Veterinary Immunology and Immunopathology, 30 ( 1992 ) 385-398 385 Elsevier Science Publishers B.V., Amsterdam Immunologic and genetic characterization of S 180, a cell line of murine origin capable of growing in different inbred strains of mice G. Alfaro a, C. Lomeli a, R. Ocadiz b, V. Ortega b, R. Barrera a, M. Ramirez b and G. Nava a aDepartarnento de Inmunologia, Instituto de Investigaciones Biomedicas, Universidad Nacional Autonoma de Mexico, 04510 Mexico D.F., Mexico bDivision de Investigacion Basica, lnstituto Nacional de Cancerologia, 14000, Mexico D.F., Mexico (Accepted 12 February 1991 ) ABSTRACT Alfaro, G., Lomeli, C., Ocadiz, R., Ortega, V., Barrera, R., Ramirez, M. and Nava, G., 1992. Immu- nologic and genetic characterization of S180, a cell line of murine origin capable of growing in different inbred strains of mice. Vet. Immunol. Immunopathol., 30: 385-398. Some of the immunologic and genetic properties of the cell line S 180 have been examined. These cells grew without restrictions in the peritoneal cavity of different inbred strains of mice and invaria- bly killed the animals. With Northern blots it was demonstrated that S180 cells contained class I mRNAs but failed to transcribe B2m genes. However, under experimental conditions, a protective humoral immune response mediated by cytotoxic antibodies and complement against S 180 cells was obtained through non-H-2 antigens in C57BL/6J mice. ABBREVIATIONS ff2m and B2m, fl2-microglobulin; CTL, cytotoxic T-lymphocytes; MHC, major histocompa- tibility complex; RPMI, Roswell Park Memorial Institute. INTRODUCTION The major histocompatibility complex (H-2) class I genes of the mouse codes for glycoproteins which are expressed on the surface of most cells (Klein et al., 1983; Hood et al., 1983; Flavell et al., 1986). Class I molecules are assembled on the cell membrane as noncovalently linked dimers, composed of a heavy chain of 40-45 kDa and a light chain of 12 kDa. Heavy chain molecules are encoded by genes located on chromosome 17, including the highly polymorphic class I classical transplantation antigens H-2K, H-2D and © 1992 Elsevier Science Publishers B.V. All rights reserved 0165-2427/92/$05.00

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Veterinary Immunology and Immunopathology, 30 ( 1992 ) 385-398 385 Elsevier Science Publishers B.V., Amsterdam
Immunologic and genetic characterization of S 180, a cell line of murine origin capable of growing in different inbred strains of mice
G. Alfaro a, C. Lomeli a, R. Ocadiz b, V. Ortega b, R. Barrera a, M. Ramirez b and G. N a v a a
aDepartarnento de Inmunologia, Instituto de Investigaciones Biomedicas, Universidad Nacional Autonoma de Mexico, 04510 Mexico D.F., Mexico
bDivision de Investigacion Basica, lnstituto Nacional de Cancerologia, 14000, Mexico D.F., Mexico
(Accepted 12 February 1991 )
ABSTRACT
Alfaro, G., Lomeli, C., Ocadiz, R., Ortega, V., Barrera, R., Ramirez, M. and Nava, G., 1992. Immu- nologic and genetic characterization of S180, a cell line of murine origin capable of growing in different inbred strains of mice. Vet. Immunol. Immunopathol., 30: 385-398.
Some of the immunologic and genetic properties of the cell line S 180 have been examined. These cells grew without restrictions in the peritoneal cavity of different inbred strains of mice and invaria- bly killed the animals. With Northern blots it was demonstrated that S180 cells contained class I mRNAs but failed to transcribe B2m genes. However, under experimental conditions, a protective humoral immune response mediated by cytotoxic antibodies and complement against S 180 cells was obtained through non-H-2 antigens in C57BL/6J mice.
ABBREVIATIONS
ff2m and B2m, fl2-microglobulin; CTL, cytotoxic T-lymphocytes; MHC, major histocompa- tibility complex; RPMI, Roswell Park Memorial Institute.
INTRODUCTION
The major histocompatibility complex (H-2) class I genes of the mouse codes for glycoproteins which are expressed on the surface of most cells (Klein et al., 1983; Hood et al., 1983; Flavell et al., 1986). Class I molecules are assembled on the cell membrane as noncovalently linked dimers, composed of a heavy chain of 40-45 kDa and a light chain of 12 kDa. Heavy chain molecules are encoded by genes located on chromosome 17, including the highly polymorphic class I classical transplantation antigens H-2K, H-2D and
© 1992 Elsevier Science Publishers B.V. All rights reserved 0165-2427/92/$05.00

386 G. ALFAROETAL.
H-2L, and the less polymorphic but genetically related Qa and T 1 a determi- nants. The light chain corresponds to fl2m and is encoded by genes which map within chromosome 2. The variable end of class I proteins can associate with antigenic determinants expressed by virally infected cells or by tumour cells and lead to its recognition by specific receptors located on the cell surface of CTLs (Zinkernagel and Doherty, 1979; Allison and Lanier, 1987).
It has been shown that the heavy chain of class I molecules cannot be ex- pressed on the cellular membrane in absence offl2m (Festenstein and Schmidt, 1981; Tanaka et al., 1988 ). The majority of such cell lines have mutations in B2m genes and are phenotypically H-2-deficient (Hyman and Stallings, 1975; Parnes and Seidman, 1982; Doyle et al., 1985; Parnes et al., 1986; Potter et al., 1987). Furthermore, loss offf2m expression correlated with histological and metastatic properties of a group of human skin tumours (Turbitt and Mackie, 1981; Lampson et al., 1983; Holden et al., 1984).
Cell lines with normal B2m genes but lacking one or more class I H-2 anti- genic determinants on the cell membrane have been described by other inves- tigators (Isakov et al., 1983; Katzav et al., 1983; Eisenbach et al., 1984; Wal- lich et al., 1985 ). Cancer cells may undergo additional phenotypic changes or secret diffusible products which prevent an efficient recognition by the im- mune system and facilitate tumour progression and the production of metas- tasis (Celis et al., 1979; North, 1984; Tryggvason et al., 1987; Tanaka et al., 1988).
S 180 cells were selected because of their capacity to grow without restric- tions in the peritoneal cavity of different inbred strains of mice. We demon- strated that escape to immune surveillance mechanisms by S 180 cells corre- lated with the lack of transcription of B2m genes. In addition, we showed that this cell line expressed non-H-2 determinants on the cell membrane which could induce an effective humoral immune response.
MATERIALS AND METHODS
Mice
Eight- to twelve-week-old mice of the following inbred strains were used: BALB/cAnN (H-2a); C.B6-H2 and C57BL/6J (H-2b); C.C3-H2 and C3HeB/ FeJ (H-2 k) and A/J (H-2a). All mice were bred in the vivarium of the Insti- tuto de Investigaciones Biomedicas.
Tumour cell lines
The S 180 cell line was isolated from a randomly bred female Swiss mouse and obtained from the ATCC (Dunham and Stewart, 1953 ). Other cell lines were YAC-1 (H-2 a) (Feny/5 et al., 1968), EL4 (H-2 b) (Gorer, 1950), B16

ff2M EXPRESSION AND GROWTH ACROSS ALLOGENEIC GROWTH 387
(H-2 b) (Fidler et al., 1978), L5178Y (H-2 d) (Fisher, 1958) and its H-2- deficient derivative LR.4 (Nava, manuscript in preparation) and BW5147 (H-2 k) (Ralph, 1973).
Some of the cell lines used were maintained in vivo growing in the perito- neal cavity of C57BL/6J (S180 and EL4) and C.C3-H2 (LR.4) mice. Cul- ture medium used for propagating cell lines in vitro (37 °C; humidified 5% CO2-air) was RPMI 1640 supplemented with 10% heat-inactivated fetal calf serum, 1 mM L-glutamine, and 1% of the following: non-essential amino acids, sodium pyruvate, and a combination of antibiotics (all these products were obtained from GIBCO Laboratories, Grand Island, NY).
Preparation of anti-S180 antiserum
Groups of C57BL/6J mice were injected intraperitoneally (i.p) four times at weekly intervals with 5 × 106 S180 cells treated with 25 #g ml-1 of Mito- mycin C (Sigma) (Benjamini et al., 1977). Animals were bled 5 days after the last immunization and the serum was collected and pooled. The specific- ity of the antiserum was tested against target and control cells by immunofluorescence.
Immunofluorescence assay
Indirect immunofluorescence was performed by incubating cell cultures with appropriate dilutions of immune or normal mouse serum for 1 h at 4 ° C. Cell suspensions were then washed with PBS containing 3% bovine serum albu- min and 0.02% sodium azide. Fluorescinated goat anti-mouse Ig (100/A; Southern Biotechnology Associates, Birmingham, AL) diluted 1/20 in the same buffer was added and the reaction mixture was incubated for another 30 min at 4 ° C. Cells were washed twice with PBS and the percentage of re- acting cells estimated using a Zeiss K7 fluorescence microscope equipped with epifluorescence illumination.
Radiolabelling of cells
5180 cells were labelled by incubating 1 × 106 cells in 0.5 ml RPMI 1640 with 200/tCi of 5~Cr (NazCrO4, 350-600 mCi mg-1 51Cr; Amersham, Ayles- bury, Buckinghamshire ) for 1 h under culture conditions. Thereafter cells were washed three times by centrifugation with cold RPMI 1640. Cells were ad- justed to the desired concentration with RPMI 1640.

388 G. ALFAROETAL
Cytotoxicity assay
The presence of cytotoxic antibodies in sera was determined by the 51Cr release assay (Celis et al., 1979). In brief, 51Cr-labelled cells were distributed among test tubes (4 X 104 cells 25/~1-1 per tube ) in an ice-water bath. A given dilution of anti-S 180 antiserum or normal mouse serum (25/zl) were added to each tube and incubated for 15 min. Thereafter, 25/zl of fresh rabbit serum, used as a complement source, were added and the volume of each tube was brought to 100/zl with RPMI 1640. All tubes were incubated for 1 h under culture conditions, followed by the addition of 1.9 ml of cold PBS to each tube. After centrifugation, 1.0 ml of supernatant was removed from each tube and transferred to fresh tubes to be counted. Controls to assess the toxicity of antibody, complement, or med ium were included.
% specific release = experimental re lease- spontaneous release X 100 total re lease-spontaneous release
DNA isolation and Southern blots
High molecular weight DNA was isolated from L5178Y, LR.4 and S180 cell lines by standard procedure (Maniatis et al., 1982 ). For Southern blots, DNAs ( 10 #g) were cleaved with appropriate restriction enzymes under con- ditions recommended by the manufacturer (Amersham), and electrop- horesed on 0.8% agarose gels. The DNA was denatured in situ and transferred to nitrocellulose filters (Schleicher and Schuell, Keene, NH ) according to the procedure described by Southern (1975). Prehybridization was in 25 mM KPO4, 5X SSC ( IX SSC is 0.15 M NaC1, 0.015 M sodium citrate), 5X Den- hardt 's solution ( 1X Denhardt 's is 0.02% bovine serum albumin, 0.02% Fi- coll, 0.02% polyvinylpyrrolidone), 0.5% NaDodSO4, 50 #g ml - l of dena- tured salmon sperm and 50% formamide. Hybridization was in the same solution but supplemented with 1% NaDodSO4 and 10% dextran sulfate. Blots were hybridized for 12 h with 5 × 108 cpm of nick translated 32p-labelled DNA fragments per miliilitre. Washing conditions were: once in 2X SSC, 0.1% NaDodSO4 at room temperature, and three times with 0.1X SSC at 50°C. DNA fragments were isolated from plasmid subclones of genomic or cDNA clones and were electrophoresed through low melting point agarose prior to extraction. Filters were exposed for several days at - 7 0 ° C to X-OMAT K film (Kodak, Guadalajara, Jalisco) with an intensifying screen.
RNA blot analysis
Total RNA was isolated from the cell lines ment ioned in the preceding sec- tion using the guanidinine thiocyanate procedure (Chirwin et al., 1979 ), pel-

ff2M EXPRESSION AND GROWTH ACROSS ALLOGENEIC GROWTH 389
leting the RNA through a CsC1 cushion. Ten micrograms of each RNA were electrophoresed through a 1.5% agarose gel containing 2.2 M formaldehyde (Maniatis et al., 1982 ). The RNA was transferred to nitrocellulose filters ac- cording to a method described by Thomas (1980) and hybridized to nick- translated DNA probes. Hybridization and wash conditions were as de- scribed in the preceding section. Filters were exposed to X-OMAT K film for several days at - 7 0 ° C with an intensifying screen.
DNA probes
A DNA probe containing a 0.66 kb HpalI fragment from a cDNA of the H- 2L a gene (Evans et al., 1982) was obtained from Dr. D. Margulies. A 0.6 kb EcoRI-KpnI fragment of genomic DNA specific for the second exon of the B2m gene was received from Dr. J. Seidman (Parnes and Seidman, 1982 ). A pBR322 derivative containing a 0.70 kb EcoRI fragment from histone H4 gene (pBR-mus-hi-l-H4-Hinfl) was kindly provided by Dr. Seyler-Tuyns (Seyler-Tuyns and Birnstiel, 1981 ).
RESULTS
Growth patterns of $180 in different strains of mice
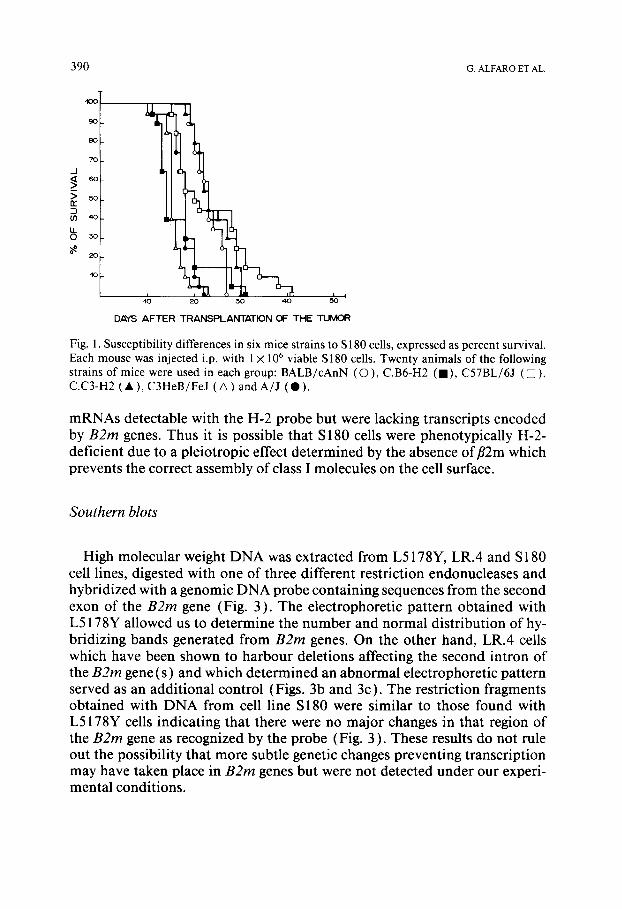
Groups of 20 animals of 6 different strains of mice, BALB/cAnN (H-2d), C.B6-H2 and C57BL/6J (H-2b), C.C3-H2 and C3HeB/FeJ (H-2 k) and A/J (H-2 a) were injected i.p. with l )< l0 6 S180 cells. Fig. l shows that all mice strains were susceptible to the tumour and died within 30 days after the trans- plant. The capacity of S 180 cells to grow in mice of different haplotypes was not determined by the size of the inoculum, since C57BL/6J and BALB/cAnN were equally susceptible to the tumor when they were injected i.p with differ- ent concentrations of cells ( 1 X l0 3 - 1 )< l05) (not shown).
Transcription of B2m and class I genes in S180 cells
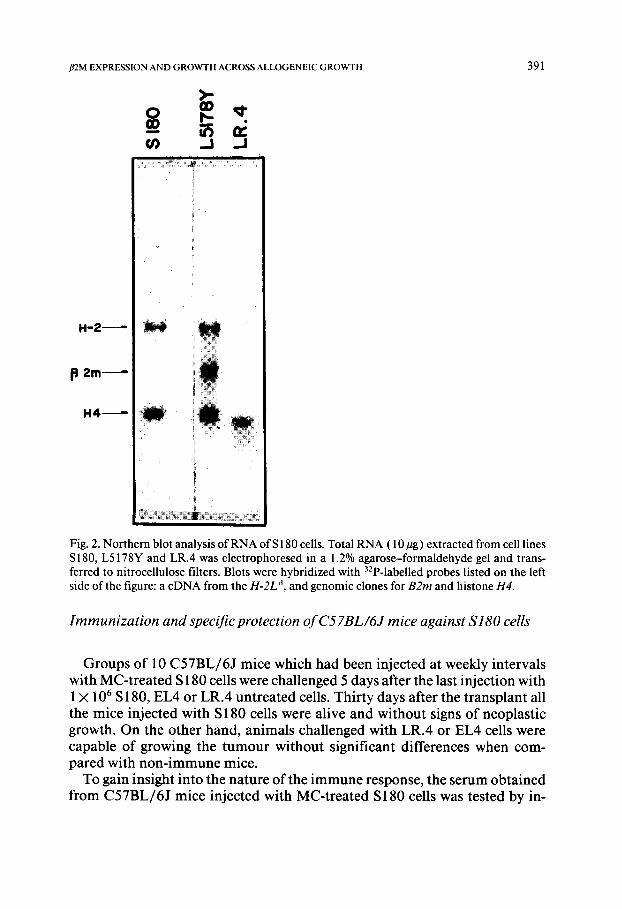
To investigate if the growth properties of S 180 cells were genetically deter- mined at the H-2 level, total RNA was extracted from L5178Y, LR.4 and S 180 cell lines, and analyzed in Northern blots with DNA probes containing sequences specific for the second exon of the B2rn gene and a cDNA from the H-2L d gene (Fig. 2). L5178Y cells, which were used as a positive control, allowed the identification of mRNAs specific for B2m and class I H-2 genes. Total RNA extracted from LR.4 cells, an H-2-deficient and promiscuous mu- tant isolated from L5178Y, did not hybridize with H-2 class I or B2m gene probes, as previously determined (in preparation). The hybridization pat- tern obtained with S180 cells showed that this cell line contained class I
3 9 0 G. ALFARO ET AL.
too
X
10 2o 30 40 5o
DAYS AFTER TRANSPLANTATION OF THE TUMOR
Fig. 1. Susceptibility differences in six mice strains to S 180 cells, expressed as percent survival. Each mouse was injected i.p. with 1 × 106 viable S 180 cells. Twenty animals of the following strains of mice were used in each group: BALB/cAnN (O), C.B6-H2 (IlL C57BL/6J ([]), C.C3-H2 ( • ), C3HeB/FeJ ( A ) and A/J ( • ).
m R N A s detectable with the H-2 probe but were lacking transcripts encoded by B2m genes. Thus it is possible that S 180 cells were phenotypically H-2- deficient due to a pleiotropic effect determined by the absence offf2m which prevents the correct assembly of class I molecules on the cell surface.
Southern blots
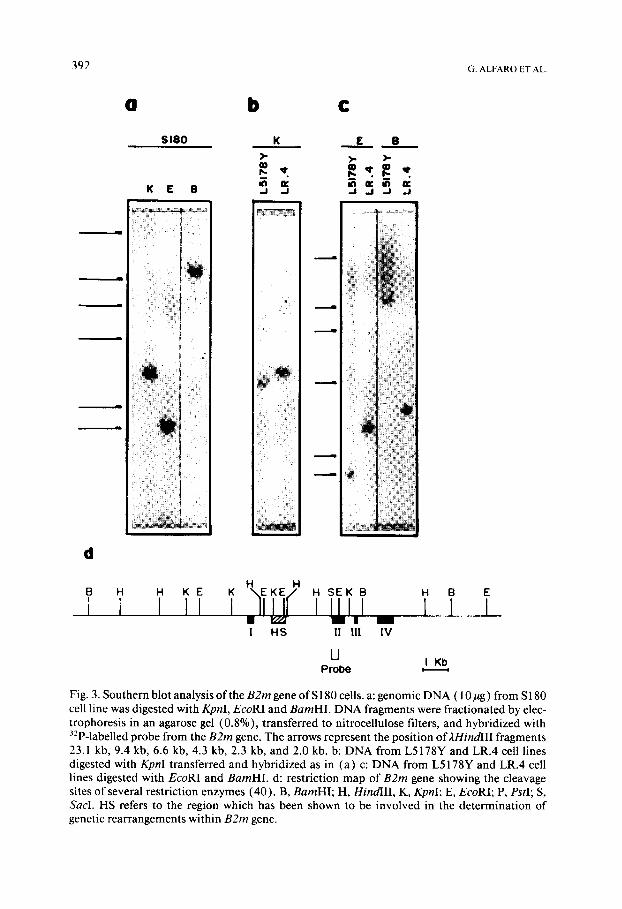
High molecular weight D N A was extracted from L5178Y, LR.4 and S180 cell lines, digested with one of three different restriction endonucleases and hybridized with a genomic D N A probe containing sequences from the second exon of the B2m gene (Fig. 3). The electrophoretic pattern obtained with L5178Y allowed us to determine the number and normal distribution of hy- bridizing bands generated from B2m genes. On the other hand, LR.4 cells which have been shown to harbour deletions affecting the second intron of the B2m gene (s) and which determined an abnormal electrophoretic pattern served as an addit ional control (Figs. 3b and 3c). The restriction fragments obtained with D N A from cell line S180 were similar to those found with L5178Y cells indicating that there were no major changes in that region of the B2m gene as recognized by the probe (Fig. 3 ). These results do not rule out the possibility that more subtle genetic changes preventing transcription may have taken place in B2m genes but were not detected under our experi- mental conditions.
ff2M EXPRESSION A N D G R O W T H ACROSS A L L O G E N E I C G R O W T H 391
>-
i ~ - .
(n .J .J
H-2
2 m - ~ -
H 4 - ~ -
U
i
Fig. 2. Northern blot analysis of RNA of $180 cells. Total RNA (10/tg) extracted from cell lines S180, L5178Y and LR.4 was electrophoresed in a 1.2% agarose-formaldehyde gel and trans- ferred to nitrocellulose filters. Blots were hybridized with 32p-labelled probes listed on the left side of the figure: a cDNA from the H-2L d, and genomic clones for B2m and histone H4.
Immunization and specific protection of C5 7BL/6J mice against $180 cells
Groups of 10 C 5 7 B L / 6 J mice which had been injected at weekly intervals with MC-treated S 180 cells were challenged 5 days after the last injection with 1 X l06 S 180, EL4 or LR.4 untreated cells. Thirty days after the transplant all the mice injected with S 180 cells were alive and without signs of neoplastic growth. On the other hand, animals challenged with LR.4 or EL4 cells were capable of growing the tumour without significant differences when com- pared with non- immune mice.
To gain insight into the nature of the immune response, the serum obtained from C 5 7 B L / 6 J mice injected with MC-treated S180 cells was tested by in-
~ 9 2 G. ALFAROETAL.
G b C
SI80
K E B
I~ qr
J J
~ ~i ~ ~!i
E B
J J J J
d
H H
i 1 I I1 i I1111 I I I I HS II Ul IV
13 I Kb
P r o D e , ,
Fig. 3. Southern blot analysis of the B2m gene of S 180 cells• a: genomic DNA ( 10 ag) from S 180 cell line was digested with KpnI, EcoRI and BamHI. DNA fragments were fractionated by elec- trophoresis in an agarose gel (0.8%), transferred to nitrocellulose filters, and hybridized with 32p-labelled probe from the B2m gene. The arrows represent the position of 2HindlII fragments 23.1 kb, 9.4 kb, 6.6 kb, 4.3 kb, 2.3 kb, and 2.0 kb. b: DNA from L5178Y and LR.4 cell lines digested with KpnI transferred and hybridized as in (a) c: DNA from L5178Y and LR.4 cell lines digested with EcoRI and BamHI. d: restriction map of B2m gene showing the cleavage sites of several restriction enzymes (40). B, BamHI; H, HindlII, K, KpnI; E, EcoRI; P, PstI; S, Sacl. HS refers to the region which has been shown to be involved in the determination of genetic rearrangements within B2m gene.
ff2M EXPRESSION AND GROWTH ACROSS ALLOGENEIC GROWTH 393
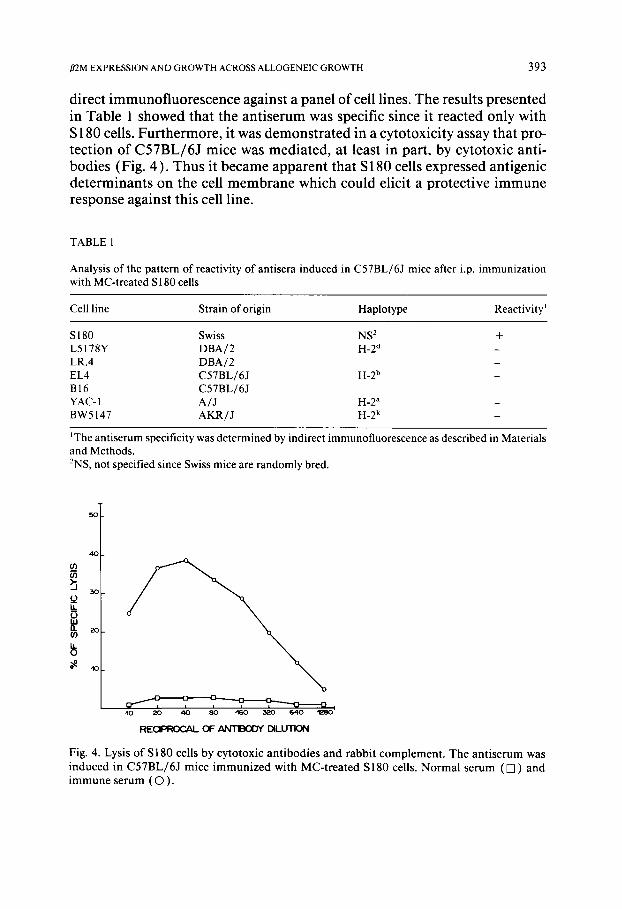
direct immunofluorescence against a panel of cell lines. The results presented in Table 1 showed that the antiserum was specific since it reacted only with S 180 cells. Furthermore, it was demonstrated in a cytotoxicity assay that pro- tection of C57BL/6J mice was mediated, at least in part, by cytotoxic anti- bodies (Fig. 4). Thus it became apparent that S 180 cells expressed antigenic determinants on the cell membrane which could elicit a protective immune response against this cell line.
TABLE 1
Analysis of the pattern of reactivity of antisera induced in C57BL/6J mice after i.p. immunization with MC-treated S 180 cells
Cell line Strain of origin Haplotype Reactivity ~
S 180 Swiss NS 2 + L5178Y DBA/2 H-2 a - LR.4 DBA/2 EL4 C57BL/6J H-2 b - B 16 C57BL/6J YAC-1 A/J H-2 a - BW5147 AKR/J H-2 k -
tThe antiserum specificity was determined by indirect immunofluorescence as described in Materials and Methods. 2NS, not specified since Swiss mice are randomly bred.
50
40
30
20
40
,.,o u o-----_, o
RECUoROCAL OF AN3IBOOY 0(LUTION
_o
,,q n (/)
Fig. 4. Lysis of S180 cells by cytotoxic antibodies and rabbit complement . The antiserum was induced in C57BL/6J mice immunized with MC-treated S180 cells. Normal serum ( [ ] ) and immune serum ( O ) .

394 G. ALFAROETAL
DISCUSSION
S 180 cells can grow without apparent restrictions in the peritoneal cavity of different inbred strains of mice. This property, which was independent of inoculum size, suggested that growth across allogeneic barriers was deter- mined by changes in the expression offl2m or class I determinants as has been shown with other cell lines of murine origin (Tanaka et al., 1988 ).
Total RNA extracted from S 180 cells and analyzed in Northern blots with two different DNA probes showed that this cell line was not transcribing B 2 m genes but expressing mRNAs encoded by class I H-2 genes. However, diges- tion of S180 DNA with three different restriction enzymes and subsequent hybridization with a genomic probe from the second exon of the B 2 m gene gave similar electrophoretic patterns to those obtained with the control cell line L5178Y. It remains possible, however, that B 2 m genes were indeed de- fective due to the presence of small mutat ions which escaped detection under our experimental conditions or because the genetic alterations were located 3' off the second B a m H I site and therefore were not recognized by the DNA probe used in these experiments (Fig. 3d). If mutat ion was the responsible mechanism for the lack of expression of B2rn genes, either both alleles were mutated or only one gene was defective and the other was deleted. However, the possibility cannot be ruled out that B 2 m genes may be repressed in re- sponse to cis or trans acting elements, as has been shown in other experimen- tal systems, (Young and Tilghman, 1984; Carrington et al., 1985; Keeney et al., 1989).
Several lines of evidence indicate that expression offl2m is essential for the correct assembly of class I proteins on the cell membrane. Most cell lines car- rying mutations in genes coding for ff2m are phenotypically H-2-deficient (Hyman and Stallings, 1975; Parnes and Seidman, 1982; Parnes et al., 1986; Potter et al., 1987). However, transfection of the H - 2 D b gene into the R1E cell line, which lacks fl2m, led to the expression of class I H - 2 D b antigenic determinants, although these were not recognized by Db-restricted CTLs or by monoclonal antibodies against the native D b protein product (Allen et al., 1986). Another exception has been described by Potter et al. ( 1984, 1985 ) who isolated a sub-line from EL4 cells expressing H - 2 D b determinants in the absence o f H - 2 I ( ° and fl2m.
MHC class I-defective cell variants have been isolated from the highly met- astatic T10 sarcoma (Katzav et al., 1983, 1984, 1985; Wallich et al., 1985) and 3LL Lewis lung carcinoma cell lines (Isakov et al., 198 l, 1983; Eisenbach et al., 1984). When transplanted into syngeneic animals, the resulting clones showed slower growth rates and often failed to produce metastasis when com- pared to their parental counterparts (Isakov et al., 1983; Eisenbach et al., 1984). In such cases, only one of two allelic forms of the H - 2 D gene were found to be responsible for the phenotype without involvement o f B 2 m genes.

ff2M EXPRESSION AND GROWTH ACROSS ALLOGENEIC GROWTH 395
On the other hand, the highly antigenic products of H-2K loci were primarily involved in the capacity to induce an efficient and specific cellular immune response (Isakov et al., 1983; Katzav et al., 1984, 1985; Wallich et al., 1985). Furthermore, some of the clones which were either H-2-deficient or expressed H2-D b products at low density on the cell membrane were successfully trans- planted across allogeneic barriers (Isakov et al., 1981; Katzav et al., 1984, 1985).
A different situation was described with cell variants derived from EL4 and RBL-5 cells (Ljunggren and K~irre, 1985; K~irre et al., 1986). These investi- gators demonstrated that clones which were either expressing low levels or did not contain class I determinants on the cell membrane failed to produce tumours at the site of injection.
Repeated injections of C57BL/6J mice with MC-treated S 180 cells resulted in the induction of a protective immune response. Subsequent challenges with S 180, EL4 or LR.4 revealed that the immune response induced under these experimental conditions was specific. This finding was supported when the antiserum obtained from immune animals was tested with other cell lines, some of which belong to unrelated differentiation pathways. Furthermore, the capacity of the antibodies in the immune serum and rabbit complement to lyse S 180 cells provided additional evidence for the participation of a hu- moral immune response in the protection of C57BL/6J mice against this cell line.
The results presented in this paper indicate that S 180 cells escape recogni- tion by immune surveillance mechanisms operating in different inbred strains of mice due, at least in part, to the lack of expression of B2m genes which may render the cell line H-2-deficient. Alternatively, if H-2 class I products were expressed on the membrane of S180 cells in the absence offl2m, these mole- cules were not recognized by the immune system of C57BL/6J mice. We would like to suggest that the antigenic determinant (s) responsible for the induction of a protective immune response in C57BL/6J mice may represent an allelic form of non-H2 determinants or an antigen expressed after malignant trans- formation as has been described by other investigators (Ostrand-Rosenberg and McCarthy, 1984; Ostrand-Rosenberg and Clements, 1987; Schreiber et al., 1988).
ACKNOWLEDGEMENTS
The authors thank the Consejo Nacional de Ciencia y Tecnologia for con- tributing funds through Grant PCCBBNA-020583. We also express our grat- itude to Carla Archer for reviewing the manuscript.

396 G. ALFARO ETAL.
REFERENCES
Allen, H., Fraser, J., Flyer, D., Calvin, S. and Flavell, R., 1986. fl2-microglobulin is not required for cell surface expression in murine class I histocompatibility antigen H - 2 D b of a truncated H - 2 D b. Proc. Natl. Acad. Sci. USA, 83: 7447-7451.
Allison, J.P. and Lanier, L.L., 1987. Structure, function, and serology of the T-cell antigen re- ceptor complex. Ann. Rev. Immunol., 5: 503-540.
Benjamini, E., Fong, S., Erickson, C., Leung, C.Y., Rennick, D. and Sciebienski, R.J., 1977. Immunity to lymphoid tumors induced in syngenic mice by immunization with mitomycin C-treated cells. J. Immunol., 118: 685-693.
Carrington, M.N., Salter, R.D., Cresswell, P. and Ting, J.P.-Y., 1985. Evidence for methylation as a regulatory mechanism in HLA-DRa gene expression. Immunogenetics, 22:219-229.
Cells, E., Hale, A.H., Russell, J.H. and Eisen, H.N., 1979. Cyclical changes in susceptibility of a myeloma tumor (LPC-1) to immune destruction. I. Changes in reactivity with cytotoxic T lymphocytes and anti-H-2 d sera. J. Immunol., 122: 954-958.
Chirwin, J.M., Przybyla, A.E., MacDonald, R.J. and Rutter, W.J., 1979. Isolation of biologi- cally active ribonucleic acid from sources enriched in ribonuclease. Biochemistry, 18: 5294- 5299.
Doyle, A., Martin, W.J., Funa, K., Gazdar, A., Carney, D., Martin, J.E., Linnoila, I., Cuttita, F., Mulshine, J., Bunn, P. and Minna, J., 1985. Markedly decreased expression of class I histocompatibility antigens, protein, and mRNA in human small cell lung cancer. J. Exp. Med.,161:l135-1151.
Dunham, L.J. and Stewart, H.L., 1953. A survey of transplantable and transmissible animal tumors. J. Natl. Cancer Inst., 13:1299-1377.
Eisenbach, L., Hollander, N., Greenfeld, L., Yakor, H., Segal, S. and Feldman, M., 1984. The differential expression of H-2K versus H-2D antigens, distinguishing high-metastatic from low-metastatic clones, is correlated with the immunogenic properties of the tumor cells. Int. J. Cancer, 34: 567-573.
Evans, G.A., Margulies, D.H., Camerini-Otero, R.D., Ozato, K. and Seidman, J.G., 1982. Structure and expression of a mouse major histocompatibility antigen gene, H - 2 L u. Proc. Natl. Acad. Sci. USA., 79:1994-I 998.
Feny6, E.M., Klein, E., Klein, G. and Swiech, K., 1968. Selection of an immunoresistant Mo- loney lymphoma subline with decreased concentration of tumor-specific surface antigens. J. Natl. Cancer Inst., 40: 69-89.
Festenstein, H. and Schmidt, W., 198 I. Variation in MHC antigenic profiles of tumor cells and its biological effects. Immunol. Rev., 60: 85-127.
Fidler, I., Gersten, P. and Hart, I., 1978. The biology of cancer invasion and metastasis. Adv. Cancer Res., 28: 149-250.
Fisher, G.A., 1958. Studies of the culture of leukemic cells in vitro. Ann. NY Acad. Sci., 76: 673.
Flavell, R.A., Allen, H., Burkly, L.C., Sherman, D.H., Waneck, G.L. and Widera, G., 1986. Molecular biology of the H-2 histocompatibility complex. Science, 233:437-443.
Gorer, P.A.0 1950. Studies in antibody response of mice to tumor inoculation. Br. J. Cancer, 4: 372.
Holden, C.A., Shaw, M., McKee, P.H., Sanderson, A.R. and MacDonald, D.M., 1984. Loss of fl2-microglobulin in eccrine porocarcinoma. Arch. Dermatol., 120: 732-735.
Hood, L., Steinmetz, M. and Malissen, B., 1983. Genes of the major histocompatibility complex of the mouse. Ann. Rev. Immunol., 1: 529-568.
Hyman, R. and Stallings, V., 1975. Characterization of a TL- variant of a homozygous TL + mouse lymphoma. Immunogenetics, 3" 75-84.
lsakov, V., Feldman, M. and Segal, S., 198 I. Control of progression of local tumor and pulmo-

ff2M EXPRESSION AND GROWTH ACROSS ALLOGENEIC GROWTH 397
nary metastasis of the 3LL Lewis lung carcinoma by different histocompatibility require- ments in mice. J. Natl. Cancer Inst., 66: 919-926.
Isakov, N., Katzav, S., Feldman, M. and Segal, S., 1983. Loss of expression of transplantation antigens encoded by the H-2K locus on Lewis lung carcinoma cells and its relevance to the tumor's metastatic properties. J. Natl. Cancer Inst., 71:139-145.
Kiirre, K., Ljunggren, H.G., Piontek, G. and Kiessling, R., 1986. Selective rejection of H-2- deficient lymphoma variants suggests alternative immune defence strategy. Nature, 319: 675- 678.
Katzav, S., DeBastelier, P., Tartakovsky, B., Feldman, M. and Segal, S., 1983. Alterations in MHC phenotypes of cloned T 10 sarcoma cells are associated with shifts from non-metastatic to metastatic cells. J. Natl. Cancer Inst., 71: 317-324.
Katzav, S., Segal, S. and Feldman, M., 1984. Immunoselection in vivo of H-2D phenotypic variants from a metastatic clone of sarcoma cells results in cell lines of altered metastatic competence. Int. J. Cancer, 33:407-415.
Katzav, S., Segal, S. and Feldman, M., 1985. Metastatic capacity of cloned T 10 sarcoma cells that differ in H-2 expression: inverse relationship to their immunogenic potency. J. Natl. Cancer Inst., 75: 307-318.
Keeney, J.B., Hedayat, M., Myers, N.M., Connolly, J.M. and Hansen, T.H., 1989. Locus-spe- cific regulation of K, D, and L class I genes in the BALB/c $49 lymphoma sublines. J. Im- munol., 143: 2364-2373.
Klein, J., Figueroa, F. and Nagy, Z.A., 1983. Genetics of the major histocompatibility complex: the final act. Ann. Rev. Immunol., 1:119-142.
Lampson, L.A., Fisher, C.A. and Whelan, J.P., 1983. Striking paucity of HLA-A, B, C, and/72- microglobulin on human neuroblastoma cell lines. J. Immunol., 130:2471-2478.
Ljunggren, H.G. and I~rre , K., 1985. Host resistance directed selectively against H-2-deficient lymphoma variants. Analysis of the mechanism. J. Exp. Med., 162:1745-1759.
Maniatis, T., Fritsch, E.F. and Sambrook, J., 1982. Molecular Cloning. A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
North, R.J., 1984. The murine antitumor immune response and its therapeutic manipulation. Adv. Immunol., 35: 89-155.
Ostrand-Rosenberg, S. and McCarthy, C., 1984. Resistance to murine 402AX teratocarcinoma: regulation by H-21A and H-3 to H-13 region genes. Immunogenetics, 19: 77-82.
Ostrand-Rosenberg, S. and Clements, V., 1987. Resistance to 402AX teratocarcinoma involves immunity to minor histocompatibility antigens. Immunogenetics, 26: 1-5.
Parnes, J.R. and Seidman, J.G., 1982. Structure of wild-type and mutant fl2-microglobulin genes. Cell, 29:661-669.
Parnes, J.R., Sizer, K.C., Seidman, J.G., Stallings, V. and Hyman, R., 1986. A mutational hot- spot within an intron of the mouse fl2-microglobulin gene. EMBO J., 5:103-111.
Potter, T.A., Boyer, C., Schmitt Verhulst, A-M., Golstein, P. and Rajan, T.V., 1984. Expression on (H-2D b) on the cell surface in the absence of detectable fl2-microglobulin. J. Exp. Med., 160:317-322.
Potter, T.A., Zeff, R.A., Schmitt Verhulst, A.-M. and Rajan, T.V., 1985. Molecular analysis of an EL4 cell line that expresses H-2D b but no H-2K b or ff2-microglobulin. Proc. Natl. Acad. Sci. USA, 82: 2950-2954.
Potter, T.A., Frankel, W., Zeff, R.A. and Rajan, T.V., 1987. Spontaneous deletion at the fl2-m locus: evidence for the site-specific genetic rearrangement. J. Immunol., 138:1270-1274.
Ralph, P., 1973. Retention of lymphocyte characteristics by myelomas and 0 +-lymphomas: sen- sitivity to cortisol and phytohemagglutinin. J. Immunol., 110: 1470-1475.
Schreiber, H., Ward, P.L., Rowley, D.A. and Stauss, H.J., 1988. Unique tumor-specific anti- gens. Ann. Rev. Immunol., 6: 465-483.

398 G. ALFAROETAL.
Seyler-Tuyns, A. and Birnstiel, M.L., 1981. Structure and expression in L-cells of a cloned H4 histone gene of the mouse. J. Mol. Biol., 151: 607-625.
Southern, E.M., 1975. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol., 98:503-517.
Tanaka, K., Yoshioka, T., Bieberich, C. and Jay, G., 1988. Role of the major histocompatibility complex class I antigens in tumor, growth and metastasis. Ann. Rev. Immunol., 6: 359-380.
Thomas, P.S., 1980. Hybridization of denatured RNA and small DNA fragments transferred to nitrocellulose. Proc. Natl. Acad. Sci. USA, 77:5201-5205.
Tryggvason, K., Hoyhtya, M. and Salo, T., 1987. Proteolytic degradation ofextracellular matrix in tumor invasion. Biochim. Biophys. Acta, 907:191-197.
Turbitt, M.L. and Mackie, R.M., 1981. Loss offl2-microglobulin from the cell surface of cuta- neous malignant and premalignant lesions. Br. J. Dermatol., 104:507-513.
Wallich, R., Bulbuc, N., H~immerling, G.J., Katzav, S., Segal, S. and Feldman, M., 1985. Abro- gation of metastatic properties of tumor cells by de novo expression of H-2K antigens follow- ing H-2 gene transfection. Nature, 315:301-305.
Young, P.R. and Tilghman, S.M., 1984. Induction of c~-fetoprotein synthesis in differentiating F9 teratocarcinoma cells is accompanied by a genome-wide loss of DNA methylation. Mol. Cell. Biol., 4: 898-907.
Zinkernagel, R.M. and Doherty, P.C., 1979. MHC-restricted cytotoxic T cells: studies on the biological role of polymorphic major transplantation antigens determining T-cell restriction- specificity, function, and responsiveness. Adv. Immunol., 27:51-177.
Related Documents











