Impact of Boron Deficiency on Xenopus laevis A Summary of Biological Effects and Potential Biochemical Roles DOUGLAS J. FORT,* ,1,2 ROBERT L. ROGERS, 1 DANIEL W. MCLAUGHLIN, 1 CHRIS M. SELLERS, 1 AND CHRISTIAN L. SCHLEKAT 3 1 Fort Environmental Laboratories, Stillwater, OK 74074; 2 Department of Zoology, Oklahoma State University, Stillwater, OK; and 3 U.S. Borax, Valencia, CA Received March 15, 2002; Accepted April 15, 2002 ABSTRACT The toxicity of boron has been understood for many years. However, limited data currently exist concerning the nutritional essentiality of B in chordates. Results from an ongoing research program evaluating the nutri- tional essentiality of B in the South African clawed frog, Xenopus laevis, found that X. laevis fed a low-B diet in a low-B culture media produced a substantially higher number of necrotic eggs and fertilized embryos than frogs fed a boron-sufficient diet. Markedly decreased embryo cell counts at mid-blastula transition and an increased frequency of abnormal gastrula- tion were also noted in embryos from adult frogs fed the B-deficient diet. By 96 h of development, none of the larvae collected from the B-deficient adults and maintained in low-boron culture media developed normally. Reproductive effects associated with B deficiency in female Xenopus included ovary atrophy, oocyte necrosis, and incomplete oocyte matura- tion. In males, a decrease in testis weight and sperm count was noted. These studies suggest that these adverse effects resulting from B deficiency could be found during gametogenesis, gamete maturation, embryonic development, and larval maturation. The studies also confirmed that B deficiency was capable of interrupting the X. laevis life cycle. Additional studies evaluating the role of B in the thyroid axis and the oocyte plasma membrane progesterone receptor provide the first line of direct evidence for a biochemical role of boron in X. laevis. Combined together, this Biological Trace Element Research 117 Vol. 90, 2002 © Copyright 2002 by Humana Press Inc. All rights of any nature, whatsoever, reserved. 0163-4984/02/90(1–3)–0117 $20.00 *Author to whom all correspondence and reprint requests should be addressed.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Impact of Boron Deficiency on Xenopus laevis
A Summary of Biological Effects and Potential Biochemical Roles
DOUGLAS J. FORT,*,1,2 ROBERT L. ROGERS,1
DANIEL W. MCLAUGHLIN,1 CHRIS M. SELLERS,1
AND CHRISTIAN L. SCHLEKAT 3
1Fort Environmental Laboratories, Stillwater, OK 74074;2Department of Zoology, Oklahoma State University, Stillwater,
OK; and 3U.S. Borax, Valencia, CA
Received March 15, 2002; Accepted April 15, 2002
ABSTRACT
The toxicity of boron has been understood for many years. However,limited data currently exist concerning the nutritional essentiality of B inchordates. Results from an ongoing research program evaluating the nutri-tional essentiality of B in the South African clawed frog, Xenopus laevis,found that X. laevis fed a low-B diet in a low-B culture media produced asubstantially higher number of necrotic eggs and fertilized embryos thanfrogs fed a boron-sufficient diet. Markedly decreased embryo cell counts atmid-blastula transition and an increased frequency of abnormal gastrula-tion were also noted in embryos from adult frogs fed the B-deficient diet.By 96 h of development, none of the larvae collected from the B-deficientadults and maintained in low-boron culture media developed normally.Reproductive effects associated with B deficiency in female Xenopusincluded ovary atrophy, oocyte necrosis, and incomplete oocyte matura-tion. In males, a decrease in testis weight and sperm count was noted.These studies suggest that these adverse effects resulting from B deficiencycould be found during gametogenesis, gamete maturation, embryonicdevelopment, and larval maturation. The studies also confirmed that Bdeficiency was capable of interrupting the X. laevis life cycle. Additionalstudies evaluating the role of B in the thyroid axis and the oocyte plasmamembrane progesterone receptor provide the first line of direct evidencefor a biochemical role of boron in X. laevis. Combined together, this
Biological Trace Element Research 117 Vol. 90, 2002
© Copyright 2002 by Humana Press Inc.All rights of any nature, whatsoever, reserved.0163-4984/02/90(1–3)–0117 $20.00
*Author to whom all correspondence and reprint requests should be addressed.

research program provides firm evidence that B is nutritionally essential inX. laevis.
Index Entries: Boron; nutritional essentiality; Xenopus; reproduction;development.
INTRODUCTION
Many trace elements that are beneficial or essential can be toxic athigh concentrations. Examples include copper, zinc, selenium, andchromium. This duality is brought about when homeostatic mechanisms,which control internal concentrations of trace elements so that sufficientquantities are available to fulfill certain biological functions, are over-whelmed at high concentrations, allowing excess trace elements to com-pete and interfere with other functions.
In general, the progress of understanding the duality of trace elementbehavior in biological systems has focused initially on the adverse andtoxic effects. For example, for over 50 yr selenium has been known or sus-pected to cause toxicity in livestock, through the ingestion of vegetationthat is naturally enriched with selenium (1). However, effects of seleniumdeficiency in livestock and in human populations were not recognized foranother 25–30 yr (1). The first biological role of selenium, which is to pro-vide the catalytic capability to the enzyme glutathione peroxidase, was notconfirmed until the mid-1980s.
Boron is an element that appears to be within this progression ofunderstanding; that is, although the toxic effects of boron are well docu-mented, understanding of boron deficiency and essentiality are at an earlystage. Boron, which occurs naturally bound to oxygen in the form of com-pounds known as borates, is known to cause adverse effects to microbes,insects, aquatic invertebrates, fish, birds, mammals, and humans. Becauseof their broad use as industrial minerals, borates and boric acid (BO3
–)have extensive toxicological dossiers. The relative toxicity of borates islow; the oral median lethal dose (LD50) of boric acid and disodium tetrab-orate for laboratory rodents is greater than 2000 mg/kg, which is similarto table salt, NaCl. The toxicology of boron, including effects ranges,observed end points, and toxicokinetics, has been extensively reviewedfrom human health (2–4) and ecological (5,6) perspectives.
In comparison to boron toxicology research, relatively little researchhas been conducted to evaluate boron’s essentiality. The major exceptionto this is in the field of agriculture, as boron has been recognized to beessential for the growth of vascular plants since the 1920s. Boron essen-tiality to plants was the subject of a recent collection of reviews (7).
Several factors have contributed to the difficulty in establishing boronas an essential nutrient to animals. One is the absence of a long-livedradioisotope that can be used as a tracer (e.g., to demonstrate the existenceof homeostatic mechanisms). A second factor is that boron is widely dis-
118 Fort et al.
Biological Trace Element Research Vol. 90, 2002

tributed. Boron is present in every environmental phase, including water,soil, minerals, and the atmosphere, and is also a constituent of laboratoryglassware (e.g., borosilicate glass). Therefore, the ubiquitous distributionof boron has presented challenges for creating conditions of boron defi-ciency. This is particularly true for heterotrophic organisms, for which theprimary source of boron is the diet.
A strategy was developed in the early 1990s to evaluate boron essen-tiality for aquatic organisms that utilize external development. Exposingdeveloping embryos in an aqueous environment allowed tighter control ofboron-exposure concentrations and diminished the importance of creatinga reduced boron diet. Additionally, the use of boron-specific resins suc-cessfully reduced boron concentrations to levels below 1 µg/L. The use ofboron-free polyethylene laboratory ware maintained the low concentra-tions. These steps produced an environment in which it was possible toexpose developing embryos to a broad range of boron concentrations.
The initial outcome of these studies showed that boron stimulated thegrowth of zebrafish (8) and rainbow trout (9), offering preliminary evi-dence for the importance of boron in vertebrate development. Growth ofrainbow trout was significantly greater at 0.119 mg B/L compared with0.02 mg B/L, and zebrafish embryos that were exposed to <0.002 mg B/Lexhibited significantly greater mortality than those exposed to 0.5 mg B/L.Although these descriptive studies demonstrated that the presence ofboron in aqueous media was beneficial, and, for zebrafish, necessary fornormal growth and development, these experiments offered limitedinsight into the biochemical mechanisms in which boron participates.
Identification of biochemical mechanisms is important in the demon-stration of nutritional necessity, both for fundamental and practical reasons.Most importantly, knowing how cells use boron helps in the understandingof the biochemical and physiological consequences of boron deficiency,knowledge of which is critical to the development of surveillance methodsfor determining the effects and occurrence of boron deficiency in humanpopulations. An understanding of boron’s biochemical function may alsoreveal mechanisms of toxicity. A mechanistic understanding of the toxiceffects of boron would reduce the uncertainty that currently exists whenextrapolating from toxic effects observed for boron in laboratory mammalsto effects in humans. Finally, identification of biochemical mechanisms thatdepend on boron is of practical importance to the field of nutrition. Orga-nizations that establish recommended daily allowances of vitamins andminerals (e.g., the US Food and Nutrition Board) use operational defini-tions for determining whether or not these substances are required for ade-quate nutrition. The first is a demonstration of a requirement for a specificbiochemical function. This can include the adverse clinical consequencesthat result from deficiency of the nutrient or a reduction in the risk ofchronic degenerative diseases that accompany adequate intake of the nutri-ent. The second factor is a method to measure the level of an indicatorlinked to the essential function of the nutrient (i.e., a marker).
Review of Boron Essentiality in Xenopus 119
Biological Trace Element Research Vol. 90, 2002

The ideal biological system for demonstrating boron essentialitywould allow for the simultaneous determination of the consequences ofboron deficiency and the identification of the biochemical basis for theseconsequences. The present article describes the use of such a model sys-tem, utilizing the African clawed frog Xenopus laevis. This amphibian is anideal test organism for these studies because it is easily cultured undercontrolled laboratory settings and because the biochemical factors affect-ing its progression through discrete developmental stages are well under-stood and easily measured. Furthermore, reproduction in adults, growth,and development of early life stages of X. laevis have been shown to beresponsive to low boron concentrations.
The present article describes effects of low boron concentrations onadult X. laevis reproduction and on the development of frog embryos andprovides evidence for a biochemical explanation for the manifestation ofdevelopmental effects.
BORON DEFICIENCY STUDIES IN XENOPUS
Establishing Boron Deficiency Status: Diet Development and the Experimental Approach to Boron Depletion
DietsLow-B (–B, 45 ± 6 µg B/kg) and B-supplemented (+B, 1850 ± 42 µg
B/kg) purified diets used to establish B deficiency were developed by Hunt(10) and obtained from the US Department of Agriculture Human NutritionResearch Center (Grand Forks, ND). These diets were originally preparedfor studying the effects of a low-B diet in rats. The general compositions ofthe diets used in these studies are provided in Table 1. In each of the studiesreported here, the composition of the –B and +B diets was identical, with theexception of the B content. In an effort to demonstrate that the +B dietsupported growth, reproduction, and development of X. laevis consistentwith the American Society of Testing and Materials (ASTM) diet (11) ofground beef liver (ASTM BL), a nonpurified ASTM BL dietary treatmentwas used in this study. ASTM BL was obtained from a local meat packagingfacility, ground, and frozen as patties until used. The ASTM BL wassupplemented with the following nutrients: 15.0 IU vitamin A palmitate, 4.0IU vitamin D3, 0.05 IU vitamin E succinate, 0.35 mg vitamin C, 0.005 mgthiamin, 0.006 mg riboflavin, 0.08 mg niacin, 0.004 mg vitamin B6, and 0.02µg vitamin B12 per gram of BL. The level of B in the ASTM BL diet wasapproximately 310 ± 10.9 µg B/kg, which is now known to be adequate forXenopus development and reproduction.
Culture WaterTo reduce B levels in the culture water used for both the embryos
and adults, amphibian culture water was prepared with ACS-grade salts(> 99.5% pure) (Sigma Chemical Co., St. Louis, MO) and deionized water
120 Fort et al.
Biological Trace Element Research Vol. 90, 2002

(< 0.1 ± 0 µg B/kg) and stored in a polyethylene plastic vessel. Low-Bculture water contained approx 0.6 ± 0.1 µg B/L. Dechlorinated tap water,in which the frogs are maintained under standard conditions (11), containsapprox 116 ± 15.2 µg B/L. Standard adequate-B culture water (11), whichis prepared with standard-grade salts (> 95% pure) and deionized waterand stored in borosilicate glass, contains 5.6 ± 0.3 µg B/L. Because X. laevisare a purely aquatic species regardless of life stage, both young and adultsare continually maintained in a low-B environment.
Animal HousingAdult Dietary Administration. Adult frogs purchased from Xenopus® I
(Ann Arbor, MI) specifically for this phase of experiments were cultured inthe low-B culture water for 120 d. Each of the three diets was fed through-out the 120-d depletion period. Laboratory equipment that minimizedcontamination (i.e., polyethylene plastic vessels instead of glass) was uti-lized throughout these studies, and the frogs were isolated from atmos-pheric B. The adult frogs for each diet were housed in separatepolyethylene plastic tanks and separated by sex.
Design. For each of the studies reviewed, 10 newly acquired male andfemale X. laevis were used per dietary regimen. Consistent culture condi-
Review of Boron Essentiality in Xenopus 121
Biological Trace Element Research Vol. 90, 2002
Table 1Composition of the Defined Boron Diets1
1 A batch of the diet was supplemented with orthoboric acid (H3BO3; Puratronic;Johnson Matthey Chemicals Ltd., Aesar, Seabrook, NH) in a boron premix (0.0046 gH3BO3 [800 µg B]/g anhydrous dextrose [ICN Biochemicals, Cleveland, OH]). Thefinal concentration of boron in the low boron diet was approx 45 µg B/kg. The boronsupplemented diet contained approx 1850 µg B/kg diet. Corn (ICN Pharmaceuticals,Costa Mesa, CA) was acid washed in accordance with Hunt (10). Caesin was obtainedfrom Tecklad, Madison, WI.
2 No analysis available.

tions and experimental design were maintained for all studies reported inthis review. The frogs were fed the –B, +B, and ASTM BL diets for 120 d, atwhich time, each of the 10 adult frogs was bred, and developmental effectstests were performed as described in the following subsections.
Boron Deficiency and Early Embryo–Larval Development
AdultsThe overall appearance of female and male frogs was similar among
each of the three diet groups throughout the 120-d depletion period(12,13). No overt differences in health or in the presence of disease werereported. Furthermore, the weight gain obtained in both the female andmale frogs during the 120-d B-depletion period was similar among each ofthe three diets (Dunnett’s test, p = 0.05). The weight gain in females fed theBL, +B, or –B diets was 61.2 ± 5.3, 59.3 ± 6.2, and 58.2 ± 7.3 g, respectively.The weight gain in males fed the BL, +B, or –B diets was 42.6 ± 3.8, 45.8 ±6.3, and 40.9 ± 8.2 g, respectively. These gains in weight, including thedifference in weight gain between female and male frogs, are consistentwith normal weight gain in Xenopus (12–16).
Reproductive Response, Fertilization, and Early Developmental EffectsA summary of B levels in various Xenopus tissue samples is provided in
Tables 2 and 3 (13–16). B concentrations in embryos from adult frogs fed theASTM BL diet (890 ± 54 µg B/kg) were similar to those of the +B embryos.Boron concentrations in embryos from adult frogs fed the +B diet for 120 dwere markedly higher than in embryos from frogs fed the –B diet (1120 ± 84µg B/kg vs 10 ± 2 µg B/kg). These results demonstrated that the +B diet wascapable of providing a significantly greater amount of boron to the adultfrogs and, subsequently, to the embryos. These results also showed thatmaternal and paternal consumption of the –B diet resulted in the productionof embryos with very low B concentrations relative to control values.
The impact of the BL, +B, and –B diets on X. laevis reproductive per-formance and early development following the 120-d dietary treatment isdepicted in Table 4 (13). Egg mass size, rates of fertilization, oocyte necro-sis, abnormal gastrulation, embryonic viability, and total embryo cellcounts were similar in the +B and ASTM BL diet groups. Thus, based onthese results, it appears that the purified rat diet used in this study willsupport normal reproductive performance in X. laevis. The egg mass sizefrom the frogs fed the –B diet for 120 d did not differ from (Dunnett’s test,p > 0.05) the egg mass collected from the frogs fed the +B diet. Fertilizationrates also did not differ (Dunnett’s test, p > 0.05) in embryos from frogs fedthe +B and –B diets.
The frequencies of necrosis, abnormal gastrulation, and embryonicviability were significantly influenced by maternal and paternal dietarytreatment. The frequencies of necrosis and of abnormal gastrulation were
122 Fort et al.
Biological Trace Element Research Vol. 90, 2002

strikingly greater (Dunnett’s test, p < 0.05) in embryos collected from frogsfed the –B diet when compared to embryos collected from frogs fed the +Bdiet. Similarly, the frequency of abnormal gastrulation in embryos fromadult frogs fed the –B diet was markedly greater than in embryos fromadults fed the +B diet. The percentage of normal-appearing blastula-stageembryos collected from adult frogs fed the –B diet was significantly less(Dunnett’s test, p < 0.05) than in embryos from frogs fed the +B diet. Thelatter values for the +B and ASTM BL dietary groups were similar to the
Review of Boron Essentiality in Xenopus 123
Biological Trace Element Research Vol. 90, 2002
Table 2Boron Levels in Tissues in Xenopus Administered a Low-Boron,
Boron-Supplemented, or Boron-Adequate Diet for 120 d1
1 Results of boron analyses in respective tissues; n = 8, 7, and 5 for oocytes,ovary, and testis samples, respectively, for each dietary regimen; data reported asmean values ± SE.
2 BL = beef liver (310 µg B/kg) (traditional diet); –B = low-boron diet (45 µgB/kg); +B = boron-supplemented diet (1850 µg B/kg).
3 Statistically less than +B dietary group (Dunnett’s test, p > 0.05).

viability frequencies for historical control embryos collected from adultsmaintained under standard culturing conditions (12–16). Although noappreciable differences based on dietary regimens were detected atNieuwkoop and Faber (NF) stages 6.5 and 7, the mean stage 7.5 embryocell count of embryos collected from frogs fed the –B diet was significantlylower (48.4 ± 5.6) (p < 0.05) than the mean cell count of embryos from the+B diet-fed frogs (98.3 ± 2.6) (13).
Developmental EffectsThe influence of low-B and B-supplemented culture media on 4-d
embryo lethality frequencies on embryos collected from frogs fed the +Band –B diets in each respective trial is presented in Table 5 (13). Overall, thefrequency of mortality in each of the treatments was significantly greater(Dunnett’s test, p < 0.05) in embryos derived from the adults fed the –B dietthan in embryos from frogs fed the +B diet. In these 120-d depletionstudies, 10 µg B/L supplementation to the embryos collected from adultsfed the –B diet did not reduce the rate of mortality, an observationconsistent with a previous study in which a 28-d boron-depletion periodwas used (12).
The effect of low-boron and boron-supplemented culture media on 4-d embryo malformation frequencies with embryos collected from frogs fedthe BL, +B, and –B diets is presented in Table 5 (13). The malformation fre-
124 Fort et al.
Biological Trace Element Research Vol. 90, 2002
Table 3Boron Levels in Immature and Mature Oocytes from Females
Maintained on a Low-Boron, Boron-Supplemented, or Boron-Adequate Dietary Regimen1
1 Normal-appearing or necrotic oocytes from adult females (n = ~50/sample) maintainedon respective diets for 120 d; data reported as mean value ± SE.
2 Staged in accordance with previous methods (11–16).3 BL = beef liver (310 µg B/kg) (traditional diet); –B = low-boron diet (45 µg B/kg); +B =
boron-supplemented diet (1850 µg B/kg).4 N = necrotic.

quencies were markedly higher (p < 0.05) in embryos derived from adultsfed the –B diet than in embryos from frogs fed the BL or +B diet. Supple-mentation of B at a concentration of 10 µg B/L to the embryos collectedfrom adults fed the –B diet did not effectively reduce the frequency of mal-formation when compared to the low-boron treatment. Thus, B repletionvia the culture water was ineffective in reducing the teratogenic effects oflow B in the embryos collected from adults fed the –B diet.
The Concentration–Response RelationshipConcentrations of ≤ 3.0 µg B/L (0.3 µM) and ≥ 53,850 µg B/L (4,980
µM) in embryo culture media induced statistically significant increases in
Review of Boron Essentiality in Xenopus 125
Biological Trace Element Research Vol. 90, 2002
Table 4Influence of 120-d Administration of Beef Liver, Boron-Supplemented,
and Low-Boron Diets on Reproduction and Early Development in X. laevis
1 Determined by the method of Nieuwkoop and Faber (17), representing sequentialembryonic stages.
2 Expressed in hours postfertilization3 BL = beef liver; +B = boron supplemented; –B = low boron. 4 Statistically different from BL diet (Dunnett’s test, p = 0.05).5 Statistically different from +B diet (Dunnett’s test, p = 0.05).
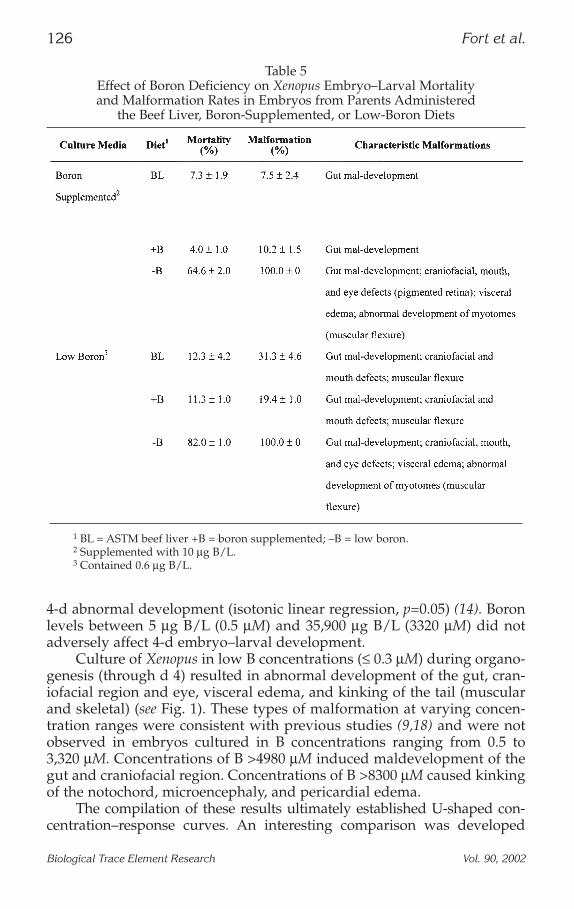
4-d abnormal development (isotonic linear regression, p=0.05) (14). Boronlevels between 5 µg B/L (0.5 µM) and 35,900 µg B/L (3320 µM) did notadversely affect 4-d embryo–larval development.
Culture of Xenopus in low B concentrations (≤ 0.3 µM) during organo-genesis (through d 4) resulted in abnormal development of the gut, cran-iofacial region and eye, visceral edema, and kinking of the tail (muscularand skeletal) (see Fig. 1). These types of malformation at varying concen-tration ranges were consistent with previous studies (9,18) and were notobserved in embryos cultured in B concentrations ranging from 0.5 to3,320 µM. Concentrations of B >4980 µM induced maldevelopment of thegut and craniofacial region. Concentrations of B >8300 µM caused kinkingof the notochord, microencephaly, and pericardial edema.
The compilation of these results ultimately established U-shaped con-centration–response curves. An interesting comparison was developed
126 Fort et al.
Biological Trace Element Research Vol. 90, 2002
Table 5Effect of Boron Deficiency on Xenopus Embryo–Larval Mortality and Malformation Rates in Embryos from Parents Administered
the Beef Liver, Boron-Supplemented, or Low-Boron Diets
1 BL = ASTM beef liver +B = boron supplemented; –B = low boron.2 Supplemented with 10 µg B/L.3 Contained 0.6 µg B/L.
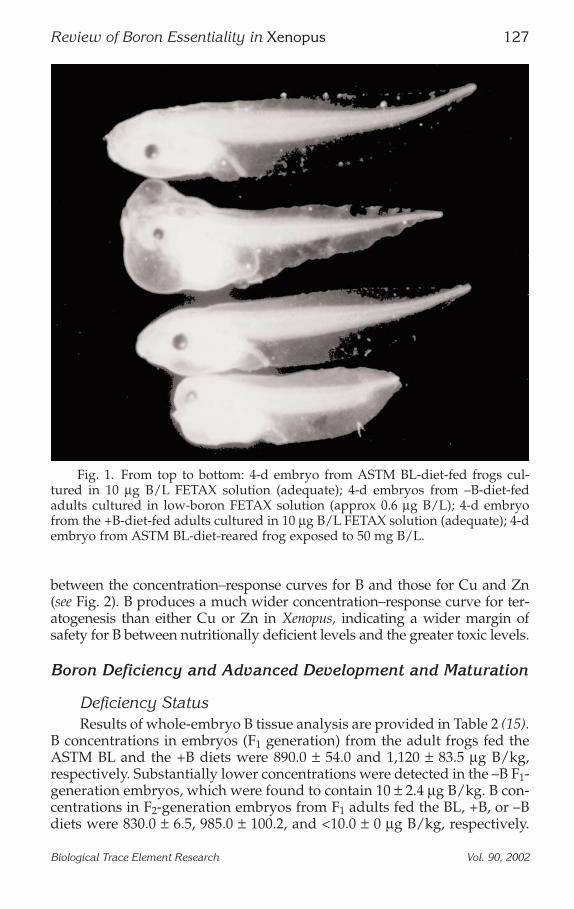
between the concentration–response curves for B and those for Cu and Zn(see Fig. 2). B produces a much wider concentration–response curve for ter-atogenesis than either Cu or Zn in Xenopus, indicating a wider margin ofsafety for B between nutritionally deficient levels and the greater toxic levels.
Boron Deficiency and Advanced Development and Maturation
Deficiency StatusResults of whole-embryo B tissue analysis are provided in Table 2 (15).
B concentrations in embryos (F1 generation) from the adult frogs fed theASTM BL and the +B diets were 890.0 ± 54.0 and 1,120 ± 83.5 µg B/kg,respectively. Substantially lower concentrations were detected in the –B F1-generation embryos, which were found to contain 10 ± 2.4 µg B/kg. B con-centrations in F2-generation embryos from F1 adults fed the BL, +B, or –Bdiets were 830.0 ± 6.5, 985.0 ± 100.2, and <10.0 ± 0 µg B/kg, respectively.
Review of Boron Essentiality in Xenopus 127
Biological Trace Element Research Vol. 90, 2002
Fig. 1. From top to bottom: 4-d embryo from ASTM BL-diet-fed frogs cul-tured in 10 µg B/L FETAX solution (adequate); 4-d embryos from –B-diet-fedadults cultured in low-boron FETAX solution (approx 0.6 µg B/L); 4-d embryofrom the +B-diet-fed adults cultured in 10 µg B/L FETAX solution (adequate); 4-dembryo from ASTM BL-diet-reared frog exposed to 50 mg B/L.
These results demonstrated that the +B diet was capable of providing asignificantly greater amount of B to the adult frogs and developing larvae.These results also showed that maternal and paternal consumption of the–B diet resulted in the production of embryos with very low B concentra-tions relative to controls. Furthermore, only slight differences in embryo Blevels in the ASTM BL and +B groups were detected between the F1 and F2generations. Only a slight generational reduction in embryo B levels wasdetected in the –B treatment, primarily as a result of the marked reductionin embryo B that occurred in the F1 generation.
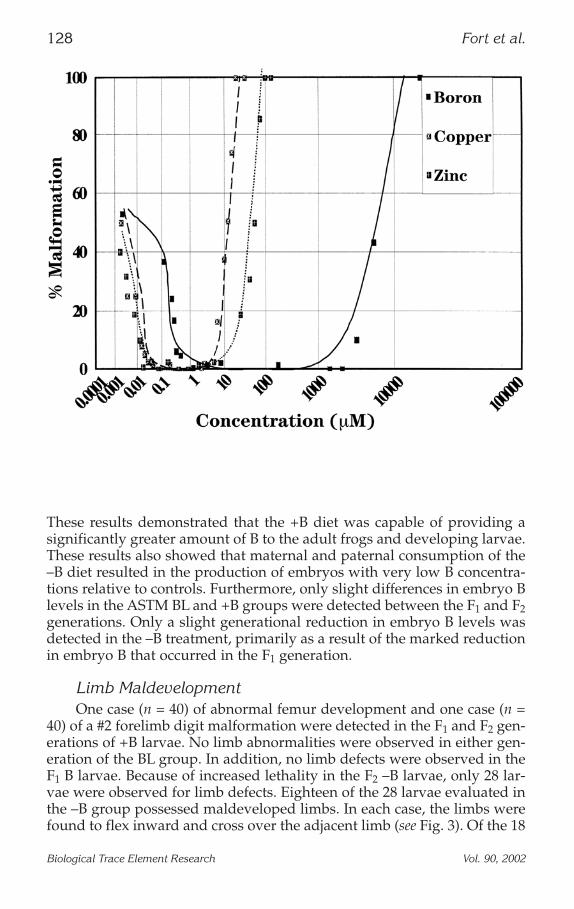
Limb MaldevelopmentOne case (n = 40) of abnormal femur development and one case (n =
40) of a #2 forelimb digit malformation were detected in the F1 and F2 gen-erations of +B larvae. No limb abnormalities were observed in either gen-eration of the BL group. In addition, no limb defects were observed in theF1 B larvae. Because of increased lethality in the F2 –B larvae, only 28 lar-vae were observed for limb defects. Eighteen of the 28 larvae evaluated inthe –B group possessed maldeveloped limbs. In each case, the limbs werefound to flex inward and cross over the adjacent limb (see Fig. 3). Of the 18
128 Fort et al.
Biological Trace Element Research Vol. 90, 2002
Fig. 2. Comparison of 4-d embryo–larval malformation concentration–responserelationships for boron, copper, and zinc in X. laevis.
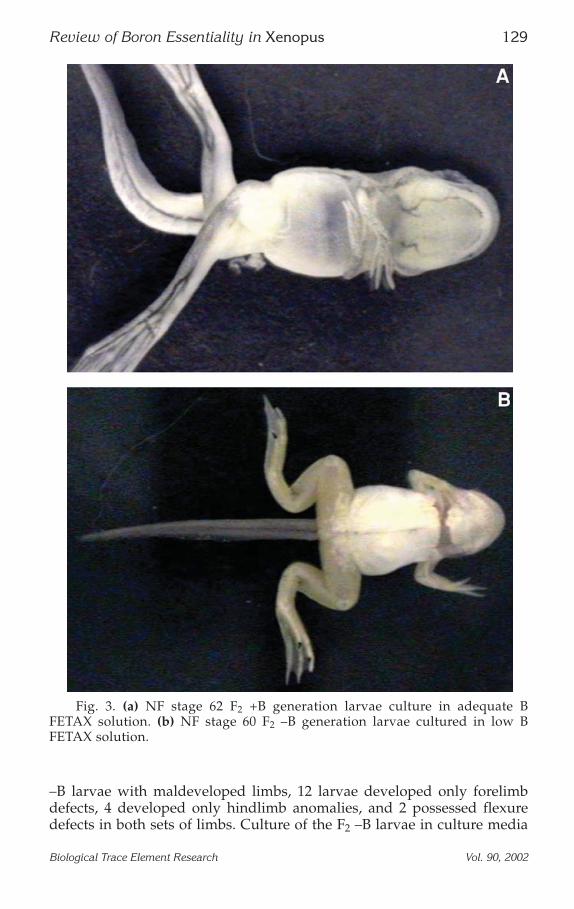
–B larvae with maldeveloped limbs, 12 larvae developed only forelimbdefects, 4 developed only hindlimb anomalies, and 2 possessed flexuredefects in both sets of limbs. Culture of the F2 –B larvae in culture media
Review of Boron Essentiality in Xenopus 129
Biological Trace Element Research Vol. 90, 2002
Fig. 3. (a) NF stage 62 F2 +B generation larvae culture in adequate BFETAX solution. (b) NF stage 60 F2 –B generation larvae cultured in low BFETAX solution.
supplemented with 100 µg B/L did not appreciably reduce the incidenceof malformation (16 of 28 malformed). In addition, coadministration of0.25 U/mL glutathione/glutathione peroxidase did not reduce the rate oflimb maldevelopment in the F2 –B larvae (17 malformed, n = 28).
Maturation, Tail Resorption, and Thyroid ActivityThe impact of maintaining parental adults and their progeny on each
of the traditional ASTM BL, –B, and +B diets on the rate of tail resorptionfrom NF stage 60 to 66 was also performed to determine the impact of Bdeficiency on metamorphic climax (14). In these studies, tail-fin length wasmeasured at each of NF stages 63, 64, 65, and 66. Under standard condi-tions, the tail resorbs at a consistent rate between stages 63 and 65 and iscompletely resorbed by stage 66, as noted in larvae from adults adminis-tered the traditional ASTM BL diet. The addition of 100 µg/L of thyroxingreatly enhances the rate of tail resorption, such that the tail is completelyresorbed by stage 64. The rate and extent of tail resorption in larvae admin-istered the +B diet and B-supplemented culture water (10 µg B/L) weresimilar to that of the ASTM BL-diet-reared frogs. However, the rate of tailresorption in larvae administered the –B diet was statistically slower thanthat observed in larvae administered either the ASTM BL or +B diets. The
130 Fort et al.
Biological Trace Element Research Vol. 90, 2002
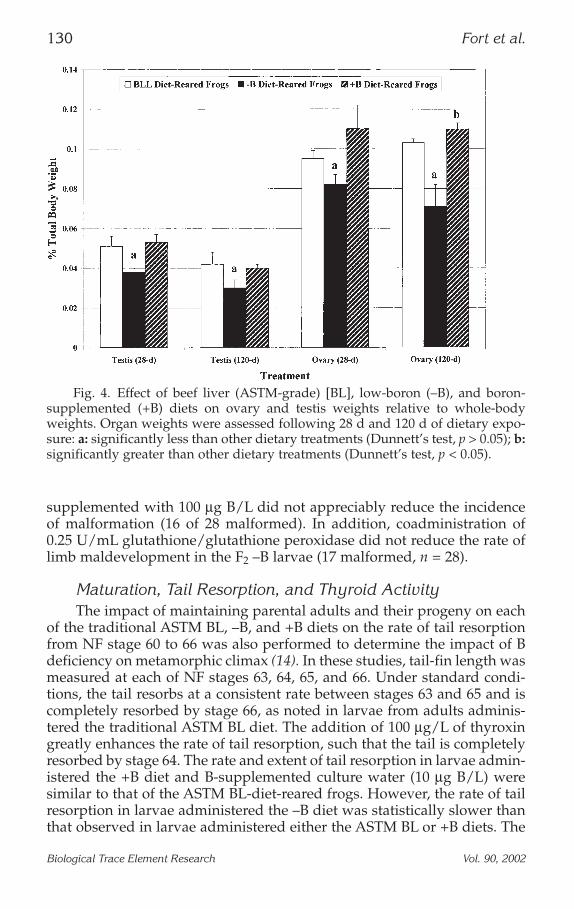
Fig. 4. Effect of beef liver (ASTM-grade) [BL], low-boron (–B), and boron-supplemented (+B) diets on ovary and testis weights relative to whole-bodyweights. Organ weights were assessed following 28 d and 120 d of dietary expo-sure: a: significantly less than other dietary treatments (Dunnett’s test, p > 0.05); b:significantly greater than other dietary treatments (Dunnett’s test, p < 0.05).

addition of 100 µg/L thyroxin and, to a lesser extent, 0.1% (w/v) iodinereversed these inhibitory effects found in the –B larvae. More interesting,however, was the impact of 100 µg B/L added to the –B cultured larvae.Addition of B at this level also reversed tail resorption inhibition associ-ated with administering the –B diet to a degree that was similar to thatobserved with the addition of iodine.
Results from tail resorption studies (14) designed to evaluate theimpact of low-B administration on maturation in Xenopus indicated theinduction of a significant slowing of the rate of tail resorption, an adversedevelopmental effect. This effect was not observed in either the +B or BLdietary treatments. Because maturation, including tail resorption, is con-trolled by the thyroid axis, these studies suggest that low-B administrationmay be interfering with the thyroid axis. Reversal of the inhibitory effectsof –B administration on tail resorption by both thyroxin and iodine furthersuggests that the effects observed were the result of disruption of the thy-roid axis. Initial measurements of triiodothreonine (T3), thyroid hormonein Xenopus, indicated that T3 levels are at least 2.5-fold lower in –B larvaethan detected in either the ASTM BL- or +B-reared larvae. The reversal oftail-fin resorption inhibition by the addition of 100 µg B/L to the culturemedia indicates that B is likely playing a role in the thyroid axis and maybe important in the synthesis of T3. These studies represent the first indi-cation of a biochemical or physiological role of B in the vertebrate bodywithout the addition of a physiological stressor.
Boron Deficiency and Reproduction
Female Assessment
Ovary Weight and Health. The effect of B depletion on ovary weight isprovided in Fig. 4 (16). Nonspecific, whole-body weight loss and theoccurrence of dietary stress-related disease were minimal across eachdietary treatment throughout the study. Results after 120 d indicated thatovary weight from the –B females was reduced significantly (Dunnett’stest, p < 0.05) when compared to ovary weights from either the +B- orASTM BL-diet-administered females. Results following 28 d of low-B-dietadministration are provided for comparison to the results described for the120-d B-depletion period. At 120 d, no significant increase in ovary sizewas noted in the +B-diet-administered females when compared to ovariesfrom the BL-diet-administered frogs. A marked increase in the size of theoviducts from the +B diet-administered females was noted when com-pared to oviducts isolated from females given either the –B or ASTM BLdietary regimen (Dunnett’s test, p < 0.05). A similar trend was noted withthe larger oviducts obtained from the +B-diet-administered females whencompared to the ASTM BL dietary group.
Oocyte Stage Distribution. The effect of B deprivation on oocyte-stagedistribution is presented in Fig. 5 (16). Results of oocyte-stage distribution
Review of Boron Essentiality in Xenopus 131
Biological Trace Element Research Vol. 90, 2002
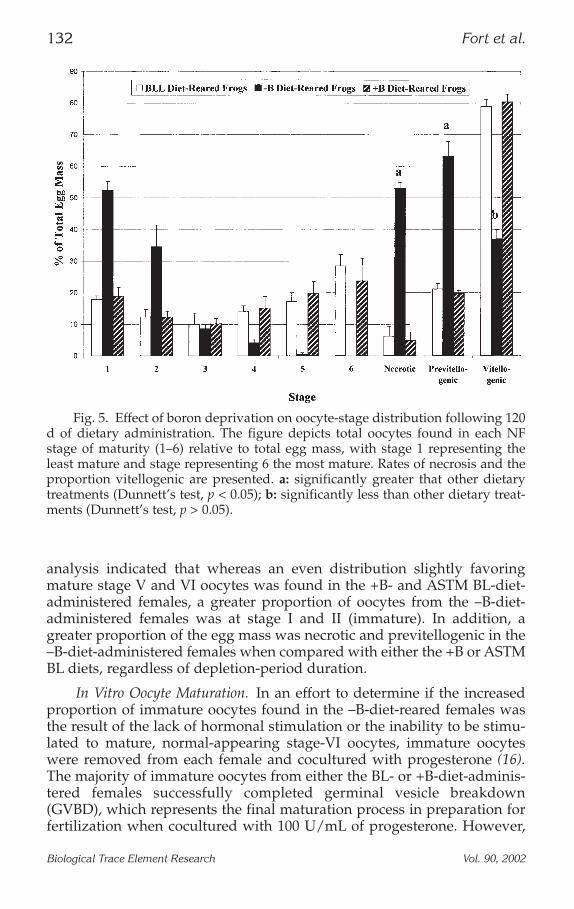
analysis indicated that whereas an even distribution slightly favoringmature stage V and VI oocytes was found in the +B- and ASTM BL-diet-administered females, a greater proportion of oocytes from the –B-diet-administered females was at stage I and II (immature). In addition, agreater proportion of the egg mass was necrotic and previtellogenic in the–B-diet-administered females when compared with either the +B or ASTMBL diets, regardless of depletion-period duration.
In Vitro Oocyte Maturation. In an effort to determine if the increasedproportion of immature oocytes found in the –B-diet-reared females wasthe result of the lack of hormonal stimulation or the inability to be stimu-lated to mature, normal-appearing stage-VI oocytes, immature oocyteswere removed from each female and cocultured with progesterone (16).The majority of immature oocytes from either the BL- or +B-diet-adminis-tered females successfully completed germinal vesicle breakdown(GVBD), which represents the final maturation process in preparation forfertilization when cocultured with 100 U/mL of progesterone. However,
132 Fort et al.
Biological Trace Element Research Vol. 90, 2002
Fig. 5. Effect of boron deprivation on oocyte-stage distribution following 120d of dietary administration. The figure depicts total oocytes found in each NFstage of maturity (1–6) relative to total egg mass, with stage 1 representing theleast mature and stage representing 6 the most mature. Rates of necrosis and theproportion vitellogenic are presented. a: significantly greater that other dietarytreatments (Dunnett’s test, p < 0.05); b: significantly less than other dietary treat-ments (Dunnett’s test, p > 0.05).
the majority of the oocytes from the –B dietary group did not under goGVBD, indicating that they were not progesterone responsive. The resultswere consistent regardless of the B-depletion period, although moremarked effects were observed following 120 d of B depletion.
Male AssessmentThe effect of B deprivation on testis weight is presented in Fig. 4.
Although not as dramatic as observed with the ovary, a significant reduc-tion in testis weight was observed as the result of administration of the –Bdiet. Unlike the observed beneficial effect of B on ovary weights in females,on statistically significant (Dunnett’s test, p < 0.05) increase in testis weightwas found in the +B-diet-administered males when compared to the BL-diet-administered adults.
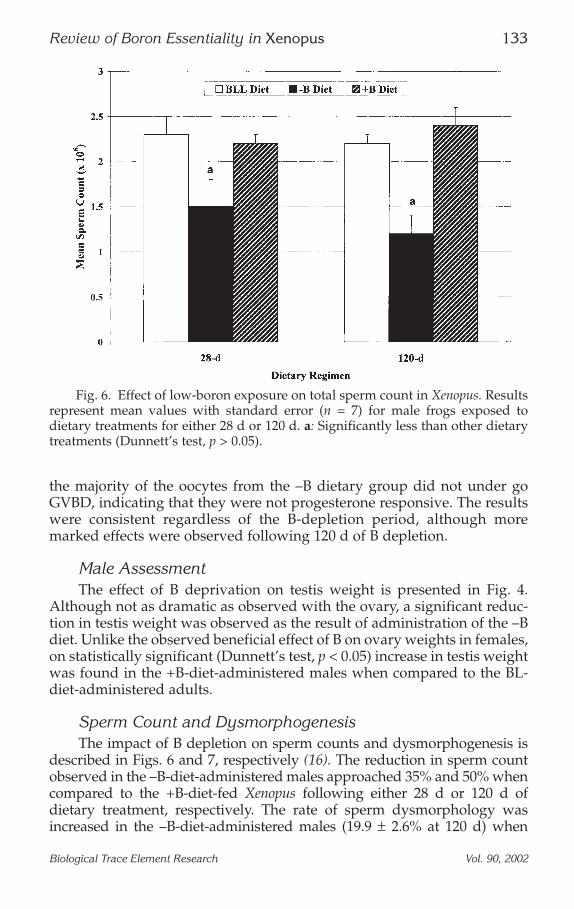
Sperm Count and DysmorphogenesisThe impact of B depletion on sperm counts and dysmorphogenesis is
described in Figs. 6 and 7, respectively (16). The reduction in sperm countobserved in the –B-diet-administered males approached 35% and 50% whencompared to the +B-diet-fed Xenopus following either 28 d or 120 d ofdietary treatment, respectively. The rate of sperm dysmorphology wasincreased in the –B-diet-administered males (19.9 ± 2.6% at 120 d) when
Review of Boron Essentiality in Xenopus 133
Biological Trace Element Research Vol. 90, 2002
Fig. 6. Effect of low-boron exposure on total sperm count in Xenopus. Resultsrepresent mean values with standard error (n = 7) for male frogs exposed todietary treatments for either 28 d or 120 d. a: Significantly less than other dietarytreatments (Dunnett’s test, p > 0.05).
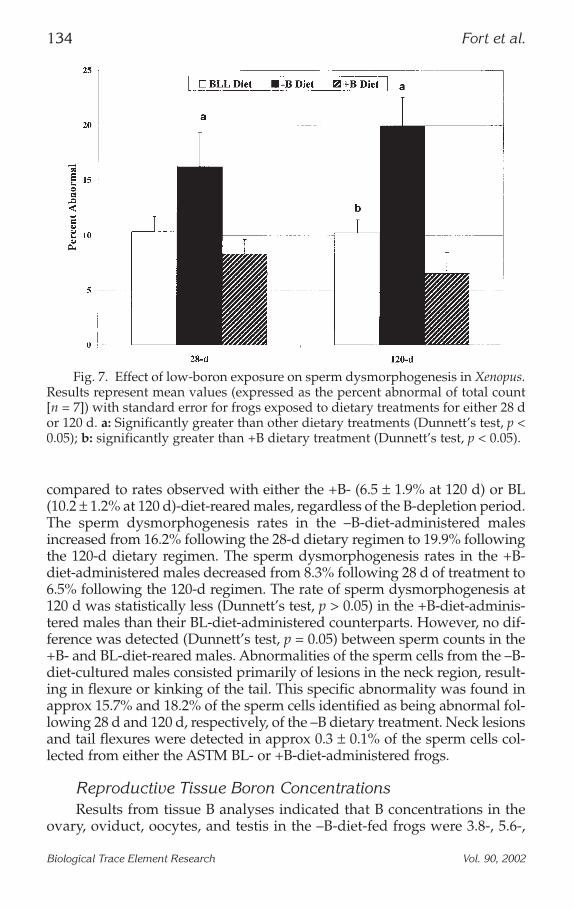
compared to rates observed with either the +B- (6.5 ± 1.9% at 120 d) or BL(10.2 ± 1.2% at 120 d)-diet-reared males, regardless of the B-depletion period.The sperm dysmorphogenesis rates in the –B-diet-administered malesincreased from 16.2% following the 28-d dietary regimen to 19.9% followingthe 120-d dietary regimen. The sperm dysmorphogenesis rates in the +B-diet-administered males decreased from 8.3% following 28 d of treatment to6.5% following the 120-d regimen. The rate of sperm dysmorphogenesis at120 d was statistically less (Dunnett’s test, p > 0.05) in the +B-diet-adminis-tered males than their BL-diet-administered counterparts. However, no dif-ference was detected (Dunnett’s test, p = 0.05) between sperm counts in the+B- and BL-diet-reared males. Abnormalities of the sperm cells from the –B-diet-cultured males consisted primarily of lesions in the neck region, result-ing in flexure or kinking of the tail. This specific abnormality was found inapprox 15.7% and 18.2% of the sperm cells identified as being abnormal fol-lowing 28 d and 120 d, respectively, of the –B dietary treatment. Neck lesionsand tail flexures were detected in approx 0.3 ± 0.1% of the sperm cells col-lected from either the ASTM BL- or +B-diet-administered frogs.
Reproductive Tissue Boron ConcentrationsResults from tissue B analyses indicated that B concentrations in the
ovary, oviduct, oocytes, and testis in the –B-diet-fed frogs were 3.8-, 5.6-,
134 Fort et al.
Biological Trace Element Research Vol. 90, 2002
Fig. 7. Effect of low-boron exposure on sperm dysmorphogenesis in Xenopus.Results represent mean values (expressed as the percent abnormal of total count[n = 7]) with standard error for frogs exposed to dietary treatments for either 28 dor 120 d. a: Significantly greater than other dietary treatments (Dunnett’s test, p <0.05); b: significantly greater than +B dietary treatment (Dunnett’s test, p < 0.05).

5.8-, and 7.0-fold less, respectively than levels detected in tissue samplescollected from the +B-diet-administered Xenopus (Table 2) (16). Theseresults indicated that the –B diet was capable of significantly reducing(Dunnett’s test, p > 0.05) tissue B levels and that the BL- and +B-diet-rearedadults possessed greater levels of B in these tissues than the –B-raisedfrogs.
Results of stage-specific analyses of B levels in both immature andmature normal-appearing oocytes, as well as necrotic oocytes, are summa-rized in Table 3. These results indicated that oocytes from the –B-diet-fedfemales contained significantly less B (Dunnett’s test, p > 0.05) thanoocytes from either the ASTM BL- or +B-diet-fed Xenopus. In addition, Bconcentrations appeared to be somewhat higher as the oocyte matured ineach of the three dietary regimens, but most appreciably in the +B dietarytreatment.
POTENTIAL BIOCHEMICAL ROLES OF BORON IN XENOPUS
The Oocyte Plasma Membrane Progesterone Receptor
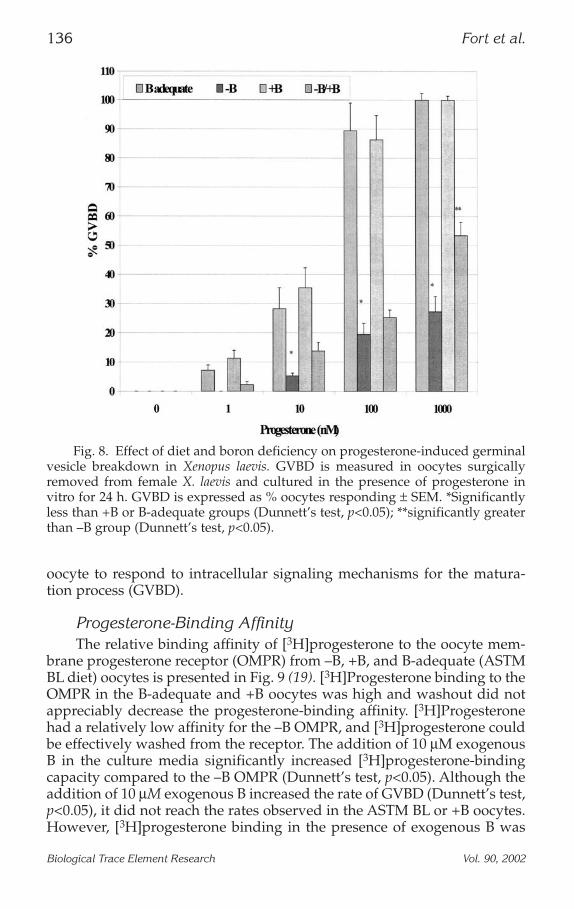
Progesterone-Induced Germinal Vesicle Breakdown Versus DietThe effect of increasing progesterone levels on GVBD in oocytes from
each dietary treatment is described in Fig. 8 (16,19). The rate of GVBDincreased with increasing progesterone concentration in oocytes fromfemales fed the ASTM BL diet and the +B diet. –B oocytes supplementedwith 10 µM of exogenous B also responded to increasing progesterone lev-els with an increase in the incidence of GVBD, however not to the extentobserved in the ASTM BL or +B oocytes. However, only a slight increase inthe incidence of GVBD was noted with increasing progesterone levels inoocytes from females fed the –B diet. At each progesterone concentration,GVBD was substantially less in the –B oocytes than in the B-adequate or+B oocytes. The –B oocytes supplemented with 10 µM of exogenous Bshowed a significantly greater response to progesterone at the 1000 nMprogesterone concentration than the –B oocytes.
Purified Oocyte Cytoplasmic Fraction MicroinjectionMicroinjection of purified oocyte cytoplasmic fraction (OCF) success-
fully induced GVBD in oocytes regardless of dietary treatment, indicatingthat the lack of GVBD in B-deficient oocytes could be reversed by cyto-plasmic factors from B-adequate oocytes (19). OCF microinjection (10 ngprotein/mL) induced GVBD in 99.3 ±, 0.8% of B-adequate oocytes in vitro,100.0% of the +B oocytes, and 97.6 ± 1.6% of the –B oocytes. These resultssuggested that B deficiency does not compromise the capacity of the
Review of Boron Essentiality in Xenopus 135
Biological Trace Element Research Vol. 90, 2002
oocyte to respond to intracellular signaling mechanisms for the matura-tion process (GVBD).
Progesterone-Binding AffinityThe relative binding affinity of [3H]progesterone to the oocyte mem-
brane progesterone receptor (OMPR) from –B, +B, and B-adequate (ASTMBL diet) oocytes is presented in Fig. 9 (19). [3H]Progesterone binding to theOMPR in the B-adequate and +B oocytes was high and washout did notappreciably decrease the progesterone-binding affinity. [3H]Progesteronehad a relatively low affinity for the –B OMPR, and [3H]progesterone couldbe effectively washed from the receptor. The addition of 10 µM exogenousB in the culture media significantly increased [3H]progesterone-bindingcapacity compared to the –B OMPR (Dunnett’s test, p<0.05). Although theaddition of 10 µM exogenous B increased the rate of GVBD (Dunnett’s test,p<0.05), it did not reach the rates observed in the ASTM BL or +B oocytes.However, [3H]progesterone binding in the presence of exogenous B was
136 Fort et al.
Biological Trace Element Research Vol. 90, 2002
Fig. 8. Effect of diet and boron deficiency on progesterone-induced germinalvesicle breakdown in Xenopus laevis. GVBD is measured in oocytes surgicallyremoved from female X. laevis and cultured in the presence of progesterone invitro for 24 h. GVBD is expressed as % oocytes responding ± SEM. *Significantlyless than +B or B-adequate groups (Dunnett’s test, p<0.05); **significantly greaterthan –B group (Dunnett’s test, p<0.05).
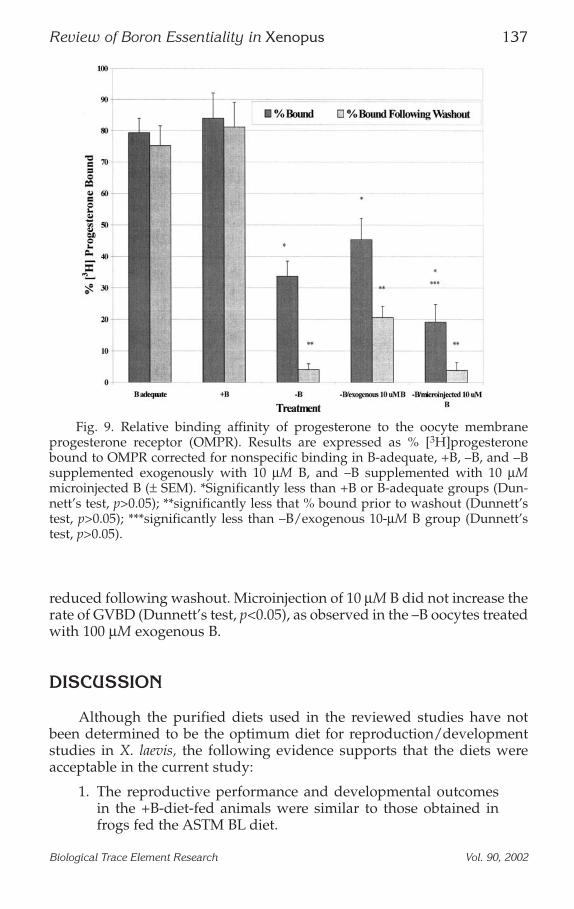
reduced following washout. Microinjection of 10 µM B did not increase therate of GVBD (Dunnett’s test, p<0.05), as observed in the –B oocytes treatedwith 100 µM exogenous B.
DISCUSSION
Although the purified diets used in the reviewed studies have notbeen determined to be the optimum diet for reproduction/developmentstudies in X. laevis, the following evidence supports that the diets wereacceptable in the current study:
1. The reproductive performance and developmental outcomesin the +B-diet-fed animals were similar to those obtained infrogs fed the ASTM BL diet.
Review of Boron Essentiality in Xenopus 137
Biological Trace Element Research Vol. 90, 2002
Fig. 9. Relative binding affinity of progesterone to the oocyte membraneprogesterone receptor (OMPR). Results are expressed as % [3H]progesteronebound to OMPR corrected for nonspecific binding in B-adequate, +B, –B, and –Bsupplemented exogenously with 10 µM B, and –B supplemented with 10 µMmicroinjected B (± SEM). *Significantly less than +B or B-adequate groups (Dun-nett’s test, p>0.05); **significantly less that % bound prior to washout (Dunnett’stest, p>0.05); ***significantly less than –B/exogenous 10-µM B group (Dunnett’stest, p>0.05).

2. The only difference in the composition of the +B and –B dietswas the B level.
3. The +B- and ASTM BL-diet-fed frogs were markedly more suc-cessful in terms of reproduction and development.
Although the dietary composition of the purified diets is differentfrom the ASTM BL diet, we know of no problems associated with the puri-fied diets in X. laevis. A similar purified diet was used successfully in thestudy of Cu deficiency in frogs (18). To specifically determine the optimumdiet for X. laevis reproduction and development, further studies will berequired in which the macronutrient (protein and lipid) sources and con-centrations are varied under a controlled feeding scenario. The micronu-trient requirements for X. laevis are somewhat better understood (2), butwill also require additional investigation. Low-B studies revealed thatfrogs fed the low-B diet in a low-B culture media produced a substantiallyhigher number of necrotic eggs and fertilized embryos that were substan-tially less viable than embryos from adult frogs fed a B-supplementedpurified diet or a meat diet. An increased frequency of abnormal gastrula-tion and markedly decreased embryo cell counts at NF stage 7.5 were alsonoted in embryos from adult frogs fed the –B diet. By 96 h of development,at which time all of the major stages of organogenesis have been com-pleted, none of the larvae collected from the –B adults and maintained inlow-B culture media developed normally. In contrast, 76% and 82% of thelarvae collected from adults fed the +B and ASTM BL diets, respectively,and maintained in 10-µg B/L culture media developed normally.
Importantly, the developmental abnormalities noted in the –Bgroup were distinct from those previously reported for copper or zinc(14,18). In this study, B deficiency induced maldevelopment of the cran-iofacial region, mouth, pigmented retina, and abnormal myotomeassembly. Addition of B (10 µg B/L) to the low-B culture water did notdecrease the adverse effects noted in embryos collected from adult frogsfed the –B diet.
Further study resulted in the development of a characteristic “U-shaped” concentration–response curve. Interestingly, the curve, whencompared to curves developed for both copper and zinc, is shifted to theright, indicating that adverse effects from B deficiency occur at higherlevels than for copper or for zinc. This also suggests that on an equimo-lar basis, B is less developmentally toxic to Xenopus than either copper orzinc. In addition, the margin of safety in which the trace nutrient is at alevel adequate for normal development (organogenesis) is appreciablygreater for B than for copper or zinc. Overall, these curves define thenutritionally essential and developmentally toxic concentration ranges ofB in Xenopus, and it appears that B is nutritionally important in lowervertebrate species. Similar concentration–response curves have beendeveloped recently for B in fish (8). Overall, these results are consistentwith findings in other species, including trout (9), zebrafish (8), and the
138 Fort et al.
Biological Trace Element Research Vol. 90, 2002

preimplantation mouse embryo (20), concerning the essential role of B. Inaddition, these studies demonstrate the usefulness of the X. laevis modelin nutrition studies.
Reproductive effects associated with B deficiency in the femaleincluded ovary atrophy, oocyte necrosis, and incomplete oocyte matura-tion. Of these findings, the latter was the most intriguing. Incomplete mat-uration of “normal-appearing” –B oocytes was marked by size andincomplete breakdown of the germinal vesicle. Normal GVBD wasobserved in oocytes from both the ASTM BL- and +B-diet-maintainedfemales. However, recent findings suggest that B deficiency resulted inincomplete oocyte maturation and that maturation could not be inducedby the administration of exogenous progesterone. Progesterone success-fully induced GVBD in +B and B-adequate X. laevis oocytes. B-Deficient X.laevis oocytes were capable of undergoing GVBD when endogenouslystimulated by microinjected purified OCF. These results indicated that theinability of the B-deficient oocytes was not associated with the cytoplasmicinduction process specifically, but rather in the progesterone receptor orsignal transduction pathways. The radio-binding studies found that prog-esterone binding to the B-deficient OMPR was greatly reduced comparedto OMPR from B-adequate or +B oocytes. Moreover, washout studiesdetermined that [3H]progesterone binding to the OMPR in B-deficientoocytes was more transient than the OMPR of B-adequate or +B oocytes.We hypothesize that B deficiency disrupts the integrity of the membrane-bound progesterone receptor. The loss of receptor stability may havereduced the effectiveness and efficiency of progesterone binding and sig-nal transduction, which leads ultimately to GVBD. Addition of exogenousB in the culture media, but not microinjected B, resulted in progesteronebinding by increased GVBD, suggesting that B may play an important rolein the structural integrity and physiological functioning of the OMPR.Thus, the extracellular portion of the transmembrane receptor may requireB to maintain physiological homology. The inability of intracellular B sup-plementation to increase GVBD suggests that B may be scavenged and notavailable to the OMPR directly. Further studies will be required to fullyelucidate how B is utilized in the Xenopus OMPR.
The OMPR in Xenopus has been characterized by Blondeau andBaulieu (21) as a 30-kDa peptide. These investigators suggested that twobinding sites were present, a higher-affinity site labeled A, which has a Kdof 1.2 µM progesterone, and a lower affinity binding site labeled B, whichhad an apparent affinity for progesterone 10-fold less than the A site. Fur-ther kinetic data suggested that the A sites have significance relateddirectly to biological activity, as progesterone binding to the A site corre-lated highly with the induction of GVBD. The specific relationshipbetween the A and B sites have not been elucidated yet. The 30-kDa pep-tide is appreciably smaller than intracellular steroid receptors, includingthe Xenopus oocyte estrogen receptor. It is possible that the intact receptoris a multiunit structure and that the 30-kDa peptide is a subunit of the
Review of Boron Essentiality in Xenopus 139
Biological Trace Element Research Vol. 90, 2002

whole receptor complex. Results from the present study suggested that Bdeficiency reduced the ability of progesterone to bind to the active site onthe OMPR. We suggest that B may play a role in maintaining the integrityof the A site of the OMPR. The role of B in the vertebrate cell membrane isfurther supported by results in zebra fish in which B deficiency results inmembrane blebbing in early B-deficient embryos (8). These results weresimilar to the exogastrulation observed in B-deficient Xenopus embryos(12,13). Dohs and Eckhert (22) also found B-deficient zebra fish to be pho-tophobic because of dystrophic cones in the pigmented retina. Poorlydeveloped outer membranes characterized this cone dystrophy with smallmyoid regions compared to B-supplemented or B-adequate specimen.
In males, a statistically significant (p > 0.05) decrease in testis weightand sperm count was noted in the –B males when compared to either theBL- or +B-diet-maintained adults. Decreased sperm counts in the –B-diet-fed males appeared to be simply the result of a general decrease in testishealth. An increase in sperm dysmorphology was also noted in the –Bmales. The primary site of dysmorphogenesis was identified in the base ofthe neck region of the sperm and characterized by a kinking of the tail.Aside from the flexure, sperm morphology appeared normal. Motility ofthe abnormal sperm was somewhat affected. Because the rate of fertiliza-tion in –B-diet-administered Xenopus recorded in previous studies(12,14,16) has only been somewhat reduced, it appears that the biologicalsignificance of these effects is not as substantial as observed in the femalesadministered the –B diet. However, these results indicate that the impactof B deficiency on male reproductive performance in higher vertebratesshould not be overlooked. It is also interesting that a B-enriched diet mightdecrease the rate of abnormal sperm cells in Xenopus and increase testisweight slightly. Similar to these enhancement effects in the male, the B-enriched diet in the female appeared to increase oviduct size and ovaryweight.
Results of B tissue analysis indicated that the –B diet reduced ovary,oviduct, oocyte, and testis B levels and that the +B-diet tissues containedthe greatest levels of B. These results were not necessarily surprising,because the +B diet contained the greatest quantity of B. The length of Bdepletion or supplementation dictated the magnitude of the tissue Bchanges, with longer depletion (120 d) reducing B levels more appreciablythan the shorter depletion period (28 d). Conversely, the longer supple-mentation period (120 d) increased tissue B concentrations when com-pared to the shorter dietary administration period. B levels in maturingoocytes generally increased from the immature (stages 1 and 2) to themature (stages 5 and 6). Oocytes from the +B- or BL-diet-administeredfemales, whether normal appearing or necrotic, contained appreciablygreater levels of B than oocytes from the –B-diet-fed group. The greaterlevels of B found in the tissue from the +B-diet-administered frogs mayaccount for the slight beneficial effects of B supplementation found onsperm dysmorphogenesis rates and ovary size.
140 Fort et al.
Biological Trace Element Research Vol. 90, 2002

Overall, several of the processes affected by B deficiency in X. laevisinvolve developmental induction processes in which an external signal isresponsible for initiating a cascade of responses leading to cellular differ-entiation and, in some cases, morphogenesis. We have found (16,19) B defi-ciency to cause impaired oocyte maturation (GVBD), vitellogenesis,abnormal development of the notochord and eye (12–14), multigenera-tional limb defects (15), and interference with metamorphic events (resorp-tion of the tail and skin maturation) (12,14).
CONCLUSIONS
Each of the studies conducted to date suggest that the adverse effectsfound in X. laevis resulting from B deficiency could be found during game-togenesis, gamete maturation, embryonic development, and larval matu-ration (12–15). These studies confirmed that B deficiency was capable ofinterrupting the life cycle of X. laevis. Additional studies evaluating therole of B in the thyroid axis and the oocyte plasma membrane progesteronereceptor provide the first line of direct evidence for a biochemical role of Bin X. laevis. Combined together, this research program provides clear evi-dence that B is nutritionally essential in X. laevis.
ACKNOWLEDGMENTS
The authors wish to thank Deanne Fort for her assistance in the prepa-ration of this manuscript. The authors would also like to acknowledge Dr.Philip Strong and U.S. Borax for their support of this research program.
REFERENCES
1. P. A. Goyer, Selenium, in Casarett and Doull’s Toxicology: The Basic Science of Poisons, 5thed. C. D. Klaassen, ed., McGraw-Hill, New York, pp. 718–720 (1996).
2. B. D. Culver, P. L. Strong, and F. J. Murray, Boron, in Patty’s Toxicology, Vol. 3, (E. Bing-ham, B. Cohrssen, and C. H. Powell, eds.) Wiley, New York, pp. 519–582 (2001).
3. US EPA, Toxicological Review of Boron and Compounds US Environmental ProtectionAgency, Washington, DC (2001).
4. S. A. Hubbard, Comparative toxicology of borates, Biol. Trace Element Res. 66, 343–358(1998).
5. ECOTOC, Ecotoxicology of Some Inorganic Borates, European Centre for Ecotoxicologyand Toxicology of Chemicals, Brussels, p. 99 (1997).
6. R. Eisler, Boron, in Handbook of Chemical Risk Assessment, Vol. 3: Metalloids, Radiation,Cumulative Index to Chemicals and Species, R. Eisler, ed., Lewis, Boca Raton, FL, pp.1567–1612 (2001).
7. B. Dell, P. H. Brown, and R. W. Bell, Boron in Soils and Plants: Reviews, Kluwer Acade-mic, Boston, p. 218 (1997).
Review of Boron Essentiality in Xenopus 141
Biological Trace Element Research Vol. 90, 2002

8. R. I. Rowe, C. Bouzan, S. Nabili, and C. D. Eckhert, The response of trout and zebrafishembryos to low and high born concentrations is u-shaped, Biol. Trace Element Res. 66,261–270 (1998).
9. C. D. Eckhert, Boron stimulates embryonic trout growth, J. Nutr 128, 2488–2493 (1998).10. C. D. Hunt, Biochemical effects of physiological amounts of dietary boron, J. Trace Ele-
ments Exp. Med. 9, 185–213 (1996).11. American Society for Testing and Materials. Standard Guide for Conducting the Frog
Embryo Teratogenesis Assay—Xenopus (FETAX). ASTM, Philadelphia (1998).12. D. J. Fort, T. L. Propst, E. L. Stover, P. L. Strong, and F. J. Murray, Adverse effects of
insufficient boron in Xenopus, Biol. Trace Element Res. 66, 237–259 (1998).13. D. J. Fort, E. L. Stover, P. L. Strong, F. J. Murray, and C. L. Keen, Chronic feeding of a
low boron diet adversely effects reproduction and development in Xenopus laevis. J.Nutr. 129, 2055–2060 (1999).
14. D. J. Fort, T. L. Propst, E. L. Stover, F. J. Murray, and P. L. Strong, Adverse effects fromlow dietary and environmental boron exposure on reproduction, development, andmaturation in Xenopus laevis. J. Trace Elements Exp. Med. 12, 175–186 (1999).
15. D. J. Fort, E. L. Stover, R. L. Rogers, H. F. Copley, L. A. Morgan, and E. R. Foster, Chronicboron or copper deficiency induces limb teratogenesis in Xenopus. Biol. Trace ElementRes. 77, 173–187 (2000).
16. D. J. Fort, E. L. Stover, P. L. Strong, and F. J. Murray, Effect of boron deprivation onreproductive parameters in Xenopus laevis. J. Trace Elements Exp. Med. 12:187–204, 1999.
17. P. D. Nieuwkoop and J. Faber. Normal Table of Xenopus laevis (Daudin), 2nd ed. NorthHolland, Amsterdam (1975).
18. D. J. Fort, E. L. Stover, C. M. Lee, and W. J. Adams, Adverse developmental and repro-ductive effects of copper deficiency in Xenopus laevis Biol. Trace Element Res. 77, 159–172(2000).
19. D. J. Fort, Boron deficiency disables Xenopus laevis maturation events, Biol. Trace ElementRes. 85, 157–169 (2002).
20. L. Lanoue, P. Strong, and C. L. Keen, Adverse effects of a low boron environment on thepreimplantation development of mouse embryos in vitro, J. Trace Elements Exp. Med. 12,235–250 (1999).
21. J. P. Blondeau and E. E. Baulieu, Progesterone receptor characterized by photoaffinitylabeling in the plasma membrane of Xenopus laevis oocytes, Biochem. J. 219, 785–792(1984).
22. S. Dohs and C. D. Eckhert, Photoreceptor dystrophy in boron depleted zebrafish, inTrace Elements in Man and Animals, Vol. 10, A. M. Roussel, R. A. Anderson, and A. E.Favrier, eds., Kluwer Academic/Plenum, New York, pp. 1082–1083 (1999).
142 Fort et al.
Biological Trace Element Research Vol. 90, 2002
Related Documents


![[ 149 ] the growth of the hindlimb bud of xenopus laevis and its ...](https://static.cupdf.com/doc/110x72/586789b31a28ab44568b868b/-149-the-growth-of-the-hindlimb-bud-of-xenopus-laevis-and-its-.jpg)








