Psychopharmacology (2004) 172:341–351 DOI 10.1007/s00213-003-1663-z ORIGINAL INVESTIGATION Maria Antonietta Casu · Carla Pisu · Carla Lobina · Luca Pani Immunocytochemical study of the forebrain serotonergic innervation in Sardinian alcohol-preferring rats Received: 3 July 2003 / Accepted: 15 September 2003 / Published online: 21 November 2003 # Springer-Verlag 2003 Abstract Rationale: The anxiolytic effect of ethanol is generally considered to be causally related to the devel- opment of alcohol dependence, and serotonin (5-HT) has been involved in both alcohol abuse and anxiety disor- ders. Several lines of evidence suggest an inverse relationship between alcohol abuse and central serotoner- gic neurotransmission. Objectives: When tested in the elevated plus-maze, selectively bred Sardinian alcohol- preferring (sP) rats display a higher degree of anxiety than Sardinian alcohol-non-preferring rats (sNP); this behavior is reversed by voluntary ethanol intake. The present study examined whether sP rats differed with respect to the 5- HT innervation in different forebrain areas. Methods: We performed an immunohistochemistry study using an antibody raised against serotonin transporter (SERT), a marker for 5-HT fibers, coupled with an unbiased stereology, the method used to count the number of 5- HT neurons in the raphe nuclei. Results: The SERT- positive innervation density was found to be significantly lower in the medial-prefrontal cortex and in the shell of the nucleus accumbens of the ethanol-naive sP rats (sP-N) when compared with the sNP and unselected Wistar rats. No differences were found in the caudate putamen and hippocampus. The stereological analysis showed a sig- nificant difference in the number of 5-HT neurons in the dorsal but not in the median raphe of sP-N rats, compared with sNP and Wistar rats. Analysis of the cell body cross- sectional area revealed no differences among the three lines of rats either in the dorsal or in the median raphe. In sP rats that had voluntarily drunk ethanol for 14 consecutive days (sP-exp), no differences were found in the 5-HT innervation relative to sP-N animals. Conclu- sions: These results indicate a selective reduction of innervation in the medial portion of the mesocorticolim- bic 5-HT system in sP rats, suggesting that this geneti- cally determined difference may be involved in the contrasting alcohol preference and consumption of sP and sNP animals. Keywords 5-HT · Animal model · Alcohol · Immunohistochemistry Introduction Anxiety disorders (e.g. panic attacks, social phobia, generalized anxiety, etc.) are generally considered to be involved in the development of dependence on—and relapse into—alcohol abuse. This may partly be due to the anxiolytic effect of ethanol described in both men and animals (Prunell et al. 1994; LaBuda and Fuchs 2000) and has led to the hypothesis that anxiety may constitute a critical factor in the etiology of alcohol drinking. The fact that anxiety may trigger an abnormal alcohol intake has been strengthened by findings on genetically selected alcohol-preferring rats, showing their greater innate degree of anxiety relative to alcohol-non-preferring rats. Genetically selected alcohol-preferring rats represent an informative animal model for the study of the neuro- chemical and behavioral correlates of ethanol abuse. Several alcohol-preferring and non-preferring rat lines are available worldwide, including the Sardinian alcohol- preferring (sP) and Sardinian alcohol-non-preferring (snP) rats (Colombo 1997). The daily ethanol intake of a sP rat is over 6 g/kg, the preference ratio for ethanol to water reaching 80–100%, while sNP rats avoid drinking ethanol solution (Colombo et al. 1995). Furthermore, when tested in the elevated plus-maze, sP rats display a higher degree of anxiety than sNP rats, this behavior being reversed by the voluntary intake of 10% ethanol solution (Colombo et al. 1995). Among other neurotransmitters, serotonin (5-HT) has been found to be involved in both alcohol abuse and M. A. Casu ( ) ) · C. Pisu · C. Lobina · L. Pani Neuroscienze S.C.A.R.L, Via Palabanda 9, 09125 Cagliari, Italy e-mail: [email protected] Tel.: +39-070-2548079 Fax: +39-070-254275 L. Pani Institute of Neurogenetic and Neuropharmacology, C.N.R., Cagliari, Italy

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Psychopharmacology (2004) 172:341–351DOI 10.1007/s00213-003-1663-z
O R I G I N A L I N V E S T I G A T I O N
Maria Antonietta Casu · Carla Pisu · Carla Lobina ·Luca Pani
Immunocytochemical study of the forebrain serotonergic innervationin Sardinian alcohol-preferring rats
Received: 3 July 2003 / Accepted: 15 September 2003 / Published online: 21 November 2003� Springer-Verlag 2003
Abstract Rationale: The anxiolytic effect of ethanol isgenerally considered to be causally related to the devel-opment of alcohol dependence, and serotonin (5-HT) hasbeen involved in both alcohol abuse and anxiety disor-ders. Several lines of evidence suggest an inverserelationship between alcohol abuse and central serotoner-gic neurotransmission. Objectives: When tested in theelevated plus-maze, selectively bred Sardinian alcohol-preferring (sP) rats display a higher degree of anxiety thanSardinian alcohol-non-preferring rats (sNP); this behavioris reversed by voluntary ethanol intake. The present studyexamined whether sP rats differed with respect to the 5-HT innervation in different forebrain areas. Methods: Weperformed an immunohistochemistry study using anantibody raised against serotonin transporter (SERT), amarker for 5-HT fibers, coupled with an unbiasedstereology, the method used to count the number of 5-HT neurons in the raphe nuclei. Results: The SERT-positive innervation density was found to be significantlylower in the medial-prefrontal cortex and in the shell ofthe nucleus accumbens of the ethanol-naive sP rats (sP-N)when compared with the sNP and unselected Wistar rats.No differences were found in the caudate putamen andhippocampus. The stereological analysis showed a sig-nificant difference in the number of 5-HT neurons in thedorsal but not in the median raphe of sP-N rats, comparedwith sNP and Wistar rats. Analysis of the cell body cross-sectional area revealed no differences among the threelines of rats either in the dorsal or in the median raphe. InsP rats that had voluntarily drunk ethanol for 14consecutive days (sP-exp), no differences were found inthe 5-HT innervation relative to sP-N animals. Conclu-
sions: These results indicate a selective reduction ofinnervation in the medial portion of the mesocorticolim-bic 5-HT system in sP rats, suggesting that this geneti-cally determined difference may be involved in thecontrasting alcohol preference and consumption of sP andsNP animals.
Keywords 5-HT · Animal model · Alcohol ·Immunohistochemistry
Introduction
Anxiety disorders (e.g. panic attacks, social phobia,generalized anxiety, etc.) are generally considered to beinvolved in the development of dependence on—andrelapse into—alcohol abuse. This may partly be due to theanxiolytic effect of ethanol described in both men andanimals (Prunell et al. 1994; LaBuda and Fuchs 2000) andhas led to the hypothesis that anxiety may constitute acritical factor in the etiology of alcohol drinking. The factthat anxiety may trigger an abnormal alcohol intake hasbeen strengthened by findings on genetically selectedalcohol-preferring rats, showing their greater innatedegree of anxiety relative to alcohol-non-preferring rats.Genetically selected alcohol-preferring rats represent aninformative animal model for the study of the neuro-chemical and behavioral correlates of ethanol abuse.Several alcohol-preferring and non-preferring rat lines areavailable worldwide, including the Sardinian alcohol-preferring (sP) and Sardinian alcohol-non-preferring (snP)rats (Colombo 1997). The daily ethanol intake of a sP ratis over 6 g/kg, the preference ratio for ethanol to waterreaching 80–100%, while sNP rats avoid drinking ethanolsolution (Colombo et al. 1995). Furthermore, when testedin the elevated plus-maze, sP rats display a higher degreeof anxiety than sNP rats, this behavior being reversed bythe voluntary intake of 10% ethanol solution (Colombo etal. 1995).
Among other neurotransmitters, serotonin (5-HT) hasbeen found to be involved in both alcohol abuse and
M. A. Casu ()) · C. Pisu · C. Lobina · L. PaniNeuroscienze S.C.A.R.L,Via Palabanda 9, 09125 Cagliari, Italye-mail: [email protected].: +39-070-2548079Fax: +39-070-254275
L. PaniInstitute of Neurogenetic and Neuropharmacology,C.N.R., Cagliari, Italy

anxiety. There is evidence to suggest an inverse relation-ship between alcohol abuse and central serotonergicneurotransmission. Deficiency in brain 5-HT and itsmetabolites has been reported in human alcohol abusersas well as in genetically selected alcohol-preferring rats(Cloninger 1987; Murphy et al. 1987; Eriksson andHumble 1990; Zhou et al. 1990; McBride et al. 1993).Accordingly, it is well known that ethanol consumptioncan be reduced by drugs that increase central 5-HTavailability by either inducing 5-HT release or byblocking 5-HT uptake (Sellers et al. 1992), or bycompound selective blockers of 5-HT receptor subtypessuch as 5-HT2, 5-HT3, and 5-HT4 (Meert et al. 1990;Panocka and Massi 1992; Panocka et al. 1993, 1995;Lovinger 1999). sP rats have been shown to display fewerbehavioral responses to the stimulation of central 5-HT2Areceptors in comparison with sNP rats (Ciccocioppo et al.1995), suggesting a reduced 5-HT system function in sPrats. For instance, decreased 5-HT and 5-HIAA contentwas found in some brain areas of alcohol-preferring rats(P) (Murphy et al. 1982) when compared with theircorresponding alcohol-non-preferring line (NP) and, morerecently, lower 5-HT levels were reported by Devoto etal. (1998) in cortical but not in mesolimbic areas of sPrats. Altered binding density of 5-HT1A and of 5-HT2Asites and fewer 5-HT immunostained fibers in severalbrain areas of P rats compared with NP rats have beenobserved (McBride et al. 1993, 1994). Finally, lower[3H]ketanserin binding levels in the medial prefrontalcortex of sP rats have also been found (Ciccocioppo et al.1999b).
Thus, the present study was conducted to furtherinvestigate any possible differences in the forebrain 5-HTinnervation between sP and sNP rats. With this aim, weused immunohistochemistry techniques with an antibodyraised against the 5-HT transporter (SERT) as a markerfor serotonergic fibers, and with an antibody raisedagainst tryptophan hydroxylase (TPH) as a marker forserotonergic neurons in the raphe nuclei, in order toexamine whether sP, sNP and Wistar (W) rats exhibitedsimilar 5-HT innervation in different forebrain structureswhere 5-HT neurons—whose cell bodies are mainlylocated in the raphe area—project. We also intended tostudy whether voluntary and chronic ethanol intake couldmodify this innervation.
Materials and methods
Animals
sP and sNP male rats from the 52nd generation (February 2002) andapproximately 3 months old, and W rats (Charles River, Calco, LC,Italy) were used. Rats were derived from a population of sP andsNP rats that underwent caesarian derivation at Charles River(Lyon, France) for the production of specific pathogen freeindividuals. Animals were housed in groups of five in standardplastic cages with wood-chip bedding. The animal facility wasunder a reversed 12-h/12-h light/dark cycle, at a constant temper-ature of 22+2�C and relative humidity of 60%. Ethanol-naive ratshad free access to tap water while ethanol-experienced rats
consumed ethanol under the two-bottle, free-choice procedure.Two graduated bottles containing water and 10% (v/v) ethanol,respectively, were continuously offered for 14 consecutive days.Ethanol and water intakes were recorded daily at 0800 hours.Bottles were refilled every day with fresh solution, and theirposition interchanged at random to avoid the development ofposition preference. In detail, the daily ethanol intake of sP rats washigher than 6 g/kg, while the daily ethanol intake of sNP rats waslower than 1.0 g/kg (Gessa et al. 1991).
All experimental protocols were approved by the ethicscommittee at the University of Cagliari and performed in strictaccordance with the E.C. regulations for the care and use ofexperimental animals (CEE N� 86/609).
Immunocytochemistry
Ethanol-experienced sP-exp (n=5), ethanol-naive sP (n=5), sNP(n=5) and W (n=5) rats were anaesthetized with equithesin (2.5 mg/kg, i.p.) and perfused transcardially with 4% paraformaldehyde in0.1% phosphate buffer (PB), pH 7.4. The brains were subsequentlypost-fixed in the same fixative for 2 h and cryoprotected overnightwith a solution of 30% sucrose in 0.1 M PB at 4�C. Coronalsections of 40 �m were cut by cryostat (Leica 3050, LeicaMicrosystems, Nussloch, Germany) and processed simultaneouslyto avoid differences in antibody penetration. Free-floating cyto-chemical stainings were performed in strict accordance with thegeneral methodological procedures indicated by Casu et al. (2002).Controls were performed by subtracting the primary antibody in theimmunocytochemical procedure.
Immunostaining for SERT and 5-HT
Briefly, after rinsing in phosphate buffered saline (PB 0.2 M: 50 ml/l; NaCl: 8.8 g/l; KCl 0.2 g/l) with 0.2% Triton X-100 (PBS+T),sections were incubated with 0.3% of H2O2 in PBS and, afterextensive washing, with a blocking solution containing 1% bovineserum albumin (BSA) and 20% normal goat serum in PBS+T toreduce background. Sections were then incubated overnight at 4�Cwith a mouse anti-SERT antibody (1:2000; from Chemicon,Temecula, CA, USA) or a rabbit anti-5-HT (1:2000 from Sigma,Saint Louis, MO, USA). After rinsing, sections were incubated witha goat anti-mouse (for SERT) and a goat anti-rabbit (for 5-HT)biotinylated IgG (1:200; Vector, Burlingame, CA, USA) for 1 h,followed by an avidin-biotin complex (1:200; Vectastain ABC kitfrom Vector, Burlingame, CA, USA) for an additional hour. AfterABC incubation and washing, sections were exposed to 3,3’-diaminobenzidine (0.06% in PBS) containing 1% cobalt chlorideand 1% nickel ammonium sulfate for 15 min. Immunostaining wasdeveloped by adding 5 �l H2O2 (0.1% in PBS) to each 500 �l of3,3’-diaminobenzidine. After washing in PBS+T, all sections weremounted on gelatin-coated glass slides, air-dried, dehydrated inascending concentrations of ethanol, cleared with xylene, andcoverslipped with Entellan. The mounted sections were examinedunder a BX-60 Olympus light-microscope (Olympus Optical Co.GmbH, Hamburg, Germany).
Quantitative analysis
Cingulate cortex
The region of analysis of the cingulate cortex corresponded tobregma +3/+2.5 mm as indicated in the Paxinos and Watson atlas(Paxinos and Watson 1986) and ten alternated sections per animalwere selected for immunostaining. In these sections, the cingulatecortex area includes cingulate cortex area 1 (Cg1), area 3 (Cg3) andinfralimbic cortex according to the Paxinos and Watson terminol-ogy (Paxinos and Watson 1986). To ensure that these cortical areaswere quantified in all samples, measurements were always carriedout along a vertical line approximately at the III, IV cortical layer
342

level, crossing the Cg1, Cg3 and infralimbic cortex. The analysiswas made taking six fields, two for each cortical area. Themicrographs were all taken from the Cg3.
NAcc and caudate-putamen
The region of analysis corresponds to bregma +2.2/+1.5 mm forcore and shell and +1.5/0.5 mm for the caudate-putamen. Theanalysis was carried out by taking ten alternated sections per animaland five randomly chosen fields through the core, shell or caudate-putamen for each section.
Hippocampus
The region of analysis corresponds to bregma �2.8/�4.5 mm. Toensure that equivalent hippocampal regions were quantified in allsamples, measurements were always carried out along a verticalline crossing the hilus just medial to the inner limit of CA3. A totalof ten fields were used to sample the layers of the hippocampalformation.
The immunostaining in all areas was quantified using an imageanalysis system (KS 300; Karl Zeiss Vision GmbH, Hallbergmoos,Germany). The analysis was carried out by measuring thepercentage of the area occupied by SERT or 5-HT positive fiberscompared with a 2000-�m2 standardized area (IR area %), using a�100 objective lens and a Zeiss Sound Vision digital microscopevideo camera (total magnification on computer screen 7500�).Gray values, measured in immunostaining negative areas of thesections, were subtracted as background from the resulting binarypicture (Bjorklund et al. 1997). The analysis procedure was strictlycarried out as a blind study.
Unbiased cell counts of 5-HT neurons in the raphe nuclei
To label neurons for stereology, 40-�m-thick free-floating coronalcryosections were incubated overnight at 4�C in a 1:1000 dilutionof tryptophan hydroxylase (TPH) antibody from Sigma (St. Louis,MO), visualized with a goat anti-mouse biotinylated IgG (1:400;Vector, Burlingame, CA, USA) followed by an avidin-biotincomplex (1:400; Vectastain ABC kit from Vector, Burlingame, CA,USA) and the 3,3’-diaminobenzidine. Immunostaining was devel-oped by adding 5 �l H2O2 (0.1% in PBS) to each 500 �l of 3,3’-diaminobenzidine and counterstained with cresyl violet withoutinterfering with the brownish TPH immunostaining. The sectionthickness after processing ranged from 18 mm to 22 �m. Eachmounted section was numbered following the rostrocaudal levelcorresponding to the rat brain atlas, and every fifth section was usedfor cell number estimation. Dedicated software (Stereologer,System Planning and Analysis, INC., Alexandria, VA) was linkedto drive a motorized stage in three axes (X, Y, Z) on a BX-60Olympus light-microscope (Olympus Optical Co. GmbH, Ham-burg, Germany). The total number of 5-HT neurons in the dorsaland medial raphe of sP, sNP and Wistar rats was estimated bymeans of the Optical Fractionator, which combines the opticaldissector with the fractionator sampling scheme, giving a directestimation of the number of 3-D objects unbiased by their shape,size and orientation (Sterio 1984; Gundersen et al. 1988; Mounton2002). A systematic random sample of neurons was achieved bypositioning a sampling grid over the entire dorsal and medial rapheon each section. This grid was divided into counting frames thatwere equidistant from each other. Movement between coordinateswas effected by a computer-controlled stepping motor attached tothe X and Y axes of the stage. The optimal dimensions of the eachcounting frame and the grid size were established following arestricted pilot study. The sampling fraction was delimited at lowpower and neurons had to have a distinct nucleus in order to becounted. Cells were sampled with a 40� oil-immersion objectivethrough a defined depth with a guard zone of 2 �m. Opticaldissector counting rules were used to count the total TPH-positive
neurons (N) in each dissector volume, which was determined bymultiplying the area of the counting frame by the dissector height.
N ¼X
Q�t
h
1asf
1ssf
where S Q� is the total number of particles counted, t is the meansection thickness, h is the height of the optical dissector, asf is thearea sampling fraction and ssf is the section sampling fraction. Thecoefficient of error (CE) for each estimation and animal rangedfrom 0.05 to 0.1.
Cell body size measurement
The region of analysis corresponding to bregma coordinates �7.04to �9.30 mm was chosen in order to evaluate the cell body size ofneurons in the dorsal and median raphe, known to be populated byserotonergic neurons. For each section (eight sections for each rat),the cell body cross-sectional area (�m2) was determined, using a�40 objective lens (total magnification on computer screen 3000�),only in those neurons where nuclei could be observed. The cellbody cross-sectional area was quantified using an image analysissystem (KS 300; Karl Zeiss Vision GmbH, Hallbergmoos,Germany). An average of 50 cells in the dorsal and 30 in themedian raphe were quantified for each rat.
Statistical analysis
All data were expressed as mean +SEM and analyzed using one-way analysis of variance (ANOVA). When a significant interaction(P<0.05) was found, the Newman-Keuls post-hoc test was used.
Results
Specificity of immunoreactivity
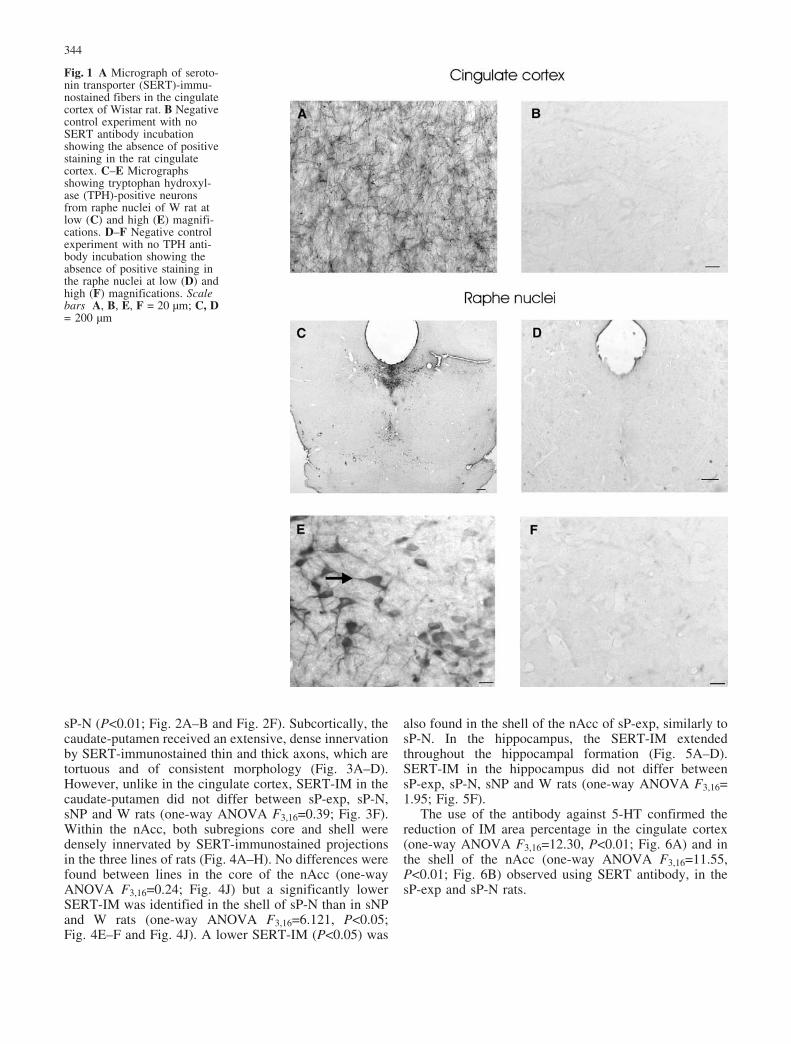
SERT-immunostaining (IM) was densely present in theentire forebrain. Figure 1A shows SERT-IM in thecingulate cortex of W rats. SERT axons extended throughall regions of cerebral cortex. A negative control exper-iment with no SERT antibody incubation showed theexpected absence of positive staining in the cingulatecortex (Fig. 1B).
TPH-immunostained neurons were visualized through-out the entire dorsal and median raphe (Fig. 1C–E). Thenegative control experiment with no TPH antibodyincubation showed the expected absence of positivestaining in the raphe nuclei (Fig. 1D–F).
SERT-IM in the cingulate cortex, nAcc,caudate-putamen and hippocampus
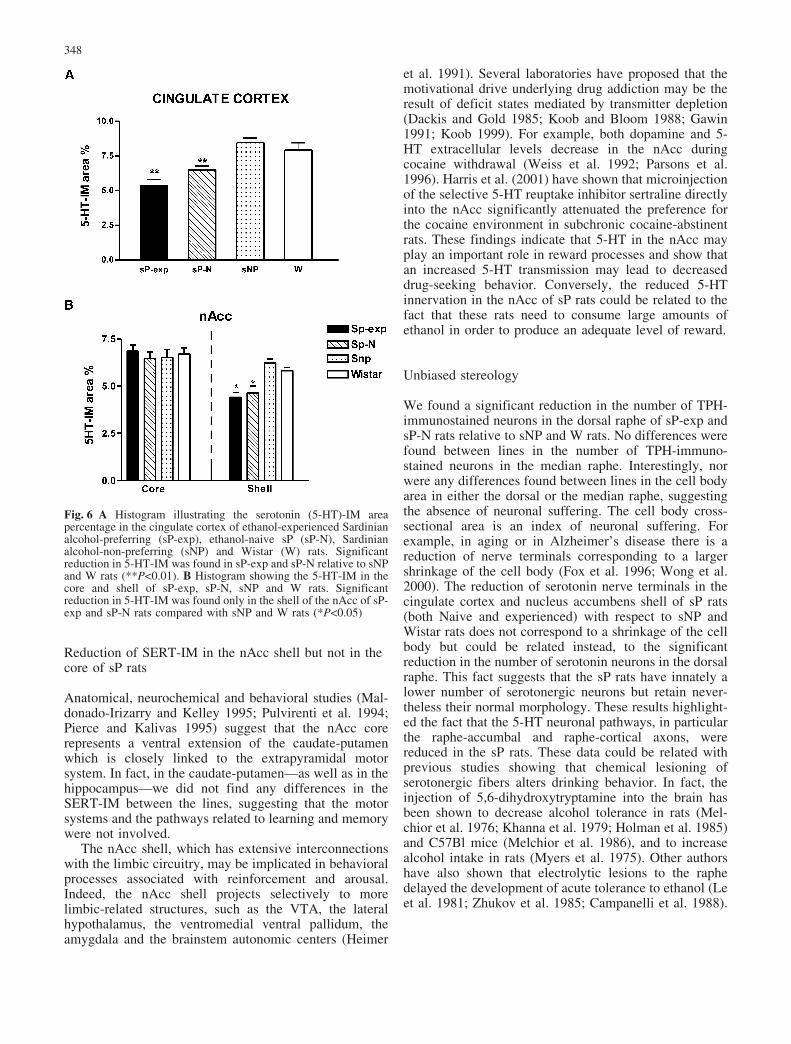
SERT-immunostained thin and thick fibers denselyinnervated all the regions of the cingulate cortex of sP-exp, sP-N, sNP and W rats (Fig. 2A–D). Significantdifferences between the rat lines were identified (one-wayANOVA F 3,16=26.92). The SERT-IM was lower in thecingulate cortex of the sP-N rats than sNP and W rats(P<0.01; Fig. 2F). Moreover, ethanol was unable tochange the SERT-IM in the cingulate cortex. In fact, alower SERT-IM was also found in the sP-exp, similarly to
343
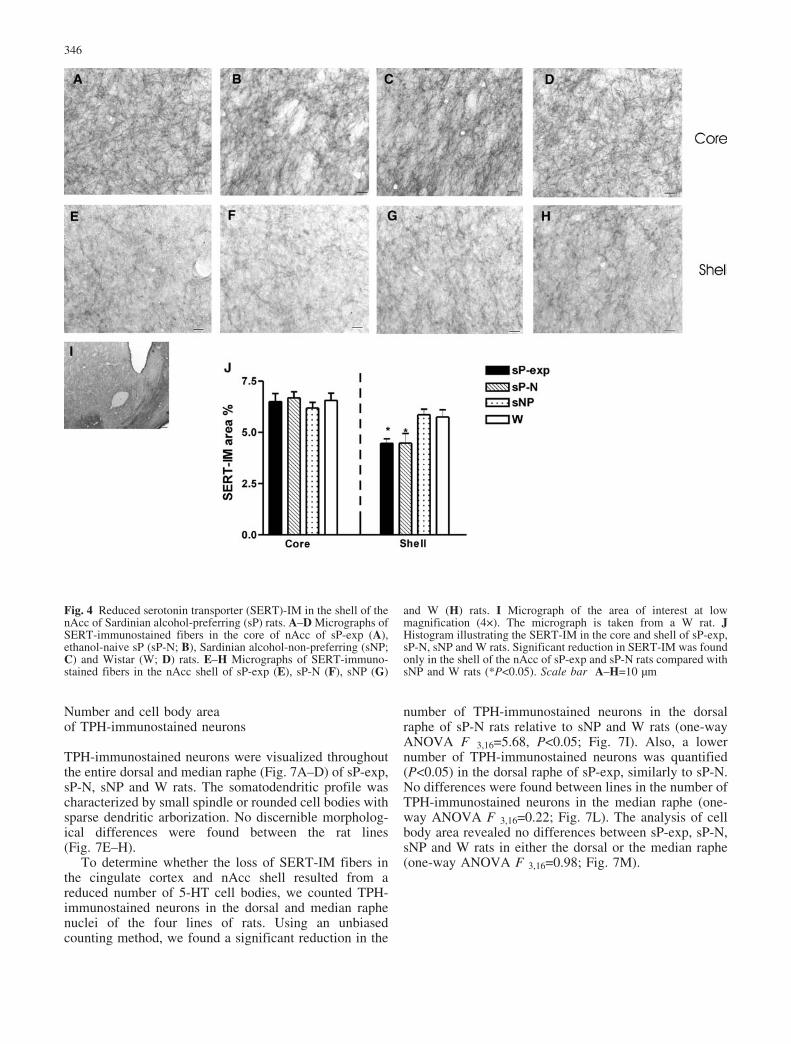
sP-N (P<0.01; Fig. 2A–B and Fig. 2F). Subcortically, thecaudate-putamen received an extensive, dense innervationby SERT-immunostained thin and thick axons, which aretortuous and of consistent morphology (Fig. 3A–D).However, unlike in the cingulate cortex, SERT-IM in thecaudate-putamen did not differ between sP-exp, sP-N,sNP and W rats (one-way ANOVA F3,16=0.39; Fig. 3F).Within the nAcc, both subregions core and shell weredensely innervated by SERT-immunostained projectionsin the three lines of rats (Fig. 4A–H). No differences werefound between lines in the core of the nAcc (one-wayANOVA F3,16=0.24; Fig. 4J) but a significantly lowerSERT-IM was identified in the shell of sP-N than in sNPand W rats (one-way ANOVA F3,16=6.121, P<0.05;Fig. 4E–F and Fig. 4J). A lower SERT-IM (P<0.05) was
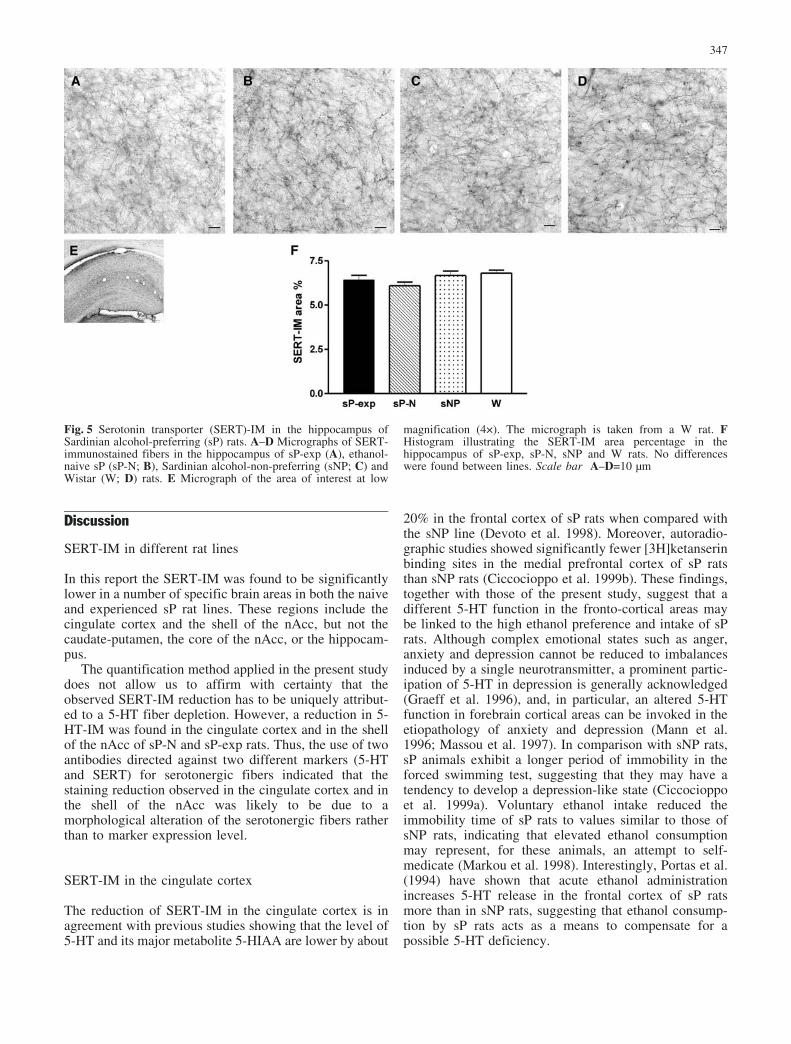
also found in the shell of the nAcc of sP-exp, similarly tosP-N. In the hippocampus, the SERT-IM extendedthroughout the hippocampal formation (Fig. 5A–D).SERT-IM in the hippocampus did not differ betweensP-exp, sP-N, sNP and W rats (one-way ANOVA F3,16=1.95; Fig. 5F).
The use of the antibody against 5-HT confirmed thereduction of IM area percentage in the cingulate cortex(one-way ANOVA F3,16=12.30, P<0.01; Fig. 6A) and inthe shell of the nAcc (one-way ANOVA F3,16=11.55,P<0.01; Fig. 6B) observed using SERT antibody, in thesP-exp and sP-N rats.
Fig. 1 A Micrograph of seroto-nin transporter (SERT)-immu-nostained fibers in the cingulatecortex of Wistar rat. B Negativecontrol experiment with noSERT antibody incubationshowing the absence of positivestaining in the rat cingulatecortex. C–E Micrographsshowing tryptophan hydroxyl-ase (TPH)-positive neuronsfrom raphe nuclei of W rat atlow (C) and high (E) magnifi-cations. D–F Negative controlexperiment with no TPH anti-body incubation showing theabsence of positive staining inthe raphe nuclei at low (D) andhigh (F) magnifications. Scalebars A, B, E, F = 20 �m; C, D= 200 �m
344

Fig. 2 Reduced serotonin transporter (SERT)-immunostaining(IM) in the cingulate cortex of Sardinian alcohol-preferring (sP)rats. A–D Micrographs of SERT-immunostained fibers in thecingulate cortex of sP-exp (A), ethanol-naive sP (sP-N; B),Sardinian alcohol-non-preferring (sNP; C) and Wistar (W; D) rats.E Micrograph of the area of interest at low magnification (4�). The
micrograph is taken from a W rat. F Histogram illustrating theSERT-IM area percentage in the cingulate cortex of sP-exp, sP-N,sNP and W rats. Significant reduction in SERT-IM was found insP-exp and sP-N relative to sNP and W rats (**P<0.01). Scale barA–D=10 �m
Fig. 3 Serotonin transporter (SERT)-IM in the caudate-putamen ofSardinian alcohol-preferring (sP) rats. A–D Micrographs of SERT-immunostained fibers in the caudate-putamen of sP-exp (A),ethanol-naive sP (sP-N; B), Sardinian alcohol-non-preferring (sNP;C) and Wistar (W; D) rats. E Micrograph of the area of interest at
low magnification (4�). The micrograph is taken from a W rat. FHistogram illustrating the SERT-IM area percentage in the caudate-putamen of sP-exp, sP-N, sNP and W rats. No differences werefound between lines. Scale bar A–D=10 �m
345
Number and cell body areaof TPH-immunostained neurons
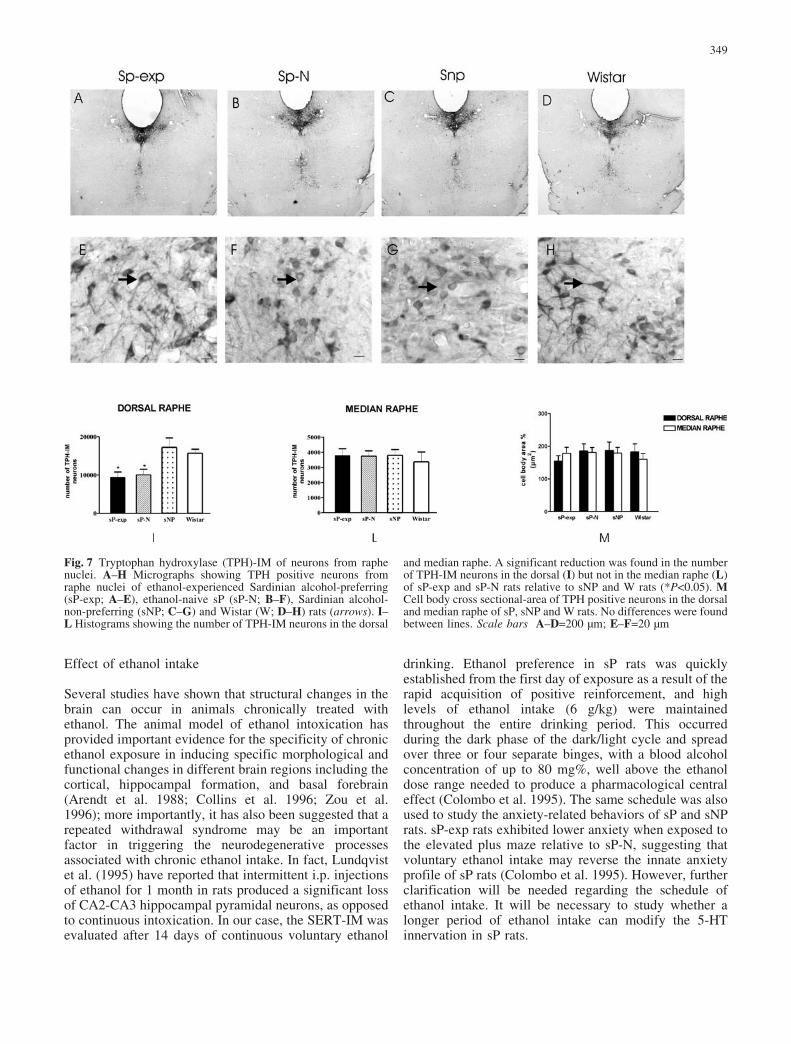
TPH-immunostained neurons were visualized throughoutthe entire dorsal and median raphe (Fig. 7A–D) of sP-exp,sP-N, sNP and W rats. The somatodendritic profile wascharacterized by small spindle or rounded cell bodies withsparse dendritic arborization. No discernible morpholog-ical differences were found between the rat lines(Fig. 7E–H).
To determine whether the loss of SERT-IM fibers inthe cingulate cortex and nAcc shell resulted from areduced number of 5-HT cell bodies, we counted TPH-immunostained neurons in the dorsal and median raphenuclei of the four lines of rats. Using an unbiasedcounting method, we found a significant reduction in the
number of TPH-immunostained neurons in the dorsalraphe of sP-N rats relative to sNP and W rats (one-wayANOVA F 3,16=5.68, P<0.05; Fig. 7I). Also, a lowernumber of TPH-immunostained neurons was quantified(P<0.05) in the dorsal raphe of sP-exp, similarly to sP-N.No differences were found between lines in the number ofTPH-immunostained neurons in the median raphe (one-way ANOVA F 3,16=0.22; Fig. 7L). The analysis of cellbody area revealed no differences between sP-exp, sP-N,sNP and W rats in either the dorsal or the median raphe(one-way ANOVA F 3,16=0.98; Fig. 7M).
Fig. 4 Reduced serotonin transporter (SERT)-IM in the shell of thenAcc of Sardinian alcohol-preferring (sP) rats. A–D Micrographs ofSERT-immunostained fibers in the core of nAcc of sP-exp (A),ethanol-naive sP (sP-N; B), Sardinian alcohol-non-preferring (sNP;C) and Wistar (W; D) rats. E–H Micrographs of SERT-immuno-stained fibers in the nAcc shell of sP-exp (E), sP-N (F), sNP (G)
and W (H) rats. I Micrograph of the area of interest at lowmagnification (4�). The micrograph is taken from a W rat. JHistogram illustrating the SERT-IM in the core and shell of sP-exp,sP-N, sNP and W rats. Significant reduction in SERT-IM was foundonly in the shell of the nAcc of sP-exp and sP-N rats compared withsNP and W rats (*P<0.05). Scale bar A–H=10 �m
346
Discussion
SERT-IM in different rat lines
In this report the SERT-IM was found to be significantlylower in a number of specific brain areas in both the naiveand experienced sP rat lines. These regions include thecingulate cortex and the shell of the nAcc, but not thecaudate-putamen, the core of the nAcc, or the hippocam-pus.
The quantification method applied in the present studydoes not allow us to affirm with certainty that theobserved SERT-IM reduction has to be uniquely attribut-ed to a 5-HT fiber depletion. However, a reduction in 5-HT-IM was found in the cingulate cortex and in the shellof the nAcc of sP-N and sP-exp rats. Thus, the use of twoantibodies directed against two different markers (5-HTand SERT) for serotonergic fibers indicated that thestaining reduction observed in the cingulate cortex and inthe shell of the nAcc was likely to be due to amorphological alteration of the serotonergic fibers ratherthan to marker expression level.
SERT-IM in the cingulate cortex
The reduction of SERT-IM in the cingulate cortex is inagreement with previous studies showing that the level of5-HT and its major metabolite 5-HIAA are lower by about
20% in the frontal cortex of sP rats when compared withthe sNP line (Devoto et al. 1998). Moreover, autoradio-graphic studies showed significantly fewer [3H]ketanserinbinding sites in the medial prefrontal cortex of sP ratsthan sNP rats (Ciccocioppo et al. 1999b). These findings,together with those of the present study, suggest that adifferent 5-HT function in the fronto-cortical areas maybe linked to the high ethanol preference and intake of sPrats. Although complex emotional states such as anger,anxiety and depression cannot be reduced to imbalancesinduced by a single neurotransmitter, a prominent partic-ipation of 5-HT in depression is generally acknowledged(Graeff et al. 1996), and, in particular, an altered 5-HTfunction in forebrain cortical areas can be invoked in theetiopathology of anxiety and depression (Mann et al.1996; Massou et al. 1997). In comparison with sNP rats,sP animals exhibit a longer period of immobility in theforced swimming test, suggesting that they may have atendency to develop a depression-like state (Ciccocioppoet al. 1999a). Voluntary ethanol intake reduced theimmobility time of sP rats to values similar to those ofsNP rats, indicating that elevated ethanol consumptionmay represent, for these animals, an attempt to self-medicate (Markou et al. 1998). Interestingly, Portas et al.(1994) have shown that acute ethanol administrationincreases 5-HT release in the frontal cortex of sP ratsmore than in sNP rats, suggesting that ethanol consump-tion by sP rats acts as a means to compensate for apossible 5-HT deficiency.
Fig. 5 Serotonin transporter (SERT)-IM in the hippocampus ofSardinian alcohol-preferring (sP) rats. A–D Micrographs of SERT-immunostained fibers in the hippocampus of sP-exp (A), ethanol-naive sP (sP-N; B), Sardinian alcohol-non-preferring (sNP; C) andWistar (W; D) rats. E Micrograph of the area of interest at low
magnification (4�). The micrograph is taken from a W rat. FHistogram illustrating the SERT-IM area percentage in thehippocampus of sP-exp, sP-N, sNP and W rats. No differenceswere found between lines. Scale bar A–D=10 �m
347
Reduction of SERT-IM in the nAcc shell but not in thecore of sP rats
Anatomical, neurochemical and behavioral studies (Mal-donado-Irizarry and Kelley 1995; Pulvirenti et al. 1994;Pierce and Kalivas 1995) suggest that the nAcc corerepresents a ventral extension of the caudate-putamenwhich is closely linked to the extrapyramidal motorsystem. In fact, in the caudate-putamen—as well as in thehippocampus—we did not find any differences in theSERT-IM between the lines, suggesting that the motorsystems and the pathways related to learning and memorywere not involved.
The nAcc shell, which has extensive interconnectionswith the limbic circuitry, may be implicated in behavioralprocesses associated with reinforcement and arousal.Indeed, the nAcc shell projects selectively to morelimbic-related structures, such as the VTA, the lateralhypothalamus, the ventromedial ventral pallidum, theamygdala and the brainstem autonomic centers (Heimer
et al. 1991). Several laboratories have proposed that themotivational drive underlying drug addiction may be theresult of deficit states mediated by transmitter depletion(Dackis and Gold 1985; Koob and Bloom 1988; Gawin1991; Koob 1999). For example, both dopamine and 5-HT extracellular levels decrease in the nAcc duringcocaine withdrawal (Weiss et al. 1992; Parsons et al.1996). Harris et al. (2001) have shown that microinjectionof the selective 5-HT reuptake inhibitor sertraline directlyinto the nAcc significantly attenuated the preference forthe cocaine environment in subchronic cocaine-abstinentrats. These findings indicate that 5-HT in the nAcc mayplay an important role in reward processes and show thatan increased 5-HT transmission may lead to decreaseddrug-seeking behavior. Conversely, the reduced 5-HTinnervation in the nAcc of sP rats could be related to thefact that these rats need to consume large amounts ofethanol in order to produce an adequate level of reward.
Unbiased stereology
We found a significant reduction in the number of TPH-immunostained neurons in the dorsal raphe of sP-exp andsP-N rats relative to sNP and W rats. No differences werefound between lines in the number of TPH-immuno-stained neurons in the median raphe. Interestingly, norwere any differences found between lines in the cell bodyarea in either the dorsal or the median raphe, suggestingthe absence of neuronal suffering. The cell body cross-sectional area is an index of neuronal suffering. Forexample, in aging or in Alzheimer’s disease there is areduction of nerve terminals corresponding to a largershrinkage of the cell body (Fox et al. 1996; Wong et al.2000). The reduction of serotonin nerve terminals in thecingulate cortex and nucleus accumbens shell of sP rats(both Naive and experienced) with respect to sNP andWistar rats does not correspond to a shrinkage of the cellbody but could be related instead, to the significantreduction in the number of serotonin neurons in the dorsalraphe. This fact suggests that the sP rats have innately alower number of serotonergic neurons but retain never-theless their normal morphology. These results highlight-ed the fact that the 5-HT neuronal pathways, in particularthe raphe-accumbal and raphe-cortical axons, werereduced in the sP rats. These data could be related withprevious studies showing that chemical lesioning ofserotonergic fibers alters drinking behavior. In fact, theinjection of 5,6-dihydroxytryptamine into the brain hasbeen shown to decrease alcohol tolerance in rats (Mel-chior et al. 1976; Khanna et al. 1979; Holman et al. 1985)and C57Bl mice (Melchior et al. 1986), and to increasealcohol intake in rats (Myers et al. 1975). Other authorshave also shown that electrolytic lesions to the raphedelayed the development of acute tolerance to ethanol (Leet al. 1981; Zhukov et al. 1985; Campanelli et al. 1988).
Fig. 6 A Histogram illustrating the serotonin (5-HT)-IM areapercentage in the cingulate cortex of ethanol-experienced Sardinianalcohol-preferring (sP-exp), ethanol-naive sP (sP-N), Sardinianalcohol-non-preferring (sNP) and Wistar (W) rats. Significantreduction in 5-HT-IM was found in sP-exp and sP-N relative to sNPand W rats (**P<0.01). B Histogram showing the 5-HT-IM in thecore and shell of sP-exp, sP-N, sNP and W rats. Significantreduction in 5-HT-IM was found only in the shell of the nAcc of sP-exp and sP-N rats compared with sNP and W rats (*P<0.05)
348
Effect of ethanol intake
Several studies have shown that structural changes in thebrain can occur in animals chronically treated withethanol. The animal model of ethanol intoxication hasprovided important evidence for the specificity of chronicethanol exposure in inducing specific morphological andfunctional changes in different brain regions including thecortical, hippocampal formation, and basal forebrain(Arendt et al. 1988; Collins et al. 1996; Zou et al.1996); more importantly, it has also been suggested that arepeated withdrawal syndrome may be an importantfactor in triggering the neurodegenerative processesassociated with chronic ethanol intake. In fact, Lundqvistet al. (1995) have reported that intermittent i.p. injectionsof ethanol for 1 month in rats produced a significant lossof CA2-CA3 hippocampal pyramidal neurons, as opposedto continuous intoxication. In our case, the SERT-IM wasevaluated after 14 days of continuous voluntary ethanol
drinking. Ethanol preference in sP rats was quicklyestablished from the first day of exposure as a result of therapid acquisition of positive reinforcement, and highlevels of ethanol intake (6 g/kg) were maintainedthroughout the entire drinking period. This occurredduring the dark phase of the dark/light cycle and spreadover three or four separate binges, with a blood alcoholconcentration of up to 80 mg%, well above the ethanoldose range needed to produce a pharmacological centraleffect (Colombo et al. 1995). The same schedule was alsoused to study the anxiety-related behaviors of sP and sNPrats. sP-exp rats exhibited lower anxiety when exposed tothe elevated plus maze relative to sP-N, suggesting thatvoluntary ethanol intake may reverse the innate anxietyprofile of sP rats (Colombo et al. 1995). However, furtherclarification will be needed regarding the schedule ofethanol intake. It will be necessary to study whether alonger period of ethanol intake can modify the 5-HTinnervation in sP rats.
Fig. 7 Tryptophan hydroxylase (TPH)-IM of neurons from raphenuclei. A–H Micrographs showing TPH positive neurons fromraphe nuclei of ethanol-experienced Sardinian alcohol-preferring(sP-exp; A–E), ethanol-naive sP (sP-N; B–F), Sardinian alcohol-non-preferring (sNP; C–G) and Wistar (W; D–H) rats (arrows). I–L Histograms showing the number of TPH-IM neurons in the dorsal
and median raphe. A significant reduction was found in the numberof TPH-IM neurons in the dorsal (I) but not in the median raphe (L)of sP-exp and sP-N rats relative to sNP and W rats (*P<0.05). MCell body cross sectional-area of TPH positive neurons in the dorsaland median raphe of sP, sNP and W rats. No differences were foundbetween lines. Scale bars A–D=200 �m; E–F=20 �m
349

In conclusion, our observations revealed a causalrelationship between the alteration of the 5-HT systemand alcohol preference in sP rats that could be related tothe fact that the anxiolytic effects of ethanol mayconstitute a component of its reinforcing properties,which then promote and maintain ethanol preference insP rats. Serotonin neurons are unlikely to be the onlyneuronal system involved in alcohol abuse. Dopamineand noradrenaline (Casu et al. 2002), g-amino butyricacid, and many peptidergic systems are also to be takeninto consideration. The reduction of serotonergic inner-vation in different brain regions of sP rats provides a clueamong multiple transmitter systems for finding thenumerous neuronal pathways that lead to alcohol drinkingbehavior and abuse.
References
Arendt T, Henning D, Gray JA, Marchbanks R (1988) Loss ofneurons in the rat basal forebrain cholinergic projection systemafter prolonged intake of ethanol. Brain Res Bull 21:563–570
Bjorklund L, Stromberg I (1997) Dopaminergic innervation ofstriatal grafts placed into different sites of normal striatum:differences in the tyrosine hydroxylase immunoreactive growthpattern. Exp Brain Res 113:13–23
Campanelli C, Dzung Z, Khanna J, Kalant H (1988) Effect of raphelesions on the development of acute tolerance to ethanol andpentobarbital. Psycopharmacology 96:454–457
Casu MA, Dinucci D, Colombo G, Gessa GL, Pani L (2002)Reduced DAT and DBH immunostaining in the limbic systemof Sardinian alcohol-preferring rats. Brain Res 948:192–202
Ciccocioppo R, Panocka I, Stefanini E, Gessa GL, Massi M (1995)Low responsiveness to agents evoking 5HT2 receptor-mediatedbehaviors in Sardinian alcohol-preferring rats. PharmacolBiochem Behav 51:21–27
Ciccocioppo R, Panocka I, Froldi R, Colombo G, Gessa GL, MassiM (1999a) Antidepressant-like effect of ethanol revealed in theforced swimming test in Sardinian alcohol-preferring rats.Psychopharmacology 144:151–157
Ciccocioppo R, Angeletti S, Colombo G, Gessa GL, Massi M.(1999b) Autoradiographic analysis of 5HT2A binding sites in thebrain of Sardinian alcohol-preferring and -nonpreferring rats.Eur J Pharmacol 373:13–19
Cloninger CR (1987) Neurogenetic adaptive mechanisms inalcoholism. Science 236:410–416
Collins MA, Corso TD, Neafsey EJ (1996) Neuronal degenerationin rat cerebrocortical and olfactory regions during subchronic“binge” intoxication with ethanol: possible explanation forolfactory deficits in alcoholics. Alcohol Clin Exp Res 20:284–292
Colombo G (1997) Ethanol drinking behaviour in Sardinianalcohol-preferring rats. Alcohol Alcohol 32:443–453
Colombo G, Agabio R, Lobina C, Reali R, Zocchi A, Fadda F,Gessa GL (1995) Sardinian alcohol-preferring rats: a geneticanimal model of anxiety. Physiol Behav 57:1181–1185
Dackis CA, Gold MS (1985) New concepts in cocaine addiction:the dopamine depletion hypothesis. Neurosci Biobehav Rev9:469–477
Devoto P, Colombo G, Stefanini E, Gessa GL (1998) Serotonin isreduced in the frontal cortex of Sardinian ethanol-preferringrats. Alcohol Alcohol 33:226–229
Fox NC, Freeborough PA, Rossor MN (1996) Visualisation andquantification of rates of atrophy in Alzheimer’s disease.Lancet 348:94–97
Gawin FH (1991) Cocaine addiction: psychology and neurophys-iology. Science 251:1580–1586
Eriksson E, Humble M (1990) The biological basis of psychiatrictreatment. In: Pohl R, Gershon S (eds) Progress in basic clinicalpharmacology 3. Karger, Amsterdam, pp 66–119
Gessa GL, Colombo G, Fadda F (1991) Rat lines geneticallyselected for differences in voluntary ethanol consumption. In:Meltzer HY, Nerozzi D (eds) Current practices and futuredevelopments in the pharmacotherapy of mental disorders.Elsevier, Amsterdam, pp 193–200
Graeff FG, Guimares FS, De Andrade TG, Deakin JFW (1996)Role of 5-HT in stress, anxiety and depression. PharmacolBiochem Behav 54:129–141
Gundersen HJG, Bagger P, Bendsten TF, Evans S, Korbo L,Marcussen N, Meller A, Nielsen K, Nyengaard B, Serensen FB,Vesterby A, West M (1988) The new stereological tools:disector, fractionator, nucleator and point sampled interceptsand their use in pathological research and diagnosis. APMIS96:857–881
Harris GC, Altomare K, Aston-Jones G (2001) Preference for acocaine-associated environment is attenuated by augmentedaccumbal serotonin in cocaine withdrawn rats. Psychopharma-cology 156:14–22
Heimer L, Zahm DS, Churcill L, Kalivas PW, Wohltmann C (1991)Specificity in the projection patterns of accumbal core and shellin the rat. Neuroscience 41:89–125
Holman RB, Snape BM (1985) Effects of ethanol on 5-hydroxy-tryptamine release from rat corpus striatum in vivo. Alcohol2:249–253
Khanna JM, Kalant H, Le AD, LeBlanc AE (1979) Effect ofmodification of brain serotonin (5-HT) on ethanol tolerance.Alcohol Clin Exp Res 3:353–358
Koob GF (1999) The role of the striatopallidal and extendedamygdala systems in drug addiction. Ann N Y Acad Sci877:445–460
Koob GF, Bloom FE (1988) Cellular and molecular mechanisms ofdrug dependence. Science 242:715–723
LaBuda CJ, Fuchs PN (2000) Aspirin attenuates the anxioliticactions of ethanol. Alcohol 21:287–290
Le AD, Khanna JM, Kalant H, LeBlanc AE (1981) The effect oflesions in the dorsal, median and magnus raphe nuclei on thedevelopment of tolerance to ethanol. Pharmacology 218:525–529
Lovinger DM (1999) 5-HT3 receptors and the neural actions ofalcohols: an increasingly exciting topic. Neurochem Int35:125–130
Lundqvist C, Alling C, Knoth R, Volk B (1995) Intermittentethanol exposure of adult rats: hippocampal cell loss after onemonth of treatment. Alcohol Alcohol 30:737–748
Maldonado-Irizarry CS, Kelley AE (1995) Excitatory amino acidreceptors within nucleus accumbens subregions differentiallymediate spatial learning in the rat. Behav Pharmacol 6:527–539
Mann JJ, Malone KM, Diehl DJ, Perel J, Cooper TB, Mintun MA(1996) Demonstration in vivo of reduced serotonin responsivityin the brain of untreated depressed patients. Am J Psychiatry153:174–182
Markou A, Kosten TR, Koob GF (1998) Neurobiological similar-ities in depression and drug dependence: a self medicationhypothesis. Neuropsychopharmacology 18:135–174
Massou JM, Trichard C, Attar-Levy D, Feline A, Corruble E,Beaufils B, Martinot JL (1997) Frontal 5HT2 receptors studiedin depressive patients during chronic treatment by selectiveserotonin reuptake inhibitors. Psychopharmacology 133:99–101
McBride WJ, Chernet E, Rabold JA, Lumeng L, Li TK (1993)Serotonin-2 receptors in the CNS of alcohol-preferring and -nonpreferring rats. Pharmacol Biochem Behav 46:631–636
McBride WJ, Guan XM, Chernet E, Lumeng L, Li TK (1994)Regional serotonin1A receptors in the CNS of alcohol-preferringand -nonpreferring rats. Pharmacol Biochem Behav 49:7–12
Meert TF, Awouters F, Melis WJC, Janssen PAJ (1990) Ritanserinreduces alcohol intake in rats given the choice between 3%alcohol and water. Pharmacology 9:63–69
350

Melchior CL, Myers RD (1976) Genetic differences in ethanoldrinking of the rat following injection of 6-OHDA, 5,6-DHT or5,7-DHT in to the cerebral ventricles. Pharmacol BiochemBehav 5:66–72
Melchior CL, Tabakoff B (1986) The effect of 5,7 dihydroxy-tryptamine treatment on the response to ethanol in mice.Pharmacology 24:955–961
Myers RD, Melchior CL (1975) Alcohol drinking in the rat afterdestruction of serotoninergic and catecholaminergic neurons inthe brain. Res Commun Chem Pathol Pharmacol 10:363–378
Mounton PR (2002) Principles and practices of unbiased stereol-ogy, an introduction for bioscientists. The Johns HopkinsUniversity Press, Baltimore
Murphy JM, McBride WJ, Lumeng L, Li TK (1982) Regionallevels of monoamines in alcohol-preferring and –nonpreferringlines of rats. Pharmacol Biochem Behav 16:145–149
Murphy JM, McBride WJ, Lumeng L, Li TK (1987) Contents ofmonoamines in forebrain regions of alcohol-preferring (P) and -nonpreferring (NP) lines of rats. Pharmacol Biochem Behav26:389–392
Panocka I, Massi M (1992) Long-lasting suppression of alcoholpreference in rats following serotonin receptor blockade byritanserin. Brain Res Bull 28:493–496
Panocka I, Ciccocioppo R, Polidori C, Massi M (1993) The nucleusaccumbens is a site of action for the inhibitory effect ofritanserin on ethanol intake in rats. Pharmacol Biochem Behav46:857–862
Panocka I, Ciccocioppo R, Polidori C, Pompei P, Massi M (1995)The 5HT4 receptor antagonist, GR113808, reduces ethanolintake in alcohol-preferring rats. Pharmacol Biochem Behav52:255–259
Parsons LH, Koob GF, Weiss F (1996) Extracellular serotonin isdecreased in the nucleus accumbens during withdrawal fromcocaine self-administration. Behav Brain Res 73:225–228
Paxinos G, Watson C (1986) The rat brain in stereotaxiccoordinates, 2nd edn. Academic Press, New York
Pierce RC, Kalivas PW (1995) Amphetamine produced sensitizedincreases in locomotion and extracellular dopamine preferen-
tially in the nucleus accumbens shell of rats administeredrepeated cocaine. J Pharmacol Exp Ther 275:1019–1029
Portas CM, Devoto P, Gessa GL (1994) Effect of ethanol onextracellular 5-hydroxytryptamine output in rat frontal cortex.Eur J Pharmacol 270:123–125
Prunell M, Escorihuela RM, Fernandez-Teruel A, Nunez JF,Tobena A (1994) Differential interactions between ethanoland Ro15-4513 on two anxiety tests in rats. PharmacolBiochem Behav 47:147–151
Pulvirenti L, Berrier R, Kriefeldt M, Koob GF (1994) Modulationof locomotor activity by NMDA receptors in the nucleusaccumbens core and shell regions of the rat. Brain Res664:231–236
Sellers EM, Higgins GA, Sobell MB (1992) 5-HT and alcoholabuse. Trends Pharmacol Sci 13:69–75
Sterio DC (1984) The unbiased estimation of number and sizes ofarbitrary particles using the disector. J Microsc 134:127–36
Weiss F, Markou A, Lorang MT, Koob GF (1992) Basalextracellular dopamine levels in the nucleus accumbens aredecreased during cocaine withdrawal after unlimited-accessself-administration. Brain Res 593:314–328
Wong TP, Marchese G, Casu MA, Ribeiro-Da Silva A, Cuello AC,De Koninck Y (2000) Loss of presynaptic and postsynapticstructures is accompanied by compensatory increase in actionpotential-dependent synaptic input to layer V neocorticalpyramidal neurons in aged rats. J Neurosci 20:8596–8606
Zhou FC, Bledsoe S, Li TK, Lumeng L (1991) Immunostainedserotoninergic fibers are decreased in selected brain regions ofalcohol-preferring rats. Alcohol 8:425–429
Zhukov VN, Varkov AI, Burov YV (1985) Effect of destruction ofthe brain serotoninergic system on the alcohol consumption byrats in the early period of experimental alcoholism. Bull ExpBiol Med 99 576–577
Zou JY, Martinez DB, Neafsey EJ, Collins MA (1996) Bingeethanol-induced brain damage in rats: effect of inhibitors ofnitric oxide synthase. Alcohol Clin Exp Res 20:1406–1411
351
Related Documents











