The Journal of Histotechnology / Vol. 33, No. 1 / March 2010 41 Abstract The present studies describe the development of immuno- histochemical methods for characterizing the presence of adenosine A 2A receptors on basal ganglia neurons in humans and rats. Antigen retrieval methods were incorporated to decrease the expression of nonspecific tissue binding and to amplify antigen sensitivity. Morphology of the tissue sec- tions was not disturbed by antigen retrieval, and the quality of staining was enhanced substantially. Conditions were optimized for visualizing adenosine A 2A receptor antibody in sections of rat and human basal ganglia, and comparing the distributions of A 2A receptor labeling across multiple striatal cell populations. The retrograde tracer cholera toxin B was injected into different pallidal regions to visualize projec- tions from multiple striatal subregions, and to determine if the cholera toxin B-labeled striatal neurons also contained adenosine A 2A receptor immunoreactivity. Apparent double- labeled neurons were observed in both nucleus accumbens and neostriatum. Thus, these immunohistochemical methods provided evidence that striatal neurons that topographically project to distinct pallidal areas also contain adenosine A 2A receptor immunoreactivity. Development of immunohis- tochemical methods can contribute to our understanding of the role that basal ganglia receptors play in modulating the forebrain circuitry involved in motor functions and effort- related motivational processes. ( The J Histotechnol 33(1): 41–47, 2010) Submitted November 30, 2009; accepted February 8, 2010. Key words: antigen retrieval, pallidum, receptors, striatum Introduction Histological analyses of the microstructure of the basal ganglia can complement studies in a number of disciplines, such as physiology, behavioral neuroscience, neuropharma- cology, and neuropathology, which all provide different approaches aimed at understanding the mechanisms by which various conditions alter brain function (1). One particular area of the basal ganglia that is of tremendous research inter- est is the caudate/putamen, collectively known as the neo- striatum in humans and rats because of the striations apparent in the tissue. Approximately 90% of striatal neurons are medium-sized spiny neurons that use gamma-aminobutyric acid (GABA) as their neurotransmitter and have dense den- dritic spines arranged in a distinctive pattern (1). The GABAergic neurons of the striatum are output neurons that are organized into two parallel, histochemically distinct sys- tems. In one system, the “direct” pathway, the neurons pref- erentially express substance P and dynorphin, and the axons extend to the internal segment of the globus pallidus and the substantia nigra pars reticulata. In contrast, the other system, the “indirect” pathway, has neurons that express enkephalin, and the axons extend to the external globus pallidus (entope- duncular nucleus in the rat) and the subthalamic nucleus (1,2). It is thought that the direct pathway system is largely modulated by dopamine (DA) D1 and adenosine A 1 recep- tors, whereas the indirect system is more likely to be modu- lated by DA D2 and adenosine A 2A receptors (3–5). Understanding how this circuitry works, and how it is modu- lated by various transmitters, is essential to comprehending Address reprint requests to Regina Vontell, Institute of Reproductive and Developmental Biology, Laboratory of Fetal Medicine and Perinatal Brain Injury, Imperial College London, IRDB Building Level 5, Du Cane Road, London W12 0NN, UK. E-mail: [email protected] Immunocytochemistry Studies of Basal Ganglia Adenosine A 2A Receptors in Rat and Human Tissue Regina Vontell 1–4 , Kristen N. Segovia 1 , Adrienne J. Betz 1,5 , Susana Mingote 1,6 , Kristin Goldring 2 , Richard W. Cartun 4 , and John D. Salamone 1 1 Department of Psychology, University of Connecticut, Storrs, CT 2 UK Parkinson’s Disease Society Tissue Bank, Division of Neuroscience and Mental Health, Imperial College, London, UK 3 Institute of Reproductive and Developmental Biology, Laboratory of Fetal Medicine and Perinatal Brain Injury, Imperial College, London, UK 4 Department of Immunopathology, Hartford Hospital, Hartford, CT 5 Department of Psychology, Quinnipiac University, New Haven, CT 6 Department of Psychiatry, Columbia University, New York, NY

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The Journal of Histotechnology / Vol. 33, No. 1 / March 2010 41
Abstract The present studies describe the development of immuno-
histochemical methods for characterizing the presence of adenosine A
2A receptors on basal ganglia neurons in humans
and rats. Antigen retrieval methods were incorporated to decrease the expression of nonspecific tissue binding and to amplify antigen sensitivity. Morphology of the tissue sec-tions was not disturbed by antigen retrieval, and the quality of staining was enhanced substantially. Conditions were optimized for visualizing adenosine A
2A receptor antibody in
sections of rat and human basal ganglia, and comparing the distributions of A
2A receptor labeling across multiple striatal
cell populations. The retrograde tracer cholera toxin B was injected into different pallidal regions to visualize projec-tions from multiple striatal subregions, and to determine if the cholera toxin B-labeled striatal neurons also contained adenosine A
2A receptor immunoreactivity. Apparent double-
labeled neurons were observed in both nucleus accumbens and neostriatum. Thus, these immunohistochemical methods provided evidence that striatal neurons that topographically project to distinct pallidal areas also contain adenosine A
2A
receptor immunoreactivity. Development of immunohis-tochemical methods can contribute to our understanding of the role that basal ganglia receptors play in modulating the forebrain circuitry involved in motor functions and effort-related motivational processes. ( The J Histotechnol 33(1):41–47, 2010)
Submitted November 30, 2009; accepted February 8, 2010.
Key words: antigen retrieval , pallidum , receptors , striatum
Introduction Histological analyses of the microstructure of the basal
ganglia can complement studies in a number of disciplines, such as physiology, behavioral neuroscience, neuropharma-cology, and neuropathology, which all provide different approaches aimed at understanding the mechanisms by which various conditions alter brain function (1). One particular area of the basal ganglia that is of tremendous research inter-est is the caudate/putamen, collectively known as the neo-striatum in humans and rats because of the striations apparent in the tissue. Approximately 90% of striatal neurons are medium-sized spiny neurons that use gamma-aminobutyric acid (GABA) as their neurotransmitter and have dense den-dritic spines arranged in a distinctive pattern (1). The GABAergic neurons of the striatum are output neurons that are organized into two parallel, histochemically distinct sys-tems. In one system, the “direct” pathway, the neurons pref-erentially express substance P and dynorphin, and the axons extend to the internal segment of the globus pallidus and the substantia nigra pars reticulata. In contrast, the other system, the “indirect” pathway, has neurons that express enkephalin, and the axons extend to the external globus pallidus (entope-duncular nucleus in the rat) and the subthalamic nucleus (1,2). It is thought that the direct pathway system is largely modulated by dopamine (DA) D1 and adenosine A
1 recep-
tors, whereas the indirect system is more likely to be modu-lated by DA D2 and adenosine A
2A receptors (3–5).
Understanding how this circuitry works, and how it is modu-lated by various trans mitters, is essential to comprehending
Address reprint requests to Regina Vontell, Institute of Reproductive and Developmental Biology, Laboratory of Fetal Medicine and Perinatal Brain Injury, Imperial College London, IRDB Building Level 5, Du Cane Road, London W12 0NN, UK. E-mail: [email protected]
Immunocytochemistry Studies of Basal Ganglia Adenosine A
2A Receptors in Rat and
Human Tissue Regina Vontell 1–4 , Kristen N. Segovia 1 , Adrienne J. Betz 1 , 5 , Susana Mingote 1 , 6 , Kristin Goldring 2 , Richard W. Cartun 4 , and John D. Salamone 1 1 Department of Psychology, University of Connecticut, Storrs, CT 2 UK Parkinson’s Disease Society Tissue Bank, Division of Neuroscience and Mental Health, Imperial College, London, UK 3 Institute of Reproductive and Developmental Biology, Laboratory of Fetal Medicine and Perinatal Brain Injury, Imperial College, London, UK 4 Department of Immunopathology, Hartford Hospital, Hartford, CT 5 Department of Psychology, Quinnipiac University, New Haven, CT 6 Department of Psychiatry, Columbia University, New York, NY

IHC of Basal Ganglia Adenosine A2A Receptors / Vontell et al.42
normal motor functions, as well as movement disorders such as Parkinson’s disease (6).
The antagonistic interaction between adenosine A 2A
and DA D2 receptors is an important consideration for Parkinson’s disease, because prolonged administration of L-Dopa, which is the most common treatment, induces dyskinesias, visual hallucinations, and psychotic symptoms (7). Adenosine has both anxiolytic and hypnotic effects, whereas blocking endogenous adenosine through the use of caffeine or other methylxanthines increases alertness and improves cognitive function (8). Administration of more selective adenosine A
2A receptor antagonists, such as KW-6002, KF17837, or
MSX-3, can produce anti-Parkinsonian effects in animal models (6,9).
Our laboratory investigates the behavioral effects of drugs that act upon adenosine A
2A receptors and how these drugs
interact with the forebrain circuitry involved in motor func-tions and effort-related motivational processes. Mingote and coworkers (10) investigated the relationship between an A
2A
agonist CGS-21680 and effort-related processes modulated by the nucleus accumbens, which is a striatal subregion. This investigation examined how intra-accumbens injections of CGS-21680 decreased lever-pressing behavior on tasks with higher work requirements. Along with the motivational impairment, this study also noted an increase of extracellular GABA in the ventral pallidum induced by administration of CGS-21680 into the accumbens. A bilateral injection of the GABA
A agonist muscimol into the ventral pallidum, when
combined with the contralateral injection of CGS-21680 into the nucleus accumbens, substantially suppressed operant response on a schedule with high work requirements. Thus, it was concluded that the effect observed in DA-depleted rats could be mimicked by stimulating the accumbens adenosine A
2A receptors.
Rosin and colleagues (4) characterized the distinct distri-bution of adenosine A
2A receptor immunoreactivity by using
a monoclonal antibody they made against human A 2A
ade-nosine receptors. The A
2A immunoreactivity showed a dif-
fuse pattern of labeling in the neostriatum, nucleus accumbens core and shell, and the olfactory tubercles (5). The ability to visualize adenosine A
2A receptor distribution patterns is
essential in comprehending the functions of the basal gan-glia. To further characterize the anatomical location of ade-nosine A
2A receptors in basal ganglia circuitry, in the current
study we used both single-labeling and double-labeling immunohistochemical methods to localize adenosine A
2A
receptors on striatal neurons on paraffin embedded sections of rat brain. First, we visualized the distribution of A
2A ade-
nosine receptors and incorporated the use of antigen retrieval (AR) to see whether we could increase the sensitivity of the antibody cloned by Rosen and coworkers (5).
Once we optimized the adenosine A 2A
receptor antibody, we then incorporated the method to a double-labeling tech-nique by using a DA marker, anti-tyrosine hydroxylase (TH), and anti-adenosine A
2A receptor. This method was used to
locate where dopaminergic fibers and A 2A
adenosine receptors overlap by double labeling the A
2A receptors with TH. We then
applied the same protocol of double-labeling with enkephalin and adenosine A
2A receptors markers to demonstrate that
enkephalin-containing neurons also contain adenosine A 2A
receptors. Furthermore, we incorporated double-labeling with substance P and adenosine A
2A receptor to demonstrate that
substance P-containing neurons did not contain adenosine A 2A
receptors.
After we optimized the sensitivity of the adenosine A 2A
antibody and developed a double labeling protocol, we used the method for free-floating cryostat sections that were injected with retrograde tracers. This was important to sub-stantiate the behavioral data found by Mingote and cowork-ers (10). In the last part of the study, we collaborated with the UK Parkinson’s Tissue Bank to determine whether the pat-terns of adenosine A
2A receptor immunoreactivity and recep-
tor specificity found in rat basal ganglia are similar to those seen in human basal ganglia. The optimization of markers, such as adenosine A
2A receptors, will enhance studies that
seek to comprehend the dysfunctions that can result from alterations in striatal neurochemistry, not only in Parkinson’s disease, but also for effort-related dysfunctions, such as anergia, fatigue, or psychomotor slowing in depression and other psychiatric or neurological disorders (11).
Materials and Methods Animal protocols were approved by the University of
Connecticut Institutional Animal Care and Use Committee, and the studies were conducted according to the United States NIH guidelines for animal care and use.
Adenosine A 2A
Receptor Visualization and Double-Labeling on Paraffin Sections
Rat brains were perfused with 4.0% paraformaldehyde and fixed in formalin for 2 days. The brains were cut into three coronal sections, ranging from 3.0 mm to 5.0 mm in thickness, and placed into tissue processing cassettes. The brain sections were rinsed in water for 10 min, processed with a series of dehydrating ethyl alcohols solutions, and then cleared in n-butyl alcohol. The processing was com-pleted by infiltrating the tissue in paraffin and embedding each section in a square mold. The sections were then cut on a Leitz Microtome at 7–10 μm, placed on ‘plus’ glass slides (Fisher Scientific, Pittsburgh, PA), and allowed to dry for 3–4 days. Deparaffinization was completed by heating the slides in a 60°C oven and removing excess wax with Citrosolv (Fisher Scientific). The slides were rehydrated by using two changes of 100% ethyl alcohol followed by two changes of 95% ethyl alcohol and one change of 70% ethyl alcohol. Endogenous peroxidase activity was quenched by placing the slides into 3.0% hydrogen peroxide for 5 min followed by a rinse in tap water. Some of the slides were immersed in preheated AR, pH 6.0 (DAKO, Carpinteria, CA), for 20 min and then cooled with cold running tap water.
After three rinses in phosphate-buffered saline (PBS), the primary antibody monoclonal A
2A receptor (1:1000;
Millipore, Temecula, CA) was applied and incubated over-night. The slides were rinsed in PBS buffer and incubated in the secondary antibody anti-mouse/horseradish peroxidase (HRP; Envision, DAKO) for 1 h. After three rinses in PBS buffer, the reactions were visualized with a purple chromo-gen (VIP Substrate Kit; Vector Laboratories, Burlingame, CA) for 10 min. Once the ideal conditions were determined for the A
2A receptor visualization, a few of the slides were
chosen for double-labeling with TH. These slides were sub-jected to another solution of preheated AR, pH 6.0 (DAKO), for 20 min and then cooled with cold running tap water.

The Journal of Histotechnology / Vol. 33, No. 1 / March 2010 43
After three rinses in PBS, the primary antibodies, monoclo-nal anti-tyrosine hydroxylase (1:500; Vision BioSystems, New Castle, UK) was applied and incubated overnight. The slides were rinsed in PBS buffer and incubated in the sec-ondary antibody anti-mouse/HRP (Envision; DAKO) for 1 h. After three rinses in PBS buffer, the reactions were visu-alized with the chromogen 3,3′-diaminobenzidine tetrahy-drochloride (DAB; DAKO) for 10 min. Finally, the sections were dehydrated, cleared in Citrosolv (Fisher Scientific), and coverslipped.
The Bond Max (Vision BioSystems) was used for the dou-ble-labeling of adenosine A
2A receptors and enkephalin or
adenosine A 2A
receptors and substance P. The slides were dewaxed, dehydrated, and subjected to AR treatment 1 (Vision BioSystems). The primary antibody monoclonal A
2A
receptor (1:1000; Millipore) was applied and incubated for 6 h. The slides were rinsed in PBS buffer and incubated in the secondary antibody anti-mouse/HRP (Vision BioSystems) for 1 h. After three rinses in PBS buffer, the reactions were visualized with the red chromogen (Bond Polymer Refine Red Detection; Vision BioSystems) for 10 min. Once the ideal conditions were determined for the A
2A receptor visual-
ization, a few of the slides were chosen for double-labeling with anti-enkephalin or anti-substance P. After three rinses in PBS, the primary antibodies, polyclonal enkephalin (1:1000; Immunostar, Hudson, WI), or polyclonal substance P (1:1000; AbD Serotec, Raleigh, NC), were applied and incubated for 6 h. The slides were rinsed in PBS buffer and incubated in the secondary antibody anti-rabbit/HRP (Vision BioSystems) for 1 h. After three rinses in PBS buffer, the reactions were visualized with the chromogen DAB (Vision BioSystem) for 10 min. Finally, the sections were dehydrated, cleared in Citrosolv (Fisher Scientific), and coverslipped.
Retrograde Labeling Once the A
2A receptors on the striatal neurons were visu-
alized, the pallidal projection zones for these neurons were investigated. Retrograde transport of cholera toxin B (CTB) allowed the characterization of neurons that project from one brain region to another. In the second groups of studies, CTB was injected into different pallidal regions (e.g., globus pallidus vs. ventral pallidum) to determine the presence of retrograde tracer in different striatal subregions (e.g., neo-striatum vs. nucleus accumbens). In the third part of this study the investigation focused on whether the CTB-labeled striatal neurons also contained adenosine A
2A receptor immu-
noreactivity. For the A 2A
receptor labeling, an AR agent was incorporated to decrease the expression of nonspecific tissue binding and to amplify antigen sensitivity.
CTB Visualization on Floating Sections Rat brains were perfused with 4.0% paraformaldehyde
and fixed in formalin for 2 days. The brains were cut into coronal sections 40 μm thick with a vibratome (Lietz). Transported CTB was visualized with a standard immunocy-tochemistry procedure modified for the detection of CTB in free-floating sections. Endogenous peroxidase activity was blocked with a 20-min incubation in 0.5% hydrogen perox-ide + 1% methanol in PBS at room temperature on a shaker table. Sections were rinsed (3×) in PBS, permeabilized and blocked in 0.2% Triton +10% normal goat serum (Vector Laboratories) for 1 h in PBS. Sections were incubated in the
primary antibody rabbit anti-CTB (1:5000; Vector Laboratories) for 40 h at 4°C. After the rinses (3 × 10 min) tissue was incubated in the secondary antibody, biotinylated goat-anti-rabbit (1:200; Vector Laboratories) for 2 h at room temperature. Tissue was again rinsed (3×, 10 min) and incu-bated in PBS, Triton, avidin-biotin complex solution (ABC; Vector Laboratories) for 1 h at room temperature. After this rinse (3 × 10 min), tissue sections were incubated in a buffer, H
2 O
2 , DAB solution with nickel (Vector Laboratories) for 2
to 10 min at room temperature.
Double-Labeling on Floating Sections Endogenous peroxidase activity was quenched by placing
the sections into 3.0% hydrogen peroxide for 5 min followed by 3 rinses in PBS. The sections were then mounted on plus slides and allowed to dry for 10 min. All of the slides were immersed in preheated AR, pH 6.0 (DAKO), for 20 min and then cooled with cold running tap water. After three rinses in PBS the primary antibody monoclonal anti-A
2A (1:000;
Millipore) was applied to the sections and incubated over-night. The slides were rinsed in PBS buffer (3 × 5 min) and incubated in the secondary antibody anti-mouse/HRP (Envision; DAKO) for 1 h. After three rinses in PBS buffer, the reactions were visualized with the purple chromogen (VIP Substrate Kit; Vector Laboratories) for 10 min. Finally, the sections were allowed to dry, placed into Citrosolv (Fisher Scientific), and coverslipped.
Adenosine A2A
Receptor Visualization on Human Tissue Tissue samples were supplied by the Parkinson’s Disease
Society Tissue Bank at Imperial College London, funded by the Parkinson’s Disease Society of the United Kingdom (charity registered in England and Wales No. 258,197 and in Scotland No. SCO37554), with approval from the National Research Ethics Committee (07/MRE09/72). Human brains were fixed in 4% formalin for 5 weeks, then different ana-tomical positions were selected and processed with a Leica Tissue Processor. The blocks were sectioned on a Leica Microtome with the mega-chuck at 8 μm and mounted on ‘plus’ glass slides (Newcomber Supply, Middleton, WI), and allowed to dry for 3–4 days. After routine deparaffinization and hydration to water, endogenous peroxidase activity was quenched by placing the slides into 3.0% hydrogen peroxide. The immunohistochemical procedures varied in pretreatment (with AR and without AR); some of the slides were immersed in preheated AR pH 6.0 (DAKO) for 30 min and then cooled with cold running tap water. After the slides were blocked in 10% normal horse serum (Vector Laboratories) for 20 min, the primary monoclonal adenosine A
2A antibody was applied
(1:1000; Millipore), and the slides were incubated overnight. This step was followed by exposure to the secondary anti-body biotinylated horse-anti-mouse for 1 h (1:100; Vector Laboratories) and then avidin-biotin complex for 1 h (1:200, ABC; Vector Laboratories). The reactions were visualized with a purple chromogen (VIP Substrate Kit; Vector Lab-oratories) for 10 min. Finally, the sections were dehydrated, cleared in xylene, and coverslipped.
Western Blot Analysis Two samples of frozen homogenate from control rat brains
were thawed and solubilized in sodium dodecyl sulfate sam-ple buffer. Human striatum samples from two different donors, which had been snap-frozen in isopentane, were

IHC of Basal Ganglia Adenosine A2A Receptors / Vontell et al.44
solubilized in sodium dodecyl sulfate sample buffer. The cells were measured by a Thermo spectrophotometer and then loaded into a 12% Precise Protein Gel (Thermo-Fisher Scientific, Leicestershire, UK) with 50 μg/mL in each lane. The proteins were transferred electrophoretically with Tris-HEPES running buffer (Thermo-Fisher Scientific) at a con-stant current of 80 V/263 mA for 90 min to a Western polyvinylidene fluoride membrane (Fisher Scientific). Nonspecific antibody binding was blocked by incubating the blot in 5% milk + 0.1% Tween made in PBS. Blots were incubated for 2 h with monoclonal adenosine A
2A antibody
(1:1000; Millipore) diluted with PBS that contained 10% fetal calf serum (Sigma-Aldrich, York, UK). After incuba-tion with the primary antibody the membrane was washed in a series of PBS rinses (5 × 5 min) and blocked again with 5% milk + 0.1% Tween made in PBS. After the blocking step, the membrane was then incubated for 1 h with biotinylated horse anti-mouse IgG (1:3000, Vector). After washing in PBS, the blot was visualized by Femto Maximum Sensitivity Substrate Kit (Thermo-Fisher Scientific).
Results Patterns of adenosine A
2A receptor immunoreactivity in
the striatum and pallidum with use of the monoclonal anti A
2A was similar to those described by Rosin and coworkers
(4 ). The distribution of A 2A
receptors clearly showed a con-sistent pattern of labeling in the striatum, nucleus accum-bens core and shell, and the olfactory tubercles. The quality of the morphology of the tissue sections was not disturbed by the use of AR, and specific staining was increased substan-tially as the nonspecific staining in the cortex was not pres-ent ( Figure 1 ). Double-labeling A
2A and TH demonstrated
the overlapping of the adenosine and dopamine receptors ( Figure 2 ). The sensitivity of the A
2A adenosine antibody was
apparent because there were areas that were specific for A 2A
but not TH and vice versa. Specificity of the antibody was observed by double-labeling adenosine A
2A receptors with
antienkephalin or adenosine A 2A
receptors with substance P. Double-labeling with anti-adenosine A
2A receptor and
antienkephalin identified a population of neurons that con-tain enkephalin and have adenosine A
2A receptors on the
cell membrane, whereas, the double-labeling of the ade-nosine A
2A receptor antibody and the anti-substance P anti-
body demonstrated cell bodies of substance P-containing
neurons that were not outlined with A 2A
receptors ( Figure 3 ). Injections of CTB into the ventral pallidum labeled the ven-tral striatal region near the anterior commissure, including nucleus accumbens core and adjacent anteroventromedial portions of the neostriatum . In contrast, injections of CTB into the globus pallidus dorsal to the ventral pallidum labeled cells in the neostriatum dorsal to the nucleus accum-bens ( Figure 4 ).
Enhanced visibility of cellular detail and morphology was observed without damaging the floating sections with the use of AR. AR increased the signal-to-noise ratio by releasing the aldehyde-protein linkage, and reduced nonspecific stain-ing ( Figure 5 ). Although AR was applied to vibratome sec-tions that were stored in phosphate buffer, there was no evidence of tearing or fraying of the tissue section. Drying the sections after the peroxide quench did not appear to affect the morphology of the tissue because cell bodies appeared intact without excessive shrinking attributable to dehydra-tion. Problems did arise when taking the slides out of AR because sometimes the sections needed to be remounted onto the slide. Some nonspecific staining was observed in the cor-tex, perhaps because of the thickness of the section or excess biotin from the visualization of the CTB retrograde tracer. Double-labeling studies indicated that striatopallidal neurons labeled with CTB also displayed adenosine A
2A receptor
Figure 1. Patterns of the adenosine A 2A
receptors in the striatum are seen using an immunohistochemical reaction of anti-adenosine A
2A receptor
antibody with and without antigen retrieval pretreatment (4×) on paraffin sections. In A, antigen retrieval pretreatment was not applied and the stain-ing is faint with non-specific staining in the cortex. Whereas, in B, the tissue was exposed to antigen retrieval and the distribution of the A
2A receptors is
clearly seen.
Figure 2. Immunohistochemical staining of adenosine A 2A
receptors and dopamine receptors using anti-adenosine A
2A (C) and anti-tyrosine hydrox-
ylase (A) taken at 1× on paraffin sections. The same images taken at 4× to show the diffuse labeling in the striatum, nucleus accumbens core and shell, along with the olfactory tubercles of the dopamine receptors (B, brown). Double labeling the two receptors is seen in images D and E (1× and 20×), the red-brown coloring shows the overlapping of the two receptors (black arrows), whereas, the purple coloring signifies areas that may have only adenosine A
2A receptors (blue arrow) and the red arrow indicates that there
may be areas that have only dopamine receptors.

The Journal of Histotechnology / Vol. 33, No. 1 / March 2010 45
immunoreactivity. Greater magnification of the double-labeled cells revealed, in the striatial regions, nicely identi-fied CTB cell bodies with A
2A receptors on the cell membrane
( Figure 6 ). The section of the ventral striatum, including the nucleus accumbens core, showed immunoreactivity of anti A
2A and the retrograde tracer CTB, which was injected into
the ventral pallidum ( Figure 7 ). This finding is consistent with the hypothesis that both ventral and dorsal striatopal-lidal neurons can also express adenosine A
2A receptors.
The same protocol was applied to paraffin-embedded mega-block human samples containing caudate, putamen, ventral pallidum, and globus pallidus. Intense specific immu-noreactivity of the adenosine A
2A receptors was found in the
sections that were pretreated with AR whereas the sections that did not receive any pretreatment had faint to no immu-noreactivity ( Figure 8 ). Nuclear detail seemed to be drasti-cally improved with the AR pretreatment, as the visualization of cell bodies that express adenosine A
2A receptors (i.e., the
same cell that has DA D2 receptors and is enkephalin posi-tive) could be distinguished from those that lack adenosine A
2A receptors (i.e., the same cell that has DA D1 receptors
and is substance P positive; Figure 8C ).
Figure 3. Double-labeling of adenosine A 2A
receptors with either enkepha-lin-containing neurons (A and B) or substance P-containing neurons (C and D). In A (2×), the distribution of A
2A receptors are stained using anti-
adenosine A 2A
receptor antibody, colorized with Novo Red (red) and the enkephalin-containing neurons are detected with anti-enkephalin using DAB chromogen (brown). B is a higher magnification (20×) of A; black arrows point to enkephalin-containing neurons, which also contain A
2A
receptors. In C, the adenosine A 2A
receptors are detected with NovoRed (red), whereas, the substance P-containing neurons are visualized with DAB (1X). Black arrows point to cell bodies of substance P-containing neu-rons, which are not outlined with A
2A receptors in D (40×).
Figure 4. The striatal subregions (imaged at 4×) are shown after the retro-grade tracer CTB is injected into the globus pallidus (A, GP). The neostria-tum dorsal to the nucleus accumbens (Nac) is labeled after the CTB is injected into the globus pallidus (B).
Figure 5. An immunohistochemical reaction of anti-adenosine A 2A
anti-body in the striatum region from vibratome sections with (B) and without (A) antigen retrieval pretreatment, imaged at 20×. In B, specific staining was increased using antigen retrieval.
Figure 6. The striatopallidal neurons were first labelled with the retrograde tracer cholera toxin B (CTB, brown) and then labeled with adenosine A
2A
receptor antibody (purple). In A, the section of the striatum dorsal to the nucleus accumbens shows the immunoreactivity of the anti-A
2A and the ret-
rograde tracer CTB that was injected into the globus pallidus (2×). Image B is the same section as A magnified at 40×, arrows are pointing to retro-gradely labeled with CTB double-labeled with A
2A receptors.
Figure 7. A section of the ventral striatum showing the immunoreactivity of anti- A
2A and the retrograde tracer CTB that was injected into the ventral
pallidum (2×). The nucleus accumbens core is circled. Image B is the mag-nified image at 40×, arrows are pointing to retrogradely labeled with CTB, double-labeled with A
2A receptors. Reproduced with permission (10).
Copyright © Society for Neuroscience.
IHC of Basal Ganglia Adenosine A2A Receptors / Vontell et al.46
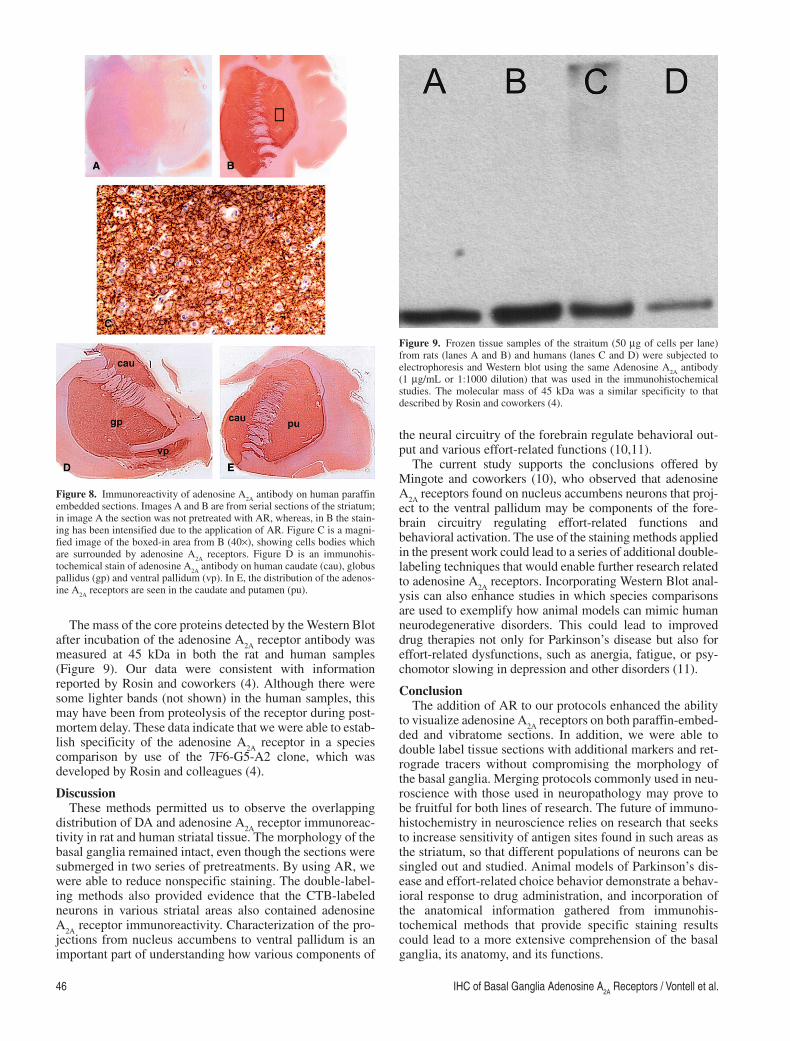
The mass of the core proteins detected by the Western Blot after incubation of the adenosine A
2A receptor antibody was
measured at 45 kDa in both the rat and human samples (Figure 9 ). Our data were consistent with information reported by Rosin and coworkers (4 ). Although there were some lighter bands (not shown) in the human samples, this may have been from proteolysis of the receptor during post-mortem delay. These data indicate that we were able to estab-lish specificity of the adenosine A
2A receptor in a species
comparison by use of the 7F6-G5-A2 clone, which was developed by Rosin and colleagues (4).
Discussion These methods permitted us to observe the overlapping
distribution of DA and adenosine A 2A
receptor immunoreac-tivity in rat and human striatal tissue. The morphology of the basal ganglia remained intact, even though the sections were submerged in two series of pretreatments. By using AR, we were able to reduce nonspecific staining. The double-label-ing methods also provided evidence that the CTB-labeled neurons in various striatal areas also contained adenosine A
2A receptor immunoreactivity. Characterization of the pro-
jections from nucleus accumbens to ventral pallidum is an important part of understanding how various components of
the neural circuitry of the forebrain regulate behavioral out-put and various effort-related functions (10,11).
The current study supports the conclusions offered by Mingote and coworkers (10), who observed that adenosine A
2A receptors found on nucleus accumbens neurons that proj-
ect to the ventral pallidum may be components of the fore-brain circuitry regulating effort-related functions and behavioral activation. The use of the staining methods applied in the present work could lead to a series of additional double-labeling techniques that would enable further research related to adenosine A
2A receptors. Incorporating Western Blot anal-
ysis can also enhance studies in which species comparisons are used to exemplify how animal models can mimic human neurodegenerative disorders. This could lead to improved drug therapies not only for Parkinson’s disease but also for effort-related dysfunctions, such as anergia, fatigue, or psy-chomotor slowing in depression and other disorders (11).
Conclusion The addition of AR to our protocols enhanced the ability
to visualize adenosine A 2A
receptors on both paraffin-embed-ded and vibratome sections. In addition, we were able to double label tissue sections with additional markers and ret-rograde tracers without compromising the morphology of the basal ganglia. Merging protocols commonly used in neu-roscience with those used in neuropathology may prove to be fruitful for both lines of research. The future of immuno-histochemistry in neuroscience relies on research that seeks to increase sensitivity of antigen sites found in such areas as the striatum, so that different populations of neurons can be singled out and studied. Animal models of Parkinson’s dis-ease and effort-related choice behavior demonstrate a behav-ioral response to drug administration, and incorporation of the anatomical information gathered from immunohis-tochemical methods that provide specific staining results could lead to a more extensive comprehension of the basal ganglia, its anatomy, and its functions.
Figure 9. Frozen tissue samples of the straitum (50 μg of cells per lane) from rats (lanes A and B) and humans (lanes C and D) were subjected to electrophoresis and Western blot using the same Adenosine A
2A antibody
(1 μg/mL or 1:1000 dilution) that was used in the immunohistochemical studies. The molecular mass of 45 kDa was a similar specificity to that described by Rosin and coworkers (4).
Figure 8. Immunoreactivity of adenosine A 2A
antibody on human paraffin embedded sections. Images A and B are from serial sections of the striatum; in image A the section was not pretreated with AR, whereas, in B the stain-ing has been intensified due to the application of AR. Figure C is a magni-fied image of the boxed-in area from B (40×), showing cells bodies which are surrounded by adenosine A
2A receptors. Figure D is an immunohis-
tochemical stain of adenosine A 2A
antibody on human caudate (cau), globus pallidus (gp) and ventral pallidum (vp). In E, the distribution of the adenos-ine A
2A receptors are seen in the caudate and putamen (pu).

The Journal of Histotechnology / Vol. 33, No. 1 / March 2010 47
Acknowledgments The authors express their deepest appreciation to the
donors and their families for donating human brain tissue for research. This work was supported by a grant from the National Institute of Mental Health (MH078023) and by the UK Parkinson’s Disease Society Tissue Bank.
References 1. Nestler EJ, Hyman SE, Malenka RC: Molecular Neurophar-
macology. A Foundation for Clinical Neuroscience . San Francisco, CA: McGraw-Hill Company; 2001, pp. 3, 4, 134, 376−377.
2. Kase H, Richardson PJ, Jenner P: Adenosine receptors and Parkinson’s disease. San Diego, CA: Academic Press; 2000, pp. 5−13, 211−214.
3. Ferre S: Adenosine-dopamine in the ventral striatum. Implications for the treatment of schizophrenia. Psychophar-macology (Berl) 133:107–120, 1997.
4. Rosin DL, Robeva A, Woodward RL, Guyenet PC, Linden J: Immunohistochemical localization of adenosine A2A recep-tors in the rat central nervous system. J Comp Neurol 401:163–186, 1998.
5. Rosin DL, Hettinger BD, Lee A, Linden J: Anatomy of A2A receptors in brain: Morphological substrates for integration of striatal function. Neurology 61:12–18, 2003.
6. Correa M, Wisniecki A, Betz A, Dobson DR, O’Neill MF, O’Neill MJ, et al.: The adenosine A2A antagonist KF17837 reverses the locomotor suppression and tremulous jaw move-ments induced by haloperidol in rats: Possible relevance to parkinsonism. Behav Brain Res 148:47–54, 2004.
7. Rascol O, Payoux P, Fabienne O, Ferreira J, Brefel-Courbon C, Monstruc JL: Limitations of current Parkinson’s therapy. Ann Neurol 53:3–15, 2003.
8. Shimada J, Suzuki F: Medicinal Chemistry of Adenosine Receptors in Brain and Periphery. Adenosine Receptors and Parkinson’s Disease . San Diego, CA: Academic Press; 2000, pp. 31–35.
9. Ishiwari K, Madson LJ, Farrar AM, Mingote SM, Valenta JP, DiGianvittorio MD, et al.: Injections of the selective adenosine A2A antagonist MSX-3 into the nucleus accumbens core atten-uate the locomotor suppression induced by haloperidol in rats. Behav Brain Res 178:190–199, 2007.
10. Mingote S, Font L, Farrar AM, Vontell R, Worden L, Stopper CM, et al.: Adenosine receptors regulate exertion of effort through actions on the ventral striatopallidal pathway. J Neurosci 28:9037–9046, 2008.
11. Salamone JD, Correa M, Farrar A, Mingote SM: Effort-related functions of nucleus accumbens dopamine and associated fore-brain circuits. Psychopharmacology (Berl) 191:461–482, 2007.
Related Documents











