Pharmacogenomics (Epub ahead of print) ISSN 1462-2416 part of Pharmacogenomics Review 10.2217/PGS.15.65 © 2015 Future Medicine Ltd Chronic HCV infection comprises a broad spectrum of liver disease, ranging from no or minimal activity to active hepatitis that in time may progress to severe liver fibrosis, cirrhosis and hepatocellular carcinoma if left untreated. This review describes the impact of genetic variants of interleukin 28B ( IL28B; also known as interferon-lambda 3), inosine triphosphate pyrophosphatase (ITPA) and patatin-like phospholipase domain-containing 3 (PNPLA3) on therapeutic outcome and liver disease severity in HCV-infected patients. Keywords: genetic variant • HCV • histology • IL28B • inosine triphosphate pyrophosphatase • interferon-λ • ITPA • ITPase • natural history • PNPLA3 • therapy Hepatitis C virus (HCV) HCV was discovered in 1989, following an intensive search for the major etiologic agent associated with non-A, non-B hepatitis [1] . The HCV virus is a positive stranded RNA virus and belongs to the family of flaviviri- dae [2] . There are six major genotypes, and recently a seventh genotype was reported, all with different global geographic distri- butions [3–5] . If left untreated, HCV virus infection over time may progress to severe liver fibrosis, cirrhosis and hepatocellular carcinoma [6–8] . Interleukin 28B (IL28B) genetic variants & impact on interferon- based HCV therapy In 2009, a genome-wide association study (GWAS) demonstrated that several genetic variants in close proximity to the IL28B (also known as interferon-λ3) gene pre- dicted greater likelihood of achieving a sustained virological response (SVR), in other words, undetectable HCV RNA 24 weeks after completion of therapy, follow- ing treatment with pegylated interferon- α (pegINF- α) and ribavirin among adher- ent HCV genotype 1 infected patients (Figure 1) . The strongest association, among a predominantly Caucasian population, was noted for the single nucleotide polymor- phism (SNP) rs12979860 where the CC variant had an almost two-fold increased likelihood of achieving SVR as compared with the TT variant [9] . The differences in C allele frequencies, with a greater frequency in Asian and European populations as com- pared with populations of African origin, could to a major extent explain the previ- ously recognized racial differences in treat- ment response [9,10] . Furthermore the CC variant of rs12979860 also was associated with a greater first phase decline in HCV RNA (i.e., the reduction in HCV RNA dur- ing the first days of interferon therapy) as well as spontaneous clearance of the viral infection in HCV genotype 1, but some- what counterintuitive, also with a higher baseline viral load [9,11,12] . Regarding patients infected with HCV genotype 2 or 3, the IL28B C allele also has been reported to be associated with greater first phase decline in HCV RNA as well as higher baseline viral load [12,13] . However, there is discordance regarding the potential benefit of the C allele regarding treatment outcome for these patients when treated with INF- α and ribavirin. Some studies Impact of IL28B, ITPA and PNPLA3 genetic variants on therapeutic outcome and progression of hepatitis C virus infection Karolina Rembeck 1 & Martin Lagging* ,1 Department of Infectious Medicine, Institute of Biomedicine at Sahlgrenska Academy, University of Gothenburg, Guldhedsgatan 10B, SE-413 46, Gothenburg, Sweden *Author for correspondence: Tel.: +46 313 424 658 Fax: +46 3141 1256 [email protected] For reprint orders, please contact: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Pharmacogenomics (Epub ahead of print) ISSN 1462-2416
part of
PharmacogenomicsReview
10.2217/PGS.15.65 © 2015 Future Medicine Ltd
Pharmacogenomics
Review 2015/06/2616
10
2015
Chronic HCV infection comprises a broad spectrum of liver disease, ranging from no or minimal activity to active hepatitis that in time may progress to severe liver fibrosis, cirrhosis and hepatocellular carcinoma if left untreated. This review describes the impact of genetic variants of interleukin 28B (IL28B; also known as interferon-lambda 3), inosine triphosphate pyrophosphatase (ITPA) and patatin-like phospholipase domain-containing 3 (PNPLA3) on therapeutic outcome and liver disease severity in HCV-infected patients.
Keywords: genetic variant • HCV • histology • IL28B • inosine triphosphate pyrophosphatase • interferon-λ • ITPA • ITPase • natural history • PNPLA3 • therapy
Hepatitis C virus (HCV)HCV was discovered in 1989, following an intensive search for the major etiologic agent associated with non-A, non-B hepatitis [1]. The HCV virus is a positive stranded RNA virus and belongs to the family of flaviviri-dae [2]. There are six major genotypes, and recently a seventh genotype was reported, all with different global geographic distri-butions [3–5]. If left untreated, HCV virus infection over time may progress to severe liver fibrosis, cirrhosis and hepatocellular carcinoma [6–8].
Interleukin 28B (IL28B) genetic variants & impact on interferon-based HCV therapyIn 2009, a genome-wide association study (GWAS) demonstrated that several genetic variants in close proximity to the IL28B (also known as interferon-λ3) gene pre-dicted greater likelihood of achieving a sustained virological response (SVR), in other words, undetectable HCV RNA 24 weeks after completion of therapy, follow-ing treatment with pegylated interferon-α (pegINF-α) and ribavirin among adher-ent HCV genotype 1 infected patients (Figure 1). The strongest association, among
a predominantly Caucasian population, was noted for the single nucleotide polymor-phism (SNP) rs12979860 where the CC variant had an almost two-fold increased likelihood of achieving SVR as compared with the TT variant [9]. The differences in C allele frequencies, with a greater frequency in Asian and European populations as com-pared with populations of African origin, could to a major extent explain the previ-ously recognized racial differences in treat-ment response [9,10]. Furthermore the CC variant of rs12979860 also was associated with a greater first phase decline in HCV RNA (i.e., the reduction in HCV RNA dur-ing the first days of interferon therapy) as well as spontaneous clearance of the viral infection in HCV genotype 1, but some-what counterintuitive, also with a higher baseline viral load [9,11,12].
Regarding patients infected with HCV genotype 2 or 3, the IL28B C allele also has been reported to be associated with greater first phase decline in HCV RNA as well as higher baseline viral load [12,13]. However, there is discordance regarding the potential benefit of the C allele regarding treatment outcome for these patients when treated with INF-α and ribavirin. Some studies
Impact of IL28B, ITPA and PNPLA3 genetic variants on therapeutic outcome and progression of hepatitis C virus infection
Karolina Rembeck1 & Martin Lagging*,1
Department of Infectious Medicine, Institute of Biomedicine at Sahlgrenska Academy, University of Gothenburg, Guldhedsgatan 10B, SE-413 46, Gothenburg, Sweden *Author for correspondence: Tel.: +46 313 424 658 Fax: +46 3141 1256 [email protected]
For reprint orders, please contact: [email protected]

10.2217/PGS.15.65 Pharmacogenomics (Epub ahead of print)
Chromosome 19 q13.
13
IFN-λ3(IL28B)
IFN-λ4 IFN-λ2(IL28A)
IFN-λ1(IL29)
rs8099917(T/G)
Exon 5 1234
rs12979860(C/T)
rs117648444(A/G)
rs368234815(ss469415590)
(TT/∆G)
Figure 1. Genomic location of IL28B (also known as IFN-λ3) and IFN-λ4 on chromosome 19.
future science group
Review Rembeck & Lagging
have reported that CC variant of rs12979860, as compared with the TT variant, is associated with increased likelihood of achieving SVR among Cau-casian patients [14,15], whereas others have failed to demonstrate such an a ssociation [13,16].
Another SNP, rs8099917, located in proximity to and in strong linkage disequilibrium (i.e., a non-random association of two alleles at two loci) with rs12979860 [12,17], also has been reported to be of significance regarding treatment outcome, especially among populations of Asian origin, where the CC variant of rs12979860 predominates. For HCV geno-type 1 infected patients, the rs8099917 TT variant is associated with favorable treatment outcome, sponta-neous virus clearance and greater first phase decline in HCV RNA, but also with a higher baseline viral load as compared with the TG and GG variants [17–19]. As described for the rs12979860 CC variant, the association between TT variant of rs8099917 and a
greater first phase decline was also observed among HCV genotypes 2 and 3 infected patients [12].
IL28B genetic variants & impact on direct acting antiviral-based HCV therapyIn the setting of direct acting antiviral (DAA) regi-mens, the rs12979860 CC variant was associated with favorable outcome following triple therapy with the first generation protease inhibitors, telapre-vir and boceprevir [20,21], but with the introduction of interferon-free regimens the importance of the IL28B genetic variants appears to be diminishing [22]. Although, in one interferon-sparing trial among HCV genotype 1 infected patients evaluating faldaprevir and deleobuvir, with and without ribavirin, the CC variant of rs12979860 was associated with increased likelihood of achieving SVR in comparison with non-CC variants [23]. Similarly in the OPTIMIST-1 study, when simeprevir and sofosbuvir were given for 8 weeks

10.2217/PGS.15.65future science group
Impact of IL28B, ITPA and PNPLA3 genetic variants on therapeutic outcome & progression of HCV infection Review
without ribavirin among HCV genotype 1 infected patients, IL28B genetic variants impacted on outcome (SVR 93, 84 and 64% for rs12979860 CC, CT and TT, respectively) [24]. In the latter study, this differ-ence was largely overcome when the treatment dura-tion was extended to 12 weeks (SVR 100, 97 and 92% for rs12979860 CC, CT and TT, respectively) [24]. Other studies investigating interferon-free HCV regi-mens have failed to demonstrate any impact of IL28B genetic variants on outcome [25], possibly due to more appropriate duration of therapy.
IL28B genetic variants & impact on liver histopathologyIn the setting of liver disease progression prior to therapeutic intervention, the GG variant of rs8099917 reportedly has been associated with milder fibrosis, less necroinflammatory activity and slower fibrosis progression rate in European HCV genotype non-1 infected patients [26]. The CC variant of rs12979860 was in the setting of Scandinavian HCV genotype 3 infected patients, significantly associated with higher ALT and AST to platelet ratio index (APRI) score, as compared with the CT and TT variants, indicative of a higher degree of inflammation and fibrosis [16]. Simi-larly in Japanese patients infected with HCV genotype 1 or 2, the otherwise favorable T allele of rs8099917 was associated with more severe inflammation activity and fibrosis [27]. In a study of primarily HCV genotype 1 infected patients, 276 paired liver biopsies were eval-uated, with a median of 4 years elapsed between the biopsies. In this study, the CC variant of rs12979860 was associated with greater hepatic inflammation grade and higher ALT, but not with fibrosis progression [28]. In the NORDynamIC study enrolling HCV genotype 2 or 3 infected patients, the otherwise favorable SNPs in proximity to IL28B were associated with more severe liver histopathology [29,30]. In this latter study, the CC variant of rs12979860 in HCV genotype 3 infected patients was significantly associated with higher nor-malized ALT, higher APRI score, more severe portal inflammation and higher grade of steatosis as compared with the CT or TT variants [29]. Similarly, the TT variant of rs8099917 was significantly associated with higher normalized ALT and APRI score as well more pronounced interface hepatitis grade and higher fibro-sis stage among HCV genotype 3 infected patients [30]. Likewise in the real-life trial by Ydreborg et al., HCV genotype 3 infected patients with the CC variant of rs12979860 had significantly higher APRI score and higher liver stiffness values as measured by means of Fibroscan [31]. Thus, regarding the natural course of HCV-associated liver disease, the less favorable genetic variants with regard to spontaneous resolution of HCV
infection and therapeutic outcome, i.e. the GG variant of rs8099917 and the TT variant of rs12979860, may be protective of liver disease progression, especially in the presence of HCV genotype 3 infection.
Interestingly the CC variant of rs12979860, among HCV genotype 3-infected patients, has been associ-ated with higher baseline viral load [29] indicating that the changes in the liver histopathology observed may have been secondary to a direct viral cytopathic effect [32–34]. HCV genotype 3-induced steatosis, which also is associated with higher HCV RNA levels, has previously been described to entail enhanced fibrosis progression [32,35–37].
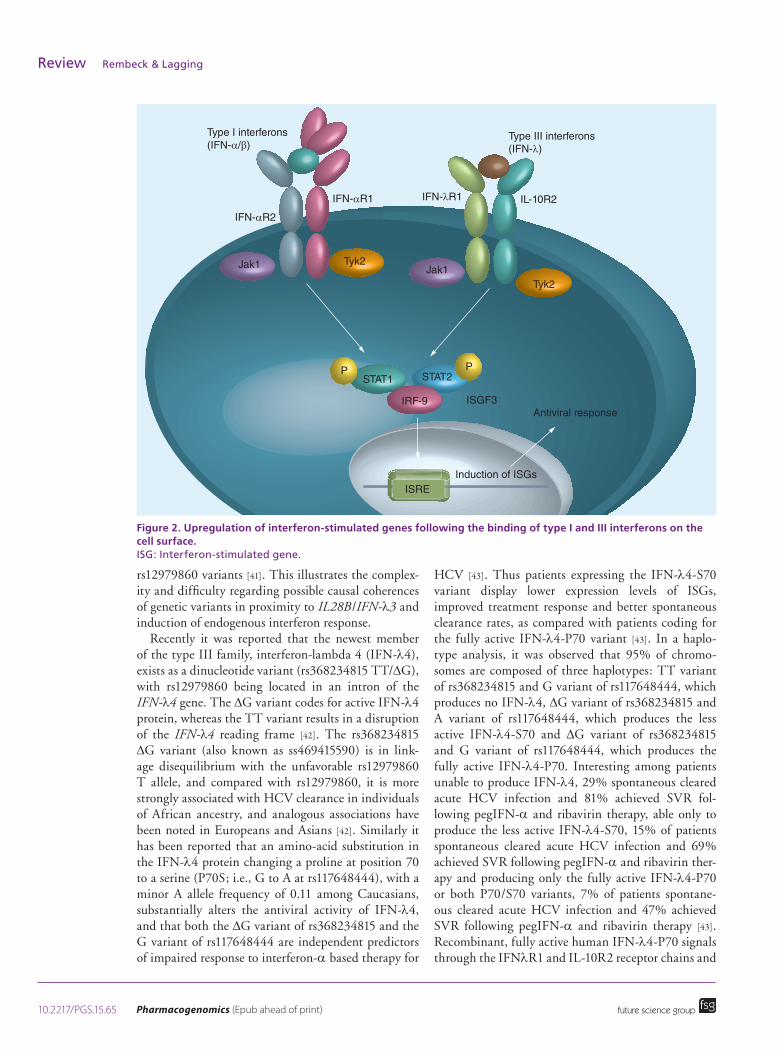
Possible molecular mechanism of action of genetic variants in proximity to IL28BThe genes in proximity to rs8099917 and rs12979860 code for the type III or interferon-λ family, i.e. IL28A (IFN-λ2), IL28B (IFN-λ3) and IL29 (IFN-λ1) genes, as well as the recently discovered interferon-λ4 [38]. The type III or lambda (λ) interferons are cytokines that are secreted in response to viral infections, and bind to receptors on uninfected cells, modulating the janus kinase-signal transducer and activator of tran-scription signaling pathway, and thus inducing the transcription of interferon-stimulated genes (ISGs) (Figure 2) [39].
The CC variant of rs12979860 has been reported to be associated with lower intrahepatic expression of ISGs, for example, the expression of IFI27, ISG15, RSAD2, HTATIP2, etc., in a study of 109 patients with chronic hepatitis C genotype 1–4 infection [40]. In this same study, higher ISG expression also was noted among nonresponders to interferon and ribavi-rin therapy irrespective of IL28B genotype. However, the finding of significantly lower ISG expression did not correlate causally with the expression of IL28B/IFN-λ3, where TT variant of rs12979860 was asso-ciated with lower IL28B/IFN-λ3 expression [40]. The latter finding is consistent with the results from two other, independent studies where the IL28B mRNA expression levels in peripheral blood mononuclear cells were measured in HCV genotype 1 infected patients, as well as in whole blood a healthy cohort [18,19]. These studies reported significantly lower mRNA expression levels of IL28B/IFN-λ3 in subjects with the rs8099917 GG genotype [18,19]. In contrast, another larger study enrolling HCV genotype 1 infected patients (n = 80) found no association between IL28/IFN-λ3 expression levels and rs12979860 genotype [9]. Also, a study mea-suring the intrahepatic levels of ISGs among Japanese HCV genotype-1 infected patients found an associa-tion between the G allele of rs8099917 and higher ISG levels, but this was not significant in the setting of the
10.2217/PGS.15.65 Pharmacogenomics (Epub ahead of print)
PP
IFN-αR2
IFN-αR1 IFN-λR1 IL-10R2
Type III interferons(IFN-λ)
Type I interferons(IFN-α/β)
Jak1 Jak1Tyk2
Tyk2
Induction of ISGsISRE
STAT1 STAT2
IRF-9 ISGF3Antiviral response
Figure 2. Upregulation of interferon-stimulated genes following the binding of type I and III interferons on the cell surface. ISG: Interferon-stimulated gene.
future science group
Review Rembeck & Lagging
rs12979860 variants [41]. This illustrates the complex-ity and difficulty regarding possible causal coherences of genetic variants in proximity to IL28B/IFN-λ3 and induction of endogenous interferon response.
Recently it was reported that the newest member of the type III family, interferon-lambda 4 (IFN-λ4), exists as a dinucleotide variant (rs368234815 TT/ΔG), with rs12979860 being located in an intron of the IFN-λ4 gene. The ΔG variant codes for active IFN-λ4 protein, whereas the TT variant results in a disruption of the IFN-λ4 reading frame [42]. The rs368234815 ΔG variant (also known as ss469415590) is in link-age disequilibrium with the unfavorable rs12979860 T allele, and compared with rs12979860, it is more strongly associated with HCV clearance in individuals of African ancestry, and analogous associations have been noted in Europeans and Asians [42]. Similarly it has been reported that an amino-acid substitution in the IFN-λ4 protein changing a proline at position 70 to a serine (P70S; i.e., G to A at rs117648444), with a minor A allele frequency of 0.11 among Caucasians, substantially alters the antiviral activity of IFN-λ4, and that both the ΔG variant of rs368234815 and the G variant of rs117648444 are independent predictors of impaired response to interferon-α based therapy for
HCV [43]. Thus patients expressing the IFN-λ4-S70 variant display lower expression levels of ISGs, improved treatment response and better spontaneous clearance rates, as compared with patients coding for the fully active IFN-λ4-P70 variant [43]. In a haplo-type analysis, it was observed that 95% of chromo-somes are composed of three haplotypes: TT variant of rs368234815 and G variant of rs117648444, which produces no IFN-λ4, ΔG variant of rs368234815 and A variant of rs117648444, which produces the less active IFN-λ4-S70 and ΔG variant of rs368234815 and G variant of rs117648444, which produces the fully active IFN-λ4-P70. Interesting among patients unable to produce IFN-λ4, 29% spontaneous cleared acute HCV infection and 81% achieved SVR fol-lowing pegIFN-α and ribavirin therapy, able only to produce the less active IFN-λ4-S70, 15% of patients spontaneous cleared acute HCV infection and 69% achieved SVR following pegIFN-α and ribavirin ther-apy and producing only the fully active IFN-λ4-P70 or both P70/S70 variants, 7% of patients spontane-ous cleared acute HCV infection and 47% achieved SVR following pegIFN-α and ribavirin therapy [43]. Recombinant, fully active human IFN-λ4-P70 signals through the IFNλR1 and IL-10R2 receptor chains and

10.2217/PGS.15.65
ITP
ITPaseRibavirin monophosphate(RMP)
IDP
IMPIMPDH
(-)
XMP Adenylo-succinate
GMP AMP
GDP ADP
GTP ATP
Figure 3. ITP to GTP/ATP biochemical pathway.
future science group
Impact of IL28B, ITPA and PNPLA3 genetic variants on therapeutic outcome & progression of HCV infection Review
thus upregulates ISG induction [44]. This higher ISG expression secondary to the production of the fully active IFN-λ4-P70, may subsequently lead to lower levels of viral replication. This lower viral production in turn paradoxically may hamper the triggering of adaptive immune responses such as intrahepatic lym-phocyte degranulation activity [45], which theoretically otherwise would aid in the resolution of infection.
Inosine triphosphate pyrophosphatase (ITPA) genetic variants & impact on HCV therapyThe ITPA gene is located on the chromosome 20 and encodes the enzyme inosine triphosphate pyrophos-phatase (ITPase), which is involved in purine metab-olism. ITPase converts inosine triphosphate (ITP) to inosine monophosphate (IMP) (Figure 3) [46,47], and other potential substrates include deoxyinosine triphosphate (dITP) and xanthosine triphosphate (XTP). Thus the presence of ITPase is essential in order to prevent intracellular accumulation of rogue nucleotides such as ITP, dITP and XTP, which oth-erwise might be falsely incorporated into RNA and DNA producing mistranslation, enzyme inhibition and genetic instability [48–50].
Two allelic variants of the ITPA gene have been associated with reduced ITPase activity, subsequently resulting in increased intracellular concentrations of ITP. The first involves a proline to threonine substitu-tion where the minor A allele variant of rs1127354 is associated with reduced ITPase activity in contrast to the wild-type C variant. The second allelic variant is a splicing altering SNP in the second intron where the minor C variant of rs7270101 variant entails reduced enzymatic activity in contrast to the wild-type A allele [46,47,51,52].
The first reports of reduced ITPase activity leading to accumulation of ITP in human erythrocytes were published in the 1960s [53]. The general consequences of this altered metabolic activity remain to be clari-fied, but reduced ITPase activity has been associated with increased risk of adverse drug reactions [54]. In studies using human cell lines, normal ITPase activ-ity has been reported to protect against DNA damage, possibly secondary to cleansing of ITP and dITP from the intracellular nucleotide pool [55]. ITP is generated continuously in all cells through nucleotide recycling, and thus it is not surprising that ITPase is evolu-tionarily conserved. Expression of ITPase has been reported from all human tissues thus far tested (liver, erythrocytes, spleen, placenta, brain, etc.) with highest levels observed in the heart, thyroid gland and skeletal muscle [48].
In 2010 a GWAS of HCV genotype 1 infected patients revealed that genetic variants associated with
reduced ITPase activity, A variant of rs1127354 and C variant of rs7270101, noted among approximately a third of patients, were associated with reduced hemo-globin decline 4 weeks after the initiation of treatment with pegIFN-α and ribavirin [56]. Further subsequent reports have confirmed this association [57–59], includ-ing studies with protease inhibitors, for example, tela-previr or simeprevir, given in combination with inter-feron and ribavirin [60], and in the interferon-sparing regimen containing faldaprevir, deleobuvir and ribavi-rin [61]. Additionally, reduced ITPase activity has been associated with a greater platelet reduction at week 4, possibly secondary to reduced erythropoietin produc-tion secondary to less anemia [62]. The mechanism by which reduced ITPase activity prevents ribavirin-induced anemia has been hypothesized to be secondary to less ATP depletion as ITP may substitute for GTP in the generation of AMP by adenylosuccinate synthe-tase, which in turn prevents erythrocyte membrane oxidative damage that mediates premature erythrocyte removal [63,64]. Aside from avoidance of ATP deple-tion, reduced ITPase activity additionally has been reported to be associated with reduced ribavirin plasma concentration after 4 weeks of therapy, which also may contribute to protection against anemia [65].
The reports of the impact of reduced ITPase activ-ity on treatment outcome in HCV patients have been conflicting, possibly secondary to variations in ribavi-rin dosing and adherence monitoring. In the setting of HCV genotypes 1, 2 or 3 infected patients treated with higher, weight-based ribavirin dosing in combination with pegIFN-α, reduced ITPase has not been reported to influence treatment outcome [57,59,66]. In contrast, other studies have reported significant associations between reduced ITPase activity and increased likeli-hood of achieving SVR. For example, an Italian study

10.2217/PGS.15.65 Pharmacogenomics (Epub ahead of print) future science group
Review Rembeck & Lagging
enrolling HCV genotype 1–4 infected patients, treated with pegIFN-α and ribavirin, reported increased SVR rates in patients with reduced ITPase activity when all patients were included in the analysis [58]. How-ever, this association was no longer significant after subdivision by infecting HCV genotype [58]. The study by Kurosaki et al. noted an association between increased likelihood of achieving SVR and ITPA A variant of rs1127354 carriage among a subset of Japanese HCV genotype 1 infected patients with the favorable IL28B rs8099917 TT variant, but improved ribavirin adherence could not be ruled out as a possible confounder [67]. In the NORDynamIC study enroll-ing HCV genotype 2 or 3 infected patients receiving lower, conventional 800 mg daily dose of ribavirin in combination with pegIFN-α, a ribavirin-like associa-tion between polymorphisms entailing reduced ITPase activity and treatment efficacy mediated by reduced relapse risk was reported among adherent patients [65].
Patatin-like phospholipase domain-containing 3 (PNPLA3) & impact on liver histopathologyThe PNPLA3 gene is located on chromosome 22 and encodes a 481 amino acid long protein that belongs to the patatin-like phospholipase family, involved in lipid metabolism [68,69]. In humans, PNPLA3 pre-dominately is expressed in the liver, presumably in the hepatocytes, but also is present in skin and adipose tis-sue [70].
In 2008, a GWAS revealed that a genetic variant of PNPLA3, a cytosine to guanine substitution entailing an amino acid change from isoleucine to methionine at residue 148 (PNPLA3 148M), was associated with more pronounced hepatic steatosis in a study enroll-ing more than 2000 individuals of varying ethnicity, including African-Americans, European-Americans and Hispanics [69]. Subsequent reports have confirmed this association between the PNPLA3 148M homo-zygotes, in other words, GG variant of rs738409, and increased incidence of nonalcoholic liver disease and more rapid progression to advanced steatohepatitis and hepatic fibrosis [71,72]. Furthermore, this association has been corroborated in the setting of alcoholic liver disease and hepatocellular carcinoma [73,74]. In patients with HCV infection, homozygotes for the PNPLA3 148M similarly has been reported to be associated with more steatosis, fibrosis, cirrhosis and hepatocellular carcinoma predominantly in Mediterranean popula-tions [75–78].
The mechanism of action through which PNPLA3 148M results in steatosis remains unclear. The criti-cal amino acid change of isoleucine to methionine at residue 148 has been proposed to result in reduced
enzymatic hydrolyses of glycerol lipids, which subse-quently leads to induction of steatosis [68,79]. An alter-native hypothesized mechanism of action is that the substitution entails acyl-transferase activity leading to increased triglyceride synthesis [80].
The frequency of the PNPLA3 148M allele (G vari-ant of rs738409) varies across ethnicities, with the high-est prevalence noted among Hispanics (49%), with an approximate homozygote frequency of 25% [69]. The prevalence of PNPLA3 148M homozygotes in the Ital-ian population has been reported to be approximately 10% (8–14%) [75,78,81]. In contrast, the homozygote frequency in Germany appears to be lower (5.5%) [82].
PNPLA3 & impact on HCV therapyPrevious diverging reports have suggested that homozy-gosity for the PNPLA3 148M variant, aside from being associated with more pronounced steatosis, also either negatively impacts therapeutic outcome [78], or is not independently associated with treatment failure [75,76]. In the NORDynamIC study that enrolled an HCV genotype 2 or 3 infected cohort of Nordic patients, the likelihood of achieving SVR following pegIFN-α and ribavirin treatment was not associated with PNPLA3 genotype, but the low prevalence of homozygosis for the PNPLA3 148M variant in the study cohort may have obscured any possible association [83].
Future clinical relevance of host genetic variantsThe clinical relevance of IL28B genotyping in the set-ting of the new interferon-free HCV regimens likely will diminish, but potentially may prove usefully in tailoring duration and choice of treatment regi-men [23]. In contrast reduced ITPase activity report-edly is associated with a ribavirin-like reduced risk of relapse [65], and thus future studies may determine whether ITPA genetic testing is useful for the selection of patients benefiting from the addition of ribavirin to DAA-based regimens.
The clinical utility of IL28B for prediction of the nat-ural course of HCV-related liver disease may remain of clinically importance, particularly for HCV genotype 3 infected patients. The CC variant of rs12979860 and the TT variant of rs8099917 variants in HCV genotype 3 infected patients reportedly are associated with more pronounced liver histopathology [29,30], and thus these patients may benefit from earlier initiation of treatment. Similarly the utility of PNPLA3 genetic testing may be of benefit for prognostication among certain ethnic groups, for example, Hispanics and south European populations, but appears less benefi-cial among north Europeans, due to the low prevalence of PNPLA3 148M in this population [83].

10.2217/PGS.15.65future science group
Impact of IL28B, ITPA and PNPLA3 genetic variants on therapeutic outcome & progression of HCV infection Review
ConclusionHost genetics, for example, IL28B, ITPA and PNPLA3, impact on therapeutic outcome and liver disease severity in HCV-infected patients, especially in the setting of interferon-based therapy. Their impact on DAA-based treatment for HCV has diminished, but in the setting of shorter duration they appear to influence outcome following otherwise highly effica-cious interferon-sparing regimes. In this light, there-fore, further investigation of their relevance for HCV and other viral infections may be warranted.
Future perspectiveHost genetic factors likely will remain significant for prediction of the natural course of HCV-related liver
disease, and may continue to be of importance for tai-loring therapy, especially with regards to duration and possible benefit of the addition of ribavirin.
Financial & competing interests disclosureThe Swedish Medical Research Council and ALF Funds at Sahl-
grenska University Hospital supported this work. The funders
had no role in the decision to publish or preparation of the
manuscript. The authors have no other relevant affiliations or
financial involvement with any organization or entity with a
financial interest in or financial conflict with the subject mat-
ter or materials discussed in the manuscript apart from those
disclosed.
No writing assistance was utilized in the production of this
manuscript.
References1 Choo QL, Kuo G, Weiner AJ, Overby LR, Bradley DW,
Houghton M. Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. Science 244(4902), 359–362 (1989).
2 Robertson B, Myers G, Howard C et al. Classification, nomenclature, and database development for hepatitis C virus (HCV) and related viruses: proposals for standardization. International Committee on Virus Taxonomy. Arch. Virol. 143(12), 2493–2503 (1998).
3 Simmonds P, Bukh J, Combet C et al. Consensus proposals for a unified system of nomenclature of hepatitis C virus genotypes. Hepatology 42(4), 962–973 (2005).
4 Simmonds P, Holmes EC, Cha TA et al. Classification of hepatitis C virus into six major genotypes and a series of subtypes by phylogenetic analysis of the NS-5 region. J. Gen. Virol. 74(Pt 11), 2391–2399 (1993).
5 Smith DB, Bukh J, Kuiken C et al. Expanded classification of hepatitis C virus into 7 genotypes and 67 subtypes: updated criteria and genotype assignment web resource. Hepatology 59(1), 318–327 (2014).
6 Kiyosawa K, Sodeyama T, Tanaka E et al. Interrelationship of blood transfusion, non-A, non-B hepatitis and hepatocellular carcinoma: analysis by detection of antibody to hepatitis C virus. Hepatology 12(4 Pt 1), 671–675 (1990).
7 Tong MJ, El-Farra NS, Reikes AR, Co RL. Clinical outcomes after transfusion-associated hepatitis C. N. Engl. J. Med. 332(22), 1463–1466 (1995).
8 Roudot-Thoraval F, Bastie A, Pawlotsky JM, Dhumeaux D. Epidemiological factors affecting the severity of hepatitis C virus-related liver disease: a French survey of 6,664 patients. The Study Group for the Prevalence and the Epidemiology of Hepatitis C Virus. Hepatology 26(2), 485–490 (1997).
9 Ge D, Fellay J, Thompson AJ et al. Genetic variation in IL28B predicts hepatitis C treatment-induced viral clearance. Nature 461(7262), 399–401 (2009).
10 Thomas DL, Thio CL, Martin MP et al. Genetic variation in IL28B and spontaneous clearance of hepatitis C virus. Nature 461(7265), 798–801 (2009).
11 Tillmann HL, Thompson AJ, Patel K et al. A polymorphism near IL28B is associated with spontaneous clearance of acute hepatitis C virus and jaundice. Gastroenterology 139(5), 1586–1592, 1592 e1581 (2010).
12 Bochud PY, Bibert S, Negro F et al. IL28B polymorphisms predict reduction of HCV RNA from the first day of therapy in chronic hepatitis C. J. Hepatol. 55(5), 980–988 (2011).
13 Stenkvist J, Sonnerborg A, Weiland O. HCV RNA decline in chronic HCV genotype 2 and 3 during standard of care treatment according to IL28B polymorphism. J. Viral Hepat. 20(3), 193–199 (2013).
Executive summary
Possible molecular mechanism of action of genetic variants in proximity to IL28B• Interleukin 28B (IL28B) genetic variants, located on chromosome 19, impact on the spontaneous clearance,
therapeutic response and natural history of hepatitis C virus (HCV) infection. This may be mediated by inability to produce interferon-λ4 (IFN-λ4), or the presence of less active IFN-λ4 S70 or fully active IFN-λ4 P70.
Inosine triphosphate pyrophosphatasegenetic variants & impact on HCV therapy• Inosine triphosphate pyrophosphatase genetic variants, located on chromosome 20, impact on ribavirin-
associated anemia, as well therapeutic outcome, by means of reduced relapse rate among HCV genotype 2 or 3 infected patients treated with ribavirin 800 mg daily in combination with pegIFN-α.
PNPLA3 & impact on HCV therapy• In HCV-infected populations, homozygosis for the PNPLA3 148M is associated with more pronounced
steatosis, liver fibrosis and hepatocellular carcinoma. However, the frequency of this allele varies considerably across ethnicities, and is more prevalent in Hispanic and Mediterranean populations.

10.2217/PGS.15.65 Pharmacogenomics (Epub ahead of print) future science group
Review Rembeck & Lagging
14 Sarrazin C, Susser S, Doehring A et al. Importance of IL28B gene polymorphisms in hepatitis C virus genotype 2 and 3 infected patients. J. Hepatol. 54(3), 415–421 (2011).
15 Mccarthy JJ, Li JH, Thompson A et al. Replicated association between an IL28B gene variant and a sustained response to pegylated interferon and ribavirin. Gastroenterology 138(7), 2307–2314 (2010).
16 Moghaddam A, Melum E, Reinton N et al. IL28B genetic variation and treatment response in patients with hepatitis C virus genotype 3 infection. Hepatology 53(3), 746–754 (2011).
17 Rauch A, Kutalik Z, Descombes P et al. Genetic variation in IL28B is associated with chronic hepatitis C and treatment failure: a genome-wide association study. Gastroenterology 138(4), 1338–1345, 1345 e1331–1337 (2010).
18 Suppiah V, Moldovan M, Ahlenstiel G et al. IL28B is associated with response to chronic hepatitis C interferon-alpha and ribavirin therapy. Nat. Genet. 41(10), 1100–1104 (2009).
19 Tanaka Y, Nishida N, Sugiyama M et al. Genome-wide association of IL28B with response to pegylated interferon-alpha and ribavirin therapy for chronic hepatitis C. Nat. Genet. 41(10), 1105–1109 (2009).
20 Poordad F, Bronowicki JP, Gordon SC et al. Factors that predict response of patients with hepatitis C virus infection to boceprevir. Gastroenterology 143(3), 608–618, e601–605 (2012).
21 Jacobson I, Catlett I, Marcellin P et al. 1369 Telaprevir substantially improved SVR rates across all IL28B genotypes in the advance trial. J. Hepatol. 54, S542–S543 (2011).
22 Poordad F, Lawitz E, Kowdley K et al. 1399 12-week interferon-free regimen of abt-450/r+ abt-333+ ribavirin achieved svr12 in more than 90% of treatment-naive hcv genotype-1-infected subjects and 47% of previous non-responders. J. Hepatol. 56, S549–S550 (2012).
23 Zeuzem S, Soriano V, Asselah T et al. Faldaprevir and deleobuvir for HCV genotype 1 infection. N. Engl. J. Med. 369(7), 630–639 (2013).
24 Kwo PNG, Nahass R, Bernstein D et al. A Phase 3, Randomised, open-label study to evaluate the efficacy and safety of 8 and 12 weeks of simeprevir plus sofosbuvir in treatment-naive and -experienced patients with chronic HCV genotype 1 infection without cirrhosis: Optimist-1. J. Hepatol. 62(Suppl. 2), S270 (2015).
25 Kumada H, Suzuki Y, Ikeda K et al. Daclatasvir plus asunaprevir for chronic HCV genotype 1b infection. Hepatology 59(6), 2083–2091 (2014).
26 Bochud PY, Bibert S, Kutalik Z et al. IL28B alleles associated with poor hepatitis C virus (HCV) clearance protect against inflammation and fibrosis in patients infected with non-1 HCV genotypes. Hepatology 55(2), 384–394 (2012).
27 Abe H, Ochi H, Maekawa T et al. Common variation of IL28 affects gamma-GTP levels and inflammation of the liver in chronically infected hepatitis C virus patients. J. Hepatol. 53(3), 439–443 (2010).
28 Noureddin M, Wright EC, Alter HJ et al. Association of IL28B genotype with fibrosis progression and clinical
outcomes in patients with chronic hepatitis C: a longitudinal analysis. Hepatology 58(5), 1548–1557 (2013).
29 Rembeck K, Alsio A, Christensen PB et al. Impact of IL28B-related single nucleotide polymorphisms on liver histopathology in chronic hepatitis C genotype 2 and 3. PLoS ONE 7(1), e29370 (2012).
30 Rembeck K, Westin J, Lindh M, Hellstrand K, Norkrans G, Laging M. Association between interleukin-28B-related genetic variants and liver histopathology differs between hepatitis C virus genotypes. Hepatology 56(1), 394 (2012).
31 Ydreborg M, Westin J, Rembeck K et al. Impact of Il28b-related single nucleotide polymorphisms on liver transient elastography in chronic hepatitis C infection. PLoS ONE 8(11), e80172 (2013).
32 Mirandola S, Realdon S, Iqbal J et al. Liver microsomal triglyceride transfer protein is involved in hepatitis C liver steatosis. Gastroenterology 130(6), 1661–1669 (2006).
33 Jackel-Cram C, Babiuk LA, Liu Q. Up-regulation of fatty acid synthase promoter by hepatitis C virus core protein: genotype-3a core has a stronger effect than genotype-1b core. J. Hepatol. 46(6), 999–1008 (2007).
34 Abid K, Pazienza V, De Gottardi A et al. An in vitro model of hepatitis C virus genotype 3a-associated triglycerides accumulation. J. Hepatol. 42(5), 744–751 (2005).
35 Jackel-Cram C, Babiuk LA, Liu Q. Up-regulation of fatty acid synthase promoter by hepatitis C virus core protein: genotype-3a core has a stronger effect than genotype-1b core. J. Hepatol. 46(6), 999–1008 (2007).
36 Abid K, Pazienza V, De Gottardi A et al. An in vitro model of hepatitis C virus genotype 3a-associated triglycerides accumulation. J. Hepatol. 42(5), 744–751 (2005).
37 Westin J, Nordlinder H, Lagging M, Norkrans G, Wejstal R. Steatosis accelerates fibrosis development over time in hepatitis C virus genotype 3 infected patients. J. Hepatol. 37(6), 837–842 (2002).
38 Metz P, Reuter A, Bender S, Bartenschlager R. Interferon-stimulated genes and their role in controlling hepatitis C virus. J. Hepatol. 59(6), 1331–1341 (2013).
39 Alberts B JA, Lewis J, Raff M, Roberts K, Walter P. Molecular Biology Of The Cell, Fouth Edition. Garland Science, NY, USA, 884–886 (2002).
40 Dill MT, Duong FH, Vogt JE et al. Interferon-induced gene expression is a stronger predictor of treatment response than IL28B genotype in patients with hepatitis C. Gastroenterology 140(3), 1021–1031 (2011).
41 Honda M, Sakai A, Yamashita T et al. Hepatic ISG expression is associated with genetic variation in interleukin 28B and the outcome of IFN therapy for chronic hepatitis C. Gastroenterology 139(2), 499–509 (2010).
42 Prokunina-Olsson L, Muchmore B, Tang W et al. A variant upstream of IFNL3 (IL28B) creating a new interferon gene IFNL4 is associated with impaired clearance of hepatitis C virus. Nat. Genet. 45(2), 164–171 (2013).
43 Terczynska-Dyla E, Bibert S, Duong FH et al. Reduced IFNlambda4 activity is associated with improved HCV clearance and reduced expression of interferon-stimulated genes. Nat. Commun. 5, 5699 (2014).
http://www.futuremedicine.com/action/showLinks?pmid=20176026&crossref=10.1053%2Fj.gastro.2010.02.009

10.2217/PGS.15.65future science group
Impact of IL28B, ITPA and PNPLA3 genetic variants on therapeutic outcome & progression of HCV infection Review
44 Hamming OJ, Terczynska-Dyla E, Vieyres G et al. Interferon lambda 4 signals via the IFNlambda receptor to regulate antiviral activity against HCV and coronaviruses. EMBO J. 32(23), 3055–3065 (2013).
45 Jouvin-Marche E, Macek Jilkova Z, Thelu MA et al. Lymphocytes degranulation in liver in hepatitis C virus carriers is associated with IFNL4 polymorphisms and ALT levels. J. Infect. Dis. 209(12), 1907–1915 (2014).
46 Maeda T, Sumi S, Ueta A et al. Genetic basis of inosine triphosphate pyrophosphohydrolase deficiency in the Japanese population. Mol. Genet. Metab. 85(4), 271–279 (2005).
47 Shipkova M, Lorenz K, Oellerich M, Wieland E, Von Ahsen N. Measurement of erythrocyte inosine triphosphate pyrophosphohydrolase (ITPA) activity by HPLC and correlation of ITPA genotype-phenotype in a Caucasian population. Clin. Chem. 52(2), 240–247 (2006).
48 Lin S, Mclennan AG, Ying K et al. Cloning, expression, and characterization of a human inosine triphosphate pyrophosphatase encoded by the itpa gene. J. Biol. Chem. 276(22), 18695–18701 (2001).
49 Chanda PK, Roy J, Banerjee AK. In vitro synthesis of genome length complementary RNA of vesicular stomatitis virus in the presence of inosine 5’-triphosphate. Virology 129(1), 225–229 (1983).
50 Simone PD, Pavlov YI, Borgstahl GE. ITPA (inosine triphosphate pyrophosphatase): from surveillance of nucleotide pools to human disease and pharmacogenetics. Mutat. Res. 753(2), 131–146 (2013).
51 Mohandas T, Sparkes RS, Passage MB, Sparkes MC, Miles JH, Kaback MM. Regional mapping of ADA and ITP on human chromosome 20: cytogenetic and somatic cell studies in an X/20 translocation. Cytogenet. Cell Genet. 26(1), 28–35 (1980).
52 Sumi S, Marinaki AM, Arenas M et al. Genetic basis of inosine triphosphate pyrophosphohydrolase deficiency. Hum. Genet. 111(4–5), 360–367 (2002).
53 Vanderheiden BS. Genetic studies of human erythrocyte inosine triphosphatase. Biochem. Genet. 3(3), 289–297 (1969).
54 Stocco G, Cheok MH, Crews KR et al. Genetic polymorphism of inosine triphosphate pyrophosphatase is a determinant of mercaptopurine metabolism and toxicity during treatment for acute lymphoblastic leukemia. Clin. Pharmacol. Ther. 85(2), 164–172 (2009).
55 Waisertreiger IS, Menezes MR, Randazzo J, Pavlov YI. Elevated levels of DNA strand breaks induced by a base analog in the human cell line with the P32T ITPA variant. J. Nucleic Acids. 2010, pii:872180 (2010).
56 Fellay J, Thompson AJ, Ge D et al. ITPA gene variants protect against anaemia in patients treated for chronic hepatitis C. Nature 464(7287), 405–408 (2010).
57 Eskesen AN, Melum E, Moghaddam A et al. Genetic variants at the ITPA locus protect against ribavirin-induced hemolytic anemia and dose reduction in an HCV G2/G3 cohort. Eur. J. Gastroenterol. Hepatol. 24(8), 890–896 (2012).
58 Clark PJ, Aghemo A, Degasperi E et al. Inosine triphosphatase deficiency helps predict anaemia, anaemia management and response in chronic hepatitis C therapy. J. Viral Hepat. 20(12), 858–866 (2013).
59 Thompson AJ, Santoro R, Piazzolla V et al. Inosine triphosphatase genetic variants are protective against anemia during antiviral therapy for HCV2/3 but do not decrease dose reductions of RBV or increase SVR. Hepatology 53(2), 389–395 (2011).
60 Ogawa E, Furusyo N, Kajiwara E et al. Comparative safety study on severe anemia by simeprevir- versus telaprevir-based triple therapy for chronic hepatitis C. J. Gastroenterol. Hepatol. doi:10.1111/jgh.12945 (2015) (Epub ahead of print).
61 Asselah T, Zeuzem S, Soriano V et al. 1186 itpa gene variants predict hemolytic ribavirin induced anaemia in patients treated with the interferon-free regimen of faldaprevir, BI 207127 and ribavirin in sound-C2. J. Hepatol. 58, S482 (2013).
62 Thompson AJ, Clark PJ, Singh A et al. Genome-wide association study of interferon-related cytopenia in chronic hepatitis C patients. J. Hepatol. 56(2), 313–319 (2012).
63 De Franceschi L, Fattovich G, Turrini F et al. Hemolytic anemia induced by ribavirin therapy in patients with chronic hepatitis C virus infection: role of membrane oxidative damage. Hepatology 31(4), 997–1004 (2000).
64 Hitomi Y, Cirulli ET, Fellay J et al. Inosine triphosphate protects against ribavirin-induced adenosine triphosphate loss by adenylosuccinate synthase function. Gastroenterology 140(4), 1314–1321 (2011).
65 Rembeck K, Waldenstrom J, Hellstrand K et al. Variants of the inosine triphosphate pyrophosphatase gene are associated with reduced relapse risk following treatment for HCV genotype 2/3. Hepatology 59(6), 2131–2139 (2014).
66 Holmes JA, Roberts SK, Ali RJ et al. ITPA genotype protects against anemia during peginterferon and ribavirin therapy but does not influence virological response. Hepatology 59(6), 2152–2160 (2014).
67 Kurosaki M, Tanaka Y, Tanaka K et al. Relationship between polymorphisms of the inosine triphosphatase gene and anaemia or outcome after treatment with pegylated interferon and ribavirin. Antivir. Ther. 16(5), 685–694 (2011).
68 He S, Mcphaul C, Li JZ et al. A sequence variation (I148M) in PNPLA3 associated with nonalcoholic fatty liver disease disrupts triglyceride hydrolysis. J. Biol. Chem. 285(9), 6706–6715 (2010).
69 Romeo S, Kozlitina J, Xing C et al. Genetic variation in PNPLA3 confers susceptibility to nonalcoholic fatty liver disease. Nat. Genet. 40(12), 1461–1465 (2008).
70 Huang Y, He S, Li JZ et al. A feed-forward loop amplifies nutritional regulation of PNPLA3. Proc. Natl Acad. Sci. USA 107(17), 7892–7897 (2010).
71 Valenti L, Al-Serri A, Daly AK et al. Homozygosity for the patatin-like phospholipase-3/adiponutrin I148M polymorphism influences liver fibrosis in patients with nonalcoholic fatty liver disease. Hepatology 51(4), 1209–1217 (2010).

10.2217/PGS.15.65 Pharmacogenomics (Epub ahead of print) future science group
Review Rembeck & Lagging
72 Speliotes EK, Butler JL, Palmer CD et al. PNPLA3 variants specifically confer increased risk for histologic nonalcoholic fatty liver disease but not metabolic disease. Hepatology 52(3), 904–912 (2010).
73 Stickel F, Buch S, Lau K et al. Genetic variation in the PNPLA3 gene is associated with alcoholic liver injury in caucasians. Hepatology 53(1), 86–95 (2011).
74 Corradini SG, Burza MA, Molinaro A, Romeo S. Patatin-like phospholipase domain containing 3 sequence variant and hepatocellular carcinoma. Hepatology 53(5), 1776; author reply 1777 (2011).
75 Valenti L, Rumi M, Galmozzi E et al. Patatin-like phospholipase domain-containing 3 I148M polymorphism, steatosis, and liver damage in chronic hepatitis C. Hepatology 53(3), 791–799 (2011).
76 Trepo E, Pradat P, Potthoff A et al. Impact of patatin-like phospholipase-3 (rs738409 C>G) polymorphism on fibrosis progression and steatosis in chronic hepatitis C. Hepatology 54(1), 60–69 (2011).
77 Cai T, Dufour JF, Muellhaupt B et al. Viral genotype-specific role of PNPLA3, PPARG, MTTP, and IL28B in hepatitis C virus-associated steatosis. J. Hepatol. 55(3), 529–535 (2011).
78 Valenti L, Aghemo A, Stattermayer AF et al. Implications of PNPLA3 polymorphism in chronic hepatitis C patients receiving peginterferon plus ribavirin. Aliment Pharmacol. Ther. 35(12), 1434–1442 (2012).
79 Pirazzi C, Adiels M, Burza MA et al. Patatin-like phospholipase domain-containing 3 (PNPLA3) I148M (rs738409) affects hepatic VLDL secretion in humans and in vitro. J. Hepatol. 57(6), 1276–1282 (2012).
80 Kumari M, Schoiswohl G, Chitraju C et al. Adiponutrin functions as a nutritionally regulated lysophosphatidic acid acyltransferase. Cell Metab. 15(5), 691–702 (2012).
81 Burza MA, Molinaro A, Attilia ML et al. PNPLA3 I148M (rs738409) genetic variant and age at onset of at-risk alcohol consumption are independent risk factors for alcoholic cirrhosis. Liver Int. 34(4), 514–520 (2014).
82 Kollerits B, Coassin S, Beckmann ND et al. Genetic evidence for a role of adiponutrin in the metabolism of apolipoprotein B-containing lipoproteins. Hum. Mol. Genet. 18(23), 4669–4676 (2009).
83 Rembeck K, Maglio C, Lagging M et al. PNPLA 3 I148M genetic variant associates with insulin resistance and baseline viral load in HCV genotype 2 but not in genotype 3 infection. BMC Med. Genet. 13, 82 (2012).
Related Documents











