IMMUNOLOGY BCR-dependent lineage plasticity in mature B cells Robin Graf 1 *†, Jane Seagal 2 *‡, Kevin L. Otipoby 2 *§, Kong-Peng Lam 3 #, Salah Ayoub 4 , Baochun Zhang 2,5 , Sandrine Sander 1,6 , Van Trung Chu 1,7 , Klaus Rajewsky 1,2,3 † B2 cells engage in classical antibody responses, whereas B1 cells are considered carriers of innate immunity, biased toward recognizing epitopes present on the surfaces of common pathogens and self antigens.To explore the role of B cell antigen receptor (BCR) specificity in driving B1 cell differentiation, we developed a transgenic system allowing us to change BCR specificity in B cells in an inducible and programmed manner. Mature B2 cells differentiated into bona fide B1 cells upon acquisition of a B1 cell–typical self-reactive BCR through a phase of proliferative expansion. Thus, B2 cells have B1 cell differentiation potential in addition to their classical capacity to differentiate into memory and plasma cells, and B1 differentiation can be instructed by BCR-mediated self-reactivity, in the absence of B1-lineage precommitment. B cells are the carriers of humoral immunity. On the basis of their phenotype and func- tional properties, mature B cells can be subdivided into B1 and B2 cells, with the former further divided into the CD5 + B1a and CD5 - B1b subsets. B2 cells form follicles in the spleen and lymph nodes and are responsible for generating specific antibody responses against foreign antigens, typically involving T cell– dependent affinity maturation and somatic hypermutation of the B cell antigen receptor (BCR) (1). In contrast, B1 cells are predominantly found in peritoneal and pleural cavities and produce natural antibodies, providing a “first line of defense” against common bacterial pathogens and contributing to the clearance of apoptotic cells and oxidized lipids (2, 3). Therefore, B1 cells are thought to perform innate-like functions in the immune system, predominantly expressing evolutionarily selected BCRs with low levels of somatic mutation and junctional diversity (4–6). Despite major advances in the field, the origin of B1 cells remains controversial. The “lineage model” posits that B1 and B2 cells represent sep- arate lineages that arise from different pro- genitor populations, committed to either lineage prior to BCR expression. In the case of B1 cells, this possibly even occurs prior to hematopoietic stem cell differentiation (7–10). As recently re- viewed in detail, definitive evidence for the line- age model is still lacking, with both putative BCR-negative B1 progenitors and exclusive pro- genitor commitment to B2 differentiation being controversial (11, 12). In the “selection model,” B cell progenitors are instructed by BCR speci- ficity to differentiate into B1 cells, depending on BCR-mediated recognition of common bacterial and certain self antigens (2, 13). Although strong evidence indeed indicates that certain BCR spec- ificities can exclusively drive B1 cell differentia- tion, this does not rule out a possible commitment of progenitor cells to either lineage prior to BCR expression (14). However, as demonstrated below, the existence of B1 and B2 cell–typical BCRs can be exploited to test lineage commitment and its control by BCR specificity in mature B cells. To address this issue, we generated a genetic system that would allow a BCR-dependent inter- conversion of mature B1 and B2 cells. For this purpose, we targeted two prearranged immuno- globulin heavy (IgH) chain variable (V) region gene segments—V H 12 and B1-8, respectively— in a head-to-head orientation and flanked by inverted loxP sites into the J H region of the IgH locus. Cre-mediated recombination results in in- version of this V H 12B1-8 fli cassette, leading to a situation in which some cells express B1-8 and others V H 12 IgH chains. In combination with Vk4 Ig light chains, the V H 12 and B1-8 IgH chains form B1 and B2 cell–typical BCRs, respectively (14, 15). V H 12B1-8 fli mice (hereafter termed B2 mice) and B1-8V H 12 fli mice (hereafter termed B1 mice) should predominantly develop B cells with either B1-8/Vk4 or V H 12/Vk4 BCRs (Fig. 1A). In agreement with previous work, the knocked- in BCRs allowed the developing cells to circum- vent the pro/pre–B cell stage (fig. S1, A to E) (15). When the B cells in these animals were stained with the BCR-specific anti-idiotypic antibodies Ac146 and 5C5, respectively (14, 16), essentially all splenic B cells of B2 mice were Ac146 + and of the B2 cell phenotype, characterized by their small size and a CD19 + , B220 + , IgD hi , CD5 - , CD23 +/lo , and CD43 - surface phenotype. In contrast, B cells of B1 mice were 5C5 + and acquired the B1 cell phenotype, characterized by large cell size and a CD19 hi , B220 lo , IgD lo , CD5 + , CD23 - , and CD43 + surface phenotype (Fig. 1B). This is consistent with the natural expression of this BCR that is specific for phosphatidylcholine (PtC) and ex- clusively expressed on B1 cells of wild-type mice (13) (fig. S1D). PtC is a common membrane phos- pholipid and is exposed on senescent red blood cells, which suggests that V H 12/Vk4 antibodies are involved in the clearance of these cells (17, 18). When splenic B cells of B1 and B2 mice were isolated and transduced with TAT-Cre in vitro, approximately 10% of these cells inverted the IgH insertion cassette (fig. S1G). This led to de- tectable populations of cells that switched from the expression of the original BCR to that of the new BCR, thereby confirming the functionality of the transgenic system (Fig. 1C and fig. S1F). For simplicity, we refer to B1-8V H 12 fli – and V H 12B1-8 fli –derived switched B cells as B1→B2 and B2→B1 cells, respectively. To test whether a change of BCR specificity would change the phenotype of fully mature B cells, we treated B1 and B2 mice for 2 weeks with antibodies against the interleukin-7 receptor (IL-7R) to block the influx of immature B cells into the spleen (fig. S2). Purified splenic B cells from these mice were then transduced with TAT- Cre in vitro. Only very low numbers of B1→B2 cells developed. In contrast, B2→B1 cells devel- oped into a dominant population (Fig. 1C and fig. S1F). However, although these cells acquired some B1-typical markers, their phenotypic change was only partial in vitro (fig. S3). We therefore transferred IgM a experimental (TAT-Cre trans- duced) mature B cells together with IgM b carrier B cells into immunodeficient recipients to study the consequences of the BCR switch in a more physiological environment (Fig. 2A) (19). Con- sistent with the in vitro data, only low numbers of B1→B2 cells were recovered from the recipi- ents. These were located predominantly in the peritoneal cavity and had acquired a phenotype intermediate between B1 and B2 (fig. S4). In contrast, B2→B1 cells expressing the 5C5 idio- type were readily detectable in the spleen and peritoneal cavity of the recipients 4, 8, and 30 days after transfer and became the dominant population among the IgM a experimental cells (Fig. 2B). Contributing to this dominance was an initial, transient phase of rapid proliferation, similar to the clonal expansion of PtC-specific B1 cells observed in wild-type mice over the first 3 to 4 weeks of life (Fig. 2C and fig. S5A) (4, 20). Strikingly, after completion of this expansion phase, the B2→B1 cells became resting cells that phenotypically were essentially indistinguishable from bona fide B1 cells. B2→B1 cells reproducibly increased in size, down-regulated B220 and IgD, lost CD23 and CD21 expression, and increased RESEARCH Graf et al., Science 363, 748–753 (2019) 15 February 2019 1 of 6 1 Immune Regulation and Cancer, Max Delbrück Center for Molecular Medicine in the Helmholtz Association, 13125 Berlin, Germany. 2 Program in Cellular and Molecular Medicine, Children’s Hospital, and Immune Disease Institute, Harvard Medical School, Boston, MA 02115, USA. 3 Institute for Genetics, University of Cologne, 50674 Cologne, Germany. 4 Systems Biology of Gene Regulatory Elements, Max Delbrück Center for Molecular Medicine in the Helmholtz Association Berlin, 13125 Berlin, Germany. 5 Department of Medical Oncology, Dana-Farber Cancer Institute, Harvard Medical School, Boston, MA 02215, USA. 6 Adaptive Immunity and Lymphoma, German Cancer Research Center / National Center for Tumor Diseases Heidelberg, 69120 Heidelberg, Germany. 7 Berlin Institute of Health, 10117 Berlin, Germany. *These authors contributed equally to this work. †Corresponding author. Email: [email protected] (R.G.); [email protected] (K.R.) ‡Present address: AbbVie Bioresearch Center, Worcester, MA 01605, USA. §Present address: Pandion Therapeutics, Cambridge, MA 02139, USA. #Present address: Immunology Group, Bioprocessing Technology Institute, Agency for Science, Technology and Research, 138673 Singapore. on March 14, 2020 http://science.sciencemag.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

IMMUNOLOGY
BCR-dependent lineage plasticityin mature B cellsRobin Graf1*†, Jane Seagal2*‡, Kevin L. Otipoby2*§, Kong-Peng Lam3#, Salah Ayoub4,Baochun Zhang2,5, Sandrine Sander1,6, Van Trung Chu1,7, Klaus Rajewsky1,2,3†
B2 cells engage in classical antibody responses, whereas B1 cells are considered carriers ofinnate immunity, biased toward recognizing epitopes present on the surfaces of commonpathogens and self antigens.To explore the role of B cell antigen receptor (BCR) specificity indriving B1 cell differentiation, we developed a transgenic system allowing us to change BCRspecificity in B cells in an inducible and programmedmanner.MatureB2 cells differentiated intobona fide B1 cells upon acquisition of a B1 cell–typical self-reactive BCR through a phase ofproliferative expansion.Thus, B2 cells have B1 cell differentiation potential in addition to theirclassical capacity to differentiate into memory and plasma cells, and B1 differentiation can beinstructed by BCR-mediated self-reactivity, in the absence of B1-lineage precommitment.
Bcells are the carriers of humoral immunity.On the basis of their phenotype and func-tional properties, mature B cells can besubdivided into B1 and B2 cells, with theformer further divided into the CD5+ B1a
and CD5− B1b subsets. B2 cells form follicles inthe spleen and lymph nodes and are responsiblefor generating specific antibody responses againstforeign antigens, typically involving T cell–dependent affinity maturation and somatichypermutation of the B cell antigen receptor(BCR) (1). In contrast, B1 cells are predominantlyfound in peritoneal and pleural cavities andproduce natural antibodies, providing a “firstline of defense” against common bacterialpathogens and contributing to the clearanceof apoptotic cells and oxidized lipids (2, 3).Therefore, B1 cells are thought to performinnate-like functions in the immune system,predominantly expressing evolutionarily selectedBCRs with low levels of somatic mutation andjunctional diversity (4–6).Despite major advances in the field, the origin
of B1 cells remains controversial. The “lineage
model” posits that B1 and B2 cells represent sep-arate lineages that arise from different pro-genitor populations, committed to either lineageprior to BCR expression. In the case of B1 cells,this possibly even occurs prior to hematopoieticstem cell differentiation (7–10). As recently re-viewed in detail, definitive evidence for the line-age model is still lacking, with both putativeBCR-negative B1 progenitors and exclusive pro-genitor commitment to B2 differentiation beingcontroversial (11, 12). In the “selection model,”B cell progenitors are instructed by BCR speci-ficity to differentiate into B1 cells, depending onBCR-mediated recognition of common bacterialand certain self antigens (2, 13). Although strongevidence indeed indicates that certain BCR spec-ificities can exclusively drive B1 cell differentia-tion, this does not rule out a possible commitmentof progenitor cells to either lineage prior to BCRexpression (14). However, as demonstrated below,the existence of B1 and B2 cell–typical BCRs canbe exploited to test lineage commitment and itscontrol by BCR specificity in mature B cells.To address this issue, we generated a genetic
system that would allow a BCR-dependent inter-conversion of mature B1 and B2 cells. For thispurpose, we targeted two prearranged immuno-globulin heavy (IgH) chain variable (V) regiongene segments—VH12 and B1-8, respectively—in a head-to-head orientation and flanked byinverted loxP sites into the JH region of the IgHlocus. Cre-mediated recombination results in in-version of this VH12B1-8
fli cassette, leading to asituation in which some cells express B1-8 andothers VH12 IgH chains. In combination withVk4 Ig light chains, the VH12 and B1-8 IgH chainsform B1 and B2 cell–typical BCRs, respectively(14, 15). VH12B1-8
fli mice (hereafter termed B2mice) and B1-8VH12
fli mice (hereafter termedB1 mice) should predominantly develop B cellswith either B1-8/Vk4 or VH12/Vk4 BCRs (Fig. 1A).In agreement with previous work, the knocked-in BCRs allowed the developing cells to circum-vent the pro/pre–B cell stage (fig. S1, A to E) (15).When the B cells in these animals were stained
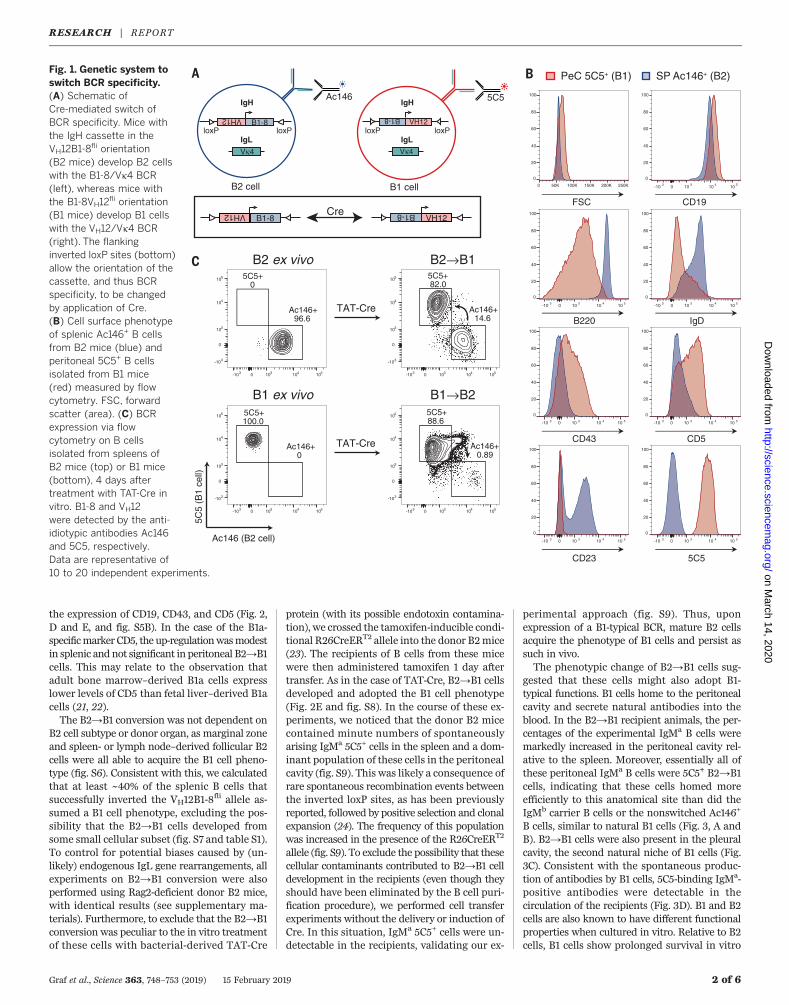
with the BCR-specific anti-idiotypic antibodiesAc146 and 5C5, respectively (14, 16), essentiallyall splenic B cells of B2 mice were Ac146+ and ofthe B2 cell phenotype, characterized by their smallsize and a CD19+, B220+, IgDhi, CD5−, CD23+/lo,and CD43− surface phenotype. In contrast, B cellsof B1 mice were 5C5+ and acquired the B1 cellphenotype, characterized by large cell size anda CD19hi, B220lo, IgDlo, CD5+, CD23−, and CD43+
surface phenotype (Fig. 1B). This is consistentwith the natural expression of this BCR that isspecific for phosphatidylcholine (PtC) and ex-clusively expressed on B1 cells of wild-type mice(13) (fig. S1D). PtC is a commonmembrane phos-pholipid and is exposed on senescent red bloodcells, which suggests that VH12/Vk4 antibodiesare involved in the clearance of these cells (17, 18).When splenic B cells of B1 and B2 mice wereisolated and transduced with TAT-Cre in vitro,approximately 10% of these cells inverted theIgH insertion cassette (fig. S1G). This led to de-tectable populations of cells that switched fromthe expression of the original BCR to that of thenew BCR, thereby confirming the functionalityof the transgenic system (Fig. 1C and fig. S1F).For simplicity, we refer to B1-8VH12
fli– andVH12B1-8
fli–derived switched B cells as B1→B2and B2→B1 cells, respectively.To test whether a change of BCR specificity
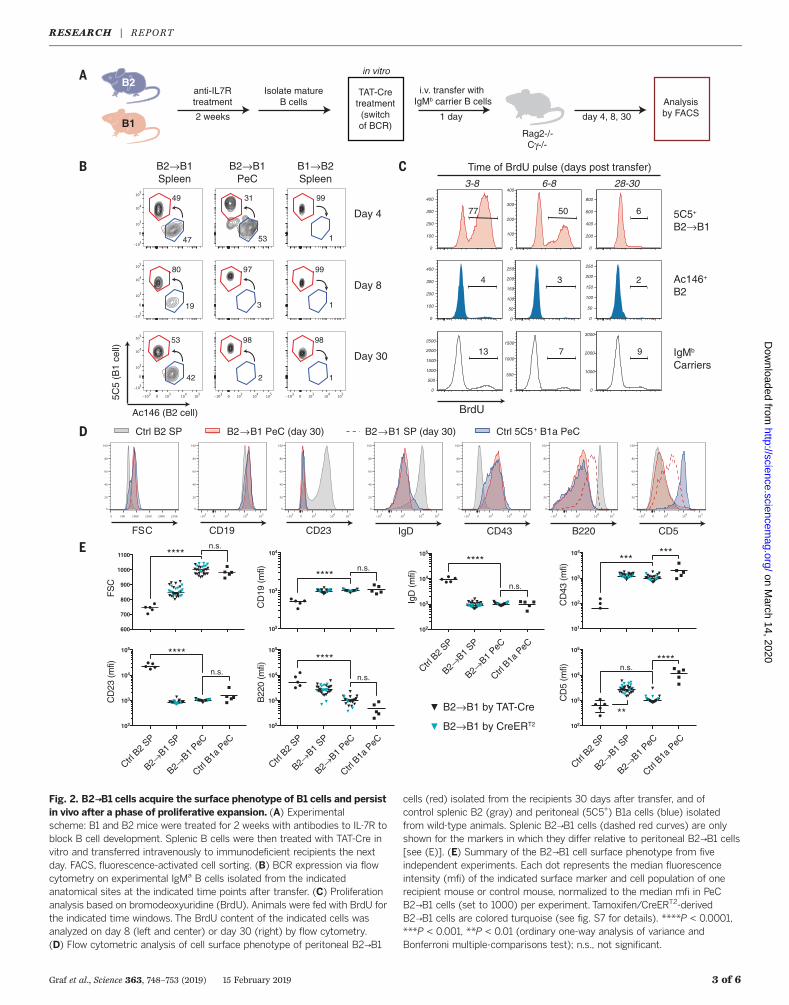
would change the phenotype of fully matureB cells, we treated B1 and B2 mice for 2 weekswith antibodies against the interleukin-7 receptor(IL-7R) to block the influx of immature B cellsinto the spleen (fig. S2). Purified splenic B cellsfrom these mice were then transduced with TAT-Cre in vitro. Only very low numbers of B1→B2cells developed. In contrast, B2→B1 cells devel-oped into a dominant population (Fig. 1C andfig. S1F). However, although these cells acquiredsome B1-typical markers, their phenotypic changewas only partial in vitro (fig. S3). We thereforetransferred IgMa experimental (TAT-Cre trans-duced) mature B cells together with IgMb carrierB cells into immunodeficient recipients to studythe consequences of the BCR switch in a morephysiological environment (Fig. 2A) (19). Con-sistent with the in vitro data, only low numbersof B1→B2 cells were recovered from the recipi-ents. These were located predominantly in theperitoneal cavity and had acquired a phenotypeintermediate between B1 and B2 (fig. S4). Incontrast, B2→B1 cells expressing the 5C5 idio-type were readily detectable in the spleen andperitoneal cavity of the recipients 4, 8, and 30days after transfer and became the dominantpopulation among the IgMa experimental cells(Fig. 2B). Contributing to this dominance wasan initial, transient phase of rapid proliferation,similar to the clonal expansion of PtC-specificB1 cells observed in wild-type mice over the first3 to 4 weeks of life (Fig. 2C and fig. S5A) (4, 20).Strikingly, after completion of this expansionphase, the B2→B1 cells became resting cells thatphenotypically were essentially indistinguishablefrom bona fide B1 cells. B2→B1 cells reproduciblyincreased in size, down-regulated B220 and IgD,lost CD23 and CD21 expression, and increased
RESEARCH
Graf et al., Science 363, 748–753 (2019) 15 February 2019 1 of 6
1Immune Regulation and Cancer, Max Delbrück Center forMolecular Medicine in the Helmholtz Association, 13125Berlin, Germany. 2Program in Cellular and MolecularMedicine, Children’s Hospital, and Immune Disease Institute,Harvard Medical School, Boston, MA 02115, USA. 3Institutefor Genetics, University of Cologne, 50674 Cologne,Germany. 4Systems Biology of Gene Regulatory Elements,Max Delbrück Center for Molecular Medicine in theHelmholtz Association Berlin, 13125 Berlin, Germany.5Department of Medical Oncology, Dana-Farber CancerInstitute, Harvard Medical School, Boston, MA 02215, USA.6Adaptive Immunity and Lymphoma, German CancerResearch Center / National Center for Tumor DiseasesHeidelberg, 69120 Heidelberg, Germany. 7Berlin Institute ofHealth, 10117 Berlin, Germany.*These authors contributed equally to this work.†Corresponding author. Email: [email protected](R.G.); [email protected] (K.R.) ‡Present address:AbbVie Bioresearch Center, Worcester, MA 01605, USA.§Present address: Pandion Therapeutics, Cambridge, MA 02139,USA. #Present address: Immunology Group, BioprocessingTechnology Institute, Agency for Science, Technology andResearch, 138673 Singapore.
on March 14, 2020
http://science.sciencem
ag.org/D
ownloaded from
the expression of CD19, CD43, and CD5 (Fig. 2,D and E, and fig. S5B). In the case of the B1a-specificmarker CD5, the up-regulationwasmodestin splenic and not significant in peritoneal B2→B1cells. This may relate to the observation thatadult bone marrow–derived B1a cells expresslower levels of CD5 than fetal liver–derived B1acells (21, 22).The B2→B1 conversion was not dependent on
B2 cell subtype or donor organ, as marginal zoneand spleen- or lymph node–derived follicular B2cells were all able to acquire the B1 cell pheno-type (fig. S6). Consistent with this, we calculatedthat at least ~40% of the splenic B cells thatsuccessfully inverted the VH12B1-8
fli allele as-sumed a B1 cell phenotype, excluding the pos-sibility that the B2→B1 cells developed fromsome small cellular subset (fig. S7 and table S1).To control for potential biases caused by (un-likely) endogenous IgL gene rearrangements, allexperiments on B2→B1 conversion were alsoperformed using Rag2-deficient donor B2 mice,with identical results (see supplementary ma-terials). Furthermore, to exclude that the B2→B1conversion was peculiar to the in vitro treatmentof these cells with bacterial-derived TAT-Cre
protein (with its possible endotoxin contamina-tion), we crossed the tamoxifen-inducible condi-tional R26CreERT2 allele into the donor B2mice(23). The recipients of B cells from these micewere then administered tamoxifen 1 day aftertransfer. As in the case of TAT-Cre, B2→B1 cellsdeveloped and adopted the B1 cell phenotype(Fig. 2E and fig. S8). In the course of these ex-periments, we noticed that the donor B2 micecontained minute numbers of spontaneouslyarising IgMa 5C5+ cells in the spleen and a dom-inant population of these cells in the peritonealcavity (fig. S9). This was likely a consequence ofrare spontaneous recombination events betweenthe inverted loxP sites, as has been previouslyreported, followed by positive selection and clonalexpansion (24). The frequency of this populationwas increased in the presence of the R26CreERT2
allele (fig. S9). To exclude the possibility that thesecellular contaminants contributed to B2→B1 celldevelopment in the recipients (even though theyshould have been eliminated by the B cell puri-fication procedure), we performed cell transferexperiments without the delivery or induction ofCre. In this situation, IgMa 5C5+ cells were un-detectable in the recipients, validating our ex-
perimental approach (fig. S9). Thus, uponexpression of a B1-typical BCR, mature B2 cellsacquire the phenotype of B1 cells and persist assuch in vivo.The phenotypic change of B2→B1 cells sug-
gested that these cells might also adopt B1-typical functions. B1 cells home to the peritonealcavity and secrete natural antibodies into theblood. In the B2→B1 recipient animals, the per-centages of the experimental IgMa B cells weremarkedly increased in the peritoneal cavity rel-ative to the spleen. Moreover, essentially all ofthese peritoneal IgMa B cells were 5C5+ B2→B1cells, indicating that these cells homed moreefficiently to this anatomical site than did theIgMb carrier B cells or the nonswitched Ac146+
B cells, similar to natural B1 cells (Fig. 3, A andB). B2→B1 cells were also present in the pleuralcavity, the second natural niche of B1 cells (Fig.3C). Consistent with the spontaneous produc-tion of antibodies by B1 cells, 5C5-binding IgMa-positive antibodies were detectable in thecirculation of the recipients (Fig. 3D). B1 and B2cells are also known to have different functionalproperties when cultured in vitro. Relative to B2cells, B1 cells show prolonged survival in vitro
Graf et al., Science 363, 748–753 (2019) 15 February 2019 2 of 6
Fig. 1. Genetic system toswitch BCR specificity.(A) Schematic ofCre-mediated switch ofBCR specificity. Mice withthe IgH cassette in theVH12B1-8
fli orientation(B2 mice) develop B2 cellswith the B1-8/Vk4 BCR(left), whereas mice withthe B1-8VH12
fli orientation(B1 mice) develop B1 cellswith the VH12/Vk4 BCR(right). The flankinginverted loxP sites (bottom)allow the orientation of thecassette, and thus BCRspecificity, to be changedby application of Cre.(B) Cell surface phenotypeof splenic Ac146+ B cellsfrom B2 mice (blue) andperitoneal 5C5+ B cellsisolated from B1 mice(red) measured by flowcytometry. FSC, forwardscatter (area). (C) BCRexpression via flowcytometry on B cellsisolated from spleens ofB2 mice (top) or B1 mice(bottom), 4 days aftertreatment with TAT-Cre invitro. B1-8 and VH12were detected by the anti-idiotypic antibodies Ac146and 5C5, respectively.Data are representative of10 to 20 independent experiments.
Cells
Potential Cre
B1 cellB2 cell
VH12
B1-8
B1-8
VH12
VH12
B1-8
Vκ4
B1-8
VH12
Vκ4
5C5Ac146
A
5C5
(B1
cell)
Ac146 (B2 cell)
TAT-Cre
TAT-Cre
B2 ex vivo
B1 ex vivo
B2→B1
B1→B2
B
FSC CD19
B220 IgD
CD43 CD5
CD23 5C5
C
PeC 5C5+ (B1) SP Ac146+ (B2)
IgH IgH
IgL IgLloxP loxP loxP loxP
0 50K 100K 150K 200K 250K
0
20
40
60
80
100
mix1_4425 PeC_017.fcs5C5+23932
0-10 3 10 3 10 4 10 5
0
20
40
60
80
100
0-10 3 10 3 10 4 10 5
0
20
40
60
80
100
0-10 3 10 3 10 4 10 5
0
20
40
60
80
100
0-10 3 10 3 10 4 10 5
0
20
40
60
80
100
0-10 3 10 3 10 4 10 5
0
20
40
60
80
100
0-10 3 10 3 10 4 10 5
0
20
40
60
80
100
0-10 3 10 3 10 4 10 5
0
20
40
60
80
100
5C5+82.0
Ac146+14.6
0-103 103 104 105
0
-103
103
104
105
5C5+88.6
Ac146+0.89
0-103 103 104 105
0
-103
103
104
105
5C5+0
Ac146+96.6
0-103 103 104 105
0
-103
103
104
105
5C5+100.0
Ac146+0
0-103 103 104 105
0
-103
103
104
105
RESEARCH | REPORTon M
arch 14, 2020
http://science.sciencemag.org/
Dow
nloaded from
Graf et al., Science 363, 748–753 (2019) 15 February 2019 3 of 6
A
Day 8
Day 30
Day 4
B2→B1Spleen 3-8 6-8
5C5+
B2→B1
Ac146+
B2
IgMb
Carriers
28-30
0
100
200
300
400
77
0
500
1000
1500
2000
2500
13
0
100
200
300
400
50
0
500
1000
1500
7
0
50
100
150
200
250
3
0
100
200
300
400
4
0
1000
2000
3000
9
0
50
100
150
200
250
2
0
200
400
600
800
6
BrdU
Time of BrdU pulse (days post transfer)
Ctrl 5C5+ B1a PeCCtrl B2 SP B2→B1 PeC (day 30)
anti-IL7Rtreatment
Isolate matureB cells
TAT-Cretreatment(switchof BCR)
i.v. transfer with IgMb carrier B cells
Rag2-/-Cγ-/-
Analysisby FACS2 weeks 1 day day 4, 8, 30
B2
B1
in vitro
CB
D
E
5C5
(B1
cell)
Ac146 (B2 cell)
49
47
99
1
99
1
80
19
53
42
98
1
31
53
97
3
98
2
B1→B2Spleen
B2→B1PeC
FSC IgDCD19 CD43 B220CD23 CD5
B2→B1 SP (day 30)
600
700
800
900
1000
1100
FS
C
102
103
104
105
IgD
(m
fi)
101
102
103
104
CD
43 (
mfi)
102
103
104
105
B22
0 (m
fi)
102
103
104
105
CD
23 (
mfi)
102
103
104
CD
19 (
mfi)
102
103
104
105
CD
5 (m
fi)
Ctrl B
2 SP
B2→B1
SP
B2→B1
PeC
Ctrl B
1a P
eC
Ctrl B
2 SP
B2→B1
SP
B2→B1
PeC
Ctrl B
1a P
eC
Ctrl B
2 SP
B2→B1
SP
B2→B1
PeC
Ctrl B
1a P
eC
Ctrl B
2 SP
B2→B1
SP
B2→B1
PeC
Ctrl B
1a P
eC
B2→B1 by TAT-Cre
B2→B1 by CreERT2
****
****n.s.
****
n.s.
***
****
n.s.
****
n.s.
****n.s.
n.s.***
**
Fig. 2. B2→B1 cells acquire the surface phenotype of B1 cells and persistin vivo after a phase of proliferative expansion. (A) Experimentalscheme: B1 and B2 mice were treated for 2 weeks with antibodies to IL-7R toblock B cell development. Splenic B cells were then treated with TAT-Cre invitro and transferred intravenously to immunodeficient recipients the nextday. FACS, fluorescence-activated cell sorting. (B) BCR expression via flowcytometry on experimental IgMa B cells isolated from the indicatedanatomical sites at the indicated time points after transfer. (C) Proliferationanalysis based on bromodeoxyuridine (BrdU). Animals were fed with BrdU forthe indicated time windows. The BrdU content of the indicated cells wasanalyzed on day 8 (left and center) or day 30 (right) by flow cytometry.(D) Flow cytometric analysis of cell surface phenotype of peritoneal B2→B1
cells (red) isolated from the recipients 30 days after transfer, and ofcontrol splenic B2 (gray) and peritoneal (5C5+) B1a cells (blue) isolatedfrom wild-type animals. Splenic B2→B1 cells (dashed red curves) are onlyshown for the markers in which they differ relative to peritoneal B2→B1 cells[see (E)]. (E) Summary of the B2→B1 cell surface phenotype from fiveindependent experiments. Each dot represents the median fluorescenceintensity (mfi) of the indicated surface marker and cell population of onerecipient mouse or control mouse, normalized to the median mfi in PeCB2→B1 cells (set to 1000) per experiment. Tamoxifen/CreERT2-derivedB2→B1 cells are colored turquoise (see fig. S7 for details). ****P < 0.0001,***P < 0.001, **P < 0.01 (ordinary one-way analysis of variance andBonferroni multiple-comparisons test); n.s., not significant.
RESEARCH | REPORTon M
arch 14, 2020
http://science.sciencemag.org/
Dow
nloaded from
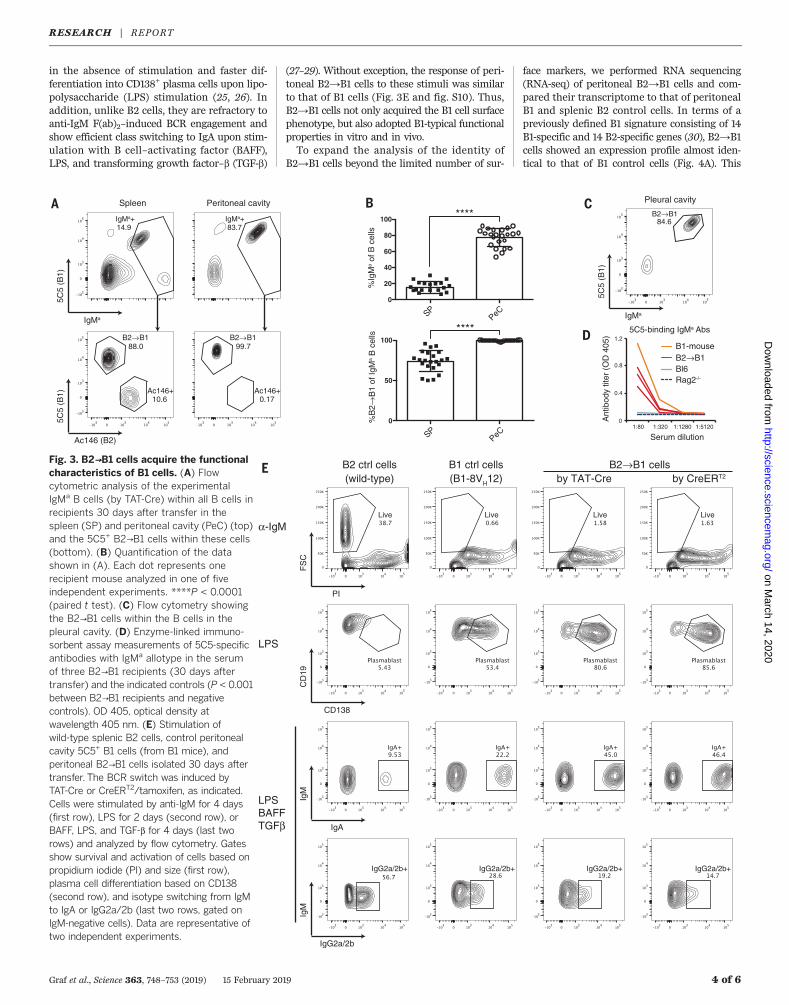
in the absence of stimulation and faster dif-ferentiation into CD138+ plasma cells upon lipo-polysaccharide (LPS) stimulation (25, 26). Inaddition, unlike B2 cells, they are refractory toanti-IgM F(ab)2–induced BCR engagement andshow efficient class switching to IgA upon stim-ulation with B cell–activating factor (BAFF),LPS, and transforming growth factor–b (TGF-b)
(27–29). Without exception, the response of peri-toneal B2→B1 cells to these stimuli was similarto that of B1 cells (Fig. 3E and fig. S10). Thus,B2→B1 cells not only acquired the B1 cell surfacephenotype, but also adopted B1-typical functionalproperties in vitro and in vivo.To expand the analysis of the identity of
B2→B1 cells beyond the limited number of sur-
face markers, we performed RNA sequencing(RNA-seq) of peritoneal B2→B1 cells and com-pared their transcriptome to that of peritonealB1 and splenic B2 control cells. In terms of apreviously defined B1 signature consisting of 14B1-specific and 14 B2-specific genes (30), B2→B1cells showed an expression profile almost iden-tical to that of B1 control cells (Fig. 4A). This
Graf et al., Science 363, 748–753 (2019) 15 February 2019 4 of 6
E
α-IgM
LPS
LPSBAFFTGFβ
FS
C
PI
CD
19
CD138
IgM
IgA
IgM
IgG2a/2b
B2 ctrl cells(wild-type)
B1 ctrl cells(B1-8VH12)
B2→B1 cells by TAT-Cre by CreERT2
A
5C5
(B1)
Ac146 (B2)
5C5
(B1)
IgMa
Spleen Peritoneal cavity B
%B
2→B
1 of
IgM
a B
cel
ls%
IgM
a of
B c
ells
5C5
(B1)
IgMa
C Pleural cavity
D
84.6
B2→B1 B2→B1
B2→B1
SPPeC
IgMa+ IgMa+14.9 83.7
88.0 99.7
Ac146+10.6
Ac146+0.17
Live Live Live Live
IgG2a/2b+ IgG2a/2b+ IgG2a/2b+ IgG2a/2b+
****
0
20
40
60
80
100
0
50
100
SPPeC
****
1:80 1:320 1:1280 1:5120 0
0.4
0.8
1.2 B1-mouseB2→B1 Bl6Rag2-/-
Ant
ibod
y tit
er (
OD
405
)
Serum dilution
5C5-binding IgMa Abs
Fig. 3. B2→B1 cells acquire the functionalcharacteristics of B1 cells. (A) Flowcytometric analysis of the experimentalIgMa B cells (by TAT-Cre) within all B cells inrecipients 30 days after transfer in thespleen (SP) and peritoneal cavity (PeC) (top)and the 5C5+ B2→B1 cells within these cells(bottom). (B) Quantification of the datashown in (A). Each dot represents onerecipient mouse analyzed in one of fiveindependent experiments. ****P < 0.0001(paired t test). (C) Flow cytometry showingthe B2→B1 cells within the B cells in thepleural cavity. (D) Enzyme-linked immuno-sorbent assay measurements of 5C5-specificantibodies with IgMa allotype in the serumof three B2→B1 recipients (30 days aftertransfer) and the indicated controls (P < 0.001between B2→B1 recipients and negativecontrols). OD 405, optical density atwavelength 405 nm. (E) Stimulation ofwild-type splenic B2 cells, control peritonealcavity 5C5+ B1 cells (from B1 mice), andperitoneal B2→B1 cells isolated 30 days aftertransfer. The BCR switch was induced byTAT-Cre or CreERT2/tamoxifen, as indicated.Cells were stimulated by anti-IgM for 4 days(first row), LPS for 2 days (second row), orBAFF, LPS, and TGF-b for 4 days (last tworows) and analyzed by flow cytometry. Gatesshow survival and activation of cells based onpropidium iodide (PI) and size (first row),plasma cell differentiation based on CD138(second row), and isotype switching from IgMto IgA or IgG2a/2b (last two rows, gated onIgM-negative cells). Data are representative oftwo independent experiments.
RESEARCH | REPORTon M
arch 14, 2020
http://science.sciencemag.org/
Dow
nloaded from
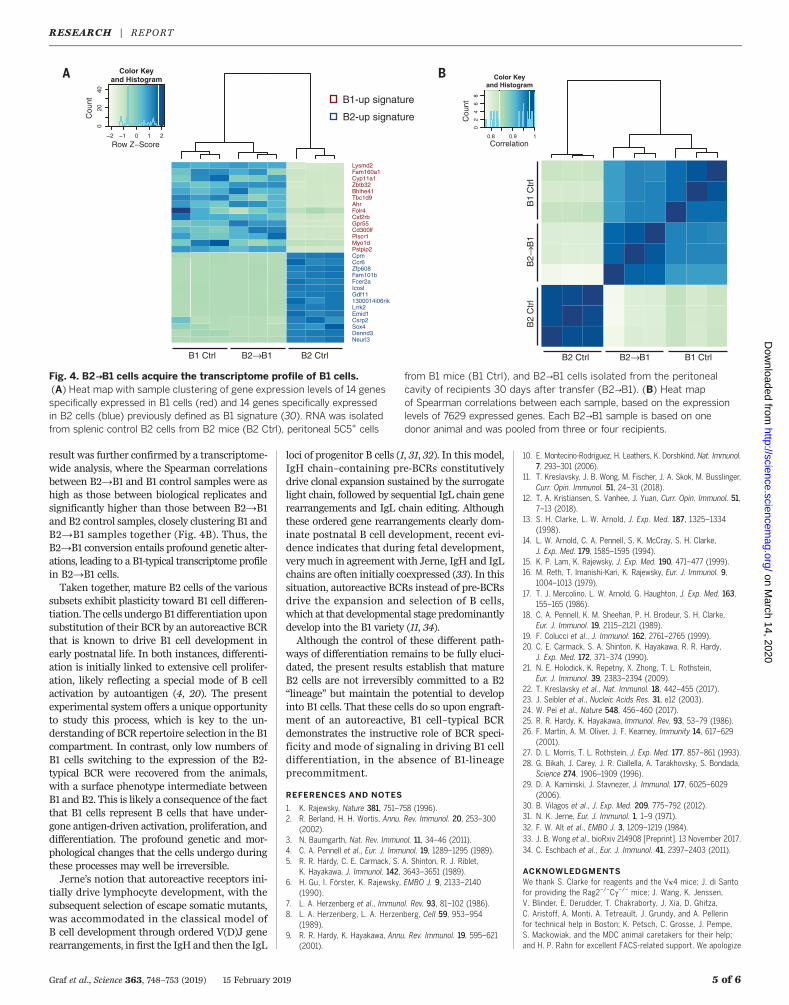
result was further confirmed by a transcriptome-wide analysis, where the Spearman correlationsbetween B2→B1 and B1 control samples were ashigh as those between biological replicates andsignificantly higher than those between B2→B1and B2 control samples, closely clustering B1 andB2→B1 samples together (Fig. 4B). Thus, theB2→B1 conversion entails profound genetic alter-ations, leading to a B1-typical transcriptome profilein B2→B1 cells.Taken together, mature B2 cells of the various
subsets exhibit plasticity toward B1 cell differen-tiation. The cells undergo B1 differentiation uponsubstitution of their BCR by an autoreactive BCRthat is known to drive B1 cell development inearly postnatal life. In both instances, differenti-ation is initially linked to extensive cell prolifer-ation, likely reflecting a special mode of B cellactivation by autoantigen (4, 20). The presentexperimental system offers a unique opportunityto study this process, which is key to the un-derstanding of BCR repertoire selection in the B1compartment. In contrast, only low numbers ofB1 cells switching to the expression of the B2-typical BCR were recovered from the animals,with a surface phenotype intermediate betweenB1 and B2. This is likely a consequence of the factthat B1 cells represent B cells that have under-gone antigen-driven activation, proliferation, anddifferentiation. The profound genetic and mor-phological changes that the cells undergo duringthese processes may well be irreversible.Jerne’s notion that autoreactive receptors ini-
tially drive lymphocyte development, with thesubsequent selection of escape somatic mutants,was accommodated in the classical model ofB cell development through ordered V(D)J generearrangements, in first the IgH and then the IgL
loci of progenitor B cells (1, 31, 32). In this model,IgH chain–containing pre-BCRs constitutivelydrive clonal expansion sustained by the surrogatelight chain, followed by sequential IgL chain generearrangements and IgL chain editing. Althoughthese ordered gene rearrangements clearly dom-inate postnatal B cell development, recent evi-dence indicates that during fetal development,verymuch in agreement with Jerne, IgH and IgLchains are often initially coexpressed (33). In thissituation, autoreactive BCRs instead of pre-BCRsdrive the expansion and selection of B cells,which at that developmental stage predominantlydevelop into the B1 variety (11, 34).Although the control of these different path-
ways of differentiation remains to be fully eluci-dated, the present results establish that matureB2 cells are not irreversibly committed to a B2“lineage” but maintain the potential to developinto B1 cells. That these cells do so upon engraft-ment of an autoreactive, B1 cell–typical BCRdemonstrates the instructive role of BCR speci-ficity and mode of signaling in driving B1 celldifferentiation, in the absence of B1-lineageprecommitment.
REFERENCES AND NOTES
1. K. Rajewsky, Nature 381, 751–758 (1996).2. R. Berland, H. H. Wortis, Annu. Rev. Immunol. 20, 253–300
(2002).3. N. Baumgarth, Nat. Rev. Immunol. 11, 34–46 (2011).4. C. A. Pennell et al., Eur. J. Immunol. 19, 1289–1295 (1989).5. R. R. Hardy, C. E. Carmack, S. A. Shinton, R. J. Riblet,
K. Hayakawa, J. Immunol. 142, 3643–3651 (1989).6. H. Gu, I. Förster, K. Rajewsky, EMBO J. 9, 2133–2140
(1990).7. L. A. Herzenberg et al., Immunol. Rev. 93, 81–102 (1986).8. L. A. Herzenberg, L. A. Herzenberg, Cell 59, 953–954
(1989).9. R. R. Hardy, K. Hayakawa, Annu. Rev. Immunol. 19, 595–621
(2001).
10. E. Montecino-Rodriguez, H. Leathers, K. Dorshkind, Nat. Immunol.7, 293–301 (2006).
11. T. Kreslavsky, J. B. Wong, M. Fischer, J. A. Skok, M. Busslinger,Curr. Opin. Immunol. 51, 24–31 (2018).
12. T. A. Kristiansen, S. Vanhee, J. Yuan, Curr. Opin. Immunol. 51,7–13 (2018).
13. S. H. Clarke, L. W. Arnold, J. Exp. Med. 187, 1325–1334(1998).
14. L. W. Arnold, C. A. Pennell, S. K. McCray, S. H. Clarke,J. Exp. Med. 179, 1585–1595 (1994).
15. K. P. Lam, K. Rajewsky, J. Exp. Med. 190, 471–477 (1999).16. M. Reth, T. Imanishi-Kari, K. Rajewsky, Eur. J. Immunol. 9,
1004–1013 (1979).17. T. J. Mercolino, L. W. Arnold, G. Haughton, J. Exp. Med. 163,
155–165 (1986).18. C. A. Pennell, K. M. Sheehan, P. H. Brodeur, S. H. Clarke,
Eur. J. Immunol. 19, 2115–2121 (1989).19. F. Colucci et al., J. Immunol. 162, 2761–2765 (1999).20. C. E. Carmack, S. A. Shinton, K. Hayakawa, R. R. Hardy,
J. Exp. Med. 172, 371–374 (1990).21. N. E. Holodick, K. Repetny, X. Zhong, T. L. Rothstein,
Eur. J. Immunol. 39, 2383–2394 (2009).22. T. Kreslavsky et al., Nat. Immunol. 18, 442–455 (2017).23. J. Seibler et al., Nucleic Acids Res. 31, e12 (2003).24. W. Pei et al., Nature 548, 456–460 (2017).25. R. R. Hardy, K. Hayakawa, Immunol. Rev. 93, 53–79 (1986).26. F. Martin, A. M. Oliver, J. F. Kearney, Immunity 14, 617–629
(2001).27. D. L. Morris, T. L. Rothstein, J. Exp. Med. 177, 857–861 (1993).28. G. Bikah, J. Carey, J. R. Ciallella, A. Tarakhovsky, S. Bondada,
Science 274, 1906–1909 (1996).29. D. A. Kaminski, J. Stavnezer, J. Immunol. 177, 6025–6029
(2006).30. B. Vilagos et al., J. Exp. Med. 209, 775–792 (2012).31. N. K. Jerne, Eur. J. Immunol. 1, 1–9 (1971).32. F. W. Alt et al., EMBO J. 3, 1209–1219 (1984).33. J. B. Wong et al., bioRxiv 214908 [Preprint]. 13 November 2017.34. C. Eschbach et al., Eur. J. Immunol. 41, 2397–2403 (2011).
ACKNOWLEDGMENTSWe thank S. Clarke for reagents and the Vk4 mice; J. di Santofor providing the Rag2−/−Cg−/− mice; J. Wang, K. Jenssen,V. Blinder, E. Derudder, T. Chakraborty, J. Xia, D. Ghitza,C. Aristoff, A. Monti, A. Tetreault, J. Grundy, and A. Pellerinfor technical help in Boston; K. Petsch, C. Grosse, J. Pempe,S. Mackowiak, and the MDC animal caretakers for their help;and H. P. Rahn for excellent FACS-related support. We apologize
Graf et al., Science 363, 748–753 (2019) 15 February 2019 5 of 6
A
Neurl3Dennd3Sox4Csrp2Emid1Lrrk21300014i06rikGdf11IcoslFcer2aFam101bZfp608Ccr6CpmPstpip2Myo1dPlscr1Cd300lfGpr55Csf2rbFolr4AhrTbc1d9Bhlhe41Zbtb32Cyp11a1Fam160a1Lysmd2
−2 −1 0 1 2
Row Z−Score
020
40
Color Keyand Histogram
Cou
nt
0.8 0.9 1
Correlation
02
46
8
Color Keyand Histogram
Cou
nt
B
B1-up signature
B2-up signature
B1 Ctrl B2→B1 B2 Ctrl B2 Ctrl B2→B1 B1 Ctrl
B1
Ctr
l B
2→B
1 B
2 C
trl
Fig. 4. B2→B1 cells acquire the transcriptome profile of B1 cells.(A) Heat map with sample clustering of gene expression levels of 14 genesspecifically expressed in B1 cells (red) and 14 genes specifically expressedin B2 cells (blue) previously defined as B1 signature (30). RNA was isolatedfrom splenic control B2 cells from B2 mice (B2 Ctrl), peritoneal 5C5+ cells
from B1 mice (B1 Ctrl), and B2→B1 cells isolated from the peritonealcavity of recipients 30 days after transfer (B2→B1). (B) Heat mapof Spearman correlations between each sample, based on the expressionlevels of 7629 expressed genes. Each B2→B1 sample is based on onedonor animal and was pooled from three or four recipients.
RESEARCH | REPORTon M
arch 14, 2020
http://science.sciencemag.org/
Dow
nloaded from

to colleagues whose work we could not cite because of spacerestrictions. Funding: Supported by NIH grant AI054636 andthe European Research Council (ERC Advanced Grant 268921 toK.R.). Author contributions: J.S. and K.L.O. started theexperimental work at the Immune Disease Institute, HarvardMedical School; R.G. substantially broadened and completedthe study at the MDC; K.-P.L. generated the VH12B1-8
fli mousestrain; S.A. helped to generate the RNA-seq library; B.Z. helpedwith cell transfer experiments; S.S. provided reagents; V.T.C.
provided scientific and experimental advice; R.G. and K.R.wrote the manuscript; and K.R. supervised the project.Competing interests: The authors declare no competingfinancial interest. Data and materials availability: The RNA-seq data are available in the Gene Expression Omnibus (GEO)database (www.ncbi.nlm.nih.gov/geo) under accession numberGSE124827. The materials required to use the transgenicsystem described here will be available through K.R. under amaterial transfer agreement from the MDC.
SUPPLEMENTARY MATERIALS
www.sciencemag.org/content/363/6428/748/suppl/DC1Materials and MethodsFigs. S1 to S10Table S1References (35–40)
22 July 2018; accepted 17 January 201910.1126/science.aau8475
Graf et al., Science 363, 748–753 (2019) 15 February 2019 6 of 6
RESEARCH | REPORTon M
arch 14, 2020
http://science.sciencemag.org/
Dow
nloaded from

BCR-dependent lineage plasticity in mature B cells
and Klaus RajewskyRobin Graf, Jane Seagal, Kevin L. Otipoby, Kong-Peng Lam, Salah Ayoub, Baochun Zhang, Sandrine Sander, Van Trung Chu
DOI: 10.1126/science.aau8475 (6428), 748-753.363Science
, this issue p. 748ScienceBCR was swapped in, in the absence of B1 lineage precommitment.which BCR specificities can be changed. Mature B2 cells differentiated into functional B1 cells when a self-reactive B1
provide support for the selection model using a transgenic system inet al.different B cell antigen receptors (BCRs). Graf by''lineage model,'' wherein both subsets have distinct progenitors, and the ''selection model,'' in which fates are directed
instead produce ''natural'' antibodies as part of the innate immune system. Two models to explain this split exist: theforeign antigens in secondary lymphoid organs. B1 cells, found predominantly in the peritoneal and pleural cavities,
Immunological B cells are generally divided into two major subsets. B2 cells generate specific antibodies againstB1 or B2? The BCR decides
ARTICLE TOOLS http://science.sciencemag.org/content/363/6428/748
MATERIALSSUPPLEMENTARY http://science.sciencemag.org/content/suppl/2019/02/13/363.6428.748.DC1
REFERENCES
http://science.sciencemag.org/content/363/6428/748#BIBLThis article cites 39 articles, 15 of which you can access for free
PERMISSIONS http://www.sciencemag.org/help/reprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAAS.ScienceScience, 1200 New York Avenue NW, Washington, DC 20005. The title (print ISSN 0036-8075; online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science. No claim to original U.S. Government WorksCopyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of
on March 14, 2020
http://science.sciencem
ag.org/D
ownloaded from
Related Documents











