Molecular Microbiology (2004) 51(5), 1413–1423 doi:10.1046/j.1365-2958.2004.03914.x © 2004 Blackwell Publishing Ltd Blackwell Science, LtdOxford, UKMMIMolecular Microbiology1365-2958Blackwell Publishing Ltd, 200351 514131423Original ArticleKluyveromyces lactis chromosomal replicatorC. Irene et al. Accepted 24 October, 2003. *For correspondence. E-mail [email protected]; Tel. (+39) 6 4991 2243; Fax (+39) 6 4991 2351. Identification of the sequences required for chromosomal replicator function in Kluyveromyces lactis Carmela Irene, 1 Clelia Maciariello, 1 Francesco Cioci, 2 Giorgio Camilloni, 2 Carol S. Newlon 3 and Lucia Fabiani 1 * 1 Dipartimento Biologia Cellulare e dello Sviluppo and 2 Dipartimento Genetica e Biologia Molecolare, Università ‘La Sapienza’, Piazzale A. Moro, 5, Roma, Italy. 3 Department of Microbiology and Molecular Genetics, UMDNJ–New Jersey Medical School, Newark, NJ 07103, USA. Summary The analysis of replication intermediates of a Kluyver- omyces lactis chromosomal autonomous replicating sequence (ARS), KARS101, has shown that it is active as a chromosomal replicator. KARS101 contains a 50 bp sequence conserved in two other K. lactis ARS elements. The deletion of the conserved sequence in KARS101 completely abolished replicator activity, in both the plasmids and the chromosome. Gel shift assays indicated that this sequence binds proteins present in K. lactis nuclear extracts, and a 40 bp sequence, previously defined as the core essential for K. lactis ARS function, is required for efficient bind- ing. Reminiscent of the origin replication complex (ORC), the binding appears to be ATP dependent. A similar pattern of protection of the core was seen with in vitro footprinting. KARS101 also functions as an ARS sequence in Saccharomyces cerevisiae . A com- parative study using S. cerevisiae nuclear extracts revealed that the sequence required for binding is a dodecanucleotide related to the S. cerevisiae ARS consensus sequence and essential for S. cerevisiae ARS activity. Introduction The replicator sequences required for eukaryotic origins were first identified in Saccharomyces cerevisiae as chro- mosomal DNA fragments capable of promoting high- frequency transformation and the autonomous replication of plasmids. They are called autonomously replicating sequence (ARS) elements. They are modular in structure, consisting of an essential domain A and elements in the B and C domains that stimulate ARS activity (reviewed by Newlon and Theis, 1993; Newlon, 1996). Domain A con- tains an 11 bp consensus sequence, the ARS consensus sequence (ACS), that acts as the core of the recognition sequence for the replication initiator protein, the six-sub- unit origin recognition complex (ORC). The ACS and ele- ment B1, localized 3 ¢ to the T-rich strand of the ACS, contribute to ORC binding. ORC serves as the ‘landing pad’ for the recruitment of proteins involved in the regula- tion of the initiation of DNA replication (reviewed by Bell and Dutta, 2002). Saccharomyces cerevisiae is an excellent model for eukaryotic DNA replication, but its replicators appear to be different from replicators in other eukaryotes. These replicators are larger, varying from a few hundred base- pairs in some unicellular organisms, e.g. Schizosaccharo- myces pombe , Physarum polycephalum and Yarrowia lipolytica (Bénard et al ., 1996; Dubey et al ., 1996; Vernis et al ., 1999), up to several kilobasepairs in mammals (Aladjem et al ., 1998). The analysis of S. pombe replica- tors revealed that they are 0.5–1.5 kb in length and are composed of multiple redundant elements important for ARS function (Kim and Huberman, 1998; 1999; Okuno et al ., 1999). Recent results indicate that at least part of the redundancy results from the presence of multiple binding sites for ORC (Kong and DePamphilis, 2002; Takahashi et al ., 2003), making S. pombe replicators anal- ogous to S. cerevisiae compound replicators, which con- tain multiple matches to the ACS, each of which must be altered to abolish replicator activity (Theis and Newlon, 2001). Replication initiation events in some mammalian replication origins appear to be distributed through large ‘initiation zones’ (reviewed by DePamphilis, 1999). Despite the differences in origin structure among different organisms, the conservation of replication initiation pro- teins, including ORC, from yeast to mammals suggests a common feature in origin structure (reviewed by Bogan et al ., 2000). The yeast Kluyveromyces lactis provides a good system for the study of chromosomal replicators (Fabiani et al .,

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular Microbiology (2004)
51
(5), 1413–1423 doi:10.1046/j.1365-2958.2004.03914.x
© 2004 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology1365-2958Blackwell Publishing Ltd, 200351
514131423
Original Article
Kluyveromyces lactis chromosomal replicatorC. Irene
et al.
Accepted 24 October, 2003. *For correspondence. [email protected]; Tel. (+39) 6 4991 2243; Fax (+39) 6 49912351.
Identification of the sequences required for chromosomal replicator function in
Kluyveromyces lactis
Carmela Irene,
1
Clelia Maciariello,
1
Francesco Cioci,
2
Giorgio Camilloni,
2
Carol S. Newlon
3
andLucia Fabiani
1
*
1
Dipartimento Biologia Cellulare e dello Sviluppo and
2
Dipartimento Genetica e Biologia Molecolare, Università ‘La Sapienza’, Piazzale A. Moro, 5, Roma, Italy.
3
Department of Microbiology and Molecular Genetics, UMDNJ–New Jersey Medical School, Newark, NJ 07103, USA.
Summary
The analysis of replication intermediates of a
Kluyver-omyces lactis
chromosomal autonomous replicatingsequence (ARS),
KARS101
, has shown that it is activeas a chromosomal replicator.
KARS101
contains a50 bp sequence conserved in two other
K. lactis
ARSelements. The deletion of the conserved sequence in
KARS101
completely abolished replicator activity, inboth the plasmids and the chromosome. Gel shiftassays indicated that this sequence binds proteinspresent in
K. lactis
nuclear extracts, and a 40 bpsequence, previously defined as the core essential for
K. lactis
ARS function, is required for efficient bind-ing. Reminiscent of the origin replication complex(ORC), the binding appears to be ATP dependent. Asimilar pattern of protection of the core was seen with
in vitro
footprinting.
KARS101
also functions as anARS sequence in
Saccharomyces cerevisiae
. A com-parative study using
S. cerevisiae
nuclear extractsrevealed that the sequence required for binding is adodecanucleotide related to the
S. cerevisiae
ARSconsensus sequence and essential for
S. cerevisiae
ARS activity.
Introduction
The replicator sequences required for eukaryotic originswere first identified in
Saccharomyces cerevisiae
as chro-mosomal DNA fragments capable of promoting high-
frequency transformation and the autonomous replicationof plasmids. They are called autonomously replicatingsequence (ARS) elements. They are modular in structure,consisting of an essential domain A and elements in theB and C domains that stimulate ARS activity (reviewed byNewlon and Theis, 1993; Newlon, 1996). Domain A con-tains an 11 bp consensus sequence, the ARS consensussequence (ACS), that acts as the core of the recognitionsequence for the replication initiator protein, the six-sub-unit origin recognition complex (ORC). The ACS and ele-ment B1, localized 3
¢
to the T-rich strand of the ACS,contribute to ORC binding. ORC serves as the ‘landingpad’ for the recruitment of proteins involved in the regula-tion of the initiation of DNA replication (reviewed by Belland Dutta, 2002).
Saccharomyces cerevisiae
is an excellent model foreukaryotic DNA replication, but its replicators appear tobe different from replicators in other eukaryotes. Thesereplicators are larger, varying from a few hundred base-pairs in some unicellular organisms, e.g.
Schizosaccharo-myces pombe
,
Physarum polycephalum
and
Yarrowialipolytica
(Bénard
et al
., 1996; Dubey
et al
., 1996; Vernis
et al
., 1999), up to several kilobasepairs in mammals(Aladjem
et al
., 1998). The analysis of
S. pombe
replica-tors revealed that they are 0.5–1.5 kb in length and arecomposed of multiple redundant elements important forARS function (Kim and Huberman, 1998; 1999; Okuno
et al
., 1999). Recent results indicate that at least part ofthe redundancy results from the presence of multiplebinding sites for ORC (Kong and DePamphilis, 2002;Takahashi
et al
., 2003), making
S. pombe
replicators anal-ogous to
S. cerevisiae
compound replicators, which con-tain multiple matches to the ACS, each of which must bealtered to abolish replicator activity (Theis and Newlon,2001). Replication initiation events in some mammalianreplication origins appear to be distributed throughlarge ‘initiation zones’ (reviewed by DePamphilis, 1999).Despite the differences in origin structure among differentorganisms, the conservation of replication initiation pro-teins, including ORC, from yeast to mammals suggests acommon feature in origin structure (reviewed by Bogan
et al
., 2000).The yeast
Kluyveromyces lactis
provides a good systemfor the study of chromosomal replicators (Fabiani
et al
.,
1414
C. Irene
et al.
© 2004 Blackwell Publishing Ltd,
Molecular Microbiology
,
51
, 1413–1423
1990). We showed previously that a 103 bp fragment con-taining
KARS101
was a replicator in both
K. lactis
and
S.cerevisiae
. This fragment contains a 12 bp sequence thatappears to function as the essential ACS in
S. cerevisiae
,but it only contributes to ARS activity in
K. lactis
. Furtheranalysis of
KARS101
revealed that the essential core of
KARS101
is 40 bp in length, at least twice as large as theessential core of
S. cerevisiae
ARS elements, and thatdifferent sequences are required for ARS function in thetwo organisms (Fabiani
et al
., 1996). We recently identi-fied in the circular plasmid
pKD1
, which is able to trans-form
K. lactis
and
S. cerevisiae
, a 159 bp sequence thatpromotes autonomous replication in both yeasts. Thisfragment contains a sequence related to the
S. cerevisiae
ACS and a region of 53% identity to
KARS101
essentialcore. Examination of
pKD1
replication intermediates bytwo-dimensional electrophoresis revealed a correlationbetween the ARS element and the origin of DNA replica-tion (Fabiani
et al
., 2001).In this paper, we analyse
KARS101
replication interme-diates to examine its role as an origin of DNA replicationin its chromosomal context. In addition, we define the
cis
-acting sequences required for
K. lactis
origin function bysubstitution at the chromosomal level of the
KARS101
wild-type sequence with a mutated copy having a com-plete deletion of the core previously shown to be essentialfor
K. lactis
ARS function (Fabiani
et al
., 1996). We alsodemonstrate, by
in vitro
analysis, that the sequenceessential for origin function is the binding site for protein(s)that are likely to be involved in the initiation of DNAreplication.
Results
KARS101
is an efficient chromosomal origin of DNA replication
Kluyveromyces lactis
ARS sequence,
KARS101
, is anefficient replicator in plasmids (Fabiani
et al
., 1990; 1996).
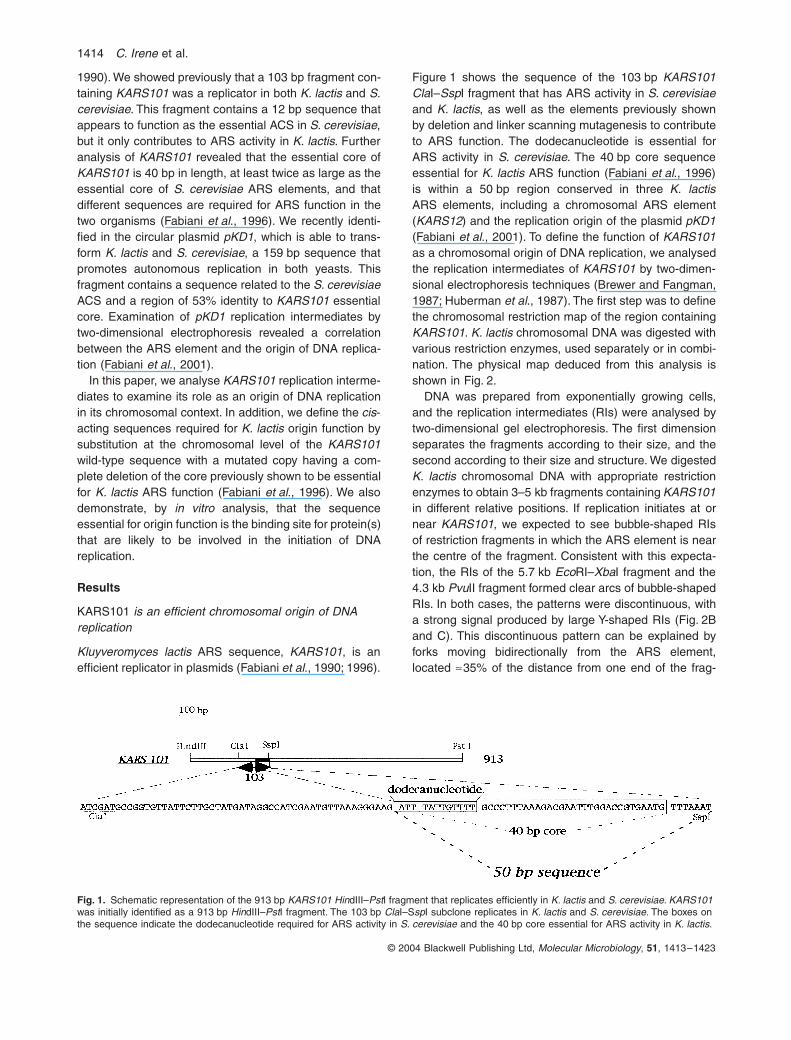
Figure 1 shows the sequence of the 103 bp
KARS101Cla
I–
Ssp
I fragment that has ARS activity in
S. cerevisiae
and
K. lactis
, as well as the elements previously shownby deletion and linker scanning mutagenesis to contributeto ARS function. The dodecanucleotide is essential forARS activity in
S. cerevisiae
. The 40 bp core sequenceessential for
K. lactis
ARS function (Fabiani
et al
., 1996)is within a 50 bp region conserved in three
K. lactis
ARS elements, including a chromosomal ARS element(
KARS12
) and the replication origin of the plasmid
pKD1
(Fabiani
et al
., 2001). To define the function of
KARS101
as a chromosomal origin of DNA replication, we analysedthe replication intermediates of
KARS101
by two-dimen-sional electrophoresis techniques (Brewer and Fangman,1987; Huberman
et al
., 1987). The first step was to definethe chromosomal restriction map of the region containing
KARS101
.
K. lactis
chromosomal DNA was digested withvarious restriction enzymes, used separately or in combi-nation. The physical map deduced from this analysis isshown in Fig. 2.
DNA was prepared from exponentially growing cells,and the replication intermediates (RIs) were analysed bytwo-dimensional gel electrophoresis. The first dimensionseparates the fragments according to their size, and thesecond according to their size and structure. We digested
K. lactis
chromosomal DNA with appropriate restrictionenzymes to obtain 3–5 kb fragments containing
KARS101
in different relative positions. If replication initiates at ornear
KARS101
, we expected to see bubble-shaped RIsof restriction fragments in which the ARS element is nearthe centre of the fragment. Consistent with this expecta-tion, the RIs of the 5.7 kb
Eco
RI–
Xba
I fragment and the4.3 kb
Pvu
II fragment formed clear arcs of bubble-shapedRIs. In both cases, the patterns were discontinuous, witha strong signal produced by large Y-shaped RIs (Fig. 2Band C). This discontinuous pattern can be explained byforks moving bidirectionally from the ARS element,located
ª
35% of the distance from one end of the frag-
Fig. 1.
Schematic representation of the 913 bp
KARS101 Hin
dIII–
Pst
I fragment that replicates efficiently in
K. lactis
and
S. cerevisiae. KARS101
was initially identified as a 913 bp
Hin
dIII–
Pst
I fragment. The 103 bp
Cla
I–S
sp
I subclone replicates in
K. lactis
and
S. cerevisiae
. The boxes on the sequence indicate the dodecanucleotide required for ARS activity in
S. cerevisiae
and the 40 bp core essential for ARS activity in
K. lactis
.

Kluyveromyces lactis
chromosomal replicator
1415
© 2004 Blackwell Publishing Ltd,
Molecular Microbiology
,
51
, 1413–1423
ment to the other. If the two forks move at equal rates,then RIs should be bubble-shaped for the first 70% ofreplication and Y-shaped for the remainder. These twopatterns and others not shown suggest that
KARS101
isan efficient origin of DNA replication. The pattern of Y-shaped RIs seen in the analysis of the 4.8 kb
Eco
RI
–Xho
Ifragment, which has
KARS101
close to one end, providedfurther support for the idea that replication initiates at ornear
KARS101
(Fig. 2D). In this case, the RIs areexpected to be predominantly Y-shaped because replica-tion initiates so close to the end of the fragment.
Essential role of the core sequence in KARS101origin function
After the demonstration that KARS101 was an efficientorigin of DNA replication in its chromosomal location, wewished to define the sequences that are essential for K.lactis replicator function. We deleted from the chromo-some a 50 bp sequence containing the essential core ofKARS101 to test its role in replicator activity (see Fig. 1).
For this purpose, we isolated from a K. lactis genomicbank a plasmid, pCC10, carrying a KARS101 fragment ofabout 6 kb (Fig. 3A) that included the EcoRI and XbaIsites shown on the map in Fig. 2. This fragment was largeenough to facilitate the homologous recombination nec-essary for the substitution of the wild-type KARS101sequence with the deletion mutant.
We inserted a selectable marker, the S. cerevisiaeURA3 gene, in the XhoI site at a distance of 0.7 kb fromKARS101 (Fig. 3A). To test whether the URA3 insertioncompromised plasmid ARS activity, we subcloned theKARS101 HindIII–BglII fragments with and without theURA3 insertion in the centromere-containing plasmid,pLF1. Both plasmids were able to transform K. lactisefficiently. Therefore, the insertion of URA3 close toKARS101 does not affect its ARS activity (Fig. 3B).
Fig. 2. Chromosomal map and two-dimensional gel analysis of rep-lication intermediates of KARS101. The restriction map of the chro-mosomal region containing KARS101 is shown at the top. The map was defined by Southern analysis of genomic DNA, using the 913 bp KARS101 HindIII–PstI fragment as probe. The box represents the 103 bp sequence, and the black box inside represents the essential core. DNA for two-dimensional analysis was isolated as described in Experimental procedures. The first dimension was run in 0.4% aga-rose gel at 1.2 V cm-1 for 28 h, and the second dimension was in a 1% agarose gel at 5 V cm-1 for 10 h.A. Schematic diagram showing the replication intermediates detected after hybridization.B, C and D. The two-dimensional patterns obtained after digestion with EcoRI–XbaI (B), PvuII (C) and EcoRI–XhoI (D). The blots were probed with the 913 bp HindIII–PstI fragment described above.
Fig. 3. Isolation of the KARS101 genomic sequence and effect of the insertion of the URA3 gene on replication activity of KARS101.A. The restriction map of KARS101 genomic fragment present in the plasmid pCC10 identified by colony hybridization. The S. cerevisiae URA3 gene was inserted in the XhoI site.B. The ARS phenotypes of the 1.3 and 2.4 kb HindIII–BglII fragments without and with URA3.C. The two-dimensional pattern obtained after digestion with PvuII of chromosomal DNA isolated from the K. lactis strain carrying the URA3 insertion near KARS101. The gel was run and probed as described in the legend to Fig. 2.

1416 C. Irene et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1413–1423
To test whether the URA3 insertion affected the activityof KARS101 at the chromosomal level, we transformed K.lactis with the 5.0 kb PvuII genomic fragment with theURA3 gene inserted in the XhoI site (Fig. 3C). We usedSouthern analysis to identify a K. lactis transformantcarrying URA3 at the correct position, and analysedKARS101 replication intermediates by two-dimensionalgel electrophoresis (Fig. 3C). The two-dimensional pat-tern obtained shows the typical bubble arc, indicating thatthe KARS101 PvuII fragment is replicated by an internalactive origin. Therefore, the URA3 insertion does not com-promise KARS101 origin function at the chromosomallevel.
Finally, we constructed a plasmid, pLF101, in which the50 bp region of homology shown in Fig. 1 was deletedfrom the URA3-marked KARS101 as described in Exper-imental procedures (Fig. 4A). This plasmid was unable totransform K. lactis at high frequency, demonstrating theessential role of the 50 bp sequence in plasmid ARSactivity. The 5.0 kb PvuII fragment obtained from this plas-mid was used to transform K. lactis. We used Southernanalysis to identify a transformant strain with theKARS101 deletion. Figure 4B shows the two-dimensionalagarose gel pattern obtained by analysis of the mutantstrain. Deletion of the 50 bp sequence containing theKARS101 core abolished the activity of the replicator, asshown by the absence, after a long exposure, of bubble-shaped RIs. The RIs detected were Y-shaped, indicatingthat the mutant fragment is replicated by an external origin(Fig. 4B, left). To show that the failure to detect a replica-tion bubble arc in the KARS101 deletion fragment was nota result of loss of replication bubbles from the DNA prep-aration, we analysed the replication intermediates ofanother K. lactis chromosomal origin (KARS12), and areplication bubble was readily detected (Fig. 4B, right).
These results demonstrate that sequences essential forchromosomal replicator activity are contained within the50 bp region containing the core sequence previouslyshown to be essential for K. lactis ARS activity. In addition,
they show that KARS12 is active as a chromosomal rep-lication origin.
Role of the core in protein binding using K. lactisnuclear extract
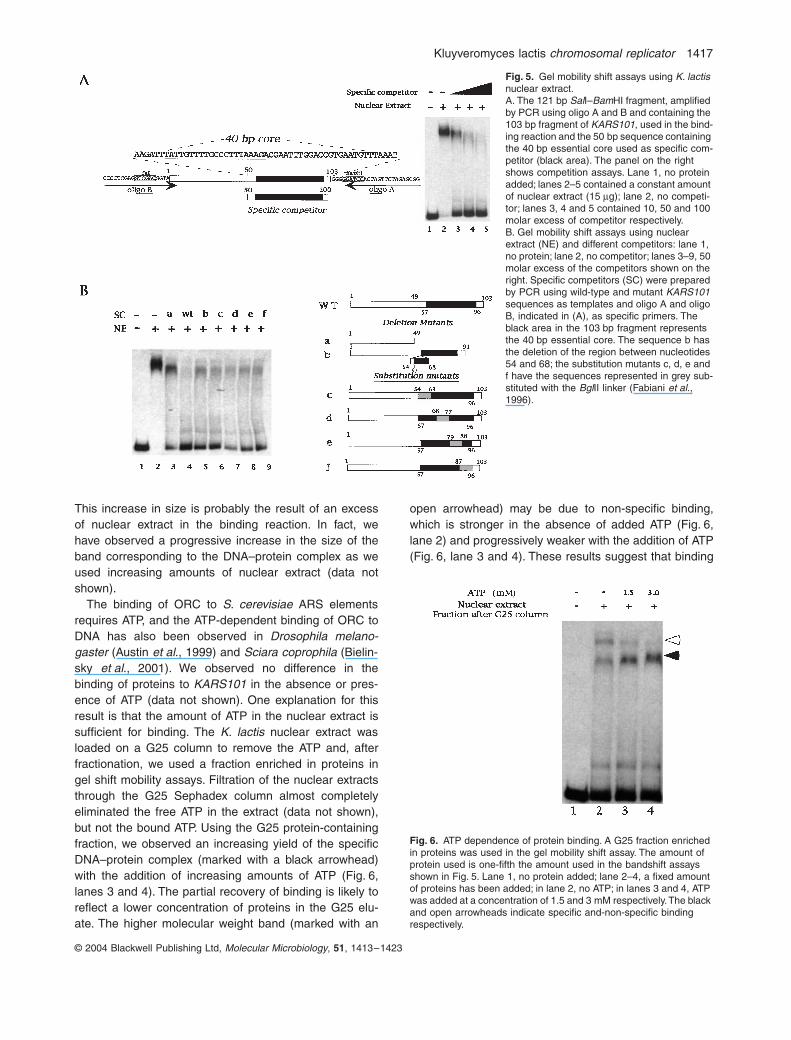
We hypothesized that the KARS101 core could representthe binding site of a protein complex essential for theinitiation of DNA replication in K. lactis. To test this idea,we prepared a nuclear extract from exponentially growingK. lactis cells and analysed, by gel retardation assays,protein binding to the 103 bp KARS101 ClaI–SspI frag-ment. Using increasing amounts of nuclear extract, con-taining up to 20 mg of protein, we found that the additionof 10 mg of protein was sufficient to shift completely theband corresponding to 0.5 ng of the KARS101 fragment.Figure 5A demonstrates that the 50 bp fragment contain-ing the essential core of KARS101 competes with thebinding. An excess of 100-fold (Fig. 5A, lane 5) wasrequired to abrogate the binding completely. This resultwas confirmed using as DNA substrate two reassociatedsynthetic oligos corresponding to the sequence used ascompetitor.
To define better the sequence important for proteinbinding, we used as competitors KARS101 substitutionmutant sequences previously shown to eliminateKARS101 activity in plasmids (Fabiani et al., 1996)(Fig. 5B). Only the wild-type sequence was able to com-pete efficiently for binding (Fig. 5B, lane 4); no competitionwas observed using the sequence between nucleotides 1and 49 (Fig. 5B, lane 3), and only a faint competition wasobserved using a fragment with a complete deletion of thedodecanucleotide (Fig. 5B, lane 5) or different linker sub-stitution mutations (Fig. 5B, lanes 6–9). Therefore, theintact core sequence is required for efficient binding.These results were confirmed using the mutatedsequences as probes.
In Fig. 5A and B, the complex is larger in the laneslacking competitor DNA (lanes 2) than in other lanes.
Fig. 4. Map of the KARS101 core deletion mutant and two-dimensional analysis of the mutant K. lactis strain.A. Map of the fragment used to replace the chromosomal copy of KARS101.B. The two-dimensional patterns obtained after PvuII digestion of chromosomal DNA isolated from the mutant K. lactis strain carrying a com-plete deletion of the 40 bp core sequence and URA3 as selectable marker. The filter was first probed with the 913 bp KARS101 fragment described in the legend to Fig. 2 (pattern on the left) and then with the 594 bp PstI–BglII frag-ment containing KARS12 (pattern on the right). The white boxes along the fragment represent KARS101 and KARS12; the black box inside KARS12 represents the 50 bp sequence with homology to the KARS101 core sequence.
Kluyveromyces lactis chromosomal replicator 1417
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1413–1423
This increase in size is probably the result of an excessof nuclear extract in the binding reaction. In fact, wehave observed a progressive increase in the size of theband corresponding to the DNA–protein complex as weused increasing amounts of nuclear extract (data notshown).
The binding of ORC to S. cerevisiae ARS elementsrequires ATP, and the ATP-dependent binding of ORC toDNA has also been observed in Drosophila melano-gaster (Austin et al., 1999) and Sciara coprophila (Bielin-sky et al., 2001). We observed no difference in thebinding of proteins to KARS101 in the absence or pres-ence of ATP (data not shown). One explanation for thisresult is that the amount of ATP in the nuclear extract issufficient for binding. The K. lactis nuclear extract wasloaded on a G25 column to remove the ATP and, afterfractionation, we used a fraction enriched in proteins ingel shift mobility assays. Filtration of the nuclear extractsthrough the G25 Sephadex column almost completelyeliminated the free ATP in the extract (data not shown),but not the bound ATP. Using the G25 protein-containingfraction, we observed an increasing yield of the specificDNA–protein complex (marked with a black arrowhead)with the addition of increasing amounts of ATP (Fig. 6,lanes 3 and 4). The partial recovery of binding is likely toreflect a lower concentration of proteins in the G25 elu-ate. The higher molecular weight band (marked with an
open arrowhead) may be due to non-specific binding,which is stronger in the absence of added ATP (Fig. 6,lane 2) and progressively weaker with the addition of ATP(Fig. 6, lane 3 and 4). These results suggest that binding
Fig. 5. Gel mobility shift assays using K. lactis nuclear extract.A. The 121 bp SalI–BamHI fragment, amplified by PCR using oligo A and B and containing the 103 bp fragment of KARS101, used in the bind-ing reaction and the 50 bp sequence containing the 40 bp essential core used as specific com-petitor (black area). The panel on the right shows competition assays. Lane 1, no protein added; lanes 2–5 contained a constant amount of nuclear extract (15 mg); lane 2, no competi-tor; lanes 3, 4 and 5 contained 10, 50 and 100 molar excess of competitor respectively.B. Gel mobility shift assays using nuclear extract (NE) and different competitors: lane 1, no protein; lane 2, no competitor; lanes 3–9, 50 molar excess of the competitors shown on the right. Specific competitors (SC) were prepared by PCR using wild-type and mutant KARS101 sequences as templates and oligo A and oligo B, indicated in (A), as specific primers. The black area in the 103 bp fragment represents the 40 bp essential core. The sequence b has the deletion of the region between nucleotides 54 and 68; the substitution mutants c, d, e and f have the sequences represented in grey sub-stituted with the BglII linker (Fabiani et al., 1996).
Fig. 6. ATP dependence of protein binding. A G25 fraction enriched in proteins was used in the gel mobility shift assay. The amount of protein used is one-fifth the amount used in the bandshift assays shown in Fig. 5. Lane 1, no protein added; lane 2–4, a fixed amount of proteins has been added; in lane 2, no ATP; in lanes 3 and 4, ATP was added at a concentration of 1.5 and 3 mM respectively. The black and open arrowheads indicate specific and-non-specific binding respectively.
1418 C. Irene et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1413–1423
of proteins in the nuclear extract to KARS101 requiresATP.
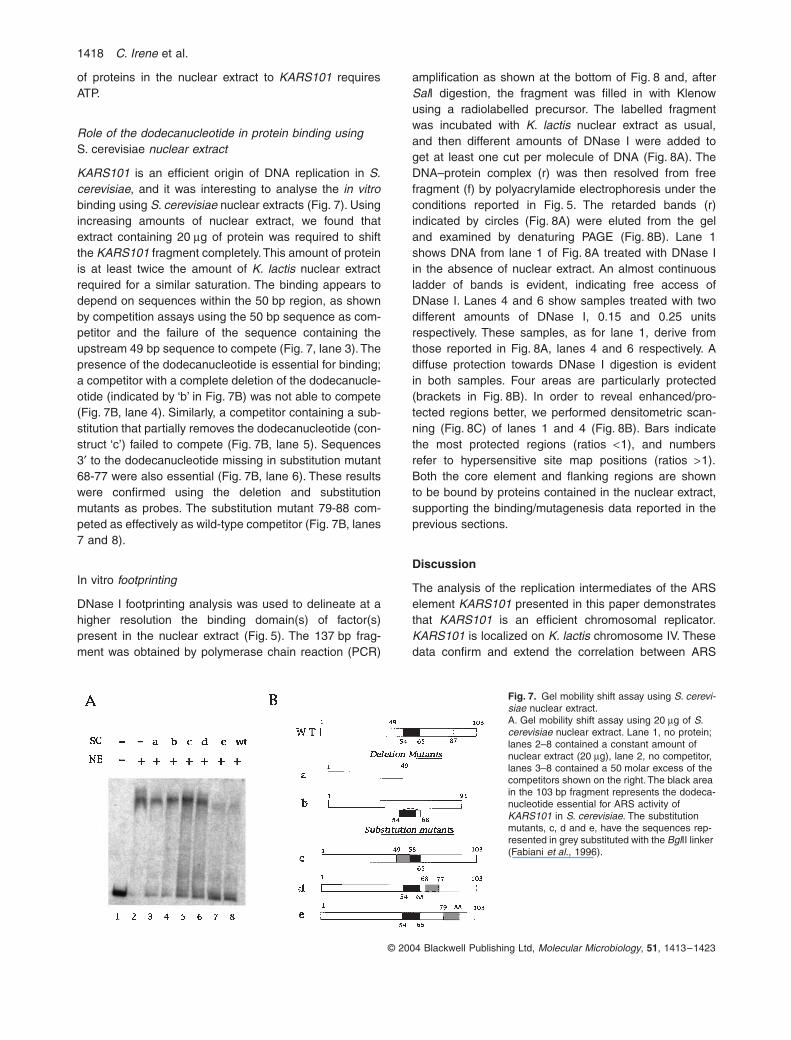
Role of the dodecanucleotide in protein binding usingS. cerevisiae nuclear extract
KARS101 is an efficient origin of DNA replication in S.cerevisiae, and it was interesting to analyse the in vitrobinding using S. cerevisiae nuclear extracts (Fig. 7). Usingincreasing amounts of nuclear extract, we found thatextract containing 20 mg of protein was required to shiftthe KARS101 fragment completely. This amount of proteinis at least twice the amount of K. lactis nuclear extractrequired for a similar saturation. The binding appears todepend on sequences within the 50 bp region, as shownby competition assays using the 50 bp sequence as com-petitor and the failure of the sequence containing theupstream 49 bp sequence to compete (Fig. 7, lane 3). Thepresence of the dodecanucleotide is essential for binding;a competitor with a complete deletion of the dodecanucle-otide (indicated by ‘b’ in Fig. 7B) was not able to compete(Fig. 7B, lane 4). Similarly, a competitor containing a sub-stitution that partially removes the dodecanucleotide (con-struct ‘c’) failed to compete (Fig. 7B, lane 5). Sequences3¢ to the dodecanucleotide missing in substitution mutant68-77 were also essential (Fig. 7B, lane 6). These resultswere confirmed using the deletion and substitutionmutants as probes. The substitution mutant 79-88 com-peted as effectively as wild-type competitor (Fig. 7B, lanes7 and 8).
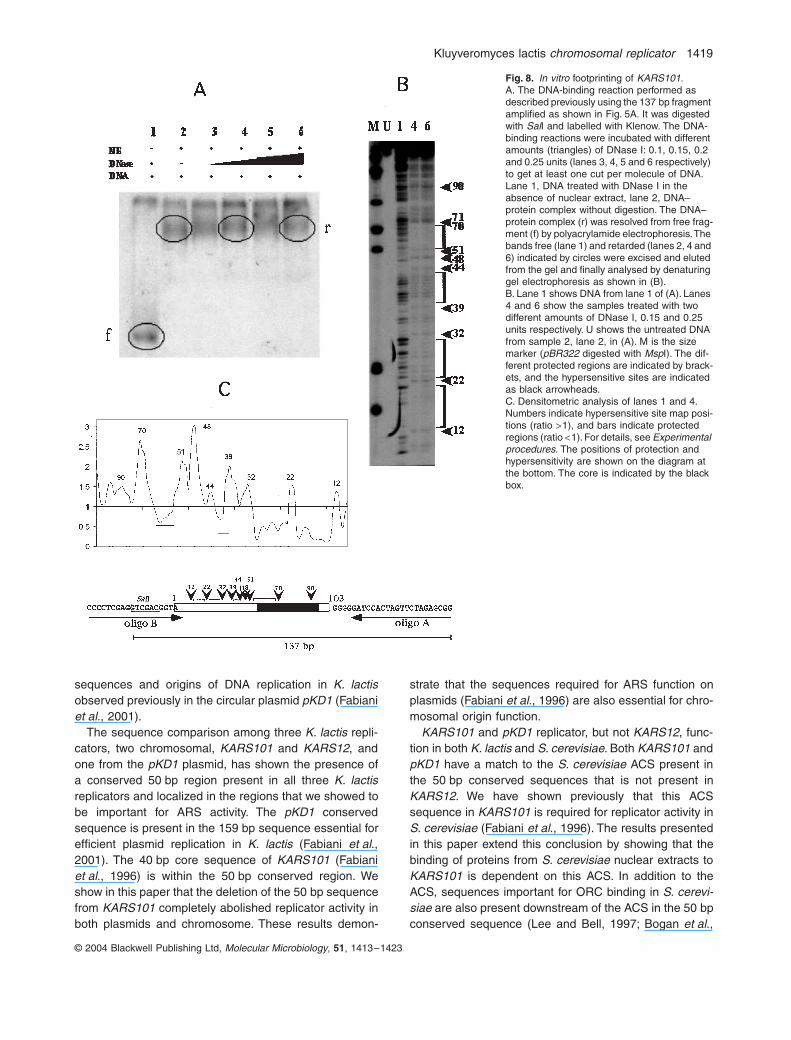
In vitro footprinting
DNase I footprinting analysis was used to delineate at ahigher resolution the binding domain(s) of factor(s)present in the nuclear extract (Fig. 5). The 137 bp frag-ment was obtained by polymerase chain reaction (PCR)
amplification as shown at the bottom of Fig. 8 and, afterSalI digestion, the fragment was filled in with Klenowusing a radiolabelled precursor. The labelled fragmentwas incubated with K. lactis nuclear extract as usual,and then different amounts of DNase I were added toget at least one cut per molecule of DNA (Fig. 8A). TheDNA–protein complex (r) was then resolved from freefragment (f) by polyacrylamide electrophoresis under theconditions reported in Fig. 5. The retarded bands (r)indicated by circles (Fig. 8A) were eluted from the geland examined by denaturing PAGE (Fig. 8B). Lane 1shows DNA from lane 1 of Fig. 8A treated with DNase Iin the absence of nuclear extract. An almost continuousladder of bands is evident, indicating free access ofDNase I. Lanes 4 and 6 show samples treated with twodifferent amounts of DNase I, 0.15 and 0.25 unitsrespectively. These samples, as for lane 1, derive fromthose reported in Fig. 8A, lanes 4 and 6 respectively. Adiffuse protection towards DNase I digestion is evidentin both samples. Four areas are particularly protected(brackets in Fig. 8B). In order to reveal enhanced/pro-tected regions better, we performed densitometric scan-ning (Fig. 8C) of lanes 1 and 4 (Fig. 8B). Bars indicatethe most protected regions (ratios <1), and numbersrefer to hypersensitive site map positions (ratios >1).Both the core element and flanking regions are shownto be bound by proteins contained in the nuclear extract,supporting the binding/mutagenesis data reported in theprevious sections.
Discussion
The analysis of the replication intermediates of the ARSelement KARS101 presented in this paper demonstratesthat KARS101 is an efficient chromosomal replicator.KARS101 is localized on K. lactis chromosome IV. Thesedata confirm and extend the correlation between ARS
Fig. 7. Gel mobility shift assay using S. cerevi-siae nuclear extract.A. Gel mobility shift assay using 20 mg of S. cerevisiae nuclear extract. Lane 1, no protein; lanes 2–8 contained a constant amount of nuclear extract (20 mg), lane 2, no competitor, lanes 3–8 contained a 50 molar excess of the competitors shown on the right. The black area in the 103 bp fragment represents the dodeca-nucleotide essential for ARS activity of KARS101 in S. cerevisiae. The substitution mutants, c, d and e, have the sequences rep-resented in grey substituted with the BglII linker (Fabiani et al., 1996).
Kluyveromyces lactis chromosomal replicator 1419
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1413–1423
sequences and origins of DNA replication in K. lactisobserved previously in the circular plasmid pKD1 (Fabianiet al., 2001).
The sequence comparison among three K. lactis repli-cators, two chromosomal, KARS101 and KARS12, andone from the pKD1 plasmid, has shown the presence ofa conserved 50 bp region present in all three K. lactisreplicators and localized in the regions that we showed tobe important for ARS activity. The pKD1 conservedsequence is present in the 159 bp sequence essential forefficient plasmid replication in K. lactis (Fabiani et al.,2001). The 40 bp core sequence of KARS101 (Fabianiet al., 1996) is within the 50 bp conserved region. Weshow in this paper that the deletion of the 50 bp sequencefrom KARS101 completely abolished replicator activity inboth plasmids and chromosome. These results demon-
strate that the sequences required for ARS function onplasmids (Fabiani et al., 1996) are also essential for chro-mosomal origin function.
KARS101 and pKD1 replicator, but not KARS12, func-tion in both K. lactis and S. cerevisiae. Both KARS101 andpKD1 have a match to the S. cerevisiae ACS present inthe 50 bp conserved sequences that is not present inKARS12. We have shown previously that this ACSsequence in KARS101 is required for replicator activity inS. cerevisiae (Fabiani et al., 1996). The results presentedin this paper extend this conclusion by showing that thebinding of proteins from S. cerevisiae nuclear extracts toKARS101 is dependent on this ACS. In addition to theACS, sequences important for ORC binding in S. cerevi-siae are also present downstream of the ACS in the 50 bpconserved sequence (Lee and Bell, 1997; Bogan et al.,
Fig. 8. In vitro footprinting of KARS101.A. The DNA-binding reaction performed as described previously using the 137 bp fragment amplified as shown in Fig. 5A. It was digested with SalI and labelled with Klenow. The DNA-binding reactions were incubated with different amounts (triangles) of DNase I: 0.1, 0.15, 0.2 and 0.25 units (lanes 3, 4, 5 and 6 respectively) to get at least one cut per molecule of DNA. Lane 1, DNA treated with DNase I in the absence of nuclear extract, lane 2, DNA–protein complex without digestion. The DNA–protein complex (r) was resolved from free frag-ment (f) by polyacrylamide electrophoresis. The bands free (lane 1) and retarded (lanes 2, 4 and 6) indicated by circles were excised and eluted from the gel and finally analysed by denaturing gel electrophoresis as shown in (B).B. Lane 1 shows DNA from lane 1 of (A). Lanes 4 and 6 show the samples treated with two different amounts of DNase I, 0.15 and 0.25 units respectively. U shows the untreated DNA from sample 2, lane 2, in (A). M is the size marker (pBR322 digested with MspI). The dif-ferent protected regions are indicated by brack-ets, and the hypersensitive sites are indicated as black arrowheads.C. Densitometric analysis of lanes 1 and 4. Numbers indicate hypersensitive site map posi-tions (ratio >1), and bars indicate protected regions (ratio <1). For details, see Experimental procedures. The positions of protection and hypersensitivity are shown on the diagram at the bottom. The core is indicated by the black box.

1420 C. Irene et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1413–1423
2000). Linker substitution mutations in these sequencesabolished ARS activity in S. cerevisiae (Fabiani et al.,1996) and also diminished the ability of KARS101 to bindproteins from the S. cerevisiae nuclear extract. The pres-ence of these sequences explains the ability of these K.lactis replicators to function in S. cerevisiae, and the prox-imity of the essential sequences for the initiation of repli-cation in both K. lactis and S. cerevisiae may reflect animportant role played by adjacent regions promoting rep-licator activity in both organisms. These sequences maybe important for the recruitment of proteins involved in theinitiation of DNA replication.
The 50 bp sequences in all three K. lactis replicatorsare A+T rich, with an identity of 60% between KARS101and KARS12 and 50% between KARS101 and pKD1. Thehigher homology is limited, for all of them, to the regionimportant for origin function, and the more conservednucleotides are the As and Ts. They are characterized notby a specific conserved sequence but by repeats of threeor more consecutive adenines or thymines. The presenceof these repeats reflects a similarity with S. pombe ARSsequences, in which the presence of A and T stretches isa common feature among the different origins analysed.However, S. pombe replicator structure is more complex.S. pombe replicators appear to have a modular organiza-tion. They are much longer than the S. cerevisiae ARSelement, a few hundred basepairs, and do not have aconsensus sequence essential for replication. Replicatoractivity in S. pombe seems to depend not on a specificsequence, but on the number and length of A and Tstretches (Kim and Huberman, 1998; 1999; Okuno et al.,1999). It has been shown recently that S. pombe ORCinteracts with the adenine/thymine stretches (Takahashiand Masukata, 2001; Takahashi et al., 2003) through theN-terminal domain of the SpOrc4 protein (Kong andDePamphilis, 2001; 2002; Lee et al., 2001).
In addition to stretches rich in adenines and thymines,the 50 bp sequence important for KARS101 replicatoractivity also contains direct and inverted repeats. Invertedrepeat sequences are a common feature of prokaryoticand eukaryotic control regions including replication ori-gins. Inverted repeats (IR) have the potential to form cru-ciforms and stem–loop structures, and these structurescan influence chromatin structure. It has been shown pre-viously that histones and/or nucleosomes bind less effi-ciently to inverted repeats, stem–loop and cruciform DNA(reviewed by Pearson et al., 1996). These structures canhave different roles, a direct one by making a nucleosome-free DNA more accessible to DNA-binding proteins, or anindirect one by phasing nucleosomes in a replication-permissive manner. In a recent paper, Novac et al. (2002)showed a direct correlation between cruciform structureand binding of protein involved in the regulation of repli-cation initiation in mammalian cells.
We hypothesized that the 50 bp sequence essential forthe replicator activity of KARS101 could represent thebinding site of protein/s involved in the initiation of DNAreplication. In this paper, we show by gel retardation assaythat this sequence is required for efficient binding of pro-teins in K. lactis nuclear extracts. Using different mutantsequences as competitors, we showed that the sequencepreviously defined as the K. lactis core is required forefficient binding. These results provide support for theidea that the proteins interacting with the core sequenceare involved in replication initiation.
The K. lactis ORC homologue is a good candidate for thebinding activity. The K. lactis ORC1 gene was isolated andcompletely sequenced (Gavin et al., 1995). The protein is50% identical and 68% similar to S. cerevisiae Orc1p. Theregion conserved among Orc1p in diverse organisms iswithin the regions involved in ATP binding and hydrolysis.ORC binding requires ATP in different organisms, and weexamined this requirement in the binding properties ofKARS101. The results suggest that the binding of K. lactisnuclear proteins to KARS101 is ATP dependent.
The downstream sequences of the KARS101 essentialcore bind to proteins in vivo (data not shown) and areparticularly rich in A+T (about 80%) and, as was sug-gested for the B2 element in S. cerevisiae ARS307, theymay represent a DNA unwinding element (DUE) (Lin andKowalski, 1997). The AT richness could facilitate theunwinding of the sequence after ORC binding to allow thebinding of proteins important for the formation of the pre-replicative complex. It was shown recently by Wilmes andBell (2002) that the S. cerevisiae B2 element in ARS1 notonly has helically unstable DNA but also contains specificsequences related to the ACS that are important for pro-tein binding. The most likely candidates are the MCMproteins because their association with an easily unwoundregion of the origin is in agreement with their role as theputative eukaryotic replicative helicase. In a comparisonof the K. lactis core sequence with the downstreamsequences involved in protein binding in vivo, we detectedthe presence of sequences in direct and inverse orienta-tion that are related to the sequences within the core.Together, these observations support our use of the K.lactis replicator KARS101 as a model to understand therole of the sequences close to the essential core, whichmay be involved in the interaction of proteins not yetdiscovered and probably important for the regulation ofthe activity of chromosomal replicators that is currentlypoorly understood.
Experimental procedures
Strains and media
Kluyveromyces lactis strain MW98-8C (uraA, arg, lys, K+)was used for transformation by electroporation (Becker and

Kluyveromyces lactis chromosomal replicator 1421
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1413–1423
Guarente, 1991). Yeast strains were grown in YPD medium(1% yeast extract, 2% peptone, 2% glucose). YPD plateswere supplemented with 1.5% Difco agar. Yeast transfor-mants were selected on –Ura plates (0.67% Difco yeastnitrogen base without amino acids, 0.5% casamino acids,1 M sorbitol, 2% glucose and 1.5% Difco agar).
Escherichia coli strain DH5a was used for amplification ofplasmid DNA (Boyer and Roulland-Dussoix, 1969). E. colitransformants were grown in Luria broth supplemented with100 mg ml-1 ampicillin (Sambrook et al., 1989).
Kluyveromyces lactis centromeric plasmid pLF1
The 527 bp HinfI fragment, containing the essentialsequences of K. lactis centromere II (Heus et al., 1993), wascloned in the AatII site of the pRS306 plasmid to make pLF1.Both plasmid and insert were treated with T4 DNA poly-merase before ligation. Various K. lactis chromosomal frag-ments were cloned in the polylinker of pLF1 and assayed forARS activity in K. lactis (Becker and Guarente, 1991).
Isolation of replicating yeast DNA
Yeast cultures were grown to mid-log phase, mixed with 0.5volumes of an ice-cold slurry of azide stop solution (0.5 MNaOH, 0.4 M EDTA, 0.2% sodium azide, pH 8.65) (McCarrolland Fangman, 1988) and centrifuged immediately at 4∞C.The cells were then washed twice with sterile water. DNAwas prepared by the method of Huberman et al. (1987).
Restriction enzyme digestion and two-dimensional gel electrophoresis
DNA (10–20 mg) was digested as described by Greenfederand Newlon (1992). The DNA was ethanol precipitated andresuspend in 50 ml of TE (10 mM Tris and 1 mM EDTA,pH 8.0). The entire digest was loaded in one well of anagarose gel for two-dimensional analysis. Two-dimensionalgels were run as described by Brewer and Fangman (1987).
Southern blot analysis
DNA from two-dimensional gels was blotted to Nytran mem-branes (Schleicher and Schuell) according to the procedureof Smith and Summers (1980). The filters were hybridized at65∞C in 5¥ SSC, 50 mM sodium pyrophosphate, 5¥ Den-hardt’s solution, 0.5% SDS and 100 mg ml-1 calf thymus DNA.Probes, isolated by restriction enzyme digestion of plasmidsor amplified by PCR, were labelled with a Multiprime RandomPrimer DNA labelling kit (Amersham). Probe was added toDNA filters after prehybridization for 1 h in hybridization solu-tion. Blots were exposed to Kodak XR film with an intensifyingscreen at -80∞C.
Isolation of a KARS101 genomic fragment from a K. lactis genomic library
The K. lactis genomic bank, a generous gift from M. Weso-lowski, was prepared by cloning Sau3A restriction fragments
from K. lactis chromosomal DNA into the BamHI site of aderivative of the YIp5 plasmid (Sikorski and Hieter, 1989) withthe S11 fragment of the pKD1 plasmid (Chen et al., 1986)cloned in the EcoRI site. We amplified the library andscreened E. coli transformants by colony hybridization usingthe 913 bp HindIII–PstI fragment to identify a genomic frag-ment containing KARS101. Plasmid pCC10 carries aKARS101 genomic fragment of about 6 kb.
Construction of the KARS101 deletion plasmid pLF101
The S. cerevisiae URA3 gene, which complements the K.lactis uraA mutation, was isolated from the YDp-U plasmid(Berben et al., 1991) on a 1.1 kb BamHI fragment. The frag-ment was blunt ended with T4 polymerase and inserted as aselectable marker in the XhoI site (previously blunt ended withT4 polymerase) of the KARS101 genomic fragment present inthe plasmid pCC10. We digested the pCC10 plasmid withSspI, blunt ended with T4 polymerase and, after digestion withXbaI, we purified the 3.1 kb fragment carrying the URA3 geneinserted in the XhoI site. This fragment was cloned in aderivative of pLF1 carrying a 49 bp fragment of KARS101extending from the ClaI site to position 49 of the 103 bpsequence (see Fig. 1). This KARS101 fragment lacks theessential core sequence. To insert the URA3 fragment, thepLF1 derivative was first digested with BglII, which cuts in thepolylinker adjacent to the KARS101 fragment, blunt endedwith T4 polymerase and finally digested with XbaI. The derivedplasmid (with the complete core deletion, URA3 as a select-able marker and a long downstream flanking sequence) wasdigested with KpnI (present in the polylinker) and, after treat-ment with T4 polymerase, with ClaI to get compatible endsuseful for cloning the upstream 3.5 kb EcoRI–blunt ClaI frag-ment derived from KARS101 genomic fragment.
Preparation of nuclear extracts from yeast
Saccharomyces cerevisiae strain AH201 and K. lactis strainMW98-8C were grown in YPD to a density of 3.5 ¥ 107
cells ml-1, and the purification of nuclear extracts was carriedout by the method of Schneider et al. (1986).
DNA-binding assays
The DNA probes were labelled with DNA polymerase I, Kle-now fragment (New England Biolabs) and then isolated byPAGE. The competitor DNA in the DNA-binding reaction waseither the plasmid pRS306 or poly(d(AG))–poly(d(CT)) (Phar-macia). Results obtained with the two competitors were sim-ilar. The typical binding reaction mixture contained 25 mMTris-HCl (pH 7.8), 5 mM MgCl2, 0.8 mM ATP, 70 mM KCl,2 mg ml-1 BSA, 5 mM dithiothreitol, 5% glycerol, 0.1 mg ml-1
competitor DNA, 0.5 ng of labelled probe (10 000 c.p.m.),corresponding to the 121 bp SalI–BamHI fragment isolatedfrom the centromeric plasmid pLF11, which carries the103 bp KARS101 ClaI–SspI fragment cloned between theClaI and SmaI sites of the polylinker, and the indicatedamount of nuclear extract (Rao and Stillman, 1995). Theprotein–DNA binding reaction was incubated for 30 min at25∞C in 20 ml final volume, and then the reaction sample was

1422 C. Irene et al.
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1413–1423
loaded onto a 5% non-denaturing polyacrylamide gel in 0.25¥Tris–borate–EDTA (TBE) (acrylamide/bisacrylamide, 29:1).The gel was run for 3 h at a constant voltage of 200 V at 4∞C.The gel was dried and then autoradiographed.
The sequence between nucleotides 50 and 100 ofKARS101 was synthesized as complementary oligonucle-otides, which were annealed in vitro and used as specificcompetitor (Fig. 5A) and as probe in the bandshift reactions(data not shown).
The sequences used as competitors in Figs 5B and 6Bwere prepared using pRS306-derived plasmids as templatesfor PCR amplification of wild-type and mutant KARS101sequences, with the primers shown in Fig. 5A. The fragmentsamplified included the wild-type 103 bp ClaI–SspI KARS101fragment (pLF11) and deletion and substitution derivatives ofthis fragment (49-58, 54-63, 68-77, 79-88 and 87-96) (Fabi-ani et al., 1996) cloned in the ClaI–SmaI sites of thepolylinker. The PCR products used as competitors in thebinding reactions were purified on 2% agarose gels, andthose used as probes were purified on a 7% native polyacry-lamide gel (50:1 acrylamide/bisacrylamide in 0.5¥ TBE) afterdigestion with SalI and BamHI and fill in with Klenow.
In vitro footprinting
After the binding reaction described above, reaction mixtureswere chilled, and different amounts of DNase I (0.1, 0.15, 0.2and 0.25 units) were added. The incubation was carried outfor 1 min at 0∞C to get at least one cut per molecule of DNA.The digestion products were separated by electrophoresis onnative polyacrylamide gels, as described by Singh et al.(1986). After autoradiography, DNA was eluted overnight atroom temperature. The recovered DNA was analysed on 15%denaturing polyacrylamide gels. The DNase footprints weredetected by autoradiography.
Electrophoretic profiles of selected lanes were subjectedto densitometric scanning, and the value of each band inten-sity was normalized to the sum of the total intensity of thelane. The data are reported as the ratio, point by point,between profiles from in vitro DNase I-treated samples andDNase I-digested naked DNA.
Acknowledgements
We thank Jim Theis, Ida Ruberti, Paola Ballario, MicheleSaliola and the members of the Newlon laboratory group forhelpful discussions throughout the study. This work was sup-ported by MURST (Ministero Università Ricerca ScientificaTecnologica – Progetti Ateneo and Progetti Facoltà), IstitutoPasteur Fondazione Cenci Bolognetti and NIH (grant numberGM35679 to C.S.N.). L.F. was supported by a CNR short-term fellowship. C.I. is a PhD student on Cellular Biology,Università ‘La Sapienza’, Roma. C.M. is a PhD student onPasteurian Sciences, Università ‘La Sapienza’, Roma.
References
Aladjem, M.I., Rodewald, L.W., Kolman, J.L., and Wahl, G.M.(1998) Genetic dissection of a mammalian replicator in thehuman b-globin. Science 281: 1005–1009.
Austin, R.J., Orr-Weaver, T.L., and Bell, S.P. (1999)Drosophila ORC specifically binds to ACE3, an origin ofDNA replication control element. Genes Dev 13: 2639–2649.
Becker, D.M., and Guarente, L. (1991) High-efficiency trans-formation of yeast by electroporation. Methods Enzymol194: 182–187.
Bell, S.P., and Dutta, A. (2002) DNA replication in eukaryoticcells. Annu Rev Biochem 71: 333–374.
Bénard, M., Lagnel, C., Pallotta, D., and Pierron, G. (1996)Mapping of a replication origin within the promoter regionof two unlinked, abundantly transcribed actin genes ofPhysarum polycephalum. Mol Cell Biol 16: 968–976.
Berben, G., Dumont, J., Gilliquet, V., Bolle, P.A., and Hilger,F. (1991) The YDp plasmids: a uniform set of vectorsbearing versatile gene disruption cassettes for Saccharo-myces cerevisiae. Yeast 7: 475–477.
Bielinsky, A.K., Blitzblau, H., Beall, E.L., Ezrokhi, M., Smith,M.S., Botchan, M.R., and Gerbi, S.A. (2001) Origin repli-cation complex binding to a metazoan replication origin.Curr Biol 11: 1427–1431.
Bogan, J.A., Natale, D.A., and DePamphilis, M.L. (2000)Initiation of eukaryotic DNA replication: conservative orliberal?. J Cell Phys 184: 139–150.
Boyer, H.W., and Roulland-Dussoix, D. (1969) A complemen-tation analysis of the restriction and modification of DNAin Escherichia coli. J Mol Biol 41: 459–472.
Brewer, B.J., and Fangman, W.L. (1987) The localization ofreplication origins on ARS plasmids in S. cerevisiae. Cell51: 463–471.
Chen, X.J., Saliola, M., Falcone, C., Bianchi, M.M., and Fuku-hara, H. (1986) Sequence organization of the circular plas-mid pKD1 from the yeast Kluyveromyces drosophilarum.Nucleic Acids Res 14: 4471–4481.
DePamphilis, M.L. (1999) Replication origins in metazoanchromosomes: fact or fiction? Bioessays 21: 5–16.
Dubey, D.D., Kim, S.M., Todorov, I.T., and Huberman, J.A.(1996) Large, complex modular structure of a fission yeastDNA replication origin. Curr Biol 6: 467–473.
Fabiani, L., Aragona, M., and Frontali, L. (1990) Isolation andsequence analysis of a K. lactis chromosomal element ableto autonomously replicate in S. cerevisiae and K. lactis.Yeast 6: 69–76.
Fabiani, L., Frontali, L., and Newlon, C.S. (1996) Identifica-tion of an essential core element and stimulatory sequencein a Kluyveromyces lactis ARS element KARS101. MolMicrobiol 19: 757–766.
Fabiani, L., Irene, C., Aragona, M., and Newlon, C.S. (2001)A DNA replication origin and a replication fork barrier usedin vivo in the circular plasmid pKD1. Mol Genet Genomics266: 326–335.
Gavin, K.A., Hidaka, M., and Stillman, B. (1995) Conservedinitiator proteins in eucaryotes. Science 270: 1667–1671.
Greenfeder, S.A., and Newlon, C.S. (1992) A replication mapof a 61 kb circular derivative of Saccharomyces cerevisiaechromosome III. Mol Biol Cell 3: 473–481.
Heus, J.J., Zonneveld, B.J., de Steensma, H.Y., and van denBerg, J.A. (1993) The consensus sequence of Kluyvero-myces lactis centromeres shows homology to functionalcentromeric DNA from Saccharomyces cerevisiae. MolGen Genet 236: 355–362.

Kluyveromyces lactis chromosomal replicator 1423
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1413–1423
Huberman, J.A., Spotila, L.D., Nawotka, K.A., El-Assouli,S.M., and Davis, R.L. (1987) The in vivo replication originof the yeast 2m plasmid. Cell 51: 473–481.
Kim, S.M., and Huberman, J.A. (1998) Multiple orientation-dependent, synergistically interacting, similar domains inthe ribosomal DNA replication origin of the fission yeast,Schizosaccharomyces pombe. Mol Cell Biol 18: 7294–7303.
Kim, S.M., and Huberman, J.A. (1999) Influence of a repli-cation enhancer on the hierarchy of origin efficiencieswithin a cluster of DNA replication origins. J Mol Biol 288:867–882.
Kong, D., and DePamphilis, M.L. (2001) Site-specific DNAbinding of the Schizosaccharomyces pombe origin recog-nition complex is determined by the Orc4 subunit. Mol CellBiol 21: 8095–8103.
Kong, D., and DePamphilis, M.L. (2002) Site-specific ORCbinding, pre-replication complex assembly and DNA syn-thesis at Schizosaccharomyces pombe replication origins.EMBO J 21: 5567–5576.
Lee, D.G., and Bell, S.P. (1997) Architecture of the yeastorigin recognition complex bound to origins of DNA repli-cation. Mol Cell Biol 17: 7159–7168.
Lee, J.K., Moon, K.Y., Jiang, Y., and Hurwitz, J. (2001) TheSchizosaccharomyces pombe origin recognition complexinteracts with multiple AT-rich regions of the replicationorigin DNA by means of the AT-hook domains of thespOrc4 protein. Proc Natl Acad Sci USA 98: 13589–13594.
Lin, S., and Kowalski, D. (1997) Functional equivalency anddiversity of cis-acting elements among yeast replicationorigins. Mol Cell Biol 17: 5473–5484.
McCarrol, R.M., and Fangman, W.L. (1988) Time of replica-tion of yeast centromeres and telomeres. Cell 54: 505–513.
Newlon, C.S. (1996) DNA replication in yeast. In DNA Rep-lication in Eucaryotic Cells. De Pamphilis, M.L. (ed.). ColdSpring Harbor, NY: Cold Spring Harbor Laboratory Press,pp. 873–914.
Newlon, C.S., and Theis, J.F. (1993) The structure and func-tion of yeast ARS elements. Curr Opin Genet Dev 3: 752–758.
Novac, O., Alvarez, D., Pearson, C.E., Price, G.B., andZannis-Hadjopoulos, M. (2002) The human cruciform-binding protein, CBP, is involved in DNA replication andassociates in vivo with mammalian replication origin. J BiolChem 277: 11174–11183.
Okuno, Y., Satoh, H., Sekiguchi, M., and Masukata, H.(1999) Clustered adenine/thymine stretches are essential
for function of a fission yeast replication origin. Mol CellBiol 19: 6699–6709.
Pearson, C.E., Zorbas, H., Price, G.B., and Zannis-Hadjopoulos, M. (1996) Inverted repeats, stem-loops andcruciforms: significance for initiation of DNA replication. JCell Biochem 63: 1–22.
Rao, H., and Stillman, B. (1995) The origin recognition com-plex interacts with a bipartite DNA binding site within yeastreplicators. Proc Natl Acad Sci USA 92: 2224–2228.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) Molecu-lar Cloning: a Laboratory Manual, 2nd edn. Cold SpringHarbor, NY: Cold Spring Harbor Laboratory Press.
Schneider, R., Gander, I., Mueller, U., Mertz, R., and Win-nacker, E.L. (1986) A sensitive and rapid gel retentionassay for nuclear factor and other DNA-binding proteins incrude nuclear extracts. Nucleic Acids Res 14: 1303–1317.
Sikorski, R.S., and Hieter, P. (1989) A system of shuttlevectors and yeast host strains designed for efficient manip-ulation of DNA in Saccharomyces cerevisiae. Genetics122: 19–27.
Singh, H., Sen, R., Baltimore, D., and Sharp, A. (1986) Anuclear factor that binds to a conserved sequence motif intranscriptional control elements of immunoglobulin genes.Nature 319: 154–158.
Smith, G.E., and Summers, M.D. (1980) The bidirectionaltransfer of DNA and RNA to nitrocellulose or diazobenzy-loxymethyl-paper. Anal Biochem 109: 123–129.
Sor, F., and Fukuhara, H. (1989) Analysis of chromosomalDNA patterns of the genus Kluyveromyces. Yeast 5: 1–10.
Takahashi, T., and Masukata, H. (2001) Interaction of fissionyeast ORC with essential adenine/thymine stretches inreplication origins. Genes Cells 6: 837–849.
Takahashi, T, Ohara, E., Nishitani H., and Masukata, H.(2003) Multiple ORC-binding sites are required for efficientMCM loading and origin firing in fission yeast. EMBO J 22:964–974.
Theis, J.F., and Newlon, C.S. (2001) Two compound replica-tion origins in Saccharomyces cerevisiae contain redun-dant origin recognition complex binding sites. Mol Cell Biol21: 2790–2801.
Vernis, L., Chasles, M., Pasero, P., Lepingle, A., Gaillardin,C., and Fournier, P. (1999) Short DNA fragments withoutsequence similarity are sites for replication in the chromo-some of the yeast Yarrowia lipolytica. Mol Biol Cell 10:757–769.
Wilmes, G.M., and Bell, S.P. (2002) The B2 element of theSaccharomyces cerevisiae ARS1 origin of replicationrequires specific sequences to facilitate pre-RC formation.Proc Natl Acad Sci USA 99: 101–106.
Related Documents











