Crop Protection 23 (2004) 117–125 Impact of environmental factors and fungicides on growth and deoxinivalenol production by Fusarium graminearum isolates from Argentinian wheat Maria L. Ramirez a,b,c , Sofia Chulze b , Naresh Magan a, * a Applied Mycology Group, Cranfield Biotechnology Centre, Cranfield University, Silsoe, Bedford MK45 4DT, UK b Departamento de Microbiologia e Inmunologia, Universidad Nacional de Rio Cuarto, Cordoba, Argentina c Consejo Nacional de Investigaciones Cient ! ıficas y T ! ecnicas (CONICET), Argentina Received 17 April 2003; received in revised form 25 July 2003; accepted 25 July 2003 Abstract The impact of five fungicides (prochloraz, propioconazole, epoxiconazole, tebuconazole and azoxystrobin, 0.5–50 mgg 1 ) on growth of two Fusarium graminearum was evaluated in relation to water activity (a W ; 0.99, 0.97, 0.95) and temperature (15 C and 25 C) on wheat-based media (in vitro). All fungicides reduce growth rates when compared to the control, and this reduction increased as the fungicide concentration increased. In general, none of the isolates was able to grow in the presence of any fungicide treatments at concentrations >15 mg ml 1 , regardless of the a W /temperature regime. The same fungicides were used in the second study on wheat grain (in situ), in order to evaluate the effect of two concentration (0.5, 5 mg ml 1 ), three a W levels (0.995, 0.99 and 0.97) and two temperatures (15 C and 25 C) and their interaction on growth rate and deoxynivalenol (DON) production by F. graminearum. All fungicides showed inhibition of growth at both concentrations in most conditions. The fungicides tested were less effective on grain in controlling growth than in in vitro studies. It was noticeable that at 15 C and 0.99 and 0.97 a W ; two of the fungicides (tebuconazole and epoxiconazole) showed growth stimulation. All fungicides showed DON stimulation or reduction in at least one of the conditions assayed. Our results show that stimulation or reduction in DON production in the presence of fungicides is influenced by complex interactions between a W ; temperature, fungicide concentration and time of incubation in both strains of F. graminearum studied. Such information is critical for effective fungicide control of Fusarium head blight of wheat. r 2003 Elsevier Ltd. All rights reserved. Keywords: Fungicides; Fusarium graminearum; Wheat; Water activity; Deoxynivalenol 1. Introduction In Argentina, the main pathogen associated with Fusarium Head Blight (FHB) is Fusarium graminearum Schw. whose perfect stage is Gibberella zeae (Schw.) Petch. and this is known since 1928 (Galich, 1996). During the last 50 years, 16 FHB epidemics of varying severity occurred in the central-north area. In the last outbreaks (1993) the highest estimated losses reached 50% in areas of no-till over maize stubble. The extent of the damage was magnified by a considerable loss in trading value of the grain resulting from low grain weight, presence of scabby grains, and mycotoxin contamination. During this severe epidemic, deoxyniva- lenol (DON) was the only toxin reported (Dalcero et al., 1997). DON, a member of the trichothecene mycoto- xins, is primarily produced by the genus Fusarium (Sutton, 1982). The occurrence of DON in cereals grain is of great concern for human health, since trichothe- cenes are associated with dermal toxicity, toxic aleukia, food refusal, vomiting and depressed immune functions (Snijders, 1990). The combination of the epidemiological characteris- tics of the pathogen, scarce genetic resistance in the host and strong environmental influence on FHB develop- ment, make it necessary to adopt several integrated measures to decrease disease damage. The use of fungicides is a complementary control measure when weather conditions are conducive to infection from anthesis to harvest. Although there are a number of ARTICLE IN PRESS *Corresponding author. Tel.: +44-01525-863539; fax: +44-0152-5863540. E-mail address: n.magan@cranfield.ac.uk (N. Magan). 0261-2194/$ - see front matter r 2003 Elsevier Ltd. All rights reserved. doi:10.1016/j.cropro.2003.07.005

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Crop Protection 23 (2004) 117–125
ARTICLE IN PRESS
*Correspondin
fax: +44-0152-5
E-mail addres
0261-2194/$ - see
doi:10.1016/j.cro
Impact of environmental factors and fungicides on growthand deoxinivalenol production by Fusarium graminearum
isolates from Argentinian wheat
Maria L. Ramireza,b,c, Sofia Chulzeb, Naresh Magana,*aApplied Mycology Group, Cranfield Biotechnology Centre, Cranfield University, Silsoe, Bedford MK45 4DT, UK
bDepartamento de Microbiologia e Inmunologia, Universidad Nacional de Rio Cuarto, Cordoba, ArgentinacConsejo Nacional de Investigaciones Cient!ıficas y T!ecnicas (CONICET), Argentina
Received 17 April 2003; received in revised form 25 July 2003; accepted 25 July 2003
Abstract
The impact of five fungicides (prochloraz, propioconazole, epoxiconazole, tebuconazole and azoxystrobin, 0.5–50 mg g�1) ongrowth of two Fusarium graminearum was evaluated in relation to water activity (aW; 0.99, 0.97, 0.95) and temperature (15�C and
25�C) on wheat-based media (in vitro). All fungicides reduce growth rates when compared to the control, and this reduction
increased as the fungicide concentration increased. In general, none of the isolates was able to grow in the presence of any fungicide
treatments at concentrations >15mgml�1, regardless of the aW/temperature regime. The same fungicides were used in the second
study on wheat grain (in situ), in order to evaluate the effect of two concentration (0.5, 5 mgml�1), three aW levels (0.995, 0.99 and
0.97) and two temperatures (15�C and 25�C) and their interaction on growth rate and deoxynivalenol (DON) production by
F. graminearum. All fungicides showed inhibition of growth at both concentrations in most conditions. The fungicides tested were
less effective on grain in controlling growth than in in vitro studies. It was noticeable that at 15�C and 0.99 and 0.97 aW; two of thefungicides (tebuconazole and epoxiconazole) showed growth stimulation. All fungicides showed DON stimulation or reduction in at
least one of the conditions assayed. Our results show that stimulation or reduction in DON production in the presence of fungicides
is influenced by complex interactions between aW; temperature, fungicide concentration and time of incubation in both strains of
F. graminearum studied. Such information is critical for effective fungicide control of Fusarium head blight of wheat.
r 2003 Elsevier Ltd. All rights reserved.
Keywords: Fungicides; Fusarium graminearum; Wheat; Water activity; Deoxynivalenol
1. Introduction
In Argentina, the main pathogen associated withFusarium Head Blight (FHB) is Fusarium graminearum
Schw. whose perfect stage is Gibberella zeae (Schw.)Petch. and this is known since 1928 (Galich, 1996).During the last 50 years, 16 FHB epidemics of varyingseverity occurred in the central-north area. In the lastoutbreaks (1993) the highest estimated losses reached50% in areas of no-till over maize stubble. The extent ofthe damage was magnified by a considerable loss intrading value of the grain resulting from low grainweight, presence of scabby grains, and mycotoxin
g author. Tel.: +44-01525-863539;
863540.
s: [email protected] (N. Magan).
front matter r 2003 Elsevier Ltd. All rights reserved.
pro.2003.07.005
contamination. During this severe epidemic, deoxyniva-lenol (DON) was the only toxin reported (Dalcero et al.,1997). DON, a member of the trichothecene mycoto-xins, is primarily produced by the genus Fusarium
(Sutton, 1982). The occurrence of DON in cereals grainis of great concern for human health, since trichothe-cenes are associated with dermal toxicity, toxic aleukia,food refusal, vomiting and depressed immune functions(Snijders, 1990).The combination of the epidemiological characteris-
tics of the pathogen, scarce genetic resistance in the hostand strong environmental influence on FHB develop-ment, make it necessary to adopt several integratedmeasures to decrease disease damage. The use offungicides is a complementary control measure whenweather conditions are conducive to infection fromanthesis to harvest. Although there are a number of

ARTICLE IN PRESS
able 1
ctive ingredient, product name and company of the fungicides during
vitro and in vivo studies
ctive ingredient Product
name
Company
ebuconazole 250 g l�1 (25% ww�1) Folicurs Bayer
ropioconazole 250 g l�1 (21.7% ww�1) Tilts Ciba-Geige
poxiconazole 125 g l�1 (12.1% ww�1) Opuss Basf
rochloraz 450 g l�1 (45% ww�1) Sportaks AgrEvo
zoxystrobin 250 g l�1 (23.9% ww�1) Amistars Zeneca
M.L. Ramirez et al. / Crop Protection 23 (2004) 117–125118
compounds with in vitro activity against FHB patho-gens, control of this disease in the field has often beendifficult (Milus and Parson, 1994). In both laboratorystudies with pure cultures of phytopathogens and fieldtrials with crop plants, the overall evidence concerningthe effectiveness of fungicide is contradictory. There isalso growing concern that sub-lethal doses of somefungicides may lead to an increase in mycotoxinproduction by Fusarium species (Milus and Parson,1994; D’Mello et al., 1998b). Such effects have beenobserved in several in vitro studies with some com-pounds inhibiting the growth of Fusarium species butcausing increased trichothecene production (Moss andFrank, 1985; Placinta et al., 1996; D’Mello et al., 1997,1998a; Matthies et al., 1999; Hope et al., 2002).However, all previous reports, with the exception ofHope et al. (2002), took no account of the interactionsbetween the efficacy of the fungicides and key environ-mental factors such as aW or temperature. These havebeen demonstrate to be key parameters determininggermination, growth and mycotoxin production byseveral Fusarium species (Magan and Lacey, 1984;Mar!ın et al., 1995a, b, 1996).The objectives of this work were to evaluate the
efficacy of five fungicides (tebuconazole, propiconazole,epoxiconazole, prochloraz and azoxystrobin) underdifferent interacting water availability and temperatureregimes on (i) growth and (ii) DON production by twostrains of F. graminearum isolated from Argentinianwheat.
2. Materials and methods
2.1. Fungal isolates
Two strains of F. graminearum (RC17-2, and RC22-2)isolated from Argentinian wheat were used. Thesestrains are held in the Universidad Nacional de RioCuarto, Argentina (RC) Culture Collection. Cultureswere maintained in 15% glycerol at �80�C.
2.2. In vitro effect of environmental conditions and
fungicides on growth
Five different fungicides were used (Table 1). Dilu-tions of the fungicides were prepared by mixingappropriate amounts of each fungicide (based onpercentage of the active ingredient) in sterile distilledwater and stored at 4�C until used.A 2% milled wheat agar was used in these studies.
The aW of the basic medium was adjusted to 0.99, 0.97and 0.95 with glycerol. The media were autoclaved at120�C for 20min before cooling to 50�C at which timethe fungicides were added to obtain the requiredconcentration (0.5, 15, 25 and 50 mgml�1). Flasks of
T
A
in
A
T
P
E
P
A
molten media were thoroughly shaken, prior to pouringinto 9 cm sterile Petri dishes, to ensure that an evendispersion of the fungicide treatment was obtained.Treatments and replicates were inoculated centrally witha 3mm diameter agar disk taken from the margin of a5–7-day-old colony of each isolate grown on maltextract agar (MEA). Inoculated plates of the same aWwere sealed in polyethylene bags. The aW of representa-tive samples of media were checked with an AqualabSeries 3 (Labcell Ltd., Basingstoke, Hants, UK). Theexperiments were carried out with three replicates pertreatment and repeated once at 15�C and 25�C for 20days.Assessment of growth was made every day during the
incubation period and two diameters of the growingcolonies measured at right angles to each other until thecolony reached the edge of the plate. The radii of thecolonies were plotted against time, and linear regressionapplied in order to obtain the growth rate (mmday�1) asthe slope of the line.
2.3. In situ effect of environmental conditions and
fungicides on growth and don production
Wheat grains were irradiated with 12 kGy of gammairradiation and stored aseptically at 4�C. The graincontained no fungal infection or contamination but hadretained germinative capacity. The initial aW of the grainwas 0.766.Four hundred gram of irradiated wheat was weighed
into sterile beakers and rehydrated to the required aW(0.995, 0.99 and 0.97) by the addition of sterile distilledwater using a moisture absorption curve. An appro-priated aliquot of the water was replaced by a suitablefungicide treatment (tebuconazole, propiconazole, epox-iconazole, prochloraz and azoxystrobin), to give thegrain a final concentration of 0.5 and 5 mg g�1 of eachfungicide. At the same time controls containing wateronly were also prepared. Flasks were subsequentlyrefrigerated at 4�C for 48 h with periodic shaking toallow absorption and equilibration. At the end of thisperiod the aW was checked with an Aqualab Series 3(Labcell Ltd., Basingstoke, Hants, UK). The rehydrated
ARTICLE IN PRESS
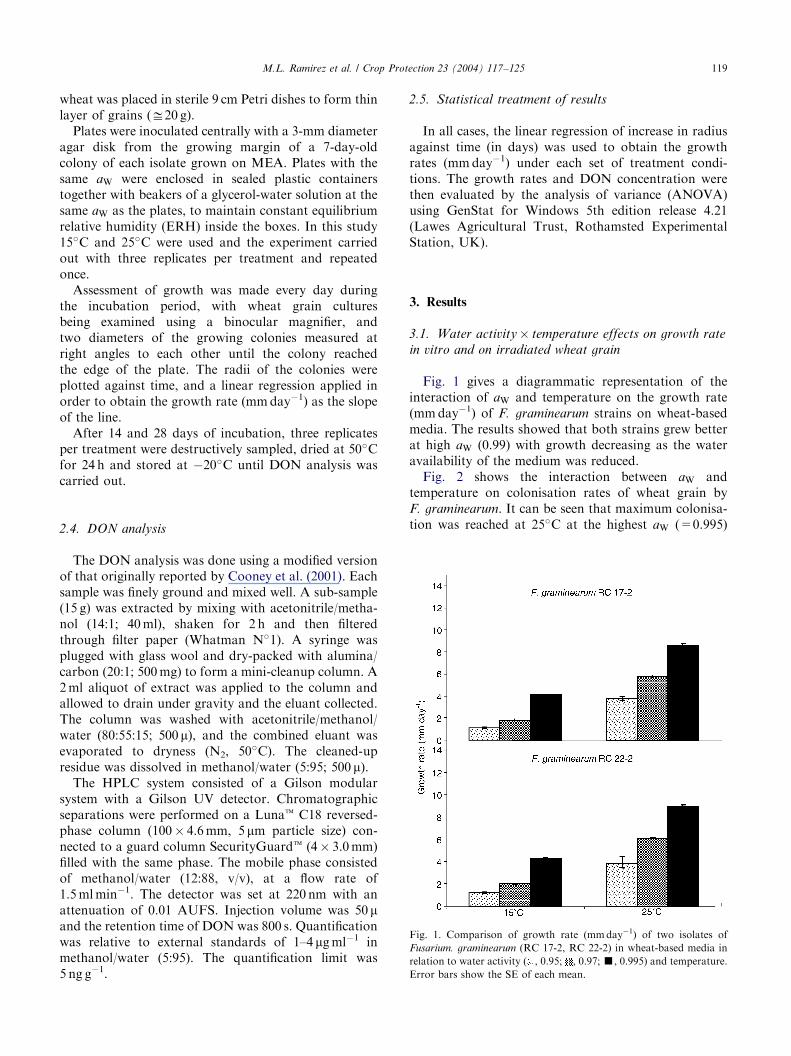
Fig. 1. Comparison of growth rate (mmday�1) of two isolates of
Fusarium. graminearum (RC 17-2, RC 22-2) in wheat-based media in
relation to water activity ( , 0.95; , 0.97;’, 0.995) and temperature.
Error bars show the SE of each mean.
M.L. Ramirez et al. / Crop Protection 23 (2004) 117–125 119
wheat was placed in sterile 9 cm Petri dishes to form thinlayer of grains (D20 g).Plates were inoculated centrally with a 3-mm diameter
agar disk from the growing margin of a 7-day-oldcolony of each isolate grown on MEA. Plates with thesame aW were enclosed in sealed plastic containerstogether with beakers of a glycerol-water solution at thesame aW as the plates, to maintain constant equilibriumrelative humidity (ERH) inside the boxes. In this study15�C and 25�C were used and the experiment carriedout with three replicates per treatment and repeatedonce.Assessment of growth was made every day during
the incubation period, with wheat grain culturesbeing examined using a binocular magnifier, andtwo diameters of the growing colonies measured atright angles to each other until the colony reachedthe edge of the plate. The radii of the colonies wereplotted against time, and a linear regression applied inorder to obtain the growth rate (mmday�1) as the slopeof the line.After 14 and 28 days of incubation, three replicates
per treatment were destructively sampled, dried at 50�Cfor 24 h and stored at �20�C until DON analysis wascarried out.
2.4. DON analysis
The DON analysis was done using a modified versionof that originally reported by Cooney et al. (2001). Eachsample was finely ground and mixed well. A sub-sample(15 g) was extracted by mixing with acetonitrile/metha-nol (14:1; 40ml), shaken for 2 h and then filteredthrough filter paper (Whatman N�1). A syringe wasplugged with glass wool and dry-packed with alumina/carbon (20:1; 500mg) to form a mini-cleanup column. A2ml aliquot of extract was applied to the column andallowed to drain under gravity and the eluant collected.The column was washed with acetonitrile/methanol/water (80:55:15; 500 m), and the combined eluant wasevaporated to dryness (N2, 50�C). The cleaned-upresidue was dissolved in methanol/water (5:95; 500 m).The HPLC system consisted of a Gilson modular
system with a Gilson UV detector. Chromatographicseparations were performed on a Lunat C18 reversed-phase column (100� 4.6mm, 5 mm particle size) con-nected to a guard column SecurityGuardt (4� 3.0mm)filled with the same phase. The mobile phase consistedof methanol/water (12:88, v/v), at a flow rate of1.5mlmin�1. The detector was set at 220 nm with anattenuation of 0.01 AUFS. Injection volume was 50 mand the retention time of DON was 800 s. Quantificationwas relative to external standards of 1–4 mgml�1 inmethanol/water (5:95). The quantification limit was5 ng g�1.
2.5. Statistical treatment of results
In all cases, the linear regression of increase in radiusagainst time (in days) was used to obtain the growthrates (mmday�1) under each set of treatment condi-tions. The growth rates and DON concentration werethen evaluated by the analysis of variance (ANOVA)using GenStat for Windows 5th edition release 4.21(Lawes Agricultural Trust, Rothamsted ExperimentalStation, UK).
3. Results
3.1. Water activity� temperature effects on growth rate
in vitro and on irradiated wheat grain
Fig. 1 gives a diagrammatic representation of theinteraction of aW and temperature on the growth rate(mmday�1) of F. graminearum strains on wheat-basedmedia. The results showed that both strains grew betterat high aW (0.99) with growth decreasing as the wateravailability of the medium was reduced.Fig. 2 shows the interaction between aW and
temperature on colonisation rates of wheat grain byF. graminearum. It can be seen that maximum colonisa-tion was reached at 25�C at the highest aW (=0.995)
ARTICLE IN PRESSM.L. Ramirez et al. / Crop Protection 23 (2004) 117–125120
level examined. Mycelial extension rates across wheatgrain were higher in comparison with the levels obtainedin wheat-based media. The analysis of variance of theeffect of single (isolate, aW and temperature) two- andthree-way interaction showed that all factors alone andonly the aW� temperature interaction were statisticallysignificant on both wheat-based agar medium andirradiated wheat grain (Table 2).
Fig. 2. Comparison of growth rate (mmday�1) of two isolates of
Fusarium graminearum (RC 17-2, RC 22-2) in irradiated wheat grain in
relation to water activity ( , 0.97; , 0.99;’, 0.995) and temperature.
Error bars show the SE of each mean.
Table 2
Analysis of variance of effect of water activity (aW), temperature (T), and
graminearum in wheat-based media and wheat grain
Source of variation dfa Wheat-base media
MSb
aW 2 48.220
T 1 127.383
i 1 0.256
aW �T 2 2.804
aW � i 2 0.0292
T � i 1 0.0389
aW �T � i 2 0.00623
�Significant Po0:001:��Significant Po0:05:
aDegrees of freedom.bMean square.cF-Snedecor.
3.2. Effect of fungicides, water activity, temperature and
their interactions on growth
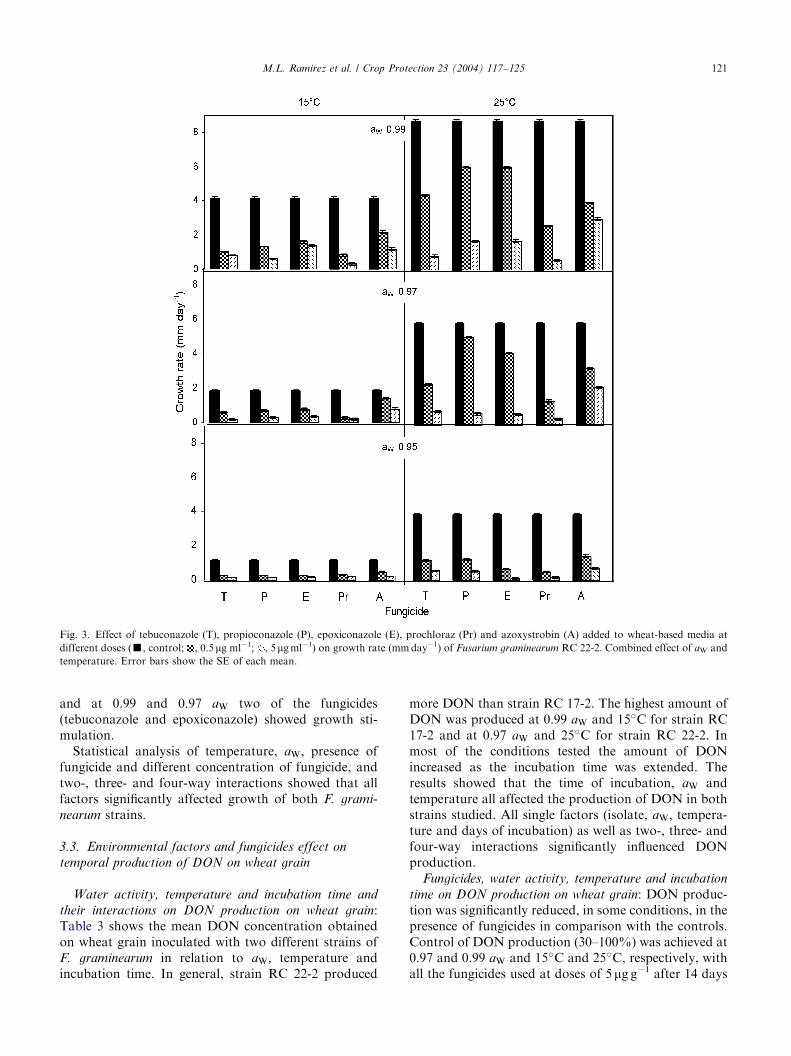
In vitro studies: The impact of five fungicides(tebuconazole, propiconazole, epoxiconzole, prochlorazand azoxystrobin) at 0.5–50 mgml�1 concentration ongrowth of F. graminearum was evaluated in relation toaW (0.99, 0.97 and 0.95) and temperature (15�C and25�C) in wheat-based media (Fig. 3). Inhibition ofmycelial growth increased as fungicide concentrationwas increased. In general, none of the isolates were ableto grow in the presence of any fungicide at concentra-tions >15 mgml�1, regardless of the aW/temperatureregime (data not shown). Most of the fungicides weremore effective in controlling fungal growth at 25�Crather than 15�C.Analysis of variance revealed that at 25�C all single
factors (fungicide, aW; isolate and fungicide concentra-tions) as well as two-, three- and four-way interactionshad a significant influence on growth rates. However,the same factors at 15�C had no significant influences ongrowth rates (P > 0:05).
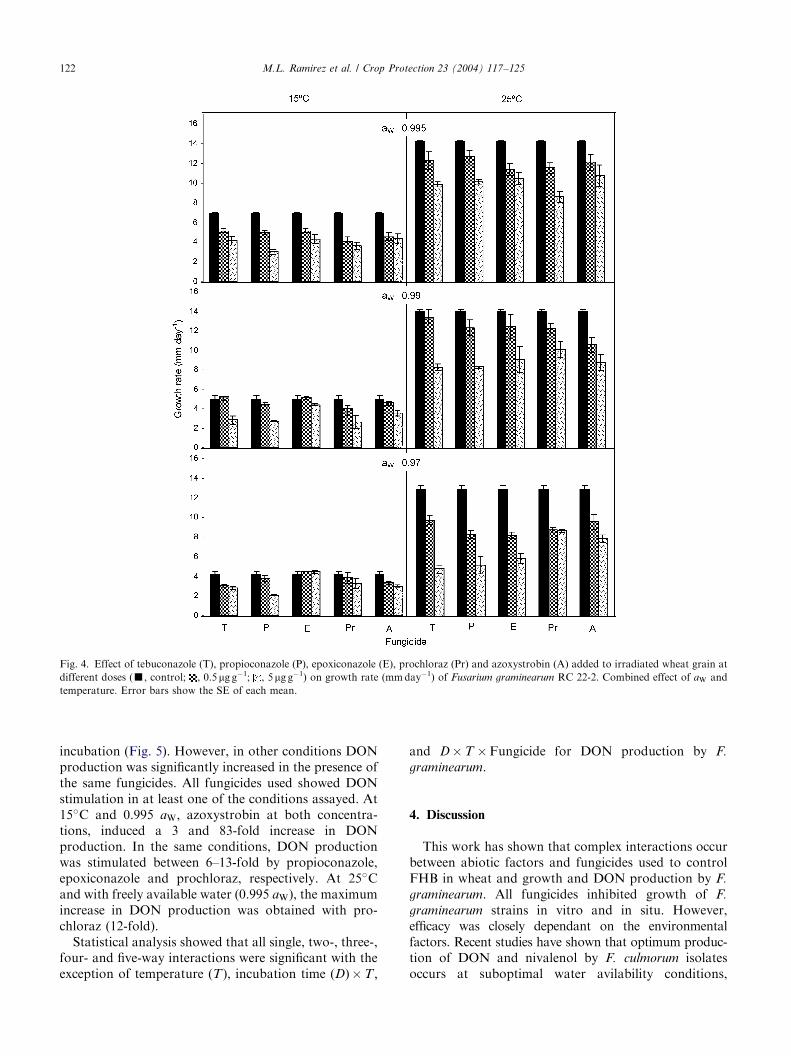
In situ studies: The same fungicides were used in thestudy on wheat grain, in order to evaluate the effect oftwo concentrations (0.5 and 5 mg g�1), three aW (0.995,0.99 and 0.97) and two temperatures (15�C and 25�C)and their interaction on growth rate by F. graminearum.All fungicides showed inhibition of growth at bothconcentrations in most conditions. The fungicides testedwere less effective on grain in controlling growth than inin vitro studies (Fig. 4).The inhibition of growth was dependent on the
concentration of fungicide used, and this generallyincreased as the concentration increased regardless ofthe temperature/aW regime used. At 25�C, for all fun-gicide treatments the levels of inhibition increased asaW decreased. At 15�C the effect was less pronounced,
different isolates (i), and their interactions on growth of Fusarium
Wheat grain
Fc MS F
1620.597� 13.497 137.421�
4281.144� 595.976 6068.116�
8.598�� 0.994 10.120��
94.252� 2.103 21.411�
0.983 0.323 3.289
1.308 0.120 1.226
0.209 0.138 1.403
ARTICLE IN PRESS
Fig. 3. Effect of tebuconazole (T), propioconazole (P), epoxiconazole (E), prochloraz (Pr) and azoxystrobin (A) added to wheat-based media at
different doses (’, control; , 0.5 mg ml�1; , 5mgml�1) on growth rate (mmday�1) of Fusarium graminearum RC 22-2. Combined effect of aW and
temperature. Error bars show the SE of each mean.
M.L. Ramirez et al. / Crop Protection 23 (2004) 117–125 121
and at 0.99 and 0.97 aW two of the fungicides(tebuconazole and epoxiconazole) showed growth sti-mulation.Statistical analysis of temperature, aW; presence of
fungicide and different concentration of fungicide, andtwo-, three- and four-way interactions showed that allfactors significantly affected growth of both F. grami-
nearum strains.
3.3. Environmental factors and fungicides effect on
temporal production of DON on wheat grain
Water activity, temperature and incubation time and
their interactions on DON production on wheat grain:Table 3 shows the mean DON concentration obtainedon wheat grain inoculated with two different strains ofF. graminearum in relation to aW; temperature andincubation time. In general, strain RC 22-2 produced
more DON than strain RC 17-2. The highest amount ofDON was produced at 0.99 aW and 15�C for strain RC17-2 and at 0.97 aW and 25�C for strain RC 22-2. Inmost of the conditions tested the amount of DONincreased as the incubation time was extended. Theresults showed that the time of incubation, aW andtemperature all affected the production of DON in bothstrains studied. All single factors (isolate, aW; tempera-ture and days of incubation) as well as two-, three- andfour-way interactions significantly influenced DONproduction.
Fungicides, water activity, temperature and incubation
time on DON production on wheat grain: DON produc-tion was significantly reduced, in some conditions, in thepresence of fungicides in comparison with the controls.Control of DON production (30–100%) was achieved at0.97 and 0.99 aW and 15�C and 25�C, respectively, withall the fungicides used at doses of 5 mg g�1 after 14 days
ARTICLE IN PRESS
Fig. 4. Effect of tebuconazole (T), propioconazole (P), epoxiconazole (E), prochloraz (Pr) and azoxystrobin (A) added to irradiated wheat grain at
different doses (’, control; , 0.5 mg g�1; , 5mg g�1) on growth rate (mmday�1) of Fusarium graminearum RC 22-2. Combined effect of aW and
temperature. Error bars show the SE of each mean.
M.L. Ramirez et al. / Crop Protection 23 (2004) 117–125122
incubation (Fig. 5). However, in other conditions DONproduction was significantly increased in the presence ofthe same fungicides. All fungicides used showed DONstimulation in at least one of the conditions assayed. At15�C and 0.995 aW; azoxystrobin at both concentra-tions, induced a 3 and 83-fold increase in DONproduction. In the same conditions, DON productionwas stimulated between 6–13-fold by propioconazole,epoxiconazole and prochloraz, respectively. At 25�Cand with freely available water (0.995 aW), the maximumincrease in DON production was obtained with pro-chloraz (12-fold).Statistical analysis showed that all single, two-, three-,
four- and five-way interactions were significant with theexception of temperature (T), incubation time (D)�T ;
and D�T �Fungicide for DON production by F.
graminearum.
4. Discussion
This work has shown that complex interactions occurbetween abiotic factors and fungicides used to controlFHB in wheat and growth and DON production by F.
graminearum. All fungicides inhibited growth of F.
graminearum strains in vitro and in situ. However,efficacy was closely dependant on the environmentalfactors. Recent studies have shown that optimum produc-tion of DON and nivalenol by F. culmorum isolatesoccurs at suboptimal water avilability conditions,
ARTICLE IN PRESS
Fig. 5. Effect of tebuconazole ( ), propioconazole ( ), epoxiconazole ( ), prochloraz (&) and azoxystrobin (’) added to irradiated wheat grain at
different concentration on deoxynivalenol production (ng g�1) by Fusarium graminearum RC 22-2 after 14 days incubation, at different aW levels and
temperatures. Means of all replicates in experiments.
Table 3
Mean deoxynivalenol concentrations (mg g�1) on wheat grains at three water activities (aW) inoculated with two different strains of Fusarium
graminearum at 15�C and 25�C after 14 and 28 days of incubation (7standard error of mean)
F. graminearum T (�C) Deoxynivalenol (ng g�1)
Strain 0.97 aW 0.99 aW 0.995 aW
14 days 28 days 14 days 28 days 14 days 28 days
RC 17-2 15 159 (729) 457 (731) 431 (747) 22,006 (72754) 13,674 (73583) 2763 (7426)
25 1516 (7398) 777 (735) 27 (73) 247 (712 1203 (757) 1053 (761)
RC 22-2 15 nda 122 (728) 181 (725) 20,997 (71535) 323 (717) 948 (736)
25 2556 (7155) 147,724 (72,630) 182 (716) 540 (712) 1250 (7114) 1826 (7107)
aNot detected.
M.L. Ramirez et al. / Crop Protection 23 (2004) 117–125 123
and sometimes only after 7–14 days incubation (Hopeand Magan, 2003). Studies on irradiated wheat grainshowed that fungicides were less effective than in wheat-
base media. In vitro screening on wheat-base mediashow that doses X15 mgml�1 of fungicide can inhibitgrowth completely. Our results suggest that the effect of

ARTICLE IN PRESSM.L. Ramirez et al. / Crop Protection 23 (2004) 117–125124
fungicides on artificial substrate may not accuratelyrepresent the real situation on a natural substrate. Theuse of irradiated wheat seeds that have retainedviability, may be a more useful system for fungicidescreening.Among the fungicides used in the present study,
tebuconazole, propiconazole and epoxiconazole belongto the triazole group, and prochloraz is an imidazolederivate. These type of fungicides interfere with themetabolism of fungal pathogens, mainly by inhibition ofergosterol biosynthesis (Hewitt, 1998). Ergosterol bio-synthesis-inhibiting fungicides often cause strikingmorphological malformations, irregular cell wall thick-ening and excessive branching in fungi. They fail toinhibit spore germination or initial cell growth and dryweight increase. They have no immediate effects onrespiratory metabolism or macromolecule synthesis.Even though ergosterol biosynthesis is quite sensitiveto inhibition by these toxicants, mycelial growth andvarious aspect of metabolism (respiration, protein andnucleic acid synthesis) are only mildly affected for aperiod of time after the curtailment of the synthesis ofsterol (Siegel, 1981). The other fungicide studied wasaxozystrobin. This compound belongs to the strobiluringroup of agrochemical fungicides. These metabolites actby blocking electron transport in the mitochondrialrespiratory chain in fungi. This severely reduces theaerobic energy production, thereby inhibiting growth ofthe fungus and thus forming the basis for the use ofstrobilurins as effective fungicides. With the new modeof action the strobilurins are an important addition tothe existing fungicide range, particularly for cereals inwhich recent broad-spectrum fungicide products havebeen largely based on sterol biosynthesis inhibitors.Some of these fungicides (tebuconazole, propiconazoleand prochloraz) were included in the present studybecause they have been reported to reduce the severityof FHB-induced losses in Argentina (Galich, 1996).Although the fungicides have different modes of action,in the present studies all were able to inhibit fungalgrowth in almost all the conditions assayed. Only twofungicides (tebuconazole and epoxiconazole) during thein situ study, in two different conditions (0.99 and 0.97aW at 15�C) showed growth stimulation.All fungicides showed DON stimulation or reduction
in at least one of the conditions assayed. Our resultsshowed that the stimulation or reduction in DONproduction in the presence of fungicides is influenced bycomplex interactions between aW; temperature, fungi-cide concentration and time of incubation in bothstrains of F. graminearum studied.Previous in vitro studies with F. graminearum have
shown an increased production of toxin after exposureto sub-lethal concentrations of fungicides. Matthies et al.(1999) reported that production of 3-Acetylated deox-ynivalenol was increased when compared with mycelium
growth for F. graminearum cultured in a liquid mediumcontaining sub-inhibitory concentration of tebuconazole(0.1–0.5 mgml�1). In contrast, the toxin was reduceddramatically when the concentration of the fungicide inthe medium was increased to 1 mgml�1 at whichconcentration mycelial growth was significantly inhib-ited. Matthies et al. (1999) found that concentrations of>3 and >2 mgml�1 of tebuconazole alone and withtriadimenol inhibited growth of F. graminearum inculture media. At sub-lethal concentrations these andtriadimenol alone, stimulated zearalenol and DONproduction. The second fungicide increased thesemycotoxins 17- and 30-fold. Increased mycotoxinproduction following exposure to sub-lethal doses offungicide have also been demonstrated with F. sporo-
trichoides and F. culmorum (Moss and Frank, 1985;Placinta et al., 1996; D’Mello et al., 1997).Unfortunately, all these studies were carried out only
in media (solid and liquid) with freely available water(0.995 aW), which makes direct comparison with thepresent work more difficult. Also the previous reporttook no account of the interactions between the efficacyof the fungicides examined and aW or temperature,which have been demonstrated to be key parametersdetermining germination, growth and mycotoxin pro-duction by several Fusarium species (Magan and Lacey,1984; Marin et al., 1995a,b, 1996).There is only one comparable study on the effect of
environmental factors and fungicides on growth of F.
culmorum and DON production (Hope et al., 2002). Intheir study, DON productions by isolates of F.
culmorum from different parts of Europe were signifi-cantly increased at reduced aW in the presence ofepoxiconazole and propioconazole.The mechanisms by which the fungicides stimulate
toxin production in Fusarium spp. are not known. Itmay be assumed that in the presence of sub-lethalconcentrations of certain fungicides the fungal strainsrespond to this stress by increased production ofsecondary metabolites including mycotoxins, as apossible mechanism.The influence of aW; temperature, fungicides and time
of incubation on growth and DON production by F.
graminearum on wheat-based media and on irradiatedwheat grains has been demonstrated in this study.However, a wide range of fungi (pathogens andsaprophytes) occur on wheat, which may have varyingtolerance to such fungicides in the field ecosystem andmay influence the efficacy of the best treatment (Maganand Lacey, 1986). Liggitt et al. (1997) have demon-strated, in a preliminary in vitro study, the differentialeffect of fungicides on growth of common saprophyticflora of wheat ears including Cladosporium and Alter-
naria species and F. culmorum. Further work isnecessary to understand the combined effects ofenvironmental factors and fungicides on growth of

ARTICLE IN PRESSM.L. Ramirez et al. / Crop Protection 23 (2004) 117–125 125
fungal pathogens and saprophytic populations presenton wheat ears and mycotoxin production.
Acknowledgements
The authors are grateful to the Consejo Nacional deInvestigaciones Cientificas y Tecnicas de la RepublicaArgentina (CONICET) for the financial support.
References
Cooney, J.M., Lauren, D.R., di Menna, M.E., 2001. Impact of
competitive fungi on trichothecene production by Fusarium
graminearum. J. Agric. Food Chem. 49, 522–526.
Dalcero, A., Torres, A., Etcheverry, M., Chulze, S., Varsavsky, E.,
1997. Occurrence of deoxynivalenol and Fusarium graminearum in
Argentinian wheat. Food Additives Contam. 14, 11–14.
D’Mello, J.P.F., Macdonald, A.M.C., Bonte, L., 1997. The effects of
difenoconazole on 3-acetyl deoxynivalenol synthesis by Fusarium
culmorum: implications for cereal quality. In: Crop Protection &
Food Quality: Meeting Customers Needs, Proceedings of BCPC
and ANPP Conference, Kent, UK, pp. 463–466.
D’Mello, J.P.F., Macdonald, A.M.C., Postel, D., Dijksma, W.T.P.,
1998a. 3-Acetyl deoxynivalenol and esterase production in a
fungicide-insensitive strain of Fusarium culmorum. Mycotoxin
Res. 14, 8–9.
D’Mello, J.P.F., Macdonald, A.M.C., Postel, D., Dijksma, W.T.P.,
Dujardin, A., Placinta, C.M., 1998b. Pesticide use and mycotoxin
production in Fusarium and Aspergillus phytopathogens. Eur. J.
Plant Pathol. 104, 741–751.
Galich, MT., 1996. Fusarium Head blight in Argentina. In: Duvin,
H.J., Gilchrist, L., Reeves, J., McNab, A. (Eds.), Fusarium Head
Scab: Global Status and Future Prospects. Proceedings of a
Workshop Held at CIMMYT, Mexico.
Hewitt, A.F., 1998. Fungicides in Plant Protection. CAB International,
Wallingford.
Hope, R., Magan, N., 2003. Two dimensional environmental profiles
of growth, deoxynivalenol and nivalenol production by Fusarium
culmorum on a wheat-based substrate. Lett. Appl. Microbiol. 37,
70–74.
Hope, RJ., Colleate, A., Baxster, E.S., Magan, N., 2002. Interactions
between environmental stress and fungicides effect growth and
mycotoxin production by Fusarium culmorum isolates from wheat
grain. Eur. J. Plant Pathol. 108, 685–690.
Liggitt, J., Jenkinson, P., Parry, D.W., 1997. The role of saprophytic
microflora in the development of Fusarium head blight of winter
wheat caused by Fusarium culmorum. Crop Prot. 16, 679–685.
Magan, N., Lacey, J., 1984. Water relations of some Fusarium species
from infected wheat ears and grain. Trans. Br. Mycol. Soc. 83,
281–285.
Magan, N., Lacey, J., 1986. The phylloplane microflora of ripening
wheat and effect of late fungicide aplications. Ann. Appl. Biol. 109,
117–128.
Mar!ın, S., Sanchis, V., Vinas, I., Canela, R., Magan, N., 1995a. Effect
of water activity and temperature on growth and fumonisin B1 and
B2 production by Fusarium proliferatum and F. moniliforme on
maize grain. Lett. Appl. Microbiol. 21, 298–301.
Mar!ın, S., Sanchis, V., Magan, N., 1995b. Water activity, temperature
and pH effects on growth of Fusarium moniliforme and Fusarium
proliferatum isolates from maize. Can. J. Microbiol. 41, 1063–1070.
Mar!ın, S., Sanchis, V., Teixeido, R., Saenz, A.J., Ramos, A.J., Magan,
N., 1996. Water activity, temperature relationships and micro-
conidial germination of Fusarium moniliforme and Fusarium
proliferatum from maize. Can. J. Microbiol. 42, 1045–1050.
Matthies, A., Walker, F., Buchenauer, H., 1999. Interference of select
fungicides, plant growth retardants as well as piperonyl butoxide
and 1-aminobenzotriazole in trichothecene production of Fusarium
graminearum (strain 4528) in vitro. Z. Pflanzenkr. Pflanzenschutz
106, 198–212.
Milus, E.A., Parson, C.E., 1994. Evaluation of foliar fungicides
for controlling Fusarium head blight of wheat. Plant Dis. 78,
697–699.
Moss, M.O., Frank, J.M., 1985. Influence of the fungicide tridemorh
on T-2 toxin production by Fusarium sporotrichioides. Trans. Br.
Mycol. Soc. 84, 585–590.
Placinta, C.M., Macdonald, A.M.C., D’Mello, J.P.F., Harling, R.,
1996. The influence of carbendazim on mycotoxin production in
Fusarium sporotrichioides. In: Proceedings of The Brighton Crop
Protection Conference British Crop Protection Council, Farnham,
UK, pp. 415–416.
Siegel, M.R., 1981. Sterol-inhibiting fungicides: effects on sterol
biosynthesis and sites of action. Plant Dis. 65, 986–989.
Snijders, C.H.A., 1990. Fusarium head blight and mycotoxin
contamination of wheat, a review. Neth. J. Plant Pathol. 96,
187–198.
Sutton, J.C., 1982. Epidemeology of wheat blight and maize ear rot
caused by Fusarium graminearum. Can. J. Plant Pathol. 4,
195–209.
Related Documents











