The Journal of Immunology Immune Modulation by Zoledronic Acid in Human Myeloma: An Advantageous Cross-Talk between Vg9Vd2 T Cells, ab CD8 + T Cells, Regulatory T Cells, and Dendritic Cells Barbara Castella,* Chiara Riganti, † Francesca Fiore,* ,‡ Francesca Pantaleoni,* Maria Elisa Canepari,* Silvia Peola,* Myriam Foglietta,* ,‡ Antonio Palumbo, ‡ Amalia Bosia, † Marta Coscia,* ,‡ Mario Boccadoro, ‡ and Massimo Massaia* ,‡ Vg9Vd2 T cells play a major role as effector cells of innate immune responses against microbes, stressed cells, and tumor cells. They constitute ,5% of PBLs but can be expanded by zoledronic acid (ZA)-treated monocytes or dendritic cells (DC). Much less is known about their ability to act as cellular adjuvants bridging innate and adaptive immunity, especially in patients with cancer. We have addressed this issue in multiple myeloma (MM), a prototypic disease with several immune dysfunctions that also affect gd T cells and DC. ZA-treated MM DC were highly effective in activating autologous gd T cells, even in patients refractory to stimulation with ZA-treated monocytes. ZA inhibited the mevalonate pathway of MM DC and induced the intracellular accu- mulation and release into the supernatant of isopentenyl pyrophosphate, a selective gd T cell activator, in sufficient amounts to induce the proliferation of gd T cells. Immune responses against the tumor-associated Ag survivin (SRV) by MHC-restricted, SRV-specific CD8 + ab T cells were amplified by the concurrent activation of gd T cells driven by autologous DC copulsed with ZA and SRV-derived peptides. Ancillary to the isopentenyl pyrophosphate-induced gd T cell proliferation was the mevalonate- independent ZA ability to directly antagonize regulatory T cells and downregulate PD-L2 expression on the DC cell surface. In conclusion, ZA has multiple immune modulatory activities that allow MM DC to effectively handle the concurrent activation of gd T cells and MHC-restricted CD8 + ab antitumor effector T cells. The Journal of Immunology, 2011, 187: 1578–1590. A key role is played by gd T cells in innate immune responses against microbes, stressed cells, and tumor cells (1). Most circulating gd T cells use the same TCR V region pair Vg9-Vd2 (Vg9Vd2 T cells, hereinafter referred to as gd T cells) to sense the presence of infectious agents via the re- cognition of intermediate metabolites produced by the microbial nonmevalonate (non-Mev) pathway of isoprenoid synthesis (col- lectively termed phosphoantigens) (2, 3). Synthetic phosphoanti- gen analogs, such as bromohydrin pyrophosphate (BrHPP), have been generated to selectively stimulate gd T cells and clinically exploit their effector functions against pathogens and tumor cells (4, 5). The Mev pathway of mammalian cells also generates in- termediate phosphorylated metabolites, such as isopentenyl py- rophosphate (IPP), which activate gd T cells as efficiently as their natural ligands (6, 7). Cell stress and transformation increase Mev activity and accelerate the formation of intracellular phosphory- lated metabolites, whose accumulation in excess of physiological levels is detected by gd T cells and renders the immune system able to sense potential threats such as stressed or transformed cells (6). IPP levels are increased by drugs such as aminobisphos- phonates (NBP) that selectively target farnesyl-pyrophosphate synthase (FPPS) in the Mev pathway (6–8). When NBP are used to target the Mev pathway of professional APC, such as monocytes (MC) and dendritic cells (DC), a quick and robust proliferative expansion of gd T cells with antitumor activity is induced (9–11). gd T cells also act as cellular adjuvants bridging innate and adaptive immunity via mutual interactions with DC (12, 13). We have shown in healthy individuals that MHC-restricted antiviral immune responses are improved by the concurrent activation of CD8 + ab T cells and gd T cells driven by autologous DC copulsed with zoledronic acid (ZA) and influenza virus-derived peptides (11). Similar data have been reported in healthy individuals after copulsing DC with ZA and melanoma-derived peptides (14). Multiple myeloma (MM) is a disease with several immune dysfunctions that do not spare gd T cells and DC (9, 15–19). Approximately 50% of MM patients at diagnosis have gd T cells refractory to stimulation with BrHPP or NBP mediated by MC (9, 15–17). These patients are categorized as nonresponders (NR) and regarded as ineligible for gd T cell-based immune interven- tions (16). Cabillic et al. (20) have recently shown that gd T cell *Laboratorio di Ematologia Oncologica, Centro di Ricerca in Medicina Sperimen- tale, 10126 Torino, Italy; † Dipartimento di Genetica, Biologia e Biochimica, Uni- versita ´ degli Studi di Torino, 10126 Torino, Italy; and ‡ Divisione Universitaria di Ematologia, Ospedale San Giovanni Battista di Torino, Universita ` degli Studi di Torino, 10126 Torino, Italy Received for publication August 4, 2010. Accepted for publication June 3, 2011. This work was supported by Regione Piemonte (Ricerca Sanitaria, Ricerca Scientif- ica e Progetto Strategico ImmOnc), Ministero dell’Istruzione, dell’Universita ´ e della Ricerca (Roma, Italy), Compagnia San Paolo di Torino (Torino, Italy), Fondazione Neoplasie Sangue Onlus (Torino, Italy), Associazione per lo Studio e la Cura delle Malattie del Sangue (Torino, Italy), Fondazione Cassa di Risparmio di Torino (Pro- getto Alfieri, Torino, Italy), and Novartis Farma S.p.A. (Origgio, Italy). Address correspondence and reprint requests to Prof. Massimo Massaia, Divisione Universitaria di Ematologia, Ospedale San Giovanni Battista, Via Genova 3, 10126 Torino, Italy. E-mail address: [email protected] The online version of this article contains supplemental material. Abbreviations used in this article: BrHPP, bromohydrin pyrophosphate; CM, central memory; CMVp, CMV-derived peptide NLVPMVATV; CTRL, control; DC, dendritic cell; EM, effector memory; f-DC, fast DC; FPP, farnesyl-pyrophosphate; FPPS, FPP synthase; HIVp, HIV-derived peptide SLYNTVATL; iDC, immature DC; IPP, iso- pentenyl pyrophosphate; MC, monocyte; mDC, mature DC; Mev, mevalonate; MM, multiple myeloma; NBP, aminobisphosphonate; NR, nonresponder; PCL, plasma cell leukemia; R, responder; Sim, simvastatin; SRV, survivin; SRVp, SRV-derivedpeptide LMLGEFLKL; Treg, regulatory T cell; ZA, zoledronic acid. Copyright Ó 2011 by The American Association of Immunologists, Inc. 0022-1767/11/$16.00 www.jimmunol.org/cgi/doi/10.4049/jimmunol.1002514

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The Journal of Immunology
Immune Modulation by Zoledronic Acid in Human Myeloma:An Advantageous Cross-Talk between Vg9Vd2 T Cells, abCD8+ T Cells, Regulatory T Cells, and Dendritic Cells
Barbara Castella,* Chiara Riganti,† Francesca Fiore,*,‡ Francesca Pantaleoni,*
Maria Elisa Canepari,* Silvia Peola,* Myriam Foglietta,*,‡ Antonio Palumbo,‡
Amalia Bosia,† Marta Coscia,*,‡ Mario Boccadoro,‡ and Massimo Massaia*,‡
Vg9Vd2 T cells play a major role as effector cells of innate immune responses against microbes, stressed cells, and tumor cells.
They constitute ,5% of PBLs but can be expanded by zoledronic acid (ZA)-treated monocytes or dendritic cells (DC). Much less
is known about their ability to act as cellular adjuvants bridging innate and adaptive immunity, especially in patients with cancer.
We have addressed this issue in multiple myeloma (MM), a prototypic disease with several immune dysfunctions that also affect
gd T cells and DC. ZA-treated MM DC were highly effective in activating autologous gd T cells, even in patients refractory to
stimulation with ZA-treated monocytes. ZA inhibited the mevalonate pathway of MM DC and induced the intracellular accu-
mulation and release into the supernatant of isopentenyl pyrophosphate, a selective gd T cell activator, in sufficient amounts to
induce the proliferation of gd T cells. Immune responses against the tumor-associated Ag survivin (SRV) by MHC-restricted,
SRV-specific CD8+ ab T cells were amplified by the concurrent activation of gd T cells driven by autologous DC copulsed with ZA
and SRV-derived peptides. Ancillary to the isopentenyl pyrophosphate-induced gd T cell proliferation was the mevalonate-
independent ZA ability to directly antagonize regulatory T cells and downregulate PD-L2 expression on the DC cell surface.
In conclusion, ZA has multiple immune modulatory activities that allow MM DC to effectively handle the concurrent activation of
gd T cells and MHC-restricted CD8+ ab antitumor effector T cells. The Journal of Immunology, 2011, 187: 1578–1590.
Akey role is played by gd T cells in innate immuneresponses against microbes, stressed cells, and tumorcells (1). Most circulating gd T cells use the same TCR
V region pair Vg9-Vd2 (Vg9Vd2 T cells, hereinafter referred to asgd T cells) to sense the presence of infectious agents via the re-cognition of intermediate metabolites produced by the microbialnonmevalonate (non-Mev) pathway of isoprenoid synthesis (col-lectively termed phosphoantigens) (2, 3). Synthetic phosphoanti-gen analogs, such as bromohydrin pyrophosphate (BrHPP), have
been generated to selectively stimulate gd T cells and clinicallyexploit their effector functions against pathogens and tumor cells(4, 5). The Mev pathway of mammalian cells also generates in-termediate phosphorylated metabolites, such as isopentenyl py-rophosphate (IPP), which activate gd T cells as efficiently as theirnatural ligands (6, 7). Cell stress and transformation increase Mevactivity and accelerate the formation of intracellular phosphory-lated metabolites, whose accumulation in excess of physiologicallevels is detected by gd T cells and renders the immune systemable to sense potential threats such as stressed or transformed cells(6). IPP levels are increased by drugs such as aminobisphos-phonates (NBP) that selectively target farnesyl-pyrophosphatesynthase (FPPS) in the Mev pathway (6–8). When NBP areused to target the Mev pathway of professional APC, such asmonocytes (MC) and dendritic cells (DC), a quick and robustproliferative expansion of gd T cells with antitumor activity isinduced (9–11).gd T cells also act as cellular adjuvants bridging innate and
adaptive immunity via mutual interactions with DC (12, 13). Wehave shown in healthy individuals that MHC-restricted antiviralimmune responses are improved by the concurrent activation ofCD8+ ab T cells and gd T cells driven by autologous DC copulsedwith zoledronic acid (ZA) and influenza virus-derived peptides(11). Similar data have been reported in healthy individuals aftercopulsing DC with ZA and melanoma-derived peptides (14).Multiple myeloma (MM) is a disease with several immune
dysfunctions that do not spare gd T cells and DC (9, 15–19).Approximately 50% of MM patients at diagnosis have gd T cellsrefractory to stimulation with BrHPP or NBP mediated by MC(9, 15–17). These patients are categorized as nonresponders (NR)and regarded as ineligible for gd T cell-based immune interven-tions (16). Cabillic et al. (20) have recently shown that gd T cell
*Laboratorio di Ematologia Oncologica, Centro di Ricerca in Medicina Sperimen-tale, 10126 Torino, Italy; †Dipartimento di Genetica, Biologia e Biochimica, Uni-versita degli Studi di Torino, 10126 Torino, Italy; and ‡Divisione Universitaria diEmatologia, Ospedale San Giovanni Battista di Torino, Universita degli Studi diTorino, 10126 Torino, Italy
Received for publication August 4, 2010. Accepted for publication June 3, 2011.
This work was supported by Regione Piemonte (Ricerca Sanitaria, Ricerca Scientif-ica e Progetto Strategico ImmOnc), Ministero dell’Istruzione, dell’Universita e dellaRicerca (Roma, Italy), Compagnia San Paolo di Torino (Torino, Italy), FondazioneNeoplasie Sangue Onlus (Torino, Italy), Associazione per lo Studio e la Cura delleMalattie del Sangue (Torino, Italy), Fondazione Cassa di Risparmio di Torino (Pro-getto Alfieri, Torino, Italy), and Novartis Farma S.p.A. (Origgio, Italy).
Address correspondence and reprint requests to Prof. Massimo Massaia, DivisioneUniversitaria di Ematologia, Ospedale San Giovanni Battista, Via Genova 3, 10126Torino, Italy. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: BrHPP, bromohydrin pyrophosphate; CM, centralmemory; CMVp, CMV-derived peptide NLVPMVATV; CTRL, control; DC, dendriticcell; EM, effector memory; f-DC, fast DC; FPP, farnesyl-pyrophosphate; FPPS, FPPsynthase; HIVp, HIV-derived peptide SLYNTVATL; iDC, immature DC; IPP, iso-pentenyl pyrophosphate; MC, monocyte; mDC, mature DC; Mev, mevalonate; MM,multiple myeloma; NBP, aminobisphosphonate; NR, nonresponder; PCL, plasma cellleukemia; R, responder; Sim, simvastatin; SRV, survivin; SRVp, SRV-derived peptideLMLGEFLKL; Treg, regulatory T cell; ZA, zoledronic acid.
Copyright� 2011 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/11/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1002514

proliferation can be reinstated in NR patients with solid tumorsusing ZA-treated DC. However, MM DC are inclined to stimulateT cells with inhibitory functions (21, 22), and whether they canreinstate gd T cell proliferation in NR patients, and effectivelydrive the concurrent activation of Ag-specific ab T cells and gdT cells, is an unanswered question not only in MM, but also inpatients with cancer.The aim of this work was: 1) to determine whether MM DC are
as effective as normal DC in activating autologous gd T cells inboth responders (R) and NR, and 2) to determine whether immuneresponses against tumor-associated Ags in MM can be amplifiedby the mutual interactions between gd T cells and MHC-restrictedCD8+ ab T cells driven by autologous DC copulsed with ZA andsurvivin (SRV). SRV was selected as a prototypic tumor-associated Ag on account of its aberrant expression in MM andbecause SRV-specific cytotoxic CD8+ ab T cells are tolerized butnot deleted from the MM T cell repertoire (23).
Materials and MethodsCells and reagents
Peripheral blood samples were drawn fromMM at diagnosis only. Differentseries of experiments were performed on a total number of 22 patients. Thecontrols (CTRL) were healthy blood donors kindly provided by the localBlood Bank (Fondazione Strumia). Samples were collected after informedconsent and approval by the local Institutional Review Boards.
After isolation on a Ficoll-Hypaque density gradient, PBMC were keptunfractionated or further processed to purify CD14+ cells and generatedifferent APC preparations. Aliquots of PBMC and CD14-depleted cells(i.e., PBL) were kept at 37˚C or frozen in liquid nitrogen to be sub-sequently used without further manipulation or as a source of T cells incoculture experiments with autologous APC.
The standard culture medium was RPMI 1640 (Euroclone, Milano,Italy), containing 10% FCS (Euroclone), 2 mM L-glutamine, 100 U/mlpenicillin, and 100 mg/ml streptomycin.
Flow cytometry
The following mAbs were used: anti-CD3, anti-CD8, anti-CD4, anti-CD25,anti-CD54 (Caltag Laboratories, Burlingame, CA); anti–pan-TCR gd, anti–TCR-Vg9, anti–TCR-ab, anti-CD56, anti-CD80, anti–HLA-DR (BectonDickinson, San Jose, CA); anti-CD3, anti-CD14 (Dako SpA, Milano,Italy); anti-CD83 (BD Pharmingen International, San Diego, CA), anti-CD86 (Chemicon, Chandlers Ford, Hampshire, U.K.), anti–HLA-A2(Proimmune, Oxford, U.K.); anti-CD1a, CD45RA (Valter Occhiena, Tor-ino, Italy), anti-CD27 (Ancell, Byport, MN); anti-FOXP3, anti-CD279(PD-1), anti–B7-H1 (PD-L1), and anti–B7-DC (PD-L2; eBioscience, SanDiego, CA).
Two- and three-color flow cytometry were performed with the appro-priate combinations of FITC, r-PE– (PE), Tricolor- (Tri), PerCP complex-(PerCP), or allophycocyanin-conjugated Abs, a FACScan cell sorter, andCellQuest software (Becton Dickinson, Mountain View, CA).
Regulatory T cell (Treg) cells were identified by incubation with PerCP-conjugated anti-CD4 and PE-conjugated CD25 at 4˚C for 20 min in thedark. After washing, they were fixed and permeabilized at 4˚C for 40–60min (according to the manufacturer’s instructions), washed twice withpermeabilization buffer, and then incubated at 4˚C in the dark for 30 minwith 20 ml anti-Foxp3 FITC or isotype CTRL mAb. After washes withPerm/Wash buffer, cells were fixed with 250 ml 1% paraformaldehyde-PBSbuffer.
Total counts of specific cell subsets per well were determined by mul-tiplying total counts of viable cells per well by the percentage of cells ofinterest, the latter being identified by two- or three-color flow cytometry andappropriate gating.
APC generation
MC were purified using CD14 MicroBeads and LS columns (MiltenyiBiotec, Bologna, Italy), and incubated for 24 h in 24-well plates at 13 106/ml in the presence (MCZA+) or absence of 5 mM ZA (MCZA2; kindlyprovided by Novartis Farma, Origgio, Italy).
Conventional immature DC (iDC) andmature DC (mDC) were generatedfrom CD14+ cells and incubated for 24 h in the presence (iDCZA+, mDCZA+)or absence of 5 mM ZA (iDCZA2, mDCZA2), as previously reported (11,22).
Fast DC (f-DC) were generated according to Dauer et al. (24). PurifiedCD14+ cells were cultured in standard culture medium at 0.5–1.5 3 106
cells/ml supplemented with GM-CSF (1000 U/ml) and IL-4 (500 U/ml) inflat-bottom 6-well plates for 24 h. After washing, they were incubated fora further 24 h with IL-1b (10 ng/ml), TNF-a (1000 IU/ml), IL-6 (10 ng/ml), and PGE2 (1 mg/ml) in the presence (f-DCZA+) or absence (f-DCZA2)of 5 mM ZA.
APC were always washed before mixing with autologous PBL or T cells.In selected experiments, supernatants were collected for quantification ofextracellular IPP levels and to investigate their ability to induce autologousgd T cell proliferation.
The internalization capability of iDC, mDC, and f-DC was tested byusing FITC-conjugated dextran (FITC-dextran; 10 kDa; Sigma Aldrich,Milano, Italy).
gd T cell proliferation
CTRL and MM patients were classed as R or NR according to their pro-liferative PBMC responses to 1 mMZA + 10 IU/ml IL-2 (Eurocetus, Milan,Italy), as previously reported (9).
Cryopreserved or freshly isolated PBL were added at the ratio of 5:1 toautologous iDC/mDC or MC/f-DC, respectively, and cultured at 1 3 106/ml with 10 IU/ml IL-2. On day 7, percentages and total counts of viable gdT cells and Tregs were calculated with the trypan blue staining assay andby flow cytometry, as previously reported (11, 25).
In selected experiments, PBMC and PBL from CTRL R were stimulatedfor 7 d with the supernatants obtained from autologous f-DCZA2 and f-DCZA+ and 10 IU/ml IL-2 to evaluate their ability to induce gd T cellproliferation.
Modulation of SRV-specific immune responses
The HLA-A*0201+–restricted, SRV-derived peptide LMLGEFLKL(SRVp; Proimmune) was used to investigate MHC-restricted immuneresponses mediated by CD8+ ab T cells. iDC were generated from HLA-A*0201+ CTRL and MM, and left untreated or pulsed with 10 mM SRVpfor 2 h at room temperature in serum-free medium. After washing, iDCmaturation was induced in the presence or absence of 5 mM ZA as reportedearlier. On the same day, CD3+ cells were purified from cryopreservedautologous PBMC (Miltenyi Pan T Cell Isolation Kit), mixed at the DC:Tratio of 1:10 in round-bottom plates, and incubated for 10 d in the presenceof 10 IU/ml IL-2, replenished every 3 d. On day 10, T cells were re-stimulated for additional 10 d with a second batch of freshly generatedmDC. On days 10 and 20, the frequency of SRV-specific CD8+ ab T cellswas determined by flow cytometry with a commercially available PE-labeled kit (Pro5 MHC Pentamer; Proimmune), according to the manu-facturer’s instructions. Because Pro5 Pentamers can bind nonspecifically toB cells, both CD8 and CD19 mAbs were added after pentamer staining,and a minimum number of 500,000 events was acquired after backgatingon viable CD8+ CD192 cells.
f-DC were also left untreated or pulsed with SRVp, and incubated with(f-DCZA+) or without 5 mM ZA (f-DCZA2) as reported earlier for mDC.CD3+ cells were purified from cryopreserved autologous PBMC and in-cubated for 14 d with f-DC at the DC:T ratio of 1:10 in round-bottomplates in the presence of 10 IU/ml IL-2 that was replenished every 3 d. Oneround of 14-d stimulation was used based on side-by-side experimentsshowing that it was equivalent to two rounds of 10-d stimulation (see laterand Supplemental Fig. 4).
The cytotoxic activity of CD8+ ab T cells was tested against the TAP-deficient, SRVp-loaded HLA-A2+ T2 cell line, the HLA-A2+ SRV+ mye-loma cell line RPMI 8226, and HLA-A2+ myeloma cells isolated from theperipheral blood of a plasma cell leukemia (PCL) patient, as previouslyreported (11).
The specificity of anti-SRV immune responses was assessed by using theHLA-A2–restricted CMV- (NLVPMVATV; Proimmune) and HIV-derived(SLYNTVATL; Proimmune) peptides (CMVp, HIVp) to load f-DCZA+ andf-DCZA2, and by determining the frequency and total counts of viableCD8+ ab T cells specifically reacting against the SRVp, CMVp, and HIVpafter 14-d stimulation.
Quantification of the Mev pathway in APC
MC, mDC, and f-DCwere incubated at 13 106/ml for 24 h with 1 mCi [3H]acetate (3600 mCi/mmol; Amersham International, Bucks, U.K.). The in-tracellular synthesis of farnesyl-pyrophosphate (FPP) and cholesterol wasmeasured with the methanol/hexane extraction method (26). Intracellularand extracellular IPP levels were measured according to Benford et al. (27)with minor modifications. A total of 300 ml cells lysate or culture super-natant was diluted 1:2 into an ice-cold acetonitrile solution containing 100
The Journal of Immunology 1579

mM NaVO4 and centrifuged at 1200 3 g for 5 min at 4˚C. After lyophili-zation under vacuum, samples were resuspended in 20 ml dimethylhexyl-amine and separated by TLC. Gels were exposed to an iodine-saturatedatmosphere for 2 h, spots corresponding to each isoprenoid species wereisolated, and radioactivity uptake was measured by liquid scintillationcounting (Ultima Gold; Perkin Elmer). The titration curves for each iso-prenoids species were performed using three serial dilutions of [3H]cho-lesterol (Amersham), [3H]farnesyl pyrophosphate (Perkin Elmer), and [14C]IPP (Perkin Elmer), and counting them with liquid scintillation. The ra-dioactivity (in terms of cpm) of each sample was referred to the respectivetitration curve to obtain an index of the amount of cholesterol, FPP, and IPP,neosynthesized from the radiolabeled precursor and normalized to thenumber of cells per milliliter. According to the titration curve, the results areexpressed as fmol/1 3 106 cells for cholesterol, FPP, and intracellular IPP,and as pmol/1 3 106 cells for extracellular IPP.
Isolation of Tregs and proliferative assay
Tregs were purified from PBMC by immunomagnetic separation with theCD4+CD25+ T Regulatory Cells Isolation kit (Miltenyi Biotec, Bologna,Italy), according to the manufacturer’s instructions. In brief, CD4+ cellswere isolated by negative selection after depletion of cells expressing theCD8, CD11b, CD16, CD36, and CD56 Ags. CD25+ cells were then se-lected by positive selection with magnetic beads directly conjugated to ananti-CD25 Ab. From 5 3 105 to 1.8 3 106 CD4+ CD25+ cells (purity 76–95%) were obtained from buffy coats containing each from 200–3003 106
PBMC. Treg function was assessed as previously described (28). AllogenicPBMC were irradiated at 3000 rad and used as accessory cells, and 1 mg/ml soluble anti-CD3 (OKT3 Orthoclone; Janssen-Cilag, Cologno Monz-ese, Italy) was used to induce polyclonal T cell activation. Tregs (CD4+
CD25+) and CD4+CD252 cells were cultured alone (50 3 105/well) or incombination (1:1 ratio) in U-bottom 96-well plates in the presence orabsence of 5 mM ZA. Cell proliferation was evaluated on day 3 by adding1 mCi [3H]thymidine deoxyribose (Amersham) to each well during the last4 h of culture and measuring its uptake with a scintillation counter.
Statistical analysis
Results are expressed as mean values 6 SE. Differences between samplegroups were evaluated with the two-tailed nonparametric Mann–WhitneyU test for paired samples or with the Fisher exact test for unpaired data.Correlations were assessed by Pearson correlation coefficient. The sig-nificance cutoff value was p , 0.05. The p values are reported in thefigures.
ResultsmDCZA+ are potent inducers of gd T cell proliferationirrespective of R/NR status
The first series of experiments was deliberately confined to CTRLand MM R to identify the most effective APC inducing gd T cellproliferation in a susceptible population. As expected, no signifi-cant proliferation of gd T cells was observed with IL-2 alone inthe absence of ZA (Fig. 1). The highest gd T cell counts wereobserved after stimulation with ZA-treated mDC in both CTRL(p , 0.01) and MM (p , 0.3). MM (Fig. 1A, right panel) dis-played significantly higher counts than CTRL (Fig. 1A, left panel)after stimulation with both ZA-treated MC (p , 0.05) and ZA-treated mDC (p , 0.005).A significant increase of central memory (CM) and effector
memory (EM) gd T cells was detected after stimulation with ZA+IL-2 versus IL-2 alone in CTRL and MM R (p , 0.05). In MM,CM and EM gd T cell counts were significantly higher than inCTRL after both IL-2 and ZA+IL-2 stimulation (both p , 0.05).CM was the predominant subset in CTRL R, whereas EM waspredominant in MM R (Fig. 1B).Approximately 50% of MM patients at diagnosis display min-
imal or no gd T cell proliferation after PBMC stimulation with 1mM ZA + 10 IU/ml IL-2 (NR). Less than 5% of our CTRL wereNR. In view of the very effective gd T cell stimulatory capacity ofmDCZA+, we performed a side-by-side experiment with PBMCZA+
and MCZA+ in the two NR groups. mDCZA+ restored gd T cellproliferation in both groups (p , 0.0003; Fig. 1C). Flow cytom-
etry of gd T cells after stimulation with PBMCZA+ and mDCZA+ inone representative MM NR is shown in Fig. 1D.Immunophenotyping showed that mDCZA+ induced the expan-
sion of gd T cells equipped with surface receptors to exert co-stimulatory (HLA-DR, CD86, CD80), but not inhibitory (PD-1,PD-L1), activity on immune responses (Fig. 1E) in MM R and NR.
SRV-specific immune responses mediated by CD8+ ab T cellsare upregulated by the concurrent gd T cell proliferation
The baseline frequency of SRV-specific CD8+ ab T cells in theperipheral blood was significantly higher in MM than in CTRL(0.416 0.05 versus 0.076 0.03%; p, 0.0001; Supplemental Fig.1). The frequency and total counts of SRV-specific CD8+ ab T cellswas then investigated after stimulation with mDCSRVp2ZA2,mDCSRVp2ZA+, mDCSRVp+ZA2, and mDCSRVp+ZA+. Results fromrepresentative experiments in the CTRL and MMR and NR groupsare shown in Fig. 2A and 2B, and the pooled data are shown in Fig.2C. Given the equivalent gd T cell proliferation and number ofCD8+ ab T cells induced by mDCZA+, R and NR individuals weregrouped together in CTRL andMM. Total counts were significantlyhigher after stimulation with mDCSRVp+ZA+ compared with stimu-lation with mDCSRVp+ZA2 (p , 0.05 in both CTRL and MM), in-dicating that the concurrent activation of gd T cells and CD8+ abT cells was not mutually detrimental.The cytotoxic activity of SRV-specific CD8+ ab T cells was
tested against the HLA-A2+ SRVp-pulsed T2 cell line (T2SRVp+).As expected, the highest cytotoxicity was observed when T cellswere stimulated with mDCSRVp+ZA+ in both CTRL and MM (p ,0.05 compared with cytotoxic values after stimulation withmDCSRVp+ZA2 in both groups) (Fig. 2D). Similar results wereobserved with RPMI 8226, a myeloma-derived SRV+ HLA-A2+
cell line (Supplemental Fig. 2).
f-DCZA+ are as effective as conventional mDCZA+ in inducinggd T cell proliferation and upregulating SRV-specific immuneresponses
The production of conventional mDC is a time-consuming, labor-intensive, and costly process that requires enough PBMC togenerate the mDC needed for two rounds of stimulation. Toovercome these limitations, we used f-DCZA+ to stimulate autol-ogous gd T cells. f-DC were generated in 48 h from CTRL andMM irrespective of their R/NR status. The immunophenotype andendocytic capacity of immature and mature before (f-DCZA2) andafter (f-DCZA+) ZA treatment are shown in Supplemental Table Iand Supplemental Fig. 3. A side-by-side comparison in CTRL Rbetween the ability of f-DCZA+ and mDCZA+ to stimulate autol-ogous gd T cells and induce the expansion of SRV-specific CD8+
ab T cells is shown in Supplemental Fig. 4. The total number ofgd T cells was not statistically different either on day 10 (after oneround) or 20 (after two rounds), whereas the number of SRV-specific CD8+ ab T cells per well was even higher after stimu-lation with f-DCZA+. We also compared the total numbers of gdT cells and SRV-specific CD8+ ab T cells per well after two 10-d stimulations as opposed to only one of 14 d without detectingany difference (Supplemental Fig. 4). Based on these results, f-DCZA+ were also used to stimulate gd T cells in MM patients.Indeed, they effectively induced the proliferative expansion of gdT cells in CTRL R and NR (p , 0.04 and p , 0.03, respectively),and in MM R and NR (both p, 0.04; Fig. 3A). Flow cytometry ofreinstated T cell proliferation in representative CTRL and MMNR are shown in Fig. 3B. f-DCZA+ also amplified SRV-specificimmune responses in both CTRL and MM R and NR. Flowcytometry of representative samples is shown in Fig. 3C and 3D.The specificity of MHC-restricted immune responses generated
1580 IMMUNOMODULATION BY ZOLEDRONIC ACID IN MM

by the costimulation of gd T cells and CD8+ ab T cells is dem-onstrated in Fig. 3E. CTRL T cells were stimulated with f-DCZA+
and f-DCZA+ loaded or unloaded with the HLA-A2–restrictedSRVp, CMVp, and HIVp. The frequency of specific CD8+ abT cells was determined with the corresponding PE-labeled pen-tamers. As expected, only CD8+ ab T cells specific for the peptideused to load f-DCZA2 and f-DCZA+ were increased after 14-d in-cubation. Pooled data reporting total counts of SRV-specific CD8+
ab T cells and cytotoxicity against the RPMI 8226 myeloma cellline are shown in Fig. 3F and 3G, respectively. The most effec-tive anti-SRV activity was observed after stimulation withf-DCSRVp+ZA+ in both CTRL and MM.Fig. 3H shows that CD8+ ab T cells elicited by f-DCSRVp+ZA+
are also able to recognize and kill primary myeloma cells. Thespecificity of this antitumor immune response is corroborated by
the observation that T cells do not exert any cytotoxicity whenCMVp is used to load f-DC.
Accelerated Mev activity in mDC and f-DC, and enhanced IPPproduction after ZA treatment
The ability of ZA-treated APC to activate cells is dependent ontheir capacity to inhibit FPPS in the Mev pathway (10). We havecompared the Mev activity rate in MC, mDC, and f-DC at baselineand after treatment with 5 mM ZA (i.e., the conditions used tostimulate T cells). Baseline cholesterol and both intracellular andextracellular IPP levels progressively increased from MC to f-DC,with mDC displaying intermediate values (Fig. 4). Unexpectedly,extracellular IPP concentrations were 1000 times greater than theintracellular concentrations. ZA-induced FPPS inhibition de-creased the production of cholesterol and FPP, and further in-
FIGURE 1. mDCZA+ are very potent inducers of gd T cell proliferation irrespective of R/NR status. A, Total counts of viable gd T cells per well in CTRL
and MM R after 7-d stimulation with IL-2 and untreated (ZA2) or ZA-treated (ZA+) PBMC, MC, iDC, and mDC. Cultures were initiated at 1 3 106/ml
with 2 3 105 PBMC or PBL and MC, iDC, and mDC (5:1 ratio) in a final volume of 200 ml/well. Multiple wells were set up as required. Bars represent the
mean values 6 SE from 10 CTRL and 5 (MC, iDC, and mDC) to 10 (PBMC) MM. Differences between untreated and ZA-treated cells are statistically
significant in CTRL (PBMC: p , 0.02; MC: p , 0.01; iDC: p , 0.03; mDC: p , 0.01), and MM (PBMC: p , 0.03; MC: p , 0.02; iDC: p , 0.02; mDC:
p , 0.03). Differences between CTRL and MM are statistically significant after stimulation with ZA-treated MC (p , 0.05) and ZA-treated mDC (p ,0.005). B, Phenotypic analysis of gd T cell subsets in CTRL and MM R after 7-d stimulation with IL-2 and ZA-treated mDC alone or in combination. Bars
represent the mean values 6 SE from six CTRL and six MM R. Cultures were set up as described earlier. CM was the predominant subset in the CTRL,
whereas EM was predominant in MM. The latter showed significantly higher CM and EM counts than CTRL (p , 0.05 and p , 0.05, respectively). C,
Total counts of viable gd T cells after stimulation of PBMC, MC, and mDC untreated (ZA2) or pretreated with ZA (ZA+) in CTRL (left panel) and MM
(right panel) NR. Cultures were set up as described earlier. Bars represent the mean values 6 SE of three side-by-side experiments performed in three
CTRL and three MM NR. Differences are statistically significant in CTRL and MM (both p , 0.0003) NR. D, Flow cytometric comparison of gd T cell
expansion after stimulation with ZA-treated PBMC (PBMCZA+) and mDC (mDCZA+) in one representative MM NR. E, Flow cytometry of the expression
of costimulatory (HLA-DR, CD86, CD80) and inhibitory (PD-1, PD-L1) molecules on gd T cells after stimulation with mDCZA+ in representative MM R
and NR.
The Journal of Immunology 1581
creased intracellular and extracellular IPP levels. All differenceswere statistically different between untreated and ZA-treated cells.The most striking effect was the increase of extracellular IPP inthe supernatants of f-DC to concentrations almost 10 times greaterthan those in the supernatants of ZA-treated MC and mDC.To further demonstrate the key role played by the Mev pathway,
mDC and f-DC were treated with ZA in the presence or absenceof simvastatin (Sim). Sim prevents ZA-induced IPP accumulationby specifically inhibiting hydroxy-methylglutaryl-coenzyme A re-ductase, the rate-limiting enzyme in the Mev pathway acting up-stream from FPPS (29, 30). As predicted (9, 22, 30), Simabrogated the ability of mDCZA+ and f-DCZA+ to induce theproliferation of gd T cells (data not shown; see also Figs. 5, 6).
The IPP released in the supernatants of f-DCZA+ is sufficient toinduce gd T cell proliferation
PBMC and PBL from CTRL R were incubated for 7 d with thesupernatants collected from f-DCZA2 and f-DCZA+, and supple-mented with 10 IU/ml IL-2. Final dilutions were 1, 10, and 20%corresponding to extracellular IPP concentrations of 14, 144, and288 nM, respectively (Fig. 5B, left panel). Standard cultures withIL-2 and ZA+IL-2 served as CTRL (Fig. 5A). As expected, gdT cell proliferation was induced only in PBMC, because PBL aredepleted of MC and cannot generate adequate IPP amounts to
stimulate gd T cells (9, 10). By contrast, gd T cell proliferation
was equivalent in PBMC and PBL after incubation with f-DCZA+
supernatants, indicating that the concentration of extracellular IPP
was sufficient to induce gd T cell proliferation, even in the ab-
sence of MC, as previously reported for exogenous IPP (2). In
confirmation, PBMC and PBL from CTRL R were stimulated with
exogenous IPP at concentrations including those detected in f-
DCZA+ supernatants (from 14 to 280 nM; Fig. 5B, right panel).
Exogenous IPP induced the proliferation of gd T cells, but with
lower efficiency than f-DCZA+ supernatants, especially if used in
the presence of MC (PBMC versus PBL) and at concentrations
equivalent to those of f-DCZA+ supernatants (i.e., from 1028 to
1027 M). It also induced gd T cell proliferation in PBMC and PBL
as effectively as f-DCZA+ supernatants when used at higher con-
centrations (.1026 M). These results suggest that other cytokines
and/or growth factors may be present in f-DCZA+ supernatants and
promote gd T cell proliferation in association with the low en-
dogenous IPP concentrations. It is also possible that MC prefer-
entially use exogenous IPP to the disadvantage of gd T cells,
because higher concentrations are required to induce gd T cell
proliferation in PBMC versus PBL.Extracellular IPP was also measured in the supernatants of f-
DCZA+ cultured in the presence or absence of Sim. As predicted,
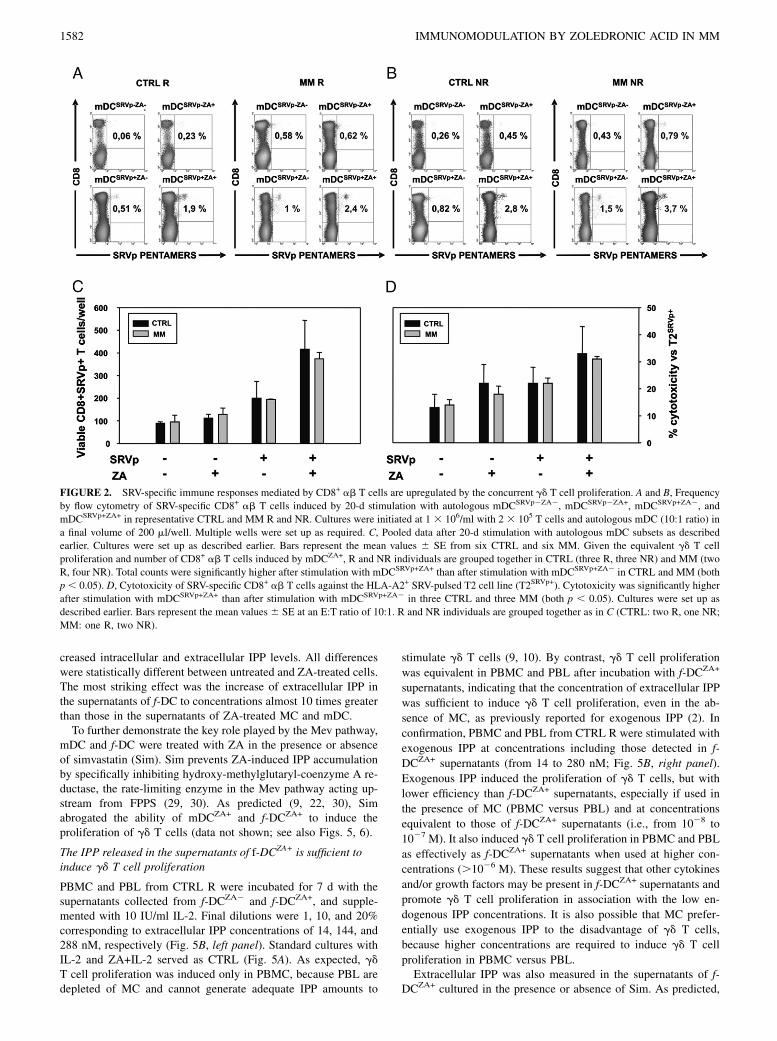
FIGURE 2. SRV-specific immune responses mediated by CD8+ ab T cells are upregulated by the concurrent gd T cell proliferation. A and B, Frequency
by flow cytometry of SRV-specific CD8+ ab T cells induced by 20-d stimulation with autologous mDCSRVp2ZA2, mDCSRVp2ZA+, mDCSRVp+ZA2, and
mDCSRVp+ZA+ in representative CTRL and MM R and NR. Cultures were initiated at 1 3 106/ml with 2 3 105 T cells and autologous mDC (10:1 ratio) in
a final volume of 200 ml/well. Multiple wells were set up as required. C, Pooled data after 20-d stimulation with autologous mDC subsets as described
earlier. Cultures were set up as described earlier. Bars represent the mean values 6 SE from six CTRL and six MM. Given the equivalent gd T cell
proliferation and number of CD8+ ab T cells induced by mDCZA+, R and NR individuals are grouped together in CTRL (three R, three NR) and MM (two
R, four NR). Total counts were significantly higher after stimulation with mDCSRVp+ZA+ than after stimulation with mDCSRVp+ZA2 in CTRL and MM (both
p , 0.05). D, Cytotoxicity of SRV-specific CD8+ ab T cells against the HLA-A2+ SRV-pulsed T2 cell line (T2SRVp+). Cytotoxicity was significantly higher
after stimulation with mDCSRVp+ZA+ than after stimulation with mDCSRVp+ZA2 in three CTRL and three MM (both p , 0.05). Cultures were set up as
described earlier. Bars represent the mean values 6 SE at an E:T ratio of 10:1. R and NR individuals are grouped together as in C (CTRL: two R, one NR;
MM: one R, two NR).
1582 IMMUNOMODULATION BY ZOLEDRONIC ACID IN MM

FIGURE 3. f-DCZA+ are as effective as conventional mDCZA+ in inducing gd T cell proliferation and upregulating SRV-specific immune response. A,
Total counts of viable gd T cells after stimulation with untreated (ZA2) or ZA-treated (ZA+) f-DC in CTRL and MM R and NR. Cultures were initiated at
13 106/ml with 23 105 T cells and autologous f-DC (10:1 ratio) in a final volume of 200 ml/well. Multiple wells were set up as required. Bars represent the
mean values 6 SE from five CTRL (three R, two NR) and five MM (three R, two NR). ZA-treated f-DC induced gd T cell proliferation in both CTRL and
MM R and NR. Differences between untreated and ZA-treated f-DC are statistically significant in CTRL R and NR (p , 0.04 and p , 0.03, respectively),
and in MM R and NR (both p , 0.04), whereas differences between CTRL and MM are not statistically different. B, Representative flow cytometry of
reinstated gd T cell proliferation in individual CTRL and MM NR after stimulation with ZA-treated f-DC (f-DCZA+). C and D, Flow cytometry of SRV-
specific CD8+ cells by pentamer staining after 14-d stimulation of autologous T cells with f-DCSRVp+ZA2 and f-DCSRVp+ZA+ in CTRL and MM R and NR.
E, Frequency of pentamer-specific CD8+ ab T cells after stimulation of CTRL T cells with autologous f-DCZA2 and f-DCZA+ loaded or unloaded with
HLA-A2–restricted SRVp, CMVp, and HIVp. Pentamer staining shows that there is no cross-reactivity, and only CD8+ ab T cells specific for the cor-
responding peptide used to load f-DCZA2 and f-DCZA+ are increased after 14-d incubation. F, Pooled data reporting total counts of SRV-specific CD8+ ab
T cells. Cultures were set up as described earlier. Bars represent the mean values 6 SE from five CTRL and five MM. Given the equivalent gd T cell
proliferation and number of CD8+ ab T cells induced by f-DCZA+, R and NR individuals are grouped together in CTRL (two R, three NR) and MM (two R,
three NR). Total counts were significantly higher after stimulation with f-DCSRVp+ZA+ than after stimulation with f-DCSRVp+ZA2 in CTRL and MM (both
The Journal of Immunology 1583

Sim almost set to zero extracellular IPP concentrations (Fig. 5C)and the supernatant of f-DCZA+Sim+ failed to induce any pro-liferation of gd T cells irrespective of whether MC were present inthe responding population (Fig. 5D).
Antagonism of ZA and gd T cells versus Tregs
gd T cell proliferation can be suppressed by Tregs (17, 31), andtheir excess over gd T cells has been proposed as an explanationfor defective ZA-induced gd T cell proliferation in MM NR (17).Treg and gd T cell counts and ratios were not significantly dif-ferent in function of R/NR status in the peripheral blood of CTRLand MM patients (data not shown). We also looked for correla-tions in individual CTRL and MM R between baseline Treg andgd T cell values, and the increase of gd T cells after stimulationwith ZA+IL-2. The only statistically significant finding was a di-rect correlation between baseline gd T cell counts and their foldincrease after stimulation in CTRL R, whereas no correlation wasdetected with Tregs (data not shown).Total counts of viable Tregs were also calculated after 7-d PBMC
stimulation in the presence or absence of IL-2, ZA, or both (Fig. 6A).These are the only culture conditions that discriminate between theR/NR status, which is no longer detectable aftermDCZA+ or f-DCZA+
stimulation. IL-2 is a major Treg growth factor (32); therefore, theirincrease after incubationwith 10 IU/ml IL-2was expected. Notably,the increase was higher in MM R than in MM NR, which rules outenhanced Treg IL-2 reactivity in the latter. The increase was sig-nificantly blunted when ZAwas added to IL-2 (ZA+IL-2 versus IL-2) in both MM R (p , 0.05) and NR (p , 0.05), even if gd T cellproliferation occurred in the former only. The fact that Treg counts
were also higher after incubation in RPMI 1640 than after stimu-lation with ZA alone (p, 0.05 in both R and NR), a condition thatdoes not lead to any gd T cell expansion even in MM R, providesfurther evidence that ZA re strains Treg expansion via gd T cell-dependent and -independent mechanisms. Flow cytometry of Tregafter PBMC stimulation in representative MM R and NR is illus-trated in Supplemental Fig. 5A.To further investigate the mechanisms exploited by ZA to an-
tagonize Tregs independently of gd T cells, we tested the abilityof Tregs to regulate the proliferation of autologous CD4+CD252
driven by irradiated accessory cells and OKT3 in the presence orabsence of ZA (Fig. 6B). CD4+CD252 proliferated in response tostimulation with anti-CD3 mAb and irradiated accessory cells,whereas Tregs were anergic. Proliferation was significantlyinhibited (p , 0.002) when CD4+CD252 and Tregs were cocul-tured at 1:1 cell ratio. This inhibition was reverted when ZA wasadded (p , 0.03). This effect was not dependent on gd T cellsbecause their proliferation was never observed under these cultureconditions (data not shown). Tregs were purified, pretreated with 5mM ZA for 24 h, and washed before mixing with autologousCD4+CD252 T cells, to further demonstrate a direct effect. Theresults in Supplemental Fig. 5B (left panel) indicate that Tregswere still able to suppress the proliferative response of CD4+
CD252 T cells. Sim was also added in two experiments to confirmthat the ZA-induced anti-Treg activity did not require any gd
T cell proliferation and was independent of the inhibition of theMev pathway (Supplemental Fig. 5B, right panel).Previous reports have shown that MM DC are inclined to
stimulate T cells with inhibitory functions (21, 22). Thus, Treg
FIGURE 4. Accelerated Mev activity in mDC
and f-DC resulting in enhanced intracellular
(IPPintra) and extracellular IPP (IPPextra) pro-
duction after ZA treatment. Quantification of
the Mev activity in MC, mDC, and f-DC at
baseline (ZA2) and after treatment with 5 mM
ZA (ZA+). Bars represent the mean values6 SE
from three CTRL. mDC and f-DC showed an
accelerated baseline Mev activity compared
with MC, as shown by the increased production
of cholesterol, FPP, IPPintra, and IPPextra. Cho-
lesterol, FPP, and IPPintra levels are shown in the
left panels and are expressed as fmol/1 3 106
cells. IPPextra levels are shown in the right
panels and are expressed as pmol/1 3 106 cells.
ZA induced a decreased production of choles-
terol and FPP, and an increase of IPPintra and
IPPextra levels in MC (p , 0.05), mDC (p ,0.04), and f-DC (p , 0.0001). The highest
levels of IPPextra were reached in the super-
natants of f-DCZA+ (p , 0.001 versus MC, p ,0.02 versus mDC). Supernatants of mDCZA+
also contained higher levels of IPPextra than
MCZA+ (p , 0.002).
p , 0.05). G, Cytotoxicity against the HLA-A2+ SRV+ RPMI 8226 myeloma cell line. Bars represent the mean values 6 SE of experiments from three
CTRL (two R, one NR) and from one MM NR at an E:T ratio of 10:1. Cytotoxicity was higher after stimulation with f-DCSRVp+ZA+ in both CTRL and MM
(p , 0.05 in CTRL; p value not evaluated in MM because data are from one experiment only); (H) cytotoxic activity of SRVp-specific CD8+ ab T cells
against primary HLA-A2+ myeloma cells isolated from the peripheral blood of a PCL patient. T cells were stimulated for 14 d with autologous f-DCZA2 and
f-DCZA+ loaded or unloaded with SRVp and CMVp as CTRL. CD8+ ab T cells stimulated with f-DCSRVp+ZA+ recognize and kill primary myeloma cells,
whereas no cytotoxicity is observed after stimulation with the other f-DC subsets, including f-DCCMVp+ZA+ .
1584 IMMUNOMODULATION BY ZOLEDRONIC ACID IN MM

counts were evaluated in MM R and NR after stimulation withZA-treated and untreated mDC and f-DC (Fig. 6C). As expected,ZA induced the proliferation of gd T cells, and this was uniformlyassociated with lower Treg counts in both MM R and NR. Toinvestigate whether ZA could also antagonize Tregs independentlyof gd T cell proliferation in these cultures, we evaluated Tregcounts on day 7 after stimulation with f-DCZA+ and f-DCZA2 inthe presence or absence of Sim. Sim alone had no effect on Tregsand did not impair the ability of ZA to antagonize Tregs (Fig. 6D,upper panels) even if Sim completely abrogated gd T cell pro-liferation (Fig. 6D, lower panels). Altogether, these results in-dicate that ZA antagonizes Tregs independently of gd T cellproliferation and Mev pathway inhibition.
ZA downregulates PD-L2 expression in mDC and f-DC
PD-L1 and PD-L2 expression was evaluated in MC, mDC, and f-DC under basal conditions and after ZA treatment. PD-L1 wasconstitutively expressed on MC, mDC, and f-DC, and its expres-sion was not modified by ZA (data not shown, Fig. 7B). PD-L2was expressed by a smaller proportion of resting MC and was notchanged after ZA treatment (Fig. 7A). By contrast, PD-L2 wasexpressed by most mDC and f-DC under basal conditions, and this
expression was significantly downmodulated by ZA in CTRLmDC (p , 0.001) and f-DC (p , 0.05), and MM mDC (p , 0.05;Fig. 7A).Sim alone had no effect on PD-L2 expression and did not prevent
ZA-induced PD-L2 downregulation (Fig. 7B, upper row). It alsohad no effect on the expression of PD-L1, whether alone or inassociation with ZA (Fig. 7B, lower row). These data indicate thatZA modulates the cell surface immunophenotype of mDC and f-DC independently from the Mev pathway.
Inhibition of gd T cell expansion by Sim prevents theamplification of SRV-specific CD8+ ab T cells
The generation of SRV-specific CD8+ ab T cells after stimulationwith mDCSRVp+ZA+ was evaluated in the absence or presence ofSim. As reported earlier, Sim prevented the IPP-dependent pro-liferation of gd T cells (Fig. 8A), which, in turn, was associatedwith a failed amplification of SRV-specific CD8+ ab T cells (Fig.8B). These results provide indirect evidence that activated of gdT cells can improve adaptive immune responses.
Discussiongd T cells play an important role in immunosurveillance in severalways. Their contribution as effector cells of innate immunity is
FIGURE 5. The amount of IPP in the supernatants of f-DCZA+ is sufficient to induce gd T cell proliferation independently from MC. A, PBMC and PBL
were stimulated for 7 d with IL-2 and ZA+IL-2, and served as CTRL. Cultures were initiated as reported in Fig. 1A. Bars represent the mean values 6 SE
from six CTRL R. As expected, gd T cells did not proliferate in the absence of MC (PBL). B, The supernatants (SN) of f-DCZA2 and f-DCZA+ generated
from the same series of six CTRL R shown in Awere used to side-by-side stimulate autologous PBMC and PBL (left panel). Final dilutions were 1, 10, and
20%, which are equivalent to IPP concentrations of 14, 140, and 280 nM, respectively. gd T cell proliferation was detected only after stimulation with SN f-
DCZA+, irrespective of the presence (PBMC) or absence (PBL) of MC. Exogenous IPP was also used to stimulate PBMC and PBL at concentrations from 10
nM to 10 mM (from 1028 to 1025 M) to include concentrations in the range of those detected in f-DCZA+ supernatants (right panel). A comparable gd T cell
proliferation in PBMC and PBL was observed only at high concentrations.10 mM (i.e.,.1025 M). Cultures were initiated with 23 105 PBMC or PBL as
R cells in 200 ml/well. C, f-DC treatment with Sim abrogates ZA-induced IPP production and yields to supernatants devoid of IPP. Results are from one of
two experiments. D, Comparison of the ability of supernatants of ZA-treated f-DC cultured for 24 h in the absence (f-DCZA+Sim2) or presence (f-DCZA+Sim+)
of Sim to induce autologous gd T cell proliferation. Results are from one of two experiments. PBMC and purified T cells were used to confirm that
extracellular IPP induces gd T cell proliferation in the absence of MC. Final dilutions and IPP concentrations are as for B.
The Journal of Immunology 1585
very well recognized, and several studies are investigating thepossibility of exploiting their effector properties in infections andcancer (1, 5). Much less is known about their ability to act ascellular adjuvants bridging innate and adaptive immunity, espe-cially in patients with cancer. We have addressed this issue inMM, a prototypic disease characterized by multiple immunedysfunctions that also involves gd T cells and DC (9, 15–19).The first series of experiments was deliberately confined to
CTRL and MMR to identify the APC most capable of inducing theproliferation of autologous gd T cells. mDCZA+ generated fromMM R were even more efficient than their normal counterparts ininducing this proliferation. After 7-d stimulation, there was a pref-erential expansion of CM and EM gd T cells in MM R patients thatwas significantly greater than that observed in CTRL. By analogywith conventional CD8+ cells (33, 34), CM gd T cells are creditedas the preferred subset for adoptive immunotherapy because theyhave self-renewal capacity and more effective antitumor activity
in vivo than terminally differentiated CD45RA+ (TEMRA) cells,even though the latter are more potent in vitro (35).Approximately 50% ofMM patients at diagnosis are classified as
NR because gd-T cell proliferation is not detected after challengingPBMC for 7 d with ZA+IL-2 (9, 15–17). A similar proportion hasbeen reported in patients with other lymphoproliferative disordersand metastatic solid cancer (17). gd T cells from MM NR produceIFN-g and exert cytotoxic activity against myeloma cells onceactivated by ZA+IL-2 (9), but NR patients are typically excludedfrom gd T cell-based adoptive immunotherapy trials because of theimpossibility of increasing the number of cells and establishinga favorable E:T cell ratio in vivo or ex vivo.In view of the high efficiency of mDCZA+, we tested their ability
to induce gd T cell proliferation in CTRL and MM NR. Occa-sionally, healthy individuals (,5% in our experience) are alsorefractory to ZA stimulation. mDCZA+ were very effective in bothCTRL and MM NR, and resulted in the proliferative expansion of
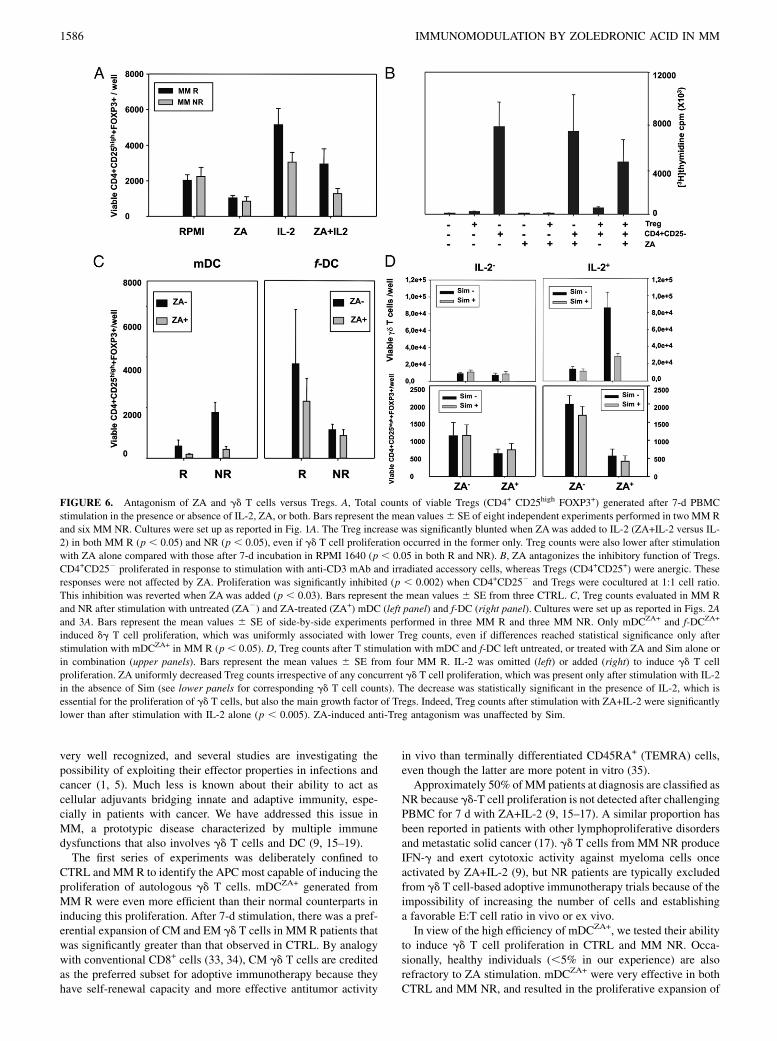
FIGURE 6. Antagonism of ZA and gd T cells versus Tregs. A, Total counts of viable Tregs (CD4+ CD25high FOXP3+) generated after 7-d PBMC
stimulation in the presence or absence of IL-2, ZA, or both. Bars represent the mean values6 SE of eight independent experiments performed in two MM R
and six MM NR. Cultures were set up as reported in Fig. 1A. The Treg increase was significantly blunted when ZAwas added to IL-2 (ZA+IL-2 versus IL-
2) in both MM R (p , 0.05) and NR (p , 0.05), even if gd T cell proliferation occurred in the former only. Treg counts were also lower after stimulation
with ZA alone compared with those after 7-d incubation in RPMI 1640 (p , 0.05 in both R and NR). B, ZA antagonizes the inhibitory function of Tregs.
CD4+CD252 proliferated in response to stimulation with anti-CD3 mAb and irradiated accessory cells, whereas Tregs (CD4+CD25+) were anergic. These
responses were not affected by ZA. Proliferation was significantly inhibited (p , 0.002) when CD4+CD252 and Tregs were cocultured at 1:1 cell ratio.
This inhibition was reverted when ZA was added (p , 0.03). Bars represent the mean values 6 SE from three CTRL. C, Treg counts evaluated in MM R
and NR after stimulation with untreated (ZA2) and ZA-treated (ZA+) mDC (left panel) and f-DC (right panel). Cultures were set up as reported in Figs. 2A
and 3A. Bars represent the mean values 6 SE of side-by-side experiments performed in three MM R and three MM NR. Only mDCZA+ and f-DCZA+
induced dg T cell proliferation, which was uniformly associated with lower Treg counts, even if differences reached statistical significance only after
stimulation with mDCZA+ in MM R (p , 0.05). D, Treg counts after T stimulation with mDC and f-DC left untreated, or treated with ZA and Sim alone or
in combination (upper panels). Bars represent the mean values 6 SE from four MM R. IL-2 was omitted (left) or added (right) to induce gd T cell
proliferation. ZA uniformly decreased Treg counts irrespective of any concurrent gd T cell proliferation, which was present only after stimulation with IL-2
in the absence of Sim (see lower panels for corresponding gd T cell counts). The decrease was statistically significant in the presence of IL-2, which is
essential for the proliferation of gd T cells, but also the main growth factor of Tregs. Indeed, Treg counts after stimulation with ZA+IL-2 were significantly
lower than after stimulation with IL-2 alone (p , 0.005). ZA-induced anti-Treg antagonism was unaffected by Sim.
1586 IMMUNOMODULATION BY ZOLEDRONIC ACID IN MM

gd T cells phenotypically indistinguishable from that observed inCTRL and MM R (HLA-DR+, CD80+, CD86+, PD-12, PD-L12).The lack of PD-1 expression sets gd T cells stimulated by mDCZA+
apart from invariant NKT cells stimulated by synthetic analogsof their natural ligands. Unlike gd T cells, invariant NKT cellsrapidly upregulate PD-1 expression after stimulation and retainthis expression for at least 30 d (36). PD-1 expression has beenimplicated in the induction and maintenance of tolerance in a va-riety of settings, and interpreted as a marker of T cells unfitted toexert antitumor activity because of their functional exhaustion (37).Cabillic et al. (20) have also reported in solid tumors that gd T cellproliferation can be reinstated in NR patients using ZA-treated DC.The next step was to determine whether gd T cells induced by
mDCZA+ could improve immune responses against tumor-as-sociated Ags mediated by CD8+ ab T cells not only in R MM,
but also in those NR MM in which gd T cell proliferation wasreinstated. SRV was selected as a prototypic tumor-associated Agbecause it is expressed by myeloma cells and contributes to theirsurvival (38). SRV-specific T cells have been tolerized but notdeleted from the MM T cell repertoire (23), and their effectorfunctions can be reinstated by appropriate SRV presentation andT cell stimulation (39, 40). Moreover, SRV is also expressed bymany other tumor cells of hematopoietic and epithelial origin (41).The frequency of SRV-specific CD8+ ab T cells in the pe-
ripheral blood of MM was higher than in CTRL, suggestinga previous exposure and selective recruitment in the early phasesof the disease. Their frequency was significantly enhanced in bothCTRL and MM after stimulation with mDCSRVp+ZA+, leading toenhanced cytotoxicity against the appropriate HLA*0201+ SRV+
target cells irrespective of the initial R/NR status. These data
FIGURE 7. ZA downregulates PD-L2 expression in mDC and f-DC. A, Cell surface expression of PD-L2 on untreated (ZA2) or ZA-treated (ZA+) MC,
mDC, and f-DC in CTRL (left panel) and MM (right panel). PD-L2 was expressed by a smaller proportion of resting MC and remained unchanged after ZA
treatment, whereas it was expressed by the majority of mDC and f-DC under basal conditions, and this expression was uniformly downmodulated by ZA in
both CTRL and MM. Downmodulation reached a statistical significance in CTRL mDC (p , 0.001) and f-DC (p , 0.05), and MM mDC (p , 0.05). Bars
represent the mean values 6 SE from five CTRL and five MM. B, Flow cytometry of PD-L1 and PD-L2 expression after mDC treatment with ZA, Sim, or
their combination. Representative data from one of three experiments. PD-L2 expression was not modified by Sim (upper row, left panel), whereas it was
downmodulated by ZA (upper row, center panel). Sim did not prevent ZA-induced PD-L2 downregulation (upper row, right panel). Sim and ZA did not
affect PD-L1 expression, either alone or in association (lower row).
FIGURE 8. Inhibition of gd T cell expansion by Sim prevents the amplification of SRV-specific CD8+ ab T cells. A, Total counts of viable gd T cells
after 10-, 14-, and 20-d stimulation with untreated (Sim2) or Sim-treated (Sim+) mDCSRVp+ZA+. As expected, gd T cell proliferation was abrogated by Sim
treatment. Differences were statistically different on days 14 (p , 0.005) and 20 (p , 0.01). B, Total counts of viable SRV-specific CD8+ ab T cells after
10-, 14-, and 20-d stimulation with Sim2 or Sim+ mDCSRVp+ZA+. As for gd T cells, the expansion of SRV-specific CD8+ ab T cells was abrogated by Sim.
Differences are statistically different on days 14 (p , 0.005) and 20 (p , 0.005). Bars represent the mean values 6 SEM from three CTRL R.
The Journal of Immunology 1587

indicate that mDCSRVp+ZA+ of CTRL and MM can handle theconcurrent activation of autologous gd T cells and tumor-specificCD8+ ab T cells with a mutual edge on their immune perform-ances. The final outcome of these reciprocally gainful interactionsis the generation of a composite population of innate and adaptiveantitumor effector cells, embodying both CM and EM gd T cellsand CD8+ ab T cells.The generation of conventional mDCZA+ by standard protocols
is a labor-intensive, time-consuming, and costly procedure thatrequires adequate cell inputs that, in turn, are dependent on suf-ficient blood samples. To corroborate the translational value of ourfindings, we conducted a subsequent series of experiments usingDC generated according to an alternative protocol yielding mDCfrom CD14+ cells within only 48 h of in vitro culture (f-DC) (24,42). This strategy has been adapted and validated for large-scaleproduction under GMP conditions in patients with advancedprostate cancer (43), but the ability of f-DC to induce the pro-liferation of gd T cells and concurrently activate MHC-restrictedCD8+ ab T cells is unknown. We have shown that f-DC can besuccessfully generated from CD14+ cells in both CTRL and MMR and NR. f-DCZA+ were as effective as mDCZA+ in inducing gdT cell proliferation in CTRL and MM irrespective of their R/NRstatus. One round of stimulation and a 14-d incubation weresufficient to maximize the frequency of SRV-specific CD8+ cells,as shown by side-by-side experiments comparing mDCZA+ and f-DCZA+. The cytotoxic function of these cells was confirmed bytheir ability to recognize and kill the appropriate HLA*0102+
SRV+ target cells, including primary PCL cells, after 14 d ofstimulation by f-DCZA+. We selected this time period as the lon-gest affordable with a single round of stimulation because Daueret al. (42) and Jarnjak-Jankovic et al. (43) have shown that 5–7 d are sufficient for f-DC to induce primary and secondary Ag-specific T cell immune responses, and because the kinetics ofSRV-specific CD8+ ab T cell expansion significantly increasedfrom days 10 to 14, but not from days 14 to 20 (see also Fig. 8B,Supplemental Fig. 4).As stated earlier, gd T cells are activated by IPP, an intermediate
Mev metabolite mimicking their natural ligands (6, 8–10). Underbasal conditions, the highest IPP production was observed in mDCand f-DC, the latter showing significantly higher values. As pre-dicted, IPP concentrations further increased after ZA treatment,and the most striking increase was detected in the supernatants off-DCZA+ in which extracellular IPP concentrations were 10 timesgreater than those detected in MCZA+ and mDCZA+ supernatants.Unexpectedly, IPP concentrations in the supernatants were 1000times greater than the intracellular ones. Thus, IPP is producedand released in the supernatant in sufficient amounts to induce gdT cell proliferation, even in the absence of MC. These results arein line with previous studies showing that gd T cell activation byexogenous IPP is TCR mediated and APC independent (2). Wehave also observed that exogenous IPP induces the proliferation ofgd T cells, but with a lower efficiency than f-DCZA+ supernatants,especially if used at low concentrations and in the presence ofMC. So far, most studies have investigated the activity of IPPat high concentrations on purified gd T cells, gd T cell lines, orgd T cell clones, whereas few data are available for IPP at lowconcentrations on MC-containing populations. The lower effi-ciency of exogenous IPP compared with f-DCZA+ supernatants,especially in PBMC, could reflect either an MC-mediated IPPsequestration to the disadvantage of gd T cells or the presenceof additional soluble factors in f-DCZA+ supernatants. Studies arecurrently in progress to address this issue.The importance of IPP in f-DCZA+ supernatants was confirmed
by the experiments in which Sim was used as an irreversible and
selective hydroxy-methylglutaryl-coenzyme A reductase inhibitorto prevent intracellular IPP accumulation (11, 30), and its releasein the supernatants. Indeed, Sim treatment blunted the prolif-erative activation of gd T cells induced by mDCZA+ and f-DCZA+
by setting almost to zero extracellular IPP concentrations.Activated gd T cells are enabled to behave as cellular adju-
vants in several ways, including the expression of costimulatoryreceptors and appropriate cytokine production. The frequency ofSRV-specific CD8+ ab T cells was uniformly higher in cultures inwhich gd T cells had proliferated, and this increase was mostlyevident after gd T cells had become the predominant population.These data confirm previous results in healthy individuals in-dicating that gd T cell proliferation anticipates and is not detri-mental to the subsequent increase of Ag-specific CD8+ ab T cells(11, 14). Activated CM and EM gd T cells are enabled to behaveas cellular adjuvants in several ways, including the expression ofcostimulatory receptors and the production of IFN-g, which pro-motes the functional maturation of DC and the developmentof antitumor cell-mediated immune responses (14, 23, 43, 44).mDCZA+ and f-DCZA+ themselves may contribute to Th1 polari-zation, because we have previously shown that mDCZA+ producemore IL-12 and less IL-6 compared with mDCZA+ (11).gd T cells antagonized the IL-2–induced proliferation of Tregs.
Low-dose IL-2 is indispensable to sustain ZA-induced gd T cellproliferation, but it is also the main growth factor for Tregs (32),and mDC from MM patients are intrinsically inclined to promotethe growth of Tregs (21, 22). Our results demonstrate that gdT cell proliferation induced by PBMCZA+, mDCZA+, and f-DCZA+
uniformly restrained IL-2–induced Treg expansion. Gong et al.(45) have recently shown a similar anti-Treg activity in PBMCfrom nonhuman primates after proliferative expansion of gdT cells induced by BrHPP and IL-2, provided that MC and/orother accessory cells were included in the cultures. A new find-ing from our experiments was that ZA counteracted Treg in-dependently from gd T cell proliferation. Lower Treg counts weredetected in MM NR after stimulation of PBMC with ZA+IL-2compared with IL-2 alone, even if gd T cell proliferation didnot occur in these patients. Likewise, PBMC stimulation with ZAalone, which is not sufficient to induce gd T cell proliferation,yielded significantly lower Treg counts compared with mediumalone. ZA also exerted a functional inhibition of Treg activity, asshown by the experiments in which ZA was added to Tregs andautologous CD4+CD252 T cells stimulated by OKT3 in thepresence of irradiated accessory cells. No proliferative expansionof gd T cells was observed in these experiments because cellswere cultured for 3 d only, and IL-2 was not added to the cultures,further supporting the ability of ZA to counteract Treg activityin the absence of gd T cell proliferation. Moreover, Tregs werefunctionally impaired even when they were pretreated for 24 h andwashed free of ZA before mixing with autologous CD4+ CD252
T cells, suggesting that ZA can directly antagonize Tregs in theabsence of gd T cells. Interestingly, Sim treatment did not abro-gate this anti-Treg activity, indicating that FPPS inhibition andintracellular IPP accumulation are unlikely to play a role.We did not disclose major differences in Treg counts and sen-
sitivity to IL-2 stimulation between MM R and NR, though anexcess of Treg over gd T cells has been proposed as a mechanismdetermining the NR status of MM and other cancer patients (17).Any role of Treg in this setting is further complicated by thediscordant data about their counts and functional status in MM,because both increased numbers of dysfunctional Treg, and nor-mal numbers of immune-competent cells have been reported (46).An unexpected finding was PD-L2 downregulation on mDCZA+
and f-DCZA+. Tumor cells and pathogens exploit the PD-1/PD-L
1588 IMMUNOMODULATION BY ZOLEDRONIC ACID IN MM

pathway to evade host immunity, and several in vitro and in vivodata indicate that downregulation of the PD-1/PD-L pathway maybe a profitable way of breaking tolerance and restoring effectiveAg-specific immune responses (47). Most studies have focused onthe inhibition of PD-L1 solely by means of blocking Abs. To thebest of our knowledge, this is the first report of pharmacologicaldownmodulation of PD-L2 ever reported. Like Treg antagonism,this downregulation was not abrogated by Sim, indicating that ZAhas immune modulatory activities not solely dependent on itsability to induce intracellular IPP accumulation and gd T cellproliferation. It has recently been reported that the antiangiogenicactivity of ZA can also be independent from its ability to target theMev pathway (48).In conclusion, we have deciphered a virtuous cross-talk between
gd T cells, Tregs, DC, and CD8 ab T cells that is fostered by theproliferative activation of gd T cells initiated by the large amountsof IPP generated by ZA-treated mDC and f-DC. Ancillary to IPPgeneration are the ZA’s abilities to antagonize Tregs and down-regulate PD-L2 on DC. These ancillary activities are independentfrom Mev pathway inhibition and gd T cell proliferation. Alto-gether, these immunomodulatory events can reinstate gd T cellproliferation and upregulate SRV-specific immune responses inMM patients irrespective of their initial NR/R status. These datamay help us to further understand the role of gd T cells as cellularbridges between innate and adaptive immunity, and to fine-tunethe development of gd T cell-based adoptive immunotherapystrategies in MM and other diseases.
AcknowledgmentsWe are indebted to Emilia Sormano from the Blood Bank of Azienda Ospe-
daliera San Giovanni Battista, Turin, Italy, for providing buffy coats from
healthy donors.
DisclosuresM.M. received research grant support and scientific adviser board and lec-
ture honoraria from Novartis Farma S.p.A. (Origgio, Italy). The other
authors have no financial conflicts of interest.
References1. Beetz, S., D. Wesch, L. Marischen, S. Welte, H. H. Oberg, and D. Kabelitz.
2008. Innate immune functions of human gammadelta T cells. Immunobiology
213: 173–182.2. Tanaka, Y., C. T. Morita, Y. Tanaka, E. Nieves, M. B. Brenner, and B. R. Bloom.
1995. Natural and synthetic non-peptide antigens recognized by human gamma
delta T cells. Nature 375: 155–158.3. Jomaa, H., J. Feurle, K. Luhs, V. Kunzmann, H. P. Tony, M. Herderich, and
M. Wilhelm. 1999. Vgamma9/Vdelta2 T cell activation induced by bacterial low
molecular mass compounds depends on the 1-deoxy-D-xylulose 5-phosphatepathway of isoprenoid biosynthesis. FEMS Immunol. Med. Microbiol. 25: 371–
378.4. Espinosa, E., C. Belmant, F. Pont, B. Luciani, R. Poupot, F. Romagne, H. Brailly,
M. Bonneville, and J. J. Fournie. 2001. Chemical synthesis and biological ac-tivity of bromohydrin pyrophosphate, a potent stimulator of human gamma delta
T cells. J. Biol. Chem. 276: 18337–18344.5. Bonneville, M., and E. Scotet. 2006. Human Vgamma9Vdelta2 T cells: prom-
ising new leads for immunotherapy of infections and tumors. Curr. Opin.
Immunol. 18: 539–546.6. Gober, H. J., M. Kistowska, L. Angman, P. Jeno, L. Mori, and G. De Libero.
2003. Human T cell receptor gammadelta cells recognize endogenous mevalo-
nate metabolites in tumor cells. J. Exp. Med. 197: 163–168.7. Roelofs, A. J., K. Thompson, S. Gordon, and M. J. Rogers. 2006. Molecular
mechanisms of action of bisphosphonates: current status. Clin. Cancer Res. 12:6222s–6230s.
8. Monkkonen, H., P. D. Ottewell, J. Kuokkanen, J. Monkkonen, S. Auriola, and
I. Holen. 2007. Zoledronic acid-induced IPP/ApppI production in vivo. Life Sci.81: 1066–1070.
9. Mariani, S., M. Muraro, F. Pantaleoni, F. Fiore, B. Nuschak, S. Peola,M. Foglietta, A. Palumbo, M. Coscia, B. Castella, et al. 2005. Effector gam-
madelta T cells and tumor cells as immune targets of zoledronic acid in multiplemyeloma. Leukemia 19: 664–670.
10. Roelofs, A. J., M. Jauhiainen, H. Monkkonen, M. J. Rogers, J. Monkkonen, andK. Thompson. 2009. Peripheral blood monocytes are responsible for gamma-delta T cell activation induced by zoledronic acid through accumulation of IPP/DMAPP. Br. J. Haematol. 144: 245–250.
11. Fiore, F., B. Castella, B. Nuschak, R. Bertieri, S. Mariani, B. Bruno,F. Pantaleoni, M. Foglietta, M. Boccadoro, and M. Massaia. 2007. Enhancedability of dendritic cells to stimulate innate and adaptive immunity on short-termincubation with zoledronic acid. Blood 110: 921–927.
12. Conti, L., R. Casetti, M. Cardone, B. Varano, A. Martino, F. Belardelli, F. Poccia,and S. Gessani. 2005. Reciprocal activating interaction between dendritic cellsand pamidronate-stimulated gammadelta T cells: role of CD86 and inflammatorycytokines. J. Immunol. 174: 252–260.
13. Devilder, M.-C., S. Maillet, I. Bouyge-Moreau, E. Donnadieu, M. Bonneville,and E. Scotet. 2006. Potentiation of antigen-stimulated V g 9V d 2 T cell cy-tokine production by immature dendritic cells (DC) and reciprocal effect on DCmaturation. J. Immunol. 176: 1386–1393.
14. Takahara, M., M. Miyai, M. Tomiyama, M. Mutou, A. J. Nicol, and M. Nieda.2008. Copulsing tumor antigen-pulsed dendritic cells with zoledronate effi-ciently enhance the expansion of tumor antigen-specific CD8+ T cells viaVgamma9gammadelta T cell activation. J. Leukoc. Biol. 83: 742–754.
15. Kunzmann, V., E. Bauer, J. Feurle, F. Weissinger, H. P. Tony, and M. Wilhelm.2000. Stimulation of gammadelta T cells by aminobisphosphonates and in-duction of antiplasma cell activity in multiple myeloma. Blood 96: 384–392.
16. Wilhelm, M., V. Kunzmann, S. Eckstein, P. Reimer, F. Weissinger, T. Ruediger,and H. P. Tony. 2003. Gammadelta T cells for immune therapy of patients withlymphoid malignancies. Blood 102: 200–206.
17. Kunzmann, V., B. Kimmel, T. Herrmann, H. Einsele, and M. Wilhelm. 2009.Inhibition of phosphoantigen-mediated gammadelta T-cell proliferation by CD4+ CD25+ FoxP3+ regulatory T cells. Immunology 126: 256–267.
18. Brown, R. D., B. Pope, A. Murray, W. Esdale, D. M. Sze, J. Gibson, P. J. Ho,D. Hart, and D. Joshua. 2001. Dendritic cells from patients with myeloma arenumerically normal but functionally defective as they fail to up-regulate CD80(B7-1) expression after huCD40LT stimulation because of inhibition by trans-forming growth factor-beta1 and interleukin-10. Blood 98: 2992–2998.
19. Ratta, M., F. Fagnoni, A. Curti, R. Vescovini, P. Sansoni, B. Oliviero, M. Fogli,E. Ferri, G. R. Della Cuna, S. Tura, et al. 2002. Dendritic cells are functionallydefective in multiple myeloma: the role of interleukin-6. Blood 100: 230–237.
20. Cabillic, F., O. Toutirais, V. Lavoue, C. T. de La Pintiere, P. Daniel, N. Rioux-Leclerc, B. Turlin, H. Monkkonen, J. Monkkonen, K. Boudjema, et al. 2010.Aminobisphosphonate-pretreated dendritic cells trigger successful Vgamma9V-delta2 T cell amplification for immunotherapy in advanced cancer patients.Cancer Immunol. Immunother. 59: 1611–1619.
21. Banerjee, D. K., M. V. Dhodapkar, E. Matayeva, R. M. Steinman, andK. M. Dhodapkar. 2006. Expansion of FOXP3high regulatory T cells by humandendritic cells (DCs) in vitro and after injection of cytokine-matured DCs inmyeloma patients. Blood 108: 2655–2661.
22. Fiore, F., B. Nuschak, S. Peola, S. Mariani, M. Muraro, M. Foglietta, M. Coscia,B. Bruno, M. Boccadoro, and M. Massaia. 2005. Exposure to myeloma celllysates affects the immune competence of dendritic cells and favors the in-duction of Tr1-like regulatory T cells. Eur. J. Immunol. 35: 1155–1163.
23. Grube, M., S. Moritz, E. C. Obermann, K. Rezvani, A. Mackensen,R. Andreesen, and E. Holler. 2007. CD8+ T cells reactive to survivin antigen inpatients with multiple myeloma. Clin. Cancer Res. 13: 1053–1060.
24. Dauer, M., B. Obermaier, J. Herten, C. Haerle, K. Pohl, S. Rothenfusser,M. Schnurr, S. Endres, and A. Eigler. 2003. Mature dendritic cells derived fromhuman monocytes within 48 hours: a novel strategy for dendritic cell differen-tiation from blood precursors. J. Immunol. 170: 4069–4076.
25. Mariani, S., S. Y. Hwang, M. Foglietta, L. Bonello, C. Vitale, M. Coscia,F. Fiore, B. Bruno, and M. Massaia. 2009. Comprehensive assessment of theTCRBV repertoire in small T-cell samples by means of an improved and con-venient multiplex PCR method. Exp. Hematol. 37: 728–738.
26. Seifert, S. C., and J. J. Lucas. 1988. Incorporation of mevalonate into dolicholand other isoprenoids during estrogen-induced chick oviduct differentiation.Biochim. Biophys. Acta 962: 16–24.
27. Benford, H. L., J. C. Frith, S. Auriola, J. Monkkonen, and M. J. Rogers. 1999.Farnesol and geranylgeraniol prevent activation of caspases by aminobi-sphosphonates: biochemical evidence for two distinct pharmacological classes ofbisphosphonate drugs. Mol. Pharmacol. 56: 131–140.
28. Kuniyasu, Y., T. Takahashi, M. Itoh, J. Shimizu, G. Toda, and S. Sakaguchi.2000. Naturally anergic and suppressive CD25(+)CD4(+) T cells as a function-ally and phenotypically distinct immunoregulatory T cell subpopulation. Int.Immunol. 12: 1145–1155.
29. Buhaescu, I., and H. Izzedine. 2007. Mevalonate pathway: a review of clinicaland therapeutical implications. Clin. Biochem. 40: 575–584.
30. Thompson, K., and M. J. Rogers. 2004. Statins prevent bisphosphonate-inducedgamma,delta-T-cell proliferation and activation in vitro. J. Bone Miner. Res. 19:278–288.
31. Mahan, C. S., J. J. Thomas, W. H. Boom, and R. E. Rojas. 2009. CD4+ CD25(high) Foxp3+ regulatory T cells downregulate human Vdelta2+ T-lymphocytefunction triggered by anti-CD3 or phosphoantigen. Immunology 127: 398–407.
32. Wan, Y. Y., and R. A. Flavell. 2006. The roles for cytokines in the generation andmaintenance of regulatory T cells. Immunol. Rev. 212: 114–130.
33. Klebanoff, C. A., L. Gattinoni, P. Torabi-Parizi, K. Kerstann, A. R. Cardones,S. E. Finkelstein, D. C. Palmer, P. A. Antony, S. T. Hwang, S. A. Rosenberg,et al. 2005. Central memory self/tumor-reactive CD8+ T cells confer superiorantitumor immunity compared with effector memory T cells. Proc. Natl. Acad.Sci. USA 102: 9571–9576.
The Journal of Immunology 1589

34. Gattinoni, L., C. A. Klebanoff, D. C. Palmer, C. Wrzesinski, K. Kerstann, Z. Yu,S. E. Finkelstein, M. R. Theoret, S. A. Rosenberg, and N. P. Restifo. 2005.Acquisition of full effector function in vitro paradoxically impairs the in vivoantitumor efficacy of adoptively transferred CD8+ T cells. J. Clin. Invest. 115:1616–1626.
35. Dieli, F., F. Poccia, M. Lipp, G. Sireci, N. Caccamo, C. Di Sano, and A. Salerno.2003. Differentiation of effector/memory Vdelta2 T cells and migratory routes inlymph nodes or inflammatory sites. J. Exp. Med. 198: 391–397.
36. Parekh, V. V., S. Lalani, S. Kim, R. Halder, M. Azuma, H. Yagita, V. Kumar,L. Wu, and L. V. Kaer. 2009. PD-1/PD-L blockade prevents anergy induction andenhances the anti-tumor activities of glycolipid-activated invariant NKT cells. J.Immunol. 182: 2816–2826.
37. Blank, C., and A. Mackensen. 2007. Contribution of the PD-L1/PD-1 pathway toT-cell exhaustion: an update on implications for chronic infections and tumorevasion. Cancer Immunol. Immunother. 56: 739–745.
38. Romagnoli, M., V. Trichet, C. David, M. Clement, P. Moreau, R. Bataille, andS. Barille-Nion. 2007. Significant impact of survivin on myeloma cell growth.Leukemia 21: 1070–1078.
39. Andersen, M. H., L. O. Pedersen, J. C. Becker, and P. T. Straten. 2001. Identi-fication of a cytotoxic T lymphocyte response to the apoptosis inhibitor proteinsurvivin in cancer patients. Cancer Res. 61: 869–872.
40. Wobser, M., P. Keikavoussi, V. Kunzmann, M. Weininger, M. H. Andersen, andJ. C. Becker. 2006. Complete remission of liver metastasis of pancreatic cancerunder vaccination with a HLA-A2 restricted peptide derived from the universaltumor antigen survivin. Cancer Immunol. Immunother. 55: 1294–1298.
41. Andersen, M. H., I. M. Svane, J. C. Becker, and P. T. Straten. 2007. The universalcharacter of the tumor-associated antigen survivin. Clin. Cancer Res. 13: 5991–5994.
42. Dauer, M., K. Schad, J. Herten, J. Junkmann, C. Bauer, R. Kiefl, S. Endres, andA. Eigler. 2005. FastDC derived from human monocytes within 48 h effectivelyprime tumor antigen-specific cytotoxic T cells. J. Immunol. Methods 302: 145–155.
43. Jarnjak-Jankovic, S., H. Hammerstad, S. Saebøe-Larssen, G. Kvalheim, andG. Gaudernack. 2007. A full scale comparative study of methods for generationof functional dendritic cells for use as cancer vaccines. BMC Cancer 7: 119–125.
44. Gao, Y., W. Yang, M. Pan, E. Scully, M. Girardi, L. H. Augenlicht, J. Craft, andZ. Yin. 2003. Gamma delta T cells provide an early source of interferon gammain tumor immunity. J. Exp. Med. 198: 433–442.
45. Gong, G., L. Shao, Y. Wang, C. Y. Chen, D. Huang, S. Yao, X. Zhan, H. Sicard,R. Wang, and Z. W. Chen. 2009. Phosphoantigen-activated V gamma 2V delta 2T cells antagonize IL-2-induced CD4+CD25+Foxp3+ T regulatory cells inmycobacterial infection. Blood 113: 837–845.
46. Joshua, D. E., R. D. Brown, P. J. Ho, and J. Gibson. 2008. Regulatory T cells andmultiple myeloma. Clin. Lymphoma Myeloma 8: 283–286.
47. Keir, M. E., M. J. Butte, G. J. Freeman, and A. H. Sharpe. 2008. PD-1 and itsligands in tolerance and immunity. Annu. Rev. Immunol. 26: 677–704.
48. Stresing, V., P. G. Fournier, A. Bellahcene, I. Benzaıd, H. Monkkonen,M. Colombel, F. H. Ebetino, V. Castronovo, and P. Clezardin. 2011. Nitrogen-containing bisphosphonates can inhibit angiogenesis in vivo without the in-volvement of farnesyl pyrophosphate synthase. Bone 48: 259–266.
1590 IMMUNOMODULATION BY ZOLEDRONIC ACID IN MM
Related Documents











