Inflammation & Allergy - Drug Targets, 2011, 10, 000-000 1 1871-5281/11 $58.00+.00 © 2011 Bentham Science Publishers Ltd. Immune Response Towards Snake Venoms Guillermo León * , Laura Sánchez, Andrés Hernández, Mauren Villalta, María Herrera, Álvaro Segura, Ricardo Estrada and José María Gutiérrez Instituto Clodomiro Picado, Facultad de Microbiología, Universidad de Costa Rica, San José, Costa Rica Abstract: The immune response involves a complex repertoire of innate and adaptive responses to foreign agents in the organism. The present review focuses on the immune response to snake venoms, including those occurring in snakebite accidental envenomation, experimental vaccination and animal hyperimmunization for snake antivenom production. The following aspects are considered: (a) the structural characteristics of snake toxins and their relationship to immunogenicity, (b) the effects that factors such as administration route, venom dose, type of adjuvant, and individual and species characteristics of the immunized animal have on the immune response, (c) the initial venom-induced inflammatory response, (d) the process by which specific antibodies towards individual toxins are produced, and (e) the techniques currently used to evaluate the antibody response. Understanding the immune response to snake venoms is highly relevant for improving antivenom production and for gaining a more complete view of snakebite envenoming. Keywords: Snake venoms, immunogens, immunization, immune response, antivenoms. 1. INTRODUCTION Snake venoms are complex mixtures which play adaptive roles associated with prey immobilization and digestion, as well as with defense against predators [1-3]. Snake venom evolution has involved the recruitment of diverse genes and their duplication and expression in venom glands [2]. Proteomic analysis of snake venoms has demonstrated the high complexity of these secretions [2, 4, 5]. Toxins in snake venoms correspond to proteins of both enzymatic and non- enzymatic nature, which exert many different deleterious effects in the organism. In addition, snake venoms comprise a number of proteins which do not exert relevant toxic effects [3]. Snakebite envenomation is an important public health problem on a global basis [6-9]. Snake species responsible for the vast majority of envenomations in humans are classified within the families Elapidae and Viperidae [7, 10]. In addition, species belonging to the families Atractaspididae and Colubridae (sensu lato) can also induce clinically- relevant envenomations [10-14]. Upon snake venom injection, two simultaneous processes occur in the organism: (a) The development of toxic effects induced by the venom toxins, and (b) the stimulation of innate and adaptive immune responses aimed at neutralizing and removing venom proteins. When the ability of snake venom to generate local and systemic damage exceeds the capacity of the animal to assimilate and respond towards the aggression, an envenomation develops. The only scientifically-validated therapy to confront snakebite envenomation is based on the parenteral administration of antivenoms [15-18]. Antivenoms are preparations of animal derived immunoglobulins, or *Address correspondence to this author at the Instituto Clodomiro Picado, Facultad de Microbiología, Universidad de Costa Rica, San José, Costa Rica; Tel: 506-22293135; Fax: 506-22920485. E-mail: [email protected] immunoglobulin fragments, capable of binding to the venom toxins and of neutralizing their toxic effects. Their production involves the immunization of animals with snake venoms to stimulate the generation of antivenom antibodies. Then, whole immunoglobulins, or their F(ab´) 2 or Fab fragments, are purified, formulated, sterilized and dispensed in vials, as required for parenteral biopharmaceutical products [18-20]. The immune response towards snake venoms depends on the physicochemical characteristics of individual toxins, such as structure and molecular mass, on their relative abundance in the venom, on the dose of venom that is administered, and on the host factors that determine the capacity of the immune system to recognize each toxin as foreign, such as venom administration route, individual genetic background of the immunized animal, and ability to process the toxins [21]. Hereby we review various issues related with the immunogenic properties of snake venom toxins, the processes involved in the stimulation of the immune system of animals to produce an antibody-based response towards venom components, and the techniques currently used to evaluate this response. 2. SNAKE VENOMS AS IMMUNOGENS Antigens are substances capable of eliciting an immune response associated with their interaction with cells of the immune system, mostly lymphocytes and various antigen- processing cells. The ability of antigens to induce an immune response is called immunogenicity [22]. From this point of view, snake venoms have two types of components: (a) Immunogenic components, which are proteins that constitute more than 98% of the venom dry weight and include toxins able to induce relevant toxic effects as well as non-toxic proteins, and (b) non-immunogenic components, such as amino acids, nucleotides, carbohydrates, lipids and biogenic amines, which constitute less than 2% of the venom dry weight and, in general terms, are devoid of toxicity [1, 3].

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Inflammation & Allergy - Drug Targets, 2011, 10, 000-000 1
1871-5281/11 $58.00+.00 © 2011 Bentham Science Publishers Ltd.
Immune Response Towards Snake Venoms
Guillermo León*, Laura Sánchez, Andrés Hernández, Mauren Villalta, María Herrera, Álvaro Segura, Ricardo Estrada and José María Gutiérrez
Instituto Clodomiro Picado, Facultad de Microbiología, Universidad de Costa Rica, San José, Costa Rica
Abstract: The immune response involves a complex repertoire of innate and adaptive responses to foreign agents in the organism. The present review focuses on the immune response to snake venoms, including those occurring in snakebite accidental envenomation, experimental vaccination and animal hyperimmunization for snake antivenom production. The following aspects are considered: (a) the structural characteristics of snake toxins and their relationship to immunogenicity, (b) the effects that factors such as administration route, venom dose, type of adjuvant, and individual and species characteristics of the immunized animal have on the immune response, (c) the initial venom-induced inflammatory response, (d) the process by which specific antibodies towards individual toxins are produced, and (e) the techniques currently used to evaluate the antibody response. Understanding the immune response to snake venoms is highly relevant for improving antivenom production and for gaining a more complete view of snakebite envenoming.
Keywords: Snake venoms, immunogens, immunization, immune response, antivenoms.
1. INTRODUCTION
Snake venoms are complex mixtures which play adaptive roles associated with prey immobilization and digestion, as well as with defense against predators [1-3]. Snake venom evolution has involved the recruitment of diverse genes and their duplication and expression in venom glands [2]. Proteomic analysis of snake venoms has demonstrated the high complexity of these secretions [2, 4, 5]. Toxins in snake venoms correspond to proteins of both enzymatic and non-enzymatic nature, which exert many different deleterious effects in the organism. In addition, snake venoms comprise a number of proteins which do not exert relevant toxic effects [3].
Snakebite envenomation is an important public health problem on a global basis [6-9]. Snake species responsible for the vast majority of envenomations in humans are classified within the families Elapidae and Viperidae [7, 10]. In addition, species belonging to the families Atractaspididae and Colubridae (sensu lato) can also induce clinically-relevant envenomations [10-14].
Upon snake venom injection, two simultaneous processes occur in the organism: (a) The development of toxic effects induced by the venom toxins, and (b) the stimulation of innate and adaptive immune responses aimed at neutralizing and removing venom proteins. When the ability of snake venom to generate local and systemic damage exceeds the capacity of the animal to assimilate and respond towards the aggression, an envenomation develops.
The only scientifically-validated therapy to confront snakebite envenomation is based on the parenteral administration of antivenoms [15-18]. Antivenoms are preparations of animal derived immunoglobulins, or
*Address correspondence to this author at the Instituto Clodomiro Picado, Facultad de Microbiología, Universidad de Costa Rica, San José, Costa Rica; Tel: 506-22293135; Fax: 506-22920485. E-mail: [email protected]
immunoglobulin fragments, capable of binding to the venom toxins and of neutralizing their toxic effects. Their production involves the immunization of animals with snake venoms to stimulate the generation of antivenom antibodies. Then, whole immunoglobulins, or their F(ab´)2 or Fab fragments, are purified, formulated, sterilized and dispensed in vials, as required for parenteral biopharmaceutical products [18-20].
The immune response towards snake venoms depends on the physicochemical characteristics of individual toxins, such as structure and molecular mass, on their relative abundance in the venom, on the dose of venom that is administered, and on the host factors that determine the capacity of the immune system to recognize each toxin as foreign, such as venom administration route, individual genetic background of the immunized animal, and ability to process the toxins [21].
Hereby we review various issues related with the immunogenic properties of snake venom toxins, the processes involved in the stimulation of the immune system of animals to produce an antibody-based response towards venom components, and the techniques currently used to evaluate this response.
2. SNAKE VENOMS AS IMMUNOGENS
Antigens are substances capable of eliciting an immune response associated with their interaction with cells of the immune system, mostly lymphocytes and various antigen-processing cells. The ability of antigens to induce an immune response is called immunogenicity [22]. From this point of view, snake venoms have two types of components: (a) Immunogenic components, which are proteins that constitute more than 98% of the venom dry weight and include toxins able to induce relevant toxic effects as well as non-toxic proteins, and (b) non-immunogenic components, such as amino acids, nucleotides, carbohydrates, lipids and biogenic amines, which constitute less than 2% of the venom dry weight and, in general terms, are devoid of toxicity [1, 3].

2 Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 León et al.
Recently, the use of proteomic tools has allowed a detailed characterization of the protein composition of a variety of snake venoms, a field known as ‘venomics’ [4]. A number of studies have characterized the venoms of viperid species of the genera Sistrurus [23], Cerastes [24], Bitis [25], Atropoides [26], Bothriechis [27], Lachesis [28], Vipera [29], Bothrops [30-35], Crotalus [36], Echis [37] and Daboia [38]. Moreover, the venomics of some species of the families Elapidae and Colubridae have been also investigated, including those of the genera Micrurus [39], Naja [40] and Philodryas [41].
From these venomic studies it has been shown that, although viperid snake venoms are composed of dozens of proteins, all of them belong to a small group of protein families, including Zn2+-depending metalloproteinases (SVMPs), serine proteinases, phopholipases A2 (PLA2), L-amino acid oxidases, disintegrins, C-type lectins and lectin-like proteins, natriuretic peptides, cysteine-rich secretory proteins (CRISPs) and Kunitz-type protease inhibitors [42]. Each protein family has structural features, i.e. molecular mass, tertiary and quaternary structures, degree of glysosylation and other post-translational modifications, which determine their immunogenicity. In addition, different proteins are present in variable concentrations in the venoms, a factor that also influences their immunogenicity. The basic structural features and toxic effects of the most abundant snake venom components are reviewed in the following subsections.
2.1. Metalloproteinases (SVMPs)
SVMPs are Zn2+-depending enzymes present in the venoms of snakes of the families Viperidae [42], Elapidae [43], Atractaspididae [44] and Colubridae [45]. SVMPs constitute 30-60% of the total protein content in the venoms of viperids [42]. According to their domain composition, SVMPs are classified in three groups: (a) P-I SVMPs, which have molecular masses of 20-30 kDa, and are composed by a single Zn2+-binding metalloproteinase domain (catalytic domain); (b) P-II SVMPs, with molecular masses of 30-60 kDa and having a disintegrin domain in addition to the catalytic domain; and (c) P-III SVMPs, of molecular masses of 50-70 kDa and comprised by metalloproteinase, disintegrin-like and cysteine-rich domains. Some P-III SVMPs present a quaternary structure, in which a subunit comprising a typical P-III SVMP structure is linked, through a disulfide bond, to a C-type lectin-like smaller subunit [46]. In general, P-I SVMPs are not glycosylated whereas P-III SVMPs are glycosylated [46].
SVMPs play various roles in the pathophysiology of envenomation. The most notorious effect induced by SVMPs is hemorrhagic activity, an effect associated with prominent local tissue damage [47, 48] and with systemic bleeding which may lead to cardiovascular shock [49]. Such hemorrhagic activity is due to the action of SVMPs on the basement membrane that surrounds capillary vessels [48]. In general, P-III SVMPs are more potent hemorrhagic toxins than P-I SVMPs, the former inducing systemic hemorrhage whereas the latter are mostly responsible for local hemorrhage [48, 50, 51]. SVMPs also affect hemostasis, by inducing defibrinogenation through activation of factor X and prothrombin [52], or as a consequence of their
fibrinogenolytic action [53, 54]. In addition, SVMPs also participate in extracellular matrix degradation and promote an inflammatory reaction [55]. Interestingly, some SVMPs are species-specific, such as hannahtoxin from the elapid Ophiophagus hanna, that induces hemorrhage in rabbits but not in mice [56]. The immunogenicity of SVMPs greatly depends on their structural features, since high molecular mass P-III SVMPs generate a higher antibody response than P-I SVMPs [57].
2.2. Serine Proteinases
Serine proteinases have molecular masses of 25-70 kDa and are characterized by a conserved catalytic triad (Ser, His and Asp) and by the presence of abundant disulfide bonds [58]. Most venom serine proteinases show a variable extent of glycosylation [59, 60], a structural feature that influences immunogenicity. Serine proteinases constitute between 10-30% of the protein content in the whole venom in viperid species [42].
Snake venom serine proteinases exert a number of activities. Some of them are thrombin-like enzymes which cleave fibrinopeptides A or B from fibrinogen, causing the formation of microclots which are readily degraded by the fibrinolytic system, thus contributing to defibrinogenation [54, 61, 62]. Other serine proteinases are kallikrein-like enzymes [62-64] that generate bradykinin and contribute to hypotension [64, 65]. Some serine proteinases from viperid snake venoms are able to bind to thrombin receptors in platelets, inducing aggregation, whereas others are plasminogen activators [66]. Elapid venoms contain serine proteinases which activate prothrombin, thus contributing to the hemostatic disturbances characteristic of these envenomations [67].
2.3. Phospholipases (PLA2)
Snake PLA2 belong to the superfamily of secreted PLA2s. These enzymes are low molecular mass proteins (14-18 kDa) classified within classes I (elapid venoms) and II (viperid venoms) of secreted PLA2s [68]. The catalytic activity requires micromolar concentrations of Ca2+ [69], which is bound to a region formed by the carbonyl groups of Tyr28, Gly30 and Gly32 and by the carboxylic group of Asp49 [70]. Viperid snake venoms present a group of PLA2 homologues in which the invariant residue Asp49 is substituted by Lys or other residues [71]. This substitution abolishes enzymatic activity, although these proteins conserve toxicity [71]. Venom PLA2s may occur in multimeric structures, including dimeric, trimeric and pentameric molecules [72-75]. The presence of multi-subunit PLA2s has implications in terms of toxicity and immunogenicity [75] since it is likely that multimeric PLA2s are more immunogenic than their monomeric counterparts.
In viperid snake venoms, PLA2 content varies between 5-45% depending on the species [42]. PLA2s play highly diverse roles in envenomation, owing to a process of accelerated evolution that has resulted in the modification of surface residues, thus allowing these enzymes to bind to many different targets [76]. Consequently, PLA2s induce neurotoxicity, local and systemic myotoxicity, hypotension, anticoagulation, inhibition of platelet aggregation and

Immune Response Towards Snake Venoms Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 3
hemolysis among other effects [68, 77, 78]. Many of these activities result from the ability of PLA2s to hydrolyze phospholipids, although many effects are catalytically-independent and result from the interaction of PLA2s with receptors in plasma membranes of with proteins in plasma [76].
2.4. L-amino Acid Oxidase
L-amino acid oxidase (LAO) is a widely distributed enzyme in the venoms of many snake species classified in several families [79]. Its abundance varies from one species to another, from 30% in Calloselasma rhodostoma venom [80], to less than 1% in Bungarus fasciatus venom [81]. Snake venom LAOs are generally found as homodimeric glycoproteins with molecular mass of 110-150 kDa [82], although some monomeric LAOs have been isolated [81]. LAOs have been related with venom activities such as induction of apoptosis [83], inhibition of platelet aggregation [82], antimicrobial activity [84] and edema formation [85].
2.5. Disintegrins
Disintegrins are components widely distributed in the venoms of viperid species [42]. They represent between 2-10% of the whole venom in a number of viperids [42]. Disintegrins are rich in disulfide bonds and can be isolated as either monomers or dimers, or as fragments containing additional domains, such as the cysteine-rich domain characteristic of SVMPs [86, 87]. According to their length and the number of disulfide bonds, the disintegrin family has been divided into five different groups: (a) Short disintegrins composed of 41–51 residues and four disulfide bonds; (b) medium-sized disintegrins containing about 70 amino acids and six disulfide bonds; (c) long disintegrins with approximately 84-residues, cross-linked by seven disulfide bonds; (d) P-III disintegrins, that are modular proteins containing an N-terminal disintegrin-like domain of about 100 amino acids including 16 cysteine residues involved in the formation of eight disulfide bonds, and a C-terminal 110–120-residue cysteine-rich domain crosslinked by six disulfides; these disintegrins are derived from P-III SVMPs; and (e) dimeric disintegrins that are homo- and heterodimers containing subunits of about 67 residues with 10 cysteines involved in the formation of four intra-chain disulfide bonds and two interchain cystine linkages [87, 88]. The secondary structure of disintegrins is based on a series of antiparallel -sheets with the region interacting with integrins located at the connection loop [89, 90].
From a functional standpoint, disintegrins bind to integrins in cell membranes, thus exerting a number of effects depending on the cell type [87]. Some disintegrins, especially those containing the sequence Arg-Gly-Asp, are potent inhibitors of platelet aggregation through the binding of integrin IIB 3 [61, 91]. However, the role of disintegrins in the pathophysiology of envenomation remains largely unknown.
2.6. C-Type Lectins and Lectin-Like Proteins
C-type lectins (CTLs) are distributed in the venoms of species of the families Viperidae [1], Elapidae [92] and
Colubridae [41]. They have a molecular mass of ~30 kDa and a relative abundance, in viperid venoms, between 0.1 % and 24 % [42]. CTLs are composed of monomers linked by disulfide bonds. The snake venom C-type lectins include two subgroups: (a) true lectins with carbohydrate recognition domains (CRDs) that are capable of binding carbohydrates [93, 94-96], and (b) C-type lectin-like proteins (CLPs) with incomplete CRDs and, therefore, unable to bind carbohydrates. Instead, they present other biological activities such as induction or inhibition of platelet aggregation, induction of platelet agglutination, and inhibition of coagulation through their binding to clotting factors [58, 97-99]. CLPs are usually heterodimeric structures ( and subunits) which often oligomerize to form larger molecules [97].
2.7. Three-Finger Toxins
Snake venoms, particularly those from species of the family Elapidae, present a family of non-enzymatic low molecular mass components characterized by a common structure of three stranded loops extending from a central core [100]. The molecular evolution of this family of toxins has generated proteins with a wide variety of toxic effects, particularly neurotoxicity. Within this family there are -neurotoxins, -neurotoxins, fasciculins, muscarinic toxins and cardiotoxins or cytotoxins, among others [100]. The medically most relevant representatives are -neurotoxins, which induce a post-synaptic blockade at the neuromuscular junctions, as a consequence of their stong binding to the acetylcholine receptor at the motor end-plate, thus provoking muscle paralysis [1, 100]. The low molecular mass of these proteins (6-9 kDa) represents a problem for the generation of antibodies, owing to their low immunogenicty.
2.8. Other Venom Components
In addition to the family proteins listed above, snake venoms present other components whose role in toxicity has not been demonstrated for a number of them. Species of the family Atractaspididae present sarafotoxins in their venoms, which are potent cardiotoxic components of low molecular mass [101]. Owing to their small size, they are poor antigens and, therefore, raising antibodies against these proteins is a difficult task. Many snake venoms contain cysteine-rich secretory proteins (CRISPs) whose role in envenomation is not clear at present [102]. Species of the genus Dendroaspis, known as mambas, contain low molecular mass toxic components known as dendrotoxins and fasciculins [1, 103]. In addition, snake venoms contain other components such as nucleotidases, hyaluronidases, acetylcholinesterases, natriuretic factors, vascular endothelial growth factors, nerve growth factors, and bradykinin-potentiating peptides [3]. Some of these components play toxic roles of diverse sort, whereas others are apparently devoid of toxicity. The immunogenicity of these proteins varies significantly, as there are highly immunogenic non-toxic components and poorly-immunogenic toxic and non-toxic components. Thus, when an animal is immunized with crude venoms, the repertoire of antibodies generated may contain irrelevant antibodies against non-toxic components as well as neutralizing antibodies against toxic proteins.

4 Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 León et al.
2.9. Inactivated Snake Venoms
Humans or animals immunized with snake venoms can suffer pathophysiological alterations due to the toxic effects induced by venom toxins [104]. In order to avoid this problem during immunization for antivenom production, several procedures for venom detoxification have been developed. Some of them use chemical agents such as carboxymethyl cellulose [105], photooxidation in the presence of methylene blue [106], photo-oxidation [107], chelation [108, 109], formalin treatment [110, 111], iodination [112, 113], glutaraldehyde [114], and dihydrotioitic acid [115]. Others use physical agents such as X-ray radiation [116], ultraviolet radiation [117], simple heating treatment [118], gamma radiation [119-121], and ionizing radiation [122]. The goal of these treatments is to abrogate, or reduce, toxicity while maintaining immunogenicity.
The principal disadvantage of immunization using detoxified venoms is the potential risk of loosing important epitopes, thus affecting venom immunogenicity and provoking the failure of the anti-detoxified venom antibodies to neutralize the native venom [15, 17]. For example, it was found that some fractions of the Daboia russelli venom reduce their immunogenicity after detoxification with gamma radiation [119]. Thus, care should be taken when detoxifying venoms for immunization as to ensure that immunogenicity is not being hampered.
In addition to venom detoxification process, conformational epitopes from some toxins can be lost due to the action of proteinases present in the venoms [123]. If care is not taken when venom is collected and processed for storage, i.e. during centrifugation, freezing and lyophilization, the action of these proteinases may induce degradation and, consequently, may affect immunogenicity. Likewise, if venom solutions for immunization are left at room temperature for prolonged time intervals, proteolytic degradation may occur, especially in viperid venoms. Therefore, the processing of venom after collection should be carefully performed following validated protocols.
3. RECOGNITION OF SNAKE TOXINS BY THE
IMMUNE SYSTEM
Snake toxins have physicochemical characteristics that, besides conferring them with the capacity to induce toxic effects, allow their recognition by the immune system of the host as foreign molecules and, therefore, elicit an immune response. In addition, depending on how the toxins are presented to the immune system, drastically different antibody responses can be obtained. The generation of an antibody response effective to recognize and neutralize toxic proteins of venoms is the key element in the production of effective antivenoms in animals. Thus, the study of the parameters that determine such immune response is of paramount importance in the design and manufacture of antivenoms.
3.1. Administration Route and Antigen Dose
Depending on the route followed by the venom before it interacts with the immune system, different organs and cell
populations can be stimulated, generating differences in the immune response towards venom proteins. For example, while recurrent exposition to aerosols of venom powder induce IgE-based responses, which has been related with allergic reactions in snake venom handlers [124], the intramuscular or subcutaneous injection of venom in snakebite victims induces an initial IgM-based response, followed by an IgG response that, years after the snakebite, is still detectable [125-128].
Snake venom immunization can be performed by injecting the whole dose of venom in a single site [104], or by administering the dose of venom divided in small parts, each one injected in a different site [129-132]. Generally, the subcutaneous route is used for immunization [104, 131], but the intradermal [130, 132] and the intramuscular [119, 132-134] routes are also used. Moreover, some investigators have tested the oral administration of venom encapsulated in liposomes as an alternative to traditional immunization strategies [135, 136].
After venom injection, some toxins are drained by the lymphatic vessels before reaching the vascular compartment [137, 138]. Along this course, toxins pass through lymph nodes, thus stimulating the immune system to produce antibodies. Using horse as an example, during the lymphatic drainage of venom components injected subcutaneously, they can stimulate dendritic cells, macrophages and B lymphocytes located in the retrofaringeal, submandibular, prescapular, subscapular or prefemoral lymph nodes. Stimulation of cells present in these lymph nodes depends on the anatomic site where the venom is injected (Fig. 1). Thus, the selection of the anatomic regions for injection and the route by which venom is administered greatly influence the immune response in the animal.
Besides the route of administration, the dose of venom injected is an important issue that influences the way in which the immune system is stimulated. While administration of low amounts of venom may produce undetectable responses [104], injection of high amounts of venom may cause damage to the animal [109]. In both cases there is a theoretical risk of inducing immune tolerance [139]. In the past, many antivenom producers followed immunization schedules based on the injection of large amounts of venoms, with deleterious consequences for the health of animals. Nowadays, immunization programs are generally based on the injection of low amounts of venom, thus precluding the development of pathophysiological alterations while stimulating a satisfactory immune response.
When immunization is performed with whole venom, it must be considered that not all toxins are present in the venom at the same concentration [42]. In addition, on a similar weight basis, variations in the molecular mass of toxins imply that a higher number of molecules of low molecular mass are injected as compared with high molecular mass components. Therefore, due to actual differences in toxin concentration and to variations in molecular mass, each toxin is administered at a different dose, thus affecting the way in which the immune system is stimulated. In some venoms, toxicologically-relevant components are present in low concentrations, thus complicating the generation of an adequate neutralizing immune response.

Immune Response Towards Snake Venoms Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 5
3.2. Nonspecific Initial Immune Response Associated
with Inflammation
Injection of snake venoms in mammalian tissues generates inflammation, a nonspecific defensive mechanism that constitutes an early response towards foreign agents [140]. In the case of viperid snake venoms, this process is characterized by a prominent edema, an inflammatory cell infiltrate and the generation of pain [140]. This inflammatory response is orchestrated by a diverse array of mediators, including eicosanoids, nitric oxide, cytokines, matrix metalloproteinases, kinins and complement, among others [140-142]. SVMPs and PLA2s have been shown to induce conspicuous inflammatory responses in experimental animals [55, 141, 143].
Initially, an increment in microvessel permeability occurs in the injured tissue, with the consequent extravasation of fluids, thus enriching the interstitial space with plasma proteins, some of which are involved in the inflammatory reaction, such as complement proteins [144, 145], whereas others, like antibodies or enzyme inhibitors, may interact and block the deleterious effects of toxins [146-150]. For instance, 2-macroglobulin inhibits several proteinases by forming covalent complexes with these enzymes, which are then removed by phagocytic cells [48]. In addition, the release of chemotactic substances from plasma proteins, resident tissue cells and damaged cells attracts inflammatory cells, mainy neutrophils and monocytes/macrophages, and
stimulates leukocytes and endothelial cells to express adhesion molecules required for leukocyte rolling, adherence and diapedesis [140, 151]. As a result, an abundant inflammatory cell infiltrate reaches the tissue locations where venom was injected.
During the first 24 hours after envenoming, a neutrophil-based inflammatory infiltrate predominates [152, 153]. Neutrophils are phagocytic cells that preferentially exert a microbicidal activity, also contributing to the removal of necrotic debris, setting the stage for reparative and regenerative processes [154]. Moreover, activation of neutrophils generates reactive oxygen species which may participate in further tissue alterations [155]. However, neutrophils are not antigen-presenting cells and, therefore, their participation in the antivenom antibody production is marginal. At later time intervals after venom injection, the early neutrophil-based inflammatory infiltrate is replaced by an infiltrate composed predominantly by mononuclear cells, such as macrophages [140, 152, 153]. Dendritic cells are also likely to arrive to the affected tissue at later time intervals. Unlike neutrophils, macrophages and dendritic cells are antigen presenting cells which actively participate in the adaptive immune response against foreign antigens, such as venom components [156].
It has been postulated that, owing to the conspicuous inflammatory response associated with snake venom injection, a number of these mediators may contribute to the tissue damage initiated by venom toxins [157]. Although this
Fig. (1). Association between the anatomic sites where venom is injected and the location of neighboring lymph nodes. Dendritic cells, macrophages and B lymphocytes located in the retrofaringeal (a), submandibular (b), prescapular (c), subscapular (d) or prefemoral (e) lymph nodes are stimulated by venom immunogenic components that are drained by the lymphatic vessels from the injection site before reaching the central vascular compartment. In the scheme, typical subcutaneous injection sites used to immunize horses to produce snake antivenoms are numbered from 1 to 4.

6 Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 León et al.
hypothesis has not been demonstrated for many mediators [158], the idea of reducing and modulating venom-induced inflammation as part of the therapy of these envenomations has been proposed. However, care should be taken as to ensure that inflammation-associated venom inhibition and promotion of immune response to venom components are not hampered by these interventions.
3.3. Production of Antivenom Antibodies
After injection of venom dissolved in saline solution, each toxin is differentially absorbed according to its toxicokinetic properties. For example, some toxins from the venoms of the elapid Walterinnesia aegyptia and the viperid Vipera aspis are drained from the injection site to the neighboring lymph nodes [137, 138], before reaching the intravascular compartment. In blood, snake toxins can be readily detected using immunoenzymatic techniques [159-161]. In contrast, after intramuscular injection of a myotoxic PLA2 from the venom of Bothrops asper, which predominantly induces local tissue damage, a large proportion of the injected protein remains in the injected tissue [162], although it is likely that lymphatic absorption also occurs. On the other hand, the molecular mass of toxins affects their toxicokinetics, since low molecular mass neurotoxins from elapid venoms are rapidly absorbed and reach the circulation more readily than large molecular mass enzymes typical of viperid venoms [137].
According to their toxicokinetic profile, different venom components may access different populations of antigen-presenting cells. Also, there are differences in the antigen presentation process associated to the cell lineage involved in antigen presentation, i.e. subsets of dendritic cells, macrophages or B lymphocytes [163]. Therefore, it is possible that the toxicokinetic profile of individual proteins has an effect on the characteristics of the immune response towards venom toxins, although this issue has not been investigated.
Venom toxins drained from the injection site by lymphatic vessels may interact with the specific membrane immunoglobulin receptors of resting B cells present in the neighboring lymph nodes. This interaction would induce an initial B cell activation, in which IgM antibodies are the predominant isotype of the secreted immunoglobulins [22]. In human victims of Bothrops jararaca snakebite, IgM towards venom components was detected since 3 days after envenomation and were maintained at low titres for 17 more days, until becoming undetectable [127]. In addition, venom toxins in lymph nodes can be endocyted by other antigen presenting cells, such as macrophages or dendritic cells [164].
After endocytosis, venom proteins are processed by the endocytic pathway, i.e. they go through early endosomes to endolysosomes, finally reaching the lysosomal compartment. During this path, snake venom proteins are degraded by hydrolytic enzymes into peptides which are coupled to class II MHC molecules and transported to the cell surface for antigen presentation [165]. The endocytic activity of macrophages can be affected by the action of venom components. For instance, exposition to sub-lethal doses of the venom of Crotalus durissus terrificus affects
macrophage function [118, 166, 167], whereas myotoxic PLA2s from the venom of Bothrops asper stimulate phagocytosis [168]. In the case of C. d. terrificus, this immunosuppressive effect is induced by the dimeric neurotoxic PLA2 crotoxin [169, 170], specifically by its phopholipase A2 subunit [171]. Clearly, the interplay between venom toxins and immune cells affects the outcome of the immune response.
On the other hand, venom toxins remaining around the injection site can be endocyted by the cells that compose the late inflammatory infiltrate such as macrophages and dendritic cells. These cells may migrate to the regional lymph nodes and might present these antigens to the TH cells [22].
Antigen presentation predominantly occurs in lymph nodes. Here, antigen presenting cells exhibit the peptides derived form the processed venom proteins coupled with type II MHC molecules to be recognized by the TCR-CD3 complex present in the membrane of specific TH cells. Conformational stability of the protein derived peptides is an important factor for TH cell stimulation [165]. In order to induce TH activation, the interaction between the oligopeptide-MHC II complex and the TCR-CD3 complex should occur simultaneously with a co-stimulatory signal which is provided by the interaction of the proteins of the B7 family expressed in the membrane of the B lymphocyte with the CD28 molecule present in the membrane of TH cells. As result, TH cells are induced to proliferate and differentiate into memory T cells and effector T cells [22].
Activated TH cells express CD40L in their membrane, which interacts with the CD40 molecule in the B lymphocyte membrane. In addition, activated TH cells release cytokines that stimulate B lymphocytes to proliferate and differentiate into memory B cells and plasma cells. The cytokine-mediated stimulation of immune response can be affected by the action of venom proteins, such as the case of elapid neurotoxins that bind the acetylcholine receptor [172, 173].
As a corollary of TH cell stimulation, plasma cells are induced to change the isotype of antibodies from IgM or IgD to IgG, IgA or IgE. Thus, the biological properties of the generated antibodies can vary without changing their specificity. During a first envenomation, antivenom IgGs appear after antivenom IgM antibodies, but reach higher plasma concentration [127] that persist long time after envenomation [125, 126, 128]. During a second exposition to the same snake venom, memory B cells are activated and produce IgG in levels that increase faster and reach higher titres than those occuring after the first venom exposition [127]. Recurrent exposition of animals to venom stimuli, associated with large increments in IgGs, can result in the reversal of the albumin/globulin ratio in serum [174, 175].
Antivenom antibodies present in immunized humans or animals may downregulate the antibody response produced in subsequent contacts with the venom [176, 177]. This downregulation might influence the production of antivenom antibodies, thus implying that different immunization schedules may affect the extent of antivenom antibody production in immunized animals [104]. In addition, antibodies can sequester the circulating snake venom toxins, thus reducing the antivenom antibody response. This can

Immune Response Towards Snake Venoms Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 7
explain why a number of snakebite envenomated patients who received antivenom did not develop a detectable antibody response against venom proteins [127].
3.4. Specificiy of the Immune Response Towards Snake
Venoms
Snake venoms are composed by proteins whose antigenicity and abundance changes from one species to another. However, some antigenic similarities are conserved among phylogenetically-related species [178-181]. Therefore, antibodies raised by immunization with a particular snake venom, in addition to reacting with its homologous toxins, can cross-react with some toxins present in heterologous venoms and may neutralize their toxic effects [178, 182-187]. For example, anti-Crotalus adamanteus immunoglobulins react stronger towards Crotalus atrox venom than towards Crotalus adamanteus venom itself [188].
On the other hand, not all cross-reactivity implies cross-neutralization [188, 189]. Antigenic variations are responsible that antibodies raised by immunization with PLA2 from Bothrops jararaca venom do not neutralize the effects produced by PLA2 from Bothrops alternatus or Bothrops erythromelas venoms [181]. Similar results were obtained with the antibodies raised against a PLA2 from Crotalus scutulatus salvini in the neutralization of PLA2 from Crotalus scutulatus scutulatus [190]. Also, limitations of the cross-neutralization have been demonstrated in the case of whole venoms [184, 189, 191, 192].
The specificity of antivenom antibodies towards a restricted group of venoms is a limitation of the immunotherapy of snakebite envenomation. This issue brings a number of implications for the design and use of antivenoms. First of all, the selection of the venoms used in the immunization mixtures has to be carefully analyzed, taking into consideration aspects such as the medical relevance of various species, the clinical presentation of the diverse envenomation syndromes and the immunological cross-reactivity of various venoms [193]. Thus, depending on these factors, monospecific or polyspecific antivenoms have to be designed. In Australia, for example, different antivenoms are produced, although the clinical features of envenomations do not always allow the identification of the offending species [194]. In this case, venom detection kits have been developed for the identification of the species and the correct selection of antivenom [194]. Similar attempts have been performed in other parts of the world [159-161].
3.5. Adjuvants
Adjuvants are substances used to increase the immune response towards immunogens [195]. In the case of vaccines, adjuvants are used to obtain a protective and lasting response. In the case of antivenom production, adjuvants are used to produce elevated titres of antibodies with high affinity [196]. Ideally, adjuvants must be effective at promoting an immune response, should generate minimal tissue damage on the immunized animal and should have a low cost [175, 197].
Adjuvants act by several mechanisms such as (a) formation of antigen deposits associated with slow release, (b) contribution to antigen presentation, or antigen targeting, and (c) stimulation of immune mechanisms associated with cell activation and cytokine synthesis [198]. The most used adjuvants for animal immunization with snake venoms are those that form deposits from which the immunogen is slowly released [196, 199], such as aluminum salts, bentonite, sodium alginate and incomplete Freund´s adjuvant [104, 174, 200-202]. In addition, Freund’s complete adjuvant is generally employed in the first immunization for antivenom production.
The adjuvant activity of aluminium salts is attributed to its ability to gradually release the immunogen from the injection site, increasing its exposition to the immune system. Moreover, the ‘particulate character’ acquired by the immunogen adsorbed on the crystal salt surface contributes to the activation of antigen-presenting cells [203]. The group of aluminum adjuvants includes compounds that, although sharing some physicochemical properties, differ regarding their immunomodulatory properties. Aluminum hydroxide is more effective than aluminum phosphate to induce high antibody titres towards Trimeresurus flavoviridis venom in guinea pigs [200]. Bentonite is an adsorptive colloid that interacts with proteins in a similar way than aluminum compounds. However, animals immunized with snake venoms using bentonite as adjuvant develop low titres of antivenom antibodies [204]. Probably, this is the reason why it is rarely used nowadays to produce antivenoms.
Incomplete Freund´s adjuvant is composed by mineral oil and an emulsifier agent such as arlacel A. The complete Freund´s adjuvant contains, in addition, heat-killed cells of Mycobacterium tuberculosis or M. butyricum. To prepare the immunogen, the snake venom is dissolved in an aqueous phase which is then dispersed in the oily base and mixed until a stable water-in-oil emulsion is obtained. While these adjuvants promote a slow release of the antigen, the presence of bacterial cells in the complete Freund stimulates the immune system for a better antigen presentation and cell activation. Mycobacterium cells activate macrophages enhancing their ability to endocyte venom proteins that are slowly released from the injected emulsion. Additionally, macrophages increase their expression of class II MHC and membrane molecules of the B7 family. As a result, the ability of macrophages to present antigens to TH cells is improved with this adjuvant.
Freund’s adjuvant was more effective than aluminum hydroxide to induce high titres towards Crotalus durissus terrificus venom in horses [201]. Moreover, when compared to bentonite and squalene/arlacel A, Freund’s adjuvant was more effective to induce high titres towards whole venoms or neurotoxic fractions from Naja naja siamensis (kaouthia), Ophiophagus hannah and Bungarus fasciatus in rats [202]. Thus, antivenom producers generally use Freund’s complete and incomplete adjuvants for the first immunization stages. A problem associated with Freund’s adjuvants is the fact that they may induce local tissue damage associated with granuloma formation. This problem has been circumvented by an immunization strategy based on the injection of small volumes in several anatomical sites [129].

8 Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 León et al.
Controlled release of venom using liposomes has been proposed as an adjuvant strategy for immunization. Mice, rabbits and sheep were immunized with the venom of the viperid Echis carinatus, encapsuled in osmium tetroxide-treated liposomes. Animals reached a high antibody response that allowed protection in animals challenged with one minimal lethal dose [135, 205]. Similar experiences have been performed using the venoms of Bothrops jararaca [206], Crotalus durissus terrificus, Crotalus durissus collilineatus [207], Echis ocellatus [136] and purified crotoxin from C. d. terrificus [208].
IMS 3012 is an aqueous based nanoparticle adjuvant that, in comparison with incomplete Freund´s adjuvant, induces higher neutralizing titres towards venoms of the Indian snakes Naja naja, Vipera ruselli, Bungarus caeruleus and Ecuis carinatus. Unlike Freund´s adjuvant, IMS 3012 did not induce significant tissue damage [175] and, therefore, represents a promising alternative for immunization for antivenom production.
As an alternative to the depot-forming adjuvants, substances that promote leukocyte stimulation from plant-derived extracts have been studied for their adjuvanticity. An enhancement of the antibody response of rabbits immunized with the venom of Vipera russelli was obtained bymixing the venom with 2-OH-4-MeO benzoic acid extracted from Hemidesmus indicus [209]. A similar result was obtained in mice immunized with Bothrops asper venom treated with an extract from Echinacea purpurea [210].
Regardless of the adjuvant used, the molecular mass of venom antigens greatly determines their antigenic properties. For instance, venom proteins of high molecular mass (> 70 kDa), such as P-III SVMPs and LAO, tend to be good immunogens. In contrast, components with lower molecular mass (5-30 kDa), such as elapid -neurotoxins, PLA2s, P-I SVMPs and disintegrins, have been shown to be less immunogenic [57, 129, 188, 211]. An exception to this is the venom of Ophiophagus hannah which has highly immunogenic components of ~25 kDa [192]. Toxin alkylation has been suggested as an effective strategy to increase the magnitude of the immune response towards some venom components of low molecular mass [212], although the risk of affecting epitopes by these modifications has to be carefully considered.
3.6. Immune Response in Vaccination with Snake
Venoms
In 1887, Henry Sewall proposed the possibility to develop immunity in pigeons repeatedly exposed to sublethal doses of Sistrurus catenatus tergeminus (formerly Crotalophorus tergeminus) snake venom. In comparison with non-vaccinated animals, immunization induced an increased resistance to the venom. However, when using low challenge doses of venom, individuals were affected by the action of venom and, at higher doses, they did not survive [213].
A major attempt to produce a vaccine against snake venom to be used in humans was performed by the group of Sawai in 1968. In this investigation, humans from the Japanese islands Amami and Rykyu were immunized with dihydrotioitic acid (DHTA)-detoxified venom of the Habu,
Trimeresurus flavoviridis. Although high titres of anti-venom antibodies were generated in the plasma of immunized subjects, antivenom antibody concentration did not reach the levels obtained after heterologous antivenom administration [214].
Immunizations using the proteinase-containing fractions HR-1 and HR-2 of T. flavoviridis venom were performed by Sadahiro and collaborators in 1978. They tested their immunogens in monkeys. Immunized animals developed high titres of anti-venom antibodies; however, when challenged with venom after immunization, only animals receiving a challenge dose lower than 5 mg venom/animal survived. It was concluded that the level of protection was not enough to have prophylactic effectiveness. On the other hand, although theoretical calculations suggested that 1.3-2.6 U/mL of anti HR-1 and HR-2 antibodies would be effective to abrogate the toxicity induced in a snakebite envenomation, in practice this level of antibodies was not enough to prevent the toxic manifestations induced by the venom and, therefore, antivenom administration was necessary [215].
Snake venoms are composed of a variety of components thus having a high biochemical and pharmacological complexity [3]. Accordingly, in contrast with diphthaeria and tetanus toxins, the attempts to produce effective and safe vaccines for human use have not been successful [15]. Therefore, neither exposition to venom in accidental snakebites nor immunization with detoxified venoms, i.e. ‘venoids’, generate an immune response that would protect the immunized person in an actual snakebite envenomation [128]. In the case of local tissue damage induced by Bothrops asper venom, which is poorly neutralized by the intravenous administration of antivenoms [216], it was shown that even in mice receiving the antivenom prior to venom injection, thus simulating the condition of vaccinated animals, reduction of the extent of myonecrosis and hemorrhage was achieved only to a partial extent [217].
3.7. Production of Hyperimmune Plasma to Antivenom
Production
Immunization of animals to produce snake antivenoms requires the consideration of many factors influencing the immune response towards snake venoms, some of which have been reviewed above. In addition, other factors affect this response, such as the health status of the immunized animals, the quality of the venoms used, the design of the mixture of venoms, and the immunization protocols [17, 18, 193, 218].
The immunogenic profile of snake venoms varies from one species to another. Moreover, there are variations in venom composition and antigenicity within the geographical range of a single species, as well as according to the age of the specimens [30, 219, 220]. Therefore, to ensure that an antivenom formulation is effective in neutralizing the effects of the venoms of specimens throughout the distribution range of a species, the venom pool used to immunize animals must be collected from a number of individuals representative of the population of snakes from a particular geographic zone. Such venom pools, used for immunization and quality control tests, should be characterized

Immune Response Towards Snake Venoms Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 9
biochemically and pharmacologically [221]. Thus, the correct selection, preparation and characterization of venom pools for immunization are of paramount importance in antivenom production.
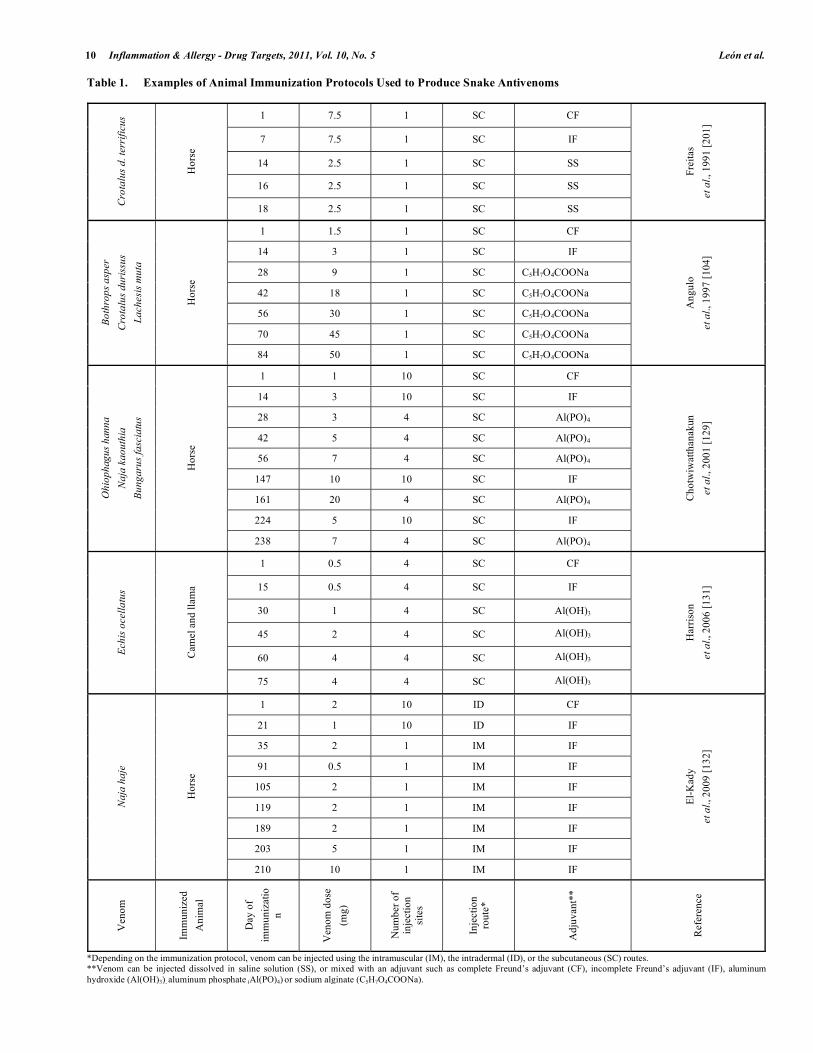
The production of animal hyperimmune plasma used for antivenom manufacture is based on the repeated injection of venoms following immunization protocols in which route of injection, venom dose, types of adjuvants and vehicles are detailed. Traditionally, the design of immunization protocols for antivenom production has been performed empirically, by evaluating the immunological and clinical responses resultant from venom injections. Because of this, there is a high diversity of immunization protocols reported in the literature, some of which are summarized in Table 1.
Immunization can be performed either with the venom of a single species or with venoms of several species, resulting in the production of monospecific or polyspecific antivenoms, respectively. While it has been argued that immunization with a venom mixture can enhance the antigenic repertoire, thus improving the antibody response [218] others propose that the use of a mixture of venoms might dilute relevant antigens and impair the immune response [193]. It is likely that, owing to the large variation in snake venom composition, no general conclusions can be reached on this issue, and that a case by case analysis has to be performed on an experimental basis. Nevertheless, the combination of venoms of highly different composition, such as elapid and viperid venoms, for immunization of animals, may complicate the development of a satisfactory immune response. In these cases, the preparation of mixtures of plasma derived from animals immunized with different venoms represents an alternative. Thus, antivenoms could be polyspecific, but individual animals are immunized with one or few venoms.
In some cases, the immune response towards a particular venom can be similar if it is injected alone or mixed with other venoms [222]. However, in other cases, a higher immune response was obtained when using venom of a single species than when using a mixture of venoms [189]. This apparent contradiction can be partially explained if one of the venoms included in the immunization mixture has an immunosupressive effect, such as the one described for the venoms of Crotalus durissus terrificus [118, 167] or Lachesis muta [223]. Thus, immunosupression by some venoms is another variable that has to be considered when designing venom mixtures for immunization.
DNA immunization has been suggested to stimulate the immune system to produce antivenom antibodies [224]. According to this strategy, cDNA encoding venom toxins are cloned into plasmids to generate constructs which are used to transform bacterial cells. The recombinant plasmids are purified from transformed bacterial cultures and used to coat tungsten microparticules which are intradermically injected into animals. An additional application of this strategy is based in the design of epitope string constructs in which relevant epitopes are encoded in plasmids and used for immunization [225]. By following variants of this general procedure, it has been possible to induce antibody production in animals towards venom toxins from Bothrops jararaca [226], Bothrops asper [227], Crotalus durissus
[228] and Naja kaouthia [229]. However, the use of DNA immunization to produce hyperimmune plasma for antivenom manufacture still requires more development.
3.8. Individual and Species Related Characteristics of the
Antibody Response
Immune response to snake toxins is influenced by the genetic background of the animal being immunized. Analysis of the response of people envenomated by Bothrops jararaca revealed a heterogeneous antibody profile in which each patient recognizes different venom components [127]. In the case of immunization for antivenom production, Angulo and co-workers documented significant differences, in the time-course of antibody response and in the final titre, in six horses injected with the venoms of Bothrops asper, Crotalus durissus and Lachesis muta. Similar differences were observed in the individual response of horses to different venoms [104]. These findings agreed with previous reports in which individual differences were demonstrated [174, 230, 231]. Due to such individual variation in the immune response, less than 20% of the horses immunized in Thailand with the Naja naja siamensis venom reached the antibody titre required to produce therapeutically effective antivenoms [202].
The majority of antivenoms are produced in horses, although some antivenom manufacturers use sheep [17, 18]. In addition, other species such as goats, camels, llamas, donkeys, hens and rabbits have been also used albeit to a more limited extent [17, 131, 232-234]. There are differences in the properties of imunoglobulins between species. In horses, the antibody responsible for most of the antivenom neutralizing activity corresponds to an immunoglobulin class traditionally known as IgG(T) [235-240]. In the current nomenclature, IgG(T) is classified as IgG3 and IgG5 [241].
Since equine IgG(T) is a highly glycosylated immunoglobulin, it has been suggested that it must have a higher immunogenicity than ovine derived IgG and, therefore, is likely to generate a higher incidence of adverse reactions, especially serum sickness [242, 243]. This hypothesis was tested in laboratory studies by different groups and contradictory results were obtained [244, 245]. Thus, the impact of the characteristics of IgG of several species on the issue of adverse reactions to antivenoms remains to be investigated.
The use of camelid immunoglobulins to formulate antivenoms has been proposed as an alternative to horses and sheep, especially for those regions where camels are common and adapt well, such as in northern Africa and the Middle East or, in the case of llamas, in the highlands of South America [131, 234, 246, 247]. Camelids immunized with snake venoms produce two types of antibodies: (a) heterotetrameric IgGs (~160 kDa), corresponding to the IgG1 subclass composed of two identical heavy chains and two identical light chains, and (b) homodimeric IgG (~100 kDa), known as heavy-chain antibodies, corresponding to the IgG2 and IgG3 subclasses composed only by two identical heavy chains without light chains [246, 248, 249]. All of the three IgG sub-classes are involved in the immune response to toxins and possess venom-neutralizing activity [248].
10 Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 León et al.
Table 1. Examples of Animal Immunization Protocols Used to Produce Snake Antivenoms
1 7.5 1 SC CF
7 7.5 1 SC IF
14 2.5 1 SC SS
16 2.5 1 SC SS
Cro
talu
s d
. te
rrif
icu
s
Hor
se
18 2.5 1 SC SS
Frei
tas
et
al.,
199
1 [2
01]
1 1.5 1 SC CF
14 3 1 SC IF
28 9 1 SC C5H7O4COONa
42 18 1 SC C5H7O4COONa
56 30 1 SC C5H7O4COONa
70 45 1 SC C5H7O4COONa
Bo
thro
ps
asp
er
Cro
talu
s d
uri
ssu
s
La
ch
esi
s m
uta
Hor
se
84 50 1 SC C5H7O4COONa
Ang
ulo
et
al.,
199
7 [1
04]
1 1 10 SC CF
14 3 10 SC IF
28 3 4 SC Al(PO)4
42 5 4 SC Al(PO)4
56 7 4 SC Al(PO)4
147 10 10 SC IF
161 20 4 SC Al(PO)4
224 5 10 SC IF
Oh
iop
hag
us
han
na
Na
ja k
ao
uth
ia
Bu
ng
aru
s fa
scia
tus
Hor
se
238 7 4 SC Al(PO)4
Cho
twiw
atth
anak
un
et
al.,
200
1 [1
29]
1 0.5 4 SC CF
15 0.5 4 SC IF
30 1 4 SC Al(OH)3
45 2 4 SC Al(OH)3
60 4 4 SC Al(OH)3
Ech
is o
cell
atu
s
Cam
el a
nd ll
ama
75 4 4 SC Al(OH)3 H
arri
son
et
al.,
200
6 [1
31]
1 2 10 ID CF
21 1 10 ID IF
35 2 1 IM IF
91 0.5 1 IM IF
105 2 1 IM IF
119 2 1 IM IF
189 2 1 IM IF
203 5 1 IM IF
Na
ja h
aje
Hor
se
210 10 1 IM IF
El-
Kad
y
et
al.,
200
9 [1
32]
Ven
om
Imm
uniz
ed
Ani
mal
Day
of
imm
uniz
atio
n
Ven
om d
ose
(mg)
Num
ber
of
inje
ctio
n si
tes
Inje
ctio
n ro
ute*
Adj
uvan
t**
Ref
eren
ce
*Depending on the immunization protocol, venom can be injected using the intramuscular (IM), the intradermal (ID), or the subcutaneous (SC) routes. **Venom can be injected dissolved in saline solution (SS), or mixed with an adjuvant such as complete Freund’s adjuvant (CF), incomplete Freund’s adjuvant (IF), aluminum hydroxide (Al(OH)3), aluminum phosphate (Al(PO)4) or sodium alginate (C5H7O4COONa).

Immune Response Towards Snake Venoms Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 11
Camelid immunoglobulins can be purified by several techniques used to produce antivenoms [131, 244]. Moreover, formulations obtained by these methods are effective in the neutralization of the local effects induced by the venom of Echis ocellatus [131] and Bothrops mattogrossensis [234]. In addition, camelid plasma contains a non-IgG protein which is a potent inhibitor of hemorrhagic activity of some viperid venoms [131].
Hens have been immunized with the venoms of snakes of the genera Daboia [133], Echis [134], Bitis [250], Bungarus [251], and Naja [250, 251], and represent a potential source of antibodies for antivenom production. Hens respond towards the snake venoms producing an immunoglobulin known as IgY which can be purified from the egg yolks by caprylic acid precipitation [252, 253], dilution and salting-out precipitation [134, 250] or dilution and chromatography [133]. IgY is a ~180 kDa protein composed by two heavy chains and two light chains. In addition, a shortened IgY of ~120 kDa has been described in anseriform birds such as ducks and geese [254].
Proponents of hens for the production of antivenom argue that mammalian immunoglobulins have less genetic variability than IgY antibodies, and that mammalian-derived antivenoms usually have a higher content of non-IgG proteins than antivenoms obtained from egg yolk IgY, thus having a higher probability of inducing adverse reactions to antivenom therapy than avian-derived antivenoms [133, 134]. However, the plasma of human normal volunteers presents a higher titre of heterophilic anti-IgY antibodies than anti-equine or anti-cattle IgG antibodies [255]. This finding predicts a major incidence of adverse reactions to hen-derived antivenom than to mammalian-derived antivenom. Controlled clinical trials are required to test these hypotheses.
4. EVALUATION OF THE ANTIBODY RESPONSE
TOWARDS SNAKE VENOMS
The assays used to assess the antibody response towards snake venoms can be classified as (a) In vivo assays, which evaluate the ability of antivenom antibodies to neutralize different toxic effects induced by the venom in animal models, and (b) in vitro assays, which evaluate the ability of the antivenom to form immune complexes with venom proteins, as tested by immunochemical assays or by the in vitro neutralization of enzymatic activities of the venom.
Antibody response to venom immunization can be evaluated in animal models, usually mice, by two principal strategies: (a) Preincubation of venom and serum prior to testing: A challenge dose of venom is mixed with the serum of the immunized animal, and the mixture is incubated before testing in animals, or (b) independent injection of venom and antivenom: A challenge dose of venom is injected in the test animal and then the serum is administered. Controls consist of animals treated with venom mixed with saline solution instead of serum or, in the case of independent injection, saline solution is administered after envenomation instead of serum [216, 256].
Depending on the model, different injection routes can be used. In general, the intravenous route is employed [17], although some laboratories use the intraperitoneal route [17,
257], especially when dealing with snake venoms that have coagulant activity. Usually, in-process and final quality control tests assess the neutralization of the lethal effect of snake venoms [17]. However, since the pathophysiology of envenomation includes other toxic activities in addition to lethality, such as hemorrhagic, coagulant, defibrinogenating and necrotizing activities, the assessment of the development of immune response to venoms should include the neutralization of these activities [184, 257, 258], especially when a new antivenom is being developed Owing to the problems associated with in vivo tests, neutralization assays are characterized by low reproducibility and high frequency of invalid tests, i.e. lethality neutralization assay [259]. In spite of this, the neutralization of lethal activity of venoms by the sera, or purified antibodies, of immunized animals remains as the gold standard for assessing antibody development in antivenom-producing laboratories.
Following the current global trend to replace in vivo assays for in vitro tests, efforts have been performed to substitute the use of mice by a variety of alternative assays [17, 260, 261]. A number of successful attempts have been based on the use of neutralization of enzymatic activities [262], passive haemagglutination [263], ELISA [264], or HPLC [265], among others.
Some researchers have used immunoassays to assess the antivenom antibody titres [264, 266, 267]. A significant correlation between ELISA titre and anti-lethal ED50 was found using a competitive assay and Bothrops alternatus venom [268]. However, the correlation of ELISA with the lethality neutralization assay using venoms of Micrurus nigrocinctus [269], Bothrops jararaca and Crotalus durissus terrificus [267, 270] was lower than that obtained by Rial and co-workers [268]. Therefore, owing to the large variability in snake venom composition, and to the predominance of different toxins in the lethality assay, no generalizations can be made on the potential usefulness of imunoassays in the assessment of antivenom potency, and specific designs should be tested for each venom-antivenom system.
In recent years, proteomic techniques have been used to determine the snake venom components that are recognized by antivenoms, a field known as ‘antivenomics [27, 42, 193]. The first stage in antivenomic analysis is the identification of the toxins that are present in the venom. For this, venom proteins are separated using reverse-phase HPLC. Then, the separated fractions are analyzed by SDS-PAGE. The N-termnal sequences of the toxins are used to determine the protein family to which they belong. If there is heterogeneity or the N-termini are blocked, electrophoretic bands are subjected to in-gel tryptic digestion and the obtained peptides are analyzed by MALDI-TOF mass fingerprinting and determination of the amino acid sequence of peptide ions by collision-induced dissociation tandem mass spectrometry [4, 193].
For the antivenomic analysis, venoms are incubated with antivenoms and the immunocomplexes are then removed by centrifugation. Thus, non-immunodepleted venom proteins remain in the supernatant and are identified by the proteomic analysis described above. Antivenomic analysis of several viperid venoms indicates that the molecular mass plays a key role in the immunogenicity of snake toxins. In the case of

12 Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 León et al.
venoms from Bothrops species, venom components of molecular mass higher than 30 kDa, such as P-II and P-III SVMPs, L–amino acid oxidase and serine proteinases, are readily recognized and immunoprecipitated by antivenom antibodies. In contrast, venom components of less than 20 kDa, such as PLA2 are usually immunodepleted only to a partial extent [57, 271]. Moreover, venom components of less than 5 kDa, such as some disintegrins and bradykinin-potentiating peptides (BPPs), do not produce a detectable antibody response [27, 31, 42].
ACKNOWLEDGEMENTS
This study was supported by Vicerrectoría de Investigación, Universidad de Costa Rica (project 741-A9-003). The authors thank our colleagues at Instituto Clodomiro Picado for their collaboration.
REFERENCES
[1] Meier, J.; Stocker, K.F. Biology and distribution of venomous snakes of medical importance and the composition of snake venoms. In: Handbook of Clinical Toxicology of Animal Venoms and Poisons; Meier, J., White, J., Eds.; CRC Press: Boca Raton, 1995; pp. 367-412.
[2] Fry, B.G.; Vidal, N.; Norman, J.A.; Vonk, F.J.; Scheib, H.; Ramjan, S.F.; Kuruppu, S.; Fung, K.; Hedges, S.B.; Richardson, M.K.; Hodgson, W.C.; Ignjatovic, V.; Summerhayes, R; Kochva, E. Early evolution of the venom system in lizards and snakes. Nature, 2006; 439, 584-588.
[3] Mackessy, S.P. The field of reptile toxinology. Snakes, lizards, and their venoms In: Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press: Boca Raton, 2009; pp. 3-23.
[4] Calvete, J.J.; Suárez, P.; Sanz, L. Snake venomics. Strategy and applications. J. Mass Spectrom., 2007, 42, 1405-1414.
[5] Serrano, S.M.T.; Fox, J.W. Exploring snake venom proteomes: multifaceted analyses for complex toxin mixtures. Proteomics, 2008, 8, 909-920.
[6] Chippaux, JP. Snake-bites: appraisal of the global situation. Bull. World Health Org., 1998, 76, 515-524.
[7] Gutiérrez, J.M.; Theakston, R.D.G.; Warrell, D.A. Confronting the neglected problem of snake bite envenoming: The need for a global partnership. PLoS Med, (On Line) 2006, 3(6), 727-731. http://plosmedicine.org/ [Accessed on: 06 June 2006].
[8] Gutiérrez, J.M.; Williams, D.; Fan, H.W.; Warrell, D.A. Snakebite envenoming from a global perspective: the need of an integrated approach. Toxicon, 2010, 56, 1223-1235.
[9] Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lallo, D.G.; de Silva, H.J. The global burden of snakebite: a literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med, [On Line] 2008, 5(11), 1591-1604. http://plosmedicine.org/ [Accessed on: 04 November 2008].
[10] Warrell, D.A. Envenomings and Their Treatments. In: Clinical
features of envenoming from snake bites; Bon, C., Goyffon, M., Eds.; Fondation Marcel Mérieux: Lyon, 1996; pp. 63-76.
[11] Ribeiro, L.A.; Puorto, G.; Jorge, M.T. Bites by the colubrid snake Philodryas olfersii: a clinical and epidemiological study of 43 cases. Toxicon, 1999, 37, 943-948.
[12] Hill, R.E.; Mackessy, S.P. Characterization of venom (Duvernoy’s secretion) from twelve species of colubrid snakes and partial sequence of four venom proteins. Toxicon, 2000, 38, 1663-1687.
[13] Peichoto, M.E.; Teibler, P.; Ruíz, R.; Leiva, L.; Acosta, O. Systemic pathological alterations caused by Philodryas
patagoniensis colubrid snake venom in rats. Toxicon, 2006, 48, 520-528.
[14] Weinstein, S.; Keyler, D. Local envenoming by the Western hognose snake (Heterodon nasicus): A case report and review of medically significant Heterodon bites. Toxicon, 2009, 54, 354-360.
[15] Chippaux, J.P.; Goyffon, M. Venoms, antivenoms and immunotherapy. Toxicon, 1998, 36, 823-846.
[16] World Health Organization. Rabies and Envenomigs. A Neglected
Public Health Geneva, 2007. [17] Theakston, R.; Warrell, D.; Griffiths, E. Report of a WHO
workshop on the standardization and control of antivenoms. Toxicon, 2003, 41, 541-557.
[18] Gutiérrez, J; León, G. Snake Antivenoms. Technological, clinical and public health issues. In Animal Toxins: State of the Art.
Perspectives in Health and Biotechnology; de Lima, M.E., Pimenta, A.M.C., Martin-Euclaire, M.F., Zingalli, R.B., Rochat, H., Eds.; Editora UFMG: Belo Horizonte, 2009; pp. 393-421.
[19] Lalloo, D.G.; Theakston, R.D.G. Snake antivenoms. J. Toxicol.
Clin. Toxicol., 2003, 41, 277-290. [20] World Health Organization. Guidelines for the Production, Control
and Regulation of Snake Antivenom Immunoglobulins. Geneva, 2010.
[21] Schunk, M.; Macallum, G. Applications and optimization of immunization procedures. ILAR J, 2005, 46, 241-257.
[22] Delves, P.J.; Roitt, I.M. The immune system. Advances in Immunology. N. Engl. J. Med., 2000, 343, 37-49.
[23] Juárez, P.; Sanz, L.; Calvete, J.J. Snake venomics: characterization of protein families in Sistrurus barbouri venom by cysteine mapping, N-terminal sequencing, and tandem mass spectrometry analysis. Proteomics, 2004, 4, 327-338.
[24] Bazaa, A.; Marrakchi, N.; El-Ayeb, M.; Sanz, L.; Calvete, J.J. Snake venomics: comparative analysis of the venoms proteomes of the Tunisian snakes Cerastes cerastes, Cerastes vipera and Macrovipera lebetina. Proteomics, 2005, 5, 4223-4235.
[25] Calvete, J.J.; Marcinkiewicz, C.; Sanz, L. Snake venomics of Bitis gabonica gabonica. Protein family composition, subunit organization of venom toxins, and characterization of dimeric disintegrins bitisgabonin-1 and bitisgabonin-2. J. Proteome Res., 2007, 6, 326-336.
[26] Angulo, Y.; Escolano, J.; Lomonte, B.; Gutiérrez, J.M.; Sanz, L.; Calvete, J.J. Snake venomics of Central American pitvipers: clues for rationalizing the distinct envenomation profiles of Atropoides
nummifer and Atropoides picadoi. J. Proteome Res., 2008, 7, 708-719.
[27] Lomonte, B.; Escolano, J.; Fernández, J.; Sanz, L.; Angulo, Y.; Gutiérrez, J.M.; Calvete, J.J. Snake venomics and antivenomics of the arboreal neotropical pitvipers Bothriechis lateralis and Bothriechis schelegelii. J. Proteome Res., 2008, 7, 2445-2457.
[28] Sanz, L.; Escolano, J.; Ferretti, M.; Biscoglio, M.J.; Rivera, E.; Crescenti, E.J.; Angulo, Y.; Lomonte, B.; Gutiérrez, J.M.; Calvete, J.J. Snake venomics of the South and Central American Bushmasters. Comparison of the toxin composition of Lachesis
muta gathered form proteomic versus transcriptomic analysis. J. Proteomics, 2008, 71, 46-60.
[29] Sanz, L.; Ayvazyan, N.; Calvete, J.J. Snake venomics of the Armenian vipers Macrovipera lebetina obusta and Vipera raddei. J. Proteomics, 2008, 71, 198-209.
[30] Alape-Girón, A.; Sanz, L.; Escolano, J.; Flores-Díaz, M.; Madrigal, M.; Sasa, M.; Calvete, J.J. Snake venomics of the lancead pitviper Bothrops asper: geographic, individual and ontogenic variations. J.
Proteome Res., 2008, 7, 3556-3571. [31] Gutiérrez, J.M.; Sanz, L.; Escolano, J.; Fernández, J.; Lomonte, B.;
Angulo, Y.; Rucavado, A.; Warrell, D.A.; Calvete, J.J. Snake venomics of the lesser Antillean pit vipers Bothrops caribbaeus and Bothrops lanceolatus: correlation with toxicological activities and immunoreactivity of a heterologous antivenom. J. Proteome
Res., 2008, 7, 4396-4408. [32] Tashima, A.K.; Sanz, L.; Camargo, A.C.; Serrano, S.M.; Calvete,
J.J. Snake venomics of the Brazilian pitvipers Bothrops cotiara and Bothrops fonsecai. Identification of taxonomy markers. J.
Proteomics, 2008, 71, 473-485. [33] Calvete, J.J.; Borges, A.; Segura, A.; Flores-Díaz, M.; Alape-Girón,
A.; Gutiérrez, J.M.; Diez, N.; De Sousa, L.; Kiriakos, D.; Sánchez, E.; Faks, J.G.; Escolano, J.; Sanz, L. Snake venomics and antivenomics of Bothrops colombiensis, a medically important pitviper of the Bothrops atrox-asper complex endemic to Venezuela: Contributing to its taxonomy and snakebite management. J. Proteomics, 2009, 72, 227-240.
[34] Nuñez, V.; Cid, P.; Sanz, L.; De La Torre, P.; Angulo, Y.; Lomonte, B.; Gutiérrez, J.M.; Calvete, J.J. Snake venomics and antivenomics of Bothrops atrox venoms from Colombia and the Amazon regions of Brazil, Perú and Ecuador suggest the

Immune Response Towards Snake Venoms Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 13
occurrence of geographic variation of venom phenotype by a trend towards paedomorphism. J. Proteomics, 2009, 73, 57-78.
[35] Valente, R.H.; Guimarães, P.R.; Junqueira, M.; Neves-Ferreira, A.G.; Soares, M.R.; Chapeaurouge, A.; Trugilho, M.R.; León, I.R.; Rocha, S.L.; Oliveira-Carvalho, A.L.; Wermelinger, L.S.; Dutra, D.L.; Leão, L.I.; Junqueira-de-Azevedo, I.L.; Ho, P.L.; Zingali, R.B.; Perales, J.; Domont, G.B. Bothrops insularis venomics: a proteomic analysis supported by transcriptomic-generated sequence data. J. Proteomics, 2009, 72, 241-255.
[36] Calvete, J.J.; Fasoli, E.; Sanz, L.; Boschetti, E.; Righetti, P.G. Exploring the venom proteome of the western diamondback rattlesnake, Crotalus atrox, via snake venomics and combinatorial peptide ligand library approaches. J. Proteome Res., 2009, 8, 3055-3067.
[37] Wagstaff, S.C.; Sanz, L.; Juárez, P.; Harrison, R.A.; Calvete, J.J. Combined snake venomics and venom gland transcriptomic analysis of the ocellated carpet viper, Echis ocellatus. J.
Proteomics, 2009, 71, 609-623. [38] Risch, M.; Georgieva, D.; von Bergen, M.; Jehmlich, N.; Genov,
N.; Arni, R.K.; Betzel, C. Snake venomics of the Siamese Russell´s viper (Daboia russelli siamensis). Relation to pharmacological activities. J. Proteomics, 2009, 72, 256-269.
[39] Olamendi-Portugal, T.; Batista, C.V.F.; Restano, R.R.; Pando, V.; Villa-Hernández, O.; Zavaleta-Matínez-Vargas, A.; Salas-Arruz, M.C.; Rodríguez de la Vega, R.C.; Becerri, B.; Possani, L.D. Proteomic analysis of the venom from the fish eating coral snake Micrurus surinamensis: Novel toxins, their functions and phylogeny. Proteomics, 2008, 8, 1919-1932.
[40] Li, S.; Wang, J.; Zhang, X.; Ren, Y.; Wang, N.; Zhao, K.; Chen, X.; Zhao, C.; Li, X.; Shao, J.; Yin, J.; West, M.B.; Xu, N.; Liu, S. Proteomic characterization of two snake venoms: Naja naja atra and Agkistrodon halys. Biochem. J., 2004, 384, 119-127.
[41] Ching, A.T.; Rocha, M.M.; Paes-Leme, A.F.; Pimenta, D.C.; de Fátima de Furtado, M.; Serrano, S.M.; Ho, P.L.; Junqueira-de-Azevedo, I.L. Some aspects of the venom proteome of the Colubridae snake Philodryas olfersii revealed from a Duvernoy´s (venom) gland transcriptome. FEBS Lett., 2006, 580, 4417-4422.
[42] Calvete, J.J.; Sanz, L.; Angulo, Y.; Lomonte, B.; Gutiérrez, J.M. Venoms, venomics and antivenomics. FEBS Lett., 2009, 583, 1736-1743.
[43] Tan, N.; Saifuddin, N. Isolation and characterization of a hemorrhagin from the venom of Ophiophagus hannah (king cobra). Toxicon, 1990, 28, 385-392.
[44] Ovadia, M. Isolation and characterization of a hemorrhagic factor from the venom of the snake Atractaspis engeddensis (Atractaspididae). Toxicon, 1987, 25, 621-630.
[45] Fox, J.W.; Serrano, S.M. Timeline of key events in snake venom metalloproteinase research. J. Proteomics, 2009, 72, 200-209.
[46] Fox, J.W.; Serrano, S.M.T. Snake venom metalloproteinases. In: Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press: Boca Raton, 2009; pp. 95-113.
[47] Gutiérrez, J.M.; Rucavado, A. Snake venom metalloproteinases: their role in the pathogenesis of local tissue damage. Biochimie, 2000, 82, 841-850.
[48] Gutiérrez, J.M.; Rucavado, A.; Escalante, T.; Díaz, C. Hemorrhage induced by snake venom metalloproteinases: biochemical and biophysical mechanisms involved in microvessel damage. Toxicon, 2005, 45, 997-1011.
[49] Gutiérrez, J.M.; Rucavado, A.; Escalante, T. Snake venom metalloproteinases. Biological roles and participation in the pathophysiology of envenomation. In: Handbook of Venoms and
Toxins of Reptiles; Mackessy, S.P. Ed.; CRC Press: Boca Raton, 2009, pp. 115-138.
[50] Franceschi, A.; Rucavado, A.; Mora, N.; Gutiérrez, J.M. Purification and characterization of BaH4, a hemorrhagic metalloproteinase from the venom of the snake Bothrops asper. Toxicon, 2000, 38, 63-77.
[51] Escalante, T.; Nuñez, J.; Moura da Silva, A.; Rucavado, A.; Theakston, R.D.; Gutiérrez, J.M. Pulmonary hemorrhage induced by jararhagin, a metalloproteinase from Bothrops jararaca snake venom. Toxicol. Appl. Pharmacol., 2003, 193, 17–28.
[52] Moura-da-Silva, A.M.; Serrano, S.M.T.; Fox, J.W.; Gutiérrez, J.M. Snake venom metalloproteinases. Structure, function and effects on snakebite pathology. In: Animal Toxins: State of the Art. Perspectives in Health and Biotechnology; de Lima, M.E.,
Pimenta, A.M.C., Martin-Euclaire, M.F., Zingalli, R.B., Rochat, H., Eds.; Editora UFMG: Belo Horizonte, 2009; pp. 525-546.
[53] Stocker, W.; Bode, W. Structural features of a superfamily of zinc-endopeptidases: the metzincins. Curr. Op. Struct. Biol., 1995, 5, 383-390.
[54] Swenson, S.; Markland, F.S. Snake venom fibrin(ogen)olytic enzymes. Toxicon, 2005, 45, 1021–1039.
[55] Teixeira, C.F.P.; Fernandes, C.M.; Zuliani, J.P.; Zamunér, S.R. Inflammatory effects of snake venom metalloproteinases. Mem.
Inst. Oswaldo Cruz, 2005, 100, 181-184. [56] Weissenberg, S.; Ovadia, M.; Kochva, E. Species specific
sensitivity towards the hemorrhagin of Ophiophagus hannah (Elapidae). Toxicon, 1987, 25, 475-481.
[57] Gutiérrez, J.M.; Sanz, L.; Flores-Díaz, M.; Figueroa, L.; Madrigal, M.; Herrera, M.; Villalta, M.; León, G.; Estrada, R.; Borges, A.; Alape-Girón, A.; Calvete, J.J. Impact of regional variation in Bothrops asper venom on the design of antivenoms: Integrating antivenomics and neutralization approaches. J. Proteome Res., 2009, 9, 564-577.
[58] Braud, S.; Bon, C.; Wisner, A. Snake venom proteins acting on hemostasis. Biochimie, 2000, 82, 851-859.
[59] Jin, Y.; Lee, W.H.; Zhang, Y. Molecular cloning of serine proteases from elapids snake venoms. Toxicon, 2007, 49, 1200-1207.
[60] Paes-Leme, A.F.; Prezoteo, B.C.; Yamashiro, E.T.; Bertholim, L.; Tashima, A.K.; Klitzke, C.F.; Camargo, A.C.; Serrano, S.M. Bothrops protease A, a unique highly glycosilated serine proteinase, is a potent, specific fibrinogenolytic agent. J. Thromb. Haemost., 2008, 6, 1363-1372.
[61] Markland, F.S. Snake venoms and the hemostatic system. Toxicon, 1998, 36, 1749-1800.
[62] Matsui, T.; Fujimura, Y.; Titani, K. Snake venom proteases afecting hemostasis and thrombosis. Biochim. Biophys. Acta, 2000, 1477, 146-156.
[63] Pirkle, H.; Markland, F.S.; Theodor, I. Thrombin-like enzymes of snake venom: actions on prothrombin. Thromb. Res., 1976, 8, 619-627.
[64] Komori, Y.; Nikai, T.; Sugihara, H. Biochemical and physiological studies on a kallikrein-like enzyme from the venom of Crotalus
viridis viridis (Prairie rattlesnake). Biochim. Biophys. Acta, 1988, 967, 92-102.
[65] Iwanaga, S.; Oshima, G.; Suzuki, T. Proteinases from the venom Agkistrodon halys blomhoffii. Meth. Enzymol., 1976, 45, 459-468.
[66] Phillips, D.J.; Swenson, S.D.; Markland, F.S. Thrombin-like snake venom serine proteinases. In: Handbook of Venoms and Toxins of
Reptiles; Mackessy, S.P. Ed.; CRC Press: Boca Raton, 2009, pp. 139-154.
[67] Kini, R.M. The intriguing world of prothrombin activators from snake venoms. Toxicon, 2005, 45, 1133-1145.
[68] Doley, R.; Zhou, X.; Kini, R.M. Snake venom phospholipase A2 enzymes. In: Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press: Boca Raton, 2009; pp. 173-205.
[69] Arni, R.K.; Ward, R.J. Phospholipase A2: A structural review. Toxicon, 1996, 34, 827-841.
[70] Magro, A.J.; Murakami, M.T.; Marcussi, S.; Soares, A.M.; Arni, R.K.; Fontes, M.R. Crystal structure of an acidic platelet aggregation inhibitor and hypotensive phospholipase A2 in the monomeric and dimeric states: insights into its oligomeric state. Biochem. Biophys. Res. Comm., 2004, 323, 24-31.
[71] Lomonte, B.; Angulo, Y.; Sasa, M.; Gutiérrez, J.M. The phospholipase A2 homologues of snake venoms: Biological activities and their posible adaptive role. Prot. Pept. Letters, 2009, 16, 860-876.
[72] Fraenkel-Conrat, H.; Singer, B. Fractionation and Composition of Crotoxin. Arch. Biochem. Biophys., 1956, 60, 64-73.
[73] Tsai, I.H.; Lu, P.J.; Wang, Y.M.; Ho, C.L.; Liaw, L.L. Molecular cloning and characterization of a neurotoxic phospholipase A2 from the venom of Taiwan habu (Trimeresurus mucrosquamatus). Biochem. J., 1995, 311, 895-900.
[74] Tsai, I.H.; Lu, P.J.; Su, J.C. Two types of Russell´s viper revealed by variation in phospholipases A2 from venom of the subspecies. Toxicon, 1996, 34, 99-109.
[75] Bon, C. Multicomponent neurotoxic phospholipases A2. In: Venom
Phospholipase A2 Enzymes. Structure, Function and Mechanism; Kini, R.M. Ed.; Wiley: Chichester, 1997, pp. 269-85.

14 Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 León et al.
[76] Kini, R.M. Excitement ahead: structure, function and mechanism of snake venom phospholipase A2 enzymes. Toxicon, 2003, 42, 827-840.
[77] Kuruppu S, Smith AI, Isbister GK, Hodgson, W.C. Neurotoxins from Australo-Papuan elapids: a biochemical and pharmacological perspective. Crit. Rev. Toxicol. 2008, 38, 73-86.
[78] Tsai, I.H.; Hseu, M.J. Snake venom neurotoxic phospholipases A2. In Animal Toxins: State of the Art. Perspectives in Health and Biotechnology; de Lima, M.E., Pimenta, A.M.C., Martin-Euclaire, M.F., Zingalli, R.B., Rochat, H., Eds.; Editora UFMG: Belo Horizonte, 2009; pp. 481-494.
[79] Du, K.Y.; Clement, K.J. Snake venom L-amino acid oxidases. Toxicon, 2002, 40, 659-665.
[80] Ponnudurai, G.; Chung, M.; Tan, N.H. Purificaction and properties of the L-amino acid oxidase from Malayan pit viper (Calloselasma
rhodostoma) venom. Arch. Biochem. Biophys., 1994, 113, 373-378. [81] Wei, J.F.; Yang, H.W.; Wei, X.L.; Qiao, L.Y.; Wang, W.Y.; He,
S.H. Purification, characterization and biological activities of the L-amino acid oxidase from Bungarus fasciatus snake venom. Toxicon, 2009, 54, 262-271.
[82] Zhong, S.R.; Jin, Y.; Wu, J.B.; Jia, Y.H.; Xu, G.L.; Wang, G.C.; Xiong, Y.L.; Lu, Q.M. Purification and characterization of a new L-amino acid oxidase from Daboia russellii siamensis venom. Toxicon, 2009, 54, 763-771.
[83] Suhr, S.M.; Kim, D.S. Identification of the snake venom substance that induces apoptosis. Biochem. Biophys. Res. Comm., 1996, 224, 134-139.
[84] Stiles, B.G.; Sexton, F.W.; Weinstein, S.A. Antibacterial effects of different snake venoms: purification and characterization of antibacterial proteins from Pseudechis australis (Australian King Brown or Mulga snake) venom. Toxicon, 1991, 29, 1129-1141.
[85] Ali, S.A.; Stoeva, S.; Abbasi, A.; Alam, J.M.; Kayed, R.; Faigle, M.; Neumeister, B.; Voelter, W. Isolation, structural and functional characterization of an apoptosis-inducing L-amino acid oxidase from Leaf Nosed Viper (Eristocophis macmahoni) snake venom. Arch. Biochem. Biophys., 2000, 384, 216-226.
[86] Calvete, J.J.; Moreno-Murciano, M.P.; Sanz, L.; Jürgens, M.; Schrader, M.; Raida, M.; Benjamín, D.C.; Fox, J.W. The disulfide bond pattern of catrocollastatin C a disintegrin-like/cysteine-rich protein isolated from Crotalus atrox venom. Protein Sci., 2000, 9, 1365-1373.
[87] Calvete, J.J.; Marcinkiewicz, C.; Monleón, D.; Esteve, V.; Celda, B.; Juárez, P.; Sanz, L. Snake venom disintegrins: evolution of structure and function. Toxicon, 2005, 45, 1063-1074.
[88] Calvete, J.J.; Moreno-Murciano, M.P.; Theakston, R.D.G.; Kisile, D.G.; Marcinkiewicz, C. Snake venom disintegrins: novel dimeric disintegrins and structural diversification by disulphide bond engineering. Biochem. J., 2003, 372, 725-734.
[89] Blobel, C.P.; White, J.M. Structure, function and evolutionary relationship of proteins containing a disintegrin domain. Curr. Opin. Cell. Biol., 1992, 4, 760-765.
[90] Wermelinger, L.; Geraldo, R.; Frattani, F.; Rodrigues, C.R.; Juliano, M.A.; Castro, H.C.; Zingali, R.B. Integrin inhibitors from snake venoms: Exploring the relationship between the structure and activity of RGD-peptides. Arch. Biochem. Biophys., 2009, 482, 25-32.
[91] Moiseeva, N.; Bau, R.; Swenson, S.D.; Markland, F.S.; Choe, J.Y.; Liu, Z.J.; Allaire, M. Structure of acostatin, a dimeric disintegrins from Southern cooperhead (Agkistrodon contortrix contortrix) at 1.7 A resolution. Acta Crystallogr. D. Biol. Crystallogr., 2008, 64, 466-470.
[92] Zha, H-G.; Lee, W-H.; Zhang, Y. Cloning of cDNAs encoding C-type lectins from Elapidae snakes Bungarus fasciatus and Bungarus multicinctus. Toxicon, 2001, 39, 1887-1892.
[93] Zingali, R.; Correa-Netto, C.; Guimaraes-Gomes, V. Snake venom C-type lectins and C-type lectins-like proteins. In Animal Toxins: State of the Art. Perspectives in Health and Biotechnology; de Lima, M.E., Pimenta, A.M.C., Martin-Euclaire, M.F., Zingalli, R.B., Rochat, H., Eds.; Editora UFMG: Belo Horizonte, 2009; pp. 589-606.
[94] Lomonte, B.; Rojas, G.; Gutiérrez, J.M.; Ramírez, G. Isolation of a galactose-binding lectin from the venom of the snake Bothrops godmani (Godmann´s pit viper). Toxicon, 1990, 28, 75-81.
[95] Nikai, T.; Kato, S.; Komori, Y.; Sugihara, H. Amino acid sequence and biological properties of the lectin from the venom of
Trimeresurus okinavensis (Himehabu). Toxicon, 2000, 38: 707-711.
[96] Guimaraes-Gomes, V.; Oliveira-Carvalho, A.; Junqueira-de-Azevedo, I.; Dutra, D.L.; Pujol-Luz, M.; Castro, H.C.; Ho, P.L.; Zingali, R.B. Cloning, characterization and structural analysis of a C-type lectin from Bothrops insularis (BiL) venom. Arch. Biochem. Biophys. 2004, 432, 1-11.
[97] Lu, Q.; Navdaev, A.; Clemetson, J.; Clemetson, K. Snake venom C-type lectins interacting with platelet receptors.Structure-function relationships and effects on haemostasis. Toxicon, 2005, 45,1089-1098.
[98] Kilpatrick, D.C. Animal lectins: a historical introduction and overview. Biochim. Biophys. Acta, 2002, 1572,187-197.
[99] Álvarez, P.; Girao, M.; Rangel, C.; Carla, H. Structure–function inferences based on molecular modeling, sequence-based methods and biological data analysis of snake venom lectins. Toxicon, 2006, 48, 690-701.
[100] Hedge, R.P.; Rajagopalan, N.; Doley, R.; Kini, R.M. Snake venom three-finger toxins. In: Handbook of Venoms and Toxins of
Reptiles; Mackessy, S.P. Ed.; CRC Press: Boca Raton, 2009; pp. 287-301.
[101] Bdolah, A. Sarafotoxins, the snake venom homologs of endothelins. In: Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P. Ed.; CRC Press: Boca Raton, 2009, 303-315.
[102] Heyborne, W.H.; Mackessy, S.P. Cysteine-rich secretory proteins in reptile venoms. In: Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P. Ed.; CRC Press: Boca Raton, 2009, 325-336.
[103] Harvey, A.L. Fasciculins. Toxins from mamba venoms that inhibit acetylcholinesterase. In: Handbook of Venoms and Toxins of
Reptiles; Mackessy, S.P. Ed.; CRC Press: Boca Raton, 2009, 317-324.
[104] Angulo, Y.; Estrada, R.; Gutiérrez, J.M. Clinical and laboratory alterations in horses during immunization with snake venoms for the production of polyvalent (Crotalinae) antivenom. Toxicon, 1997, 35, 81-90.
[105] Moroz, C.; Goldblum, N.; Vries, A. Preparation of Vipera palestinae antineurotoxin using caroximethyl-cellulose-bound neurotoxin as antigen. Nature, 1963, 16, 697-698.
[106] Kocholaty, W. Detoxification of Crotalus atrox venom by photooxidation in the presence of methylene blue. Toxicon, 1966, 3, 175-186.
[107] Kocholaty, W.; Ashley, B. Detoxification of Russell`s viper (Vipera russelli) and water moccasin (Agkistrodon piscivorus) venoms by photooxidation. Toxicon, 1966, 3, 187-194.
[108] Higashi, H.G.; Guidolin, R.; Nishikawa, A.K.; Yamaguchi, I.K.; Lima, M.L.S.R.; Morais, J.F.; Dias Da Silva, W. Bothropic venoms pre-treated with inhibitors for active enzymatic sites of proteases and chelating substances preserve their immunogenicity. Mem. Inst. Butantan, 1989, 51, 107-115.
[109] León, G.; Estrada, R.; Chaves, F.; Rojas, G.; Ovadia, M.; Gutiérrez, J.M. Inhibition by CaNa2EDTA of local tissue damage induced by Bothrops asper (terciopelo) venom: Application in horse immunization for antivenom production. Toxicon, 1998, 36, 321-331.
[110] Grasset, E. Anavenoms and their use in the preparation of antivenomous sera. Trans. Royal. Soc. Trop. Med., 1945, 38, 463-488.
[111] Costa, L. Comparative study of equine immune response towards venom from Crotalus durissus terrificus, in natura, treated with formaldehyde and submited to heat treatment. Vac. Soros, 1985, 1, 24-29.
[112] Heneine, L; Cardoso, V.; Daniel, J.; Heneine, I.F. Detoxification of the T z fraction from a scorpion (Tityus serrulatus, Lutz and Mello)
venom by iodination and some immunogenic properties of derivates. Toxicon, 1986, 24, 501-505.
[113] Daniel, J.; Heneine, L.; Tavares, C.; Nascimento, M.C.; Heneine, I.F. Generation of protective immune sera by Crotalus durissus
terrificus detoxified by controlled iodination. Braz. J. Med. Biol. Res., 1987, 20, 713-720.
[114] Guidolin, R.; Dias da Silva, W.; Higashi, H.; Caricat, C.P.; Lima, M.L.S.R.; Morais, J.F.; Pinto, J.R.; Marcelino, J.R. Hyperimmunization of horses with bothropic and crotalic venoms treated with glutaraldehyde. Mem. Inst. Butantan, 1989, 51, 85-90.
[115] Sawai, Y.; Kawamura, Y. Study on the toxoids against the venoms of certain Asian snakes. Toxicon, 1969, 7, 19-24.

Immune Response Towards Snake Venoms Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 15
[116] Flowers, H. The effects of X-irradiation on the biological activity of cottonmouth moccasin (Ancistrodon piscivorus) venom. Toxicon, 1963, 1, 131-136.
[117] Tejasen, P.; Ottolenghi, A. The effect of ultra-violet light on the toxicity and the enzymatic and antigenic activities of snake venom. Toxicon, 1970, 8, 225-230.
[118] Rangel-Santos, A.C.; Mota, I. Effect of heating on the toxic, immunogenic and immunosuppressive activities of Crotalus durissus terrificus venom. Toxicon, 2000, 38, 1451-1457.
[119] Hati, R.; Mandal, M.; Hati, K. Active immunization of rabbit with gamma irradiated russell`s viper venom toxoid. Toxicon, 1990, 28, 895-902.
[120] Cardi, B.; Andrade, H.; Rogero, J.; Nascimento, N. Differential biodistribution of native and 2kGy 60Co irradiated crotoxin in tissues of CBA/J mice. Nat. Toxins, 1998, 6, 19-25.
[121] Clissa, P.B., do Nascimento N, Rogero JR. Toxicity and immunogenicity of Crotalus durissus terrificus venom treated with different doses of gamma rays. Toxicon, 1999, 37, 1131-1141.
[122] Nascimento, N.; Seebart, C.; Francis, B.; Rogero, J.R.; Kaiser, I.I. Influence of ionizing radiation on crotoxin: biochemical and immunological aspects. Toxicon, 1996, 34, 123-131.
[123] Sousa, J.R.F.; Monteiro, R.Q.; Castro, H.C.; Zingali, R.B. Proteolytic action of Bothrops jararaca venom upon its own constituents. Toxicon, 2001, 39, 787-792.
[124] Medeiros, C.R.; Barbaro, K.C.; Lira, M.S.; França, F.O.; Zaher, V.L.; Kokron, C.M.; Kalil, J.; Castro, F.F. Predictors of Bothrops jararaca venom allergy in snake handlers and snake venom handlers. Toxicon, 2008, 51, 672-680.
[125] Theakston, R.D.G.; Pugh, R.N.H.; Reid, H.A. Enzyme-lynked immunosorbent assay of venom antibodies in human victims of snakebite. J. Trop. Med. Hyg., 1981, 84, 109-122.
[126] Ameno, S.; Ameno, K.; Fuke, Ch.; Kiryu, T.; Ljiri, I. IgG subclass distributions of anti-horse serum antibodies and natural venom-antibodies produced in reponse to antivenom injection of snake bite in humans. Toxicon, 1990, 28, 347-350.
[127] Domingos, M.O.; Cardoso, J.L.; Moura-da-Silva, A.M.; Mota, I. The humoral immune responses of patients bitten by the snake Bothrops jararaca (Jararaca). Toxicon, 1990, 28, 723-726.
[128] Isbister, G.K.; Halkidis, L.; O´Leary, M.A.; Whitaker, R.; Cullen, P.; Mulcahy, R.; Bonnin, R.; Brown, S.G. Human anti-snake venom IgG antibodies in a previously bitten snake-handler, but no protection against local envenoming. Toxicon, 2009, 55, 646-649.
[129] Chotwiwatthanakun, Ch.; Ronachai, P.; Akesowan, S.; Sriprapat, S.; Ratanabanangkoon, K. Production of potent polyvalent antivenom against three elapid venoms using a low dose, low volume, multi-site immunization protocol. Toxicon, 2001, 39, 1487-1494.
[130] Sriprapat, S.; Aeksowan, S.; Sapsutthipas, S.; Chotwiwatthanakun, C.; Suttijitpaisal, P.; Pratanaphon, R.; Khow, O.; Sitprija, V.; Ratanabanangkoon, K. The impact of a low dose, low volume, multi-site immunization on the production of therapeutic antivenoms in Thailand. Toxicon, 2003, 41, 57-64.
[131] Harrison, R.A.; Hasson, S.S.; Harmsen, M.; Laing, G.D.; Conrath, K.; Theakston, R.D. Neutralization of venom-induced haemorrhage by IgG from camels and llamas immunized with viper venom and also by endogenous, non-IgG components in camelid sera. Toxicon, 2006, 47, 364-368.
[132] El-Kady, E.M.; Ibrahim, N.M.; Wahby, A.F. Assessment of the anti-Naja haje antibodies elicited in a low dose multi-site immunization protocol. Toxicon, 2009, 54, 450-459.
[133] Maya, C.; Vasantha, M.; Krishnan, L.K. Development of viper-venom antibodies in chicken egg yolk and assay of their antigen binding capacity. Toxicon, 2002, 40, 857-861.
[134] Paul, K.; Manjula, J.; Deepa, E.P.; Selvanayagam, Z.E.; Ganesh, K.A.; Subba Rao, P.V. Anti-Echis carinatus venom antibodies from chicken egg yolk: Isolation, purification and neutralization efficacy. Toxicon, 2007, 50, 893-900.
[135] New, R.; Theakston, R.; Zumbuehl, O.; Iddon, D.; Friend, J. Liposomal immunization against snake venoms. Toxicon, 1985, 23, 215-219.
[136] Laing, G.; Theakston, R. Immunization against Echis ocellatus
(carpet viper) venom using liposomes incorporating immunostimulants: role of lipopolysacharide in conferring protection in a mouse model. Toxicon, 1993, 31, 615-626.
[137] Ismail, M.; Abd-Elsalam, M.A.; Al-Ahaidib, M.S. Pharmacokinetics of 125I-labelled Walterinnesia aegyptia venom
and its specific antivenins: flash absorption and distribution of the venom and its toxin versus slow absorption and distribution of IgG, F(ab´)2 and F(ab) of the antivenin. Toxicon, 1998, 36, 93-114.
[138] Riviére, G.; Choumet, V.; Saliou, B.; Debray, M.; Bon, C. Absorption and elimination of viper venom after antivenom administration. J. Pharmacol. Exp. Ther., 1998, 285, 490-495.
[139] Lofthouse, Sh. Immunological aspects of controlled antigen delivery. Adv. Drug Deliv. Rev., 2002, 54, 863-870.
[140] Teixeira, C.; Cury, Y.; Moreira, V.; Picolob, G.; Chaves, F. Inflammation induced by Bothrops asper venom. Toxicon, 2009, 54, 988-997.
[141] Teixeira, C.F.P.; Landucci, E.C.T.; Antunes, E.; Chacur, M.; Cury, Y. Inflammatory effects of snake venom myotoxic phospholipases A2. Toxicon, 2003, 42, 947-962.
[142] Farsky, S.H.P.; Antunes, E.; Mello, S.B. Pro and antiinflammatory properties of toxins from animal venoms. Curr. Durg Targets Imflamm. Allergy, 2005, 4, 401-411.
[143] Costa, E.P.; Clissa, P.B.; Teixeira, C.F.P.; Moura da Silva, A.M. Importance of metalloproteinases and macrophages in viper snake envenomation-induced local inflammation. Inflammation, 2002, 26, 13-17.
[144] Farsky, S.H.P.; Gonçalves, L.R.C.; Gutiérrez, J.M.; Correa, A.P.; Rucavado, A.; Gasque, P. Tambourgi, D.V. Bothrops asper snake venom and its metalloproteinase BaP-1 activate the complement system. Role in leucocyte recruitment. Med. Inflammation, 2000, 9, 213-221.
[145] Rodrigues, F.G.; Petretski, J.H.; Kanashiro, M.M.; Lemos, L.; da Silva, W.D.; Kipnis, T.L. The complement system is involved in acute inflammation but not in the hemorrhage produced by a Bothrops atrox snake venom low molecular mass proteinase. Mol. Immunol., 2004, 40, 1149-1156.
[146] Domont, G.B.; Perales, J.; Moussatche, H. Natural anti-snake venom proteins. Toxicon, 1991, 29, 1183-1194.
[147] León, G.; Stiles, B.; Alape, A.; Rojas, G.; Gutiérrez, J.M. Comparative study on the ability of IgG and F(ab´)2 antivenoms to neutralize lethal and myotoxic effects induced by Micrurus nigrocinctus (coral snake) venom. Am. J. Trop. Med. Hyg., 1999, 62, 266-272.
[148] León, G.; Monge, M.; Rojas, E.; Lomonte, B.; Gutiérrez, J.M. Comparison between IgG and F(ab´)2 polyvalent antivenoms: neutralization of systemic effects induced by Bothrops asper venom in mice, extravasation to muscle tissue, and potential for induction of adverse reactions. Toxicon, 2001, 39, 793-801.
[149] Battellino, C.; Piazza, R.; Moura-da-Silva, A.M.; Cury, Y.; Farsky, S.H. Assesment of efficacy of bothropic antivenom therapy on microcirculatory effects induced by Bothrops jararaca snake venom. Toxicon, 2003, 41, 583-593.
[150] Escalante, T.; Rucavado, A.; Pinto, A.F.; Terra, R.M.; Gutiérrez, J.M.; Fox, J.W. Wound exudate as a proteomic window to reveal different mechanisms of tissue damage by snake venom toxins. J. Prot. Res., 2009, 8, 5120-5131.
[151] Zamunér, S.R.; Zuliani, J.P.; Fernandes, C.M.; Gutiérrez, J.M.; de-Fátima-Pereira-Teixeira, C. Inflamation induced by Bothrops asper venom: release of proinflamatory cytokines and eicosanoids, and role of adhesion molecules in leukocyte infiltration. Toxicon, 2005, 46, 806-813.
[152] Gutiérrez, J.M.; Chaves, F.; Cerdas, L. Inflammatory infiltrate in skeletal muscle injected with Bothrops asper venom. Rev. Biol. Trop., 1986, 34, 209-219.
[153] Lomonte, B.; Tarkowski, A.; Hanson, L.A. Host response to Bothrops asper snake venom. Inflammation, 1993, 17(2), 93-105.
[154] Teixeira, C.F.; Zamunér, S.R.; Zuliani, J.P.; Fernandes, C.M.; Cruz-Hofling, M.A.; Fernandes, I.; Chaves, F.; Gutiérrez, J.M. Neutrophils do not contribute to local tissue damage, but play a key role in skeletal muscle regeneration, in mice injected with Bothrops
asper snake venom. Muscle Nerve, 2003, 28(4), 449-459. [155] Zamunér, S.R.; Gutiérrez, J.M.; Muscará, M.N.; Teixeira, S.A.;
Teixeira, C.F. Bothrops asper and Bothrops jararaca snake venoms trigger microbicidal functions of peritoneal leukocytes in
vivo. Toxicon, 2001, 39(10), 1505-1513. [156] Dudziak, D.; Kamphorst, A.O.; Heidkamp, G.F.; Buchholz, V.R.;
Trumpfheller, C.; Yamazaki, S.; Cheong, C.; Liu, K.; Lee, H.W.; Park, C.G.; Steinman, R.M.; Nussenzweig, M.C. Differential antigen processing by dendritic cell subsets in vivo. Science, 2007, 315(5808), 107-111.

16 Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 León et al.
[157] Moura-da-Silva, A.M.; Laing, G.D.; Paine, M.J.; Dennison, J.M.; Politi, V.; Crampton, J.M.; Theakston, R.D. Processing of pro-tumor necrosis factor- by venom metalloproteinases: a hypothesis explaining local tissue damage following snake bite. Eur. J. Immunol., 1996, 26(9), 2000-2005.
[158] Gutiérrez, J.M.; Rucavado, A.; Chaves, F.; Díaz, C.; Escalante, T. Experimental pathology of local tissue damage induced by Bothrops asper snake venom. Toxicon, 2009, 54(7), 958-975.
[159] Trevett, A.J.; Lallo, D.G.; Nwokolo, N.C.; Theakston, D.G.; Naraqi, S.; Warrell, D.A. Venom detection kits in the management of snakebite in central province, Papua New Guinea. Toxicon, 1995, 33(5), 703-705.
[160] Selvanayagam, Z.E.; Gnanavendhan, S.G.; Ganesh, K.A.; Rajagopal, D.; Rao, P.V. ELISA for the detection of venoms from four medically important snakes of India. Toxicon, 1999, 37(5), 757-770.
[161] Van Dong, L.; Quyen le, K.; Eng, K.H.; Gopalakrishnakone, P. Immunogenicity of venoms from four common snakes in the South of Vietnam and development of ELISA kit for venom detection. J.
Immunol. Meth., 2003, 282(1-2), 13-31. [162] Moreno, E.; Gutiérrez, J.M. Body distribution of Bothrops asper
(Terciopelo) snake venom myotoxin and its relationship to pathological changes. Toxicon, 1988, 26(4), 403-409.
[163] Metlay, J.P.; Puré, E.; Steinman, R.M. Control of the immune response at the level of antigen-presenting cells: A comparison of the function of dendritic cells and B lymphocytes. Adv. Immunol., 1989, 47, 45-116.
[164] Rodríguez-Pinto, D. B cells as antigen presenting cells. Cell. Immunol., 2005, 238(2), 67-75.
[165] Thai, R.; Moine, G.; Desmadril, M.; Servent, D.; Tarride, J.L.; Ménez, A.; Léonetti, M. Antigen stability controls antigen presentation. J. Biol. Chem., 2004, 279(48), 50257-50266.
[166] Cardoso, D.F.; Mota, I. Effect of Crotalus venom on the humoral and cellular immune response. Toxicon, 1997, 35(4), 607-612.
[167] Cruz, A.H.; Mendonça, R.Z.; Petricevich, V.L. Crotalus durissus
terrificus venom interferes with morphological, functional, and biochemical changes in murine macrophage. Mediat. Inflamm., 2005, 2005(6), 349–359.
[168] Zuliani, J.P.; Gutiérrez, J.M.; Cassais-e-Silva, L.L.; Coccuzzo-Sampaio, S.; Lomonte, B.; Pereira-Teixeira, Cde.F. Activation of cellular functions in macrophages by venom secretory Asp-49 and Lys-49 phospholipases A2. Toxicon, 2005, 46(5), 523-532.
[169] Sampaio, S.C.; Brigatte, P.; Sousa-e-Silva, M.C.; dos-Santos, E.C.; Rangel-Santos, A.C.; Curi, R.; Cury, Y. Contribution of crotoxin for the inhibitory effect of Crotalus durissus terrificus snake venom on macrophage function. Toxicon, 2003, 41(7), 899-907.
[170] Rangel-Santos, A.; Lima, C.; Lopes-Ferreira, M.; Cardoso, D.F. Immunosuppresive role of principal toxin (crotoxin) of Crotalus durissus terrificus venom. Toxicon, 2004, 44(6), 609-616.
[171] Sampaio, S.C.; Rangel-Santos, A.C.; Peres, C.M.; Curi, R.; Cury, Y. Inhibitory effect of phospholipase A2 isolated from Crotalus
durissus terrificus venom on macrophage function. Toxicon, 2005, 45(5), 671-676.
[172] Skok, M.V.; Kalashnik, E.N.; Koval, L.N.; Tsetlin, V.I.; Utkin, Y.N.; Changeux, J.P.; Grailhe, R.. Functional nicotinic acetylcholine receptors are expressed in B lymphocyte-derived cell lines. Mol. Pharmacol., 2003, 64(4), 885-889.
[173] Skok, M.; Grailhe, R.; Changeux, J.P. Nicotinic receptors regulate B lymphocyte activation and immune response. Eur. J.
Pharmacol., 2005, 517(3), 246-251. [174] Estrada, R.; Robles, A.; Alvarado, J.; Rojas, E.; Gonzalez, N.;
Segura, E.; Gutiérrez, J.M. Development of antibody response and clinical and hematological alterations in horses immunized with snake venoms for the production of antivenom in Costa Rica. Mem. Inst. Butantan, 1991, 53(2), 181-190.
[175] Waghmare, A.; Deopurkar, R.L.; Salvi, N.; Khadilkar, M.; Kalolikar, M.; Gade, S.K. Comparisson of Montanide adjuvants, IMS 3012 (nanoparticle), ISA 206 and ISA 35 (emulsion based) along with incomplete Freund’s adjuvant for hyperimmunization of equines used for production of polyvalent snake antivenom. Vaccine, 2009, 27(7), 1067-1072.
[176] Heyman, B. Feedback regulation by IgG antibodies. Immunol. Lett., 2003, 88(2), 157-161.
[177] Brady, L.J. Antibody-mediated immunomodulation: a strategy to improve host responses against microbial antigens. Infect. Immun., 2005, 73(2), 671-678.
[178] Minton, S.A.Jr. Paraspecific protection by elapid and sea snake antivenins. Toxicon, 1967, 5(1), 47-55.
[179] Minton, S.A. Neutralization of old world vipers venoms by American pit viper antivenin. Toxicon, 1976, 14(2), 146-148.
[180] Chippaux, J.P.; Williams, V.; White, J. Snake venom variability: methods of study, results and interpretation. Toxicon, 1991, 29(11), 1279–1303.
[181] Ferreira, M.; Moura-Da-Silva, A.M.; Mota, I. Neutralization of different activities of venoms from nine species of Bothrops snakes by Bothrops jararaca antivenom. Toxicon, 1992, 30(12), 1591-1602.
[182] Dias da Silva, W.; Guidolin, R.; Raw, I.; Higashi, H.G.; Caricatti, C.P.; Morais J.F.; Lima, M.L.; Yamaguchi, I.K.; Nishikawa, A.K., Stephano, M.N.; Marcelino, J.R.; Pinto, J.R.; Santos, M.J. Cross-reactivity of horse monovalent antivenoms to venom of ten Bothrops species. Mem Inst Butantan, 1989, 51, 153-168.
[183] Anderson, S.; Gutiérrez, J.M.; Ownby, C.L. Comparison of the immunogenicity and antigenic composition of ten Central American snake venoms. Toxicon, 1993, 31(8), 1051-1059.
[184] Arce, V.; Rojas, E.; Ownby, C.L.; Rojas, G.; Gutiérrez, J.M. Preclinical assessment of the ability of polyvalent (Crotalinae) and anticoral (Elapidae) antivenoms produced in Costa Rica to neutralize the venoms of North American snakes. Toxicon, 2003, 41(7), 851-860.
[185] Ramos-Cerrillo, B.; de Roodt, A.; Chippaux, J.P.; Olguín, L.; Casasola, A.; Guzmán, G.; Paniagua-Solís, J.; Alagón, A.; Stock, R.P. Characterization of a new polyvalent antivenom (Antivipmyn® Africa) against African vipers and elapids. Toxicon, 2008, 52(8), 881-888.
[186] Casasola, A.; Ramos-Cerrillo, B.; de Roodt, A.; Carbajal Saucedo, A.; Chippaux, J.P.; Alagón, A.; Stock, R.P. Paraspecific neutralization of the venom of African species of cobra by analysis of equine antiserum against Naja melanoleuca: A comparative study. Toxicon, 2009, 53(6), 602-608.
[187] Segura, A.; Villalta, M.; Herrera, M.; León, G.; Harrison, R.; Durfa, N.; Nasidi, A.; Calvete, J.J.; Theakston, R.D.; Warrell, D.A.; Gutiérrez, J.M. Preclinical assesment of the efficacy of a new antivenom (EchiTab-Plus-ICP®) for the treatment of viper envenoming in sub-Saharan Africa. Toxicon, 2010, 55(2-3), 369-374.
[188] Ownby, C.; Colberg, T. Comparison of the immunogenicity and antigenic composition of several venoms of snakes in the family Crotalidae. Toxicon, 1990, 28, 189-199.
[189] Kornalík, F.; Táborská, E. Cross reactivity of mono and polyvalent antivenoms with viperidae and crotalidae snake. Toxicon, 1989, 27(10), 1135-1142.
[190] Nair, C.; Nair, B.; Elliot, W.B. Immunological comparison of Phospholipases A2 present in rattlesnake (genus Crotalus) venoms. Toxicon, 1980, 18(5-6), 675-680.
[191] Bolaños, R.; Cerdas, L.; Taylor, R. The production and characteristics of a coral snake (Micururs mipartitus hertwigi)
antivenin. Toxicon, 1975, 13(2), 139-142. [192] Chinonavanig, L.; Billings, P.B.; Matangkasombut, P.;
Ratanabanangkoon, K. Antigenic relationships and relative immunogenicities of venom proteins from six poisonous snakes of Thaliand. Toxicon, 1988, 26(9), 883-890.
[193] Gutiérrez, J.M.; Lomonte, B.; León, G.; Alape-Girón, A.; Flores-Díaz, M.; Sanz, L.; Angulo, Y.; Calvete, J.J. Snake venomics and antivenomics: Proteomic Tools in the design and control of antivenoms for the treatment of snakebite envenoming. J. Prot., 2009, 72(2), 165-182.
[194] White, J. Envenomation. Prevention and treatment in Australia. In: Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press: Boca Raton, 2009; pp. 423-451.
[195] Stills, H.F.Jr. Adjuvants and antibody production: dispelling the myths associated with Freud’s complete and other adjuvants. ILAR J., 2005, 46(3), 280-293.
[196] Jennings, V.M. Review of selected adjuvants used in antibody production. ILAR J., 1995, 37(3), 119-125.
[197] Morais, V.; Massaldi, H. Economic evaluation of snake antivenom production in the public system. J. Venom. Anim. Toxins incl. Trop.
Dis., 2006, 12(3), 497-511. [198] Gregoriadis, G. Immunological adjuvants: a role for liposomes.
Immunol. Today, 1990, 11(3), 89- 97. [199] Bowersock, T.L.; Martin, S. Vaccine delivery to animals. Adv.
Drug. Deliv. Rev., 1999, 38(2), 167-194.

Immune Response Towards Snake Venoms Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 17
[200] Sadahiro, S.; Kondo, L.; Yamamoto, A. Effects of adjuvants on the immunogenicity of Habu-venom toxoid in the monkey and the guinea pig. Jpn. J. Med. Sci. Biol., 1981, 34(3), 169-173.
[201] Freitas, T.V.; Fortes-Dias, C.L.; Diniz, C.R.; Velarde, D.T.; Freitas, C.F. Immunization of horses with Crotalus durissus terrificus
(south American rattlesnake) venom. A comparison of four different procedures. Braz. J. Med. Biol. Res., 1991, 24(3), 281-290.
[202] Sunthornandh, P.; Ratanabanangkoon, K. A comparative study of three vehicles on antibody responses against elapid snake neurotoxin immunogens. Toxicon, 1994, 32(5), 561-571.
[203] Lindblad, E.B. Aluminium adjuvants-in retrospect and prospect. Vaccine, 2004, 22(27-28), 3658-3668.
[204] Pratanaphon, R.; Akesowan, S.; Khow, O.; Sriprapat, S.; Ratanabanangkoon, K. Production of highly potent horse antivenom against the Thai cobra (Naja kaouthia). Vaccine, 1997, 15(14), 1523-1528.
[205] Theakston, R.D.G.; Zumbuehl, O.; New, R.R.C. Use of liposomes for protective immunization in sheep against Equis carinatus snake venom. Toxicon, 1985, 23(6), 921-925.
[206] Carvalho, V.T.; Gomes, R.T.; Viotti, A.P.; Freitas, T.V. Immunization with liposome-encapsulated Bothrops jararaca venom. Toxicon, 2000, 38(6), 881-886.
[207] Freitas, T.V; Tavares, A.P.; Theakston, R.D.; Laing, G.; New, R.R. Use of liposomes for protective immunisation against Crotalus
durissus (tropical rattlesnake) venom. Toxicon, 1989, 27(3), 341-347.
[208] Freitas, T.V.; Frézard, F. Encapsulation of native crotoxin in liposomes: a safe approach for the production of antivenom and vaccination against Crotalus durissus terrificus venom. Toxicon, 1997, 35(1), 91-100.
[209] Alam, M.I.; Gomes, A. Adjuvant effects and antiserum action potentiation by a (herbal) compound 2-hydroxy-4-methoxy benzoic acid isolated from the root extract of the indian medicinal plant “sarsaparilla” (Hemidesmus indicus r. br.). Toxicon, 1998, 36(10), 1423-1431.
[210] Chaves, F.; Chacón, M.; Badilla, B.; Arévalo, C. Effect of Echinacea purpurea (Asteraceae) aqueous extract on antibody response to Bothrops asper venom and immune cell response. Rev.
Biol. Trop., 2007, 55(1), 113-119. [211] Judge, R.K.; Henry, P.J.; Mirtschin, P.; Jelinek, G.; Wilce, J.A.
Toxins not neutralized by brown snake antivenom. Toxicol. Appl. Pharmacol., 2006, 213(2), 117-125.
[212] Soares, A.M.; Sestito, W.P.; Marcussi, S.; Stábeli, R.G.; Andrião-Escarso, S.H.; Cunha, O.A.; Vieira, C.A.; Giglio, J.R. Alkylation of myotoxic phospholipases A2 in Bothrops moojeni venom: a promising approach to an enhanced antivenom production. Int. J.
Biochem. Cell Biol., 2004, 36(2), 258-270. [213] Sewall, H. Experiments on the preventive inoculation of rattlesnake
venom. J. Physiol., 1887, 8(3-4), 203-210. [214] Sawai, Y. A study on the prophylactic toxoid against venom of
Asian snakes. Final report, N° FE-356-2. Contract No. DAJB17-67-C-0610, U.S. Army Research and Development Group (Far East) 1968; http://www.normanbenoit.com/Habu%20project.pdf.pdf.
[215] Sadahiro, S.; Kondo, S.; Kondo, H.; Ohsaka, A.; Fukushima, H.; Murata, R. Standardization of habu (Trimeresurus flavoviridis) snake-venom toxoid. Toxicon, 1978, 16(3), 275-282.
[216] Gutiérrez, J.M.; León, G.; Rojas, G.; Lomonte, B.; Rucavado, A.; Chaves, F. Neutralization of local tissue damage induced by Bothrops asper (terciopelo) snake venom. Toxicon, 1998, 36(11), 1529-1538.
[217] Rucavado, A.; Lomonte, B. Neutralization of myonecrosis, hemorrhage and edema induced by Bothrops asper snake venom by homologous and heterologous pre-existing antibodies in mice. Toxicon, 1996, 34(5), 567-577.
[218] Chippaux, J.; Goyffon, M. Production and use in snake antivenin. In: Handbook of Natural Toxins; Tu, A.T. Ed.; Marcel Dekker: New York, 1991, pp 529–555.
[219] Warrell, D.A. Geographical and intraspecies variation in the clinical manifestations of envenoming by snakes. In: Venomous
Snakes. Ecology, Evolution and Snakebite; Thorpe, R.S., Wüster, W., Melhotra, A. Eds.; Clarendon Press: Oxford, 1997, pp. 189-203.
[220] Fry, B.G.; Winkel, K.D.; Wickramaratna, J.C.; Hodgson, W.C.; Wuster, W. Effectiveness of snake antivenom: species and regional
venom variation and its clinical impact. J. Toxicol. Toxin. Rev., 2003, 22(1), 23-34.
[221] Camey, K.U.; Velarde, D.T.; Sanchez, E.F. Pharmacological characterization and neutralization of the venoms used in the production of Bothropic antivenom in Brazil. Toxicon, 2002, 40(5), 501-509.
[222] Raweerith, R.; Ratanabanangkoon, K. Immunochemical and biochemical comparisons of equine monovalent and polyvalent snake antivenoms. Toxicon, 2005, 45(3), 369-375.
[223] Stephano, M.A.; Guidolin, R.; Higashi, H.G.; Higashi, H.G.; Tambourgi, D.V.; Sant’Anna, O.A. The improvement of the therapeutic anti-Lachesis muta serum production in horses. Toxicon, 2005, 45(4), 467-473.
[224] Harrison, R.A. Development of venom toxin-specific antibodies by DNA immunization: rationale and strategies to improve therapy of viper envenoming. Vaccine, 2004, 22(13-14), 1648-1655.
[225] Wagstaff, S.C.; Laing, G.D.; Theakston, R.D.G.; Papaspyridis, C.; Harrison, R.A. Bioinformatics and multiepitope DNA immunization to design rational snake antivenom. PLoS Med, [On Line] 2006, 3(6), 832-844. http://plosmedicine.org/ [Accessed on: 06 June 2006].
[226] Harrison, R.A.; Moura-da-Silva, A.M.; Laing, G.D.; Wu, Y.; Richards, A.; Broadhead, A.; Bianco, A.E.; Theakston, R.D. Antibody from mice immunized with DNA enconded the carboxil-disintegrin and cysteine-rich domain (jD9) of the haemorrhagic metalloproteinase, jararhagin, inhibits the main lethal component of viper venom. Clin Exp Immunol, 2000, 121(2), 358-363.
[227] Arce-Estrada, V.; Azofeifa-Cordero, G.; Estrada, R.; Alape-Girón, A.; Flores-Díaz, M. Neutralization of venom-induced hemorrhage by equine antibodies raised by immunization with a plasmid encoding a novel P-II metalloproteinase from the lancehead pitviper Bothrops asper. Vaccine, 2009, 27(3), 460-466.
[228] Azofeifa-Cordero, G.; Arce-Estrada, V.; Flores-Díaz, M.; Alape-Girón, A. Immunization with cDNA of a novel P-III type metalloproteinase from the rattlesnake Crotalus durissus durissus
elicits antibodies which neutralize 69% of the hemorrhage induced by the whole venom. Toxicon, 2008, 52(2), 302-308.
[229] Pergolizzi, R.G.; Dragos, R.; Ropper, A.E.; Menez, A.; Crystal, R.G. Protective immunity against a-cobratoxin following a single administration of a genetic vaccine encoding a non-toxic cobratoxin variant. Hum. Gene. Ther., 2005, 16(3), 292-298.
[230] Gutiérrez, J.M.; Chaves, F.; Rojas, E.; Elizondo, J.; Avila, C.; Cerdas, L. Production of monovalent anti-Bothrops asper
antivenom: development of immune response in horses and neutralizing ability. Rev. Biol. Trop., 1988, 36(2B), 511-517.
[231] Estrada, R.; Gutiérrez, J.M.; Alvarado, J.; Robles, A.; Avila, C.; Gonzáles, N. Desarrollo de la respuesta de anticuerpos anti-fosfolipasa A2 en caballos inoculados con veneno para la producción de suero antiofídico polivalente en Costa Rica. Rev.
Biol. Trop., 1989, 37(2), 187-191. [232] Christensen, P.A. South African Snake Venoms and Antivenoms;
The South African Institute for Medical Research: Johannesburg, 1955, pp. 52-57.
[233] Russell, F.; Timmerman, W.; Meadows, P. Clinical use of antivenin prepared from goat serum. Toxicon, 1970, 8(1), 63-65.
[234] Fernández, G.P.; Segura, A.; Herrera, M.; Velasco, W.; Solano, G.; Gutiérrez, J.M.; León, G. Neutralization of Bothrops
mattogrossensis snake venom from Bolivia: Experimental evaluation of llama and donkey antivenoms produced by caprylic acid precipitation. Toxicon, 2009, 55(2-3), 642-645.
[235] Helms, C.M.; Allen, P.Z. Studies on equine immunoglobulins. II. Antigenic interrelationships among horse IgG, IgG(T) and antipneumococcal 1 component. J. Immunol., 1970, 105(5), 1253-1263.
[236] Widders, P.; Stokes, C.R.; Bourne, F.J. Investigation of the antigenic relationship between equine IgG and IgGT. Vet. Imm. Imm., 1986, 13(3), 255-259.
[237] Fernandes, I.; Takehara, H.A.; Mota, I. Isolation of IgGT from hyperimmune horse anti-snake venom serum: its protective ability. Toxicon, 1991, 29(11), 1373-1379.
[238] Fernandes, I.; Takehara, H.A.; Santos, A.C.; Cormont, F.; Latinne, D.; Barzin, H.; Mota, I. Neutralizing of bothropic and crotalic venom toxic activities by IgG(T) and IgGa subclasses isolated from immune horse serum. Toxicon, 1997, 35(6), 931-936.
[239] Fernandes, I.; Tavares, F.L.; Sano-Martins, I.S.; Takehara, H.A. Efficacy of bothropic antivenom and its IgG (T) fraction in

18 Inflammation & Allergy - Drug Targets, 2011, Vol. 10, No. 5 León et al.
restoring fibrinogen levels of Bothrops jararca envenomed mice. Toxicon, 2000, 38(7), 995-998.
[240] Grandgeorge, M.; Veron, J.L.; Lutsch, C.; Makula, M.F.; Riffard, P.; Pepin, S.; Scherrmann, J.M. Preparation of improved F(ab´)2 antivenoms. An example: New polyvalent european viper antivenom (equine). In: Envenomings and Their Treatments; Bon, C., Goyffon, M. Eds.; Editions Fondation Marcel Merieux: Lyon, 1996, pp. 161-172.
[241] Wagner, B. Immunoglobulins and immunoglobulin genes of the horse. Devel. Comp. Immunol., 2006, 30(1-2), 155-164.
[242] Sjostrom, L.; AL-Abdulla, I.H.; Rawat, S.; Smith, D.C.; Landon, J. A comparison of ovine and equine antivenoms. Toxicon, 1994, 32(4), 427-433.
[243] Landon, J.; Smith, D.S. Merits of sheep antisera for antivenom manufacture. J. Toxicol. Toxin. Rev., 2003; 22(1), 15-22.
[244] Herrera, M.; León, G.; Segura, A.; Meneses, F.; Lomonte, B.; Chippaux, J.P.; Gutiérrez, J.M. Factors associated with adverse reactions induced by caprylic acid-fractionated whole IgG preparations: comparison between horse, sheep and camel IgGs. Toxicon, 2005, 46(7), 775-781.
[245] Redwan, E.R.M.; Fahmy, A.; Hanafy, A.E.; EL-Baky, N.A. Ovine anti-rabies antibody production and evaluation. Comp. Immunol. Microbiol., 2009, 32(1), 9-19.
[246] Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hamers, C.; Songa, E.B.; Bendahman, N.; Hamers, R. Naturally occurring antibodies devoid of light chains. Nature, 1993, 363(6428), 446-448.
[247] Muyldermans, S.; Baral, T.; Retamozzo, V.C.; De Baetselier, P.; De Genst, E.; Kinne, J.; Leonhardt, H.; Magez, S.; Nguyen, V.K.; Revets, H.; Rothbauer, U.; Stijlemans, B.; Tillib, S.; Wernery, U.; Wyns, L.; Hassanzadeh-Ghassabeh, G.; Saerens, D. Camelid immunoglobulins and nanobody technology. Vet. Immunol. Immunop., 2009, 128(1-3), 178–183.
[248] Meddeb-Mouelhi, F.; Bouhaouala-Zahar, B.; Benlasfar, Z.; Hammadi, M.; Mejri, T.; Moslah, M.; Karoui, H.; Khorchani, T.; El Ayeb, M. Immunized camel sera and derived immunoglobulin subclasses neutralizing Androctonus australis hector scorpion toxins. Toxicon, 2003, 42(7), 785-791.
[249] De Simone, E.; Saccodossi, N.; Ferrari, A.; Leoni, L.; Leoni, J. Immunochemical analysis of IgG subclasses and IgM in South American camelids. Small Ruminant Res., 2006, 64(1-2), 2-9.
[250] Almeida, C.M.; da Silva, C.L.; Couto, H.P.; Escocard, Rde. C.; da Rocha, D.G.; Sentinelli, Lde. P.; Kipnis, T.L.; da Silva, W.D. Development of process to produce polyvalent IgY antibodies anti-African snake venom. Toxicon, 2008, 52(2), 293-301.
[251] Meenatchisundaram, S.; Parameswari, G.; Michael, A.; Ramalingam, S. Neutralization of the pharmacological effects of Cobra and Krait venoms by chicken egg yolk antibodies. Toxicon, 2008, 52(2), 221-227.
[252] McLaren, R.D.; Prosser, C.G.; Grieve, R.C.; Borissenko, M. The use of caprylic acid for the extraction of the immunoglobulin fraction from egg yolk of chickens immunized with ovine alpha-lactalbumin. J. Immunol. Meth., 1994, 177(1-2), 175-184.
[253] Almeida, C.M.; Kanashiro, M.M.; Rangel Filho, F.B.; Mata, M.F.; Kipnis, T.L.; da Silva, W.D. Development of snake antivenom antibodies in chickens and their purification from yolk. Vet. Rec., 1998, 143(21), 579-584.
[254] Warr, G.W.; Magor, K.E.; Higgins, D.A. IgY: clues to the origins of modern antibodies. Immunol. Today, 1995, 16(8), 392-398.
[255] Sevcik, C.; Díaz, P.; D’Suze, G. On the presence of antibodies against bovine, equine and poultry immunoglobulins in human IgG
preparations, and its implications on antivenom production. Toxicon, 2008, 51(1), 10-16.
[256] Gutiérrez, J.M.; Rojas, G.; Lomonte, B.; Gené, J.A.; Chaves, F.; Alvarado, J.; Rojas, E. Standardization of assays for testing the neutralizing ability of antivenoms. Toxicon, 1990, 28(10), 1127-1129.
[257] Bogarín, G.; Morais, J.F.; Yamaguchi, I.K.; Stephano, M.A.; Marcelino, J.R.; Nishikawa, A.K.; Guidolin, R.; Rojas, G.; Higashi, H.G.; Gutiérrez, J.M. Neutralization of crotaline snake venoms from Central and South America by antivenoms produced in Brazil and Costa Rica. Toxicon, 2000, 38(10), 1429-1441.
[258] Rojas, E.; Quesada, L.; Arce, V.; Lomonte, B.; Rojas, G.; Gutiérrez, J.M. Neutralization of four Peruvian Bothrops sp. snake venoms by polyvalent antivenoms produced in Perú and Costa Rica: preclinical assessment. Acta Trop., 2005, 93(1), 85-95.
[259] Araujo, H.P.; Bourguignon, S.C.; Boller, M.A.; Dias, A.A.; Lucas, E.P.; Santos, I.C.; Delgado, I.F. Potency evaluation of antivenoms in Brazil: The national control laboratory experience between 2000 and 2006. Toxicon, 2008, 51(4), 502-514.
[260] Meier, J.; Theakston, R.D.G. Approximate LD50 determinations of snake venoms using eight to ten experimental animals. Toxicon, 1986, 24(4), 395-401.
[261] Sells, P.G. Animal experimentation in snake venom research and in
vitro alternatives. Toxicon, 2003, 42(2), 115-133. [262] Gutiérrez, J.M.; Avila, C.; Rojas, E.; Cerdas, L. An alternative in
vitro method for testing the potency of the polyvalent antivenom produced in Costa Rica. Toxicon, 1988, 26(4), 411-413.
[263] Pradhan, S.; Kumar, S.; Singh, D.; Sood, R.C.; Sehgal, R. Development of passive haemagglutination (PHA) an haemagglutination inhibition (HAI) technique for potency estimation of Cobra antisnake venom serum (ASVS). Biologicals, 2007, 35(3), 155-160.
[264] Theakston, R.D.; Reid, H.A. Enzyme-linked immunosorbent assay (ELISA) in assessing antivenom potency. Toxicon, 1979, 17(5), 511-515.
[265] Price, J.A.; Sanny, C.G. CroFabTM total anti-venom activity measured by SE-HPLC, and anti PLA2 activity assayed in vitro at physiological pH. Toxicon, 2007, 49(6), 848-854.
[266] Heneine, L.G.; Carvalho, A.D.J.; Barbosa, C.F.; Arávio dos Santos, M.R. Development of an ELISA to assess the potency of horse therapeutic polyvalent antibothropic antivenom. Toxicon, 1998, 36(10), 1363-1370.
[267] Maria, W.S.; Cambuy, M.O.; Costa, J.O.; Velarde, D.T.; Chávez-Olórtequi, C. Neutralizing potency of horse antibothropic antivenom. Correlation between in vivo and in vitro methods. Toxicon, 1998, 36(10), 1433-1439.
[268] Rial, A.; Morais, V.; Rossi, S.; Massaldi, H. A new ELISA for determination of potency in snake antivenoms. Toxicon, 2007, 48(4), 462-466.
[269] Alape-Girón, A.; Miranda-Arrieta, K.; Cortes-Bratti, X.; Stiles, B.G.; Gutiérrez, J.M. A comparison of in vitro methods for assessing the potency of therapeutic antisera against the venom of the coral snake Micrurus nigrocinctus. Toxicon, 1997, 35(4), 573-581.
[270] Maria, W.S.; Pacheco, B.G.; Barbosa, C.F.; Velarde, D.T.; Chávez-Olórtequi, C. Determination of the neutralizing potency of horse antibothropic and anticrotalic antivenoms in blood samples collected on filter paper. Toxicon, 2001, 39(10), 1607-1609.
[271] Judge, R.K.; Henry, P.J.; Mirtschin, P.; Jelinek, K.; Wilce, J.A. Toxins not neutralized by brown snake antivenom. Toxicol. Appl. Pharm., 2006, 213(2), 117-125.
Received: August 2, 2010 Revised: February 15, 2011 Accepted: February 17, 2011
Related Documents











![Vitamin C 24/7 · aluminum, fluorine); [Levy, 2002, Curing the Incurable, pp. 280-312] Venoms (snake, spider); Klenner (1971) Observations of the dose and administration of ascorbic](https://static.cupdf.com/doc/110x72/5f0226677e708231d402d1b4/vitamin-c-247-aluminum-fluorine-levy-2002-curing-the-incurable-pp-280-312.jpg)