in 5-ohC excision comparing healthy controls (Figure 3). This
could indicate a DNA glycosylase activity with affinity for 5-ohC but not faPy. The 8-
oxoG activity in CD8+ T cells from AIDS patients was not altered (Figure 3).
Except for 11 patients who were receiving nucleoside analog(s) (see Methods),
none of the patients were taken any regular medications. Although these 11 patients did
not differ from the other patients with regard to the measured parameters (data not
shown), we can not exclude some influence from the use of these medications.
DNA glycosylase activity during HAART
HAART strongly reduces levels of plasma HIV RNA with concomitant increases in T
cell counts and beneficial effects on clinical symptoms and mortality.31 To further
examine the relationship between HIV infection, oxidative DNA damage and DNA
glycosylases, we examined DNA glycosylase activity for removal of faPy and alkylating
DNA damage in CD4+ and CD8+ T cells from 7 HIV-infected patients before and 6
months after initiating HAART. All patients received an HIV protease inhibitor
(indinavir, 800 mg thrice a day) in combination with two nucleoside analogs [zidovudine
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

11
(250 mg) and lamivudine (150 mg), twice a day]. In response to HAART there was a
marked fall in HIV RNA copies in plasma [maximal decrease: 3.02(1.92-3.98)log10,
p<0.05] and a marked increase in CD4+ [maximal increase: 150(50-215)x106/L, p<0.05]
and CD8+ T cell [maximal increase: 260(30-415)x106/L, p<0.05] counts in peripheral
blood. Concomitantly, there was a significant rise in faPy activity in CD4+ T cells (Figure
4), inversely correlated with the decrease in plasma viral load (r=0.68, p<0.05). However,
there was no normalization compared with faPy activity in healthy controls (Figure 4).
Moreover, even if faPy activity in CD8+ T cells was raised before initiating therapy,
HAART induced a further increase in activity with particularly enhancing effect in those
with the lowest faPy levels at baseline (Figure 4). In comparison, DNA glycosylase
activity for repair of alkylating damage showed no changes during HAART in either
CD4+ or CD8+ T cells (data not shown). Finally, we analysed 8-oxoG levels in T cell
subsets of 4 AIDS patients who had received HAART for 1 year. Notably, while CD4+ T
cells in AIDS patients not receiving HAART had markedly enhanced levels of 8-oxoG in
genomic DNA (Figure 1), CD4+ T cells from AIDS patients on successful HAART had 8-
oxoG levels within the range of healthy controls [34.0(29.5-36.0) 8-oxoG per 106 dG
versus 40.5(27.8-48.0) 8-oxoG per 106 dG, patients and controls, respectively].
Discussion
Several studies have reported enhanced oxidative stress in HIV infection possibly playing
a pathogenic role in this disorder.10 In the present study we show that HIV-infected
patients, and particularly those with advanced disease, are characterized by enhanced
oxidative DNA damage in CD4+ T cells as assessed by increased 8-oxoG accumulation.
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

12
Notably, this increase in oxidative DNA damage was accompanied by a marked decline
in DNA glycosylase activity for repair of oxidative base lesion in CD4+ T cells. In
contrast, CD8+ T cells of HIV-infected patients, with 8-oxoG levels similar or decreased
compared with those in healthy controls, showed enhanced capacity to repair oxidative
damage. Finally, during HAART there was a rise in glycosylase activity in CD4+ T cells
accompanied by a near normalisation of 8-oxoG levels in these cells. Although we can
not totally exclude some influence from any undiagnosed and clinical asymptomatic co-
infection on our results, our data suggest that impaired base excision repair of oxidative
damage, with subsequent accumulation of oxidative DNA base lesions in CD4+ T cells,
may play a pathogenic role in HIV infection.
The mechanisms leading to enhanced oxidative stress in CD4+ T cells during HIV
infection are complex. Increased activity of inflammatory cytokines (e.g. TNFα) and
altered intracellular glutathione redox status, characterizing HIV-infected patients,9 may
promote oxidative DNA damage in these cells. However, our findings of an inverse
correlation between viral load and glycosylase activity and near normalization of repair
capacity during HAART, suggest that direct HIV-related mechanisms may be involved in
the dysregulation of oxidative DNA damage/repair activity during HIV infection. Thus,
TNFα and ROS can increase transcription of viral proteins by activating the nuclear
factor κB.32 The HIV proteins Tat and gp120 may again directly enhance oxidative stress
in T cells possibly involving TNF dependent mechanisms,33 representing a pathogenic
loop promoting inflammation, enhanced oxidative stress and HIV replication. Moreover,
the HIV protein vpr, which may arrest T cells and induce subsequent apoptosis, has been
found to incorporate catalytically active uracil DNA glycosylase into HIV viron particle
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

13
further supporting a link between HIV replication and DNA repair mechanisms.34 In fact,
this glycosylase, which initiates base excision repair of deaminated cytosine, is important
for modulation of the virus mutation rate.35 It is therefore tempting to hypothesize that the
dysbalance between glycosylase activity and oxidative DNA damage in CD4+ T cells
during HIV infection could reflect potent interactions between viral proteins, enhanced
oxidative stress and increased activity of inflammatory cytokines overwhelming DNA
repair mechanisms.
The regulation of the various DNA glycosylases involved in repair of oxidative
damage in man has not been fully clarified. However, hNth1 transcription was shown to
increase during early and mid S-phase of cell cycle36 suggesting that increased 5ohC
DNA glycosylase activity in CD8+ T cells during HIV infection may correlate with the
enhanced spontaneous T cell proliferation observed in these patients.37 In CD4+ T cells
such a stimulus may be counteracted and overshadowed by mechanisms as discussed
above. In contrast to hNth1, the hOgg1 gene lacks TATA or CAAT boxes suggesting that
Ogg1 is a housekeeping gene with stable expression during cell cycle.38 However, the
results are somewhat conflicting, possibly reflecting different regulation in different cell
types. Thus, raised 8-oxoG levels and ischemia have been reported to enhance Ogg1
activity in human colorectal carcinoma cells and in mouse brain, respectively.39,40
Moreover, while acute ischemia and oxidative stress may promote up-regulation of DNA
glycosylase activity, persistently enhanced oxidative stress and inflammation as in HIV
infection, may potentially down-regulate or “consume” the activity of these enzymes.
Nevertheless, the mechanisms for regulation of DNA glycosylases for repair of oxidative
base lesions in T cell subsets during HIV infection are at present unclear.
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

14
In the present study we show that DNA glycosylases for repair of oxidative base
lesions are differently regulated in CD4+ and CD8+ T cells during HIV infection. In fact,
rather than decreased activity, CD8+ T cells in non-AIDS patients were characterized by
enhanced faPy, 5-ohC and 8-oxoG DNA glycosylase activity, accompanied by a non-
significant decrease in 8-oxoG level in these cells. This phenomenon could possibly
reflect that while HIV infection is characterized by a functional and numerical decline in
CD4+ T cells, the CD8+ T cells is characterized by increased numbers and enhanced
activity at least in non-AIDS patients.41
Our findings suggest that CD4+ T cells in HIV-infected patients may lack the
ability to sufficiently repair DNA damage induced by oxidative stress that may have
several consequences. Mutant forms of hOgg1 have been found in lung and kidney
tumors.42 A very recent report suggests that reduced Neil1 activity arising from mutations
and reduced expression may be involved in pathogenesis of a subset of gastric cancers.43
It is possible that the imbalance between oxidative DNA damage and repair activity in
CD4+ T cells from AIDS patients might contribute to the increased frequency of
malignancies in these patients. Moreover, while long-time exposure to moderately
increased oxidative DNA damage may pre-dispose to malignancies, markedly enhanced
oxidative DNA damage combined with decreased repair activity as found in CD4+ T cells
from HIV-infected patients, may lead to functional impairment of these cells by slowing
cell cycle progression upon activation and by enhancing apoptosis, possibly contributing
to the pathogenesis of T cell deficiency in these patients. Finally, the HIV genome is
characterized by a high frequency of mutation that at least partly is due to the high rate of
HIV replication and the low fidelity of the reverse transcriptase gene. These mutations
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

15
which involve a high frequency of base misincorporations, may lead to a several-fold
decrease in sensitivity to one or more antiretroviral drugs and is at present an increasing
challenge in HAART.44 Several oxidative base lesions are strongly premutagenic if they
remain in DNA during replication. 5-ohC and 8-oxoG are stable oxidation products with
strong miscoding properties, which produce G→T transversions45 and C→T
transitions/C→A transversions,46 respectively. Therefore, it is possible that accumulation
of mutagenic base lesions as a result of enhanced oxidative stress and impaired capacity
for repair of oxidative DNA damage in CD4+ T cells in HIV-infected patients, also may
pre-dispose for elevated mutation frequency in the HIV genome of these cells.
. In conclusion, the present study further supports a role for oxidative stress in the
pathogenesis of HIV infection by demonstrating a marked imbalance between
accumulation of oxidative DNA damage and the capacity to repair such lesions in CD4+
T cells (favouring enhanced DNA damage). These findings may represent a previous
unrecognised mechanism involved in the numerical and functional impairment of CD4+ T
cells during HIV infection.
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

16
References
1. Halliwell B. Antioxidant defence mechanisms: from the beginning to the end (of the
beginning). Free Radic Res. 1999;31:261-272.
2. Ames BN, Gold LS, Willet WC. The causes and prevention of cancer. Proc Nat Acad
Sci USA. 1195;92:5258-5265.
3. Hensley K, Robinson KA, Gabbita SP, Salsman S, Floyd RA. Reactive oxygen
species, cell signaling, and cell injury. Free Radic Biol Med. 2000;28:1456-1462.
4. Cooke MS, Evans MD, Dizdaroglu M, Lunec J. Oxidative DNA
damage:mechanisms, mutation, and disease. FASEB J. 2003;17:1195-1214.
5. Olinski R, Gackowski D, Foksinski M, Rozalski R, Roszkowski K, Jaruga P.
Oxidative DNA damage: assessment of the role in carcinogenesis, atherosclerosis,
and acquired immunodeficiency syndrome. Free Radic Biol Med. 2002;33:192-200.
6. Dröge W, Eck HP, Mihm S. Oxidant-antioxidant status in human immunodeficiency
virus infection. Methods Enzymol. 1994;233:594-601.
7. Aukrust P, Svardal AM, Müller F, et al. Increased levels of oxidized glutathione in
CD4+ lymphocytes associated with disturbed intracellular redox balance in human
immunodeficiency virus type 1 infection. Blood. 1995;86:258-267.
8. Roederer M, Staal FJ, Osada H, Herzenberg LA, Herzenberg LA. CD4 and CD8 T
cells with high intracellular glutathione levels are selectively lost as the HIV infection
progresses. Int Immunol. 1991;3:933-937.
9. Aukrust P, Muller F, Svardal AM, Ueland T, Berge RK, Froland SS. Disturbed
glutathione metabolism and decreased antioxidant levels in human immunodeficiency
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

17
virus-infected patients during highly active antiretroviral therapy--potential
immunomodulatory effects of antioxidants. J Infect Dis. 2003;188:232-238.
10. Aukrust P, Müller F. Glutathione redox disturbances in HIV infection - immunologic
and therapeutical consequences. Nutrition. 1999;15:165-167.
11. Buttke TM, Sandstrom PA. Oxidative stressas a mediator of apoptosis. Immunol
Today. 1994;15:7-10.
12. Demple B, Harrison L. Repair of oxidative damage to DNA: Enzymology and
biology. Ann Rev Biochem. 1994;63:915-948.
13. Feig DI, Sowers LC, Loeb LA. Reverse chemical mutagenesis: Identification of the
mutagenic lesions resulting from reactive oxygen species-mediated damage to DNA.
Proc Natl Acad Sci USA. 1994;91:6609-6613.
14. Kasai H, Crain PF, Kuchino Y, Nishimura S, Ootsuyama A, Tanooka H. Formation
of 8-hydroxyguanine moiety in cellular DNA by agents producing oxygen radicals
and evidence for its repair. Carcinogenesis. 1986;7:1849-1851.
15. Wagner R.J, Hu C, Ames B. Endogenous oxidative damage of deoxycytidine in
DNA. Proc Natl Acad Sci USA. 1992;89:3380-3384.
16. Seeberg E, Eide L, Bjørås M. The base excision repair pathway. Trends Biochem Sci.
1995;20:391-397.
17. Lindahl T, Wood RD. Quality control by DNA repair. Science. 1999;286:1897-1905.
18. Aspinwall R, Rothwell DG, Roldan-Arjona T, et al. Cloning and characterization of a
functional human homolog of Escherichia coli endonuclease III. Proc Natl Acad Sci
USA. 1997;94:109-114.
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

18
19. Aburatani H, Hippo Y, Ishida T, et al. Cloning and characterization of mammalian 8-
hydroxyguanine-specific DNA glycosylase/apurinic, apyrimidinic lyase, a functional
mutM homologue. Cancer Res. 1997;57:2151-2156.
20. Arai K, Morishita K, Shinmura K, et al. Cloning of a human homolog of the yeast
OGG1 gene that is involved in the repair of oxidative DNA damage. Oncogene.
1997;14:2857-61.
21. Bjørås M, Luna L, Hoff E, Haug T, Rognes T, Seeberg E. Opposite base-dependent
reactions of a human base excision repair enzyme on DNA containing 7,8-dihydro-8-
oxoguanine and abasic sites. EMBO J. 1997;16:6314-6322.
22. Bandaru V, Sunkara S, Wallace SS, Bond JP. A novel human DNA glycosylase that
removes oxidative DNA damage and is homologous to Escherichia coli endonuclease
VIII. DNA Repair. 2002;17:;517-529.
23. Kuo FC, Sklar J. Augmented expression of a human gene for 8-oxoguanine DNA
glycosylase (MutM) in B lymphocytes of the dark zone in lymph node germinal
centers. J Exp Med. 1997;186:1547-1556.
24. Lu R, Nash HM, Verdine GL. A mammalian DNA repair enzyme that excises
oxidatively damaged guanines maps to a locus frequently lost in lung cancer. Curr
Biol. 1997;7:397-407.
25. Radicella JP, Dherin C, Desmaze C, Fox MS, Boiteux S. Cloning and
characterization of hOGG1, a human homolog of the OGG1 gene of Saccharomyces
cerevisiae. Proc Natl Acad Sci USA. 1997;94:8010-8015.
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

19
26. Hazra TK, Izumi T, Boldogh I, et al. Identification and characterization of a human
DNA glycosylase for repair of modified bases in oxidatively damaged DNA. Proc
Natl Acad Sci USA. 2002;99:3523-3528.
27. Hazra TK, Kow YW, et al. Identification and characterization of a novel human DNA
glycosylase for repair of cytosine-derived lesions. J Biol Chem. 2002;277:30417-
3020.
28. Morland I, Rolseth V, Luna L, Rognes T, Bjoras M, Seeberg E. Human DNA
glycosylases of the bacterial Fpg/MutM superfamily: an alternative pathway for the
repair of 8-oxoguanine and other oxidation products in DNA. Nucleic Acids Res.
2002;30:4926-4936.
29. Hofer T, Moller L. Reduction of oxidation during the preparation of DNA and
analysis of 8-hydroxy-2'-deoxyguanosine. Chem Res Toxicol. 1998;11:882-887.
30. Boiteux S, Belleney J, Roques BP, Laval J. Two rotyameric forms of open ring 7-
methylguanine are present in alkylated polynucleotides. Nucleic Acids Res.
1984;12:5429-3549.
31. Yeni PG, Hammer SM, Hirsch MS, et al. Treatment for adult HIV infection: 2004
recommendations of the International AIDS Society-USA Panel. JAMA.
2004;292:251-265.
32. Greenspan HC, Aruoma OI, Arouma O. Could oxidative stress initiate programmed
cell death in HIV infection? A role for plant derived metabolites having synergistic
antioxidant activity. Chem Biol Interact. 1994;91:187-197.
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

20
33. Westendorp MO, Shatrov VA, Schulze-Osthoff K, et al. HIV-1 Tat potentiates TNF-
induced NF-κ B activation and cytotoxicity by altering the cellular redox state.
EMBO J. 1995;14:546-554.
34. Willetts KE, Rey F, Agostini I, et al. DNA repair enzyme uracil DNA glycosylase is
specifically incorporated into human immunodeficiency virus type 1 viral particles
through a Vpr-independent mechanism. J Virol. 1999;73:1682-1688.
35. Chen R, Le Rouzic E, Kearney JA, Mansky LM, Benichou S. Vpr-mediated
incorporation of UNG2 into HIV-1 particles is required to modulate the virus
mutation rate and for replication in macrophages. J Biol Chem. 2004;279:28419-
28425.
36. Luna L, Bjoras M, Hoff E, Rognes T, Seeberg E. Cell-cycle regulation, intracellular
sorting and induced overexpression of the human NTH1 DNA glycosylase involved
in removal of formamidopyrimidine residues from DNA. Mutat Res. 2000;460:95-
104.
37. Badley AD, Pilon AA, Landay A, Lynch DH. Mechanisms of HIV-associated
lymphocyte apoptosis. Blood. 2000;96:2951-2964.
38. Dhenaut A, Boiteux S, Radicella JP. Characterization of the hOGG1 promoter and its
expression during the cell cycle. Mutat Res. 2000;461:109-118.
39. Lin LH, Cao S, Yu L, Cui J, Hamilton WJ, Liu PK. Up-regulation of base excision
repair activity for 8-hydroxy-2'-deoxyguanosine in the mouse brain after forebrain
ischemia-reperfusion. J Neurochem. 2000;74:1098-1105.
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

21
40. Kim JI, Park YJ, Kim KH, et al. hOGG1 Ser326Cys polymorphism modifies the
significance of the environmental risk factor for colon cancer. World J Gastroenterol.
2003;5:956-960.
41. Levy JA. Pathogenesis of human immunodeficiency virus infection. Microbiol Rev.
1993;57:183-289.
42. Chevillard S, Radicella JP, Levalois C, et al. Mutations in OGG1, a gene involved in
the repair of oxidative DNA damage, are found in human lung and kidney tumours.
Oncogene. 1998;16:3083-3086.
43. Shinmura K, Tao H, Goto M, et al. Inactivating mutations of the human base excision
repair gene NEIL1 in gastric cancer. Carcinogenesis. 2004, in press.
44. Deeks SG Treatment of antiretroviral-drug-resistant HIV-1 infection. Lancet.
2003;362:2002-2011.
45. Cheng KC, Cahill DS, Kasai H, Nishimura S, Loeb LA. 8-Hydroxyguanine, an
abundant form of oxidative DNA damage, causes G →T and A→C substitutions. J
Biol Chem. 1992;267:166-172.
46. Hatahet Z, Purmal AA, Wallace SS. A novel method for site specific introduction of
single model oxidative DNA lesions into oligodeoxyribonucleotides. Nucleic Acids
Res. 1993;21:1563-1568.
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

22
Figure legends
Figure 1. Accumulation of 8-oxoG in (A) CD4+ and (B) CD8+ T cells from HIV-infected
patients. Nuclear DNA was isolated from T cell subsets of AIDS patients (n=5), non-
AIDS HIV-infected patients (n=4) and healthy controls (n=7). Levels of 8-oxoG were
quantified by HPLC-ECD after enzymatic DNA hydrolysis. * p<0.05 versus healthy
controls; # p<0.05 versus non-AIDS patients. Data are given as medians and 25th-75th
percentiles.
Figure 2. FaPy DNA glycosylase activity (A and B) and alkylbase DNA glycosylase
activity (C and D) in extracts (ng) of CD4+ and CD8+ T cells from HIV-infected patients
and controls. Extracts (ng) isolated from AIDS patients (n=7), non-AIDS HIV-infected
patients (n=8) and healthy controls (n=13) were incubated with 0,4 µg 3H-labelled faPy-
poly(dG-dC) DNA or alkylated calf thymus DNA. Horizontal lines represent median
values.
Figure 3. 5-ohC DNA glycosylase activity (A) and 8-oxoG DNA glycosylase activity (B)
in CD4+ and CD8+ T cells from HIV-infected patients. Extracts (ng), isolated from AIDS-
patients (n=7), non-AIDS HIV-infected patients (n=8) and healthy controls (n=13), were
incubated with 50 fmol duplex oligo containing a single 8-oxoG or 5-ohC residue. The
cleavage products were analysed by 20% denaturing PAGE and PhosphoImager
scanning. * p<0.05 and ** p<0.01 versus healthy controls. # p<0.05 versus AIDS
patients. Data are given as medians and 25th-75th percentiles.
Figure 4. Fapy DNA glycosylase activity in CD4+ (A) and CD8+ (B) T cells from HIV-
infected patients during HAART. Extracts (ng), isolated from 7 HIV-infected patients
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

23
before and after 6 months of HAART, were incubated with 0,4 µg 3H-labelled faPy-
poly(dG-dC) DNA. The shaded area indicates ranges in 13 healthy controls.
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

24
Table 1. Characteristics of the study group.
Non-AIDS HIV-infected patients
(n=8)
AIDS patients(n=7)
Controls(n=13)
Age (years), median and
ranges
38(32-52) 37(34-56) 37(29-61)
Males/females 6/2 5/2 9/4
CD4+ T cells(106/l)
280(95-430)
45(10-110)
760(540-980)
CD8+ T cells(106/l)
710(410-1170)
280(110-430)
400(210-550)
TNFα(pg/ml)
25(12-37)
65(40-98)
8(0-12)
HIV RNA copies/ml
plasma (x103)44.2
(22.9-108.6)510.9
(210.0-1200.0) -Data are given as medians and 25th-75th percentiles if not otherwise stated.
T cell subset counts were analysed in peripheral blood and TNFα levels in plasma.
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

25
Figure 1.
CD4+
Controls non-AIDS AIDS0
50
100
150
200*#A
*
8-ox
oG p
er 1
06 d
G
CD8+
Controls non-AIDS AIDS0
25
50
75B
8-ox
oG p
er 1
06 d
G
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom
26
Figure 2.
CD4+
Controls non-AIDS AIDS0
1
2
3A
P<0.001
P<0.001
P<0.001
Rea
leas
ed f
aPy
base
s (f
mol
) CD8+
Controls non-AIDS AIDS0.0
2.5
5.0
7.5
10.0 B P<0.001
P<0.002
Rea
leas
ed f
aPy
base
s (f
mol
)
CD4+
Controls non-AIDS AIDS0.0
0.5
1.0
1.5 C
Rel
ease
d al
kyla
ted
base
s(f
mol
)
CD8+
Controls non-AIDS AIDS0.0
0.5
1.0
1.5 D
Rel
ease
d al
kyla
ted
base
s(f
mol
)
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

27
Figure 3.
5-ohC
CD4 CD80
5
10
15
20 Controlsnon-AIDSAIDS
*
**#
**
A
CD4+ T cells CD8 + T cells
Rel
ease
d ba
ses
(fm
ol)
8-oxoG
CD4 CD80.0
0.1
0.2
0.3
0.4
0.5 B
*
**
#
CD4+ T cells CD8 + T cells
Rel
ease
d ba
ses
(fm
ol)
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom

28
Figure 4.
CD4+
0 6 months0.0
0.5
1.0
1.5
2.0A
P<0.02
Rea
leas
ed f
aPy
base
s (f
mol
)
CD8+
0 6 months0.0
2.5
5.0
7.5
10.0B P<0.02
Rea
leas
ed f
aPy
base
s (f
mol
)
For personal use only. by guest on June 1, 2013. bloodjournal.hematologylibrary.orgFrom
Related Documents











