of October 7, 2015. This information is current as Immunity in C4-Deficient Mice Component C4 Can Restore Humoral Macrophage-Derived Complement C. Carroll Heather Jezak, Li Ming Shen, David M. Knipe and Michael Mihaela Gadjeva, Admar Verschoor, Mark A. Brockman, http://www.jimmunol.org/content/169/10/5489 doi: 10.4049/jimmunol.169.10.5489 2002; 169:5489-5495; ; J Immunol References http://www.jimmunol.org/content/169/10/5489.full#ref-list-1 , 35 of which you can access for free at: cites 50 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2002 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on October 7, 2015 http://www.jimmunol.org/ Downloaded from by guest on October 7, 2015 http://www.jimmunol.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

of October 7, 2015.This information is current as
Immunity in C4-Deficient MiceComponent C4 Can Restore Humoral Macrophage-Derived Complement
C. CarrollHeather Jezak, Li Ming Shen, David M. Knipe and Michael Mihaela Gadjeva, Admar Verschoor, Mark A. Brockman,
http://www.jimmunol.org/content/169/10/5489doi: 10.4049/jimmunol.169.10.5489
2002; 169:5489-5495; ;J Immunol
Referenceshttp://www.jimmunol.org/content/169/10/5489.full#ref-list-1
, 35 of which you can access for free at: cites 50 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2002 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on October 7, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on October 7, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

Macrophage-Derived Complement Component C4 Can RestoreHumoral Immunity in C4-Deficient Mice1
Mihaela Gadjeva,*† Admar Verschoor,‡ Mark A. Brockman,§ Heather Jezak,* Li Ming Shen,*David M. Knipe,§ and Michael C. Carroll2*†‡
Mice with a disrupted C4 locus (C4�/�) have an impaired immune response to thymus-dependent Ags. To test the role of bonemarrow-derived C4 in humoral immunity, we reconstituted deficient animals with wild-type bone marrow or an enriched fractionof bone marrow-derived macrophages. C4 chimeras were immunized with 4-hydroxy-3-nitrophenyl5 conjugated to keyhole limpethemocyanin (NP5- KLH) or infected with HSV-1, and the Ab response was evaluated. Wild-type bone marrow rescued thehumoral immune response to both Ags, i.e., the soluble Ag and HSV-1, demonstrating that local C4 production is sufficient forhumoral responses. Although the C4 chimeric animals lacked detectable C4 in their sera, C4 mRNA was identified in splenicsections by in situ hybridization, and C4 protein deposits were identified in the germinal center areas of splenic follicles byimmunofluorescence staining. Macrophages derived from bone marrow produced sufficient C4 protein to restore the humoralresponse to NP5-KLH in C4-deficient animals when administered along with Ag. Cell-sorting experiments, followed by C4-specificRT-PCR, identified splenic macrophages (CD11b�, CD11c�) as a cellular source for C4 synthesis within the spleen. The Journalof Immunology, 2002, 169: 5489–5495.
A ctivation of complement modulates humoral responses(1, 2). Studies with transient depletion of complementwith cobra venom factor or with animals bearing natu-
rally occurring complement deficiencies demonstrate the impor-tance of the complement system in thymus-dependent (T-D)3 re-sponses (3–5). Similarly, genetically engineered mice strainsdeficient in complement C1qA (6), C3, C4 (7), and CD35/CD21(8–10), or mice treated with blocking Ab (11–13) or soluble re-ceptor (14) show defective responses to T-D Ags administered inthe absence of adjuvants. The reduction in Ab production corre-lates with reduced numbers of germinal centers (GC) (7, 9, 10, 15).T cells appear to be activated normally, suggesting that the defectis in the B cell response (7, 11, 16). Complement enhances B cellresponses by localization of Ag on follicular dendritic cells (FDC)(15, 17) and B cell coreceptor signaling via CD21/CD19/CD81 (9,18). Coligation of CD21/CD19 coreceptor enhances B cell recep-tor signaling (19, 20) and effectively lowers the threshold for ac-tivation. The effects of complement are dependent on covalent at-tachment of C3 to Ag or Ab (21, 22). Significantly, direct couplingof C3d multimers to protein Ag can increase the immunogenicityby greater than 1000-fold (23).
The impairment in T-D responses to Ags administered i.v. canbe restored by passive administration of purified C3 or C4 alongwith Ag (24, 25). Although the liver is the major source of serumC3 and C4, certain bone marrow (BM)-derived cells are also ca-pable of producing complement proteins (26, 27). Notably, recon-stitution of C3-deficient animals with wild-type (WT) BM can re-store the humoral response to T-D Ag administered i.v (28) orintradermally (i.d.) (29). BM-derived cells are also a source of C1q(30); thus, myeloid cells can provide a local source for comple-ment, both in the spleen and lymph nodes. These studies raiseimportant questions regarding the source and regulation of my-eloid-derived complement.
To determine whether local synthesis is sufficient to restore thehumoral response, C4-deficient mice were engrafted with WT BMor with an enriched fraction of BM-derived macrophages. BM en-graftment was sufficient to restore the humoral response to a T-D-soluble Ag 4-hydroxy-3-nitrophenyl5 conjugated to keyhole limpethemocyanin (NP5-KLH) or infectious virus (HSV-1). Local C4was produced by CD11b�/CD11c� splenic macrophages, whichalso synthesized C1q and C3. Moreover, an enriched population ofBM-derived macrophages was capable of restoring the B cell re-sponse to NP5-KLH in deficient animals. Taken together, thesedata suggest that upon stimulation, macrophages within the sec-ondary lymphoid compartment produce early complement compo-nents, which allows for triggering of the complement cascade viathe classical pathway, leading to enhancement of humoral re-sponses to T-D Ags.
Materials and MethodsMouse strains
C4�/� mice were generated by homologous recombination, as describedpreviously (7), and backcrossed with C57BL/6 (The Jackson Laboratory,Bar Harbor, ME) mice for five generations (�95% C57BL/6). Animalswere bred and maintained at the Warren Alpert Animal Facility at HarvardMedical School. STAT-1- and IFN-�R-deficient mice were purchased fromTaconic (Germantown, NY) and The Jackson Laboratory, respectively.
*Center for Blood Research, Boston, MA 02115; and Departments of †Pediatrics,‡Pathology, and §Microbiology and Molecular Genetics, Harvard Medical School,Boston, MA 02115
Received for publication July 23, 2002. Accepted for publication September 16, 2002.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by grants from the National Institutes of Health, P01AI42257 and R01 HD38749.2 Address correspondence and reprint requests to Dr. Michael C. Carroll, Center forBlood Research, 221 Longwood Avenue, LMRC501, Boston, MA 02115. E-mailaddress: [email protected] Abbreviations used in this paper: T-D, thymus dependent; BM, bone marrow;BMM, BM macrophage; DIG, digoxigenin; FDC, follicular dendritic cell; GC, ger-minal center; i.d., intradermally; KLH, keyhole limpet hemocyanin; NP5, 4-hydroxy-3-nitrophenyl5; PI, propidium iodide; PNA, peanut agglutinin; WT, wild type.
The Journal of Immunology
Copyright © 2002 by The American Association of Immunologists, Inc. 0022-1767/02/$02.00
by guest on October 7, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

Generation of BM chimeras
BM cells were collected by flushing the femurs and tibias with cold PBS/0.1 mM EDTA, followed by depletion of erythrocytes by lysis with 0.15 MNH4Cl, 10 mM KHCO3. At 7–10 wk of age, C4�/� or C57BL/6 mice werelethally irradiated using two 650-rad doses and reconstituted i.v with 10 �106 BM cells derived from gender- and age-matched C57BL/6 mice.
Immunization protocol
After transplantation, animals were rested for 6 wk and then either immu-nized with 100 �g NP5-KLH i.v. or infected with 2 � 106 PFU HSV-1(KOS1.1 strain) i.d. (16, 31). After each immunization, mice were restedfor 3 wk and then boosted. Animals were sacrificed 1 wk after the thirdboost. Serum samples were collected weekly.
Serum C4 ELISA
Immulon 1B microtiter plates (DYNEX Technologies, Chantilly, VA)were coated overnight with rat anti-murine C4 mAb 16D2 (a kind gift fromE. Kremmer, GSF National Research Center for Environment and Health,Munich, Germany) in carbonate buffer. After blocking with 5% dry milk inPBS and 0.01% Tween 20, serial dilutions of mouse serum in blockingbuffer were applied to the wells and incubated for 2 h at 37°C. Murine C4was detected with rabbit anti-human C4c (DAKO, Glostrup, Denmark) andthen followed by alkaline phosphatase-conjugated goat anti-rabbit IgG(Sigma-Aldrich, St. Louis, MO). Plates were developed by adding Sigma-Aldrich alkaline phosphatase substrate 104, and absorbance was measuredat 405 nm.
Ab titers
Immulon 1B microtiter plates (DYNEX Technologies) were coated over-night with NP5-haptenated BSA (32) (Sigma-Aldrich), blocked as de-scribed above, and serial serum dilutions were applied and incubated for3 h at 37°C. Murine IgG was detected by alkaline phosphatase-conjugatedrat anti-mouse IgG (Sigma-Aldrich). HSV-1-specific ELISA was per-formed as previously described (29).
In situ C4 hybridization
A BamHI-KpnI restriction fragment representing the 5� terminal end of C4cDNA (a kind gift from R. Ogata, Torrey Pines Institute for MolecularStudies, San Diego, CA) was subcloned into pBluescript II KS�/� (Strat-agene, La Jolla, CA). Antisense digoxigenin (DIG)-labeled mC4 tran-scripts were produced by linearizing the pBSTmC4 plasmid with Xbaand transcribing with T3 RNA polymerase I. DIG-labeled probes wereproduced with a DIG RNA labeling kit (SP/T7) (Roche Diagnostics, In-dianapolis, IN) per manufacturer’s instructions. RNase-free sections (4�m) were cut, and in situ hybridization was performed, as described pre-viously (28).
BM-derived macrophages
BM-derived macrophages were grown in L929 (American Type CultureCollection, Manassas, VA)-preconditioned DMEM (Life Technologies,Rockville, MD) medium and supplemented with 10% FCS, 5% horse se-rum (Sigma-Aldrich), 2 mM L-glutamine, and 100 U/ml penicillin/strep-tomycin (Life Technologies) until the cells were confluent. Cells werescraped with a cell lifter (Fisher Scientific, Pittsburgh, PA), replated at 1 �106 cells/well density in six-well cell culture Costar plates (Corning Glass,Corning, NY), and stimulated with 5000 IU rIFN-� (R&D Systems, Min-neapolis, MN) for 12 h.
Immunofluorescence
Spleens were snap frozen in OCT (Tissue Tek Sakura, Torrence, CA)-filledmolds (VWR, West Chester, PA) and stored at �80°C until cryosectionswere cut. Sections (5 �m) were fixed for 4 min in ice-cold acetone. Sec-tions were blocked with 2% BSA, 2% FCS, and PBS. The following stepswere conducted in the presence of the blocking buffer: sections werestained with anti-C4 rat mAb 15H12 or 16D2 (kind gifts from E. Kremmer)conjugated to Cy5 (Amersham, Arlington Heights, IL) or biotin- and FITC-conjugated peanut agglutinin (PNA; EY Laboratories, San Mateo, CA), asdescribed (9). Both mAbs produced similar results when used for immu-nofluorescence. The biotinylated 16D2 was visualized using avidin-PE(BD PharMingen, San Diego, CA).
FACS analysis and sorting
Splenocytes were incubated on ice with anti-FcR (2.4G2), before treatmentwith Abs specific for CD45.1, CD45.2, CD11b, or CD11c (all from BD
PharMingen), and analyzed using a FACSCalibur flow cytometer withCellQuest software (BD Biosciences, San Jose, CA), or sorted using aFACSVantage. When necessary, the splenocytes were depleted from B andT lymphocytes by magnetic cell sorting with microbeads before FACSanalysis or sorting. For depletion purposes, biotinylated Abs specific forCD19 or CD3 (BD PharMingen) were used, followed by streptavidinMACS beads (Miltenyi Biotec, Auburn, CA). The separation was per-formed on MACS LD-separation columns (Miltenyi Biotec), and the un-bound cells were collected and analyzed further. Cell viability was ac-counted for by propidium iodide (PI) staining and subsequent gating out ofPI positively stained cells during the analysis.
RT-PCR
Total RNA was purified from sorted cells or splenic tissue per Qiagen(Valencia, CA) RNeasy mini kit instructions. First strand synthesis wasperformed with SuperScript RT (Life Technologies). C4 RT-PCR yieldeda 472-bp-specific band after 30 rounds of amplification with the followingprimer set: sense (GGTTCTGAAGGTGCCTTGTCCC) and antisense(GTGAAGGGCAATGACCACAAAGG). C3 message was amplified after35 cycles with the following set of primers: sense (GGCTGACTCTGTGTGGGT) and antisense (TCTCTGGTTCTTTCAACTCT).
ResultsHumoral responses are restored in the WT BM3C4�/�
chimeric animals
To determine whether myeloid-derived C4 was important in theT-D humoral response, C4-deficient mice were reconstituted withWT BM (WT BM3C4�/�). Three cohorts of chimeric animalswere analyzed in these studies: 1) WT BM3WT; 2) WTBM3C4�/�; and 3) C4 �/� BM3C4�/�. C57BL/6 (CD45.2)and C4�/� mice on C57BL/6 (CD45.2) background were used asrecipients for donor marrow from C57BL/6 (CD45.1) mice. Thecongenic CD45.1 C57BL/6 strain was used to facilitate tracking ofthe donor-derived cells bearing the allotypic CD45.1 marker in therecipient mice. Mice were immunized i.v. with 100 �g of solublehaptenated NP5-KLH at days 0 and 21. This dose of Ag was pre-viously shown to be fully immunogenic (28). Adjuvants wereomitted from the immunization protocol to avoid circumventingthe role of complement. NP-specific IgG titers were determined byELISA (Fig. 1a). Alternatively, mice were infected i.d. with 2 �106 PFU of infectious HSV-1 (Fig. 1b). Consistent with previousstudies using C4-deficient mice, the immune response of C4-de-ficient chimeras to both soluble Ag (7) and infectious HSV (16)was �8- to 18-fold lower than that of WT chimeras. By contrast,nearly normal primary and secondary Ab responses were observedin C4-deficient mice reconstituted with WT BM.
To further characterize the humoral response in the chimericanimals, splenic sections were prepared, and the number of GCswas determined by PNA staining, followed by immunofluores-cence analysis. At least five animals per group were analyzed 7days after the tertiary immunization (Table I). The percentage ofGCs per follicle was similar to that observed in splenic sections ofWT BM3C4�/� chimeras and WT BM3WT chimeras (Table I,Fig. 1). By contrast, the percentage of GCs in the splenic sectionsof C4�/� BM3C4�/� chimeras was reduced. Immunohistochem-ical analysis of splenic sections identified detectable C4 deposits inthe GC areas in all of the WT chimeras, compared with �50% inWT BM3C4�/� chimeras (Table I, Fig. 2b). C4-specific stainingin the spleen of all analyzed sections was found in the GC areasand appeared to be network-like, consistent with the presence ofadducts of C4 and Ag/C3 protein captured by the FDCs.
The cellular source of complement C4
The WT BM3C4�/� chimeric animals lacked measurable C4protein in their sera based on ELISA analysis (Fig. 2a), but C4 wasdetected in splenic sections by immunofluorescence, as discussed
5490 A ROLE FOR MACROPHAGE-PRODUCED C4
by guest on October 7, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
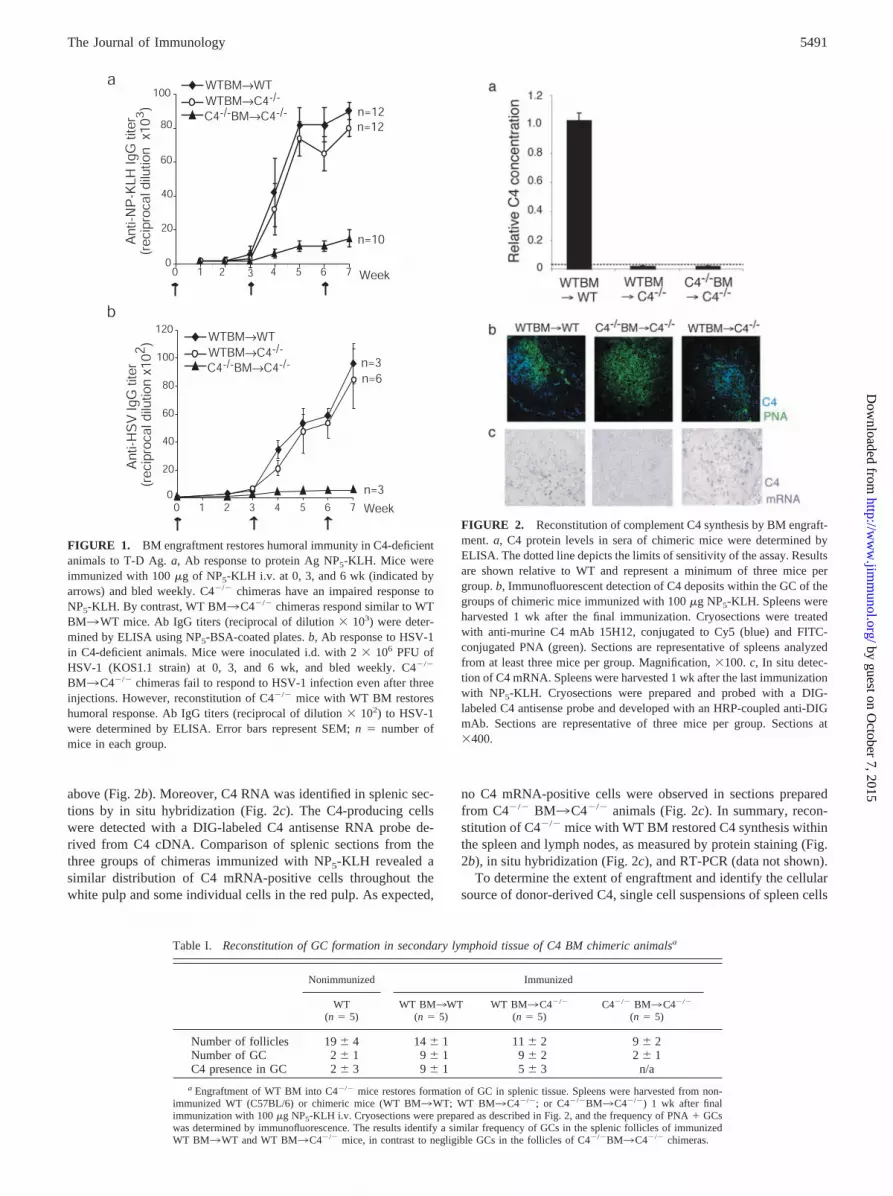
above (Fig. 2b). Moreover, C4 RNA was identified in splenic sec-tions by in situ hybridization (Fig. 2c). The C4-producing cellswere detected with a DIG-labeled C4 antisense RNA probe de-rived from C4 cDNA. Comparison of splenic sections from thethree groups of chimeras immunized with NP5-KLH revealed asimilar distribution of C4 mRNA-positive cells throughout thewhite pulp and some individual cells in the red pulp. As expected,
no C4 mRNA-positive cells were observed in sections preparedfrom C4�/� BM3C4�/� animals (Fig. 2c). In summary, recon-stitution of C4�/� mice with WT BM restored C4 synthesis withinthe spleen and lymph nodes, as measured by protein staining (Fig.2b), in situ hybridization (Fig. 2c), and RT-PCR (data not shown).
To determine the extent of engraftment and identify the cellularsource of donor-derived C4, single cell suspensions of spleen cells
Table I. Reconstitution of GC formation in secondary lymphoid tissue of C4 BM chimeric animalsa
Nonimmunized Immunized
WT(n � 5)
WT BM3WT(n � 5)
WT BM3C4�/�
(n � 5)C4�/� BM3C4�/�
(n � 5)
Number of follicles 19 � 4 14 � 1 11 � 2 9 � 2Number of GC 2 � 1 9 � 1 9 � 2 2 � 1C4 presence in GC 2 � 3 9 � 1 5 � 3 n/a
a Engraftment of WT BM into C4�/� mice restores formation of GC in splenic tissue. Spleens were harvested from non-immunized WT (C57BL/6) or chimeric mice (WT BM3WT; WT BM3C4�/�; or C4�/�BM3C4�/�) 1 wk after finalimmunization with 100 �g NP5-KLH i.v. Cryosections were prepared as described in Fig. 2, and the frequency of PNA � GCswas determined by immunofluorescence. The results identify a similar frequency of GCs in the splenic follicles of immunizedWT BM3WT and WT BM3C4�/� mice, in contrast to negligible GCs in the follicles of C4�/�BM3C4�/� chimeras.
FIGURE 1. BM engraftment restores humoral immunity in C4-deficientanimals to T-D Ag. a, Ab response to protein Ag NP5-KLH. Mice wereimmunized with 100 �g of NP5-KLH i.v. at 0, 3, and 6 wk (indicated byarrows) and bled weekly. C4�/� chimeras have an impaired response toNP5-KLH. By contrast, WT BM3C4�/� chimeras respond similar to WTBM3WT mice. Ab IgG titers (reciprocal of dilution � 103) were deter-mined by ELISA using NP5-BSA-coated plates. b, Ab response to HSV-1in C4-deficient animals. Mice were inoculated i.d. with 2 � 106 PFU ofHSV-1 (KOS1.1 strain) at 0, 3, and 6 wk, and bled weekly. C4�/�
BM3C4�/� chimeras fail to respond to HSV-1 infection even after threeinjections. However, reconstitution of C4�/� mice with WT BM restoreshumoral response. Ab IgG titers (reciprocal of dilution � 102) to HSV-1were determined by ELISA. Error bars represent SEM; n � number ofmice in each group.
FIGURE 2. Reconstitution of complement C4 synthesis by BM engraft-ment. a, C4 protein levels in sera of chimeric mice were determined byELISA. The dotted line depicts the limits of sensitivity of the assay. Resultsare shown relative to WT and represent a minimum of three mice pergroup. b, Immunofluorescent detection of C4 deposits within the GC of thegroups of chimeric mice immunized with 100 �g NP5-KLH. Spleens wereharvested 1 wk after the final immunization. Cryosections were treatedwith anti-murine C4 mAb 15H12, conjugated to Cy5 (blue) and FITC-conjugated PNA (green). Sections are representative of spleens analyzedfrom at least three mice per group. Magnification, �100. c, In situ detec-tion of C4 mRNA. Spleens were harvested 1 wk after the last immunizationwith NP5-KLH. Cryosections were prepared and probed with a DIG-labeled C4 antisense probe and developed with an HRP-coupled anti-DIGmAb. Sections are representative of three mice per group. Sections at�400.
5491The Journal of Immunology
by guest on October 7, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

were prepared from chimeras and analyzed by FACS. Over 90% ofthe CD11b� population in the spleen was of donor origin, based onexpression of the CD45.1 donor allotype (Fig. 3). This compart-ment includes myeloid-derived cells, such as splenic macrophages,dendritic cells, and neutrophils. The CD11b� splenocytes werefurther subdivided based on CD11c staining and forward and sidescatter characteristics. CD11b� R2- and CD11b� R3-gated cellswere sorted (Fig. 4a), and cytospins were prepared. Nuclear mor-phology determined with Giemsa Wright staining showed that theR2 gate included granulocytes, whereas the R3 gate was comprisedprimarily of monocytes (macrophages). To establish whether thesorted cells produced not only C4 and C3 complement compo-
nents, but also C1q, total RNA was prepared from the sorted pop-ulations and analyzed by RT-PCR. Although both granulocytesand monocytes synthesize C3 (Fig. 4b), only the monocytic pop-ulation was positive for C4 and C1q mRNA. Thus, a similar pop-ulation of macrophages produced C1q, C4, and C3.
BM-derived macrophages can produce sufficient C4 to restorehumoral immunity to NP5-KLH
To test the ability of macrophages to produce complement C4,monocytic cells were derived from cultured C57BL/6 BM. FACSanalysis of the cultured cells revealed a high frequency of macro-phages (CD11b�, F480�CD11c�, and I-Ab� cells; data not
FIGURE 3. Engraftment of C4�/�
animals with WT BM results in over 90%donor CD11b� cells in the spleens of re-cipient mice after 14 wk. a, Degree of chi-merism in CD11b� splenic populations inBM chimeras. Freshly harvested spleno-cytes were stained with Abs specific forCD45.1, CD45.2, and CD11b. PI wasadded to detect dead cells. Flow data wereanalyzed by gating on the majority ofsplenocytes based on forward and sidescatter (R1), excluding PI-positive cells(R2) (data not shown). The CD11b profile(R3) is based on combined R1 and R2gates. Representative histograms areshown from WT BM3WT, WTBM3C4�/�, and C4�/� BM3C4�/�
chimeras 14 wk after BM transplantation.b, Mean level of chimerism in WTBM3WT and WT BM3C4�/� is de-termined by the frequency of CD45.1 al-lotype-marked donor cells. Significantly,�90% of CD11b� cells are donor de-rived. Results represent analysis ofsplenocytes harvested from five mice pergroup. Note that C4�/� BM donor cellsare CD45.2; therefore, the degree of chi-merism could not be determined.
FIGURE 4. Splenic macrophages are a common cel-lular source for C1q, C3, and C4. a, A representative sort-ing experiment to distinguish between monocytes andpolymorphonuclear cells in the spleens of C57BL/6 ani-mals. The splenocytes were sorted on the basis of theirgranularity and size, determined by forward (FSC) andside scatter (SSC) characteristics and expression ofCD11b and CD11c. The CD11b�CD11c� cells (R1gated) were subdivided based on FSC and SSC character-istics into two groups, i.e., R2 and R3 gates. Cytospinswere prepared from sorted cells and analyzed followingGiemsa Wright staining. b, Detection of synthesis of com-plement components. Total RNA was prepared from 20 �104 sorted cells (R2 and R3 gates) and analyzed by RT-PCR for C4, C3, and C1q mRNA presence. As controls,total RNA from splenic tissues was prepared fromC57BL/6, C3�/�/C4�/�, and C1q�/� mice, followed byactin, C3, C4, or C1q RT-PCRs. Results are representativeof at least three separate sorting experiments.
5492 A ROLE FOR MACROPHAGE-PRODUCED C4
by guest on October 7, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

shown), confirming the expected macrophage phenotype of thecells in the cultures. C4 expression by BM macrophages (BMM)was detected by RT-PCR (Fig. 5a) after stimulation with 5000 IUof rIFN-�. Expression was STAT-1 dependent, as BMM isolatedfrom STAT-1- or IFN-�-deficient mice (data not shown) failed toup-regulate C4 mRNA, suggesting that C4 expression is tightlyregulated (Fig. 5a).
To test the functional importance of C4 expression by BMM invivo, 20 � 106 nonstimulated BMM were administered i.v. toC4-deficient mice 1 h before immunization with 100 �g NP5-KLH. At 3 wk, a comparable dose of BMM was administered, andmice were boosted with 100 �g NP5-KLH. Serum titers of anti-NPIgG were assayed weekly (Fig. 5b). As expected, the response inC4-deficient animals receiving C4�/� BMM was impaired com-pared with WT animals. By contrast, the response observed in WTand C4�/� animals that received BMM was comparable. Thus,engraftment of C4�/� mice with an enriched population of BMMwas sufficient to restore humoral response to NP5-KLH.
DiscussionMice homozygous for null alleles at C3, C4 (7), or C1q (6) locihave an impaired immune response to T-D Ags such as �X174 orNP5-KLH. BM engraftment restores this defect in C3-deficient an-imals (28). It is possible that BM-derived cells can be a source of
local complement synthesis that includes not only C3, but alsoother complement proteins. To test this possibility, C4�/� micewere engrafted with WT BM or an enriched population of BMM.We found that BM engraftment leads to restoration of humoralresponse in C4-deficient animals despite an apparent absence ofC4 protein in the serum.
Ab responses to two different T-D Ags were evaluated (solubleprotein Ag and infectious HSV-1). In both cases, WT BM engraft-ment led to reconstitution of humoral responses as measured by Abtiters (Fig. 1). Peripheral infection with HSV-1 represents a phys-iologically relevant model to study the role of complement acti-vation. Previous analysis of mice deficient in C3, C4, or CD21/35following infection with HSV-1 revealed an impaired secondaryresponse characterized by low Ab titers. These results suggest thatthe classical pathway is important in initiating complement acti-vation leading to the activation of C3. The effects of C3 are de-pendent on CD21/CD35, as a similar impairment is observed inCD21/CD35-deficient mice (16). Previously, we demonstrated thatWT BM engraftment in C3�/� mice restored the impaired Abresponse to HSV-1 (29). WT BM3C4�/� chimeric mice have aphenotype similar to WT BM3C3�/� mice with regard to HSV-1Ab titers, indicating that local complement production is adequateto enhance the antiviral response and that BM-derived cells canproduce complement components C3 and C4. These findings im-plicate locally produced complement as important to the humoralimmune response and may be helpful in the design of futurevaccines.
In contrast to WT BM3C1q�/� chimeras (30), in which WTBM engraftment led to reconstitution of circulating C1q serumlevels, engraftment did not restore C4 serum levels in WTBM3C4�/� chimeras (Fig. 2a); this finding is similar to previousobservations in WT BM3C3�/� mice (28). These variationscould be explained by the differences in regulation of C1q vs C3and C4. Macrophages, a major cellular source of C1q, are respon-sible for circulating C1q levels, while hepatocytes are the predom-inant source of serum C3 and C4 (26, 27, 30).
C4 synthesis by BM-derived cells was detected by in situ hy-bridization (Fig. 2c). C4 mRNA localized randomly throughout thesplenic white pulp, suggesting that the monocytic-producing cellswere not clustered within a subregion, such as the follicular zone.A similar pattern was previously observed for C3 mRNA (28). Inthis study, the pattern of C3 mRNA expression correlated withMOMA-2� cells based on immunohistochemistry of serial sec-tions. Moreover, MOMA-2� cells were identified as a majorsource of C3 by RT-PCR analysis of an enriched fraction of cells.We used a similar approach to sort MOMA-2� cells by magneticbeads from a suspension of splenocytes; RT-PCR analysis identi-fied C4 mRNA in MOMA-2�, but not B cell controls (results notshown). Thus, the MOMA-2� population of macrophages can ex-press both C3 and C4.
Previous experiments have identified numerous sites for C3synthesis in addition to the liver. In humans, extrahepatic C3-pro-ducing cells include monocytes/macrophages, fibroblasts, capil-lary endothelial cells, T cells, endometrium, adipocytes, osteo-blasts, and intestinal epithelial cells (33–38). Similar extrahepaticsources have been identified in the mouse (39, 40). Although stud-ied less intensely, C4 synthesis appears to be more limited than C3.Murine C4 synthesis was identified in peritoneal and resident kid-ney macrophages (41, 42). Bone marrow-derived macrophages(CD11b�CD11c�) are a cellular source of C4 in the spleen andlymph nodes (Figs. 4 and 5, and data not shown). The CD11b�
cells in WT BM3C4�/� chimeras were over 98% donor derived,as judged by the frequency of the allotypic marker (Fig. 3). Splenicpopulations were sorted based on CD11b vs CD11c expression,
FIGURE 5. Adoptive transfer of BMM in C4�/� mice restores humoralimmunity. a, C4 message is detected in BM-derived macrophages by RT-PCR upon IFN-� stimulation. BMM prepared from C57BL/6 and STAT-1�/� mice were cultured for 5 days. BMM (1 � 106 cells) were stimulatedwith 5000 IU rIFN-�; 6 h later, total RNA was prepared, followed byRT-PCR using primers for C4 and actin. Results indicate negligible levelsof C4 mRNA in BMM isolated from STAT-1�/� mice or untreated BMMisolated from C57BL/6 mice. However, treatment of C57BL/6-, but notSTAT-1�/�-derived BMM with 5000 U IFN-� induces C4 mRNA expres-sion. Results are representative of three different experiments. b, Ab re-sponse to NP5-KLH in C4-deficient animals after adoptive transfer. C4�/�
animals were engrafted with 20 � 106 BMM prepared from C57BL/6 micebefore immunization with NP5-KLH at 0 and 3 wk. Animals were bledweekly, and Ab IgG titers were determined by ELISA, as described in Fig. 1.
5493The Journal of Immunology
by guest on October 7, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

size, and granularity (43). C4 and C1q mRNAs were detected onlyin the monocytic/macrophage-sorted population (Fig. 5b), whereasgranulocytes and monocytic cells both expressed C3. This patternof differential synthesis of complement may reflect the variousphysiological activities of the cell types. Macrophages appear toproduce all the necessary complement components to enhance Bcell activation, whereas neutrophils might contribute more to al-ternative pathway-mediated inflammatory reactions.
Notably, engraftment of WT BM led to sufficient C4 synthesisto allow for local complement activation. As a consequence ofcomplement activation, C4 deposits were captured on FDCs (Fig.2b). The C4-specific staining in the splenic sections colocalizedwith PNA (Fig. 2b) and Ag (data not shown). The frequency ofGCs in the splenic follicles of immunized WT BM3WT and WTBM3C4�/� mice was comparable (Table I). However, while C4deposits were detectable within all GCs in WT BM3WT chime-ras, only �50% of the GCs of WT BM3C4�/� chimeras in-cluded detectable C4 deposits. By contrast, few GCs were ob-served in the follicles of C4�/� BM3C4�/� and nonimmunizedage-matched controls. The immunoreactive C4 protein visualizedby the immunofluorescent staining most likely represents C4-bear-ing immune complexes. These data are consistent with previouslypublished reports identifying the presence of activated products ofcomplement (C1q, C3, C4, and C5) within the GCs (6, 28, 44, 45).Taylor et al. (46) recently reported that the FDC-restricted epitope,FDC-M2, is complement C4 and that its localization in the spleenfollows complement activation and is in the form of immune com-plexes. Complement facilitates trapping of Ag-Ab complexeswithin the FDC network. C4 deposits on FDCs can be explained bydirect interaction with CD35 (9, 15). Alternatively, C4 capturemay be C3 dependent because activated C3b can covalently attachto a specific site in the C4b molecule (47).
Dermal infection of BALB/C mice with HSV-1 is associatedwith a Th1-type response characterized by IFN-� production (48).The cytokine environment within lymphoid organs of immunemice may favor induction of local complement expression. Forexample, C3 synthesis by macrophages is regulated by IFN-� andis shown to be STAT-1 dependent (49). It has been proposed thatIFN-� also increases the stability of C3 and C4 mRNA (50). Sim-ilar to C3, BMMs express higher levels of C4 message by RT-PCRupon IFN-� stimulation (Fig. 5a). The IFN-� effect is also STAT-1dependent (Fig. 5a). It is likely that a common activation pathwayexists in macrophages to ensure rapid up-regulation of comple-ment production locally following infection.
Adoptive transfer experiments of BM-derived macrophages inC4�/� mice suggest a physiological role for macrophage-producedC4 (Fig. 5b). Low levels of macrophage C4 can suffice to facilitatecomplement activation, leading to Ag tagging with C3/C4 frag-ments, capture, and retention on FDCs. The data are consistentwith the hypothesis that complement activation via the classicalpathway is required to enhance T-D responses to inert protein Agsdelivered i.v.
In summary, WT BM or BMM engraftment can provide suffi-cient C4 to allow for complement activation and restoration ofhumoral responses in WT BM3C4�/� chimeric mice. Thus, en-hancement of humoral immunity to pathogens is an importantfunction of locally produced early complement.
AcknowledgmentsWe thank Drs. R. Barrington and M. Zhang for helpful comments duringpreparation of the manuscript; Dr. T. Schneider for assistance with cell-sorting experiments; A. Burton and J. Xia for genotyping the mice; andM. Ottaviano for technical assistance.
References1. Fearon, D. T., and R. H. Carter. 1995. The CD19/CR2/TAPA-1 complex of B
lymphocytes: linking natural to acquired immunity. Annu. Rev. Immunol. 13:127.2. Carroll, M. C., and M. B. Fischer. 1997. Complement and the immune response.
Curr. Opin. Immunol. 9:64.3. Pepys, M. B. 1974. Role of complement in induction of antibody production in
vivo: effect of cobra factor and other C3-reactive agents on thymus-dependentand thymus-independent antibody responses. J. Exp. Med. 140:126.
4. Bottger, E. C., S. Metzger, D. Bitter-Suermann, G. Stevenson, S. Kleindienst, andR. Burger. 1986. Impaired humoral immune response in complement C3-defi-cient guinea pigs: absence of secondary antibody response. Eur. J. Immunol.16:1231.
5. O’Neil, K. M., H. D. Ochs, S. R. Heller, L. C. Cork, J. M. Morris, andJ. A. Winkelstein. 1988. Role of C3 in humoral immunity: defective antibodyproduction in C3-deficient dogs. J. Immunol. 140:1939.
6. Cutler, A. J., M. Botto, D. van Essen, R. Rivi, K. A. Davies, D. Gray, andM. J. Walport. 1998. T cell-dependent immune response in C1q-deficient mice:defective interferon � production by antigen-specific T cells. J. Exp. Med. 187:1789.
7. Fischer, M. B., M. Ma, S. Goerg, X. Zhou, J. Xia, O. Finco, S. Han, G. Kelsoe,R. G. Howard, T. L. Rothstein, et al. 1996. Regulation of the B cell response toT-dependent antigens by classical pathway complement. J. Immunol. 157:549.
8. Croix, D. A., J. M. Ahearn, A. M. Rosengard, S. Han, G. Kelsoe, M. Ma, andM. C. Carroll. 1996. Antibody response to a T-dependent antigen requires B cellexpression of complement receptors. J. Exp. Med. 183:1857.
9. Ahearn, J. M., M. B. Fischer, D. Croix, S. Goerg, M. Ma, J. Xia, X. Zhou,R. G. Howard, T. L. Rothstein, and M. C. Carroll. 1996. Disruption of the Cr2locus results in a reduction in B-1a cells and in an impaired B cell response toT-dependent antigen. Immunity 4:251.
10. Molina, H., V. M. Holers, B. Li, Y. Fung, S. Mariathasan, J. Goellner,J. Strauss-Schoenberger, R. W. Karr, and D. D. Chaplin. 1996. Markedly im-paired humoral immune response in mice deficient in complement receptors 1 and2. Proc. Natl. Acad. Sci. USA 93:3357.
11. Gustavsson, S., T. Kinoshita, and B. Heyman. 1995. Antibodies to murine com-plement receptor 1 and 2 can inhibit the antibody response in vivo without in-hibiting T helper cell induction. J. Immunol. 154:6524.
12. Thyphronitis, G., T. Kinoshita, K. Inoue, J. E. Schweinle, G. C. Tsokos,E. S. Metcalf, F. D. Finkelman, and J. E. Balow. 1991. Modulation of mousecomplement receptors 1 and 2 suppresses antibody responses in vivo. J. Immunol.147:224.
13. Heyman, B., E. J. Wiersma, and T. Kinoshita. 1990. In vivo inhibition of theantibody response by a complement receptor-specific monoclonal antibody.J. Exp. Med. 172:665.
14. Hebell, T., J. M. Ahearn, and D. T. Fearon. 1991. Suppression of the immuneresponse by a soluble complement receptor of B lymphocytes. Science 254:102.
15. Fang, Y., C. Xu, Y. X. Fu, V. M. Holers, and H. Molina. 1998. Expression ofcomplement receptors 1 and 2 on follicular dendritic cells is necessary for thegeneration of a strong antigen-specific IgG response. J. Immunol. 160:5273.
16. Da Costa, X. J., M. A. Brockman, E. Alicot, M. Ma, M. B. Fischer, X. Zhou,D. M. Knipe, and M. C. Carroll. 1999. Humoral response to herpes simplex virusis complement-dependent. Proc. Natl. Acad. Sci. USA 96:12708.
17. Papamichail, M., C. Gutierrez, P. Embling, P. Johnson, E. J. Holborow, andM. B. Pepys. 1975. Complement dependence of localization of aggregated IgG ingerminal centers. Scand. J. Immunol. 4:343.
18. Fischer, M. B., S. Goerg, L. Shen, A. P. Prodeus, C. C. Goodnow, G. Kelsoe, andM. C. Carroll. 1998. Dependence of germinal center B cells on expression ofCD21/CD35 for survival. Science 280:582.
19. Carter, R. H., and D. T. Fearon. 1992. CD19: lowering the threshold for antigenreceptor stimulation of B lymphocytes. Science 256:105.
20. Cherukuri, A., P. C. Cheng, and S. K. Pierce. 2001. The role of the CD19/CD21complex in B cell processing and presentation of complement-tagged antigens.J. Immunol. 167:163.
21. Isenman, D. E., and J. R. Young. 1984. The molecular basis for the difference inimmune hemolysis activity of the Chido and Rodgers isotypes of human com-plement component C4. J. Immunol. 132:3019.
22. Law, S. K., and R. P. Levine. 1977. Interaction between the third complementprotein and cell surface macromolecules. Proc. Natl. Acad. Sci. USA 74:2701.
23. Dempsey, P. W., M. E. Allison, S. Akkaraju, C. C. Goodnow, and D. T. Fearon.1996. C3d of complement as a molecular adjuvant: bridging innate and acquiredimmunity. Science 271:348.
24. Finco, O., S. Li, M. Cuccia, F. S. Rosen, and M. C. Carroll. 1992. Structuraldifferences between the two human complement C4 isotypes affect the humoralimmune response. J. Exp. Med. 175:537.
25. Ochs, H. D., R. J. Wedgwood, M. M. Frank, S. R. Heller, and S. W. Hosea. 1983.The role of complement in the induction of antibody responses. Clin. Exp. Im-munol. 53:208.
26. Alper, C. A., A. M. Johnson, A. G. Birtch, and F. D. Moore. 1969. Human C�3:evidence for the liver as the primary site of synthesis. Science 163:286.
27. Saunders, D., and M. Edidin. 1974. Sites of localization and synthesis of Ssprotein in mice. J. Immunol. 112:2210.
28. Fischer, M. B., M. Ma, N. C. Hsu, and M. C. Carroll. 1998. Local synthesis ofC3 within the splenic lymphoid compartment can reconstitute the impaired im-mune response in C3-deficient mice. J. Immunol. 160:2619.
29. Verschoor, A., M. A. Brockman, D. M. Knipe, and M. C. Carroll. 2001. Cuttingedge: myeloid complement C3 enhances the humoral response to peripheral viralinfection. J. Immunol. 167:2446.
5494 A ROLE FOR MACROPHAGE-PRODUCED C4
by guest on October 7, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

30. Petry, F., M. Botto, R. Holtappels, M. J. Walport, and M. Loos. 2001. Recon-stitution of the complement function in C1q-deficient (C1qa�/�) mice with wild-type bone marrow cells. J. Immunol. 167:4033.
31. Jacob, J., R. Kassir, and G. Kelsoe. 1991. In situ studies of the primary immuneresponse to (4-hydroxy-3-nitrophenyl)acetyl. I. The architecture and dynamics ofresponding cell populations. J. Exp. Med. 173:1165.
32. Hatcher, V., and O. Makela. 1972. Immunological cross-reactions within a familyof related haptens. Immunochemistry 9:1139.
33. Katz, Y., and R. C. Strunk. 1989. IL-1 and tumor necrosis factor: similarities anddifferences in stimulation of expression of alternative pathway of complementand IFN-�2/IL-6 genes in human fibroblasts. J. Immunol. 142:3862.
34. Pantazis, P., V. S. Kalyanaraman, and D. H. Bing. 1990. Synthesis of the thirdcomponent of complement (C3) by lectin-activated and HTLV-infected humanT-cells. Mol. Immunol. 27:283.
35. Sundstrom, S. A., B. S. Komm, H. Ponce-de-Leon, Z. Yi, C. Teuscher, andC. R. Lyttle. 1989. Estrogen regulation of tissue-specific expression of comple-ment C3. J. Biol. Chem. 264:16941.
36. Choy, L. N., B. S. Rosen, and B. M. Spiegelman. 1992. Adipsin and an endog-enous pathway of complement from adipose cells. J. Biol. Chem. 267:12736.
37. Hong, K., T. Kinoshita, P. Pramoonjago, Y. U. Kim, T. Seya, and K. Inoue. 1991.Reconstitution of C5 convertase of the alternative complement pathway withisolated C3b dimer and factors B and D. J. Immunol. 146:1868.
38. Molmenti, E. P., T. Ziambaras, and D. H. Perlmutter. 1993. Evidence for an acutephase response in human intestinal epithelial cells. J. Biol. Chem. 268:14116.
39. Circolo, A., G. Garnier, W. Fukuda, X. Wang, T. Hidvegi, A. J. Szalai,D. E. Briles, J. E. Volanakis, R. A. Wetsel, and H. R. Colten. 1999. Geneticdisruption of the murine complement C3 promoter region generates deficientmice with extrahepatic expression of C3 mRNA. Immunopharmacology 42:135.
40. Pratt, J. R., S. A. Basheer, and S. H. Sacks. 2002. Local synthesis of complementcomponent C3 regulates acute renal transplant rejection. Nat. Med. 8:582.
41. Passwell, J., G. F. Schreiner, M. Nonaka, H. U. Beuscher, and H. R. Colten. 1988.Local extrahepatic expression of complement genes C3, factor B, C2, and C4 isincreased in murine lupus nephritis. J. Clin. Invest. 82:1676.
42. Sackstein, R., and H. R. Colten. 1984. Molecular regulation of MHC class III (C4and factor B) gene expression in mouse peritoneal macrophages. J. Immunol.133:1618.
43. Lagasse, E., and I. L. Weissman. 1996. Flow cytometric identification of murineneutrophils and monocytes. J. Immunol. Methods 197:139.
44. Masuda, A. 1988. Immunohistochemical study of Warthin’s tumor with specialregard to the germinal center. Histol. Histopathol. 3:81.
45. Schwaeble, W., M. K. Schafer, F. Petry, T. Fink, D. Knebel, E. Weihe, andM. Loos. 1995. Follicular dendritic cells, interdigitating cells, and cells of themonocyte-macrophage lineage are the C1q-producing sources in the spleen: iden-tification of specific cell types by in situ hybridization and immunohistochemicalanalysis. J. Immunol. 155:4971.
46. Taylor, P. R., M. C. Pickering, M. H. Kosco-Vilbois, M. J. Walport, M. Botto,S. Gordon, and L. Martinez-Pomares. 2002. The follicular dendritic cell restrictedepitope, FDC-M2, is complement C4; localization of immune complexes inmouse tissues. Eur. J. Immunol. 32:1888.
47. Kim, Y. U., M. C. Carroll, D. E. Isenman, M. Nonaka, P. Pramoonjago,J. Takeda, K. Inoue, and T. Kinoshita. 1992. Covalent binding of C3b to C4bwithin the classical complement pathway C5 convertase: determination of aminoacid residues involved in ester linkage formation. J. Biol. Chem. 267:4171.
48. Brubaker, J. O., C. M. Thompson, L. A. Morrison, D. M. Knipe, G. R. Siber, andR. W. Finberg. 1996. Th1-associated immune responses to �-galactosidase ex-pressed by a replication-defective herpes simplex virus. J. Immunol. 157:1598.
49. Meraz, M. A., J. M. White, K. C. Sheehan, E. A. Bach, S. J. Rodig, A. S. Dighe,D. H. Kaplan, J. K. Riley, A. C. Greenlund, D. Campbell, et al. 1996. Targeteddisruption of the Stat1 gene in mice reveals unexpected physiologic specificity inthe JAK-STAT signaling pathway. Cell 84:431.
50. Mitchell, T. J., M. Naughton, P. Norsworthy, K. A. Davies, M. J. Walport, andB. J. Morley. 1996. IFN-� up-regulates expression of the complement compo-nents C3 and C4 by stabilization of mRNA. J. Immunol. 156:4429.
5495The Journal of Immunology
by guest on October 7, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
Related Documents