Immunology Letters 103 (2006) 33–38 Immunoglobulins can utilize riboflavin (Vitamin B2) to activate the antibody-catalyzed water oxidation pathway Jorge Nieva b , Lisa Kerwin a , Anita D. Wentworth a , Richard A. Lerner a,c , Paul Wentworth Jr. a,c,∗ a Department of Chemistry, The Scripps Research Institute, 10550N. Torrey Pines Road, La Jolla, CA 92037, United States b Department of Molecular and Experimental Medicine, The Scripps Research Institute, 10550N. Torrey Pines Road, La Jolla, CA 92037, United States c Oxford Glycobiology Institute, Department of Biochemistry, University of Oxford, South Parks Road, Oxford OX1 3QU, UK Received 19 November 2005; received in revised form 20 November 2005; accepted 21 November 2005 Available online 15 December 2005 Abstract We have recently discovered a reaction that all antibodies, regardless of source or antigenic specificity can catalyze, that is the reaction between singlet dioxygen ( 1 O 2 * ) and H 2 O to generate H 2 O 2 . We have named this process the antibody-catalyzed water oxidation pathway (ACWOP). As part of our ongoing investigations into the possible biological role of this pathway, we have studied whether isoalloxazine-containing cofactors, that are known to be endogenous photosensitizers via Type-II pathways to generate 1 O 2 * , such as riboflavin (RF, Vitamin B2) can trigger the ACWOP. Herein we show that regardless of the antigenic specificity or heavy and light chain composition, all antibodies and their fragments are able to intercept the 1 O 2 * generated by photo-oxidation of RF in the presence of oxygen (ambient aerobic conditions) to activate the ACWOP. The initial rate of HOOH generation by a panel of murine antibodies ranges from 0.218 to 0.998 M/min. The initial rate of antibody-catalyzed HOOH production is accelerated in D 2 O and is quenched in NaN 3 , highlighting the key intermediacy of 1 O 2 * in the process. Critically, the ACWOP is photo-activated at physiologically relevant concentrations of RF (<50nM) suggesting that this pathway may be relevant in an in vivo setting. Finally, when activated by RF the ACWOP generates oxidants that accelerate the hemolysis of sheep RBCs hinting at a pathophysiological effect of this RF-induced photo-oxidation pathway. © 2005 Elsevier B.V. All rights reserved. Keywords: Antibody; Riboflavin; Catalysis; Oxidation; Hemolysis 1. Introduction One of the most underappreciated facts in immunochemistry is that a certain subclass of antibodies are known to trans- port isoalloxazine-containing cofactors, such as riboflavin (RF), riboflavin 5 -monophosphate (FMN) and flavin adenine dinu- cleotide (FAD) in plasma [1,2]. The likely reason that this has escaped further attention is that until recently there was no apparent role for the redox or photochemical properties of flavin- containing cofactors in the biological function of immunoglob- ulins. However, we have recently shown that all antibodies, regardless of source or antigenic specificity, can catalyze the reaction between singlet oxygen ( 1 O 2 * ) and H 2 O into hydro- gen peroxide (HOOH) [3–5]. During this process oxidants never ∗ Corresponding author. Tel.: +1 858 784 2576; fax: +1 858 784 2593. E-mail address: [email protected] (P. Wentworth Jr.). before considered to occur in biology, such as dihydrogen triox- ide (HOOOH), the hydrotrioxy radical (HOOO • ) [6] and ozone (O 3 ) [7] are postulated to be formed. We have termed this path- way the antibody-catalyzed water oxidation pathway (ACWOP) [4,8]. Experiments have revealed that, upon activation, this oxi- dation pathway can be bactericidal to Gram-negative bacteria [7] and we have gained evidence of its occurrence in activated human neutrophils [7,9], during a reversed Arthus inflammatory reaction and most recently in human atherosclerotic arteries [10]. When considering the relevance of this pathway in vivo we presume that the 1 O 2 * originates from phagocytic cells, such as polymorphonuclear neutrophils, macrophages and eosinophils. However, an alternative but equally well-accepted biological source of 1 O 2 * arises from photochemical activation of pho- tosensitizers in the skin, such as occurs in porphyria or in the retinal macular as seen in patients with macular degeneration. Therefore, it was of interest to learn whether RF, itself known to generate 1 O 2 * under photochemical activation by visible light 0165-2478/$ – see front matter © 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.imlet.2005.11.020

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Immunology Letters 103 (2006) 33–38
Immunoglobulins can utilize riboflavin (Vitamin B2) to activatethe antibody-catalyzed water oxidation pathway
Jorge Nievab, Lisa Kerwina, Anita D. Wentwortha,Richard A. Lernera,c, Paul Wentworth Jr.a,c,∗
a Department of Chemistry, The Scripps Research Institute, 10550 N. Torrey Pines Road, La Jolla, CA 92037, United Statesb Department of Molecular and Experimental Medicine, The Scripps Research Institute, 10550 N. Torrey Pines Road, La Jolla, CA 92037, United States
c Oxford Glycobiology Institute, Department of Biochemistry, University of Oxford, South Parks Road, Oxford OX1 3QU, UK
Received 19 November 2005; received in revised form 20 November 2005; accepted 21 November 2005Available online 15 December 2005
Abstract
We have recently discovered a reaction that all antibodies, regardless of source or antigenic specificity can catalyze, that is the reaction betweensinglet dioxygen (1O * ) and H O to generate HO . We have named this process the antibody-catalyzed water oxidation pathway (ACWOP). Asp cofactors,t theA ments area ACWOP.T dH Pi vo setting.F ogical effecto©
K
1
iprceacurrg
riox-
ath-OP)oxi-teriaatedry
weasphils.icalo-the
ation.n toht
0d
2 2 2 2
art of our ongoing investigations into the possible biological role of this pathway, we have studied whether isoalloxazine-containinghat are known to be endogenous photosensitizers via Type-II pathways to generate1O2
* , such as riboflavin (RF, Vitamin B2) can triggerCWOP. Herein we show that regardless of the antigenic specificity or heavy and light chain composition, all antibodies and their fragble to intercept the1O2
* generated by photo-oxidation of RF in the presence of oxygen (ambient aerobic conditions) to activate thehe initial rate of HOOH generation by a panel of murine antibodies ranges from 0.218 to 0.998�M/min. The initial rate of antibody-catalyzeOOH production is accelerated in D2O and is quenched in NaN3, highlighting the key intermediacy of1O2
* in the process. Critically, the ACWOs photo-activated at physiologically relevant concentrations of RF (<50 nM) suggesting that this pathway may be relevant in an in viinally, when activated by RF the ACWOP generates oxidants that accelerate the hemolysis of sheep RBCs hinting at a pathophysiolf this RF-induced photo-oxidation pathway.2005 Elsevier B.V. All rights reserved.
eywords: Antibody; Riboflavin; Catalysis; Oxidation; Hemolysis
. Introduction
One of the most underappreciated facts in immunochemistrys that a certain subclass of antibodies are known to trans-ort isoalloxazine-containing cofactors, such as riboflavin (RF),iboflavin 5′-monophosphate (FMN) and flavin adenine dinu-leotide (FAD) in plasma[1,2]. The likely reason that this hasscaped further attention is that until recently there was nopparent role for the redox or photochemical properties of flavin-ontaining cofactors in the biological function of immunoglob-lins. However, we have recently shown that all antibodies,egardless of source or antigenic specificity, can catalyze theeaction between singlet oxygen (1O2
* ) and H2O into hydro-en peroxide (HOOH)[3–5]. During this process oxidants never
∗ Corresponding author. Tel.: +1 858 784 2576; fax: +1 858 784 2593.E-mail address: [email protected] (P. Wentworth Jr.).
before considered to occur in biology, such as dihydrogen tide (HOOOH), the hydrotrioxy radical (HOOO•) [6] and ozone(O3) [7] are postulated to be formed. We have termed this pway the antibody-catalyzed water oxidation pathway (ACW[4,8]. Experiments have revealed that, upon activation, thisdation pathway can be bactericidal to Gram-negative bac[7] and we have gained evidence of its occurrence in activhuman neutrophils[7,9], during a reversed Arthus inflammatoreaction and most recently in human atherosclerotic arteries[10].
When considering the relevance of this pathway in vivopresume that the1O2
* originates from phagocytic cells, suchpolymorphonuclear neutrophils, macrophages and eosinoHowever, an alternative but equally well-accepted biologsource of1O2
* arises from photochemical activation of phtosensitizers in the skin, such as occurs in porphyria or inretinal macular as seen in patients with macular degenerTherefore, it was of interest to learn whether RF, itself knowgenerate1O2
* under photochemical activation by visible lig
165-2478/$ – see front matter © 2005 Elsevier B.V. All rights reserved.oi:10.1016/j.imlet.2005.11.020

34 J. Nieva et al. / Immunology Letters 103 (2006) 33–38
[11], is capable of activating the water oxidation pathway ofantibodies.
Herein we show that RF is efficient at activating the water oxi-dation pathway in antibodies derived from a range of species,and with a mixture of heavy and light chain compositions in aprocess that is promoted in deuterium oxide (D2O) and inhibitedby sodium azide. The ACWOP can be activated by RF at con-centrations of the cofactor that exist under normal conditions inplasma (<50 nM), and upon activation the ROS generated by theantibodies is capable of accelerating hemolysis in vitro. Thesecombined data reinforce the possibility that this pathway maybe pathological in vivo.
2. Materials and methods
2.1. Antibodies and fragments
Whole antibodies were obtained from PharMingen: S.S.1.2(mouse IgG2a, �), N.S.8.1.1 (mouse IgG2b, �), N.S.7.1 (mouseIgG3). The following murine monoclonal antibodies wereobtained ‘in house’ using hybridoma technology and purifiedas described previously: JW38C2 (mouse IgG2a, �) [12], DC36H10 (mouse IgG2b, �), EP2 19G2 (mouse IgG2b, �) [13],EP2 22B9 (mouse IgG2b, �), EP2 25C10 (mouse IgG2b, �),MT3 38H10 (mouse IgG2b, �), MT3 40C10 (mouse IgG2b,�), MT4 7C5 (mouse IgG , �), MT4 7E4 (mouse IgG , �),N�P yF ious[ 12)w atop as-r
2
ateb as ddetp me2 pro-t atop -t erer ousld -t at.N( ast minT sureu heh stand ande is
was performed using Prism 4 for Macintosh (Graphpad SoftwareInc.).
2.3. Photoirradiation of JW38C2 in the presence of D2Oand NaN3
A solution of antibody JW38C2 (murine IgG, 20�M) in PBS(pH 7.4) and RF (60�M) was irradiated on a transilluminatorplate (white light, 2.0 mW cm−2 Fischer-Biotech transillumina-tor) at 37◦C. Hydrogen peroxide concentration was determinedas described above. The assay was also performed in the pres-ence of NaN3 (100 mM) or PBS in D2O. All experiments wererun in at least duplicate and each time point is reported as themean± S.E.M. Data analysis was performed using Prism 4 forMacintosh (Graphpad Software Inc.).
2.4. Hemolysis experiments
A 1 mL solution of whole antibody or antibody fragment(20�M) in PBS (pH 7.4), RF (60�M) and sheep red blood cells(RBC, 108 to 109 cells/mL) in a 24-well cell culture plate wasirradiated on a transilluminator plate (white light, 2.0 mW cm−2
Fischer-Biotech transilluminator) at 37◦C. At specific times,aliquots were removed (50�L) and the OD of this solution wasmeasured at 405 nm. Raw OD data were then converted to num-ber of hemolysed cells by interpolation of a standard curve. Alle pointi edu
3
3p
g el er-iHa ,t PIX( arlys dt adst isonb asiso ux( ithRr ions n-er
odyJ t toe urine
2b 2bPN 43C9 (mouse IgG2b, �) [14], PW6 1F5 (mouse IgG2b,), PW6 2F9 (mouse IgG2b, �), PW6 19A7 (mouse IgG2b, �),W6 7E9 (mouse IgG2a, �), PW6 8H10 (mouse, IgG). Antibodab and Fc fragments were prepared as described prev
3]. AmplexTM Red hydrogen peroxide assay kits (A-122ere obtained from Molecular Probes. Riboflavin and hemorphyrin IX were obtained from Sigma–Aldrich and usedeceived.
.2. Photoirradiation experiments
Photosensitizer, riboflavin (from a stock in phosphuffered saline, PBS pH 7.4) or hematoporphyrin IX (fromtock in PBS, pH 7.4), at a range of concentrations was ao a solution of antibody or antibody fragment (20�M in PBSH 7.4) in a 96-well microtiter plate well (final reaction volu00�L). The reaction well was sealed with a glass ball. The
ein/sensitizer solution was then irradiated on a transilluminlate (white light, 2.0 mW cm−2 Fischer-Biotech transillumina
or) at 37◦C. At times throughout the irradiation aliquots wemoved and the solution was assayed for HOOH as previescribed[3]. In brief, the aliquot (20�L) from the protein solu
ion added into a well of a 96-well microtiter plate (Costar Co. 3603) containing reaction buffer (80�L). Working solution
100�L, 400�M AmplexTM Red reagent and 2 U/mL HRP) when added and the plate was incubated in the dark for 30he fluorescence of the well components was then measing a Multiwell Plate Reader (EX: 563 nm; EM: 587 nm). Tydrogen peroxide concentration was determined using aard curve. All experiments were run in at least duplicateach time point is reported as the mean± S.E.M. Data analys
ly
-
d
r
y
.d
-
xperiments were run in at least duplicate and each times reported as the mean± S.E.M. Data analysis was performsing Prism 4 for Macintosh (Graphpad Software Inc.).
. Results
.1. Hydrogen peroxide generation by antibodies duringhotoirradiation of RF
To elucidate whether antibodies are able to utilize the1O2*
enerated by RF (60�M) upon photoirradiation with visiblight to activate the ACWOP we performed irradiation expments under physiologic conditions (PBS, pH 7.4, 37◦C).OOH production was quantified using the AmplexTM Redssay as previously described[3] (Fig. 1A). For comparison
he HOOH generation by antibodies in the presence of H60�M) and white light was also measured. The data clehow that RF is a valid source of1O2
* for antibodies anhat the photoirradiation of RF in the presence of IgG leo a time-dependent production in HOOH. The comparetween HPIX and RF reveals that on a mole by mole bf photosensitizer (60�M) and with the same average light fl2.0 mW cm−2) antibody JW38C2 generates HOOH faster wF (0.646�M/min, r2 = 0.984) than with HPIX (0.381�M/min,
2 = 0.999) (Fig. 1A). The progress curves for HOOH producthown inFig. 1A are corrected for the initial rate of HOOH gerated by irradiation of the sensitizer alone (RF = 0.423�M/min,2 = 0.998; HPIX = 0.002�M/min, r2 = 0.985).
Having established that HOOH is generated by antibW38C2 upon photoirradiation with RF it was of interesxplore the scope of this process. Thus, a panel of m
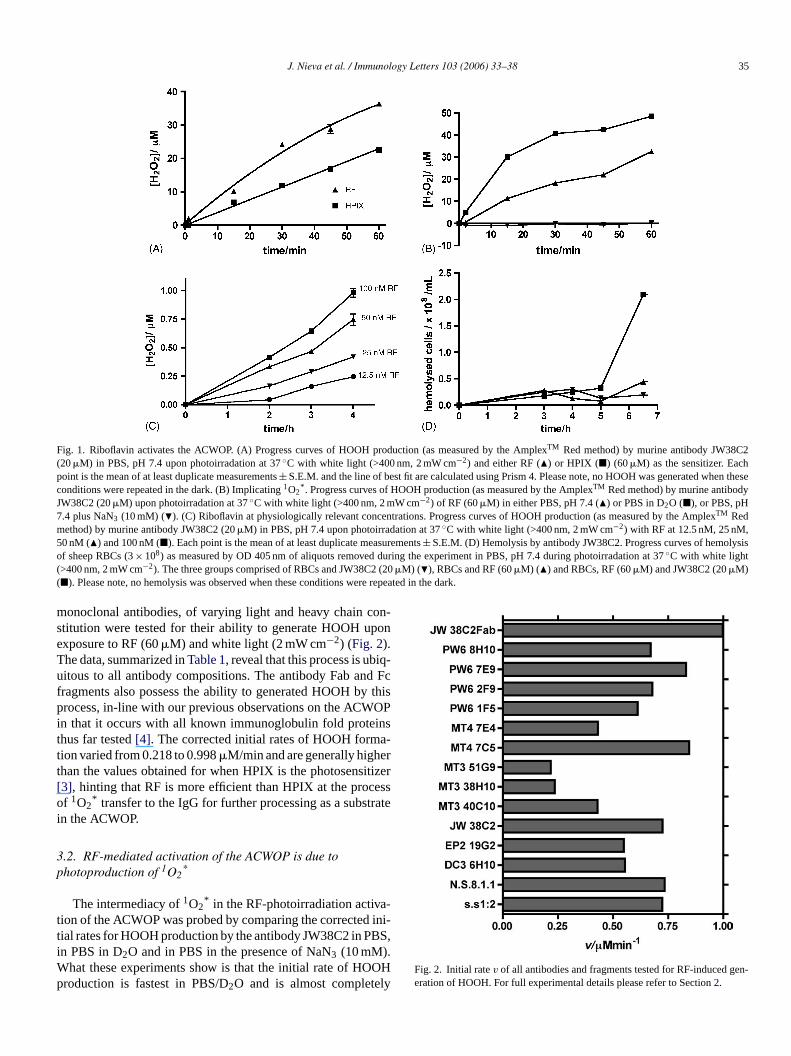
J. Nieva et al. / Immunology Letters 103 (2006) 33–38 35
Fig. 1. Riboflavin activates the ACWOP. (A) Progress curves of HOOH production (as measured by the AmplexTM Red method) by murine antibody JW38C2(20�M) in PBS, pH 7.4 upon photoirradation at 37◦C with white light (>400 nm, 2 mW cm−2) and either RF (�) or HPIX (�) (60�M) as the sensitizer. Eachpoint is the mean of at least duplicate measurements± S.E.M. and the line of best fit are calculated using Prism 4. Please note, no HOOH was generated when theseconditions were repeated in the dark. (B) Implicating1O2
* . Progress curves of HOOH production (as measured by the AmplexTM Red method) by murine antibodyJW38C2 (20�M) upon photoirradation at 37◦C with white light (>400 nm, 2 mW cm−2) of RF (60�M) in either PBS, pH 7.4 (�) or PBS in D2O (�), or PBS, pH7.4 plus NaN3 (10 mM) (�). (C) Riboflavin at physiologically relevant concentrations. Progress curves of HOOH production (as measured by the AmplexTM Redmethod) by murine antibody JW38C2 (20�M) in PBS, pH 7.4 upon photoirradation at 37◦C with white light (>400 nm, 2 mW cm−2) with RF at 12.5 nM, 25 nM,50 nM (�) and 100 nM (�). Each point is the mean of at least duplicate measurements± S.E.M. (D) Hemolysis by antibody JW38C2. Progress curves of hemolysisof sheep RBCs (3× 108) as measured by OD 405 nm of aliquots removed during the experiment in PBS, pH 7.4 during photoirradation at 37◦C with white light(>400 nm, 2 mW cm−2). The three groups comprised of RBCs and JW38C2 (20�M) (�), RBCs and RF (60�M) (�) and RBCs, RF (60�M) and JW38C2 (20�M)(�). Please note, no hemolysis was observed when these conditions were repeated in the dark.
monoclonal antibodies, of varying light and heavy chain con-stitution were tested for their ability to generate HOOH uponexposure to RF (60�M) and white light (2 mW cm−2) (Fig. 2).The data, summarized inTable 1, reveal that this process is ubiq-uitous to all antibody compositions. The antibody Fab and Fcfragments also possess the ability to generated HOOH by thisprocess, in-line with our previous observations on the ACWOPin that it occurs with all known immunoglobulin fold proteinsthus far tested[4]. The corrected initial rates of HOOH forma-tion varied from 0.218 to 0.998�M/min and are generally higherthan the values obtained for when HPIX is the photosensitizer[3], hinting that RF is more efficient than HPIX at the processof 1O2
* transfer to the IgG for further processing as a substratein the ACWOP.
3.2. RF-mediated activation of the ACWOP is due tophotoproduction of 1O2
*
The intermediacy of1O2* in the RF-photoirradiation activa-
tion of the ACWOP was probed by comparing the corrected ini-tial rates for HOOH production by the antibody JW38C2 in PBS,in PBS in D2O and in PBS in the presence of NaN3 (10 mM).What these experiments show is that the initial rate of HOOHproduction is fastest in PBS/D2O and is almost completely
Fig. 2. Initial ratev of all antibodies and fragments tested for RF-induced gen-eration of HOOH. For full experimental details please refer to Section2.
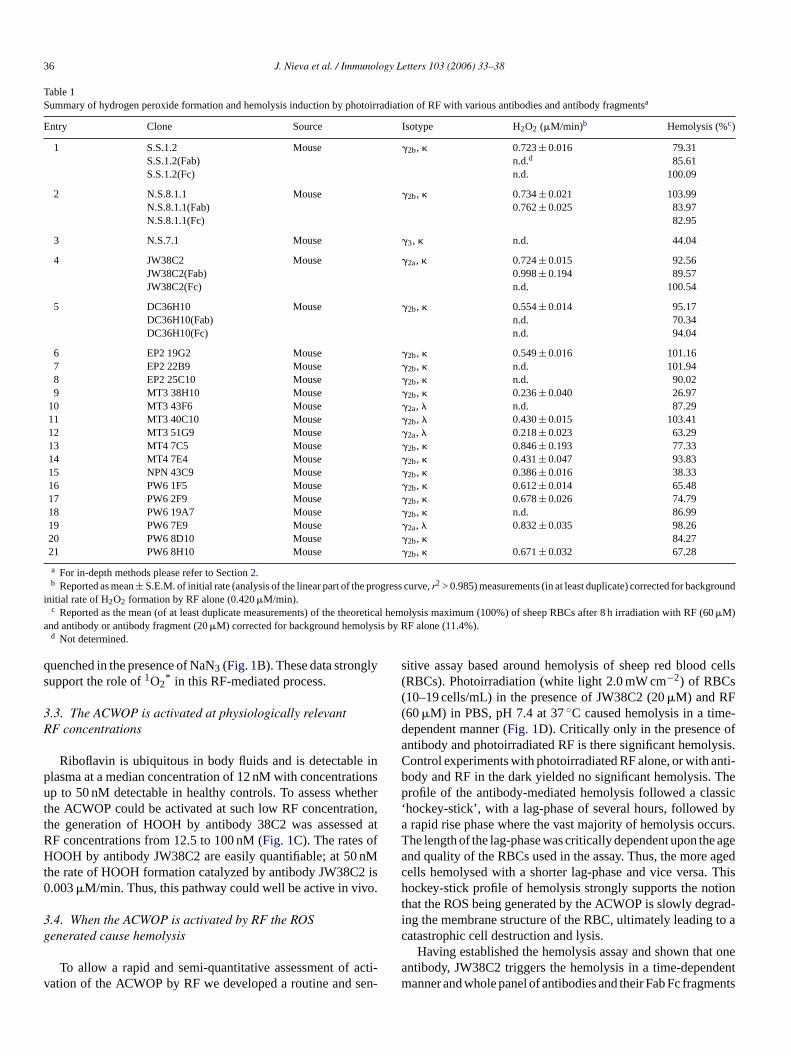
36 J. Nieva et al. / Immunology Letters 103 (2006) 33–38
Table 1Summary of hydrogen peroxide formation and hemolysis induction by photoirradiation of RF with various antibodies and antibody fragmentsa
Entry Clone Source Isotype H2O2 (�M/min)b Hemolysis (%c)
1 S.S.1.2 Mouse �2b, � 0.723± 0.016 79.31S.S.1.2(Fab) n.d.d 85.61S.S.1.2(Fc) n.d. 100.09
2 N.S.8.1.1 Mouse �2b, � 0.734± 0.021 103.99N.S.8.1.1(Fab) 0.762± 0.025 83.97N.S.8.1.1(Fc) 82.95
3 N.S.7.1 Mouse �3, � n.d. 44.04
4 JW38C2 Mouse �2a, � 0.724± 0.015 92.56JW38C2(Fab) 0.998± 0.194 89.57JW38C2(Fc) n.d. 100.54
5 DC36H10 Mouse �2b, � 0.554± 0.014 95.17DC36H10(Fab) n.d. 70.34DC36H10(Fc) n.d. 94.04
6 EP2 19G2 Mouse �2b, � 0.549± 0.016 101.167 EP2 22B9 Mouse �2b, � n.d. 101.948 EP2 25C10 Mouse �2b, � n.d. 90.029 MT3 38H10 Mouse �2b, � 0.236± 0.040 26.97
10 MT3 43F6 Mouse �2a, � n.d. 87.2911 MT3 40C10 Mouse �2b, � 0.430± 0.015 103.4112 MT3 51G9 Mouse �2a, � 0.218± 0.023 63.2913 MT4 7C5 Mouse �2b, � 0.846± 0.193 77.3314 MT4 7E4 Mouse �2b, � 0.431± 0.047 93.8315 NPN 43C9 Mouse �2b, � 0.386± 0.016 38.3316 PW6 1F5 Mouse �2b, � 0.612± 0.014 65.4817 PW6 2F9 Mouse �2b, � 0.678± 0.026 74.7918 PW6 19A7 Mouse �2b, � n.d. 86.9919 PW6 7E9 Mouse �2a, � 0.832± 0.035 98.2620 PW6 8D10 Mouse �2b, � 84.2721 PW6 8H10 Mouse �2b, � 0.671± 0.032 67.28
a For in-depth methods please refer to Section2.b Reported as mean± S.E.M. of initial rate (analysis of the linear part of the progress curve,r2 > 0.985) measurements (in at least duplicate) corrected for background
initial rate of H2O2 formation by RF alone (0.420�M/min).c Reported as the mean (of at least duplicate measurements) of the theoretical hemolysis maximum (100%) of sheep RBCs after 8 h irradiation with RF (60�M)
and antibody or antibody fragment (20�M) corrected for background hemolysis by RF alone (11.4%).d Not determined.
quenched in the presence of NaN3 (Fig. 1B). These data stronglysupport the role of1O2
* in this RF-mediated process.
3.3. The ACWOP is activated at physiologically relevantRF concentrations
Riboflavin is ubiquitous in body fluids and is detectable inplasma at a median concentration of 12 nM with concentrationsup to 50 nM detectable in healthy controls. To assess whetherthe ACWOP could be activated at such low RF concentration,the generation of HOOH by antibody 38C2 was assessed atRF concentrations from 12.5 to 100 nM (Fig. 1C). The rates ofHOOH by antibody JW38C2 are easily quantifiable; at 50 nMthe rate of HOOH formation catalyzed by antibody JW38C2 is0.003�M/min. Thus, this pathway could well be active in vivo.
3.4. When the ACWOP is activated by RF the ROSgenerated cause hemolysis
To allow a rapid and semi-quantitative assessment of acti-vation of the ACWOP by RF we developed a routine and sen-
sitive assay based around hemolysis of sheep red blood cells(RBCs). Photoirradiation (white light 2.0 mW cm−2) of RBCs(10–19 cells/mL) in the presence of JW38C2 (20�M) and RF(60�M) in PBS, pH 7.4 at 37◦C caused hemolysis in a time-dependent manner (Fig. 1D). Critically only in the presence ofantibody and photoirradiated RF is there significant hemolysis.Control experiments with photoirradiated RF alone, or with anti-body and RF in the dark yielded no significant hemolysis. Theprofile of the antibody-mediated hemolysis followed a classic‘hockey-stick’, with a lag-phase of several hours, followed bya rapid rise phase where the vast majority of hemolysis occurs.The length of the lag-phase was critically dependent upon the ageand quality of the RBCs used in the assay. Thus, the more agedcells hemolysed with a shorter lag-phase and vice versa. Thishockey-stick profile of hemolysis strongly supports the notionthat the ROS being generated by the ACWOP is slowly degrad-ing the membrane structure of the RBC, ultimately leading to acatastrophic cell destruction and lysis.
Having established the hemolysis assay and shown that oneantibody, JW38C2 triggers the hemolysis in a time-dependentmanner and whole panel of antibodies and their Fab Fc fragments

J. Nieva et al. / Immunology Letters 103 (2006) 33–38 37
were studied in this assay format for their ability to trigger thehemolysis of the sheep RBCs upon photoirradiation with RF.A summary of the data is shown inTable 1. Clearly what thehemolysis experiments show is that, relative to RF alone andregardless of heavy and light chain construction, all antibodiesand their fragments generate ROS that contribute to hemolysis.
4. Discussion
Since our discovery that antibodies have the intrinsic abilityto catalyze the conversion of1O2
* into hydrogen peroxide, wehave instigated a search for both evidence of its occurrence incomplex biological systems and its potential relevance in eitherimmune defense or immunopathology. For this pathway to beactive one has to juxtapose immunoglobulins with singlet dioxy-gen (1O2
* ). Within a biological setting1O2* can originate from
to main sources, activated leukocytes and photosensitization ofbiological molecules. With this in mind, we have turned outattention to an investigation of antibodies and biological sensi-tizers such as the isoalloxazine-containing flavin cofactors.
Riboflavin is a water-soluble vitamin that serves as a precur-sor for flavin mononucleotide and FAD[15] and is a well-studiedendogenous photosensitizer. Flavin photochemistry is an inten-sive area for research and was originally promoted because of theproposed involvement of flavin excited states in several photobi-ological and photochemical processes[11]. The RF absorptions s cet tionc oU en-d izedo n off n Rt essit Fh na uress
frag-m -IIp ateoH withH ho-t littles oure(t thaa ut as ction[ ed ationo
ti-b is
activated or whether this is simply non-specific photoreduc-tion of molecular oxygen by antibody side chains. The rate ofHOOH in this RF-photosensitized process is increased in D2O-containing buffers and is quenched in the presence of NaN3(10 mM) (Fig. 1B). Both of these effects are in-line with theknown increase in lifetime of1O2
* in deuterated buffers and thequenching of1O2
* by azide.The destruction of red blood cells via riboflavin photo-
oxidation is a well-studied multifactorial process that is notfully understood[18–21]. We decided to utilize this process as areporter assay for our ACWOP assay and found that all antibod-ies, regardless of heavy or light chain construction and regardlessof whether they were specific for RBC surface markers, accel-erate the RF-photoinduced hemolysis (Fig. 1D and Table 1).In most cases the elevation is profound. For comparison, underconditions where the RF-induced hemolysis was∼11% of thetotal sheep RBCs in a samples, the vast majority of Igs had causebetween 80 and 100% of the maximal hemolysis. While the rel-evance of the ACWOP to RF-induced hemolysis in vivo has notat all been investigated, this study hints that Ig, which are presentin vivo at >10 mg/mL could contribute in part to the RF-inducedhemolysis in the presence of a suitable photoirradiation source.
In conclusion, this report shows that the intrinsic catalyticoxidation of water by antibodies can be activated by a knownendogenous photosensitizer at concentrations of the sensitizerthat are normally observed in vivo. We have shown that thisa d-i Ig.W BCsh ossi-b areb way,b havei
A
rnerL anti-b orkw d theA
R
an
ndInt
nti-Acad
il-nce
gletSci
pectrum in aqueous media exhibits four structureless peakered at 446, 475, 265 and 220 nm with high molar extincoefficients (>104 M−1 cm−1). RF is particularly sensitive tV and visible light and induces both Type-I (oxygen indepent) and critically Type-II (oxygen-dependent) photosensitxidation mechanisms. The former involves the formatio
ree radicals through hydrogen or electron transfer betweeriplet excited state (3RF* ) and substrates. The Type-II procnvolves the formation of1O2
* by energy transfer from3RF*
o triplet molecular oxygen (3O2). The photoreactivity of Ras been linked to the lens damage and cataract formatio[16]nd the deleterious effects of light exposure to nutrient mixtuch as paranteral solutions and infant formulae[17].
Our studies have shown that immunoglobulins and theirents can intercept the1O2
* generated by RF during a Typehoto-oxidation process to activate the antibody-catalyzed wxidation pathway (Figs. 1 and 2andTable 1). The profile of theOOH generation by antibodies by RF, when comparedPIX offers an in-sight into the difference between the p
ochemistry of the two sensitizers. HPIX generates veryuperoxide (via Type-I photoxidation) compared to RF. Inxperiments, the initial ratev of HOOH formation by HPIX60�M) was 0.002�M/min as compared to 0.45�M/min byhe same concentration of RF. We have shown previouslyntibodies can generate >100 equivalents of HOOH withoignificant decrease in the observed rate of HOOH produ4]. Thus, the no-linearity observed inFig. 1A is presumed to bue to a loss of photosensitizer due to the Type-I photo-oxidf RF.
The intermediacy of1O2* in the HOOH generation by an
odies is always a critical test of whether the ACWOP
n-
F
r
t
ctivation is linked to a Type-II photo-oxidation of RF leang to the generation of1O2
* that is then intercepted by thehen the ACWOP is activated in the presence of sheep R
emolysis is accelerated relative to RF alone, hinting at a ple pathogenic role for this pathway in vivo. Further studieseing directed at furthering our understanding of this pathoth in terms of the oxidants it generates, the impact they
n immune defense and their pathological consequences.
cknowledgements
The authors would like to thank all members of the Leaboratory, especially Diane Kubitz, for their assistance inody production and purification during this project. This was supported by the Skaggs Institute for Research anLSAM foundation.
eferences
[1] Baker H, Frank O, Feingold S, Leevy CM. Vitamin distribution in humplasma proteins. Nature 1967;215:84–5.
[2] Watson CD, Ford HC. High-affinity binding of riboflavin aFAD by immunoglobulins from normal human serum. Biochem1988;16:1067–74.
[3] Wentworth AD, Jones LH, Wentworth P, Janda KD, Lerner RA. Abodies have the intrinsic capacity to destroy antigens. Proc NatlSci USA 2000;97:10930–5.
[4] Wentworth Jr P, Jones LH, Wentworth AD, Zhu X, Larsen NA, Wson IA, et al. Antibody catalysis of the oxidation of water. Scie2001;293:1806–9.
[5] Xu X, Muller PR, Goddard III WA. The gas phase reaction of sindioxygen with water: a water-catalyzed reaction. Proc Natl AcadUSA 2002;99:3376–81.

38 J. Nieva et al. / Immunology Letters 103 (2006) 33–38
[6] Wentworth Jr P, Wentworth AD, Zhu X, Wilson IA, Janda KD, Eschen-moser A, et al. Evidence for the production of trioxygen species duringantibody-catalyzed chemical modifications of antigens. Proc Natl AcadSci USA 2003;100:1490–3.
[7] Wentworth Jr P, McDunn J, Wentworth AD, Takeuchi C, Nieva J, JandaKD, et al. Evidence for antibody-catalyzed ozone formation in bacterialkilling and inflammation. Science 2002;298:2195–9.
[8] Nieva J, Wentworth P. The antibody-catalyzed water oxidationpathway—a new chemical arm to immune defense? TIBS 2004;29:274–8.
[9] Babior BM, Takeuchi C, Ruedi J, Guitierrez A, Wentworth Jr P. Inves-tigating ozone generation by human neutrophils. Proc Natl Acad SciUSA 2003;100:3920–5.
[10] Wentworth Jr P, Nieva J, Takeuchi C, Galve R, Wentworth AD, Dil-ley RB, et al. Evidence for ozone generation in human atheroscleroticarteries. Science 2003;302:1053–6.
[11] Heelis PF. The photophysical and photochemical properties of flavins(isoalloxazines). Chem Soc Rev 1982;11:15–51.
[12] Wagner J, Lerner RA, Barbas III CF. Efficient aldolase catalytic anti-bodies that use the enamine mechanism of the natural enzyme. Science1995;270:1797–800.
[13] Simeonov A, Matsushita M, Juban EA, Thompson EHZ, HoffmanTZ, Beuscher AE, et al. Blue-fluorescent antibodies. Science 2000;290:307–13.
[14] Janda KD, Schloeder D, Benkovic SJ, Lerner RA. Induction of anantibody that catalyzes the hydrolysis of an amide bond. Science1988;241:1188–91.
[15] Cooperman JM, Lopez R. Riboflavin. In: Machlin LJ, editor. Handbookof vitamins. New York: Marcel Dekker; 1991. p. 283–310.
[16] de La Rochette A, Silva E, Birlouez-Aragon I, Mancini M, Edwards A-M, Morliere P. Riboflavin photodegradation and photosensitizing effectsare highly dependent on oxygen and ascorbate concentrations. Pho-tochem Photobiol 2000;72:815–20.
[17] Lee KH, Jung MY, Kim SY. Effects of ascorbic acid on the light-inducedriboflavin degradation and color changes in milks. J Agric Food Chem1998;46:407–10.
[18] Bereswill S, Fassbinder F, V̈olzing C, Covacci A, Haas R, Kist M.Hemolytic properties and riboflavin synthesis ofHelicobacter pylori:cloning and functional characterization of the ribA gene encoding GTP-cyclohydrolase II that confers hemolytic activity toEschericia coli. MedMicrobiol Immunol (Berl) 1998;186:177–87.
[19] Misra RB, Bajpai PK, Joshi PC, Hans RK. An unusual photohaemolyticproperty of riboflavin. Food Chem Toxicol 2001;39:11–8.
[20] Ali L, Naseem L. Hemolyis of human red blood cells by combinationof riboflavin and aminophylline. Life Sci 2002;70:2013–22.
[21] Ali L, Sakhnini N, Naseem L. Hemolysis of human red bloodcells by riboflavin–Cu(II) system: enhancement by azide. Biochemistry(Moscow) 2005;70:1011–4.
Related Documents











