Introduction The human intestinal mucosal surface provides approxi- mately 400 m 2 of available surface area for intimate contact with antigen. It is now a generally accepted phenomenon that this constant exposure to dietary antigens, consisting of both proteinaceous and non-protein components, as well as a complex mix of bacterial microflora that have taken up resi- dence in the intestinal ecosystem, is associated with a sub- dued systemic immune hyporesponsiveness known as tolerance. Paradoxically, the mammalian intestinal archi- tecture has evolved with organized inductive and effector sites for immunological responsiveness to enteric antigen. So how can one reconcile conflicting requirements for immune non-responsiveness and responsiveness in the gut? In partic- ular, what are the available mechanisms for the gut mucosal immune system to discriminate, at the molecular level, between harmless food antigens and those associated with pathogenic and non-pathogenic microbes that may have inad- vertently gained entry via the oral orifice? Alternatively, does it really discriminate as discerningly as we have been led to believe? Tolerance The default immune response in the gut to nominal antigen is one of tolerance. 1 This phenomenon has been most exten- sively studied by intragastric feeding of soluble antigen in various animal models. Animals that have been previously fed an antigen, such as ovalbumin (OVA) or lysozyme, mount a suppressed cellular and humoral immune response after subsequent systemic vaccination with that antigen, usually when formulated with an adjuvant. The success of the toler- ance model hinges on the demonstration that the level of immunity generated post-vaccination in antigen-fed animals is suppressed relative to unfed controls and, unfortunately, appears to preclude the possibility that immune responses may have been primed as a consequence of antigen feeding. Tolerance models exemplify why food consumption does not always lead to clinical hypersensitivity and food allergies. They have also led to the development of oral strategies for the treatment of autoimmune disorders. 2,3 A noteworthy aspect of all tolerance induction models is that dietary feeding of a pure antigen has never been observed to com- pletely eliminate immune responses against that antigen fol- lowing vaccination. Immunity is always generated, but the magnitude of the response is reduced. Because tolerance is antigen specific and because protein antigens are rarely ever consumed singly and in a pure state in our diets, it becomes conceptually overwhelming to consider how tolerance can operate against the entire spectrum of non-self antigens being consumed daily. The dietary milieu in which a specific antigen is presented to the gut mucosa may have a profound Immunology and Cell Biology (2000) 78, 55–66 Special Feature Immune response to orally consumed antigens and probiotic bacteria JAMES CHIN, BERNADETTE TURNER, IDRIS BARCHIA and ARNO MULLBACHER* Microbiology and Immunology, Elizabeth Macarthur Agricultural Institute, Sydney, New South Wales and John Curtin School of Medical Research, Australian National University, Canberra, Australian Capital Territory, Australia Summary The gut mucosal system must fulfil conflicting roles in suppressing immune responses against orally fed antigens (tolerance) while still retaining the ability to respond to potential enteric pathogens. It must also, to a large degree, not mount an immune response against commensal enteric bacteria and the administration of large numbers of probiotic bacteria formulated as dietary supplements in food products. Contrary to this dogma, it has been found that feeding ovalbumin as a marker antigen, in association with selected probiotic bacteria, appears to prime for an intestinal immune response that is further augmented by skin vaccination. Skin immunization is known to stimulate a strong innate, humoral and cellular immune response. Such dominant immunogenic signals appear to override tolerogenic signals engendered by oral feeding of antigen. High-dose antigen feeding stimulated a strong Th2-dependent antibody response to skin vaccination but completely suppressed cytotoxic T cell responses. This was true even when ovalbumin was administered in conjunction with various selected probiotic bacteria. However, while yeast appeared to be better at priming for an enhanced humoral response, Lactobacillus fermen- tum and Staphylococcus carnosus were more effective in enhancing the postvaccinal lymphoproliferative response against ovalbumin. Key words: dietary antigen, gut immunity, lymphoproliferation, mucosal immunity, oral tolerance, probiotic bacteria. Correspondence: Dr J Chin, Immunology, Elizabeth Macarthur Agricultural Institute, PMB 8, Camden, NSW 2570, Australia. Email: [email protected] *Present address: Arno Mullbacher, John Curtin School of Medical Research, Australian National University, Canberra, ACT 0200, Australia. Received 13 September 1999; accepted 13 September 1999.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Introduction
The human intestinal mucosal surface provides approxi-mately 400 m2 of available surface area for intimate contactwith antigen. It is now a generally accepted phenomenon thatthis constant exposure to dietary antigens, consisting of bothproteinaceous and non-protein components, as well as acomplex mix of bacterial microflora that have taken up resi-dence in the intestinal ecosystem, is associated with a sub-dued systemic immune hyporesponsiveness known astolerance. Paradoxically, the mammalian intestinal archi-tecture has evolved with organized inductive and effectorsites for immunological responsiveness to enteric antigen. Sohow can one reconcile conflicting requirements for immunenon-responsiveness and responsiveness in the gut? In partic-ular, what are the available mechanisms for the gut mucosalimmune system to discriminate, at the molecular level,between harmless food antigens and those associated withpathogenic and non-pathogenic microbes that may have inad-vertently gained entry via the oral orifice? Alternatively, doesit really discriminate as discerningly as we have been led tobelieve?
Tolerance
The default immune response in the gut to nominal antigenis one of tolerance.1 This phenomenon has been most exten-sively studied by intragastric feeding of soluble antigen invarious animal models. Animals that have been previouslyfed an antigen, such as ovalbumin (OVA) or lysozyme, mounta suppressed cellular and humoral immune response aftersubsequent systemic vaccination with that antigen, usuallywhen formulated with an adjuvant. The success of the toler-ance model hinges on the demonstration that the level ofimmunity generated post-vaccination in antigen-fed animalsis suppressed relative to unfed controls and, unfortunately,appears to preclude the possibility that immune responsesmay have been primed as a consequence of antigen feeding.
Tolerance models exemplify why food consumption doesnot always lead to clinical hypersensitivity and food allergies.They have also led to the development of oral strategies forthe treatment of autoimmune disorders.2,3 A noteworthyaspect of all tolerance induction models is that dietaryfeeding of a pure antigen has never been observed to com-pletely eliminate immune responses against that antigen fol-lowing vaccination. Immunity is always generated, but themagnitude of the response is reduced. Because tolerance isantigen specific and because protein antigens are rarely everconsumed singly and in a pure state in our diets, it becomesconceptually overwhelming to consider how tolerance canoperate against the entire spectrum of non-self antigens beingconsumed daily. The dietary milieu in which a specificantigen is presented to the gut mucosa may have a profound
Immunology and Cell Biology (2000) 78, 55–66
Special Feature
Immune response to orally consumed antigens and probioticbacteria
JAMES CHIN, BERNADETTE TURNER, IDRIS BARCHIA and ARNO MULLBACHER*
Microbiology and Immunology, Elizabeth Macarthur Agricultural Institute, Sydney, New South Wales and John CurtinSchool of Medical Research, Australian National University, Canberra, Australian Capital Territory, Australia
Summary The gut mucosal system must fulfil conflicting roles in suppressing immune responses against orallyfed antigens (tolerance) while still retaining the ability to respond to potential enteric pathogens. It must also, to alarge degree, not mount an immune response against commensal enteric bacteria and the administration of largenumbers of probiotic bacteria formulated as dietary supplements in food products. Contrary to this dogma, it hasbeen found that feeding ovalbumin as a marker antigen, in association with selected probiotic bacteria, appears toprime for an intestinal immune response that is further augmented by skin vaccination. Skin immunization is knownto stimulate a strong innate, humoral and cellular immune response. Such dominant immunogenic signals appearto override tolerogenic signals engendered by oral feeding of antigen. High-dose antigen feeding stimulated astrong Th2-dependent antibody response to skin vaccination but completely suppressed cytotoxic T cell responses.This was true even when ovalbumin was administered in conjunction with various selected probiotic bacteria.However, while yeast appeared to be better at priming for an enhanced humoral response, Lactobacillus fermen-tum and Staphylococcus carnosus were more effective in enhancing the postvaccinal lymphoproliferative responseagainst ovalbumin.
Key words: dietary antigen, gut immunity, lymphoproliferation, mucosal immunity, oral tolerance, probiotic bacteria.
Correspondence: Dr J Chin, Immunology, Elizabeth MacarthurAgricultural Institute, PMB 8, Camden, NSW 2570, Australia.Email: [email protected]
*Present address: Arno Mullbacher, John Curtin School ofMedical Research, Australian National University, Canberra, ACT0200, Australia.
Received 13 September 1999; accepted 13 September 1999.
influence on tolerance induction. While tolerogenic signals(from orally dosed antigen) appear to dominate opposingimmunogenic signals engendered by peripheral vaccination,4
the choice of adjuvants and the route of vaccination may shiftthe tolerance tightrope in favour of an enhanced immuneresponse.
By ignoring measurements of immunity post-feeding andat any time point prior to systemic vaccination, oral tolerancemodels may have failed to acknowledge the possibility thatorally fed antigens can gain access systemically5 and therebyprime for an immune response. Hanson has found thatnewborn mice fed OVA within 48 h of birth become primedand not tolerized for an anti-OVA antibody response.6 Oraltolerance first becomes detectable in mice between 4 and7 days of age and remains until adult age, except for anarrow window between 23 and 26 days after birth. Thissecond period of loss of tolerizability coincides with the timeof weaning. There have been reports of priming for systemicantibody responses against dietary antigens. Bailey et al.have shown that piglets weaned onto soya at 3 weeks of agedevelop serum IgG antisoya antibodies comparable to thatgenerated by vaccination with adjuvanted soya.7 The oral tolerance model is therefore not an invariant model and anorally fed antigen can sometimes fulfil dual roles in provid-ing an immunogenic as well as a tolerogenic signal.
Can orally fed antigen prime for a systemic immuneresponse?
We adopted a classical protocol for OVA feeding in conjunc-tion with skin immunization to explore the possibility that thechoice of adjuvant and route of vaccination8 might be impor-tant in how a tolerogenic OVA-fed antigen may be seen by theimmune system as an immunogenic signal. In these experi-ments, 6–8-week-old C57BL mice were each fed three dosesof OVA on alternate days. The OVA was administered byintragastric intubation at a high (20 mg per dose) or low(200 µg per dose) dose to two groups of mice (n = 6). Unfedcontrol mice were not fed any OVA. A week later, mice wereinjected twice intradermally, 7 days apart, with OVA (100 µg)adjuvanted in the form of immune-stimulating complexes orISCOM.9 The experiment was replicated in three sets so thatimmune responses in high-dose OVA-fed mice (HDOV), low-dose OVA-fed mice (LDOV) and unfed controls could bedetermined at 7 days post-feeding (PF), at 7 days after onevaccination (post-vaccination 1 or PV1) and, finally, at 7 daysafter two vaccinations (PV2). Blood was collected fromkilled mice and serum IgG responses against OVA weredetermined by ELISA. Spleens were also removed andsplenocytes cultured in vitro with various concentrations ofOVA. Lymphoproliferative (LP) responses were determinedby the incorporation of tritiated thymidine into DNA. Thestimulation index was calculated as a ratio of the amount ofradioactivity incorporated into cells cultured in the presenceand absence of OVA, which provided an assessment of cellu-lar immunity (CMI).
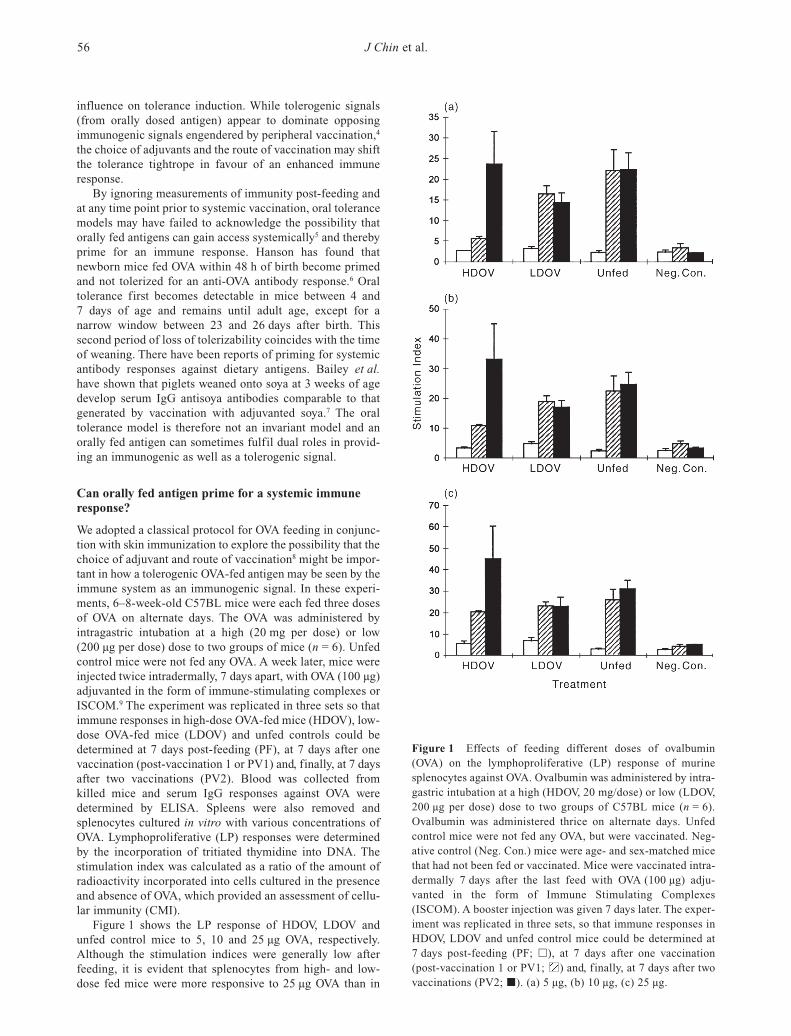
Figure 1 shows the LP response of HDOV, LDOV andunfed control mice to 5, 10 and 25 µg OVA, respectively.Although the stimulation indices were generally low afterfeeding, it is evident that splenocytes from high- and low-dose fed mice were more responsive to 25 µg OVA than in
J Chin et al.56
Figure 1 Effects of feeding different doses of ovalbumin(OVA) on the lymphoproliferative (LP) response of murinesplenocytes against OVA. Ovalbumin was administered by intra-gastric intubation at a high (HDOV, 20 mg/dose) or low (LDOV,200 µg per dose) dose to two groups of C57BL mice (n = 6).Ovalbumin was administered thrice on alternate days. Unfedcontrol mice were not fed any OVA, but were vaccinated. Neg-ative control (Neg. Con.) mice were age- and sex-matched micethat had not been fed or vaccinated. Mice were vaccinated intra-dermally 7 days after the last feed with OVA (100 µg) adju-vanted in the form of Immune Stimulating Complexes(ISCOM). A booster injection was given 7 days later. The exper-iment was replicated in three sets, so that immune responses inHDOV, LDOV and unfed control mice could be determined at7 days post-feeding (PF; h), at 7 days after one vaccination(post-vaccination 1 or PV1; ) and, finally, at 7 days after twovaccinations (PV2; j). (a) 5 µg, (b) 10 µg, (c) 25 µg.
unfed controls (Fig. 1c). This is indicative that both low- andhigh-dose OVA feeding had primed for a CMI response. Anti-bodies against OVA were not detected post-feeding in anytreatment group.
After one vaccination (Fig. 1a), LP responses against 5 µgOVA were significantly suppressed in HDOV mice (P < 0.01)compared with unfed controls, while LDOV mice were inter-mediate in their stimulation indices. Increasing amounts ofOVA in the LP assays reduced the gap differential in stimu-lation indices between the three groups and also their level ofsignificance (Fig. 1b,c). Therefore, high-dose OVA feedingwas able to induce tolerance better than low-dose feeding inresponse to a single vaccination and this was more evidentwhen the amount of OVA used in LP assays was low. TheCMI response after two vaccinations was completely differ-ent between HDOV and LDOV mice. While low-dose OVAfeeding was able to maintain a lower but non-significant levelof lymphoproliferation compared to unfed mice, high-doseOVA feeding was able to stimulate a higher LP response. Thisincreased CMI response also increased in absolute termsbased on the stimulation index with increasing levels of OVAin the culture wells. The use of splenocytes in LP culturesinstead of enriched lymphocyte subpopulations was intendedto simulate, in vitro, the complexity of processes ongoing inthe spleen in vivo. Clearly, the amount of OVA antigen pro-vided by feeding, vaccination or by exogenous addition invitro can influence CMI assessment, probably by retuning theability of antigen-presenting cells (APC) to present OVApeptide in situ. Because stimulation, as well as suppression,can be demonstrated by the availability or deprivation ofantigen in the assay system, the case for tolerance inductionbased on in vitro LP assays may be biologically meaningless,even though this is carried out regularly in many animalmodels for tolerance induction.
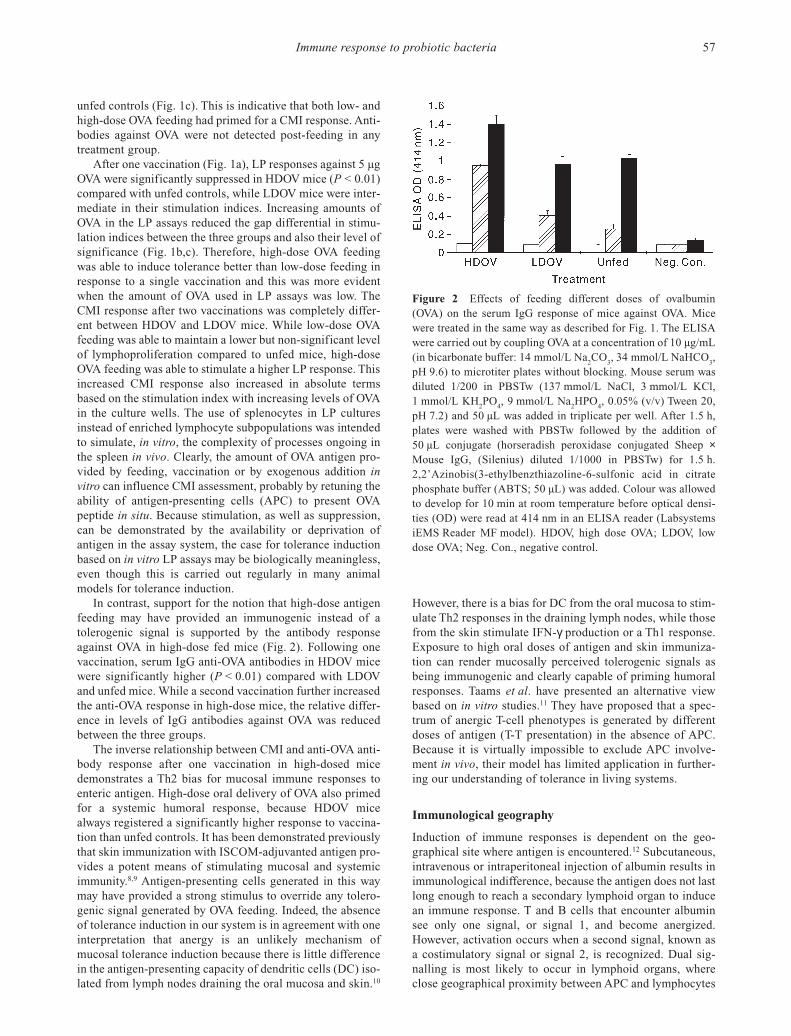
In contrast, support for the notion that high-dose antigenfeeding may have provided an immunogenic instead of atolerogenic signal is supported by the antibody responseagainst OVA in high-dose fed mice (Fig. 2). Following onevaccination, serum IgG anti-OVA antibodies in HDOV micewere significantly higher (P < 0.01) compared with LDOVand unfed mice. While a second vaccination further increasedthe anti-OVA response in high-dose mice, the relative differ-ence in levels of IgG antibodies against OVA was reducedbetween the three groups.
The inverse relationship between CMI and anti-OVA anti-body response after one vaccination in high-dosed micedemonstrates a Th2 bias for mucosal immune responses toenteric antigen. High-dose oral delivery of OVA also primedfor a systemic humoral response, because HDOV micealways registered a significantly higher response to vaccina-tion than unfed controls. It has been demonstrated previouslythat skin immunization with ISCOM-adjuvanted antigen pro-vides a potent means of stimulating mucosal and systemicimmunity.8,9 Antigen-presenting cells generated in this waymay have provided a strong stimulus to override any tolero-genic signal generated by OVA feeding. Indeed, the absenceof tolerance induction in our system is in agreement with oneinterpretation that anergy is an unlikely mechanism ofmucosal tolerance induction because there is little differencein the antigen-presenting capacity of dendritic cells (DC) iso-lated from lymph nodes draining the oral mucosa and skin.10
However, there is a bias for DC from the oral mucosa to stim-ulate Th2 responses in the draining lymph nodes, while thosefrom the skin stimulate IFN-γ production or a Th1 response.Exposure to high oral doses of antigen and skin immuniza-tion can render mucosally perceived tolerogenic signals asbeing immunogenic and clearly capable of priming humoralresponses. Taams et al. have presented an alternative viewbased on in vitro studies.11 They have proposed that a spec-trum of anergic T-cell phenotypes is generated by differentdoses of antigen (T-T presentation) in the absence of APC.Because it is virtually impossible to exclude APC involve-ment in vivo, their model has limited application in further-ing our understanding of tolerance in living systems.
Immunological geography
Induction of immune responses is dependent on the geo-graphical site where antigen is encountered.12 Subcutaneous,intravenous or intraperitoneal injection of albumin results inimmunological indifference, because the antigen does not lastlong enough to reach a secondary lymphoid organ to inducean immune response. T and B cells that encounter albuminsee only one signal, or signal 1, and become anergized.However, activation occurs when a second signal, known asa costimulatory signal or signal 2, is recognized. Dual sig-nalling is most likely to occur in lymphoid organs, whereclose geographical proximity between APC and lymphocytes
Immune response to probiotic bacteria 57
Figure 2 Effects of feeding different doses of ovalbumin(OVA) on the serum IgG response of mice against OVA. Micewere treated in the same way as described for Fig. 1. The ELISAwere carried out by coupling OVA at a concentration of 10 µg/mL(in bicarbonate buffer: 14 mmol/L Na
2CO
3, 34 mmol/L NaHCO
3,
pH 9.6) to microtiter plates without blocking. Mouse serum wasdiluted 1/200 in PBSTw (137 mmol/L NaCl, 3 mmol/L KCl,1 mmol/L KH
2PO
4, 9 mmol/L Na
2HPO
4, 0.05% (v/v) Tween 20,
pH 7.2) and 50 µL was added in triplicate per well. After 1.5 h,plates were washed with PBSTw followed by the addition of50 µL conjugate (horseradish peroxidase conjugated Sheep ×Mouse IgG, (Silenius) diluted 1/1000 in PBSTw) for 1.5 h.2,2’Azinobis(3-ethylbenzthiazoline-6-sulfonic acid in citratephosphate buffer (ABTS; 50 µL) was added. Colour was allowedto develop for 10 min at room temperature before optical densi-ties (OD) were read at 414 nm in an ELISA reader (LabsystemsiEMS Reader MF model). HDOV, high dose OVA; LDOV, lowdose OVA; Neg. Con., negative control.

facilitate two-signal mediated activation. Although thismechanism has provided an explanation for tolerance induction when antigen is administered intravenously, it isprobably invalid to argue that oral tolerance induction is sim-ilarly induced because of the entry of dietary antigens intothe blood circulation. Unlike intravenous injection of antigen,the passage of consumed antigens from the lumen to blood inthe intestine proceeds via many geographically distinct cellular structures that envelop the intestinal epithelium. Asignificant proportion of these are immune cells with thecapacity to respond immunologically and it is thereforeimportant that the induction of immunity or tolerance todietary antigens should be considered in terms of the geo-graphy and anatomical structure of the intestine.13
Cellular architecture of the gut
A single layer of epithelial cells lines the entire intestinalmucosal surface. These cells not only provide a very thinbarrier for systemic protection against intestinal lumen con-tents, but they must also fulfil an important function in bi-directional signalling between the lumen and other immunecells that occupy the mucosae. Nested behind the epithelialcells is a unique population of lymphocytes known as intra-epithelial lymphocytes (IEL).
Intestinal epithelial cells
The small intestine arises from cytodifferentiation of theendoderm in late foetal life. In mice, the multipotent stemcell located near the base of crypts of Lieberkuhn undergoseveral rounds of cell division to give rise to four principalepithelial cell lineages, consisting of mucus-secreting gobletcells, enteroendocrine cells, absorptive enterocytes andPaneth cells.14 Progenitors of the latter three cell lineagesmigrate upwards to the apical extrusion zone of their associ-ated villi, while Paneth cells differentiate by downwardmigration to the base of the crypt. Paneth cells maintain amucosal defence barrier through the production of antimi-crobial peptides, such as cryptdins and lysozyme. They alsoproduce growth factors and other molecules that regulate theproliferation and differentiation of various epithelial cell lin-eages. The other cell lineages that migrate to the apex of theirassociated villus undergo apoptosis and exfoliate, resulting indramatic cycling of the gut epithelium every 3–5 days. Theformation and involution of crypt-villus units is completed inearly postnatal life, providing the greatest possible surfacearea for absorption of nutrients.
Peyer’s patches
The epithelium is surrounded by the lamina propria and, inturn, is enveloped by smooth muscle tissue. Lymphoidfollicular aggregates, known as Peyer’s patches (PP), areorganized discretely in the small intestine with their apicalcover of M cells facing the lumen. M cells are devoid ofmicrovilli, but are formed with folds and crevices that aid inthe entrapment of bacteria. They also possess specific recep-tors that are capable of binding bacteria such as Salmonella,Shigella and Yersinia. The presence of APC in closeproximity to T cells in PP supports the contention that this is
the primary inductive site for the priming of mucosalimmune responses,15,16 including the generation of suppressorT cells.17
Exogenous DNA from plants, animals and bacteria con-stitutes a major component in our diets. Such DNA has beenshown to be rapidly absorbed through the intestinal mucosaand incorporated by lymphocytes.18 Self-antigens, such asDNA and histones, are generated in PP by germinal centrereactions that chronically induce massive cell death. Shimodaet al. have been able to demonstrate the presence of IgA auto-antibodies against DNA by fusion of PP lymphocytes withX63-Ag8 myelomas.19 Self-priming antigens and foreignDNA in the gut can maintain selection processes favourablefor the production of anti-DNA antibodies found in humanand murine models of systemic lupus erythmatosus (SLE).
Intra-epithelial lymphocytes
Two lineages of IEL that line the intestine lumen are believedto originate from thymus-dependent (TD) and extrathymic orthymo-independent (TI) precursors in the gut wall.20 The TDpopulation consists of T cells that bear CD4 or heterodimericCD8αβ molecules, Thy-1 and αβ γδ Τ-cell receptors (TCR).The TI population expresses either αβ or γδ TCR and themajority of these cells bear the CD8 homodimeric αα mole-cules; a few are double negative and only some express Thy-1.There are approximately equal numbers of TI IEL bearing theαβ or γδTCR21 and they appear to play an important role inregulating local IgA responses22,23 and in mediating naturalkiller (NK)- and antibody-dependent cell-mediated cytotoxi-city (ADCC) functions. Unlike other NK cells, TI IEL withNK function are not regulated through recognition of majorhistocompatibility complex class I molecules and thereforeconstitute a unique lymphocyte subset with enhanced anti-genic repertoire diversity for participation in defence of themucosal epithelial barrier.
Lamina propria lymphocytes
The bulk of immune cells are located in the lamina propriaand include B and T cells, macrophages, dendritic cells (Pavli90) and polymorphonuclear leucocytes. Most of the re-circulating IgA antibody producing plasma cells are found inthe lamina propria. Lamina propria T cells mostly express theαβ TCR and possess, in addition, a memory/activated pheno-type characterized by the following surface markers:CD45RO+/CD45Rblo, CD44hi, L-sello and α
4β
7hi.24 Lamina
propria T cells respond poorly in vitro to antigen or TCRstimuli, but do secrete large amounts of IFN-γ, IL-4 and IL-5.25
Dendritic cells and antigen-presenting cells
Intestinal dendritic cells, like their peripheral counterparts,26
have been shown to be able to acquire orally presentedantigen and to present these to naïve T cells.27,28 Viney et al.have examined the role of intestinal DC in tolerance induc-tion by injecting mice with Flt3L, a growth factor known toexpand the number of DC in mucosal tissue and in lymphoidorgans.29 Surprisingly, they have found that injection of Flt3L prior to low-dose feeding (10–500 µg OVA) induces
J Chin et al.58

significant levels of tolerance after vaccination, comparedwith non-Flt3L-treated mice. The expanded DC populationdid not express costimulatory markers CD80 and CD86.However, they were responsive to stimulation with bacterialLPS as an inflammatory signal and displayed significantincreased expression of MHC II, CD80, CD86 and CD40.
Encounters with bacteria antigen and immune function
Resident lumenal antigens: Non-pathogenic intestinalmicroflora
During normal vaginal delivery, the newborn human infantcomes into contact with the maternal enteric bacteria con-taminating the perineum. These bacteria rapidly colonize theinfant’s intestine (a process known as conventionalization),reaching concentrations of 1011 per gram of stool, and remainthroughout the life of the individual.30 More than 99% ofthese bacteria are obligate anaerobes with at least 400 differ-ent species. These indigenous or autochthonous flora are notpathogenic and there is a steady physiological leak of smallnumbers of bacteria into the mesenteric lymph nodes andblood, a process termed translocation.31
In a revealing analysis of epithelium–microbial inter-actions using the gut as a model of an open mammalianecosystem, Bry et al. have shown staining of fucosylated epitopes on the surface of enterocytic, goblet and Paneth celllineages in conventional mice with labelled Ulex europaeustype 1 agglutinin (UEA1).14,32 Germ-free (GF) miceexpressed fucosylated glycoconjugates in their Paneth cellsonly. Introduction of Bacteroides thetaiotaomicron (a normalcommensal in the small intestine and colon of mice andhumans) to GF mice signalled the intestine to induce andsustain a 1,2 fucosyl-transferase gene transcription and pro-duction of fucosylated glycoproteins and glycolipids.33 Thissignalling was specific and could not be elicited with otherenteric anaerobes, such as Peptostreptococcus micros andBifidobacterium infantis. Signalling occurred without theneed for attachment of Bacteroides thetaiotaomicron toepithelial cells, but was dependent on attainment of a criticalmass of bacteria so that the appropriate number of signallingmolecules can be produced to induce a host response. Similarchanges have been reported,34 except that attachment of anindigenous segmented filamentous bacteria was required toinduce fucosylation, up-regulation of MHC II, expansion ofintestinal IEL (i-IEL) and increased IgA production.35
Populations of intestinal bacteria have been mapped bymolecular techniques and shown to be remarkably constantover time within an individual, but may vary substantiallybetween humans.36 There appears to be a continuous processof selection exerted possibly by the gut immune system andmatched by selective adaptation of specific microbiotastrains. It remains to be established whether an adapted pop-ulation within an individual is engendered by imprinting withcolonizers during the initial stages of conventionalization ofthe newborn infant’s sterile gut environment.
Pathogenic bacteria
Enteric pathogens, such as enterotoxigenic Escherichia coli(ETEC) or Vibrio cholerae, produce toxins that initiate diarrhoea. Non-toxin producing enteric pathogens, such as
Salmonella typhimurium, provoke an intense intestinalinflammatory response associated with transepithelial migra-tion of polymorphonuclear leucocytes into the lumen.37 It was thought that attachment of Salmonellae to enterocytes followed by invasion and transcytosis, or entry via M cells inPeyer’s patches, represented the main virulence mechanismfor initiating inflammatory mucosal responses. However,McCormick et al. have been able to demonstrate that Salmonella strains or serotypes that elicit diffuse enteritis in humans are able to induce transepithelial signalling to neutrophils across epithelial cell monolayers.38 Strains, suchas S. enteritidis, S. pullorum, S. typhi and S. paratyphi, thatdo not normally cause enteritis as well as invasion-defectivemutants of S. typhimurium do not display transepithelial signalling. Although the identity of the signalling factor hasnot been elucidated, it is conceivable that other bacteria mayhave the ability to aberrantly initiate the epithelial signallingloop. Clinical enteric disorders, such as inflammatory boweldisease (IBD), Crohn’s disease (CD) and ulcerative colitis(UC), which appear seemingly unrelated to colonization bypathogenic micro-organisms, may have been triggered by signalling from non-pathogenic bacteria.
Oral infection of mice with Listeria monocytogenes hasbeen found to trigger the production of abundant amounts ofIL-15 by intestinal epithelial cells (i-EC) and to increase thesynthesis of IFN-γ by i-IEL.39 The αβ and γδT cell subpop-ulations in these mice remain relatively unchanged afterinfection. It is believed that IL-15 produced by i-EC prefer-entially activates αβ i-IEL expressing CD8αα, as well as γδi-IEL, which exclusively express CD8αα . The bias towards aTh1 response early in listeriosis demonstrated another formof enterocyte signalling that is substantively different fromthat initiated by diarrhoeagenic Salmonellae.
Probiotic bacteria
In recent years, the food manufacturing sector has promul-gated the importance of functional foods as dietary supple-ments for promoting health. Probiotics represent a microbialfood supplement that beneficially influences the host byimproving its intestinal microbial balance. While the compo-sition of a balanced enteric microbial population that is con-ducive to healthiness has eluded scientific quantification, theindustry appears to have no hesitation in redefining probi-otics as micro-organisms that, when ingested, exert healthbenefits beyond inherent basic nutrition. Claims of healthbenefits should be clearly distinguished between resilience todisease and the psychological feeling of wellbeing. Proof ofthe former is difficult to demonstrate, because there havebeen no reports to date that convincingly prove that animalsfed probiotics are resistant to experimental challenge with apathogen. Until such experimental verification can be pro-vided, a non-specific de facto role as vaccines cannot beascribed for probiotic bacteria. Even clinical studies, such asthose carried out by Kaila et al. where feeding of Lacto-bacillus sp. strain GG was shown to promote clinical recov-ery from acute rotavirus diarrhoea,40 provide only associativeevidence that enhanced non-specific levels of IgG, IgA andIgM actually play a role in ameliorating the disease process.It has never been shown that these antibodies have the abilityto neutralize the virus.
Immune response to probiotic bacteria 59

While the feeling of wellbeing is generally considered tobe a psychosomatic parameter, there is increasing evidencethat immunological criteria can be used to assess this condi-tion. Environmental, physiological, social and nutritionalstressors inherent in intensive livestock animal productionfacilities have long been known to influence growth perfor-mance and health. Stressors act on the sympathetic nervoussystem (SNS) and the hypothalamic-pituitary-adrenal (HPA)axis, altering hormonal levels that in turn affect production ofcytokines by immune cells.41 While acute stressors areknown to stimulate immunity, chronic stress is believed todown-regulate immune responses.42 Use of immunologicalparameters to assess immune responses to dietary supple-mentation with various nutriceuticals or dietary and nutri-tional supplements may provide scientific validation ofpotential health benefits.
Immunological triggering strategies of bacteria
Bacteria possess a repertoire of strategies for triggeringimmune responses and only in recent years have many ofthese been unravelled. An innocuous commensal of thehuman oral mucosa, Streptococcus gordonii, was geneticallymodified to express OVA on its surface. This recombinantbacteria induced de novo synthesis of both MHC class I andII molecules in a mouse dendritic cell culture system and was106-fold more efficient in mediating OVA peptide antigenpresentation than soluble OVA.43 In certain situations, bac-teria can activate T cells by furnishing costimulatory signals.The lipid-containing recombinant outer surface lipoprotein Aof Borrelia burgdorferi, but not its delipidated derivative,augmented CD3-induced T cell proliferation in a dose-dependent manner and at levels similar to that obtained withanti-CD28 antibodies.44 The discovery that bacterial DNAcontaining CpG motifs can act as a ‘danger signal’ and hasthe capacity to switch on strong Th1 humoral and cell-mediated immune responses45 adds to their immuno-stimulatory arsenal. Finally, there is now increasing evidencethat antigen presentation to T cells may not be restricted topeptides presented via class I and II molecules. Lipid or lipo-glycan antigens of bacterial origin can be recognized bymannose receptors and delivered via endosomes for presen-tation to T cells by CD1b molecules.46 Because all bacteriaand antigens derived from them have the ability to provokeimmune responsiveness, and in view of the intimate interac-tions occurring at the intestinal lumen interface, it is morelikely that microbial signals are seen by the gut immunesystem as being immunogenic.
Dietary/enteric antigens
Form of oral antigen delivery
The dogma that feeding pure protein antigens in mice doesnot prime for a systemic antibody response against the fedantigen47 has, until now, not been seriously questioned. Thesefeeding regimes do not truly mimic normal dietary con-sumption of antigens where proteins are eaten together withother components, such as oil, fats and complex carbohy-drates. Kaneko et al. have found that serum IgG responsesagainst β-lactoglobulin are detected in BALB/c mice after the
mice are fed 10 daily doses of 25 mg antigen emulsified with50 µL soybean oil.48
Strain differences in the murine response to orally fedantigen
The ability to discern dietary antigens as immunogenicsignals and not as tolerogenic cues may be strain dependent.Immunoglobulin E antibodies against casein (CN) have beenelicited as early as 7 days after 6-week-old DBA/2 mice hadbeen fed a diet containing casein (400–500 mg/day) for 63days. Antibodies have not been detected in mice fed a similarregime of OVA, nor in BALB/c and B10.A mice that hadbeen fed either CN or OVA49 Faecal IgA against casein hasalso been detected in DBA/2 mice fed with casein, but notOVA. Flow cytometric analysis of freshly isolated lympho-cytes from i-IEL, PP, MLN Spl and liver have revealed littledifference between CN- and OVA-fed mice with respect tosubpopulations stained for the B220+, CD3+, CD4+CD8α–
and CD4–CD8α+ phenotypes. However, compared with OVA-fed mice, CN-fed mice have a significantly higher level ofCD3+TCRαβ+ and a lower level of CD3+TCRγδ+ i-IEL.Levels of CD3intermediate IL-2Rβ+ lymphocytes are also signif-icantly higher in CN-fed mice.
Reverse transcriptase polymerase chain reaction (RT-PCR) analysis of freshly isolated lymphocytes from i-IEL,MLN and liver has shown relatively higher levels of IL-4 andlower levels of IFN-γ in CN mice compared with the OVA-fedgroup. Although TGF-β mRNA levels have been found to besimilar in splenocytes from the two groups, they are muchhigher in lymphocytes from PP in CN-fed compared withOVA-fed mice. αβ Intestinal IEL express a higher level ofIL-4 mRNA and the γδ i-IEL express a higher level of TGF-βin CN compared with OVA-fed mice. Higher levels of transcribed messenger TGF-β RNA is consistent with theproduction of IgA in these mice and also plays a role in systemic tolerance.50 Once again, and consistent with the biasof mucosal antigens to prime for antibody responses, TCRtriggering with anti-CD3 of lymphocytes from spleen, MLNand liver in CN mice also elicits a predominantly Th2response marked by IL-4 production associated with concur-rently enhanced levels of serum IgE.
It is difficult to understand why strain differences canexplain tolerogenic or immunogenic responses to entericantigen. DBA/2 and BALB/c mice carry the same haplotype(H-2d) of MHC class II. DBA/2 mice are Ityr, while BALB/cand B10.A mice are Itys. The Ity gene locus encodes theNramp1 gene product, which is involved in the compartmen-talization or concentrating of substrates for NO synthase andplays an important role in limiting intracellular replication ofpathogenic bacteria such as Salmonella and Mycobacteriumin macrophages.51 DBA/2 mice also lack C5, which isinvolved in chemotaxis and activation of phagocytes. None ofthese genetic determinants can adequately explain theobserved differences in strain response to orally fed antigen.
Tolerance and commensal bacteria
The composition of enteric microbiota changes dramaticallyfrom birth, through weaning to adulthood, where the com-position remains extremely stable for each individual.
J Chin et al.60

Consumption of antibiotics also changes the populationdynamics of commensal bacteria in the intestine. Moreau andCorthier have found that oral tolerance is maintained in con-ventional mice for up to 3 months after feeding, comparedwith only 20 days in germ-free mice.52 In an attempt to abro-gate tolerance by feeding OVA together with cholera holo-toxin (CT) or OVA and Escherichia coli heat-labileenterotoxin (LT), Gaboriau-Routhiau and Moreau havereported that CT + OVA feeding abrogates tolerance andelicits better antibody titres in conventional than germ-freemice.53 However, LT + OVA feeding fails to abrogate toler-ance in germ-free mice and stimulates a strong antibodyresponse against OVA. In contrast, cofeeding LT and OVA inconventional mice fails to stimulate an immune response andis successful, instead, in generating a suppressed antibodyresponse similar to that produced by feeding with OVA alone.One interpretation of these observations is that enteric bac-teria in conventional animals may maintain a level ofbystander tolerance compared to that in germ-free mice.
Using a different approach, Karlsson et al. have con-structed an OVA-producing E. coli strain (O6K13) and exam-ined whether colonization of germ-free rats with thisbacterium generates tolerance.54 Based on delayed typehypersensitivity (DTH; ear thickening) measurements, theyhave found that neonatal colonization generates suppressedDTH responses compared with adult colonization, wherethere is an enhanced response. A similar finding has beenobtained with serum antibody responses against OVA, whereneonatally colonized rats generate lower antibody responsesto OVA vaccination compared with colonized adult rats,which show an increased antibody response. An interestingobservation is that colonization of adult rats with OVA-expressing E. coli generates a strong antibody responseagainst LPS, a T-independent antigen from gram negativebacteria. Further vaccination with killed bacteria fails toincrease serum antibody titres against LPS and this lack ofincrease has been interpreted as tolerance, even though thelevel of antibody responses in these animals is as high as thatgenerated in non-colonized rats that have been vaccinatedwith bacteria. Defining tolerance as a failure to furtheraugment existing antibody levels against an orally fed antigenafter further peripheral vaccination is conceptually differentfrom demonstrating a significant drop (usually > 50%) inantibody levels in animals that have been fed and then vacci-nated with the same antigen. Interpretation of results fromtolerance induction experiments becomes difficult if toler-ance is redefined in such an ad hoc fashion, even thoughessentially similar experimental protocols have been followed.
Immune response during enteric disease
A study has been carried out by Duchmann et al. to examinewhether hyporesponsiveness of lamina propria mononuclearcells (LPMC) against gut bacteria is affected by entericdisease.55 The LPMC isolated from inflamed biopsies ofintestines from patients with IBD56 proliferated after co-culture with sonicated bacteria that had been isolated and cultured from autologous intestines. Tolerized LPMC fromneighbouring non-inflamed regions of the intestine from the same patients did not proliferate in parallel coculture experiments with autologous bacteria antigen. Interestingly,
tolerized LPMC also proliferated strongly when cultured withbacteria antigens from heterologous patients. Cellular prolif-eration was accompanied by expansion of CD8+ T cells andincreased production of IL-12, IFN-γ and IL-10, but wasinhibited by the addition of anti-MHC class II monoclonalantibodies to the culture. Lymphocytes from patients with CDhave also been found to proliferate in response to microbialantigens.57 These examples have been used frequently toargue a case that tolerance against enteric microbiota isbroken down in the event of an inflammatory mucosalresponse associated with IBD and CD. Under these condi-tions, both humoral and cellular immunity can be generatedagainst harmless colonic bacteria populations.
Effects of feeding probiotic bacteria on systemicimmunity against a co-fed marker antigen
Because non-pathogenic enteric bacteria appear to possessimmunomodulatory properties, we have conducted a numberof feeding experiments in conventional mice to obtain someinsight into the effects of probiotic bacteria on co-fed proteinantigens in generating either a tolerogenic or an immuno-genic response. The selection of bacteria species was con-fined to strains that are normally used as starter cultures forthe production of cheeses and salami, such as Streptococcusthermophilus (St), Staphylococcus carnosus (Sc), and alsoincluded yeast (Y) and Lactobacillus fermentum (Lf).
The experiments followed standard feeding protocols, asdefined previously, and included the following treatments assummarized in Table 1. Treatment groups were replicated intwo sets so that LP and antibody responses could be assayedat 7 days after the last feeding (PF analysis, without vaccina-tion) and at 7 days after the booster injection (PV).
The splenocyte lymphoproliferative response againstvarious concentrations of OVA from mice after feeding, andafter feeding and vaccination, is shown in Fig. 3. Only back-ground levels of LP responses against OVA were seen in allgroups of mice after feeding. Following vaccination, there
Immune response to probiotic bacteria 61
Table 1 Treatment schedule for feeding ovalbumin with or withoutprobiotic bacteria to 6–8 week-old C57Bl mice
Treatment Amount of bacteria and/or OVA fed per dose (mg)
OVA 20Y 20Y/OVA 20 + 20St 20St/OVA 20 + 20Sc 20Sc/OVA 20 + 20Lf 20Lf/OVA 20 + 20Unfed 0
Mice were fed 3 times per week on alternate days for 3 weeks.Mice were vaccinated intradermally with immune-stimulatingcomplex-adjuvanted ovalbumin (OVA; 100 µg) at 7 days after the lastfeed. A booster injection was given 7 days later and mice were killedafter 7 days. Y, yeast; St, Streptococcus thermophilus; Sc, Staphylo-coccus carnosus; Lf, Lactobacillus fermentum.
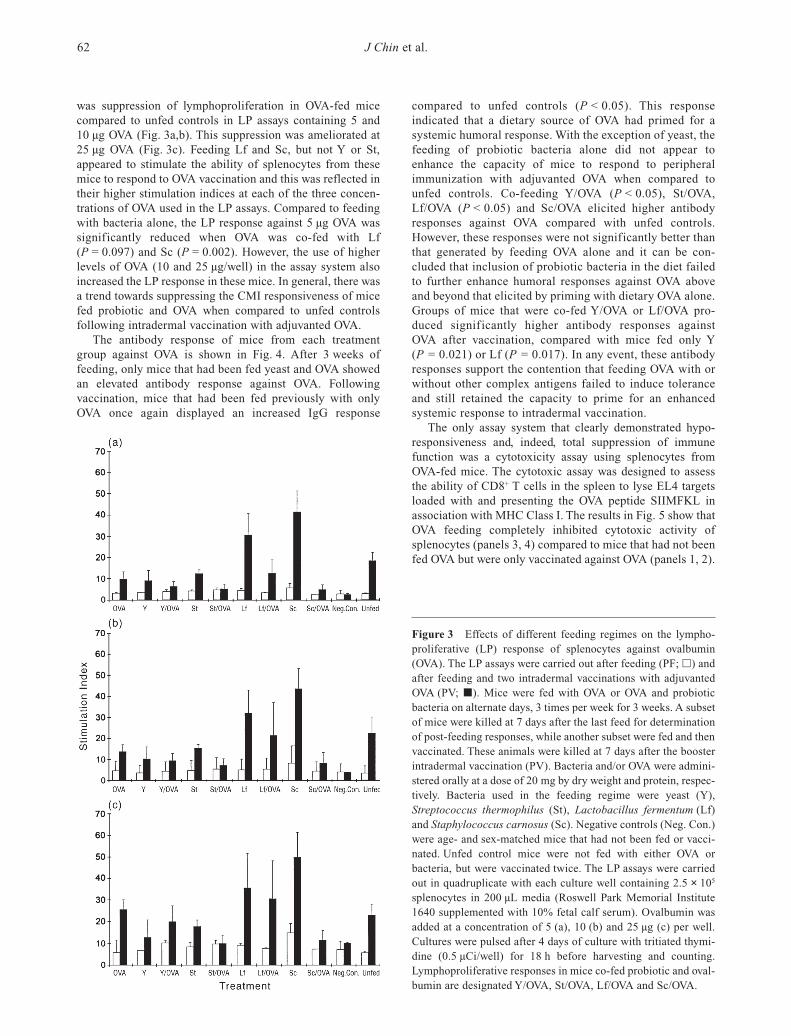
was suppression of lymphoproliferation in OVA-fed micecompared to unfed controls in LP assays containing 5 and10 µg OVA (Fig. 3a,b). This suppression was ameliorated at25 µg OVA (Fig. 3c). Feeding Lf and Sc, but not Y or St,appeared to stimulate the ability of splenocytes from thesemice to respond to OVA vaccination and this was reflected intheir higher stimulation indices at each of the three concen-trations of OVA used in the LP assays. Compared to feedingwith bacteria alone, the LP response against 5 µg OVA wassignificantly reduced when OVA was co-fed with Lf(P = 0.097) and Sc (P = 0.002). However, the use of higherlevels of OVA (10 and 25 µg/well) in the assay system alsoincreased the LP response in these mice. In general, there wasa trend towards suppressing the CMI responsiveness of micefed probiotic and OVA when compared to unfed controls following intradermal vaccination with adjuvanted OVA.
The antibody response of mice from each treatmentgroup against OVA is shown in Fig. 4. After 3 weeks offeeding, only mice that had been fed yeast and OVA showedan elevated antibody response against OVA. Following vaccination, mice that had been fed previously with onlyOVA once again displayed an increased IgG response
compared to unfed controls (P < 0.05). This response indicated that a dietary source of OVA had primed for a systemic humoral response. With the exception of yeast, thefeeding of probiotic bacteria alone did not appear toenhance the capacity of mice to respond to peripheralimmunization with adjuvanted OVA when compared tounfed controls. Co-feeding Y/OVA (P < 0.05), St/OVA,Lf/OVA (P < 0.05) and Sc/OVA elicited higher antibodyresponses against OVA compared with unfed controls.However, these responses were not significantly better thanthat generated by feeding OVA alone and it can be con-cluded that inclusion of probiotic bacteria in the diet failedto further enhance humoral responses against OVA aboveand beyond that elicited by priming with dietary OVA alone.Groups of mice that were co-fed Y/OVA or Lf/OVA pro-duced significantly higher antibody responses against OVA after vaccination, compared with mice fed only Y(P = 0.021) or Lf (P = 0.017). In any event, these antibodyresponses support the contention that feeding OVA with orwithout other complex antigens failed to induce toleranceand still retained the capacity to prime for an enhanced systemic response to intradermal vaccination.
The only assay system that clearly demonstrated hypo-responsiveness and, indeed, total suppression of immunefunction was a cytotoxicity assay using splenocytes fromOVA-fed mice. The cytotoxic assay was designed to assessthe ability of CD8+ T cells in the spleen to lyse EL4 targetsloaded with and presenting the OVA peptide SIIMFKL inassociation with MHC Class I. The results in Fig. 5 show thatOVA feeding completely inhibited cytotoxic activity ofsplenocytes (panels 3, 4) compared to mice that had not beenfed OVA but were only vaccinated against OVA (panels 1, 2).
J Chin et al.62
Figure 3 Effects of different feeding regimes on the lympho-proliferative (LP) response of splenocytes against ovalbumin(OVA). The LP assays were carried out after feeding (PF; h) andafter feeding and two intradermal vaccinations with adjuvantedOVA (PV; j). Mice were fed with OVA or OVA and probioticbacteria on alternate days, 3 times per week for 3 weeks. A subsetof mice were killed at 7 days after the last feed for determinationof post-feeding responses, while another subset were fed and thenvaccinated. These animals were killed at 7 days after the boosterintradermal vaccination (PV). Bacteria and/or OVA were admini-stered orally at a dose of 20 mg by dry weight and protein, respec-tively. Bacteria used in the feeding regime were yeast (Y),Streptococcus thermophilus (St), Lactobacillus fermentum (Lf)and Staphylococcus carnosus (Sc). Negative controls (Neg. Con.)were age- and sex-matched mice that had not been fed or vacci-nated. Unfed control mice were not fed with either OVA or bacteria, but were vaccinated twice. The LP assays were carriedout in quadruplicate with each culture well containing 2.5 × 105
splenocytes in 200 µL media (Roswell Park Memorial Institute1640 supplemented with 10% fetal calf serum). Ovalbumin wasadded at a concentration of 5 (a), 10 (b) and 25 µg (c) per well.Cultures were pulsed after 4 days of culture with tritiated thymi-dine (0.5 µCi/well) for 18 h before harvesting and counting.Lymphoproliferative responses in mice co-fed probiotic and oval-bumin are designated Y/OVA, St/OVA, Lf/OVA and Sc/OVA.
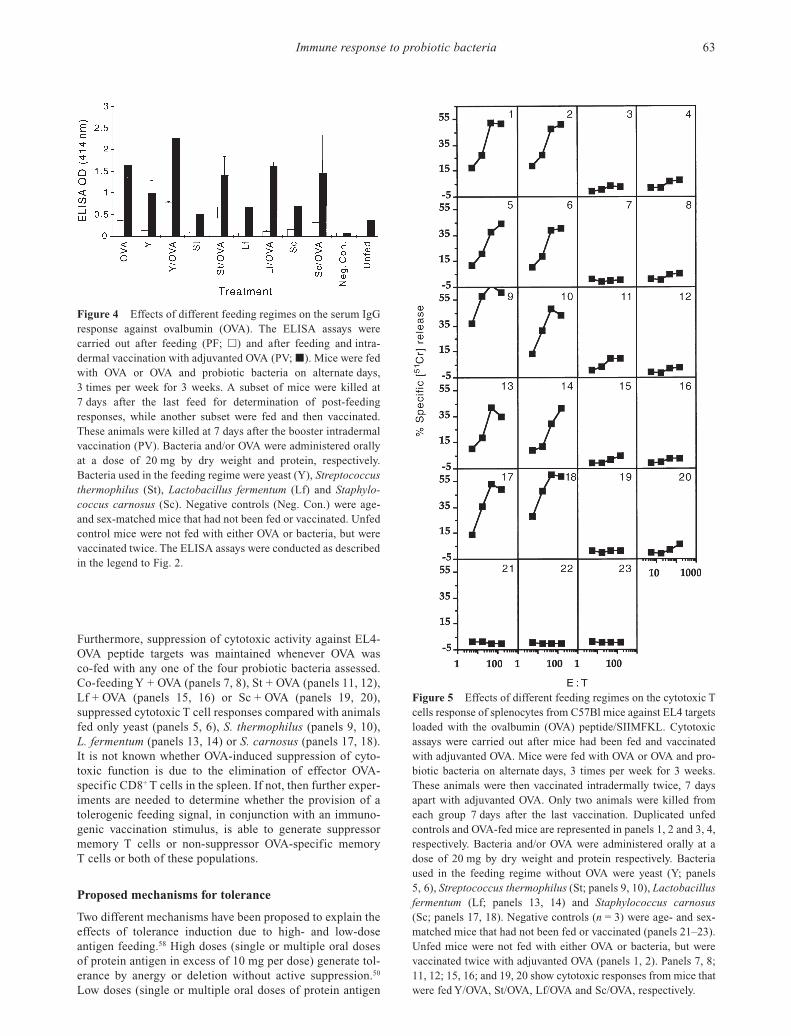
Furthermore, suppression of cytotoxic activity against EL4-OVA peptide targets was maintained whenever OVA was co-fed with any one of the four probiotic bacteria assessed.Co-feeding Y + OVA (panels 7, 8), St + OVA (panels 11, 12),Lf + OVA (panels 15, 16) or Sc + OVA (panels 19, 20), suppressed cytotoxic T cell responses compared with animalsfed only yeast (panels 5, 6), S. thermophilus (panels 9, 10), L. fermentum (panels 13, 14) or S. carnosus (panels 17, 18).It is not known whether OVA-induced suppression of cyto-toxic function is due to the elimination of effector OVA-specific CD8+ T cells in the spleen. If not, then further exper-iments are needed to determine whether the provision of atolerogenic feeding signal, in conjunction with an immuno-genic vaccination stimulus, is able to generate suppressormemory T cells or non-suppressor OVA-specific memory T cells or both of these populations.
Proposed mechanisms for tolerance
Two different mechanisms have been proposed to explain theeffects of tolerance induction due to high- and low-doseantigen feeding.58 High doses (single or multiple oral dosesof protein antigen in excess of 10 mg per dose) generate tol-erance by anergy or deletion without active suppression.50
Low doses (single or multiple oral doses of protein antigen
Immune response to probiotic bacteria 63
Figure 4 Effects of different feeding regimes on the serum IgGresponse against ovalbumin (OVA). The ELISA assays werecarried out after feeding (PF; h) and after feeding and intra-dermal vaccination with adjuvanted OVA (PV; j). Mice were fedwith OVA or OVA and probiotic bacteria on alternate days,3 times per week for 3 weeks. A subset of mice were killed at7 days after the last feed for determination of post-feedingresponses, while another subset were fed and then vaccinated.These animals were killed at 7 days after the booster intradermalvaccination (PV). Bacteria and/or OVA were administered orallyat a dose of 20 mg by dry weight and protein, respectively. Bacteria used in the feeding regime were yeast (Y), Streptococcusthermophilus (St), Lactobacillus fermentum (Lf) and Staphylo-coccus carnosus (Sc). Negative controls (Neg. Con.) were age-and sex-matched mice that had not been fed or vaccinated. Unfedcontrol mice were not fed with either OVA or bacteria, but werevaccinated twice. The ELISA assays were conducted as describedin the legend to Fig. 2.
Figure 5 Effects of different feeding regimes on the cytotoxic Tcells response of splenocytes from C57Bl mice against EL4 targetsloaded with the ovalbumin (OVA) peptide/SIIMFKL. Cytotoxicassays were carried out after mice had been fed and vaccinatedwith adjuvanted OVA. Mice were fed with OVA or OVA and pro-biotic bacteria on alternate days, 3 times per week for 3 weeks.These animals were then vaccinated intradermally twice, 7 daysapart with adjuvanted OVA. Only two animals were killed fromeach group 7 days after the last vaccination. Duplicated unfed controls and OVA-fed mice are represented in panels 1, 2 and 3, 4,respectively. Bacteria and/or OVA were administered orally at adose of 20 mg by dry weight and protein respectively. Bacteriaused in the feeding regime without OVA were yeast (Y; panels5, 6), Streptococcus thermophilus (St; panels 9, 10), Lactobacillusfermentum (Lf; panels 13, 14) and Staphylococcus carnosus(Sc; panels 17, 18). Negative controls (n = 3) were age- and sex-matched mice that had not been fed or vaccinated (panels 21–23).Unfed mice were not fed with either OVA or bacteria, but werevaccinated twice with adjuvanted OVA (panels 1, 2). Panels 7, 8;11, 12; 15, 16; and 19, 20 show cytotoxic responses from mice thatwere fed Y/OVA, St/OVA, Lf/OVA and Sc/OVA, respectively.

below 100 µg per dose) induce tolerance by the production ofcytokines, such as IL-4, IL-1059 and TGF-β.60 Elimination ofCD8+ T cells with monoclonal antibodies appears to have littleor no impact on antibody production or lymphoproliferationin high-dose tolerance induction.61 Costimulatory moleculesare believed to play a role in tolerance induction. Blockade ofB7–CTLA-4 interactions completely inhibits high-doseinduction of systemic tolerance.62 Similarly, it is not possibleto induce systemic T cell unresponsiveness to ovalbumin inCD40 ligand gene-disrupted (CD40 L–/–) mice.63 A major rolefor TGF-β1 in orally induced immune suppression was notproven when TGF-β1 knockout mice were used in toleranceinduction experiments.64 These reports demonstrate the com-plexity of immune mechanisms that operate to achieve toler-ance, even when the experimental model uses only a singlemodel antigen. The complexity of the process is compoundedwhen one takes into account the presence of commensal bac-teria and the consumption of dietary probiotics with complexmixes of antigen. It is more than likely that several of thesemechanisms probably operate in concert in vivo and theimmunological outcomes are many.65
Intestinal immunity: Discerning tolerogenic andimmunogenic signals
A number of conclusions may be drawn from toleranceinduction experimental models described in previous sec-tions:1. The provision of a tolerogenic signal by feeding high orlow doses of a single protein antigen is able to prime intesti-nal immune responses.2. The nature of the priming response is affected by thecomposition of enteric commensal bacteria and also by theprovision of dietary bacteria as probiotic supplements.3. Co-feeding ovalbumin with certain probiotics, such asyeast or S. thermophilus, can stimulate antibody responsespostfeeding.4. Depending upon the method of peripheral immunization,immunogenic signals generated by subcutaneous vaccinationwith conventional adjuvants, such as alum or completeFreund’s adjuvant, are overridden by orally induced tolero-genic signals. However, multiple intradermal vaccination,probably because of its ability to engender innate immuneresponses,8 appears to dominate over tolerogenic signals pro-vided by oral feeding of antigen. This is particularly true forhumoral responses that are Th2 dependent.5. Cellular immunity based upon in vitro lymphoprolifera-tive responses is affected by the dose of feeding antigen.Lymphoproliferative responses in vitro are more suppressedin high- than low-dose fed animals after only one intradermalvaccination. This is particularly noticeable when the concen-tration of antigen used in the LP assays is low. Increasing theconcentration of OVA in LP assays reduces the differencebetween high-dose, low-dose and unfed mice. Feeding highdoses of OVA enhances the LP response relative to unfedmice when immunogenic signals are enhanced by a secondintradermal vaccination.6. Ovalbumin-specific cytotoxic responses are completelysuppressed by feeding OVA with or without probiotic bacteriaand cannot be elicited by additional skin vaccination regimes.
Acknowledgements
The authors thank Matthew Quinnell and Susan Lemke fortheir participation in experimental feeding of mice and insample collections. MQ was involved in helping with thecytotoxicity assay. We would also like to thank Ron Tha Hlafor excellent technical assistance.
References
1 Mowat AM. The regulation of immune responses to dietaryprotein antigens. Immunol. Today 1987; 8: 93–8.
2 Weiner HL, Mackin GA, Matsui M et al. Double blind pilot trialof oral tolerization with myelin antigens in multiple sclerosis.Science 1993; 261: 1727–9.
3 Zhang JZ, Lee CSY, Lider O, Weiner HL. Suppression of adjuvant arthritis in Lewis rats by oral administration of type II collagen. J. Immunol. 1990; 145: 2489–93.
4 Melamed D, Friedman A. Modification of the immune responseby oral tolerance: Antigen requirements and interaction withimmunogenic stimuli. Cell. Immunol. 1993; 146: 412–20.
5 Husby S, Jensenius JC, Svehag SE. Passage of undegradeddietary antigen into the blood of healthy adults. Quantification,estimation of size distribution and relation of uptake to levels ofspecific antibodies. Scand. J. Immunol. 1985; 22: 83–92.
6 Hanson DG. Ontogeny of orally induced tolerance to solubleproteins in mice. 1. Priming and tolerance in newborns. J.Immunol. 1981; 127: 1518–24.
7 Bailey M, Miller BG, Telemo E, Stokes CR, Bourne FM. Spe-cific immunological unresponsiveness following active primaryresponses to proteins in the weaning diet of piglets. Int. Arch.Allergy Immunol. 1993; 101: 266–71.
8 Chin JC, San Gil F. Skin delivery of a hybrid liposome/ISCOMvaccine implicates a role for adjuvants in rapid modulation ofinflammatory cells involved in innate immunity before theenhancement of adaptive immune responses. Immunol. CellBiol. 1998; 76: 245–55.
9 Chin JC, San-Gil F, Novak M et al. Manipulating systemic andmucosal immune responses with skin-deliverable adjuvants. J.Biotech. 1996; 44: 13–19.
10 Erna JG, Wilsem V, Breve J et al. Oral tolerance is determinedat the level of draining lymph nodes. Immunobiology 1995; 194:403–14.
11 Taams LS, Van Eden W, Wauben MHM. Dose-dependent induc-tion of distinct anergic phenotypes: Multiple levels of T cellanergy. J. Immunol. 1999; 162: 1974–81.
12 Zinkernagel RM, Kelly J. How antigen influences immunity.Immunologist 1997; 5/4: 114–20.
13 Mowat AM, Viney JL. The anatomical basis of intestinal immu-nity. Immunol. Rev. 1997; 156: 145–66.
14 Bry L, Falk P, Huttner K, Ouellette A, Midtvedt T, Gordon JI.Paneth Cell differentiation in the developing intestine of normaland transgenic mice. Proc. Natl Acad. Sci. USA 1994; 91:10 335–9.
15 Liu LM, MacPherson GG. Antigen acquisition by dendriticcells: Intestinal dendritic cells acquire antigen administeredorally and can prime naive T cells in vivo. J. Exp. Med. 1993;177: 1299–306.
16 Richman L, Graeff AS, Strober W. Antigen presentation bymacrophage-enriched cells from the mouse Peyer’s patch. Cell.Immunol. 1981; 62: 110–18.
17 Ngan J, Kind LS. Suppressor T cells for IgE and IgG in Peyer’sPatches of mice made tolerant by the oral administration of ovalbumin. J. Immunol. 1978; 120: 861–5.
J Chin et al.64

18 Schubert R, Renz D, Schmitz B, Doerfler W. Foreign (M13) DNAingested by mice reaches peripheral leukocytes, spleen and livervia the intestinal wall mucosa and can be covalently linked tomouse DNA. Proc. Natl Acad. Sci. USA 1997; 94: 961–7.
19 Shimoda M, Inoue Y, Ametani A et al. Anti-DNA IgA autoanti-bodies are spontaneously generated in mouse Peyer’s patches.Immunology 1998; 95: 200–7.
20 Guy-Grand D, Cuenod-Jabri B, Malassis-Seris M, Selz F, Vassalli P. Complexity of the mouse gut T cell immune system:identification of two distinct natural killer T cell intraepitheliallineages. Eur. J. Immunol. 1996; 26: 2248–56.
21 Guy-Grand D, Cerf-Bensussan N, Malissen B, Malassis-SerisM, Briottet C, Vassalli P. Two gut intraepithelial CD8+ lympho-cyte populations with different T cell receptors: a role for the gutepithelium in T cell differentiation. J. Exp. Med. 1991; 173:471–6.
22 Fujihashi K, Taguchi R, Aicher WK et al. Immunoregulatoryfunction for murine intraepithelial lymphocytes: γδ T cells abrogate oral tolerance while αβ TCR+ T cells provide B cellhelp. J. Exp. Med. 1992; 175: 695–707.
23 Yamamoto M, Fujihashi K, Beagley KW, McGhee JR, KiyonoH. Cytokine synthesis by intestinal intraepithelial lymphocytes.J. Immunol. 1993; 150: 106–14.
24 Schieferdecker HL, Ullrich R, Hirseland H, Zeitz M. T cell differentiation antigens on lymphocytes in the human andintestinal lamina propria. J. Immunol. 1992; 149: 2816–22.
25 Breese E, Braegger CP, Corrigan CJ, Walker-Smith JA, Mac-Donald TT. Interleukin 2 and interferon-g secreting T cells innormal and diseased human intestinal mucosa. Immunology1993; 78: 127–31.
26 Steinbrink K, Matthias W, Jonuleit H, Knop J, Enk AH. Induc-tion of tolerance by IL-10 treated dendritic cells. J. Immunol.1997; 159: 4772–80.
27 Harper H, Cochrane L, Williams NA. The role of small intesti-nal antigen-presenting cells in the induction of T-cell reactivityto soluble protein antigens: association between aberrant pre-sentation in the lamina propria and oral tolerance. Immunology1996; 89: 449–53.
28 Liu LM, MacPherson GG. Rat intestinal dendritic cells:Immunostimulatory potency and phenotypic characterization.Immunology 1995; 85: 88–93.
29 Viney JL, Mowat AM, O’Malley JM, Williamson E, Fanger NA.Expanding dendritic cells in vivo enhances the induction of oraltolerance. J. Immunol. 1998; 160: 5815–25.
30 Cooperstock MS, Jedd AJ. Intestinal flora in infants. In: HentgesDJ (ed.). Human intestinal microflora in health and disease.New York: Academic Press, 1983; 79–99.
31 Berg RD. Mechanisms confining indigenous bacteria to the gastrointestinal tract. Am. J. Clin. Nutr. 1980; 33: 2472–84.
32 Bry L, Falk PG, Huttner K, Midtvedt T, Gordon JI. A model ofhost–microbial interactions in an open mammalian ecosystem.Science 1996; 273: 1380–3.
33 Gordon JI, Hooper LV, Bry L, Midtvedt T, Falk PG. Interactionsbetween epithelial cells and bacteria, normal and pathogenic.Science 1997; 276: 965.
34 Umesaki Y, Okada Y, Imaoka A, Setoyama H, Matsumoto S.Interactions between epithelial cells and bacteria, normal andpathogenic. Science 1997; 276: 964.
35 Umesaki Y, Okada Y, Matsumoto S, Imaoka A, Setoyama H.Segmented filamentous bacteria are indigenous intestinal bacte-ria that activate intraepithelial lymphocytes and induce MHCClass II molecules and fucosyl asialo GM1 glycolipids on thesmall intestinal epithelial cells in the ex-germ-free mouse.Microbiol. Immunol. 1995; 39: 555–62.
36 McCartney AL, Wang WZ, Tannock GW. Molecular analysis of the composition of the bifidobacterial and lactobacillus micro-flora of humans. Appl. Environ. Microbiol. 1996; 62: 4608–13.
37 Takeuchi A. Electron microscope studies of experimental Sal-monella infection. Am. J. Pathol. 1974; 50: 109–19.
38 McCormick BA, Miller SI, Carnes D, Madara JL. Transepithe-lial signaling to neutrophils by salmonellae: A novel virulencemechanism for gastroenteritis. Infect. Immun. 1996; 63: 2302–9.
39 Mitani A, Nishimura H, Hirose K et al. Interleukin-15 produc-tion at the early stage after oral infection with Listeria mono-cytogenes in mice. Immunology 1999; 97: 92–9.
40 Kaila M, Isolauri E, Soppi E, Virtanen E, Laine S, Arvilommi H.Enhancement of the circulating antibody secreting cell responsein human diarrhea by a human Lactobacillus strain. Pediatr. Res.1992; 32: 141–4.
41 Del Aguila LF, Claffey KP, Kirwan JP. TNF-alpha impairsinsulin signaling and insulin stimulation of glucose uptake inC2C12 muscle cells. Am. J. Physiol. 1999; 39: E849–55.
42 Olff M. Stress, Depression and immunity: The role of defenseand coping styles. Psychiatry Res. 1999; 85: 7–15.
43 Rescigno M, Citterio S, Thery C et al. Bacteria-induced neo-biosynthesis, stabilization, and surface expression of functionalclass I molecules in mouse dendritic cells. Proc. Natl Acad. Sci.USA 1998; 95: 5229–34.
44 Knigge H, Simon M, Meuer SC, Kramer MD, Wallich R. Theouter surface lipoprotein OspA of Borrelia burgdorferi providesco-stimulatory signals to normal human peripheral CD4+ andCD8+ T lymphocytes. Eur. J. Immunol. 1996; 26: 2299–303.
45 Millan C, Weeratna R, Krieg AM, Siegrist CA, Davis HL. CpGDNA can induce strong Th1 humoral and cell-mediated immuneresponses against hepatitis B surface antigen in young mice.Proc. Natl Acad. Sci. USA 1998; 95: 15 553–8.
46 Prigozy TI, Sieling PA, Clemens D et al. The mannose receptordelivers lipoglycan antigens to endosomes for presentation to Tcells by CD1b molecules. Immunity 1997; 6: 187–97.
47 Wold AE, Dahlgren UIH, Hanson LS, Mattsby-Baltzer I, Mid-vetdt T. Difference between bacterial and food antigens inmucosal immunogenicity. Infect. Immun. 1989; 57: 2666–73.
48 Kaneko T, Terasawa Y, Senou S, Nagata M, Kuwata T. Oral tolerance is not influenced by oral application of oil-emulsifiedproteins. Int. Arch. Allergy Immunol. 1997; 116: 81–2.
49 Ito K, Inagaki-Ohara K, Murosaki S et al. Murine model of IgEproduction with a predominant Th2-response by feeding proteinantigen without adjuvants. Eur. J. Immunol. 1997; 27: 3427–37.
50 Chen Y, Inobe R, Marks P, Gonnella VK, Weiner HL. Peripheraldeletion of antigen-reactive T cells in oral tolerance. Nature1995; 376: 177–9.
51 Vidal S, Malo D, Vogan K, Skamens E, Gros P. Natural resis-tance to infection with intracellular parasites: Isolation of a candidate for Bcg. Cell 1993; 73: 469–85.
52 Moreau CM, Corthier G. Effect of gastrointestinal microflora oninduction and maintenance of oral tolerance to ovalbumin inC3H/HeJ mice. 1988; 56: 2766–8.
53 Gaboriau-Routhiau V, Moreau MC. Gut flora allows recovery oforal tolerance to ovalbumin in mice after transient breakdownmediated by cholera toxin or Escherichia coli heat-labile entero-toxin. Pediatr. Res. 1996; 39: 625–9.
54 Karlsson MR, Kahu H, Hanson LA, Telemo E, Dahlgren U.Neonatal colonization of rats induces immunological toleranceto bacterial antigens. Eur. J. Immunol. 1999; 29: 109–18.
55 Duchmann R, Kaiser I, Hermann E, Mayet W, Ewe K, Meyerzum Buschenfelde KH. Tolerance exists towards resident intesti-nal flora but is broken in active inflammatory bowel disease(IBD). Clin. Exp. Immunol. 1995; 102: 448–55.
Immune response to probiotic bacteria 65

56 MacDonald TT. Breakdown of tolerance to the intestinal bacterial flora in inflammatory bowel disease (IBD). Clin. Exp.Immunol. 1995; 102: 445–7.
57 Pirzer U, Schasanhaar A, Fleischer B, Hermann E, Meyer zumBuschenfelde KH. Reactivity of infiltrating T lymphocytes withmicrobial antigens in Crohn’s disease. Lancet 1991; 338: 1238–9.
58 Friedman A, Weiner HL. Induction of anergy or active suppres-sion following oral tolerance is determined by antigen dosage.Proc. Natl Acad. Sci. USA 1994; 91: 6688–92.
59 Garofalo R, Chheda S, Mei F et al. Interleukin 10 in humanmilk. Pediatr. Res. 1995; 37: 444–8.
60 Caspi RR, Stiff LR, Morawetz R et al. Cytokine dependent modulation of oral tolerance in murine model of autoimmuneuveitis. Ann. N. Y. Acad. Sci. 1996; 778: 315–26.
61 Barone KS, Jain SL, Michael JG. Effect of in vivo depletion ofCD4+ and CD8+ cells on the induction and maintenance of oraltolerance. Cell 1995; 163: 19–24.
62 Samilova EB, Horton JL, Zhang H, Khoury SJ, Weiner HL,Chen Y. CTLA-4 is required for the induction of high dose oraltolerance. Intern. Immunol. 1998; 10: 491–503.
63 Kweon M, Fujihashi K, Wakatsuki Y et al. Mucosally inducedsystemic T cell unresponsiveness to ovalbumin requires CD40ligand–CD40 interactions. J. Immunol. 1999; 162: 1904–9.
64 Barone KS, Tolarova DD, Ormsby I, Doetschman T, Michael JG.Induction of oral tolerance in TGFβ1 null mice. J. Immunol.1998; 161: 154–60.
65 Claessen AME, Von Blomberg BME, De Groot J, Wolvers DAE,Draal G, Scheper RJ. Reversal of mucosal tolerance by subcuta-neous administration of interleukin-12 at the site of attemptedsensitization. Immunology 1996; 88: 363–7.
J Chin et al.66
Related Documents