Copyright C Blackwell Munksgaard 2002 Traffic 2002; 3: 630–640 Blackwell Munksgaard ISSN 1398-9219 Identification of RanBP2- and Kinesin-Mediated Transport Pathways with Restricted Neuronal and Subcellular Localization Timur A. Mavlyutov, Yunfei Cai and Paulo A. Ferreira* Department of Pharmacology and Toxicology, Medical College of Wisconsin, 8701 Watertown Plank Road, Milwaukee, WI 53226, USA * Corresponding author: Paulo A. Ferreira, [email protected] Ran-binding proteins, karyopherins, and RanGTPase mediate and impart directionality to nucleocyto- plasmic transport processes. This biological process re- mains elusive in neurons. RanBP2 has been localized at the nuclear pore complexes and is very abundant in the neuroretina. RanBP2 mediates the assembly of a large complex comprising RanGTPase, CRM1/exportin- 1, importin-b, KIF5-motor proteins, components of the 19S cap of the 26S proteasome, ubc9 and opsin. Here, we show RanBP2 is abundant in the ellipsoid compart- ment of photoreceptors and RanGTPase-positive par- ticles in cytoplasmic tracks extending away from the nuclear envelope of subpopulations of ganglion cells, suggesting RanBP2’s release from nuclear pore com- plexes. KIF5C and KIF5B are specifically expressed in a subset of neuroretinal cells and differentially localize with RanBP2 and importin-b in distinct compartments. The C-terminal domains of KIF5B and KIF5C, but not KIF5A, associate directly with importin-b in a RanGTPase-dependent fashion in vivo and in vitro, in- dicating importin-b is an endogenous cargo for a subset of KIF5s in retinal neurons. The KIF5 transport pathway is absent from the myoid region of a topographically distinct subclass of blue cones and the distribution of kinesin-light chains is largely distinct from its KIF5 partners. Altogether, the results identify the existence of neuronal- and subtype-specific kinesin-mediated transport pathways of importin-b-bound cargoes to and/or from RanBP2 and indicate RanBP2 itself may also constitute a scaffold carrier for some of its associ- ated partners. The implications of these findings in pro- tein kinesis and pathogenesis of degenerative neuro- pathies are discussed. Key words: importin-b, karyopherin, kinesin, neurode- generation, nucleocytoplasmic transport, protein kin- esis, Ran-binding protein-2, RanBP2, Ran, opsin, retina Received 17 April 2002, revised and accepted for publi- cation 8 May 2002 Nucleocytoplasmic transport conveys a plethora of input stimuli to the nucleus and processed outputs to the cytosol 630 (1,2). Nuclear pore complexes (NPCs) represent the gates for intercompartmental communication between the nucleus and the cytosol. About 10 6 molecules/min cross the nuclear envelope (NE) of a growing cell (3). The mechanisms by which the pace and direction of this process are integrated with the cytosolic transport machinery remain obscure. Emerging evidence supports the existence of common com- ponents and cross-talk mechanisms, which harness nuclear transport and cytosolic secretory pathways (4,5). In purified NE of liver cells, RanBP2/Nup358 seems to be the primary component of fibrils emanating from the NPC (6). RanBP2 was implicated in mediating the initial steps of import of protein cargoes from the periphery of the NPC to its central channel. To this end, an antibody against RanBP2 inhibits nuclear protein import in HeLa cells (7). Furthermore, purified RanBP2 from NEs of liver nuclei associates with the import receptor, importin-b, in the presence of RanGTP (6). The association of this binary complex with RanGAP is re- quired for the translocation of importin-b and its cargo into the NPC channel, as antibodies to RanBP2-associated Ran- GAP inhibit nuclear protein import (8). RanBP2 is highly expressed in the retina (9,10). Systematic structure-function analysis of RanBP2 indicates its role in traf- ficking (and signaling) processes is far more complex, as re- vealed by the increasing number of proteins interacting specifically with selective domains of RanBP2 (Figure 1). To this end, the RBD4-CY domains of RanBP2 promote the functional production and chaperoning of red opsin in a CY- (PPIase)-dependent fashion (11,12). The cyclophilin like do- main (CLD) interacts with subunits of the 19S regulatory par- ticle of the proteasome(13). The species-variable zinc finger- rich domain associates with the nuclear export receptor, ex- portin-1/CRM1, in a leptomycin B-independent fashion (14). The domain between the RBD2 and RBD3, JX2, associates with the neuronal KIF5C and ubiquitous KIF5B motor proteins (and their kinesin–light chain partner/s) (15). Finally, Saitoh et al. revealed that the E2-ubiquitin-like conjugating enzyme, Ubc9, interacts with an internal repeat domain between the RBD4 and CLD domains in Xenopus egg extracts (16). Ubc9 conjugates SUMO-1 with RanGAP leading to SUMO-1– RanGAP association with RanBP2 (8,16–18). Amongst these interactions, the associations of KIF5s and the 19S particle components with RanBP2 occur in a neuronal-selective fashion (13,15,19). These data suggest RanBP2 moors speci- fic nuclear export and import transport steps with the cyto- solic trafficking of cargoes mediated by some kinesin-motor proteins in the neuroretina, a process likely modulated by

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Copyright C Blackwell Munksgaard 2002Traffic 2002; 3: 630–640
Blackwell Munksgaard ISSN 1398-9219
Identification of RanBP2- and Kinesin-MediatedTransport Pathways with Restricted Neuronal andSubcellular Localization
Timur A. Mavlyutov, Yunfei Cai andPaulo A. Ferreira*
Department of Pharmacology and Toxicology, Medical
College of Wisconsin, 8701 Watertown Plank Road,
Milwaukee, WI 53226, USA
* Corresponding author: Paulo A. Ferreira,
Ran-binding proteins, karyopherins, and RanGTPasemediate and impart directionality to nucleocyto-plasmic transport processes. This biological process re-mains elusive in neurons. RanBP2 has been localizedat the nuclear pore complexes and is very abundant inthe neuroretina. RanBP2 mediates the assembly of alarge complex comprising RanGTPase, CRM1/exportin-1, importin-b, KIF5-motor proteins, components of the19S cap of the 26S proteasome, ubc9 and opsin. Here,we show RanBP2 is abundant in the ellipsoid compart-ment of photoreceptors and RanGTPase-positive par-ticles in cytoplasmic tracks extending away from thenuclear envelope of subpopulations of ganglion cells,suggesting RanBP2’s release from nuclear pore com-plexes. KIF5C and KIF5B are specifically expressed ina subset of neuroretinal cells and differentially localizewith RanBP2 and importin-b in distinct compartments.The C-terminal domains of KIF5B and KIF5C, but notKIF5A, associate directly with importin-b in aRanGTPase-dependent fashion in vivo and in vitro, in-dicating importin-b is an endogenous cargo for a subsetof KIF5s in retinal neurons. The KIF5 transport pathwayis absent from the myoid region of a topographicallydistinct subclass of blue cones and the distribution ofkinesin-light chains is largely distinct from its KIF5partners. Altogether, the results identify the existenceof neuronal- and subtype-specific kinesin-mediatedtransport pathways of importin-b-bound cargoes toand/or from RanBP2 and indicate RanBP2 itself mayalso constitute a scaffold carrier for some of its associ-ated partners. The implications of these findings in pro-tein kinesis and pathogenesis of degenerative neuro-pathies are discussed.
Key words: importin-b, karyopherin, kinesin, neurode-generation, nucleocytoplasmic transport, protein kin-esis, Ran-binding protein-2, RanBP2, Ran, opsin, retina
Received 17 April 2002, revised and accepted for publi-cation 8 May 2002
Nucleocytoplasmic transport conveys a plethora of inputstimuli to the nucleus and processed outputs to the cytosol
630
(1,2). Nuclear pore complexes (NPCs) represent the gates forintercompartmental communication between the nucleusand the cytosol. About 106 molecules/min cross the nuclearenvelope (NE) of a growing cell (3). The mechanisms bywhich the pace and direction of this process are integratedwith the cytosolic transport machinery remain obscure.Emerging evidence supports the existence of common com-ponents and cross-talk mechanisms, which harness nucleartransport and cytosolic secretory pathways (4,5).
In purified NE of liver cells, RanBP2/Nup358 seems to bethe primary component of fibrils emanating from the NPC(6). RanBP2 was implicated in mediating the initial steps ofimport of protein cargoes from the periphery of the NPC toits central channel. To this end, an antibody against RanBP2inhibits nuclear protein import in HeLa cells (7). Furthermore,purified RanBP2 from NEs of liver nuclei associates with theimport receptor, importin-b, in the presence of RanGTP (6).The association of this binary complex with RanGAP is re-quired for the translocation of importin-b and its cargo intothe NPC channel, as antibodies to RanBP2-associated Ran-GAP inhibit nuclear protein import (8).
RanBP2 is highly expressed in the retina (9,10). Systematicstructure-function analysis of RanBP2 indicates its role in traf-ficking (and signaling) processes is far more complex, as re-vealed by the increasing number of proteins interactingspecifically with selective domains of RanBP2 (Figure 1). Tothis end, the RBD4-CY domains of RanBP2 promote thefunctional production and chaperoning of red opsin in a CY-(PPIase)-dependent fashion (11,12). The cyclophilin like do-main (CLD) interacts with subunits of the 19S regulatory par-ticle of the proteasome (13). The species-variable zinc finger-rich domain associates with the nuclear export receptor, ex-portin-1/CRM1, in a leptomycin B-independent fashion (14).The domain between the RBD2 and RBD3, JX2, associateswith the neuronal KIF5C and ubiquitous KIF5B motor proteins(and their kinesin–light chain partner/s) (15). Finally, Saitohet al. revealed that the E2-ubiquitin-like conjugating enzyme,Ubc9, interacts with an internal repeat domain between theRBD4 and CLD domains in Xenopus egg extracts (16). Ubc9conjugates SUMO-1 with RanGAP leading to SUMO-1–RanGAP association with RanBP2 (8,16–18). Amongst theseinteractions, the associations of KIF5s and the 19S particlecomponents with RanBP2 occur in a neuronal-selectivefashion (13,15,19). These data suggest RanBP2 moors speci-fic nuclear export and import transport steps with the cyto-solic trafficking of cargoes mediated by some kinesin-motorproteins in the neuroretina, a process likely modulated by

Neuroretinal Mapping of RanBP2 Macroassembly Complex
Figure1: Schematic diagram of RanBP2 and its interacting protein partners. RanBP2 is comprised of several structural modulessuch as four interspersed RanGTPase binding domains (RBDn Ω 1ª4), a variable and species-specific zinc-finger cluster domain (ZnF),leucine-rich (LD), cyclophilin (CY), cyclophilin-like (CLD), junction-2 (JX2) and an internal repeat domain (W1W2). Arrows point to proteinsthat interact specifically with selective domains of RanBP2. Legend: KIF5C/KIF5B, microtubule-based motor proteins (kinesins) KIF5B andKIF5C; ropsin, red (L/M) opsin; CRM1/exportin-1, nuclear export sequence (NES) export receptor; Ubc9, E2-SUMO-1 conjugating enzyme;Imp-b, nuclear import receptor, importin-b. See introduction for details.
other RanBP2 partners. Here, we determine the spatial re-lationships between RanBP2 and some of its partners, anddefine neuronal-restricted transport pathways and cognatecargo mediated by RanBP2, KIF5B and KIF5C in the neurore-tina.
Results
Neuronal organization and lamination of the retina
To orient the reader in interpreting the images and findingsreported, we first provide a brief description of the neuronalorganization of the mammalian retina (Figure 2). The ver-tebrate retina is a well-defined neuronal and stratified tissuethat lines the posterior eyecup. It is comprised of a diversepopulation of neuronal cells with distinctive morphologicaland electrophysiological features. These cells are organizedin three major (nuclear) layers (Figure 2A). The photon (light)-capturing neurosensory cells, photoreceptors, consist of avariable population of rods and, red, green, and blue cones,each expressing a unique subclass of light-receptors. Photo-receptors are highly polarized and compartmentalized neu-rons with several distinct subcellular compartments (Figure2B). The outer segment, which contains all the matured pho-ton (light)-capturing receptors (opsins) and light transductionmachinery components, is linked by a non-motile connectingcilium to the inner segment compartment containing a polar-ized distribution of cell organelles. The apical portion of thiscompartment, ellipsoid, is very rich in mitochondria and post-Golgi trafficking components, while the basal compartment,the myoid, contains most of the biosynthetic machinery andassociated organelles (ER/Golgi). Variable lengths of outerand inner fibers connect the inner segment to the nucleus(positioned at different depths in the outer nuclear layer ofthe retina) and this to the synaptic terminal ends of photore-ceptor neurons, respectively. Photoreceptors synapse at theouter plexiform layer with some second-order neurons of theretina, comprising diverse classes of horizontal and bipolarneurons, and the latter are modulated by the interneurons,amacrine cells (Figure 2A). The nuclei of the second-orderneurons form the inner nuclear layer and are topographicallypositioned within this layer in a neuronal class-specific
631Traffic 2002; 3: 630–640
fashion. These neurons synapse at the inner plexiform layerwith a diverse population of ganglion cells (Figure 2A). Thebundles of axons emanating from these neurons form thenerve fiber layer, which synapse distantly with neurons in thethalamus region of the brain. We have employed the retinalcircuitry as a model system to dissect trafficking processesin neuronal cells in light of the neuronal variety, the distinctcellular architectures, the considerable distances cargoesmust travel in these cells and the large number of inheriteddegenerative neuropathies associated with retinal dysfunc-tion.
Distribution of RanBP2 and RanGTPase in the bovine
retina
The destruction of the spatial relationships by the homogen-ization methods previously employed (9–15), the tissue-re-strictive binding properties of RanBP2 with various partnersand the existence of a diverse neuronal population in the ret-ina prompted us to investigate first the cellular and subcellul-ar expression and distribution of RanBP2 and some of itspartners in the neuroretina. As described previously, selectivedomains of RanBP2 mediate the association with specificmolecular partners (Figure 1). We have shown that RanBP2and domains thereof associate strongly with RanGTP in reti-nal extracts (11,14,15). Antibodies against RanBP2 detected,in general, a broad cellular expression of the cognate proteinin bovine retinas. RanBP2 localization at distinct punctate lociaround the periphery of nuclei in all the three nuclear layers[outer nuclear layer (ONL), inner nuclear layer (INL) and gan-glion cell layer; Figure 3A,B] is consistent with the fen-estration of NE by nuclear pore complexes and attachmentof RanBP2 at cytoplasmic fibrils emanating from the NPC(Figure 3A,C) (6,7,20,21). However, photoreceptors and someganglion cells exhibited a striking and distinctive distributionpattern of RanBP2 (Figure 3C). RanBP2 was very abundantin the apical (ellipsoid) compartment of the inner segmentsof cones and rod photoreceptors (Figure 3A–C). RanBP2 wassignificantly more abundant at the NE of nuclei at the distaledge (first row of nuclei) of the ONL (Figure 3A–C, and Fig-ure 4). Moreover, its presence was conspicuously absentfrom the myoid compartment (Figure 3A–C), which com-prises the space between the uppermost row of nuclei of the

Mavlyutov et al.
Figure2: Neuronal organization and lamination of the mammalian retina and morphology of photoreceptors. A. The mammalianretina is a well-organized and stratified neuronal tissue comprising a variety of classes of neurons (see Results for details). The neurosensorycells, the photoreceptors, consist of rods (gray color) and red, green and blue cones (in red, green and blue color). These synapse at theouter plexiform layer (OPL) with horizontal cells (H, light blue) and rod (RB, yellow) and cone (CB, purple) bipolar cells, whose cell bodiesare located in the inner nuclear layer. The amacrine cells (A, green) located at the proximal edge of the inner nuclear layer (INL) modulatethe synaptic input of bipolar cells with the ganglion cells (orange). These establish synaptic contact at the inner plexiform layer (IPL) withcells located in the inner nuclear layer. The axons of ganglion cells coalesce and form the nerve fiber layer (NFL) in the retina. These synapsewith neurons in the thalamus (brain). B. Rod and cone photoreceptors are highly polarized and compartmentalized cells. They consist ofouter segments containing all the photoreception and phototransduction components, and these are connected to the inner segments by anon-motile connecting cilium (CC). The organelles in the inner segments have a polarized distribution. The ellipsoid is rich in mitochondriaand post-Golgi-derived vesicles, and the biosynthetic machinery is contained mostly in the myoid compartment. The nuclear compartmentis connected to the inner segment and synaptic terminal, respectively, by outer and inner fibers of variable lengths. Legend: OS, outersegments; IS, inner segments; ONL, outer nuclear layer; OPL, outer plexiform layer, INL, inner nuclear layer; IPL, inner plexiform layer; GCL,ganglion cell layer; NFL, nerve fiber layer; CC, connecting cilium.
ONL and ellipsoid region of all photoreceptor inner segments,with the exception of a subpopulation of blue cone photore-ceptors as presented below.
RanGTPase was localized in the intranuclear and perinuclearcompartments of retinal cells. However, RanGTPase was alsovery abundant throughout the myoid and ellipsoid photore-ceptor compartments and excluded from the outer segments(Figure 3D,E). Its presence was also evident at the synapticendings of the outer plexiform layer (OPL) and nerve fiberlayer (NFL) (Figure 3D). The relative abundance of Ran, andin particular, RanBP2, among retinal neurons was remarkable.In a population of large ganglion cells, RanBP2 and Ran wereextremely abundant and formed a continuous nuclear rim(Figure 3G–J) due to the extremely high abundance of NPCsat the NE, as shown by transmission electron microscopy
632 Traffic 2002; 3: 630–640
(TEM) (Figure 3K). In these cells, RanBP2 and Ran alwayscolocalized (Figure 3I–J) in spatially restricted spots (e.g. Fig-ure3G–J, open arrowheads) associated with confined fila-ment-like tracks reminiscent of ‘Plasmastrassen’ (22) or‘roads’ (23). These radiated axially from the perinuclear re-gion and funneled into a protoplasmic extension, which poss-ibly represents an axon hillock (Figure 3F–J). Some of thesespots were often juxtaposed with, and seemingly protrudingfrom, the NE (Figure 3I–J, filled arrowhead).
Localization of RanBP2 and KIF5s in bovine and
human retinas
In light of RanBP2 interaction in vitro and in vivo with the neur-onal-abundant KIF5C and ubiquitous KIF5B, but not KIF5A,microtubule-based motor proteins (15), we investigatedwhether these kinesins mediated transport pathways in a neur-

Neuroretinal Mapping of RanBP2 Macroassembly Complex
Figure3: Expression and localization of RanBP2 and RanGTPase in the bovine retina. Radial cryosections were singly immuno-stained with an affinity-purified polyclonal antibody, ZnF.20908 (7.5mg/ml) against the ZnF-rich domain of bovine RanBP2 (A–C) andmonoclonal (5mg/ml) (D) and polyclonal (1 :500) (E) antibodies against RanGTPase. B is an overlay Nomarski/Differential Interference Contrast(DIC) of the immunofluorescence image in A. RanBP2 and RanGTPase are expressed in all three major nuclear layers of the retina. Inphotoreceptors, RanBP2 is present strongly in and absent from the ellipsoid and myoid (A and C, filled arrowhead) compartments, respec-tively. RanBP2 distribution pattern around the periphery of nuclei of photoreceptors (C, open arrowheads) and other neurons is highlypunctate (C). RanGTPase is expressed and distributed in the intranuclear compartment and nuclear rims of all cell bodies, albeit to differentdegrees (D, E). RanGTPase was distributed continuously throughout the inner segments of photoreceptors (E). F–J are images of a focalplane of a retinal ganglion cell by DIC (F), immunofluorescence-labeled with anti-Ran mAb (2.5 mg/ml) (G) and anti-RanBP2 antibody(ZnF.20908, 3.5mg/ml) (H), overlay of G and H (I), and overlay of I and F (J). Ran and RanBP2 always colocalized at small and restrictedfoci associated with cytoplasmic tracks (e.g. open and filled arrowheads in F–J). Focal ‘bursts’ of RanBP2- and Ran-containing particles areseen closely associated with the NE and juxtaposed to this (e.g. filled arrowhead in F, I and J). K is a transmission electron micrograph(30000¿) of a ganglion cell nucleus with large soma depicting the large density of NPCs at the NE. Legend: ROS, rod outer segments;RIS, rod inner segments; CN, cones; ONL, outer nuclear layer; OPL, outer plexiform layer, INL, inner nuclear layer; IPL, inner plexiform layer;GC, ganglion cell layer; NFL, nerve fiber layer.
onal cell- (and/or compartment)-specific fashion in the neuror-etina. As seen in Figure 4(A–D) of a bovine retina, the H1 mAb(24), which recognizes the neuronal-abundant and recombin-ant KIF5A and KIF5C motor proteins (15), detected the expres-sion of one and/or both of these kinesins strongly in rod andcone photoreceptors and axons comprising the NFL, but not inother neurons. In all rod and the vast majority of cone photore-ceptors, KIF5C filled the intercompartmental (myoid) spacebetween the ellipsoid region and NE of nuclei, locales it inter-faced with RanBP2. Such intercompartmental space was con-spicuously absent from a small population of topographicallydistinct cones with their nuclei protruding distally from the ONLand thus, without KIF5C, and the expression of RanBP2 wascontinuous between the NE and inner segments (Figure 4A,D,white arrow). KIF5C was present in the (outer and inner) fibers,emanating from the nuclei of photoreceptors (25,26), and their
633Traffic 2002; 3: 630–640
synapses (Figure 4B–C). Punctate labeling was also seen atthe tip of ellipsoid compartment, indicating the presence ofKIF5C/KIF5A at the connecting cilium and/or basal bodies(Figure 4B, filled arrowheads), as suggested by other studiesfor other homologous kinesins (27). KIF5C/KIF5A kinesinswere conspicuously absent from the inner nuclear layer andthroughout the perikarya and NE of ganglion cells (Figure4A).
In human retinas, KIF5C/KIF5A had a similar overall distri-bution and colocalization pattern to that seen in bovine ret-inas, albeit with some noteworthy differences. In the innersegments of all photoreceptor types, KIF5C, while preponder-ant in the myoid region, was also present throughout the el-lipsoid compartment, where its distribution graduallydwindled towards the apical end of this compartment (Figure

Mavlyutov et al.
Figure4: Distribution and colocalization of RanBP2 and KIF5s in bovine and human retinas. A–F and G–J are, respectively, radialcryosections (3mm) and overlay images of retinas double-immunostained with anti-RanBP2 (green) and H1mAb (red), and anti-RanBP2(green) and H2 (red) antibodies (except in J where anti-RanBP2 and anti-H2 are depicted in red and green, respectively). E and F are imagesfrom human retinas, and all others are bovine retinas. F is an overlay of E and its DIC image. The H1 antibody immunostained strongly themyoid compartment (e.g. black arrowhead in A and B) as well as the NFL. In A and D, white arrow points to a rare (blue) photoreceptor(see also Figure 4). In this rare subclass of blue cones, KIF5 expression is absent from the myoid compartment and RanBP2 distribution(also seen in H) is uninterrupted throughout the inner segment. In B (amplified in C), note the arrows pointing to inner fibers emanatingfrom (red) cone nuclei. The NEs of these are rich in RanBP2 (open arrowheads in B and also shown in I). KIF5C interfaces with RanBP2 atthe NE and boundary region between myoid and ellipsoid compartments of bovine (B and C) and human photoreceptors (E, F, arrowheadin E). Note KIF5C presence in the cone pedicles (E, arrow) and connecting cilium and/or basal bodies (E, open arrowheads). In G, H and Inote the absence of KIF5B with H2 antibody from the myoid and presynaptic regions of photoreceptors (filled arrowheads in G). KIF5B wasabundant in the postsynaptic endings of OPL, INL, OPL, and perikarya and NE of ganglion cells. KIF5 and RanBP2 colocalized at the NE ofa ganglion cell (J). A–F, E–F and G–J, respectively, were immunostained with antibodies, ZnF.20908 (7.5mg/ml) and H1 (7.5mg/ml),ZnF.20908 (10mg/ml) and H1 (10mg/ml), and ZnF.20909 (1.5mg/ml) and H2 (15mg/ml). Nuclei are stained in blue by DAPI. Note the nucleiof photoreceptors stain stronger with DAPI than those of cells in the INL (and ganglion cells). Legend: ROS, rod outer segments; RIS, rodinner segments; CN, cones; ONL, outer nuclear layer; OPL, outer plexiform layer, INL, inner nuclear layer; IPL, inner plexiform layer; GC,ganglion cell layer; NFL, nerve fiber layer; ELM, external limiting membrane (a virtual membrane formed by plaque-bearing junctions betweenphotoreceptors and Muller glial cells).
634 Traffic 2002; 3: 630–640

Neuroretinal Mapping of RanBP2 Macroassembly Complex
Figure5: Identification of a topographically distinct subclass of blue cones (A–E) and colocalization of kinesin-light chains(KLC) with RanBP2 (F–H). Topographically distinct subsets of blue cones (compare left (arrow) and right ‘green-stained’ cones in A andimmunolabeled with anti-blue opsin JH455 Ab at 1 :1000) have KIF5C (red; H1 mAb at 10mg/ml) conspicuously absent from any compart-ment distal of the nucleus. B is a DIC image of the section in A (filled arrowhead COS, open arrowhead, myoid). An overlay image of KIF5C(H1 mAb, 10mg/ml) and red opsin (Ab.21069, 1 :1000) localization (C) shows the presence of KIF5C in the myoid compartment (openarrowhead) of a red cone (ROS, filled arrowhead) through the rod photoreceptor inner segment (arrow, C) and its outer fiber. D is an overlayof C and the same imaged by DIC. All nuclei (filled arrowheads) of red cones (Ab.21069, 1 :1000) can be traced to the uppermost nucleirow of the ONL (E). Red opsin is located in the perinuclear and myoid regions and plasma membrane of inner segments. F and G aremerged images of DIC and co-immunostained sections with anti-RanBP2 antibody (green, ZnF.20908 at 10mg/ml) and a monoclonalantibody (red, L2 at 15mg/ml) against the variable C-terminal domain of KLC. KLCs are exclusively detected in perikarya of ganglion cells,and some of these interface with RanBP2 at the nuclear envelope (F, G). Note also the aligned RanBP2 positive foci (dashed arrow) (as inFigure 3H) in a protoplasmic extension of a ganglion cell (G). Not all ganglion cells seem to express RanBP2 or KLC (G). KLC distributionwas similar with L1 monoclonal antibody (10 mg/ml) against the conserved N-terminus of KLCs (H). Legend: COS, cone outer segments;KLCs, kinesin light chains/s; ROS, rod outer segments; RIS, rod inner segments; CN, cones; ONL, outer nuclear layer; OPL, outer plexiformlayer, INL, inner nuclear layer; IPL, inner plexiform layer; GC, ganglion cell layer; NFL, nerve fiber layer.
4E,F, data not shown). In addition, a distinct punctate stainingof kinesin, and nonoverlapping with RanBP2, was observedat the apical end of all inner segments, indicating the localiz-ation of KIF5C/KIF5A at the basal bodies and/or connectingcilium of photoreceptors (Figure 4E,F open arrowheads), assuggested by other studies for related kinesins (27). KIF5A/
635Traffic 2002; 3: 630–640
KIF5C filled the calyciform synaptic endings of the cone ped-icles (Figure 4E, arrows), and possibly, rod spherules, ofphotoreceptors in a markedly homogeneous-sized vesicle-like and pseudo-orderly pattern.
Then, we investigated the distribution of KIF5B with H2 mAb

Mavlyutov et al.
(24), which recognizes the recombinant and ubiquitousKIF5B as well as the neuronal KIF5C/KIF5A isoforms (15).When used at identical concentrations to that employed forH1 mAb, all photoreceptor compartments were surprisinglydenuded of immunostaining by the H2 mAb (Figure 4G–I).This suggests that this antibody fails to detect KIF5C in retinalsections (see Discussion), albeit immunoreactivity in photore-ceptors could be observed when H2 was used at higher con-centrations (data not shown). To this end, H2 detected thepresence of KIF5B in the distal synaptic endings of the OPL,neuronal bodies throughout the INL, IPL and in a subpopula-tion of ganglion cells with large cell bodies and nuclei (Figure4G–I). In these ganglion cells, KIF5B colocalized with RanBP2at the periphery of the NE, where both proteins were veryprominent, and throughout the perikarya of these cells (Fig-ure 4G,H,J).
KIF5C transport pathway is absent from the myoid
segment of a topographically distinct subclass of blue
cones with unique RanBP2 distribution
We consistently observed a topographically distinct subclassof cones with shorter inner segments and whose nuclei al-ways protruded distally from the edge of the ONL. In thesecones, the apparent absence of the intersegmental compart-ment gave rise to an uninterrupted distribution of RanBP2between the nuclear and inner segment compartmentswhere no KIF5C presence was detected (Figure 5A, arrow).These protruding cones comprised a small population of bluecones (Figure 5A). Conversely, all other blue (Figure 5A) andred (Figure 5C,D) cones contained KIF5 in the myoid, but notthroughout the ellipsoid, compartments. In rod photorecep-tors (Figure 5C,D), KIF5 distribution was observed throughouttheir inner segments (arrow) and outer fibers. An antibodyagainst the N-terminus of bovine red opsin sequence tracedthe nuclei of red cones always to the outer edge of the ONL(Figure 5E), as supported by transgenic studies with a re-porter gene (28). Interestingly, red opsin was observedaround the perinuclear region, immediate myoid compart-ment distal of the nucleus, plasma membrane of inner seg-ments and, as expected, concentrated in outer segments(Figure 5E).
The kinesin-light chains have a distribution largely
distinct from KIF5s and RanBP2 in the retina
The coiled-coil/tail domains of KIF5C and KIF5B associatespecifically with RanBP2 (15). This association is ac-companied by the co-association of the kinesin-light chains(KLC), but these are not required in vitro for KIF5 interactionwith RanBP2 (15). Thus, we investigated the distribution pat-tern of these in relation to RanBP2 and KIF5 in the retina. ThemAb L2 (24) against the variable C-terminal end of KLC (29)detected the expression of these as abundant heterogen-eous-sized signals throughout the perikarya of ganglion cells(Figure 5F,G). Some of these granules overlapped withRanBP2 staining at the cytoplasmic face of nuclear envelopes(Figure 5F,G). In addition, KLC could not be observed in otherretinal cell types (e.g. photoreceptors) (Figure 5F). The mAb,L1 (24), against the conserved N-terminal domain of KLC
636 Traffic 2002; 3: 630–640
(29) provided similar results, with the exception of a ratherweak and diffuse distribution of KLC in the inner segment ofphotoreceptors (Figure 5H).
Importin-b is a cargo for KIF5C and KIF5B, and
colocalizes with RanBP2
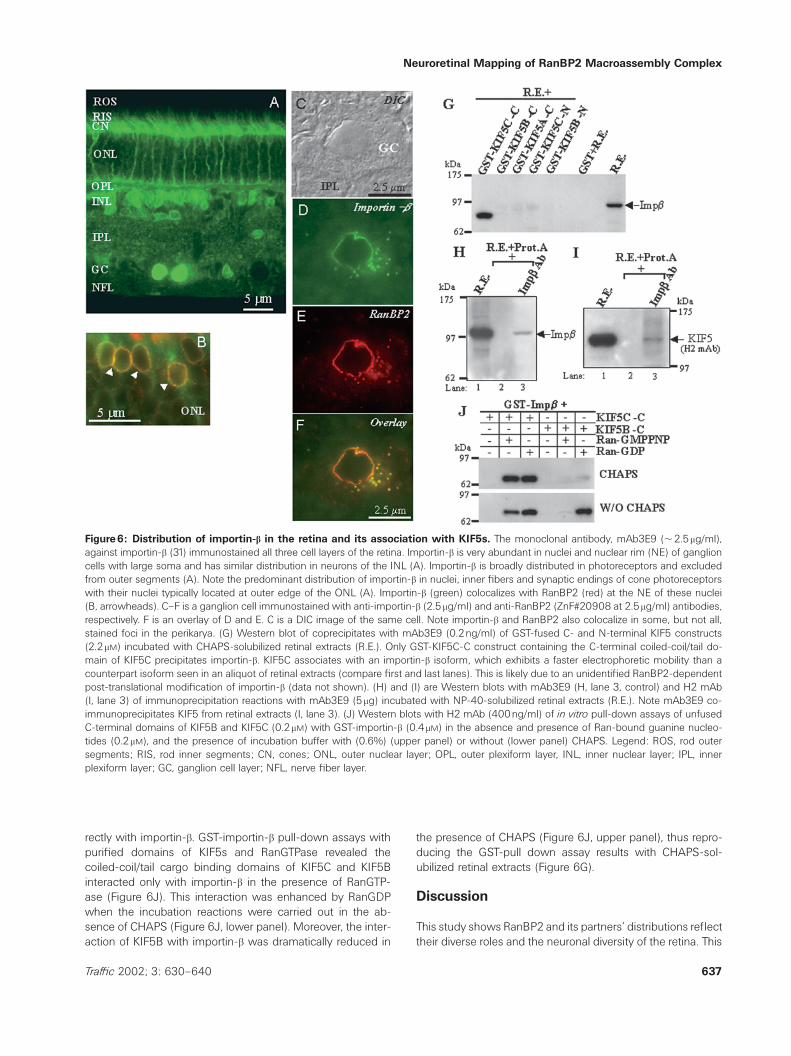
The nuclear import receptor, importin-b, was reported tointeract with RanBP2 (6,30), and we have confirmed suchspecific interaction in retinal extracts with certain RanBP2domains (manuscript in preparation). To dissect out therole of KIF5s in RanBP2-mediated nuclear-cytoplasmic traf-ficking processes in neuroretinal cells, and since no endog-enous cargoes have been yet identified for KIF5s, we inves-tigated whether importin-b was an endogenous cargo forone or more KIF5s in retinal neurons. We first determinedthe distribution pattern of importin-b among retinal neurons.Immunofluorescence microscopy with the anti-importin-b
monoclonal antibody, mAb 3E9 (31), showed importin-b
was differentially distributed in subcellular compartmentsamong retinal neurons and in photoreceptors, paralleledthat observed for some KIF5s (Figure 6A). Importin-b wasprominent in the myoid, perinuclear and intranuclear re-gions of cone nuclei at the distal edge of the ONL, innerfibers emanating from their cell bodies and synaptic end-ings (Figure 6A). In addition, importin-b was localizedmainly in the perinuclear and intranuclear cell bodies ofneurons in the inner nuclear layer (Figure 6A) typical ofthat seen in non-neuronal cells. Among some ganglioncells, its localization was remarkably intranuclear and at thenuclear rim of large ganglion cell bodies (Figure 6A).RanBP2 and importin-b strongly colocalized in punctatestructures (nuclear pores) at the rim of nuclei of cones(Figure 6B) and ganglion cells (Figure 6C–F) and restrictedfoci in the perikaryon of these latter cells (Figure 6C–F).Only a subpopulation of importin-b-immunoreactive foci inthe perikaryon contained RanBP2 (Figure 6F). These im-munoreactive foci are likely not due to the presence ofannulate lamellae structures in the perikarya, becauseanalysis by transmission electron microscopy of ganglioncells did not reveal the presence of such structures inthese neurons (data not shown).
We investigated whether importin-b constitutes an endoge-nous cargo for KIF5s, in part because of the striking similar-ities between importin-b and KIF5C distributions in the conephotoreceptors. To this end, incubation of GST-constructs,comprising just the coiled-coil/tail cargo binding and motordomains of KIF5s with retinal extracts, led to the coprecipita-tion of importin-b only with the coiled-coil/tail of KIF5C (GST-KIF5C-C) (Figure 6G). Interestingly, KIF5C associates with animportin-b isoform with a faster electrophoretic mobility thanthat seen in an aliquot of retinal extracts (Figure 6G). Thiselectrophoretic mobility change in importin-b is likely due toa RanBP2-dependent post-translational modification of im-portin-b (data not shown). Immunoprecipitation assays withimportin-b antibody, mAb3E9, coprecipitated KIF5s from reti-nal extracts, indicating they are natural partners in vivo (Fig-ure 6I). Then, we determined whether KIF5s interacted di-
Neuroretinal Mapping of RanBP2 Macroassembly Complex
Figure6: Distribution of importin-b in the retina and its association with KIF5s. The monoclonal antibody, mAb3E9 (2.5mg/ml),against importin-b (31) immunostained all three cell layers of the retina. Importin-b is very abundant in nuclei and nuclear rim (NE) of ganglioncells with large soma and has similar distribution in neurons of the INL (A). Importin-b is broadly distributed in photoreceptors and excludedfrom outer segments (A). Note the predominant distribution of importin-b in nuclei, inner fibers and synaptic endings of cone photoreceptorswith their nuclei typically located at outer edge of the ONL (A). Importin-b (green) colocalizes with RanBP2 (red) at the NE of these nuclei(B, arrowheads). C–F is a ganglion cell immunostained with anti-importin-b (2.5mg/ml) and anti-RanBP2 (ZnF.20908 at 2.5mg/ml) antibodies,respectively. F is an overlay of D and E. C is a DIC image of the same cell. Note importin-b and RanBP2 also colocalize in some, but not all,stained foci in the perikarya. (G) Western blot of coprecipitates with mAb3E9 (0.2ng/ml) of GST-fused C- and N-terminal KIF5 constructs(2.2mM) incubated with CHAPS-solubilized retinal extracts (R.E.). Only GST-KIF5C-C construct containing the C-terminal coiled-coil/tail do-main of KIF5C precipitates importin-b. KIF5C associates with an importin-b isoform, which exhibits a faster electrophoretic mobility than acounterpart isoform seen in an aliquot of retinal extracts (compare first and last lanes). This is likely due to an unidentified RanBP2-dependentpost-translational modification of importin-b (data not shown). (H) and (I) are Western blots with mAb3E9 (H, lane 3, control) and H2 mAb(I, lane 3) of immunoprecipitation reactions with mAb3E9 (5 mg) incubated with NP-40-solubilized retinal extracts (R.E.). Note mAb3E9 co-immunoprecipitates KIF5 from retinal extracts (I, lane 3). (J) Western blots with H2 mAb (400ng/ml) of in vitro pull-down assays of unfusedC-terminal domains of KIF5B and KIF5C (0.2 mM) with GST-importin-b (0.4mM) in the absence and presence of Ran-bound guanine nucleo-tides (0.2mM), and the presence of incubation buffer with (0.6%) (upper panel) or without (lower panel) CHAPS. Legend: ROS, rod outersegments; RIS, rod inner segments; CN, cones; ONL, outer nuclear layer; OPL, outer plexiform layer, INL, inner nuclear layer; IPL, innerplexiform layer; GC, ganglion cell layer; NFL, nerve fiber layer.
rectly with importin-b. GST-importin-b pull-down assays withpurified domains of KIF5s and RanGTPase revealed thecoiled-coil/tail cargo binding domains of KIF5C and KIF5Binteracted only with importin-b in the presence of RanGTP-ase (Figure 6J). This interaction was enhanced by RanGDPwhen the incubation reactions were carried out in the ab-sence of CHAPS (Figure 6J, lower panel). Moreover, the inter-action of KIF5B with importin-b was dramatically reduced in
637Traffic 2002; 3: 630–640
the presence of CHAPS (Figure 6J, upper panel), thus repro-ducing the GST-pull down assay results with CHAPS-sol-ubilized retinal extracts (Figure 6G).
Discussion
This study shows RanBP2 and its partners’ distributions reflecttheir diverse roles and the neuronal diversity of the retina. This

Mavlyutov et al.
is in contrast to other non-neuronal cells where RanBP2 hasbeen linked just to cytoplasmic fibrils emanating from NPCs(6,7,20,21). Four unique facets of RanBP2 expression and dis-tribution are reported here. First, RanBP2 was preponderant inthe ellipsoid but not the myoid compartments of (most) coneand rod photoreceptors. Thus, RanBP2 distribution in photore-ceptors strongly departed from that observed in other cells.Second, a distinct topographic subpopulation of blue coneshad a continuous distribution of RanBP2 between the nuclearenvelope and inner segment. This sort of distribution was alsoobserved for RanGTPase along all inner segments of photore-ceptors, where RanGTPase was prominent. Third, RanBP2 andRanGTPase always colocalized at discrete spots along cyto-plasmic tracks (and protoplasmic extensions) of some gan-glion cells extremely rich in NPCs and RanBP2. Some of thesespots were closely associated with the NE, as if they werepinching off from the latter, reminiscent of reports of vesiclebudding from the NE in yeast (32). This and RanBP2’s localiz-ation in the ellipsoid compartments of photoreceptors indicateRanBP2 is released from the filaments at the nuclear pores.The implications of its release are unclear, but one possibilityis that RanBP2 plays an additional role as a scaffold carrier indelivering some of its associated partners away from the nu-clear pore. An analogous scenario has been proposed for thekinesin-mediated delivery of scaffold proteins of the JNK-sig-naling pathway (33). Finally, the relative abundance of RanBP2at the NE among retinal cells was remarkable, with many gan-glion cells having the highest expression of RanBP2 and NPCsat the nuclear envelope. This may reflect largely differences inmetabolic activity between retinal neurons, as many ganglioncells were extremely rich in biosynthetic machinery (endoplas-mic reticulum, polyribosomes, etc.) compared to that seen inother neurons by transmission electron microscopy (data notshown).
In this study, we find the interactions of KIF5B/KIF5C andRanBP2 are prominently and differentially restricted to sub-populations of neurons and compartments thereof. Immuno-staining with H1 antibody showed that the KIF5C, but not theKIF5B, transport pathway is restricted to photoreceptors (andNFL). They colocalize with RanBP2 at the NE and boundaryregion of the myoid interfacing with the ellipsoid compart-ment of cones (and rods). The absence of immunostaining ofKIF5s from any photoreceptor compartment by H2 antibody(when used at the same concentration of H1) is surprisingbecause this antibody recognizes all recombinant KIF5 iso-forms (15). Still, H2 antibody recognizes predominantly KIF5in neuronal tissues, despite its slightly broader tissue-speci-ficity compared to that of H1 on Western blots of tissue ex-tracts (15). Therefore, it is plausible that epitope masking dueto post-translational modification of KIF5C (and possibly,KIF5A) plays a role in the KIF5C recognition by the H2 anti-body, thus leaving KIF5B as the only remaining KIF5 to bedetected in the retina. This is supported by the observationthat a different antibody, Pcp42, which recognizes also therecombinant isoforms of KIF5B and KIF5C, detects an evenand broad tissue-distribution of KIF5s (15). Moreover, it is alsopossible that KIF5B expression is much higher than KIF5C in
638 Traffic 2002; 3: 630–640
the retina, in particular inner retinal neurons, and thus H2 willpreferentially detect KIF5B. To this effect, immunodetectionof KIF5s can also be observed in the photoreceptors whenH2 antibody is used at significantly higher concentrations(data not shown). Altogether, these results indicate theKIF5B-mediated transport pathway (15) is absent in photore-ceptors and predominates in most other neuronal cell bodies,in particular, ganglion cells with large cell bodies. In addition,both KIF5B and KIF5C are present in the axonal-rich, nervefiber layer, as reported in other studies (34).
The restricted localization of KLCs, which largely did not over-lap with the kinesin heavy chain (KHC) partners, is also sur-prising and indicates additional roles for these accessory pro-teins other than transport receptors of cargoes, as previouslysuggested (33). Its partial colocalization with RanBP2 at thenuclear rim of ganglion cells and interaction in vivo withRanBP2 and KIF5s (15) indicate one of its functions may beto modulate this interaction and/or prime KIF5s with cargoesmediated by the RanBP2-mediated nuclear-cytoplasmic traf-ficking pathway upon their delivery and/or arrival. For the firsttime, we have identified importin-b, a coreceptor for sub-strates containing nuclear localization signals (NLS), as a di-rect cargo for KIF5C, and possibly KIF5B. This interaction oc-curs directly with the C-terminal coiled-coil tail domain of ki-nesin heavy chain in vitro, but only in the presence ofRanGTPase. This supports a cooperative mechanism be-tween these molecules [e.g. analogous to that observed forRanGTP-dependent binding of NES substrates to the NESnuclear receptor, CRM1/exportin-1 (35)], whereby aRanGTPase-dependent conformational change in importin-b
may lead to its binding to the coiled-coil/tail domain of KIF5C(and KIF5B). The interaction of KHC tail directly with its car-goes is in line with a study in Neurospora crassa, where KHCalone has been shown to bind to cargo (36). The specificityof the interaction of KIF5s with their cargoes is likely deter-mined by their unique and very C-terminal domains (15).
The distribution pattern of importin-b and its interaction withKIF5C in photoreceptors suggests that it likely constitutes anactive nuclear carrier of substrates transported from/to syn-aptic terminals and inner segments. In light of a growingnumber of mutations in transcription and other nuclear fac-tors affecting photoreceptor function and leading to congeni-tal retinopathies (http://www.sph.uth.tmc.edu/RetNet/dis-ease.htm), the KIF5C-mediated nuclear-cytoplasmic traffick-ing pathway towards the RanBP2 molecular station emergesas a candidate transport pathway for the convergence of nu-clear import of such factors, and wherein mutations maycompromise this process. Likewise, KIF5C- and/or KIF5B-de-pendent nuclear import pathways may be involved in otherdegenerative neuropathies. For example, the pathologic ex-pansion of polyglutamine (polyQ) repeats of cytosolic pro-teins in a growing spectrum of neurodegenerative neuro-pathies leads to the aberrant localization and deposit of thesein the intranuclear compartment followed by neuronal celldeath (37). As the nuclear import pathway of these aberrantproteins remains to be identified (38), and in light of KIF5s

Neuroretinal Mapping of RanBP2 Macroassembly Complex
and RanBP2 (data not shown) expression in different regionsof the brain, some KIF5s and/or RanBP2 may participate inthe aberrant nuclear import of such polyQ-expanded pro-teins.
The differential distribution of RanBP2 and KIF5C in the my-oid compartments among blue cones likely reflects differ-ences in the cytoarchitectural microtubule organization ofthese cells or compartments thereof. The virtual exclusion ofKIF5C (and ‘inclusion’ of RanBP2) from the myoid compart-ment of some blue cone photoreceptor cells indicates thereis no need for active transport of cargoes by KIF5s to/fromRanBP2. Instead, diffusion and/or facilitated transport mayaccount for such transport activity in the distal compartmentof these cells. Such differences are also mirrored by the moreextensive distribution of KIF5C throughout the rod inner seg-ments, whereas in red cones, KIF5C was mostly excludedfrom the ellipsoid segment. Thus, these data support theexistence of a spatially restricted cytoarchitecture of the my-oid region responsible for orchestrating intracellular transportevents mediated by KIF5C (and RanBP2) in most photore-ceptors.
Materials and Methods
Tissue sources and primary antibodies
Fresh bovine (Bos taurus) eyes with a postmortem time of less than 30minwere obtained from a local slaughterhouse. Human eyes with a postmor-tem time of 6–12h were provided by the Eye Bank at the Eye Instituteof the Medical College of Wisconsin. All tissue manipulation procedurescomplied with institutional and federal guidelines.
The polyclonal and affinity-purified antibodies, ZnF-20909 and ZnF-20908, against the Zn-finger-rich (ZnF) domain of bovine RanBP2 havebeen described previously (15). H1 and H2 monoclonal antibodies againstthe conventional kinesin heavy chain purified from bovine brain (19) werekindly supplied by George Bloom (University of Virginia) and Scott Brady(U.T. South-western Medical Center). L1 and L2 monoclonal antibodiesagainst the kinesin light chain (KLC) purified from bovine brain (24) werepurchased from Chemicon (Temecula, CA, USA). Anti-RanGTPase mono-clonal and polyclonal antibodies, respectively, were purchased from Trans-duction Laboratories (Lexington, KY, USA) and a gift from Peter D’Eusta-chio (New York University Medical Center). The monoclonal antibody, mAb3E9, against importin-b (31) was a generous gift from Stephen Adam(Northwestern University Medical School). The antibodies, JH455 and JH492, against human blue and red opsin were kindly supplied by JeremyNathans at John Hopkins University (21). Only results with JH455 areshown in this report. Two rabbit polyclonal antibodies (.21068 and.21069) against the first N-terminal 38 amino acids of the bovine redopsin (Genebank Accession.: AF280398, unpublished results) (28) weregenerated and antibody titers determined by ELISA (Research Genetics,Huntsville, AL, USA). They provided similar and expected results on West-ern blots (data not shown) and immunohistochemistry. Only Ab.21069 isshown in this report.
Immunohistochemistry and transmission electron microscopy
Posterior eye cups were fixed in 2% paraformaldehyde in phosphate bufferpH7.4 at 4 æC for about 1h and infused sequentially for 12h with 15 and30% sucrose. Retinas were embedded in O.C.T. medium (Miles, INC,Eikhart, IN, USA), frozen in liquid nitrogen and sectioned radially at 3mm
639Traffic 2002; 3: 630–640
in a Reichert-Jung cryostat. Retinal cryosections were mounted onto gela-tin-coated slides, incubated with 10% normal goat serum (Jackson Immu-noresearch Laboratories, West Grove, PA, USA) in incubation buffer (0.1%Triton X-100 or 0.1% Saponin in PBS, pH7.4) for 15min followed by an-other incubation for 2h at room temperature in the same buffer contain-ing the primary antibody at the concentrations described in figure legends.For single-label immunofluorescence microscopy, sections were thenrinsed thrice for 10min with incubation buffer followed by incubation inthe same buffer with goat anti-rabbit or anti-mouse secondary antibodies(2.5mg/ml) conjugated with the fluorescent dyes, Alexa488 or Alexa594(Molecular Probes, Eugene, OR, USA), for 1h at room temperature. Finally,sections were DAPI stained, washed twice with incubation buffer for about5min, mounted with Antifade mounting medium (Molecular Probes, Eug-ene, OR, USA) and cover-slipped. For double-label immunofluorescencemicroscopy, primary antibodies from different species (rabbit or mouse)were added sequentially after intermediate washing steps and additionof the species-specific fluorescent-labeled secondary antibody (2.5mg/ml)described above. Control experiments were carried out whenever appropri-ate with cognate preimmune serums, omitting the primary antibody and/or preadsorbed antibodies. Visualization of retinal sections and localizationof proteins were carried out by Nomarski and wide-field epifluorescencemicroscopy at various focal planes with an E600 Nikon research light mi-croscope. This was equipped with fluorescence, appropriate filters, a 100-Watt mercury light source, Nomarski DIC and Plan Apochromat optics(100¿- and 60¿-oil objectives with N.A 1.4, and 40¿, 20¿ and 10¿objectives). Crossover of fluorescent probes, background and autofluo-rescence were found to be negligible. Images were acquired with a SPOT-RT digital camera coupled to the microscope and driven by SPOT Imagingv3.4 software (Diagnostic Instruments, Sterling Heights, MI, USA). All im-ages were captured at nonsaturating integration levels, 12-bit mono black/white (except for DAPI staining), then pseudo-colored and whenever appli-cable, fluorescent images were merged with the same software. Overallarrangements and cropping of images were performed by importing theseto Adobe Photoshop v5.5 (Adobe, Mountain View, CA, USA). For trans-mission electron microscopy, bovine retinal sections 40nm thick were pro-cessed and poststained according Karnovsky’s method (39) and imagedwith a Hitachi TEM.
Tissue extract preparation, coimmunoprecipitation, GST-pull-
down and in vitro assays
CHAPS-solubilized retinal extracts, expression and purification of GST-fused constructs were prepared exactly as previously described (19). AllGST pull-down assays with retinal extracts were carried out with 2.2mM ofGST-fused proteins and as described in detail elsewhere (19). Preparationof these GST-fused constructs has been described (15). Co-precipitates ofretinal and in vitro (in the presence or absence of CHAPS) incubation reac-tions with GST-fused constructs at concentrations described in the figurelegends were resolved on SDS-PAGE and analyzed by Western blot withantibodies described in the results. Immunoprecipitations assays were de-scribed elsewhere (15). Nucleotide exchange of recombinant RanGTPasewas performed in the presence of 5–10mM EDTA and nucleotide, followedby the addition of 10mM MgCl2.
Acknowledgments
We thank George Bloom and Scott Brady for the generous amounts of H1and H2 monoclonal antibodies, helpful discussions and suggestions. Wethank Stephen Adam, P. Eustachio and J. Nathans for kindly supplyingantibodies against importin-b, RanGTPase, and red and blue opsin, respec-tively. We also thank Stephen Adam for providing the GST-importin-b con-struct. This work was supported by NIH grants EY11993 and EY012665to P.A.F. T.A.M. was supported in part by a postdoctoral fellowship fromFight for Sight research division of Prevant Blindness American.

Mavlyutov et al.
References
1. Gorlich D, Kutay U. Transport between the cell nucleus and the cyto-plasm. Annu Rev Cell Dev Biol 1999;15:607–660.
2. Mattaj I, Englmeier L. Nucleocytoplasmic transport: the soluble phase.Annu Rev Biochem 1999;67:265–306.
3. Gorlich D, Mattaj I. Nucleocytoplasmic transport. Science1996;271:1513–1518.
4. Siniossoglou S, Wimmer C, Rieger M, Doye V, Tekotte H, Weise C,Emig S, Segref A, Hurt EC. A novel complex of nucleoporins, whichincludes Sec13p and a Sec13p homolog, is essential for normal nu-clear pores. Cell 1996;84:265–275.
5. Nanduri J, Tartakoff A. The arrest of the secretion response in yeast:Signaling from the secretory path to the nucleus via Wsc proteins andPkc1p. Mol Cell 2001;8:281–289.
6. Delphin C, Guan T, Melchior F, Gerace L. RanGTP targets p97 toRanBP2, a filamentous protein localized at the cytoplasmic peripheryof the nuclear pore complex. Mol Biol Cell 1997;8:2379–2390.
7. Yokoyama N, Hayashi N, Seki T, Pante N, Ohba T, Nishii K, Kuma K,Hayashida T, Miyata T, Aebi U, Fukui M, Nishimoto T. A giant nu-cleopore protein that binds Ran/TC4. Nature 1995;376:184–188.
8. Mahajan R, Delphin C, Guan T, Gerace L, Melchior F. A small ubiquitin-related polypeptide involved in targeting RanGAP1 to nuclear porecomplex protein RanBP2. Cell 1997;88:97–107.
9. Ferreira PA, Hom J, Pak W. Retina-specifically expressed novel sub-types of bovine cyclophilin. J Biol Chem 1995;270:23179–23188.
10. Fauser S, Roepman R, Ferreira PA. Genomic organization, expressionand localization of murine Ran-binding protein 2 (RanBP2) gene.Mamm Genome 2000;12:406–415.
11. Ferreira PA, Nakayama T, Pak W, Travis G. Cyclophilin-related proteinRanBP2 acts as chaperone for red/green opsin. Nature1996;383:637–640.
12. Ferreira P, Nakayama T, Travis G. Interconversion of red opsin isoformsby the cyclophilin-related chaperone protein, RanBP2. Proc Natl AcadSci USA 1997;94:1556–1561.
13. Ferreira P, Yunfei C, Schick D, Roepman R. The cyclophilin-like domainmediates the association of Ran-binding protein 2 with subunits ofthe 19S regulatory complex of the proteasome. J Biol Chem1998;273:24676–24682.
14. Singh B, Patel H, Roepman R, Schick D, Ferreira PA. The zinc fingercluster domain of RanBP2 is a specific docking site for the nuclearexport factor, exportin-1. J Biol Chem 1999;274:37370–37378.
15. Cai Y, Singh B, Aslanukov A, Zhao H, Ferreira PA. The docking ofkinesins, KIF5B and KIF5C, to RanBP2 is mediated via a novel RanBP2domain. J Biol Chem 2001;276:41594–41062.
16. Saitoh H, Sparrow D, Shiomi T, Pu R, Nishimoto T, Mohun T, DassoM. Ubc9p and the conjugation of SUMO-1 to RanGAP1 and RanBP2.Curr Biol 1998;8:121–124.
17. Mahajan R, Gerace L, Melchior F. Molecular characterization of theSUMO-1 modification of RanGAP1 and its role in nuclear envelopeassociation. J Cell Biol 1998;140:259–270.
18. Manutis M, Coutavas E, Blobel G. A novel ubiquitin-like modificationmodulates the partitioning of the Ran-GTPase-activating protein Ran-GAP1 between the cytosol and the nuclear pore complex. J Cell Biol1996;135:1457–1470.
19. Ferreira PA. Characterization of RanBP2-associated molecular compo-nents in neuroretina. Methods Enzymol 2000;316:455–468.
640 Traffic 2002; 3: 630–640
20. Wilken N, Senecal J-L, Scheer U, Dabauvalle M-C. Localization of theRan-GTP binding protein RanBP2 at the cytoplasmic side of the nu-clear pore complex. Eur J Cell Biol 1995;68:211–219.
21. Wu J, Matunis MJ, Kraemer D, Blobel G, Coutavas E. Nup358, a cyto-plasmically exposed nucleoporin with peptide repeats, Ran-GTP bind-ing sites, zinc fingers, a cyclophilin a homologous domain, and a leu-cine-rich region. J Biol Chem 1995;270:14209–14213.
22. Andres KH. Untersuchungen uber den Feinbau von Spinalganglien. ZZellforsch Mikrosk Anat 1961;55:1–48.
23. Bunge M, Bunge R, Peterson E, Murray MR. A light and electron mi-croscope study of long-term organized cultures of rat dorsal root gan-glia. J Cell Biol 1967;32:439–466.
24. Pfister K, Wagner M, Stenoien DA, Brady S, Bloom G Monoclonalantibodies to kinesin heavy and light chains stain vesicle-like struc-tures, but not microtubules, in cultured cells. J Cell Biol1989;108:1453–1463.
25. Missotten L. The Ultrastructure of the Human Retina. Brussels: ArsciaUitgaven; 1965.
26. Townes-Anderson E, Dacheux R, Raviola E. Rod photoreceptors dis-sociated from the adult rabbit retina. Neuron 1988;8:320–331.
27. Beech P, Pagh-Roehl K, Noda Y, Hirokawa N, Burnside B, RosenbaumJ. Localization of kinesin superfamily proteins to the connecting ciliumof fish photoreceptors. J Cell Sci 1996;109:889–897.
28. Wang Y, Macke J, Merbs S, Zack D, Klaunberg B, Bennett J, GearhartJ, Nathans J. A locus control region adjacent to the human red andgreen visual pigments genes. Neuron 1992;9:429–440.
29. Khodjakov A, Lizunova E, Minin A, Koonce M, Gyoeva F. A specificlight-chain of kinesin associates with mitochondria in cultured cells.Mol Biol Cell 1998;9:333–343.
30. Braslavsky CI, Novak C, Gorlich D, Wittinghofer A, Kuhlmann J. Differ-ent structural and kinetic requirements for the interaction of Ran withthe Ran-binding domains from RanBP2 and importin-b. Biochemistry2000;39:11629–11639.
31. Chi N, Adam E, Adam S. Sequence and characterization of cyto-plasmic nuclear protein import factor p97. J Cell Biol 1995;130:265–274.
32. Schekman R, Orci L. Coat proteins and vesicle budding. Science1996;271:1526–1533.
33. Verhey K, Meyer D, Deehan R, Blenis J, Schanpp B, Rapoport T, Marg-olis B. Cargo of kinesin identified as JIF scaffolding proteins and as-sociated signaling molecules. J Cell Biol 2001;152:959–970.
34. Elluru R, Bloom G, Brady S. Fast axonal transport of kinesin in the ratvisual system: functionality of kinesin heavy chain isoforms. Mol BiolCell 1995;6:21–40.
35. Fornerod M, Ohno M, Yoshida M, Mattaj I. The human homologue ofyeast CRM1 is in a dynamic subcomplex with CAN/Nup214 and anovel nuclear pore component NUP88. Cell 1997;90:1051–1060.
36. Seiler S, Kirchner J, Horn A, Kallipolitou A, Woehlke G, Schliwa M.Cargo binding and regulatory sites in the tail of fungal conventionalkinesin. Nat Cell Biol 2000;2:333–338.
37. Orr H, Zoghbi H, Reversing neurodegeneration. A promise unfolds.Cell 2000;101:1–4.
38. Tao T, Tartakoff AM. Nuclear relocation of normal Huntingtin. Traffic2001;2:385–394.
39. Karnovsky MJ. A formaldehyde-glutaraldehyde fixative of high osmol-arity for use in electron microscopy. J Cell Biol 1965;27:137A.
Related Documents











