ORIGINAL ARTICLE Immunostimulatory activity of potential probiotic yeast strains in the dorsal air pouch system and the gut mucosa A. Kourelis 1 , C. Kotzamanidis 1 , E. Litopoulou-Tzanetaki 2 , J. Papaconstantinou 3 , N. Tzanetakis 3 and M. Yiangou 1 1 Department of Genetics, Development & Molecular Biology, Biology School, Aristotle University of Thessaloniki, Thessaloniki, Greece 2 Laboratory of Food Microbiology and Hygiene, Faculty of Agriculture, Aristotle University of Thessaloniki, Thessaloniki, Greece 3 Department of Biochemistry & Molecular Biology, University of Texas Medical Branch, Galveston, TX, USA Introduction Immunity in the gut ecosystem is achieved by the interac- tion of cells that participate in the architecture of the gastrointestinal system with luminal microflora. Microor- ganisms from the environment and the normal commen- sal flora of the gastrointestinal tract have a marked impact on the maturation of the mucosal immune system (Hessle et al. 1999). They have been shown to crucially influence the immune response establishment, especially the Th1 proinflammatory and Th2 ⁄ Treg anti-inflammatory and regulatory balance (Wittig and Zeitz 2003). These responses are mediated in part by the recognition of microbe-associated molecular patterns (MAMPs) from a class of pattern recognition receptors (PRRs), the Toll-like receptors (TLRs), present on intestinal epithelial (IECs), dendritic and macrophage cells (Neish 2002). Probiotics are living microorganisms which upon ingestion in adequate amounts confer health benefits to the host (FAO ⁄ WHO 2002). Probiotics fulfil this defini- tion through a variety of disparate and overlapping mech- anisms such as regulation of the intestinal homeostasis, antimicrobial activity and modulation of the innate and ⁄ or adaptive immunity. The positive influence of probiotics on the phagocytic activity of monocytes and polymorphonuclear (PMN) cells as well as their effects on IgA, chemokine and cytokine production after in vitro and in vivo studies has been recently reviewed (Delcenser- ie et al. 2008). These strain-specific beneficial probiotic effects enhance the defence mechanisms and help the host to maintain immune homeostasis (Galdeano et al. 2007). The most extensively studied probiotics belong to the genera Lactobacillus and Bifidiobacterium (Borriello et al. 2003; Tuohy et al. 2003). Despite the occurrence of yeasts Keywords air pouch, cytokines, gut mucosa, probiotic, yeast. Correspondence Yiangou Minas, Department of Genetics, Development & Molecular Biology, Biology School, Aristotle University of Thessaloniki, 54 124, Thessaloniki, Greece. E-mail: [email protected] 2009 ⁄ 1422: received 11 August 2009, revised 9 November 2009 and accepted 6 December 2009 doi:10.1111/j.1365-2672.2009.04651.x Abstract Aims: To determine the immunostimulatory activity of 15 presumptive probiotic yeast strains in the dorsal air pouch system in comparison with their activity in the gut mucosa. Methods and Results: Presumptive probiotic yeast strains previously isolated from human gastrointestinal tract and Feta cheese were further characterized genotypically and biochemically. The Saccharomyces cerevisiae 982, Saccharo- myces boulardii KK1 and Kluyveromyces lactis 630 strains exhibited in the air pouch increased polymorphonuclear cell influx and phagocytic activity as well as cytokine production with similar potency as the probiotics Ultra levure S. boulardii and Lactobacillus acidophilus NCFB 1748. Oral administration of these strains in mice results in differential activation of small intestine immune responses concerning IgA and cytokine production as well as Toll-like receptor expression. Conclusion: Besides the Saccharomyces strains 982 and KK1, the K. lactis 630 strain could also be considered as a candidate probiotic. Significance and Impact of the Study: The air pouch model may be used as an alternative and rapid method for the discrimination and selection of potential probiotic yeast strains. Journal of Applied Microbiology ISSN 1364-5072 260 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 260–271 ª 2010 The Authors

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Immunostimulatory activity of potential probiotic yeaststrains in the dorsal air pouch system and the gut mucosaA. Kourelis1, C. Kotzamanidis1, E. Litopoulou-Tzanetaki2, J. Papaconstantinou3, N. Tzanetakis3
and M. Yiangou1
1 Department of Genetics, Development & Molecular Biology, Biology School, Aristotle University of Thessaloniki, Thessaloniki, Greece
2 Laboratory of Food Microbiology and Hygiene, Faculty of Agriculture, Aristotle University of Thessaloniki, Thessaloniki, Greece
3 Department of Biochemistry & Molecular Biology, University of Texas Medical Branch, Galveston, TX, USA
Introduction
Immunity in the gut ecosystem is achieved by the interac-
tion of cells that participate in the architecture of the
gastrointestinal system with luminal microflora. Microor-
ganisms from the environment and the normal commen-
sal flora of the gastrointestinal tract have a marked impact
on the maturation of the mucosal immune system (Hessle
et al. 1999). They have been shown to crucially influence
the immune response establishment, especially the Th1
proinflammatory and Th2 ⁄ Treg anti-inflammatory and
regulatory balance (Wittig and Zeitz 2003). These
responses are mediated in part by the recognition of
microbe-associated molecular patterns (MAMPs) from a
class of pattern recognition receptors (PRRs), the Toll-like
receptors (TLRs), present on intestinal epithelial (IECs),
dendritic and macrophage cells (Neish 2002).
Probiotics are living microorganisms which upon
ingestion in adequate amounts confer health benefits to
the host (FAO ⁄ WHO 2002). Probiotics fulfil this defini-
tion through a variety of disparate and overlapping mech-
anisms such as regulation of the intestinal homeostasis,
antimicrobial activity and modulation of the innate
and ⁄ or adaptive immunity. The positive influence of
probiotics on the phagocytic activity of monocytes and
polymorphonuclear (PMN) cells as well as their effects on
IgA, chemokine and cytokine production after in vitro
and in vivo studies has been recently reviewed (Delcenser-
ie et al. 2008). These strain-specific beneficial probiotic
effects enhance the defence mechanisms and help the host
to maintain immune homeostasis (Galdeano et al. 2007).
The most extensively studied probiotics belong to the
genera Lactobacillus and Bifidiobacterium (Borriello et al.
2003; Tuohy et al. 2003). Despite the occurrence of yeasts
Keywords
air pouch, cytokines, gut mucosa, probiotic,
yeast.
Correspondence
Yiangou Minas, Department of Genetics,
Development & Molecular Biology, Biology
School, Aristotle University of Thessaloniki, 54
124, Thessaloniki, Greece.
E-mail: [email protected]
2009 ⁄ 1422: received 11 August 2009, revised
9 November 2009 and accepted 6 December
2009
doi:10.1111/j.1365-2672.2009.04651.x
Abstract
Aims: To determine the immunostimulatory activity of 15 presumptive
probiotic yeast strains in the dorsal air pouch system in comparison with their
activity in the gut mucosa.
Methods and Results: Presumptive probiotic yeast strains previously isolated
from human gastrointestinal tract and Feta cheese were further characterized
genotypically and biochemically. The Saccharomyces cerevisiae 982, Saccharo-
myces boulardii KK1 and Kluyveromyces lactis 630 strains exhibited in the air
pouch increased polymorphonuclear cell influx and phagocytic activity as well
as cytokine production with similar potency as the probiotics Ultra levure
S. boulardii and Lactobacillus acidophilus NCFB 1748. Oral administration of
these strains in mice results in differential activation of small intestine immune
responses concerning IgA and cytokine production as well as Toll-like receptor
expression.
Conclusion: Besides the Saccharomyces strains 982 and KK1, the K. lactis 630
strain could also be considered as a candidate probiotic.
Significance and Impact of the Study: The air pouch model may be used as an
alternative and rapid method for the discrimination and selection of potential
probiotic yeast strains.
Journal of Applied Microbiology ISSN 1364-5072
260 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 260–271
ª 2010 The Authors

in many dairy-related products (Fleet 1990; Jakobsen and
Narvhus 1996) and the human gastrointestinal tract
(Psomas et al. 2001; Czerucka et al. 2007), their potential
as probiotics has been overlooked. So far, only yeasts of
the genus Saccharomyces are used as probiotics. Saccharo-
myces boulardii has been shown to possess immunomodu-
latory activity (Rodrigues et al. 2000; Czerucka et al. 2007)
and along with brewer’s yeast (Saccharomyces cerevisiae,
Saccharomyces carlsbergensis; Sargent and Wickens 2004)
are the only yeast strains commercialized as medicinal
probiotics in human medicine.
In previous studies, several yeasts were isolated from Feta
cheese and infants’ gastrointestinal tract (Tzanetakis et al.
1996; Andrighetto et al. 2000; Psomas et al. 2001), and
some of them were shown to possess in vitro probiotic
properties such as acid and bile tolerance and cholesterol
removal ability (Psomas et al. 2001, 2003). In vitro pro-
biotic assays could screen among a very high number of
microorganisms but cannot predict their behaviour in a
multifactorial in vivo system. In previous study, we have
demonstrated that the architecture of the epithelium
enclosed dorsal mouse or rat air pouch (Sedgwick et al.
1983) resembles gut mucosa concerning the immune
reactivity induced by potential probiotic Lactobacillus
strains (Kourelis et al. 2010a). Mice received Lactobacillus
strains in the air pouch or orally exhibited interaction
with the air pouch barrier and the gut mucosa barrier
and immune responses such as increased PMN chemo-
taxis and phagocytic activity as well as increased cytokine
production. We suggested that the air pouch system may
be used as an alternative and rapid in vivo method for
the initial discrimination and selection of potential pro-
biotic Lactobacillus strains (Kourelis et al. 2010a). These
results provide us with the biological system to examine
the in vivo immunomodulatory activity of potential
probiotic yeast strains.
On the basis of polyphasic taxonomy, it was of interest
to further characterize the above yeast strains based on
genotypic and biochemical methods and to determine
their potential immunomodulatory activity using the air
pouch model in comparison with the gut mucosa. We
present data that three yeast strains exhibiting immuno-
stimulatory activity in the air pouch also show regulation
of small intestine immune responses concerning the
production of IgA, cytokines and TLRs.
Materials and methods
Yeast and bacterial strains
The yeast strains (Table 1) were obtained from the collec-
tion of the Laboratory of Food Microbiology and Hygiene,
Aristotle University of Thessaloniki and were isolated from
Feta cheese and infants’ gastrointestinal tract (Tzanetakis
et al. 1996; Andrighetto et al. 2000; Psomas et al. 2001).
The strains 746, 832, 982, KK1, 570, 630, 414, KK2.1,
KK2.5, KK3.1, KK4.1, KK6.5, KK6P, KK5Y3 may be consid-
ered as potential probiotics because of their ability to with-
stand low pH values (pH 3Æ0) and high bile concentration,
reduce cholesterol levels and show adhesion capacity
(Psomas et al. 2001, 2003; Kourelis et al. 2010b). The
probiotic strains Ultra levure S. boulardii (UL; Biocodex,
Table 1 Biolog metabolic pattern identifica-
tion and pulsed-field gel electrophoresis
(PFGE) chromosome profile grouping of yeast
strains
Origin Strain Previous identification* Biolog
PFGE
grouping
Feta cheese 746 Saccharomyces cerevisiae Zygosaccharomyces cidri S. cerevisiae
832 No ID
840a Saccharomyces boulardii
952 S. cerevisiae
982 S. cerevisiae
Infants’ faeces KK1 S. boulardii S. boulardii S. boulardii
Ultra Levure UL
Feta cheese 570 Kluyveromyces lactis K. lactis K. lactis
630 Kluyveromyces marxianus
Feta cheese 414 Debaryomyces hansenii D. hansenii D. hansenii
Infants’ faeces KK2.1 Candida albicans C. albicans C. albicans
KK2.5
KK3.1
KK4.1
KK6.5 Candida parapsilosis C. parapsilosis C. parapsilosis
KK6P
KK5Y3 Isaatchenkia orientalis I. orientalis I. orientalis
No ID, no identification.
*(Andrighetto et al. 2000; Psomas et al. 2001; Tzanetakis et al. 1996).
A. Kourelis et al. Characterization of candidate probiotic yeasts
ª 2010 The Authors
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 260–271 261

Gentilly, France) and Lactobacillus acidophilus NCFB 1748
(National Collection of Food Bacteria, NCFB, Scotland)
were also included as positive control strains. The strain
S. cerevisiae 952 and the reference strains S. cerevisiae
NCYC167, Debaryomyces hansenii NCYC9 (National
Collection of Yeast Cultures: NCYC, Norwich, UK) and
Candida parapsilosis CBS1954 (Centraal Bureau voor
Schimmelcultures: CBS, Baarn, the Netherlands) were used
as negative controls because they exhibited no proven in vitro
or in vivo probiotic properties (Kourelis et al. 2010b).
Culture conditions and preparation of strains
Yeasts were grown at 30�C for 20–24 h in yeast extract
glucose peptone (YEGP) broth [2% (w ⁄ v) glucose, 0Æ5%
(w ⁄ v) yeast extract, 1% (w ⁄ v) peptone]. A YEGP agar
slope [2% (w ⁄ v) glucose, 0Æ5% (w ⁄ v) yeast extract, 1%
(w ⁄ v) peptone, 1Æ5% (w ⁄ v) agar] was used for the main-
tenance of the cultures at 4�C. L. acidophilus NCFB 1748
was maintained at )80�C in MRS (de Man, Rogosa,
Sharpe) broth with 25% (v ⁄ v) glycerol and was grown in
MRS broth at 37�C for 18 h. All strains were subcultured
at least three times prior to experimental use.
Yeast or NCFB 1748 cells were harvested from 20- to
24- and 18-h cultures, respectively by centrifugation and
washed twice with sterile saline. For administration into
the air pouch, yeast or NCFB 1748 bacterial cells were
resuspended in pyrogenic-free sterile saline at 5 · 107 and
5 · 108 CFU ml)1, respectively. For oral administration
in mice, yeasts were resuspended at 109 CFU ml)1.
Biolog YT microplate identification
Yeast strains were grown on Biolog Universal Yeast Agar
– BUY (Biolog Inc., Hayward, CA, USA) at 26�C for
48 h. Yeast identification was performed according to the
manufacturer’s instructions using the Biolog Microlog 3
computer software.
Pulsed-field gel electrophoresis (PFGE)
Chromosome plugs were prepared from 24-h yeast cul-
tures (109 cells per ml) according to the protocol of DNA
extraction as described previously (Steinkamp-Zucht and
Fahrig 1995) with an adaptation regarding the lyticase
enzyme concentration which was reduced in half for the
two Kluyveromyces strains.
PFGE karyotypes were performed in a Rotaphor R23
Tank (Biometra, Goettingen, Germany). Two different run-
ning conditions were used: duration 45 h, interval 50–20 s
linear, angle 130–115� log, field strength (124–114 V) log,
temperature 14�C in 1% (w ⁄ v) agarose for the S. cerevisiae
strains and duration 84 h, interval 350–50 s log, angle 110–
100� log, field strength 120–50 V linear, temperature 11�C,
in 0Æ7% (w ⁄ v) agarose for the other yeast strains. Agarose
gels were then stained with ethidium bromide while the
Saccharomyces cerevisiae YPH80 (New England Biolabs, Inc.,
Ipswich, MA) and Hansenula wingei (Bio-Rad Laboratories
Ltd, Herts, UK) strains served as molecular size standards.
The software GelCompar 4.0 (Applied Maths, Kortrijk,
Belgium) was used for normalization and processing of
the chromosome profiles and cluster analysis, based on
the Pearson coefficient and the unweighted pair group
algorithm with arithmetic averages (UPGMA).
Animals
Balb ⁄ c mice (25–30 g) and Fischer 344 rats (130–180 g)
aged 8–14 weeks were housed in stainless-steel cages with a
12-h light ⁄ dark cycle in a controlled atmosphere. The ani-
mals were fed ad libitum with a conventional balanced diet
and had free access to water during the experimental proto-
col. Animals were sacrificed by light ether anaesthesia fol-
lowed by cervical dislocation. All experiments were
performed in an accredited animal facility (number EL 54
BIO 02, School of Biology, Aristotle University of Thessalo-
niki). The protocol complied with the current ethical regu-
lations on animal research of our university and was in
accordance with both Greek National Legislation and to EC
Ethical Regulations. All groups included in the present
study consisted of five animals (unless otherwise indicated),
and each experiment was repeated at least two times.
Air pouch formation
Air pouches were raised on the dorsum of mice and rats
as we previously described (Kourelis et al. 2010a) by sub-
cutaneous injection of 3 and 20 ml sterile air, respectively.
Same size of air pouches was achieved by refilling the air
pouches with the appropriate volume of air day by day.
Six-day air pouches were then injected with 200 ll (mice)
and 1 ml (rats) of pyrogenic-free saline containing
5 · 107 CFU ml)1 of yeast or 5 · 108 CFU ml)1 of the
NCFB 1748 strain. Control animals received in the air
pouch only pyrogenic-free saline. The above doses were
chosen according to previous studies (Coates and McColl
2001; Kourelis et al. 2010a), and the fact that a beneficial
probiotic dosage is considered to be 107–109 CFU ml)1
(van Niel et al. 2002).
Determination of PMN accumulation in yeast or NCFB
1748-treated air pouches
The PMN cells accumulated in mouse or rat air pouches
in response to yeast or NCFB 1748 strain treatment were
harvested at the maximum of the induction 3 h post
Characterization of candidate probiotic yeasts A. Kourelis et al.
262 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 260–271
ª 2010 The Authors

treatment (Kourelis et al. 2010a). PMN cells were col-
lected by centrifugation at 300 g for 10 min after injec-
tion in the rat or mouse air pouch of 10 or 2 ml of saline
respectively, followed by air pouch lavage. After centrifu-
gation, the supernatant was collected, filtered through
0Æ22-lm filter and hence referred to as the ‘air pouch
exudate’. The cell pellet containing the air pouch exudate
PMN cells was washed twice in saline and counted using
a haemocytometer while cell viability was determined by
trypan blue exclusion.
Phagocytic activity
Phagocytic experiments were performed using rat air
pouch PMN cells following the protocol of Loose et al.
(1978) with some modifications (Kourelis et al. 2010a).
Briefly, 100 ll containing 107 CFU ml)1 of baker’s yeast
(inactivated after boiling for 1 h) were co-incubated
(opsonized) with 100 ll of rat serum at 37�C for 30 min.
Then, 0Æ5 ml containing 5 · 106 cells per ml of air pouch
PMNs and 0Æ3 ml Hanks’ balanced salt solution (HBSS)
were added and incubated for additional 30 min. Samples
(100 ll) were taken at the indicated time post intervals of
5, 15 and 30 min. After stopping further phagocytosis by
adding 50 ll foetal bovine serum (FBS, Thermo Hyclone
Scientific, MA, USA), 50 ll trypan blue was also added to
visualize nonphagocytosed baker’s yeast cells. The number
of air pouch PMN cells containing baker’s yeast cells was
counted using a haemocytometer.
Detection of cytokines in air pouch exudates
Air pouch exudates were isolated from control or yeast-
treated air pouches and used for the determination of
tumour necrosis factor alpha (TNF-a), gamma interferon
(IFN-c) and interleukin 10 (IL-10). Determination of
cytokine concentration in mouse air pouch exudates was
performed using ELISA kits (eBioscience, Inc., San Diego,
CA, USA) according to the manufacturer’s instructions.
Purified cytokines included in the kit as well as air pouch
exudates isolated from bacterial lipopolysaccharide (LPS)
or Freund’s complete adjuvant (FCA) treated air pouches
were included as positive samples for the assay.
Histological samples
The yeast suspension (108 CFU per mouse) was orally
administered for 10 consecutive days to mice housed in
separate cages. The doses for oral administration used in
this study were chosen on the basis of the usual con-
sumption levels of 107–109 cells per ml for a probiotic to
have a beneficial effect (van Niel et al. 2002). The control
group received only pyrogenic-free sterile saline for the
same period. Small intestines were then removed, fixed in
4% parafolmadehyde in phosphate buffered saline (PBS)
and then embedded in paraffin blocks (Sainte-Marie
1962). Sections of 4 lm thickness were either stained with
haematoxylin–eosin or further processed for immuno-
histochemistry.
Immunohistochemical detection of IgA, cytokine and
TLR producing cells in the small intestine
The number of IgA and TLR2 producing cells was deter-
mined on histological sections by a direct immunofluo-
rescence assay. After deparaffinization and rehydration in
a graded series of ethanol, paraffin sections were incu-
bated with anti-IgA monospecific (Sigma, St Louis, MO,
USA) or TLR2 (Santa Cruz Biotechnology, Santa Cruz,
CA, USA) antibodies conjugated with fluorescein isothio-
cyanate (FITC) for 45 min and 2 h, respectively.
Cytokine, cyclooxygenase (COX) and TLR producing
cells were determined by an indirect immunofluorescence
assay. After deparaffinization and rehydration in a graded
ethanol series, paraffin sections were blocked with a 5%
(v ⁄ v) FBS in PBS solution for 30 min at room tempera-
ture (RT). The sections were then washed three times in
PBS and incubated for 2 h at RT with specific antibodies
directed against mouse TLR4, TLR9 (Abcam plc, Cam-
bridge, UK), TLR6, COX-1, COX-2, IL-5, IL-6, IL-12B
(Santa Cruz Biotechnology), IL-10, IFN-c and TNF-a(R&D systems Inc., Minneapolis, MN, USA). After four
washes in PBS, the sections were incubated for 1 h with
anti-goat or anti-rabbit (for IL-5) FITC-conjugated anti-
bodies (Sigma). The number of positive cells was counted
in 10 fields at 400· magnification. To confirm the speci-
ficity of the immunodetection, the samples were incu-
bated only with the secondary antibody, and the number
of positive cells was excluded from the number of cells
estimated for testing samples. For the final scoring, two
investigators achieved consensus.
Statistical analysis
Results are expressed as mean ± SEM. Multiple compari-
sons were performed by one-way anova followed by
Tukey’s test, and statistical significance was accepted at
values of P < 0Æ05.
Results
Biolog YT microplate identification and chromosome
length polymorphism (CLP)
Many of the probiotic properties are strain specific, and
thus reliable identification at strain level is required.
A. Kourelis et al. Characterization of candidate probiotic yeasts
ª 2010 The Authors
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 260–271 263

The yeast strains isolated from Feta cheese and infants’
gastrointestinal tract were identified by biochemical
(Application Programming Interface: API-ID32) and
molecular (randomly amplified polymorphic DNA PCR
and mitochondrial DNA restriction analysis) methods
(Tzanetakis et al. 1996; Andrighetto et al. 2000; Psomas
et al. 2001). On the basis of polyphasic taxonomy, these
strains were further characterized by Biolog YT microplate
identification and PFGE analysis. The data presented in
Table 1 show that Biolog YT method identified the strain
630, previously characterized as Kluyveromyces marxianus,
as Kluyveromyces lactis, while the strains 746 and 840apreviously characterized as S. cerevisiae were identified as
Zygosaccharomyces cidri and S. boulardii, respectively. The
strain 832 could not be identified (Table 1).
PFGE is considered as the gold standard method to
discriminate probiotics at strain level (FAO ⁄ WHO 2002).
PFGE analysis revealed polymorphism in the size and
number of chromosomes of the examined strains (Fig. 1).
The 746, 832, 840a, 952, 982 and KK1 strains had
chromosome band distribution (Fig. 1a) typical of yeast
species within the Saccharomyces sensu stricto complex
(Jespersen et al. 2000). The non-Saccharomyces strains
(Fig. 1b) showed a divergent chromosome profile accord-
ing to their species apart from the D. hansenii strains
(NCYC 9 and 414), which were clustered separately.
PFGE analysis of Isaatchenkia orientalis (KK5Y3) strain
(Fig. 1b) revealed only three bands ranging in size from
1Æ1 to 3Æ0 Mbp. The estimated chromosome number is at
least four, because of the increased fluorescence of the
�3Æ0 Mbp band. As far as we are concerned, this is the
first report of PFGE concerning I. orientalis.
Determination of the immunostimulatory activity of
yeast strains using the dorsal air pouch model
To determine the potential in vivo probiotic properties of
the yeast strains included in this study, we examined their
952
832
840a
982
KK
1
KK
2.1
KK
2.5
KK
3.1
KK
4.1
KK
6.5
KK
6ρ
KK
5Y3
NC
YC
9 41
4 H
. win
gei
50 60 70 80
92%
90 80
(a)
(b)
100 Strain
952 16
16
17
17
16
16
16
17
982
746
832
840a
UL
NCYC167 (R)
NCYC9 (R)
CBS1954 (R)(D)
570
630
KK2·5 (D)
KK4·1 (D)
KK2·1 (D)
KK3·1 (D)
Strain
KK5Y3
6 14 14 14 14 5
– >4
No chr
5 16 16 16 16
414
KK6·5 (D)
KK6P(D)
KK1
No chr
99%
81% 99%
II
II
I
III
I
90 100
570
630
CB
S19
54
H. w
inge
i
UL
746
1900
Kbp
1640 945 815 745 680 610 555 450 375 295 225
Mbp
3·13 2·70 2·35
1·81 1·66 1·37
1·05
0·08
NC
YC
167
Y
PH
80
YP
H80
Figure 1 Pulsed-field gel electrophoresis patterns and clustering of (a) Saccharomyces strains and (b) Candida, Isaatchenkia, Debaryomyces and
Kluyveromyces yeast strains. Similarities were estimated by the use of Pearson product moment correlation coefficient (r) and the unweighted pair
group algorithm with arithmetic averages (UPGMA). R, reference strain; D, diploid yeasts; Chr, chromosomes.
Characterization of candidate probiotic yeasts A. Kourelis et al.
264 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 260–271
ª 2010 The Authors
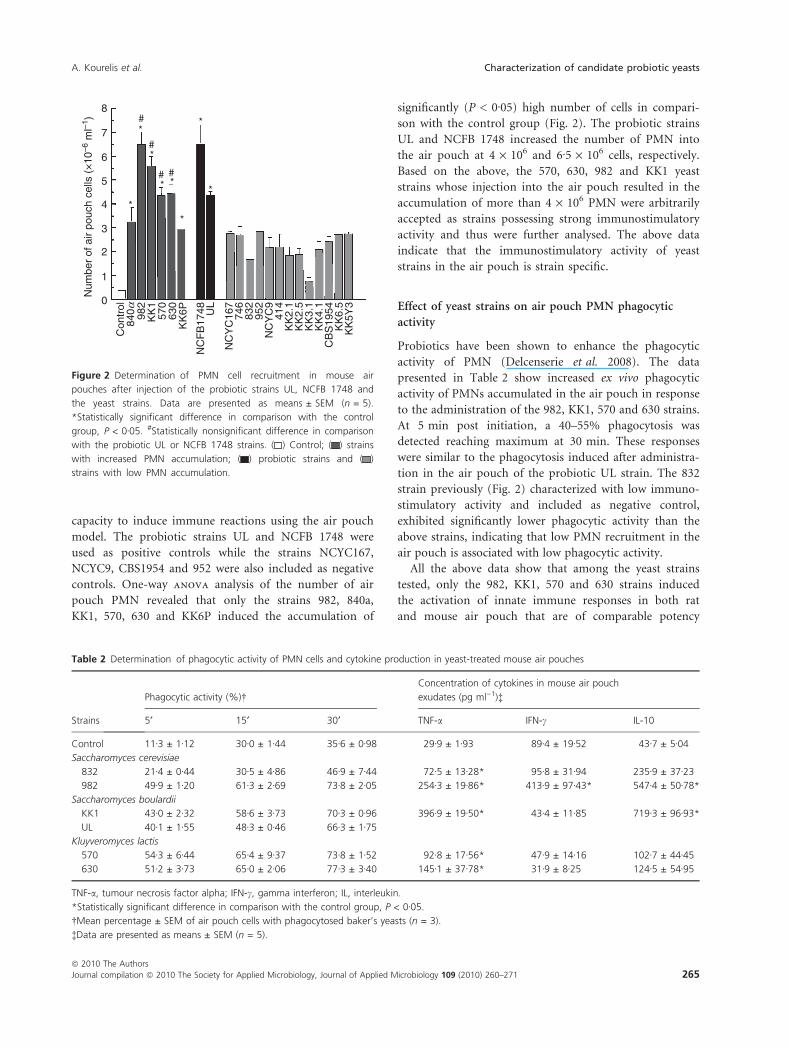
capacity to induce immune reactions using the air pouch
model. The probiotic strains UL and NCFB 1748 were
used as positive controls while the strains NCYC167,
NCYC9, CBS1954 and 952 were also included as negative
controls. One-way anova analysis of the number of air
pouch PMN revealed that only the strains 982, 840a,
KK1, 570, 630 and KK6P induced the accumulation of
significantly (P < 0Æ05) high number of cells in compari-
son with the control group (Fig. 2). The probiotic strains
UL and NCFB 1748 increased the number of PMN into
the air pouch at 4 · 106 and 6Æ5 · 106 cells, respectively.
Based on the above, the 570, 630, 982 and KK1 yeast
strains whose injection into the air pouch resulted in the
accumulation of more than 4 · 106 PMN were arbitrarily
accepted as strains possessing strong immunostimulatory
activity and thus were further analysed. The above data
indicate that the immunostimulatory activity of yeast
strains in the air pouch is strain specific.
Effect of yeast strains on air pouch PMN phagocytic
activity
Probiotics have been shown to enhance the phagocytic
activity of PMN (Delcenserie et al. 2008). The data
presented in Table 2 show increased ex vivo phagocytic
activity of PMNs accumulated in the air pouch in response
to the administration of the 982, KK1, 570 and 630 strains.
At 5 min post initiation, a 40–55% phagocytosis was
detected reaching maximum at 30 min. These responses
were similar to the phagocytosis induced after administra-
tion in the air pouch of the probiotic UL strain. The 832
strain previously (Fig. 2) characterized with low immuno-
stimulatory activity and included as negative control,
exhibited significantly lower phagocytic activity than the
above strains, indicating that low PMN recruitment in the
air pouch is associated with low phagocytic activity.
All the above data show that among the yeast strains
tested, only the 982, KK1, 570 and 630 strains induced
the activation of innate immune responses in both rat
and mouse air pouch that are of comparable potency
8 # *
*
*
* *
*
*
*
#
# #
7
6
5
4
3
Num
ber
of a
ir po
uch
cells
(×
10–6
ml–1
)
2
1
0
Con
trol
84
0a 982
KK
1 57
0 63
0 K
K6P
UL
NC
YC
167
NC
YC
9
746
832
952
414
KK
2.1
KK
2.5
KK
3.1
KK
4.1
KK
6.5
KK
5Y3
CB
S19
54
NC
FB
1748
Figure 2 Determination of PMN cell recruitment in mouse air
pouches after injection of the probiotic strains UL, NCFB 1748 and
the yeast strains. Data are presented as means ± SEM (n = 5).
*Statistically significant difference in comparison with the control
group, P < 0Æ05. #Statistically nonsignificant difference in comparison
with the probiotic UL or NCFB 1748 strains. ( ) Control; ( ) strains
with increased PMN accumulation; ( ) probiotic strains and ( )
strains with low PMN accumulation.
Table 2 Determination of phagocytic activity of PMN cells and cytokine production in yeast-treated mouse air pouches
Strains
Phagocytic activity (%)�
Concentration of cytokines in mouse air pouch
exudates (pg ml)1)�
5¢ 15¢ 30¢ TNF-a IFN-c IL-10
Control 11Æ3 ± 1Æ12 30Æ0 ± 1Æ44 35Æ6 ± 0Æ98 29Æ9 ± 1Æ93 89Æ4 ± 19Æ52 43Æ7 ± 5Æ04
Saccharomyces cerevisiae
832 21Æ4 ± 0Æ44 30Æ5 ± 4Æ86 46Æ9 ± 7Æ44 72Æ5 ± 13Æ28* 95Æ8 ± 31Æ94 235Æ9 ± 37Æ23
982 49Æ9 ± 1Æ20 61Æ3 ± 2Æ69 73Æ8 ± 2Æ05 254Æ3 ± 19Æ86* 413Æ9 ± 97Æ43* 547Æ4 ± 50Æ78*
Saccharomyces boulardii
KK1 43Æ0 ± 2Æ32 58Æ6 ± 3Æ73 70Æ3 ± 0Æ96 396Æ9 ± 19Æ50* 43Æ4 ± 11Æ85 719Æ3 ± 96Æ93*
UL 40Æ1 ± 1Æ55 48Æ3 ± 0Æ46 66Æ3 ± 1Æ75
Kluyveromyces lactis
570 54Æ3 ± 6Æ44 65Æ4 ± 9Æ37 73Æ8 ± 1Æ52 92Æ8 ± 17Æ56* 47Æ9 ± 14Æ16 102Æ7 ± 44Æ45
630 51Æ2 ± 3Æ73 65Æ0 ± 2Æ06 77Æ3 ± 3Æ40 145Æ1 ± 37Æ78* 31Æ9 ± 8Æ25 124Æ5 ± 54Æ95
TNF-a, tumour necrosis factor alpha; IFN-c, gamma interferon; IL, interleukin.
*Statistically significant difference in comparison with the control group, P < 0Æ05.
�Mean percentage ± SEM of air pouch cells with phagocytosed baker’s yeasts (n = 3).
�Data are presented as means ± SEM (n = 5).
A. Kourelis et al. Characterization of candidate probiotic yeasts
ª 2010 The Authors
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 260–271 265

with the respective responses induced by the probiotic UL
and NCFB 1748 strains.
Determination of TNF-a, IFN-c and IL-10 in air pouch
exudates
To examine whether the above yeast strains induce
increased production of cytokines in the air pouch, we
performed ELISA immunosorbent assay in the air pouch
exudates. The data in Table 2 indicate differential ability
of the yeast strains tested to induce a specific cytokine
profile, i.e. TNF-a, IFN-c and IL-10. The 982 strain
induced increased production of TNF-a, IFN-c and IL-10;
the KK1 strain induced the production of TNF-a and
IL-10, while the strains 570 and 630 induced only TNF-aproduction (Table 3). These data show strain-specific
activation of the cells forming the air pouch that in turn
produce and secrete certain profile of cytokines.
All the above air pouch model data demonstrate that
the strains 982, KK1, 570 and 630 exhibited immunostim-
ulatory activity similar to that induced by the probiotics
UL and NCFB 1748 suggesting potential activation of the
immune responses in the gut mucosa as well.
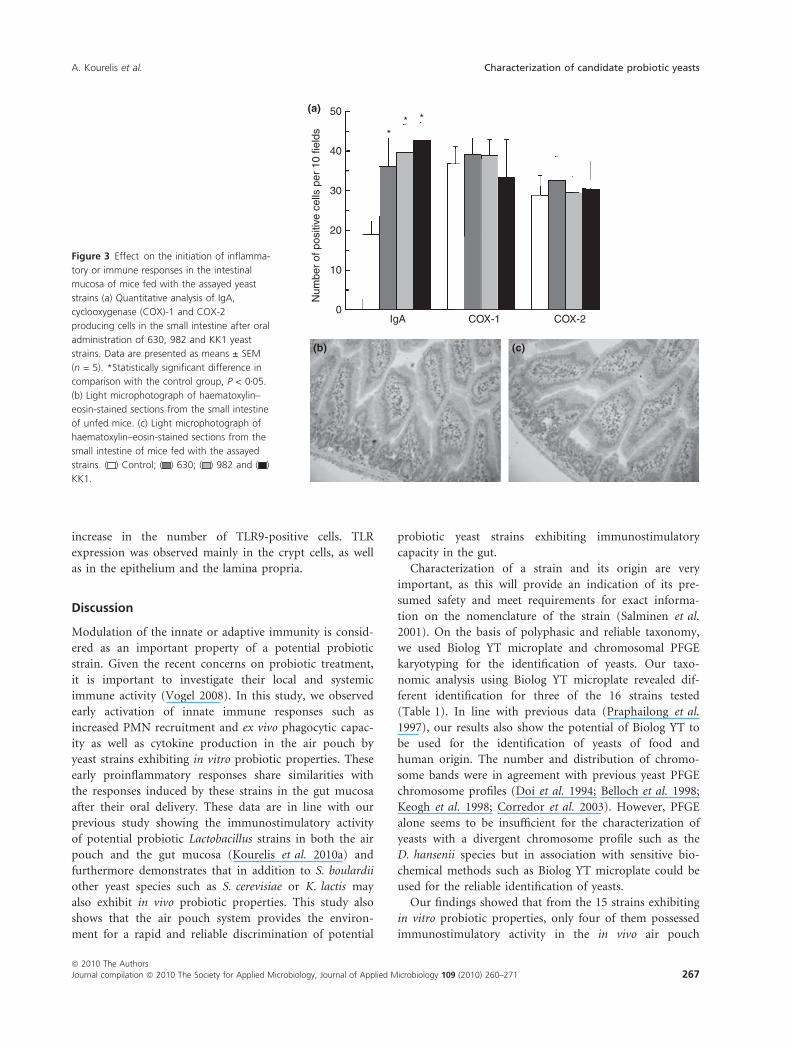
Determination of IgA, COX-1, COX-2, cytokine and TLR
producing cells in the gut mucosa
To examine whether a selected set of yeast strains (630,
982, KK1) induce similar immune responses in the air
pouch and the intestine, they were orally administered to
mice for 10 days. We have previously shown that several
Lactobacillus strains exhibit similar immunostimulatory
activity in the small and large intestine (Kourelis et al.
2010a). However, higher responses concerning cytokine
and TLR production were observed in the small intestine.
Thus, yeast strains’ immunostimulatory activity was
determined only in the small intestine. The two K. lactis
(570, 630) strains showed similar PFGE chromosome pro-
file (Fig. 1) and exhibited similar immune effects in the
air pouch indicating that they potentially belong to the
same strain. Thus, only the 630, 982 and KK1 strains
were further analysed.
All strains increased the number of IgA producing cells
in the small intestine lamina propria with the KK1 strain
exhibiting the highest response (Fig. 3a, Table 3). This
induction occurred in the absence of inflammatory
immune response because yeast oral administration did
not increase the number of cells producing the inflamma-
tory mediators COX-1 and COX-2. In addition, there was
no evidence of any morphological alteration in the small
intestine or modification in the structure of the villi
(Fig. 3b,c).
All three yeast strains induced a significant increase in
the number of TNF-a, IL-5, IL-6 and IL-12 positive cells
(P < 0Æ05) in comparison with the control group (Fig. 4a,
Table 3). Moreover, the strains 982 and KK1 increased
the number of IL-10 producing cells, while only the strain
982 caused a significant increase in the number of IFN-cproducing cells. All cytokine producing cells were
detected in the epithelium and the lamina propria
(Fig. 4b,c). The above data show that the three yeasts
exhibit differential activation of cytokine production in
the small intestine that is similar to the response induced
by these strains in the air pouch.
To determine the signalling events leading to the
induction of the above cytokine profile, we also examined
the TLR expression in small intestine cells. Oral adminis-
tration of the yeast strains 630, 982 and KK1 resulted in a
significant increase in the number of intestinal cells
expressing TLR2 and TLR6 (Fig. 5, Table 3). In addition,
the strains 630 and 982 increased the number of TLR4-
positive cells, while the 982 strain also caused a significant
Table 3 Comparison of the immune responses induced in the air pouch and the small intestine of mice after administration of yeast strains
Strains*
Air pouch Small intestine
Chemotaxis Phagocytosis
Cytokine production IgA and cytokine production TLR expression
TNF-a IFN-c IL-10 IgA TNF-a IFN-c IL-5 IL-6 IL-10 IL-12 TLR2 TLR4 TLR6 TLR9
Saccharomyces cerevisiae
832 ) ) + ) )982 + + + + + + + + + + + + + + + +
Saccharomyces boulardii
KK1 + + + ) + + + ) + + + + + ) + )UL + +
Kluyveromyces lactis
570 + + + ) )630 + + + ) ) + + ) + + ) + + + + )
TNF-a, tumour necrosis factor alpha; IFN-c, gamma interferon; IL, interleukin; TLR, Toll-like receptors.
*The yeast strains exhibiting low chemotaxis are not presented in this table.
Characterization of candidate probiotic yeasts A. Kourelis et al.
266 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 260–271
ª 2010 The Authors
increase in the number of TLR9-positive cells. TLR
expression was observed mainly in the crypt cells, as well
as in the epithelium and the lamina propria.
Discussion
Modulation of the innate or adaptive immunity is consid-
ered as an important property of a potential probiotic
strain. Given the recent concerns on probiotic treatment,
it is important to investigate their local and systemic
immune activity (Vogel 2008). In this study, we observed
early activation of innate immune responses such as
increased PMN recruitment and ex vivo phagocytic capac-
ity as well as cytokine production in the air pouch by
yeast strains exhibiting in vitro probiotic properties. These
early proinflammatory responses share similarities with
the responses induced by these strains in the gut mucosa
after their oral delivery. These data are in line with our
previous study showing the immunostimulatory activity
of potential probiotic Lactobacillus strains in both the air
pouch and the gut mucosa (Kourelis et al. 2010a) and
furthermore demonstrates that in addition to S. boulardii
other yeast species such as S. cerevisiae or K. lactis may
also exhibit in vivo probiotic properties. This study also
shows that the air pouch system provides the environ-
ment for a rapid and reliable discrimination of potential
probiotic yeast strains exhibiting immunostimulatory
capacity in the gut.
Characterization of a strain and its origin are very
important, as this will provide an indication of its pre-
sumed safety and meet requirements for exact informa-
tion on the nomenclature of the strain (Salminen et al.
2001). On the basis of polyphasic and reliable taxonomy,
we used Biolog YT microplate and chromosomal PFGE
karyotyping for the identification of yeasts. Our taxo-
nomic analysis using Biolog YT microplate revealed dif-
ferent identification for three of the 16 strains tested
(Table 1). In line with previous data (Praphailong et al.
1997), our results also show the potential of Biolog YT to
be used for the identification of yeasts of food and
human origin. The number and distribution of chromo-
some bands were in agreement with previous yeast PFGE
chromosome profiles (Doi et al. 1994; Belloch et al. 1998;
Keogh et al. 1998; Corredor et al. 2003). However, PFGE
alone seems to be insufficient for the characterization of
yeasts with a divergent chromosome profile such as the
D. hansenii species but in association with sensitive bio-
chemical methods such as Biolog YT microplate could be
used for the reliable identification of yeasts.
Our findings showed that from the 15 strains exhibiting
in vitro probiotic properties, only four of them possessed
immunostimulatory activity in the in vivo air pouch
(a) 50
** *
40
30
20
10
0N
umbe
r of
pos
itive
cel
ls p
er 1
0 fie
lds
IgA COX-1 COX-2
(b) (c)
Figure 3 Effect on the initiation of inflamma-
tory or immune responses in the intestinal
mucosa of mice fed with the assayed yeast
strains (a) Quantitative analysis of IgA,
cyclooxygenase (COX)-1 and COX-2
producing cells in the small intestine after oral
administration of 630, 982 and KK1 yeast
strains. Data are presented as means ± SEM
(n = 5). *Statistically significant difference in
comparison with the control group, P < 0Æ05.
(b) Light microphotograph of haematoxylin–
eosin-stained sections from the small intestine
of unfed mice. (c) Light microphotograph of
haematoxylin–eosin-stained sections from the
small intestine of mice fed with the assayed
strains. ( ) Control; ( ) 630; ( ) 982 and ( )
KK1.
A. Kourelis et al. Characterization of candidate probiotic yeasts
ª 2010 The Authors
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 260–271 267

model. The air pouch provides an excellent system to dis-
criminate presumptive probiotic yeast strains, because it
shows similar immune responses with the intestine and
outweighs the in vitro assays such as cell culture experi-
ments used for probiotic characterization. In addition, the
potential immunostimulatory activity in the air pouch
could be determined within 3 h after a single injection of
yeast cells and outweighs the delivery in the intestinal tract
that requires daily administration for at least 7 days. We
have demonstrated the K. lactis 570 and 630 strains, the
S. cerevisiae 982 strain and the S. boulardii KK1 strain
exhibit in the air pouch increased PMN influx, phagocytic
ability and cytokine production of comparable potency
with the probiotics UL and NCFB 1748 (Fig. 1, Table 2,
Kourelis et al. 2010a). Strains negative for in vitro probiot-
ic properties such as 952 and strains positive for in vitro
probiotic properties such as 832 were used as low
responders concerning PMN recruitment and phagocytosis
suggesting potential strain-specific regulation of immune
proinflammatory reactions in the air pouch.
Probiotic yeast strains of the species S. boulardii have
been shown to increase the phagocytic activity and cyto-
kine production (Rodrigues et al. 2000; Czerucka et al.
2007). The strains 570, 630, 982 and KK1 can positively
influence innate immune responses, indicating that these
strains may exhibit pathogen clearance activity. Although
the dorsal air pouch does not simulate the intestinal func-
tion, the air pouch lining tissue and the intestine mucosa
share similarities as they both exhibit a barrier function
that is achieved by epithelial cells and resident macro-
phages (Sedgwick et al. 1983; Wittig and Zeitz 2003;
Magalhaes et al. 2007). Furthermore, the above yeast
(b) (c)
120(a)
100
80
60
**
*
* * * *
* *
*
*
* * * *
40
20
0TNF-a IFN-g IL-5 IL-6 IL-10 IL-12B
Num
ber
of p
ositi
ve c
ells
per
10
field
s
Figure 4 Effect of the assayed yeast strains
on the cytokine profile of mice small intestine
(a) Quantitative analysis of tumour necrosis fa-
ctor alpha (TNF-a), gamma interferon (IFN-c),
interleukin (IL)-5, IL-6, IL-10 and IL-12 produc-
ing cells after oral administration of 630, 982
and KK1 yeast strains. Data are presented as
means ± SEM (n = 5). *Statistically significant
difference in comparison with the control gro-
up, P < 0Æ05. (b) Microphotography of IL-6-p-
ositive cells in the epithelium and lamina
propria of the small intestine of (control) unfe-
d mice. (c) Microphotography of cytokine-pos-
itive cells in the epithelium and lamina propria
of the small intestine of mice fed with the ass-
ayed strains. ( ) Control; ( ) 630; ( ) 982 a-
nd ( ) KK1.
80
70
60
50
*
*
*
*
*
*
*
*
*
40
30
20
10
0TLR2 TLR4 TLR6 TLR9
Num
ber
of p
ositi
ve c
ells
per
10
field
s
Figure 5 Quantitative analysis of Toll-like receptor (TLR)2, TLR4, TLR6
and TLR9 producing cells in the small intestine after oral administra-
tion of 630, 982 and KK1 yeast strains. Data are presented as
means ± SEM (n = 5). *Statistically significant difference in compari-
son with the control group, P < 0Æ05. ( ) Control; ( ) 630; ( ) 982
and ( ) KK1.
Characterization of candidate probiotic yeasts A. Kourelis et al.
268 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 260–271
ª 2010 The Authors

strains interact with the cells forming the air pouch lining
tissue and the gut-associated lymphoid tissue (GALT)
with subsequent increase in cytokine production. In addi-
tion, the cytokine profile of the 630, 982 and KK1 strains
in the air pouch was similar to the cytokine profile
observed in the small intestine after oral administration
of these strains (Table 3). Thus, the interactivity of the
yeast strains with the air pouch may be the signal initiat-
ing the proinflammatory response by the production of
species-specific cytokines. TNF-a is necessary to initiate
the crosstalk between the immune cells of the lamina pro-
pria and the intestine epithelial cells (Galdeano et al.
2007). The increase in TNF-a in the air pouch could sug-
gest that TNF-a may be required to initiate the interac-
tion among the epithelial cells, the PMN cells and the
resident macrophages of the air pouch membrane. The
precise mechanism by which 570, 630, 982 and KK1 exert
immunostimulatory activity in the air pouch remains
unclear in this study. A possible mechanism could involve
the rapid activation of TLRs (Doyle et al. 2004), the com-
plement cascade and the production of chemokines (Yam
et al. 2008). These chemotactic factors increase the
recruitment of PMNs exhibiting strong phagocytic activity
(Coates and McColl 2001) and the subsequent production
of cytokines such as TNF-a, IFN-c (Diamond et al. 1991;
Utsunomiya et al. 1998).
The interaction of yeast strains with the GALT is not
followed by the activation of an inflammatory response
because their oral delivery does not result in morphologi-
cal changes of the villi, in massive accumulation of PMNs
or in increased number of COX-1 and COX-2 producing
cells (Fig. 3). These differences in the small intestine vs
the air pouch may be attributed to the higher number of
resident immune cells and yeast antigen availability in the
air pouch than in the intestine.
Oral administration of 630, 982 and KK1 strains caused
a significant increase in the number of IgA producing
cells. Such a response has been previously reported for
the probiotic S. boulardii species (Rodrigues et al. 2000).
Increased IgA production suggests a potential role of
these strains in the immunity against invading pathogenic
microorganisms and the maintenance of intestinal
homeostasis (Fagarasan and Honjo 2004).
Interaction of yeast strains with the gut mucosa acti-
vated a strain-specific profile of cytokines and TLRs
(Fig. 4, Table 3). The strain 630 induced the production
of TNF-a, IL-5, IL-6 and IL-12 in a TLR2, -4, -6 depen-
dent manner indicating no bias towards Th1 or Th2 ⁄ Treg
intestinal immune response. Furthermore, the increased
number of IL-5 and IL-6 producing cells could also con-
tribute in the maintenance of tolerance as well as IgA
switch and secretion in the gut mucosa (Corthesy et al.
2007). The strain 982 induced the production of TNF-a,
IFN-c, IL-5, IL-6, IL-10 and IL-12 through TLR2, -4, -6,
-9. The cytokine profile of increased levels of both IL-12
and IFN-c by the 982 strain indicates that this strain may
favour the induction of a Th1 response. The strain KK1
activated the profile of TNF-a, IL-5, IL-6, IL-10 and
IL-12 through TLR2, -6. The KK1 strain may favour the
induction of a Th2 ⁄ Treg response as a result of the pro-
duction of IL-5 and IL-10. The Saccharomyces strains
(982 and KK1) may also exert a potent anti-inflammatory
activity in the intestine through the production of IL-10
that would prevent Th1-driven local and systemic inflam-
matory responses (Rennick et al. 1992). Oral administra-
tion of the KK1 strain in gluten-sensitive mice inhibited
the induction of gluten-induced enteropathy that is
characterized by small intestine inflammatory lesions and
specific Th1 responses (V. Gerakopoulos unpublished
data). Activation of combinations of certain TLRs by the
three yeast strains is required for the high discriminatory
ability of TLRs (Roeder et al. 2004). Our findings suggest
that the immunostimulatory activity of the yeast strains
examined is mediated through the interaction of TLR4
with the cell wall glucan present on the strains 630 and
982, or through TLR2, -6 heterodimer interaction with
mannan present on the strains 630, 982 and KK1 (Roeder
et al. 2004). In addition, mannan or zymosan CpG–DNA
complexes recognized by TLR9 (Anada et al. 2006; Dan
et al. 2008) may also be responsible for the immuno-
stimulatory activity of the 982 strain.
In conclusion, a combination of phenotypic, biochemi-
cal and genotypic assays should be used for the identifica-
tion of yeasts. The strains S. cerevisiae 982, S. boulardii
KK1 and K. lactis 630 could be considered as candidate
probiotics, further supporting the use of other yeast
strains besides S. boulardii as probiotics. Furthermore, the
dorsal air pouch model may be used to rapidly and effi-
ciently discriminate among presumptive probiotic yeast
strains with immunostimulatory activity. Under normal
conditions, this stimulation of the host’s mucosal immune
system by nonpathogenic microorganisms, through a
highly regulated system of TLR and cytokine expression,
may have favourable effects on the development and
maintenance of the immune system and may provide
the host with a higher capacity to resist any inflamma-
tory response. Finally, further experimentation is
required using disease animal models to confirm the
immunomodulatory activity of the above yeast strains
while final validation of the findings of this study must
await clinical trials.
References
Anada, T., Okada, N., Minari, J., Karinaga, R., Mizu, M.,
Koumoto, K., Shinkai, S. and Sakurai, K. (2006) CpG
A. Kourelis et al. Characterization of candidate probiotic yeasts
ª 2010 The Authors
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 260–271 269

DNA ⁄ zymosan complex to enhance cytokine secretion
owing to the cocktail effect. Bioorg Med Chem Lett 16,
1301–1304.
Andrighetto, C., Psomas, E., Tzanetakis, N., Suzzi, G. and
Lombardi, A. (2000) Randomly amplified polymorphic
DNA (RAPD) PCR for the identification of yeasts isolated
from dairy products. Lett Appl Microbiol 30, 5–9.
Belloch, C., Barrio, E., Garcia, M.D. and Querol, A. (1998)
Inter- and intraspecific chromosome pattern variation in
the yeast genus Kluyveromyces. Yeast 14, 1341–1354.
Borriello, S.P., Hammes, W.P., Holzapfel, W., Marteau, P.,
Schrezenmeir, J., Vaara, M. and Valtonen, V. (2003) Safety
of probiotics that contain lactobacilli or bifidobacteria.
Clin Infect Dis 36, 775–780.
Coates, N.J. and McColl, S.R. (2001) Production of chemokin-
es in vivo in response to microbial stimulation. J Immunol
166, 5176–5182.
Corredor, M., Davila, A.M., Casaregola, S. and Gaillardin, C.
(2003) Chromosomal polymorphism in the yeast species
Debaryomyces hansenii. Antonie Van Leeuwenhoek 84, 81–88.
Corthesy, B., Gaskins, H.R. and Mercenier, A. (2007) Cross-
talk between probiotic bacteria and the host immune sys-
tem. J Nutr 137, 781S–790S.
Czerucka, D., Piche, T. and Rampal, P. (2007) Review article:
yeast as probiotics – Saccharomyces boulardii. Aliment
Pharmacol Ther 26, 767–778.
Dan, J.M., Wang, J.P., Lee, C.K. and Levitz, S.M. (2008)
Cooperative stimulation of dendritic cells by Cryptococcus
neoformans mannoproteins and CpG oligodeoxynucleo-
tides. PLoS ONE 3, e2046.
Delcenserie, V., Martel, D., Lamoureux, M., Amiot, J., Boutin,
Y. and Roy, D. (2008) Immunomodulatory effects of probi-
otics in the intestinal tract. Curr Issues Mol Biol 10, 37–54.
Diamond, R.D., Lyman, C.A. and Wysong, D.R. (1991) Dispa-
rate effects of interferon-gamma and tumor necrosis fac-
tor-alpha on early neutrophil respiratory burst and
fungicidal responses to Candida albicans hyphae in vitro.
J Clin Invest 87, 711–720.
Doi, M., Homma, M., Iwaguchi, S., Horibe, K. and Tanaka, K.
(1994) Strain relatedness of Candida albicans strains iso-
lated from children with leukemia and their bedside par-
ents. J Clin Microbiol 32, 2253–2259.
Doyle, S.E., O’Connell, R.M., Miranda, G.A., Vaidya, S.A.,
Chow, E.K., Liu, P.T., Suzuki, S., Suzuki, N. et al. (2004)
Toll-like receptors induce a phagocytic gene program
through p38. J Exp Med 199, 81–90.
Fagarasan, S. and Honjo, T. (2004) Regulation of IgA synthesis
at mucosal surfaces. Curr Opin Immunol 16, 277–283.
FAO ⁄ WHO (2002) Guidelines for the Evaluation of Probiotics
in Food. London Ontario, Canada: Food and Agriculture
Organization of the United Nations and World Health
Organization Working Group Report.
Fleet, G.H. (1990) Yeasts in dairy products. J Appl Bacteriol 68,
199–211.
Galdeano, C.M., de Moreno de LeBlanc, A., Vinderola, G.,
Bonet, M.E. and Perdigon, G. (2007) Proposed model:
mechanisms of immunomodulation induced by probiotic
bacteria. Clin Vaccine Immunol 14, 485–492.
Hessle, C., Hanson, L.A. and Wold, A.E. (1999) Lactobacilli
from human gastrointestinal mucosa are strong stimulators
of IL-12 production. Clin Exp Immunol 116, 276–282.
Jakobsen, M. and Narvhus, J. (1996) Yeasts and their possible
beneficial and negative effects on the quality of dairy prod-
ucts. Int Dairy J 6, 755–768.
Jespersen, L., van der Kuhle, A. and Petersen, K.M. (2000)
Phenotypic and genetic diversity of Saccharomyces contam-
inants isolated from lager breweries and their phylogenetic
relationship with brewing yeasts. Int J Food Microbiol 60,
43–53.
Keogh, R.S., Seoighe, C. and Wolfe, K.H. (1998) Evolution of
gene order and chromosome number in Saccharomyces,
Kluyveromyces and related fungi. Yeast 14, 443–457.
Kourelis, A., Zinonos, I., Kakagianni, M., Christidou, A.,
Christoglou, N., Yiannaki, E., Testa, T., Kotzamanidis, C.
et al. (2010a) Validation of the dorsal air pouch model to
predict and examine immunostimulatory responses in the
gut. J Appl Microbiol 108, 274–284.
Kourelis, A., Kotzamanidis, C., Litopoulou-Tzanetaki, E.,
Scouras, Z.G., Tzanetakis, N. and Yiangou, M. (2010b)
Preliminary probiotic selection of dairy and human yeast
strains. J Biol Res 13, in press.
Loose, L.D., Silkworth, J.B. and Simpson, D.W. (1978) Influ-
ence of cadmium on the phagocytic and microbicidal
activity of murine peritoneal macrophages, pulmonary
alveolar macrophages, and polymorphonuclear neutrophils.
Infect Immun 22, 378–381.
Magalhaes, J.G., Tattoli, I. and Girardin, S.E. (2007) The intes-
tinal epithelial barrier: how to distinguish between the
microbial flora and pathogens. Semin Immunol 19, 106–
115.
Neish, A.S. (2002) The gut microflora and intestinal epithelial
cells: a continuing dialogue. Microbes Infect 4, 309–317.
van Niel, C.W., Feudtner, C., Garrison, M.M. and Christakis,
D.A. (2002) Lactobacillus therapy for acute infectious
diarrhea in children: a meta-analysis. Pediatrics 109, 678–
684.
Praphailong, W., Van Gestel, M., Fleet, G.H. and Heard, G.M.
(1997) Evaluation of the Biolog system for the identifica-
tion of food and beverage yeasts. Lett Appl Microbiol 24,
455–459.
Psomas, E., Andrighetto, C., Litopoulou-Tzanetaki, E.,
Lombardi, A. and Tzanetakis, N. (2001) Some probiotic
properties of yeast isolates from infant faeces and Feta
cheese. Int J Food Microbiol 69, 125–133.
Psomas, E.I., Fletouris, D.J., Litopoulou-Tzanetaki, E. and
Tzanetakis, N. (2003) Assimilation of cholesterol by yeast
strains isolated from infant feces and Feta cheese. J Dairy
Sci 86, 3416–3422.
Characterization of candidate probiotic yeasts A. Kourelis et al.
270 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 260–271
ª 2010 The Authors

Rennick, D., Berg, D. and Holland, G. (1992) Interleukin 10:
an overview. Prog Growth Factor Res 4, 207–227.
Rodrigues, A.C., Cara, D.C., Fretez, S.H., Cunha, F.Q.,
Vieira, E.C., Nicoli, J.R. and Vieira, L.Q. (2000) Saccha-
romyces boulardii stimulates sIgA production and the
phagocytic system of gnotobiotic mice. J Appl Microbiol
89, 404–414.
Roeder, A., Kirschning, C.J., Rupec, R.A., Schaller, M. and
Korting, H.C. (2004) Toll-like receptors and innate anti-
fungal responses. Trends Microbiol 12, 44–49.
Sainte-Marie, G. (1962) A paraffin embedding technique for
studies employing immunofluorescence. J Histochem
Cytochem 10, 250.
Salminen, S.J., von Wright, A.J., Ouwehand, A.C. and
Holzapfel, W.H. (2001) Safety assessment of probiotics
and starters. In Fermentation and Food Safety ed. Adams,
M.R. and Nout, M.J.R. pp. 239–251. Gaithersburg: Aspen
Publishers.
Sargent, G. and Wickens, H. (2004) Brewers’ yeast in C. diffi-
cile infection: probiotic or B-group vitamins? Pharm J 273,
230–231.
Sedgwick, A.D., Sin, Y.M., Edwards, J.C. and Willoughby, D.A.
(1983) Increased inflammatory reactivity in newly formed
lining tissue. J Pathol 141, 483–495.
Steinkamp-Zucht, A. and Fahrig, R. (1995) Monitoring of
induced chromosomal aberrations in S. cerevisiae in aga-
rose gels by pulsed field gel electrophoresis. Mutat Res 335,
285–292.
Tuohy, K.M., Probert, H.M., Smejkal, C.W. and Gibson, G.R.
(2003) Using probiotics and prebiotics to improve gut
health. Drug Discov Today 8, 692–700.
Tzanetakis, N., Hatzikamari, M. and Litopoulou-Tzanetaki, E.
(1996) Yeasts on the surface microflora of Feta cheese.
FIL-IDF Yeasts in the Dairy Industry: Positive and Nega-
tive Aspects Proceedings of the Symposium Organised by
Group F47 held in Copenhagen (Denmark), pp. 34–43.
Utsunomiya, I., Ito, M. and Oh-ishi, S. (1998) Generation of
inflammatory cytokines in zymosan-induced pleurisy in
rats: TNF induces IL-6 and cytokine-induced neutrophil
chemoattractant (CINC) in vivo. Cytokine 10, 956–963.
Vogel, G. (2008) Clinical trials. Deaths prompt a review of
experimental probiotic therapy. Science 319, 557.
Wittig, B.M. and Zeitz, M. (2003) The gut as an organ of
immunology. Int J Colorectal Dis 18, 181–187.
Yam, K.K., Pouliot, P., N’Diaye M, M., Fournier, S., Olivier,
M. and Cousineau, B. (2008) Innate inflammatory
responses to the Gram-positive bacterium Lactococcus
lactis. Vaccine 26, 2689–2699.
A. Kourelis et al. Characterization of candidate probiotic yeasts
ª 2010 The Authors
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 260–271 271
Related Documents











