Identification of protein factors and U3 snoRNAs from a Brassica oleracea RNP complex involved in the processing of pre-rRNA Hala Samaha 1,† , Vale ´ rie Delorme 1 , Frederic Pontvianne 1,‡ , Richard Cooke 1 , Francois Delalande 2 , Alain Van Dorsselaer 2 , Manuel Echeverria 1 and Julio Sa ´ ez-Va ´ squez 1,* 1 Laboratoire Ge ´ nome et De ´ veloppement des Plantes, UMR 5096 CNRS-IRD-UPVD; Perpignan France. 52 av. ¨ Paul Alduy, 66860 Perpignan-Cedex, France, and 2 Laboratoire de Spectrome ´ trie de Masse Bio-Organique De ´ partement des Sciences Analytiques, Institut Pluridisciplinaire Hubert Curien, UMR 7178 (CNRS-ULP) ECPM, 25 rue Becquerel F67087-Strasbourg-Cedex, France Received 1 July 2009; revised 18 September 2009; accepted 15 October 2009; published online 8 December 2009. * For correspondence (fax +33 4 68668499; e-mail [email protected]). † Present address: Universite ´ de Pircadie Jules Verne, Amiens, France. ‡ Present address: Washington University, St. Louis, MO, USA. SUMMARY We report on the structural characterization of a functional U3 snoRNA ribonucleoprotein complex isolated from Brassica oleracea. The BoU3 snoRNP complex (formerly NF D) binds ribosomal DNA (rDNA), specifically cleaves pre-rRNA at the primary cleavage site in vitro and probably links transcription to early pre-rRNA processing in vivo. Using a proteomic approach we have identified 62 proteins in the purified BoU3 snoRNP fraction, including small RNA associated proteins (Fibrillarin, NOP5/Nop58p, Diskerin/Cbf5p, SUS2/PRP8 and CLO/GFA1/sn114p) and 40S ribosomal associated proteins (22 RPS and four ARCA-like proteins). Another major protein group is composed of chaperones/chaperonins (HSP81/TCP-1) and at least one proteasome subunit (RPN1a). Remarkably, RNA-dependent RNA polymerase (RdRP) and Tudor staphylococcal nuclease (TSN) proteins, which have RNA- and/or DNA-associated activities, were also revealed in the complex. Furthermore, three U3 snoRNA variants were identified in the BoU3 snoRNP fraction, notably an evolutionarily conserved and variable stem loop structure located just downstream from the C-box domain of the U3 sequence structures. We conclude that the BoU3 snoRNP complex is mainly required for 40S pre-ribosome synthesis. It is also expected that U3 snoRNA variants and interacting proteins might play a major role in BoU3 snoRNP complex assembly and/or function. This study provides a basis for further investigation of these novel ribonucleoprotein factors and their role in plant ribosome biogenesis. Keywords: U3 snoRNP, pre-rRNA, processing, ribosome, Arabidopsis, proteomic. INTRODUCTION Ribosome biogenesis requires coordination between transcription and processing of ribosomal RNA precursors (pre-rRNA), import–export of ribosomal proteins (r proteins) and assembly and transport of ribosome particles (Grandi et al., 2002; Schafer et al., 2003; Tschochner and Hurt, 2003). All these processes begin in the nucleolus with 90S pre- ribosome particle formation, and end in the cytoplasm after export and final maturation of 60S and 40S subunits. One way to coordinate these complex reactions is to concentrate the factors involved in the ribosome biogenesis process in a multifunctional protein complex (reviewed in Fatica and Tollervey, 2002; Fromont-Racine et al., 2003). In the nucleolus, transcription of 18S, 5.8S and 25S–28S rRNA genes by RNA polymerase I (RNA polI) is linked to processing of the primary transcript (pre-rRNA) and 90S pre- ribosomal particle synthesis. Through transcription of the full-length pre-rRNA, the external (5¢ ETS and 3¢ ETS) and internal (ITS-1 and ITS-2) spacers are removed to generate mature rRNA (Nomura, 2001; Saez-Vasquez and Echeverria, 2006). Likewise, modification of numerous rRNA residues occurs. During this process, numerous multifunctional factors associate with the nascent transcribed pre-rRNA, including small nucleolar RNA (snoRNA) as well as ribo- somal and non-ribosomal proteins. The latter include endo ª 2009 The Authors 383 Journal compilation ª 2009 Blackwell Publishing Ltd The Plant Journal (2010) 61, 383–398 doi: 10.1111/j.1365-313X.2009.04061.x

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Identification of protein factors and U3 snoRNAs froma Brassica oleracea RNP complex involved in the processingof pre-rRNA
Hala Samaha1,†, Valerie Delorme1, Frederic Pontvianne1,‡, Richard Cooke1, Francois Delalande2, Alain Van Dorsselaer2,
Manuel Echeverria1 and Julio Saez-Vasquez1,*
1Laboratoire Genome et Developpement des Plantes, UMR 5096 CNRS-IRD-UPVD; Perpignan France. 52 av.¨Paul Alduy, 66860
Perpignan-Cedex, France, and2Laboratoire de Spectrometrie de Masse Bio-Organique Departement des Sciences Analytiques, Institut Pluridisciplinaire
Hubert Curien, UMR 7178 (CNRS-ULP) ECPM, 25 rue Becquerel F67087-Strasbourg-Cedex, France
Received 1 July 2009; revised 18 September 2009; accepted 15 October 2009; published online 8 December 2009.*For correspondence (fax +33 4 68668499; e-mail [email protected]).†Present address: Universite de Pircadie Jules Verne, Amiens, France.‡Present address: Washington University, St. Louis, MO, USA.
SUMMARY
We report on the structural characterization of a functional U3 snoRNA ribonucleoprotein complex isolated
from Brassica oleracea. The BoU3 snoRNP complex (formerly NF D) binds ribosomal DNA (rDNA), specifically
cleaves pre-rRNA at the primary cleavage site in vitro and probably links transcription to early pre-rRNA
processing in vivo. Using a proteomic approach we have identified 62 proteins in the purified BoU3 snoRNP
fraction, including small RNA associated proteins (Fibrillarin, NOP5/Nop58p, Diskerin/Cbf5p, SUS2/PRP8 and
CLO/GFA1/sn114p) and 40S ribosomal associated proteins (22 RPS and four ARCA-like proteins). Another
major protein group is composed of chaperones/chaperonins (HSP81/TCP-1) and at least one proteasome
subunit (RPN1a). Remarkably, RNA-dependent RNA polymerase (RdRP) and Tudor staphylococcal nuclease
(TSN) proteins, which have RNA- and/or DNA-associated activities, were also revealed in the complex.
Furthermore, three U3 snoRNA variants were identified in the BoU3 snoRNP fraction, notably an evolutionarily
conserved and variable stem loop structure located just downstream from the C-box domain of the U3
sequence structures. We conclude that the BoU3 snoRNP complex is mainly required for 40S pre-ribosome
synthesis. It is also expected that U3 snoRNA variants and interacting proteins might play a major role in
BoU3 snoRNP complex assembly and/or function. This study provides a basis for further investigation of these
novel ribonucleoprotein factors and their role in plant ribosome biogenesis.
Keywords: U3 snoRNP, pre-rRNA, processing, ribosome, Arabidopsis, proteomic.
INTRODUCTION
Ribosome biogenesis requires coordination between
transcription and processing of ribosomal RNA precursors
(pre-rRNA), import–export of ribosomal proteins (r proteins)
and assembly and transport of ribosome particles (Grandi
et al., 2002; Schafer et al., 2003; Tschochner and Hurt, 2003).
All these processes begin in the nucleolus with 90S pre-
ribosome particle formation, and end in the cytoplasm after
export and final maturation of 60S and 40S subunits. One
way to coordinate these complex reactions is to concentrate
the factors involved in the ribosome biogenesis process in a
multifunctional protein complex (reviewed in Fatica and
Tollervey, 2002; Fromont-Racine et al., 2003).
In the nucleolus, transcription of 18S, 5.8S and 25S–28S
rRNA genes by RNA polymerase I (RNA polI) is linked to
processing of the primary transcript (pre-rRNA) and 90S pre-
ribosomal particle synthesis. Through transcription of the
full-length pre-rRNA, the external (5¢ ETS and 3¢ ETS) and
internal (ITS-1 and ITS-2) spacers are removed to generate
mature rRNA (Nomura, 2001; Saez-Vasquez and Echeverria,
2006). Likewise, modification of numerous rRNA residues
occurs. During this process, numerous multifunctional
factors associate with the nascent transcribed pre-rRNA,
including small nucleolar RNA (snoRNA) as well as ribo-
somal and non-ribosomal proteins. The latter include endo
ª 2009 The Authors 383Journal compilation ª 2009 Blackwell Publishing Ltd
The Plant Journal (2010) 61, 383–398 doi: 10.1111/j.1365-313X.2009.04061.x

and exonucleases that process the pre-rRNA, pseudouridine
synthases and methyltransferases that catalyse pseudou-
ridylation and 2¢-O-ribose methylation of numerous rRNA
residues, and helicases and chaperones that might facilitate
folding and cleavage of RNA (Eichler and Craig, 1994;
Venema and Tollervey, 1995; Kufel et al., 1999; Barneche
et al., 2000; Comella et al., 2008).
The recent development of sensitive methods for identi-
fying proteins by mass spectrometry has largely contributed
to our knowledge of ribosome formation pathways. Thus,
biochemical characterization of the 90S pre-ribosome parti-
cle revealed that this complex contains mainly ribosomal
proteins from the 40S subunit and �35 non-ribosomal
components, including proteins associated with U3 sno-
RNA, in addition to many others factors required for 18S
rRNA synthesis (Grandi et al., 2002; Schafer et al., 2003).
Interestingly, it almost completely lacks components from
the 60S subunit (Grandi et al., 2002). Thus, the 40S synthesis
machinery is associated with the larger pre-rRNA forms,
whereas factors required for 60S formation bind later,
indicating a clear separation between 40S and 60S biosyn-
thesis pathways. A few years ago, a large U3 snoRNP
complex named the SSU processome was isolated from
Saccharomyces cerevisiae (Dragon et al., 2002). This com-
plex is required for the processing of 18S rRNA biogenesis. It
contains the U3 snoRNA, 17 U3 proteins (Utp1–17), 10
U3 snoRNA-associated proteins, Rrp5 protein and five small
subunit ribosomal proteins (Dragon et al., 2002). Further
genetic dissection of the yeast U3 snoRNP identified the Utp
complex, which is formed in the absence of U3 and interacts
directly with the rDNA 5¢ ETS region just before the assem-
bly of U3 snoRNP with the pre-rRNA (Gallagher et al., 2004).
Several pre-rRNA endonucleolytic cleavage events in the
nucleolus require U3 snoRNA and lead selectively to the
generation of 18S rRNA (Hughes and Ares, 1991; Beltrame
and Tollervey, 1992, 1995). The U3 RNA contains the
consensus C and D box, and is thus classified in the C/D
box family of snoRNA. It therefore associates with the
common C/D snoRNP proteins like Snu13p, Nop56p, Nop58
and Nop1/fibrillarin (Hartshorne and Agabian, 1994; Lafon-
taine and Tollervey, 1999; Lukowiak et al., 2000; Granneman
et al., 2002; Leary et al., 2004; Clery et al., 2007). The
U3 snoRNP is the only C/D snoRNP for which specific
components have been identified. Indeed, functional and
structural analysis of U3 sequences from diverse organisms
revealed at least five additional short elements, known as
boxes GAC A, A¢, B and C¢. Several reports have clearly
demonstrated that the C/D boxes are required for the
nucleolar localization of U3 snoRNA, whereas the GAC A
and A¢ boxes are essential for directing the specific 18S
pre-rRNA cleavage (Speckmann et al., 1999; Borovjagin and
Gerbi, 2001).
In plants, little is known about the factors involved in
ribosome biosynthesis. Proteomic approaches have been
successfully used to identify ribosomal proteins from the
purified cytosolic ribosome (Giavalisco et al., 2005; Zanetti
et al., 2005), as well as nucleolar proteins in Arabidopsis
(Pendle et al., 2005). In total, 217 nucleolar proteins were
identified in Arabidopsis, including fibrillarin, Nop56, Nop58
and some U3-specific proteins (Pendle et al., 2005). How-
ever, this number is relatively low compared with approx-
imately 4500 proteins identified in the human nucleolus
(Leung et al., 2006; Ahmad et al., 2009), suggesting the
existence of other stable and/or transitory nucleolar proteins
in plants. On the other hand, hundreds of C/D box snoRNAs
have been reported in plants, including U3 snoRNA (Leader
et al., 1997; Barneche et al., 2001; Brown et al., 2003a,b;
Chen et al., 2003), which can be folded into a secondary
structure similar to that from yeast, trypanosome and
xenopus (but not to U3 sequences from other vertebrates)
(Kiss and Solymosy, 1990; Marshallsay et al., 1990). How-
ever, plant U3 sequences display particular features with
respect to U3 snoRNA from yeast and metazoans (Kiss and
Solymosy, 1990; Marshallsay et al., 1990). Indeed, although
the U3 snoRNA structures from plants and other organisms
are conserved, the sequence identity between plant
U3 snoRNA and those of human, xenopus, trypanosome
and yeast is less than 35%. In contrast, sequence identity
between Brassica, Arabidopsis, rice (Oryza sativa), wheat
(Triticum spp.), maize (Zea mays), Solanum and Lotus
japonica (see below for secondary structures) is greater
than 66%. Among all potential U3 snoRNA proteins
described in plants (reviewed recently in Saez-Vasquez and
Medina, 2008), fibrillarin is the best characterized (Pih et al.,
2000; Barneche et al., 2001). Interestingly, the homologues
of U3 nucleolar proteins, such as Nop56p, Nop58 and
fibrillarin, were identified in the Arabidopsis nuclear matrix,
together with nucleolin, histone deacetylase, translation
factor eIF-1, two chaperones and other non-nucleolar or
ribosome-related proteins (Calikowski et al., 2003). Never-
theless, in the absence of any experimentation it would be
premature to propose any functional relationship between
U3 snoRNP and other non-nucleolar proteins.
Previously, a large U3 snoRNP (NFD, renamed in this
paper as BoU3 snoRNP) was described in our laboratory
after its purification from cauliflower (Brassica oleracea var.
botrytis) inflorescence. It was characterized by its capacity to
bind pre-rRNA and accurately produce the first cleavage of
the pre-rRNA in vitro (Saez-Vasquez et al., 2004a,b). Inter-
estingly, the Brassica U3 snoRNP also binds to rDNA in vitro.
Indeed, the BoU3 snoRNP complex was purified based on its
sequence-specific rDNA binding activity to a double-
stranded rDNA cluster motif, A123B (see Figure 1b), located
just upstream from the primary cleavage site in the pre-RNA
5¢ ETS (Caparros-Ruiz et al., 1997). Thus, the BoU3 snoRNP
most probably links RNA polI transcription to processing
and maturation of pre-RNA. This might occur by a mecha-
nism in which the U3 snoRNP processing complex first
384 Hala Samaha et al.
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2010), 61, 383–398
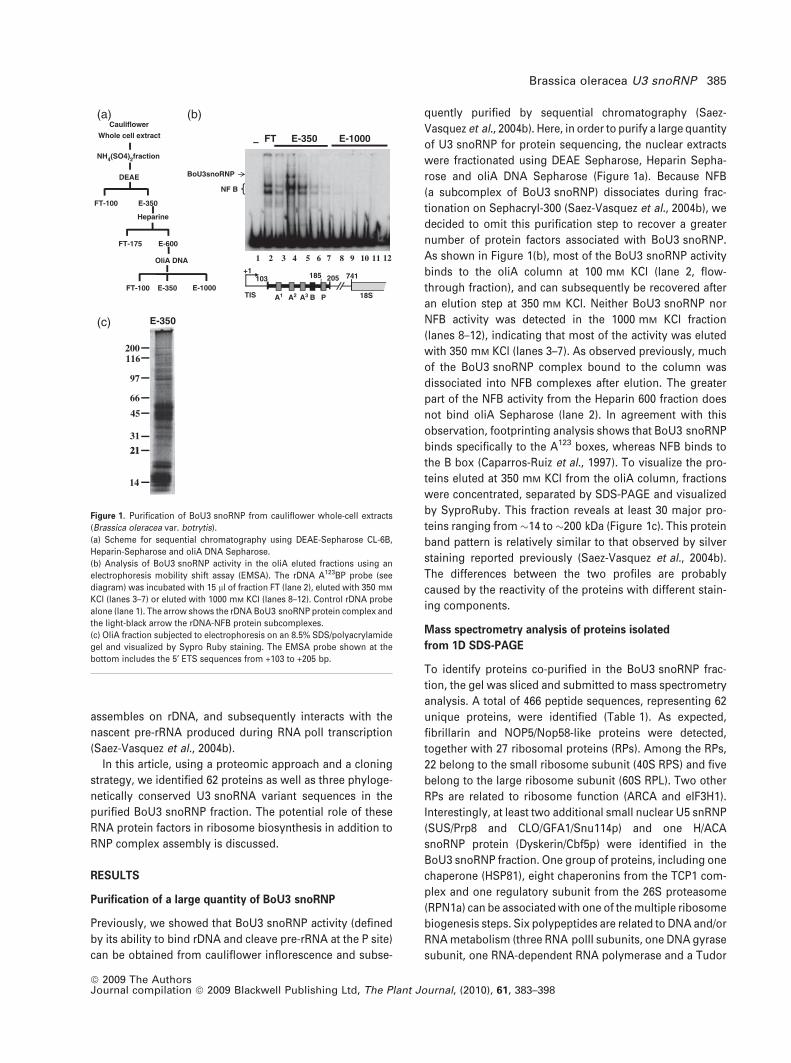
assembles on rDNA, and subsequently interacts with the
nascent pre-rRNA produced during RNA polI transcription
(Saez-Vasquez et al., 2004b).
In this article, using a proteomic approach and a cloning
strategy, we identified 62 proteins as well as three phyloge-
netically conserved U3 snoRNA variant sequences in the
purified BoU3 snoRNP fraction. The potential role of these
RNA protein factors in ribosome biosynthesis in addition to
RNP complex assembly is discussed.
RESULTS
Purification of a large quantity of BoU3 snoRNP
Previously, we showed that BoU3 snoRNP activity (defined
by its ability to bind rDNA and cleave pre-rRNA at the P site)
can be obtained from cauliflower inflorescence and subse-
quently purified by sequential chromatography (Saez-
Vasquez et al., 2004b). Here, in order to purify a large quantity
of U3 snoRNP for protein sequencing, the nuclear extracts
were fractionated using DEAE Sepharose, Heparin Sepha-
rose and oliA DNA Sepharose (Figure 1a). Because NFB
(a subcomplex of BoU3 snoRNP) dissociates during frac-
tionation on Sephacryl-300 (Saez-Vasquez et al., 2004b), we
decided to omit this purification step to recover a greater
number of protein factors associated with BoU3 snoRNP.
As shown in Figure 1(b), most of the BoU3 snoRNP activity
binds to the oliA column at 100 mM KCl (lane 2, flow-
through fraction), and can subsequently be recovered after
an elution step at 350 mM KCl. Neither BoU3 snoRNP nor
NFB activity was detected in the 1000 mM KCl fraction
(lanes 8–12), indicating that most of the activity was eluted
with 350 mM KCl (lanes 3–7). As observed previously, much
of the BoU3 snoRNP complex bound to the column was
dissociated into NFB complexes after elution. The greater
part of the NFB activity from the Heparin 600 fraction does
not bind oliA Sepharose (lane 2). In agreement with this
observation, footprinting analysis shows that BoU3 snoRNP
binds specifically to the A123 boxes, whereas NFB binds to
the B box (Caparros-Ruiz et al., 1997). To visualize the pro-
teins eluted at 350 mM KCl from the oliA column, fractions
were concentrated, separated by SDS-PAGE and visualized
by SyproRuby. This fraction reveals at least 30 major pro-
teins ranging from�14 to �200 kDa (Figure 1c). This protein
band pattern is relatively similar to that observed by silver
staining reported previously (Saez-Vasquez et al., 2004b).
The differences between the two profiles are probably
caused by the reactivity of the proteins with different stain-
ing components.
Mass spectrometry analysis of proteins isolated
from 1D SDS-PAGE
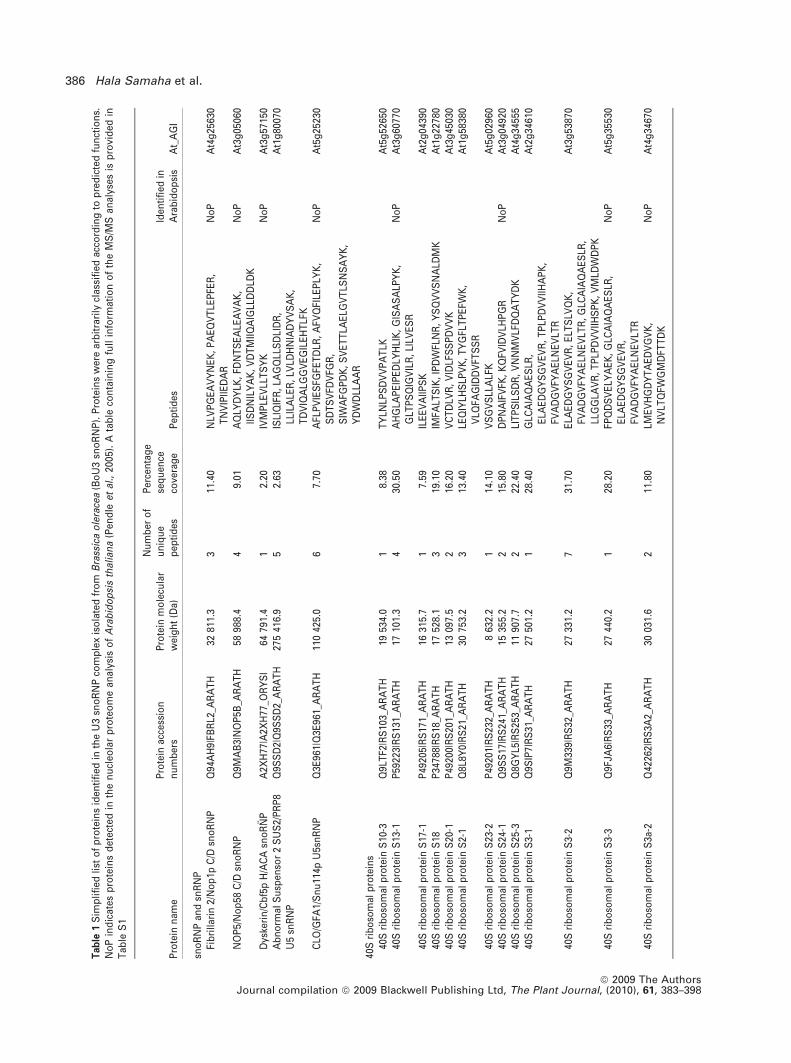
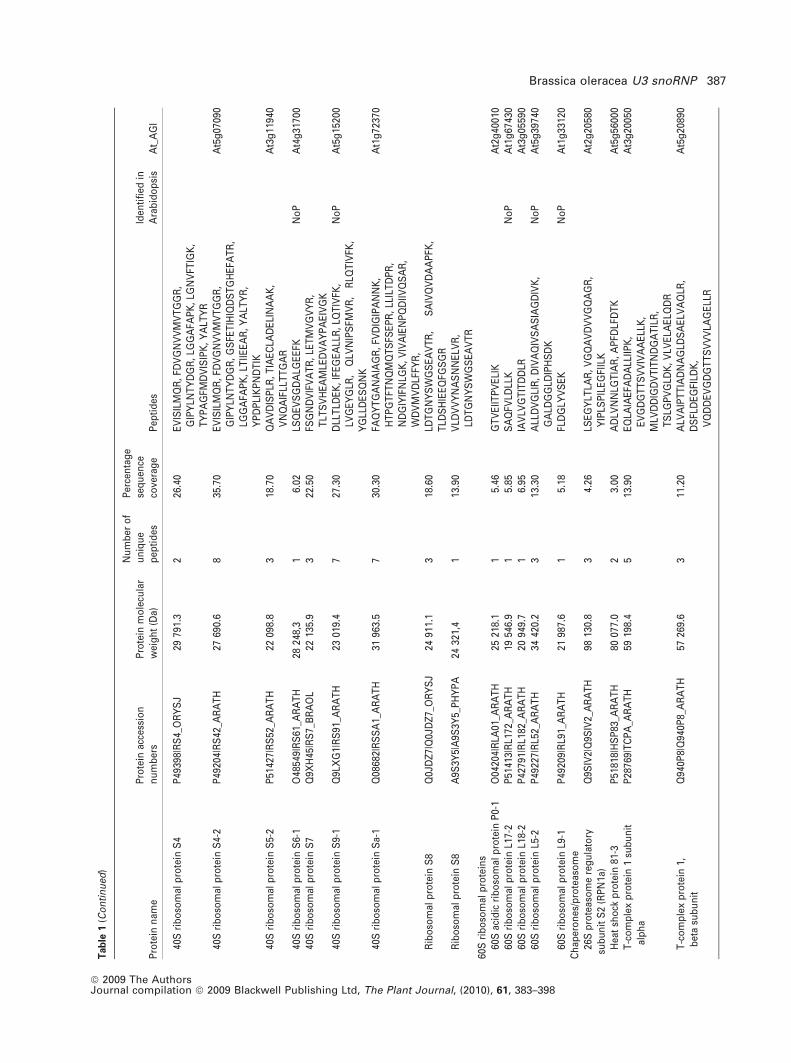
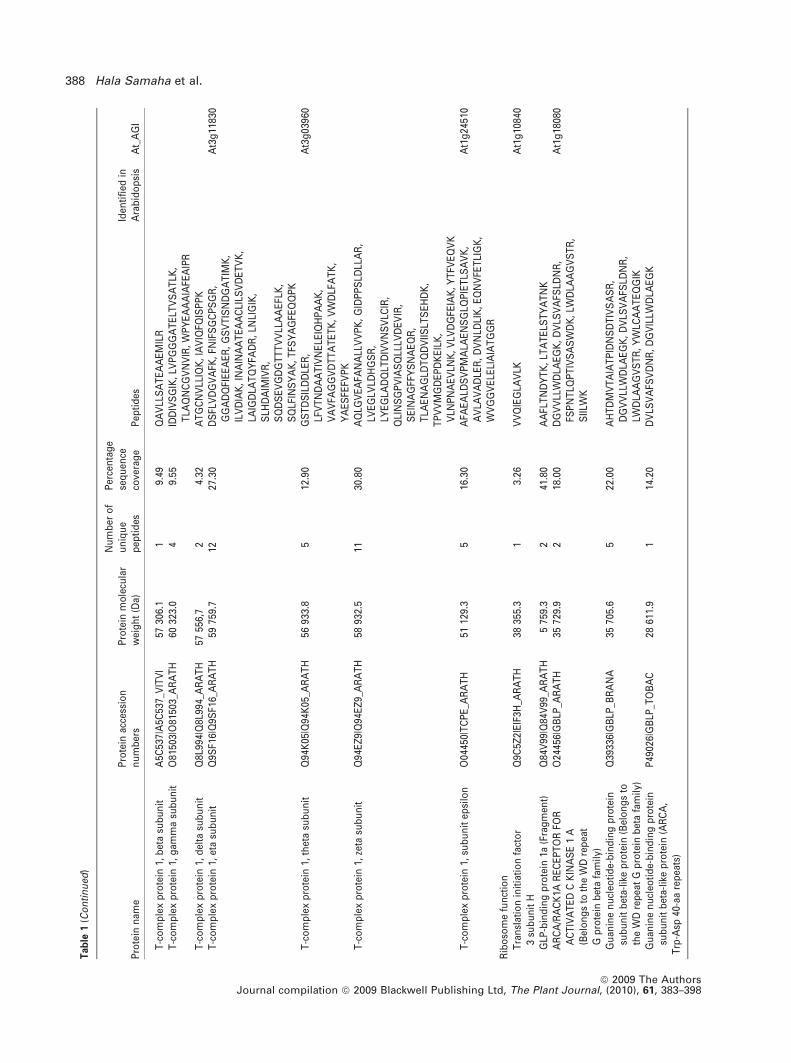
To identify proteins co-purified in the BoU3 snoRNP frac-
tion, the gel was sliced and submitted to mass spectrometry
analysis. A total of 466 peptide sequences, representing 62
unique proteins, were identified (Table 1). As expected,
fibrillarin and NOP5/Nop58-like proteins were detected,
together with 27 ribosomal proteins (RPs). Among the RPs,
22 belong to the small ribosome subunit (40S RPS) and five
belong to the large ribosome subunit (60S RPL). Two other
RPs are related to ribosome function (ARCA and eIF3H1).
Interestingly, at least two additional small nuclear U5 snRNP
(SUS/Prp8 and CLO/GFA1/Snu114p) and one H/ACA
snoRNP protein (Dyskerin/Cbf5p) were identified in the
BoU3 snoRNP fraction. One group of proteins, including one
chaperone (HSP81), eight chaperonins from the TCP1 com-
plex and one regulatory subunit from the 26S proteasome
(RPN1a) can be associated with one of the multiple ribosome
biogenesis steps. Six polypeptides are related to DNA and/or
RNA metabolism (three RNA polII subunits, one DNA gyrase
subunit, one RNA-dependent RNA polymerase and a Tudor
Cauliflower
Whole cell extract
DEAE
NH4(SO4)2fraction
–
BoU3snoRNP
NF B
Heparine
FT-175
FT-100
E-600
FT-100 E-1000E-350
OliA DNA
FT E-350 E-1000
E-350
1 2 3 4 5 6 7 8 9 10 11 12
A1 A2 A3 B P 18S
103 741+1
185
TIS
205
E-350
116
97
200
66
45
31
2121
14
(a) (b)
(c)
Figure 1. Purification of BoU3 snoRNP from cauliflower whole-cell extracts
(Brassica oleracea var. botrytis).
(a) Scheme for sequential chromatography using DEAE-Sepharose CL-6B,
Heparin-Sepharose and oliA DNA Sepharose.
(b) Analysis of BoU3 snoRNP activity in the oliA eluted fractions using an
electrophoresis mobility shift assay (EMSA). The rDNA A123BP probe (see
diagram) was incubated with 15 ll of fraction FT (lane 2), eluted with 350 mM
KCl (lanes 3–7) or eluted with 1000 mM KCl (lanes 8–12). Control rDNA probe
alone (lane 1). The arrow shows the rDNA BoU3 snoRNP protein complex and
the light-black arrow the rDNA-NFB protein subcomplexes.
(c) OliA fraction subjected to electrophoresis on an 8.5% SDS/polyacrylamide
gel and visualized by Sypro Ruby staining. The EMSA probe shown at the
bottom includes the 5¢ ETS sequences from +103 to +205 bp.
Brassica oleracea U3 snoRNP 385
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2010), 61, 383–398
Tab
le1
Sim
plifi
edlis
to
fp
rote
ins
iden
tifi
edin
the
U3
sno
RN
Pco
mp
lex
iso
late
dfr
om
Bra
ssic
ao
lera
cea
(Bo
U3
sno
RN
P).
Pro
tein
sw
ere
arb
itra
rily
clas
sifi
edac
cord
ing
top
red
icte
dfu
nct
ion
s.N
oP
ind
icat
esp
rote
ins
det
ecte
din
the
nu
cleo
lar
pro
teo
me
anal
ysis
of
Ara
bid
op
sis
thal
ian
a(P
end
leet
al.,
2005
).A
tab
leco
nta
inin
gfu
llin
form
atio
no
fth
eM
S/M
San
alys
esis
pro
vid
edin
Tab
leS
1
Pro
tein
nam
eP
rote
inac
cess
ion
nu
mb
ers
Pro
tein
mo
lecu
lar
wei
gh
t(D
a)
Nu
mb
ero
fu
niq
ue
pep
tid
es
Per
cen
tag
ese
qu
ence
cove
rag
eP
epti
des
Iden
tifi
edin
Ara
bid
op
sis
At_
AG
I
sno
RN
Pan
dsn
RN
PFi
bri
llari
n2/
No
p1p
C/D
sno
RN
PQ
94A
H9|
FBR
L2_A
RA
TH
3281
1.3
311
.40
NLV
PG
EA
VY
NE
K,
PA
EQ
VT
LEP
FER
,T
NV
IPIIE
DA
RN
oP
At4
g25
630
NO
P5/
No
p58
C/D
sno
RN
PQ
9MA
B3|
NO
P5B
_AR
AT
H58
988.
44
9.01
AQ
LYD
YLK
,FD
NT
SE
ALE
AV
AK
,IIS
DN
ILY
AK
,V
DT
MIIQ
AIG
LLD
DLD
KN
oP
At3
g05
060
Dys
keri
n/C
bf5
pH
/AC
Asn
oR
NP
A2X
H77
|A2X
H77
_OR
YS
I64
791.
41
2.20
IVM
PLE
VLL
TS
YK
No
PA
t3g
5715
0A
bn
orm
alS
usp
enso
r2
SU
S2/
PR
P8
U5
snR
NP
Q9S
SD
2|Q
9SS
D2_
AR
AT
H27
541
6.9
52.
63IS
LIQ
IFR
,LA
GQ
LLS
DLI
DR
,LL
ILA
LER
,LV
LDH
NIA
DY
VS
AK
,T
DV
IQA
LGG
VE
GIL
EH
TLF
K
At1
g80
070
CLO
/GFA
1/S
nu
114p
U5s
nR
NP
Q3E
961|
Q3E
961_
AR
AT
H11
042
5.0
67.
70A
FLP
VIE
SFG
FET
DLR
,A
FVQ
FILE
PLY
K,
SD
TS
VFD
VFG
R,
SIW
AFG
PD
K,
SV
ET
TLA
ELG
VT
LSN
SA
YK
,Y
DW
DLL
AA
R
No
PA
t5g
2523
0
40S
rib
oso
mal
pro
tein
s40
Sri
bo
som
alp
rote
inS
10-3
Q9L
TF2
|RS
103_
AR
AT
H19
534.
01
8.38
TY
LNLP
SD
VV
PA
TLK
At5
g52
650
40S
rib
oso
mal
pro
tein
S13
-1P
5922
3|R
S13
1_A
RA
TH
1710
1.3
430
.50
AH
GLA
PE
IPE
DLY
HLI
K,
GIS
AS
ALP
YK
,G
LTP
SQ
IGV
ILR
,LI
LVE
SR
No
PA
t3g
6077
0
40S
rib
oso
mal
pro
tein
S17
-1P
4920
5|R
S17
1_A
RA
TH
1631
5.7
17.
59IL
EE
VA
IIPS
KA
t2g
0439
040
Sri
bo
som
alp
rote
inS
18P
3478
8|R
S18
_AR
AT
H17
528.
13
19.1
0IM
FALT
SIK
,IP
DW
FLN
R,
YS
QV
VS
NA
LDM
KA
t1g
2278
040
Sri
bo
som
alp
rote
inS
20-1
P49
200|
RS
201_
AR
AT
H13
097.
52
16.2
0V
CT
DLV
R,
VID
LFS
SP
DV
VK
At3
g45
030
40S
rib
oso
mal
pro
tein
S2-
1Q
8L8Y
0|R
S21
_AR
AT
H30
753.
23
13.4
0LE
QIY
LHS
LPV
K,
TY
GFL
TP
EFW
K,
VLQ
FAG
IDD
VFT
SS
RA
t1g
5838
0
40S
rib
oso
mal
pro
tein
S23
-2P
4920
1|R
S23
2_A
RA
TH
863
2.2
114
.10
VS
GV
SLL
ALF
KA
t5g
0296
040
Sri
bo
som
alp
rote
inS
24-1
Q9S
S17
|RS
241_
AR
AT
H15
355.
22
15.8
0D
PN
AIF
VFK
,K
QFV
IDV
LHP
GR
No
PA
t3g
0492
040
Sri
bo
som
alp
rote
inS
25-3
Q8G
YL5
|RS
253_
AR
AT
H11
907.
72
22.4
0LI
TP
SIL
SD
R,
VN
NM
VLF
DQ
AT
YD
KA
t4g
3455
540
Sri
bo
som
alp
rote
inS
3-1
Q9S
IP7|
RS
31_A
RA
TH
2750
1.2
128
.40
GLC
AIA
QA
ES
LR,
ELA
ED
GY
SG
VE
VR
,T
PLP
DV
VIIH
AP
K,
FVA
DG
VFY
AE
LNE
VLT
R
At2
g34
610
40S
rib
oso
mal
pro
tein
S3-
2Q
9M33
9|R
S32
_AR
AT
H27
331.
27
31.7
0E
LAE
DG
YS
GV
EV
R,
ELT
SLV
QK
,FV
AD
GV
FYA
ELN
EV
LTR
,G
LCA
IAQ
AE
SLR
,LL
GG
LAV
R,T
PLP
DV
VIIH
SP
K,V
MLD
WD
PK
At3
g53
870
40S
rib
oso
mal
pro
tein
S3-
3Q
9FJA
6|R
S33
_AR
AT
H27
440.
21
28.2
0FP
QD
SV
ELY
AE
K,
GLC
AIA
QA
ES
LR,
ELA
ED
GY
SG
VE
VR
,FV
AD
GV
FYA
ELN
EV
LTR
No
PA
t5g
3553
0
40S
rib
oso
mal
pro
tein
S3a
-2Q
4226
2|R
S3A
2_A
RA
TH
3003
1.6
211
.80
LME
VH
GD
YT
AE
DV
GV
K,
NV
LTQ
FWG
MD
FTT
DK
No
PA
t4g
3467
0
386 Hala Samaha et al.
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2010), 61, 383–398
Tab
le1
(Co
nti
nu
ed)
Pro
tein
nam
eP
rote
inac
cess
ion
nu
mb
ers
Pro
tein
mo
lecu
lar
wei
gh
t(D
a)
Nu
mb
ero
fu
niq
ue
pep
tid
es
Per
cen
tag
ese
qu
ence
cove
rag
eP
epti
des
Iden
tifi
edin
Ara
bid
op
sis
At_
AG
I
40S
rib
oso
mal
pro
tein
S4
P49
398|
RS
4_O
RY
SJ
2979
1.3
226
.40
EV
ISIL
MQ
R,
FDV
GN
VV
MV
TG
GR
,G
IPY
LNT
YD
GR
,LG
GA
FAP
K,
LGN
VFT
IGK
,T
YP
AG
FMD
VIS
IPK
,Y
ALT
YR
40S
rib
oso
mal
pro
tein
S4-
2P
4920
4|R
S42
_AR
AT
H27
690.
68
35.7
0E
VIS
ILM
QR
,FD
VG
NV
VM
VT
GG
R,
GIP
YLN
TY
DG
R,
GS
FET
IHIQ
DS
TG
HE
FAT
R,
LGG
AFA
PK
,LT
IIEE
AR
,Y
ALT
YR
,Y
PD
PLI
KP
ND
TIK
At5
g07
090
40S
rib
oso
mal
pro
tein
S5-
2P
5142
7|R
S52
_AR
AT
H22
098.
83
18.7
0Q
AV
DIS
PLR
,T
IAE
CLA
DE
LIN
AA
K,
VN
QA
IFLL
TT
GA
RA
t3g
1194
0
40S
rib
oso
mal
pro
tein
S6-
1O
4854
9|R
S61
_AR
AT
H28
248,
31
6.02
LSQ
EV
SG
DA
LGE
EFK
No
PA
t4g
3170
040
Sri
bo
som
alp
rote
inS
7Q
9XH
45|R
S7_
BR
AO
L22
135.
93
22.5
0FS
GN
DV
IFV
AT
R,
LET
MV
GV
YR
,T
LTS
VH
EA
MLE
DV
AY
PA
EIV
GK
40S
rib
oso
mal
pro
tein
S9-
1Q
9LX
G1|
RS
91_A
RA
TH
2301
9.4
727
.30
DLL
TLD
EK
,IF
EG
EA
LLR
,LQ
TIV
FK,
LVG
EY
GLR
,Q
LVN
IPS
FMV
R,
RLQ
TIV
FK,
YG
LLD
ES
QN
K
No
PA
t5g
1520
0
40S
rib
oso
mal
pro
tein
Sa-
1Q
0868
2|R
SS
A1_
AR
AT
H31
963.
57
30.3
0FA
QY
TG
AN
AIA
GR
,FV
DIG
IPA
NN
K,
HT
PG
TFT
NQ
MQ
TS
FSE
PR
,LL
ILT
DP
R,
ND
GIY
IFN
LGK
,V
IVA
IEN
PQ
DIIV
QS
AR
,W
DV
MV
DLF
FYR
,
At1
g72
370
Rib
oso
mal
pro
tein
S8
Q0J
DZ
7|Q
0JD
Z7_
OR
YS
J24
911.
13
18.6
0LD
TG
NY
SW
GS
EA
VT
R,
SA
IVQ
VD
AA
PFK
,T
LDS
HIE
EQ
FGS
GR
Rib
oso
mal
pro
tein
S8
A9S
3Y5|
A9S
3Y5_
PH
YP
A24
321,
41
13.9
0V
LDV
VY
NA
SN
NE
LVR
,LD
TG
NY
SW
GS
EA
VT
R60
Sri
bo
som
alp
rote
ins
60S
acid
icri
bo
som
alp
rote
inP
0-1
O04
204|
RLA
01_A
RA
TH
2521
8.1
15.
46G
TV
EIIT
PV
ELI
KA
t2g
4001
060
Sri
bo
som
alp
rote
inL1
7-2
P51
413|
RL1
72_A
RA
TH
1954
6.9
15.
85S
AQ
FVLD
LLK
No
PA
t1g
6743
060
Sri
bo
som
alp
rote
inL1
8-2
P42
791|
RL1
82_A
RA
TH
2094
9.7
16.
95IA
VLV
GT
ITD
DLR
At3
g05
590
60S
rib
oso
mal
pro
tein
L5-2
P49
227|
RL5
2_A
RA
TH
3442
0.2
313
.30
ALL
DV
GLI
R,
DIV
AQ
IVS
AS
IAG
DIV
K,
GA
LDG
GLD
IPH
SD
KN
oP
At5
g39
740
60S
rib
oso
mal
pro
tein
L9-1
P49
209|
RL9
1_A
RA
TH
2198
7.6
15.
18FL
DG
LYV
SE
KN
oP
At1
g33
120
Ch
aper
on
es/p
rote
aso
me
26S
pro
teas
om
ere
gu
lato
rysu
bu
nit
S2
(RP
N1a
)Q
9SIV
2|Q
9SIV
2_A
RA
TH
9813
0.8
34.
26LS
EG
YLT
LAR
,V
GQ
AV
DV
VG
QA
GR
,Y
IPLS
PIL
EG
FIIL
KA
t2g
2058
0
Hea
tsh
ock
pro
tein
81-3
P51
818|
HS
P83
_AR
AT
H80
077.
02
3.00
AD
LVN
NLG
TIA
R,
AP
FDLF
DT
KA
t5g
5600
0T
-co
mp
lex
pro
tein
1su
bu
nit
alp
ha
P28
769|
TC
PA
_AR
AT
H59
198.
45
13.9
0E
QLA
IAE
FAD
ALL
IIPK
,E
VG
DG
TT
SV
VIV
AA
ELL
K,
MLV
DD
IGD
VT
ITN
DG
AT
ILR
,T
SLG
PV
GLD
K,
VLV
ELA
ELQ
DR
At3
g20
050
T-c
om
ple
xp
rote
in1,
bet
asu
bu
nit
Q94
0P8|
Q94
0P8_
AR
AT
H57
269.
63
11.2
0A
LVA
IPT
TIA
DN
AG
LDS
AE
LVA
QLR
,D
SFL
DE
GFI
LDK
,V
QD
DE
VG
DG
TT
SV
VV
LAG
ELL
R
At5
g20
890
Brassica oleracea U3 snoRNP 387
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2010), 61, 383–398
Tab
le1
(Co
nti
nu
ed)
Pro
tein
nam
eP
rote
inac
cess
ion
nu
mb
ers
Pro
tein
mo
lecu
lar
wei
gh
t(D
a)
Nu
mb
ero
fu
niq
ue
pep
tid
es
Per
cen
tag
ese
qu
ence
cove
rag
eP
epti
des
Iden
tifi
edin
Ara
bid
op
sis
At_
AG
I
T-c
om
ple
xp
rote
in1,
bet
asu
bu
nit
A5C
537|
A5C
537_
VIT
VI
5730
6.1
19.
49Q
AV
LLS
AT
EA
AE
MIL
RT
-co
mp
lex
pro
tein
1,g
amm
asu
bu
nit
O81
503|
O81
503_
AR
AT
H60
323.
04
9.55
IDD
IVS
GIK
,LV
PG
GG
AT
ELT
VS
AT
LK,
TLA
QN
CG
VN
VIR
,W
PY
EA
AA
IAFE
AIP
RT
-co
mp
lex
pro
tein
1,d
elta
sub
un
itQ
8L99
4|Q
8L99
4_A
RA
TH
5755
6,7
24.
32A
TG
CN
VLL
IQK
,IA
VIQ
FQIS
PP
KT
-co
mp
lex
pro
tein
1,et
asu
bu
nit
Q9S
F16|
Q9S
F16_
AR
AT
H59
759.
712
27.3
0D
SFL
VD
GV
AFK
,FN
IFS
GC
PS
GR
,G
GA
DQ
FIE
EA
ER
,G
SV
TIS
ND
GA
TIM
K,
ILV
DIA
K,
INA
INA
AT
EA
AC
LILS
VD
ET
VK
,LA
IGD
LAT
QY
FAD
R,
LNLI
GIK
,S
LHD
AIM
IVR
,S
QD
SE
VG
DG
TT
TV
VLL
AA
EFL
K,
SQ
LFIN
SY
AK
,T
FSY
AG
FEQ
QP
K
At3
g11
830
T-c
om
ple
xp
rote
in1,
thet
asu
bu
nit
Q94
K05
|Q94
K05
_AR
AT
H56
933.
85
12.9
0G
ST
DS
ILD
DLE
R,
LFV
TN
DA
AT
IVN
ELE
IQH
PA
AK
,V
AV
FAG
GV
DT
TA
TE
TK
,V
WD
LFA
TK
,Y
AE
SFE
FVP
K
At3
g03
960
T-c
om
ple
xp
rote
in1,
zeta
sub
un
itQ
94E
Z9|
Q94
EZ
9_A
RA
TH
5893
2.5
1130
.80
AQ
LGV
EA
FAN
ALL
VV
PK
,G
IDP
PS
LDLL
AR
,LV
EG
LVLD
HG
SR
,LY
EG
LAD
QLT
DIV
VN
SV
LCIR
,Q
LIN
SG
PV
IAS
QLL
LVD
EV
IR,
SE
INA
GFF
YS
NA
EQ
R,
TLA
EN
AG
LDT
QD
VIIS
LTS
EH
DK
,T
PV
VM
GD
EP
DK
EIL
K,
VLN
PN
AE
VLN
K,V
LVD
GFE
IAK
,YT
FVE
QV
KT
-co
mp
lex
pro
tein
1,su
bu
nit
epsi
lon
O04
450|
TC
PE
_AR
AT
H51
129.
35
16.3
0A
FAE
ALD
SV
PM
ALA
EN
SG
LQP
IET
LSA
VK
,A
VLA
VA
DLE
R,
DV
NLD
LIK
,E
QN
VFE
TLI
GK
,W
VG
GV
ELE
LIA
IAT
GG
R
At1
g24
510
Rib
oso
me
fun
ctio
nT
ran
slat
ion
init
iati
on
fact
or
3su
bu
nit
HQ
9C5Z
2|E
IF3H
_AR
AT
H38
355.
31
3.26
VV
QIE
GLA
VLK
At1
g10
840
GLP
-bin
din
gp
rote
in1a
(Fra
gm
ent)
Q84
V99
|Q84
V99
_AR
AT
H5
759.
32
41.8
0A
AFL
TN
DY
TK
,LT
AT
ELS
TY
AT
NK
AR
CA
/RA
CK
1AR
EC
EP
TO
RFO
RA
CT
IVA
TE
DC
KIN
AS
E1
A(B
elo
ng
sto
the
WD
rep
eat
Gp
rote
inb
eta
fam
ily)
O24
456|
GB
LP_A
RA
TH
3572
9.9
218
.00
DG
VV
LLW
DLA
EG
K,
DV
LSV
AFS
LDN
R,
FSP
NT
LQP
TIV
SA
SW
DK
,LW
DLA
AG
VS
TR
,S
IILW
K
At1
g18
080
Gu
anin
en
ucl
eoti
de-
bin
din
gp
rote
insu
bu
nit
bet
a-lik
ep
rote
in(B
elo
ng
sto
the
WD
rep
eat
Gp
rote
inb
eta
fam
ily)
Q39
336|
GB
LP_B
RA
NA
3570
5.6
522
.00
AH
TD
MV
TA
IAT
PID
NS
DT
IVS
AS
R,
DG
VV
LLW
DLA
EG
K,
DV
LSV
AFS
LDN
R,
LWD
LAA
GV
ST
R,
YW
LCA
AT
EQ
GIK
Gu
anin
en
ucl
eoti
de-
bin
din
gp
rote
insu
bu
nit
bet
a-lik
ep
rote
in(A
RC
A,
Trp
-Asp
40-a
are
pea
ts)
P49
026|
GB
LP_T
OB
AC
2861
1.9
114
.20
DV
LSV
AFS
VD
NR
,D
GV
ILLW
DLA
EG
K
388 Hala Samaha et al.
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2010), 61, 383–398

Tab
le1
(Co
nti
nu
ed)
Pro
tein
nam
eP
rote
inac
cess
ion
nu
mb
ers
Pro
tein
mo
lecu
lar
wei
gh
t(D
a)
Nu
mb
ero
fu
niq
ue
pep
tid
es
Per
cen
tag
ese
qu
ence
cove
rag
eP
epti
des
Iden
tifi
edin
Ara
bid
op
sis
At_
AG
I
Pu
tati
veu
nch
arac
teri
zed
pro
tein
WD
40d
om
ain
(Po
pu
lus
tric
ho
carp
a)A
9P92
6|A
9P92
6_P
OP
TR
3587
2.0
112
.80
DG
VIL
LWD
LAE
GK
,FS
PN
TLQ
PT
IVS
AS
WD
K
DN
A/R
NA
rela
ted
acti
viti
esT
ud
or
do
mai
n-c
on
tain
ing
pro
tein
(TS
N)
Q8V
ZG
7|Q
8VZ
G7_
AR
AT
H11
466
6.8
33.
81D
LGLE
LVE
NG
LAK
,IP
AV
VE
YV
LSG
HR
,T
NV
AT
VLL
EA
GLA
KA
t5g
0735
0
RN
A-d
irec
ted
RN
Ap
oly
mer
ase
O82
189|
O82
189_
AR
AT
H10
263
7.3
10.
88Y
GV
FFP
QK
At2
g19
920
DN
A-d
irec
ted
RN
Ap
oly
mer
ase
IIsu
bu
nit
RP
B1
P18
616|
RP
B1_
AR
AT
H27
721.
71
4.58
NA
TLF
FNIL
LRA
t4g
3580
0
DN
A-d
irec
ted
RN
Ap
oly
mer
ase
IIsu
bu
nit
RP
B2
P38
420|
RP
B2_
AR
AT
H13
983.
02
16.8
0LD
LAG
PLL
GG
LFR
,LL
ICA
LGR
At4
g21
710
DN
A-d
irec
ted
RN
Ap
oly
mer
ase
IIsu
bu
nit
RP
B3-
AQ
3921
1|R
PB
3A_A
RA
TH
3543
4.0
14.
08A
SQ
LVLN
AID
LLK
At2
g15
430
DN
Ag
yras
esu
bu
nit
AQ
9CA
F6|G
YR
A_A
RA
TH
104
522.
66
8.42
ELL
QA
FID
FR,
FHP
HG
DT
AV
YD
SLV
R,
FTD
ES
SS
LTE
QIT
K,
GG
DP
ALV
LNN
LYR
,LI
EQ
EA
IELK
,N
AA
GT
PLV
QIL
SM
SE
GE
R
At3
g10
690
Oth
ers
Tu
bu
linal
ph
a-2/
alp
ha-
4ch
ain
P29
510|
TB
A2_
AR
AT
H49
556.
84
11.1
0E
IVD
LCLD
R,
FDLM
YA
K,
LVS
QV
ISS
LTA
SLR
,T
VG
GG
DD
AFN
TFF
SE
TG
AG
KA
t1g
0482
0
Tu
bu
linb
eta-
4ch
ain
P24
636|
TB
B4_
AR
AT
H49
639.
78
22.5
0A
VLM
DLE
PG
TM
DS
LR,
FPG
QLN
SD
LR,
GH
YT
EG
AE
LID
SV
LDV
VR
,LA
VN
LIP
FPR
,LH
FFM
VG
FAP
LTS
R,
NS
SY
FVE
WIP
NN
VK
,V
SE
QFT
AM
FR,
YLT
AS
AV
FR
At5
g44
340
Co
ato
mer
sub
un
itg
amm
aQ
0WW
26|C
OP
G_A
RA
TH
9860
5.0
11.
35S
IAT
LAIT
TLL
KA
t4g
3445
0E
nd
og
luca
nas
e17
O81
416|
GU
N17
_AR
AT
H24
575.
81
5.45
QV
DY
ILG
DN
PLR
At4
g02
290
NA
DP
-sp
ecifi
cg
luta
tam
ate
deh
ydro
gen
ase
Q9C
8I0|
Q9C
8I0_
AR
AT
H69
670.
91
2.08
LAG
QFQ
GS
FTG
PR
At1
g33
420
P27
SJ
(Fra
gm
ent)
Q5G
1J7|
Q5G
1J7_
HY
PP
E26
207.
21
4.56
SE
SS
GT
TE
LFT
RP
atel
lin-1
Q56
WK
6|P
AT
L1_A
RA
TH
6402
7.0
11.
75T
FGS
IITS
PR
At1
g72
150
Brassica oleracea U3 snoRNP 389
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2010), 61, 383–398

domain-containing protein or TSN). Finally, only seven
detected proteins were not classed in any of the above
groups. The identified proteins were distributed in seven
classes, as shown in Figure 2. Approximately 79% of the
proteins identified in the BoU3 snoRNP fraction are directly
related to biogenesis and the structure and/or function of the
ribosome (Figure 2 and Discussion).
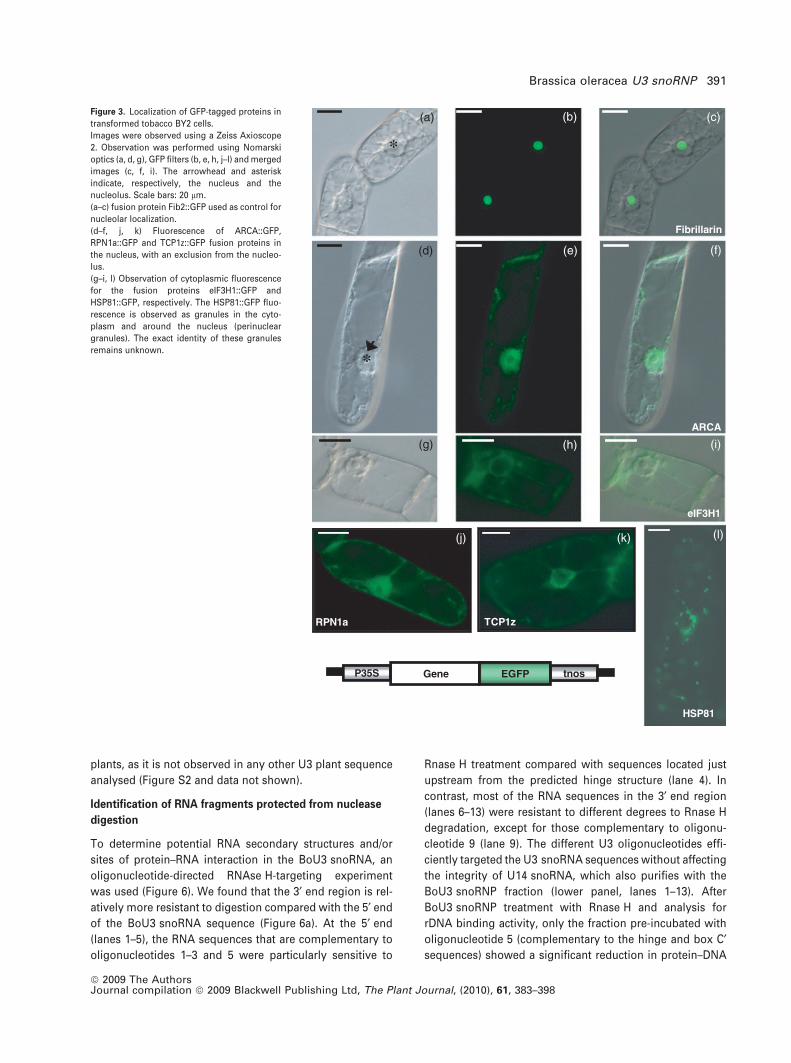
Cellular localization of BoU3 snoRNP_GFP fused proteins
In order to establish a functional connection between ribo-
some biogenesis and the identified BoU3 snoRNP proteins,
the subcellular localization of several proteins was deter-
mined. Fibrillarin/Nop1p, Nop56p, and some small and large
subunit ribosomal proteins (see Table 1) were also identified
in the proteomic analysis of nucleoli purified from Arabid-
opsis cell cultures (Pendle et al., 2005), and in some cases
the nucleolar localization was confirmed using GFP::protein
fusions expressed transiently in Arabidopsis thaliana and
tobacco (Nicotiana tabacum) cells (Barneche et al., 2001;
Pendle et al., 2005; Pontvianne et al., 2007). Our goal was to
search for novel proteins that could play a role in certain
nuclear and/or nucleolar steps of plant ribosome biogenesis.
We examined the cellular localization of HSP81, TCP, RPN1a
and eIF3H1 (Figures 3 and S3) subunits because the coun-
terparts in archaeon and yeast cells have been implicated in
pre-rRNA processing and/or ribosomal biogenesis (Ruggero
et al., 1998; Valasek et al., 2001; Schlatter et al., 2002;
Stavreva et al., 2006), but not in those from higher eukary-
otes. Moreover, we tested the cellular localization of ARCA
(Zanetti et al., 2005), a protein associated with the 40S
ribosome subunit (Figure 3). This was carried out by cloning
the cDNA sequence upstream from the enhanced green
fluorescent protein (eGFP) coding sequence to create a
chimeric sequence under the control of the 35S promoter.
The constructs were then transformed stably into BY2 cells.
Fibrillarin was used as a control for nucleolar localization
(Figure 3a–c). Observation of GFP fluorescence by micros-
copy indicates that ARCA, RPN1a and TCP-1z show nuclear
localization with a clear exclusion from the nucleolus (Fig-
ures 3d–f, j, k and S3), whereas eIF3H1 and HSP81 encode
cytoplasmic proteins (Figures 3g–i,l and S3).
Identification and analysis of plant U3 snoRNA sequences
The U3 snoRNAs co-purifying in the BoU3 snoRNP fraction
were cloned by RT-PCR. Sequence analysis of these clones
revealed three conserved and distinct U3 sequences,
named BoU3A, BoU3B and BoU3C (Figure 4). As shown in
Figure 4(b), the BoU3 sequences can be folded into a
similar triple stem-loop structure as that proposed for
Arabidopsis U3 snoRNA (Marshallsay et al., 1990). In all
sequences we identified the characteristic GAC A¢, A, B, C
and D boxes as observed in the U3 sequences from yeast,
trypanosome and vertebrates (Segault et al., 1992; Hart-
shorne and Agabian, 1994; Speckmann et al., 1999; Boro-
vjagin and Gerbi, 2001). However, significant nucleotide
variance between BoU3A, BoU3B and BoU3C is also
observed (Figure 4a,b). Nucleotide changes are observed in
the 5¢-terminal portion (hairpins 1 and 2), the hinge struc-
ture and the 3¢ terminal portion (hairpins 3 and 4).
Remarkably, we observed considerable variation in the
short stem structure located downstream from the C box
(hairpin 4). However, all the variations are structurally
compensatory and, consequently, the hairpin structure is
maintained (Figure 4b).
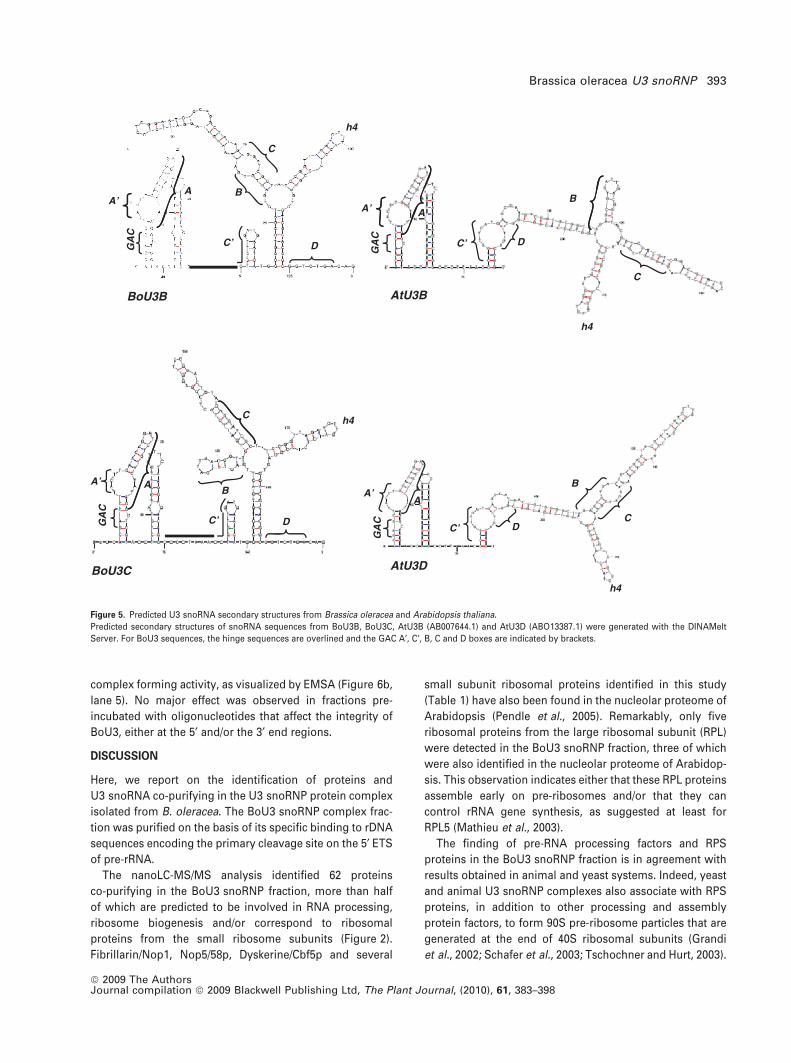
Phylogenetic analysis showed that the BoU3 sequences
are classified in the U3 group from dicotyledonous plants
(Figure S1). Furthermore, it is probable that U3 sequences
from Brassica and Arabidopsis diverged from a recent
duplication event in dicotyledonous plants. Moreover, anal-
ysis of U3 sequences folded using the DINAMelt Server
(Markham and Zuker, 2005) revealed a conserved two-U3
structural domain from both monocotyledonous and dicot-
yledonous plants (Figures 5 and S2). Despite a few structural
differences in the 5¢ and central regions, hairpin 4 is
maintained in the large stem loop in the 3¢ terminal region
in all predicted U3 structures (Figures 5 and S2). Another
interesting feature in cruciferous plants emerged from this
analysis. The large stem loop located in the 3¢ end region of
the B. oleracea and A. thaliana U3 sequences can be folded
into a single stem loop structure with two (BoU3A and
BoU3B) or three (BoU3C) hairpin structures. Interestingly,
the cruciform structure seems to be specific to cruciferous
DNA/RNAactivities
10%
Others11%
snoRNP/snRNP
8%
40S Ribosomalproteins
35%
Ribosomefunction
10%
60S Ribosomalproteins
8%
Chaperones/proteasome
18%
Figure 2. Distribution of the proteins identified
in the BoU3 snoRNP fraction according to the
biological process to which they are assigned: 22
small subunit ribosomal proteins (RPSs); five
large subunit ribosomal proteins (RPLs); ten
chaperones/proteasomes (one HSP 81, eight
TCP subunits and one RPN1a); five snoRNA/
snRNP (three small snoRNPs and two small
nuclear snRNPs); six ribosome function-related
proteins (ARCA and eIF3H1); six proteins related
to DNA and/or RNA metabolism (RNA polII
subunits, DNA gyrase, TSN and RdRP); and
seven unclassified polypeptides.
390 Hala Samaha et al.
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2010), 61, 383–398
plants, as it is not observed in any other U3 plant sequence
analysed (Figure S2 and data not shown).
Identification of RNA fragments protected from nuclease
digestion
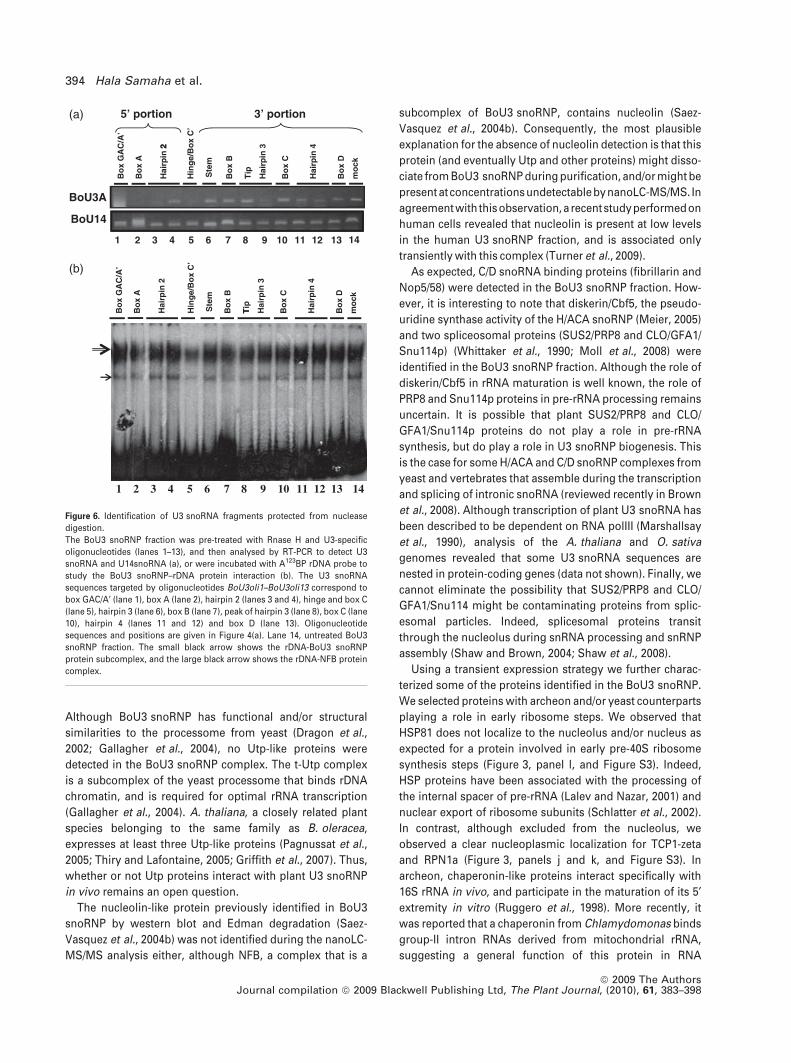
To determine potential RNA secondary structures and/or
sites of protein–RNA interaction in the BoU3 snoRNA, an
oligonucleotide-directed RNAse H-targeting experiment
was used (Figure 6). We found that the 3¢ end region is rel-
atively more resistant to digestion compared with the 5¢ end
of the BoU3 snoRNA sequence (Figure 6a). At the 5¢ end
(lanes 1–5), the RNA sequences that are complementary to
oligonucleotides 1–3 and 5 were particularly sensitive to
Rnase H treatment compared with sequences located just
upstream from the predicted hinge structure (lane 4). In
contrast, most of the RNA sequences in the 3¢ end region
(lanes 6–13) were resistant to different degrees to Rnase H
degradation, except for those complementary to oligonu-
cleotide 9 (lane 9). The different U3 oligonucleotides effi-
ciently targeted the U3 snoRNA sequences without affecting
the integrity of U14 snoRNA, which also purifies with the
BoU3 snoRNP fraction (lower panel, lanes 1–13). After
BoU3 snoRNP treatment with Rnase H and analysis for
rDNA binding activity, only the fraction pre-incubated with
oligonucleotide 5 (complementary to the hinge and box C¢sequences) showed a significant reduction in protein–DNA
Fibrillarin
ARCA
eIF3H1
RPN1a
*
*
TCP1z
HSP81
(a)
(d)
(g)
(j) (k) (l)
(i)(h)
(f)(e)
(c)(b)
Gene EGFP tnosP35S
Figure 3. Localization of GFP-tagged proteins in
transformed tobacco BY2 cells.
Images were observed using a Zeiss Axioscope
2. Observation was performed using Nomarski
optics (a, d, g), GFP filters (b, e, h, j–l) and merged
images (c, f, i). The arrowhead and asterisk
indicate, respectively, the nucleus and the
nucleolus. Scale bars: 20 lm.
(a–c) fusion protein Fib2::GFP used as control for
nucleolar localization.
(d–f, j, k) Fluorescence of ARCA::GFP,
RPN1a::GFP and TCP1z::GFP fusion proteins in
the nucleus, with an exclusion from the nucleo-
lus.
(g–i, l) Observation of cytoplasmic fluorescence
for the fusion proteins eIF3H1::GFP and
HSP81::GFP, respectively. The HSP81::GFP fluo-
rescence is observed as granules in the cyto-
plasm and around the nucleus (perinuclear
granules). The exact identity of these granules
remains unknown.
Brassica oleracea U3 snoRNP 391
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2010), 61, 383–398

1 23GAC Box A’ Box A
4 67
89
1011
Hinge BxoBCxoB
Box C
1213
Box D
Hairpin 3
Hairpin 1
Hairpin 2
Hairpin 4
(a)
(b)
Figure 4. Sequences and secondary structures of BoU3 snoRNA.
(a) Sequence alignment of BoU3A, BoU3B and BoU3C clones isolated from BoU3 snoRNP. The GAC A¢, C¢, B, C and D sequences are boxed. Sequences overlined
and numbered (1–13) correspond to oligonucleotides BoU3oli1–BoU3oli13 used in the RNaseH digestion experiments (see Figure 6).
(b) The BoU3 snoRNA structure model derived from BoU3C and obtained manually based on the U3 snoRNA structure from Arabidopsis thaliana. Nucleotide
substitutions and/or insertions/deletions found in BoU3A and BoU3B sequences are indicated by arrows and are set in lower case. The GAC A¢, C¢, B, C and D
sequences are boxed, and the hinge sequences are underlined. Hairpin 4, containing major conservative substitutions, is encircled.
392 Hala Samaha et al.
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2010), 61, 383–398
complex forming activity, as visualized by EMSA (Figure 6b,
lane 5). No major effect was observed in fractions pre-
incubated with oligonucleotides that affect the integrity of
BoU3, either at the 5¢ and/or the 3¢ end regions.
DISCUSSION
Here, we report on the identification of proteins and
U3 snoRNA co-purifying in the U3 snoRNP protein complex
isolated from B. oleracea. The BoU3 snoRNP complex frac-
tion was purified on the basis of its specific binding to rDNA
sequences encoding the primary cleavage site on the 5¢ ETS
of pre-rRNA.
The nanoLC-MS/MS analysis identified 62 proteins
co-purifying in the BoU3 snoRNP fraction, more than half
of which are predicted to be involved in RNA processing,
ribosome biogenesis and/or correspond to ribosomal
proteins from the small ribosome subunits (Figure 2).
Fibrillarin/Nop1, Nop5/58p, Dyskerine/Cbf5p and several
small subunit ribosomal proteins identified in this study
(Table 1) have also been found in the nucleolar proteome of
Arabidopsis (Pendle et al., 2005). Remarkably, only five
ribosomal proteins from the large ribosomal subunit (RPL)
were detected in the BoU3 snoRNP fraction, three of which
were also identified in the nucleolar proteome of Arabidop-
sis. This observation indicates either that these RPL proteins
assemble early on pre-ribosomes and/or that they can
control rRNA gene synthesis, as suggested at least for
RPL5 (Mathieu et al., 2003).
The finding of pre-RNA processing factors and RPS
proteins in the BoU3 snoRNP fraction is in agreement with
results obtained in animal and yeast systems. Indeed, yeast
and animal U3 snoRNP complexes also associate with RPS
proteins, in addition to other processing and assembly
protein factors, to form 90S pre-ribosome particles that are
generated at the end of 40S ribosomal subunits (Grandi
et al., 2002; Schafer et al., 2003; Tschochner and Hurt, 2003).
A’A
C
B
A’ AB
h4
BoU3B AtU3B
C’ DGA
C
GA
C
C’
C
D
h4
C h4
A’ A
C’
B
DGA
C
A’
GA
C
A
C’
B
CD
BoU3C AtU3D
h4
Figure 5. Predicted U3 snoRNA secondary structures from Brassica oleracea and Arabidopsis thaliana.
Predicted secondary structures of snoRNA sequences from BoU3B, BoU3C, AtU3B (AB007644.1) and AtU3D (ABO13387.1) were generated with the DINAMelt
Server. For BoU3 sequences, the hinge sequences are overlined and the GAC A¢, C¢, B, C and D boxes are indicated by brackets.
Brassica oleracea U3 snoRNP 393
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2010), 61, 383–398
Although BoU3 snoRNP has functional and/or structural
similarities to the processome from yeast (Dragon et al.,
2002; Gallagher et al., 2004), no Utp-like proteins were
detected in the BoU3 snoRNP complex. The t-Utp complex
is a subcomplex of the yeast processome that binds rDNA
chromatin, and is required for optimal rRNA transcription
(Gallagher et al., 2004). A. thaliana, a closely related plant
species belonging to the same family as B. oleracea,
expresses at least three Utp-like proteins (Pagnussat et al.,
2005; Thiry and Lafontaine, 2005; Griffith et al., 2007). Thus,
whether or not Utp proteins interact with plant U3 snoRNP
in vivo remains an open question.
The nucleolin-like protein previously identified in BoU3
snoRNP by western blot and Edman degradation (Saez-
Vasquez et al., 2004b) was not identified during the nanoLC-
MS/MS analysis either, although NFB, a complex that is a
subcomplex of BoU3 snoRNP, contains nucleolin (Saez-
Vasquez et al., 2004b). Consequently, the most plausible
explanation for the absence of nucleolin detection is that this
protein (and eventually Utp and other proteins) might disso-
ciate from BoU3 snoRNP during purification, and/or might be
presentatconcentrationsundetectablebynanoLC-MS/MS. In
agreementwith thisobservation,a recentstudyperformedon
human cells revealed that nucleolin is present at low levels
in the human U3 snoRNP fraction, and is associated only
transiently with this complex (Turner et al., 2009).
As expected, C/D snoRNA binding proteins (fibrillarin and
Nop5/58) were detected in the BoU3 snoRNP fraction. How-
ever, it is interesting to note that diskerin/Cbf5, the pseudo-
uridine synthase activity of the H/ACA snoRNP (Meier, 2005)
and two spliceosomal proteins (SUS2/PRP8 and CLO/GFA1/
Snu114p) (Whittaker et al., 1990; Moll et al., 2008) were
identified in the BoU3 snoRNP fraction. Although the role of
diskerin/Cbf5 in rRNA maturation is well known, the role of
PRP8 and Snu114p proteins in pre-rRNA processing remains
uncertain. It is possible that plant SUS2/PRP8 and CLO/
GFA1/Snu114p proteins do not play a role in pre-rRNA
synthesis, but do play a role in U3 snoRNP biogenesis. This
is the case for some H/ACA and C/D snoRNP complexes from
yeast and vertebrates that assemble during the transcription
and splicing of intronic snoRNA (reviewed recently in Brown
et al., 2008). Although transcription of plant U3 snoRNA has
been described to be dependent on RNA polIII (Marshallsay
et al., 1990), analysis of the A. thaliana and O. sativa
genomes revealed that some U3 snoRNA sequences are
nested in protein-coding genes (data not shown). Finally, we
cannot eliminate the possibility that SUS2/PRP8 and CLO/
GFA1/Snu114 might be contaminating proteins from splic-
esomal particles. Indeed, splicesomal proteins transit
through the nucleolus during snRNA processing and snRNP
assembly (Shaw and Brown, 2004; Shaw et al., 2008).
Using a transient expression strategy we further charac-
terized some of the proteins identified in the BoU3 snoRNP.
We selected proteins with archeon and/or yeast counterparts
playing a role in early ribosome steps. We observed that
HSP81 does not localize to the nucleolus and/or nucleus as
expected for a protein involved in early pre-40S ribosome
synthesis steps (Figure 3, panel l, and Figure S3). Indeed,
HSP proteins have been associated with the processing of
the internal spacer of pre-rRNA (Lalev and Nazar, 2001) and
nuclear export of ribosome subunits (Schlatter et al., 2002).
In contrast, although excluded from the nucleolus, we
observed a clear nucleoplasmic localization for TCP1-zeta
and RPN1a (Figure 3, panels j and k, and Figure S3). In
archeon, chaperonin-like proteins interact specifically with
16S rRNA in vivo, and participate in the maturation of its 5¢extremity in vitro (Ruggero et al., 1998). More recently, it
was reported that a chaperonin from Chlamydomonas binds
group-II intron RNAs derived from mitochondrial rRNA,
suggesting a general function of this protein in RNA
2
5’ portion 3’ portion
BoU3A
BoU14
Bo
x G
AC
/A’
Hai
rpin
2
Bo
x B
Tip
Hai
rpin
3
Bo
x C
Hai
rpin
4
Bo
x D
mo
ck
Bo
x A
Hin
ge/
Bo
x C
’
Ste
m
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Bo
x G
AC
/A’
Hin
ge/
Bo
x C
’
Hai
rpin
2
Bo
x B
Tip
Hai
rpin
3
Bo
x C
Hai
rpin
4
Bo
x D
mo
ck
Bo
x A
Ste
m
T
1 2 3 4 5 6 7 8 9 10 11 12 13 14
(a)
(b)
Figure 6. Identification of U3 snoRNA fragments protected from nuclease
digestion.
The BoU3 snoRNP fraction was pre-treated with Rnase H and U3-specific
oligonucleotides (lanes 1–13), and then analysed by RT-PCR to detect U3
snoRNA and U14snoRNA (a), or were incubated with A123BP rDNA probe to
study the BoU3 snoRNP–rDNA protein interaction (b). The U3 snoRNA
sequences targeted by oligonucleotides BoU3oli1–BoU3oli13 correspond to
box GAC/A¢ (lane 1), box A (lane 2), hairpin 2 (lanes 3 and 4), hinge and box C
(lane 5), hairpin 3 (lane 6), box B (lane 7), peak of hairpin 3 (lane 8), box C (lane
10), hairpin 4 (lanes 11 and 12) and box D (lane 13). Oligonucleotide
sequences and positions are given in Figure 4(a). Lane 14, untreated BoU3
snoRNP fraction. The small black arrow shows the rDNA-BoU3 snoRNP
protein subcomplex, and the large black arrow shows the rDNA-NFB protein
complex.
394 Hala Samaha et al.
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2010), 61, 383–398

metabolism (Balczun et al., 2006). Thus, the fact that chap-
eronin-like proteins co-purify in the BoU3 snoRNP indicates
that the TCP1 complex might play a major role in 5¢ ETS
pre-rRNA processing. Moreover, in agreement with the
observed nuclear localization of RPN1, it has been shown
in yeast cells that the inhibition of the proteasome affects
the accumulation of 90S pre-ribosomes, the production of
mature rRNA, and the distribution of early and late pre-rRNA
processing factors (Stavreva et al., 2006).
In Arabidopsis thaliana, mutation of the RPN1a gene
causes embryo lethality (Brukhin et al., 2005). A similar
situation was observed for other protein factors related to
ribosome biogenesis, including the plant mutants for SWA1
and TOZ genes that encode the Arabidopsis Utp15 and
Utp13 proteins (see above) that are both essential for
gametogenesis and embryogenesis (Thiry and Lafontaine,
2005; Griffith et al., 2007). The role of these factors in the
processing of 18S rRNA has been confirmed, at least for
SWA1 (reviewed in Saez-Vasquez and Medina, 2008). On the
other hand, our results describe the nuclear localization of
the ARCA protein (Figure 3, panels d–f). In the ribosome,
ARCA locates on the head of the 40S ribosomal subunit,
where it plays a major role in translation (Giavalisco et al.,
2005; Zanetti et al., 2005). The reason for its nuclear local-
ization remains unclear, but it is known that the repeated
WD40 motifs, also found in ARCA protein sequences, act as a
site for protein–protein interaction and serve as platforms
for the assembly of protein complexes (Miles et al., 2005).
The specific role of TCP1-zeta, RPN1a and ARCA proteins in
ribosome biogenesis in plants remains an open issue.
Among all the proteins identified in this study, the
detection of TSN and RNA-directed RNA polymerase (RdRP)
in the BoU3 snoRNP fraction is particularly intriguing. In
higher eukaryotic cells, the TSN protein is both an RNA and
DNA binding protein, but lacks the nucleolytic active site
residues of Staphylococcal nuclease (Abe et al., 2003).
Interestingly, the Tudor domain from the SMM protein has
been implicated in direct interaction with the GAR domain of
fibrillarin (Jones et al., 2001), suggesting that TSN and
fibrillarin from B. oleracea might interact in vivo, and may
consequently co-purify in the BoU3 snoRNP fraction. On the
other hand, it is well known that RdRPs play a key role in
RNA-mediated gene silencing (Wassenegger and Krczal,
2006). RdRP/RDR2 shows a nucleolar localization, and has
been involved in silencing of rDNA in the epigenetic
phenomenon known as nucleolar dominance (Preuss et al.,
2008). So far no interaction between RdRP and rRNA
processing factors has been reported. However, we can
reasonably hypothesize that processing of pre-rRNA and its
conversion into dsRNA, which directs rDNA silencing, might
share a number of pre-rRNA binding factors.
Another major finding of this study concerns the U3
snoRNA sequences detected in the BoU3 snoRNP fraction
(Figure 4). What is the role of hairpin 4 and U3 snoRNA
variants in the BoU3 snoRNP fraction? The hairpin 4 struc-
ture is present in the U3 sequences from yeast to humans.
Despite the fact that the role of GAC A¢, A, B, C and D boxes is
relatively well established, little is known about the structure
of hairpin 4 in these species (Segault et al., 1992; Hartshorne
and Agabian, 1994; Speckmann et al., 1999). Nevertheless, it
is possible that this hairpin contributes to the recruitment of
specific U3 snoRNA binding proteins. This is the case for an
RNA element flanking the B/C box, which is required for the
optimal binding of the hU3-55K protein (Granneman et al.,
2002). On the other hand, a biform (BoU3A and BoU3B) and
a cruciform secondary structure (BoU3C) were predicted
at the 3¢ end of the BoU3 snoRNA sequences identified
(Figure 5). Remarkably, the BoU3B cruciform structure is
also predicted for some U3 snoRNA sequences from
A. thaliana, but not in other monocotyledons and/or dicoty-
ledons (Figure S2). Thus, the cruciform structure seems to
be specific to ‘cruciferous plants’. Although the functional
significance of these structures in vivo remains unclear,
nuclease protection experiments (Figure 6) indicate that
the 3¢ region of BoU3 is highly structured and/or protected
from digestion by RNA binding factors co-purifying in the
BoU3 snoRNP fraction, in contrast with the 5¢ region.
In conclusion, structural analysis of the functional BoU3
snoRNP complex allowed us to identify novel proteins and
RNA structures that are potentially involved in rRNA synthe-
sis and/or other ribosome assembly steps in plants. Some
of the identified factors are certainly contaminant proteins
co-eluting throughout the biochemical steps of purification,
but it is also clear that many of the identified nuclear proteins
(TCP1-zeta, RPN1a and ARCA) could be related in one way
or another to ribosome biogenesis. Finally, it is clear that
this analysis revealed unexpected protein factors (TSN and
RdrRP) and RNA structures that might play a major role in
RNA synthesis and RNP complex assembly, not only in
plants but also in other higher eukaryotic organisms.
EXPERIMENTAL PROCEDURES
Purification of BoU3 snoRNP
Cauliflower protein extracts precipitated with ammonium sulphatewere fractionated using DEAE-sepharose CL-6B, Heparin sepharoseand oliA DNA sepharose, as described previously (Saez-Vasquezet al., 2004b), and as shown in Figure 1a. The heparin F600 fractionwas dialysed against buffer II (50 mM Tris–HCl, pH 8, 6 mM MgCl2,15% glycerol, 1 mM EDTA and 2% NP40) containing 100 mM KCl andsubjected to oliA DNA sepharose equilibrated with buffer II-100. Thecolumn was washed with buffer II-100 and proteins were eluted withbuffer II-350, and then with buffer II-1000. Fractions were dialysedagainst buffer II-100 and were stored at )80�C. The numbers afterbuffer II and/or fraction F indicate the concentration of KCl to be inthe mM range.
DNA binding activity
For DNA binding assays, the A123BP fragment was 5¢-end labelledusing Klenow fragment and [a-32P]dCTP. Between 5 and 10 fmol of
Brassica oleracea U3 snoRNP 395
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2010), 61, 383–398

gel-purified A123BP fragment was mixed with 15 ll of the purifiedprotein samples. After incubation of the reaction mixtures for20 min on ice, binding products were analysed by electrophoresismobility shift assay (EMSA), as described previously (Saez-Vasquezet al., 2004b).
Rnase H digestion
RNase H treatment was performed according to Kass et al. (1990).Briefly, 15 ll of the purified BoU3 snoRNP fraction were mixed with1 ll (100 lM) of oligonucleotide BoU3Oli1-13 (Appendix S2) and1 ll (5 U ml)1) Rnase H (USB, http://www.usbweb.com). Afterincubation (10 min at 37�C and 20 min at 30�C), the reaction mix-tures were treated with 1 ll (2.2 U ll)1) of Dnase I (Worthington,http://www.worthington-biochem.com) for 3 min at 37�C, followedby 7 min at 30�C. The remaining U3 snoRNAs were analyzed using1 ll of the reaction mix and by performing a one-step RT-PCR with aplatinum Taq system (Invitrogen, http://www.invitrogen.com). Toverify that oligonucleotides BoU3Oli1–BoU3Oli13 specifically directthe cleavage of U3 snoRNA, the integrity of U14 snoRNA treatmentwas controlled using oligonucleotides 5¢-AtU14 and 3¢-AtU14. Totest the BoU3 snoRNP binding activity after the total and/or partialremoval of U3 snoRNA, the S300 peak fraction (Saez-Vasquez et al.,2004b) was mixed with oligonucleotides BoU3Oli1–BoU3Oli13 andRnase H. After 10 min at 37�C and 20 min at 30�C, the reactionmixtures were incubated with the 5¢-end-labelled A123BP fragmentand then analysed by EMSA.
Analysis of U3 snoRNA sequences
BoU3A, BoU3B and BoU3C from the BoU3 snoRNP fractionwere cloned by RT-PCR and then sequenced on a ABI3100 DNAsequencer using an ABI PRISM Big Dye Terminator CycleSequencing Ready Reaction Kit (Applied Biosystems, http://www.appliedbiosystems.com). The other plant U3 snoRNA se-quences (Appendix S1) were obtained from the RNA functionaldatabase (http://www.ncrna.org/frnadb). The phylogenetic tree forU3 snoRNA was generated by CLUSTALW 2.0.10 (Larkin et al., 2007)using the neighbour-joining method coupled with 1000 bootstraptests. U3 snoRNA secondary structures were predicted using theDINAMelt Server DINAMelt web server for nucleic acid meltingprediction (Markham and Zuker, 2005).
Cloning and cellular localization of GFP fusion proteins
Complete cDNA sequences for TCP1 (RAFL_08-18-F20), HSP81(RAFL_09-06-O18), RPN1a (RAFL_09-17-J10), ARCA (RAFL_04-13-D02) and eIF3H1 (RAFL_05-13-M08) were obtained from the RIKENBioresource Center (Yokohama, Japan). Gateway cassettes witheach coding sequence were generated by PCR following Invitro-gen’s instructions. PCR products were introduced first intopDONR207 and then cloned by recombination into pK7FWG2 toproduce TCP1::GFP, HSP81::GFP, RPN1a::GFP, ARCA::GFP,eIF3H1::GFP and FIB2::GFP plasmids. Constructs were introducedinto the Agrobacterium tumefaciens LBA4404 strain, and were usedto transform tobacco BY2 cells as described previously (Chabouteet al., 2000). Observations were performed using Nomarski opticsand a GFP filter with a Zeiss Axioscope 2 microscope (Zeiss, http://www.zeiss.com). Images were taken with a Leica DC350FX camera(Leica, http://www.leica.com).
In-gel digestion and mass spectrometry analysis
In-gel digestion and mass spectrometry analyses were performedon a CapLC (Waters, http://www.waters.com) coupled to a hybrid
quadrupole orthogonal acceleration time-of-flight tandem massspectrometer (Q-TOF 2; Waters), as described in Appendix S3.
Protein identification
The MS and MS/MS data were analysed using a local Mascot server(MASCOT 2.0; MatrixScience, http://www.matrixscience.com) bycomparison with a composite target–decoy database, including theUniProt protein sequences of Viridiplantae (554 832 sequences;January 2009), human keratins, porcine Trypsin and all corre-sponding reversed sequences (1 109 972 entries in total).
Searches were performed with a mass tolerance of 250 ppm inMS mode and 0.4 Da in MS/MS mode for nanoLC-MS/MS data. Onemissed cleavage per peptide was allowed and variable modifica-tions were taken into account, such as carbamidomethylation ofcysteine, oxidation of methionine and N-acetylation (protein N-ter).Neither protein molecular weight nor isoelectric point constrainswere applied (see Figure S4).
Mascot results were loaded in MuDPIT mode into SCAFFOLD
(Proteome Software, http://www.proteomesoftware.com). Resultswere subjected to the following filtering criteria. For the identifica-tion of proteins with two peptides or more, a Mascot ion score ofabove 25 was required. In the case of single peptide hits, the score ofthe unique peptide must be greater (minimal ‘difference score’ of 0)than the 95% Mascot significance threshold. The spectra of thosesingle peptide hits are provided in Figure S5). The target–decoydatabase search allows us to control and estimate the false-positiveidentification rate of our study (Peng et al., 2003; Elias and Gygi,2007). Thus, the final catalogue of proteins presents an estimatedfalse-positive rate of below 1%.
ACKNOWLEDGEMENTS
This work was supported by the Centre National de la RechercheScientifique. FP and HS were supported by fellowships from theMinistere de l’Enseignement et de la Recherche and the Lebanesegovernment, respectively. The authors also thank C. Chaparro forhelp in obtaining U3 snoRNA sequences and in the generation ofthe phylogenetic tree.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the onlineversion of this article:Figure S1. Phylogenetic relationships of different plant U3 snoRNAsequences.Figure S2. Predicted secondary structures of plant snoRNAsequences.Figure S3. Localization of GFP-tagged RPN1a TCP1z and HSP81proteins.Figure S4. Distribution of the errors measured on the precursor ionmasses.Figure S5. MS/MS spectra of single peptide hits.Table S1. Complete list of proteins identified in the U3 snoRNPcomplex isolated from Brassica oleracea.Appendix S1. Sequences used for phylogenetic and structuralanalysis.Appendix S2. Oligonucleotides used in this work.Appendix S3. Mass spectrometry analysis and protein identification.Please note: As a service to our authors and readers, this journalprovides supporting information supplied by the authors. Suchmaterials are peer-reviewed and may be re-organized for onlinedelivery, but are not copy-edited or typeset. Technical supportissues arising from supporting information (other than missingfiles) should be addressed to the authors.
396 Hala Samaha et al.
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2010), 61, 383–398

REFERENCES
Abe, S., Sakai, M., Yagi, K., Hagino, T., Ochi, K., Shibata, K. and Davies, E.
(2003) A Tudor protein with multiple SNc domains from pea seedlings:
cellular localization, partial characterization, sequence analysis, and phy-
logenetic relationships. J. Exp. Bot. 54(384), 971–983.
Ahmad, Y., Boisvert, F.M., Gregor, P., Cobley, A. and Lamond, A.I. (2009)
NOPdb: Nucleolar Proteome Database – 2008 update. Nucleic Acids Res.
37(Database issue), D181–D184.
Balczun, C., Bunse, A., Schwarz, C., Piotrowski, M. and Kuck, U. (2006)
Chloroplast heat shock protein Cpn60 from Chlamydomonas reinhardtii
exhibits a novel function as a group II intron-specific RNA-binding protein.
FEBS Lett. 580(18), 4527–4532.
Barneche, F., Steinmetz, F. and Echeverria, M. (2000) Fibrillarin genes encode
both a conserved nucleolar protein and a novel small nucleolar RNA
involved in ribosomal RNA methylation in Arabidopsis thaliana. J. Biol.
Chem. 275(35), 27212–27220.
Barneche, F., Gaspin, C., Guyot, R. and Echeverria, M. (2001) Identification of
66 box C/D snoRNAs in Arabidopsis thaliana: extensive gene duplications
generated multiple isoforms predicting new ribosomal RNA 2¢-O-methyl-
ation sites. J. Mol. Biol. 311(1), 57–73.
Beltrame, M. and Tollervey, D. (1992) Identification and functional analysis of
two U3 binding sites on yeast pre-ribosomal RNA. EMBO J. 11(4), 1531–
1542.
Beltrame, M. and Tollervey, D. (1995) Base pairing between U3 and the pre-
ribosomal RNA is required for 18S rRNA synthesis. EMBO J. 14(17), 4350–
4356.
Borovjagin, A.V. and Gerbi, S.A. (2001) Xenopus U3 snoRNA GAC-Box A’ and
Box A sequences play distinct functional roles in rRNA processing. Mol.
Cell. Biol. 21(18), 6210–6221.
Brown, J.W., Echeverria, M. and Qu, L.H. (2003a) Plant snoRNAs: functional
evolution and new modes of gene expression. Trends Plant Sci. 8(1), 42–49.
Brown, J.W., Echeverria, M., Qu, L.H., Lowe, T.M., Bachellerie, J.P., Hut-
tenhofer, A., Kastenmayer, J.P., Green, P.J., Shaw, P. and Marshall, D.F.
(2003b) Plant snoRNA database. Nucleic Acids Res. 31(1), 432–435.
Brown, J.W., Marshall, D.F. and Echeverria, M. (2008) Intronic noncoding
RNAs and splicing. Trends Plant Sci. 13(7), 335–342. Epub 2008 Jun 2012.
Brukhin, V., Gheyselinck, J., Gagliardini, V., Genschik, P. and Grossniklaus, U.
(2005) The RPN1 subunit of the 26S proteasome in Arabidopsis is essential
for embryogenesis. Plant Cell, 17(10), 2723–2737.
Calikowski, T.T., Meulia, T. and Meier, I. (2003) A proteomic study of the
arabidopsis nuclear matrix. J. Cell. Biochem. 90(2), 361–378.
Caparros-Ruiz, D., Lahmy, S., Piersanti, S. and Echeverria, M. (1997) Two
ribosomal DNA-binding factors interact with a cluster of motifs on the 5¢external transcribed spacer, upstream from the primary pre-rRNA pro-
cessing site in a higher plant. Eur. J. Biochem. 247(3), 981–989.
Chaboute, M.E., Clement, B., Sekine, M., Philipps, G. and Chaubet-Gigot, N.
(2000) Cell cycle regulation of the tobacco ribonucleotide reductase small
subunit gene is mediated by E2F-like elements. Plant Cell, 12(10), 1987–
2000.
Chen, C.L., Liang, D., Zhou, H., Zhuo, M., Chen, Y.Q. and Qu, L.H. (2003) The
high diversity of snoRNAs in plants: identification and comparative study
of 120 snoRNA genes from Oryza sativa. Nucleic Acids Res. 31(10), 2601–
2613.
Clery, A., Senty-Segault, V., Leclerc, F., Raue, H.A. and Branlant, C. (2007)
Analysis of sequence and structural features that identify the B/C motif of
U3 small nucleolar RNA as the recognition site for the Snu13p-Rrp9p pro-
tein pair. Mol. Cell. Biol. 27(4), 1191–1206.
Comella, P., Pontvianne, F., Lahmy, S., Vignols, F., Barbezier, N., Debures, A.,
Jobet, E., Brugidou, E., Echeverria, M. and Saez-Vasquez, J. (2008) Char-
acterization of a ribonuclease III-like protein required for cleavage of
the pre-rRNA in the 3¢ETS in Arabidopsis. Nucleic Acids Res. 36(4), 1163–
1175.
Dragon, F., Gallagher, J.E., Compagnone-Post, P.A. et al. (2002) A large
nucleolar U3 ribonucleoprotein required for 18S ribosomal RNA biogene-
sis. Nature, 417(6892), 967–970.
Eichler, D.C. and Craig, N. (1994) Processing of eukaryotic ribosomal RNA.
Prog. Nucleic Acid Res. Mol. Biol. 49, 197–239.
Elias, J.E. and Gygi, S.P. (2007) Target-decoy search strategy for increased
confidence in large-scale protein identifications by mass spectrometry.
Nat. Methods, 4(3), 207–214.
Fatica, A. and Tollervey, D. (2002) Making ribosomes. Curr. Opin. Cell Biol.
14(3), 313–318.
Fromont-Racine, M., Senger, B., Saveanu, C. and Fasiolo, F. (2003) Ribosome
assembly in eukaryotes. Gene, 313(2003), 17–42.
Gallagher, J.E., Dunbar, D.A., Granneman, S., Mitchell, B.M., Osheim, Y.,
Beyer, A.L. and Baserga, S.J. (2004) RNA polI transcription and pre-rRNA
processing are linked by specific SSU processome components. Genes
Dev. 18(20), 2506–2517.
Giavalisco, P., Wilson, D., Kreitler, T., Lehrach, H., Klose, J., Gobom, J. and
Fucini, P. (2005) High heterogeneity within the ribosomal proteins of the
Arabidopsis thaliana 80S ribosome. Plant Mol. Biol. 57(4), 577–591.
Grandi, P., Rybin, V., Bassler, J. et al. (2002) 90S pre-ribosomes include the
35S pre-rRNA, the U3 snoRNP, and 40S subunit processing factors but
predominantly lack 60S synthesis factors. Mol. Cell, 10(1), 105–115.
Granneman, S., Pruijn, G.J., Horstman, W., van Venrooij, W.J., Luhrmann, R.
and Watkins, N.J. (2002) The hU3-55K protein requires 15.5K binding to the
box B/C motif as well as flanking RNA elements for its association with the
U3 small nucleolar RNA in vitro. J. Biol. Chem. 277(50), 48490–48500.
Griffith, M.E., Mayer, U., Capron, A., Ngo, Q.A., Surendrarao, A., McClinton,
R., Jurgens, G. and Sundaresan, V. (2007) The TORMOZ gene encodes a
nucleolar protein required for regulated division planes and embryo
development in Arabidopsis. Plant Cell, 19(7), 2246–2263.
Hartshorne, T. and Agabian, N. (1994) A common core structure for U3 small
nucleolar RNAs. Nucleic Acids Res. 22(16), 3354–3364.
Hughes, J.M. and Ares, M. Jr (1991) Depletion of U3 small nucleolar RNA
inhibits cleavage in the 5¢ external transcribed spacer of yeast pre-ribo-
somal RNA and impairs formation of 18S ribosomal RNA. EMBO J. 10(13),
4231–4239.
Jones, K.W., Gorzynski, K., Hales, C.M., Fischer, U., Badbanchi, F., Terns, R.M.
and Terns, M.P. (2001) Direct interaction of the spinal muscular atrophy
disease protein SMN with the small nucleolar RNA-associated protein fi-
brillarin. J. Biol. Chem. 276(42), 38645–38651.
Kass, S., Tyc, K., Steitz, J.A. and Sollner-Webb, B. (1990) The U3 small
nucleolar ribonucleoprotein functions in the first step of preribosomal RNA
processing. Cell, 60(6), 897–908.
Kiss, T. and Solymosy, F. (1990) Molecular analysis of a U3 RNA gene locus in
tomato: transcription signals, the coding region, expression in transgenic
tobacco plants and tandemly repeated pseudogenes. Nucleic Acids Res.
18(8), 1941–1949.
Kufel, J., Dichtl, B. and Tollervey, D. (1999) Yeast Rnt1p is required for
cleavage of the pre-ribosomal RNA in the 3¢ ETS but not the 5¢ ETS. RNA,
5(7), 909–917.
Lafontaine, D.L. and Tollervey, D. (1999) Nop58p is a common component of
the box C+D snoRNPs that is required for snoRNA stability. RNA, 5(3), 455–
467.
Lalev, A.I. and Nazar, R.N. (2001) A chaperone for ribosome maturation.
J. Biol. Chem. 276(20), 16655–16659.
Larkin, M.A., Blackshields, G., Brown, N.P. et al. (2007) Clustal W and Clustal
X version 2.0. Bioinformatics, 23(21), 2947–2948.
Leader, D.J., Clark, G.P., Watters, J., Beven, A.F., Shaw, P.J. and Brown, J.W.
(1997) Clusters of multiple different small nucleolar RNA genes in plants
are expressed as and processed from polycistronic pre-snoRNAs. EMBO J.
16(18), 5742–5751.
Leary, D.J., Terns, M.P. and Huang, S. (2004) Components of U3 snoRNA-
containing complexes shuttle between nuclei and the cytoplasm and dif-
ferentially localize in nucleoli: implications for assembly and function. Mol.
Biol. Cell, 15(1), 281–293.
Leung, A.K., Trinkle-Mulcahy, L., Lam, Y.W., Andersen, J.S., Mann, M. and
Lamond, A.I. (2006) NOPdb: Nucleolar Proteome Database. Nucleic Acids
Res. 34(Database issue), D218–D220.
Lukowiak, A.A., Granneman, S., Mattox, S.A., Speckmann, W.A., Jones, K.,
Pluk, H., Venrooij, W.J., Terns, R.M. and Terns, M.P. (2000) Interaction of
the U3-55k protein with U3 snoRNA is mediated by the box B/C motif of U3
and the WD repeats of U3-55k. Nucleic Acids Res. 28(18), 3462–3471.
Markham, N.R. and Zuker, M. (2005) DINAMelt web server for nucleic
acid melting prediction. Nucleic Acids Res. 33(Web Server issue), W577–
W581.
Marshallsay, C., Kiss, T. and Filipowicz, W. (1990) Amplification of plant U3
and U6 snRNA gene sequences using primers specific for an upstream
promoter element and conserved intragenic regions. Nucleic Acids Res.
18(12), 3459–3466.
Brassica oleracea U3 snoRNP 397
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2010), 61, 383–398

Mathieu, O., Yukawa, Y., Prieto, J.L., Vaillant, I., Sugiura, M. and Tourmente,
S. (2003) Identification and characterization of transcription factor IIIA and
ribosomal protein L5 from Arabidopsis thaliana. Nucleic Acids Res. 31(9),
2424–2433.
Meier, U.T. (2005) The many facets of H/ACA ribonucleoproteins. Chromo-
soma, 114(1), 1–14.
Miles, T.D., Jakovljevic, J., Horsey, E.W., Harnpicharnchai, P., Tang, L. and
Woolford, J.L. Jr (2005) Ytm1, Nop7, and Erb1 form a complex necessary
for maturation of yeast 66S preribosomes. Mol. Cell. Biol. 25(23), 10419–
10432.
Moll, C., von Lyncker, L., Zimmermann, S., Kagi, C., Baumann, N., Twell, D.,
Grossniklaus, U. and Gross-Hardt, R. (2008) CLO/GFA1 and ATO are novel
regulators of gametic cell fate in plants. Plant J. 56(6), 913–921.
Nomura, M. (2001) Ribosomal RNA genes, RNA polymerases, nucleolar
structures, and synthesis of rRNA in the yeast Saccharomyces cerevisiae.
Cold Spring Harb. Symp. Quant. Biol. 66, 555–565.
Pagnussat, G.C., Yu, H.J., Ngo, Q.A., Rajani, S., Mayalagu, S., Johnson, C.S.,
Capron, A., Xie, L.F., Ye, D. and Sundaresan, V. (2005) Genetic and
molecular identification of genes required for female gametophyte devel-
opment and function in Arabidopsis. Development 132(3), 603–614.
Pendle, A.F., Clark, G.P., Boon, R., Lewandowska, D., Lam, Y.W., Andersen, J.,
Mann, M., Lamond, A.I., Brown, J.W. and Shaw, P.J. (2005) Proteomic
analysis of the Arabidopsis nucleolus suggests novel nucleolar functions.
Mol. Biol. Cell, 16(1), 260–269.
Peng, J., Elias, J.E., Thoreen, C.C., Licklider, L.J. and Gygi, S.P. (2003) Evalu-
ation of multidimensional chromatography coupled with tandem mass
spectrometry (LC/LC-MS/MS) for large-scale protein analysis: the yeast
proteome. J. Proteome Res. 2(1), 43–50.
Pih, K.T., Yi, M.J., Liang, Y.S., Shin, B.J., Cho, M.J., Hwang, I. and Son, D.
(2000) Molecular cloning and targeting of a fibrillarin homolog from Ara-
bidopsis. Plant Physiol. 123(1), 51–58.
Pontvianne, F., Matia, I., Douet, J., Tourmente, S., Medina, F.J., Echeverria, M.
and Saez-Vasquez, J. (2007) Characterization of AtNUC-L1 reveals a central
role of nucleolin in nucleolus organization and silencing of AtNUC-L2 gene
in Arabidopsis. Mol. Biol. Cell, 18(2), 369–379.
Preuss, S.B., Costa-Nunes, P., Tucker, S. et al. (2008) Multimegabase silen-
cing in nucleolar dominance involves siRNA-directed DNA methylation
and specific methylcytosine-binding proteins. Mol. Cell, 32(5), 673–684.
Ruggero, D., Ciammaruconi, A. and Londei, P. (1998) The chaperonin of the
archaeon Sulfolobus solfataricus is an RNA-binding protein that partici-
pates in ribosomal RNA processing. EMBO J. 17(12), 3471–3477.
Saez-Vasquez, J. and Echeverria, M. (2006) Polymerase I transcription. In
Regulation of Transcription in Plants (Grasser, K.D., ed.). Oxford, UK:
Blackwell, pp. 162–183.
Saez-Vasquez, J. and Medina, F.J. (2008) The plant nucleolus. In Botanical
Research: Incorporating Advances in Plant Pathology, Vol. 47 (Kader,
J.-C. and Delseny, M., eds). San Diego: Elsevier Academic Press Inc,
pp. 1–46.
Saez-Vasquez, J., Caparros-Ruiz, D., Barneche, F. and Echeverria, M. (2004a)
Characterization of a crucifer plant pre-rRNA processing complex. Bio-
chem. Soc. Trans. 32(Pt 4), 578–580.
Saez-Vasquez, J., Caparros-Ruiz, D., Barneche, F. and Echeverria, M. (2004b)
A plant snoRNP complex containing snoRNAs, fibrillarin, and nucleolin-like
proteins is competent for both rRNA gene binding and pre-rRNA pro-
cessing in vitro. Mol. Cell. Biol. 24(16), 7284–7297.
Schafer, T., Strauss, D., Petfalski, E., Tollervey, D. and Hurt, E. (2003) The path
from nucleolar 90S to cytoplasmic 40S pre-ribosomes. EMBO J. 22(6),
1370–1380.
Schlatter, H., Langer, T., Rosmus, S., Onneken, M.L. and Fasold, H. (2002) A
novel function for the 90 kDa heat-shock protein (Hsp90): facilitating nu-
clear export of 60 S ribosomal subunits. Biochem. J. 362(Pt 3), 675–684.
Segault, V., Mougin, A., Gregoire, A., Banroques, J. and Branlant, C. (1992) An
experimental study of Saccharomyces cerevisiae U3 snRNA conformation
in solution. Nucleic Acids Res. 20(13), 3443–3451.
Shaw, P.J. and Brown, J.W. (2004) Plant nuclear bodies. Curr. Opin. Plant Biol.
7(6), 614–620.
Shaw, D.J., Eggleton, P. and Young, P.J. (2008) Joining the dots: production,
processing and targeting of U snRNP to nuclear bodies. Biochim. Biophys.
Acta, 1783(11), 2137–2144.
Speckmann, W., Narayanan, A., Terns, R. and Terns, M.P. (1999) Nuclear
retention elements of U3 small nucleolar RNA. Mol. Cell. Biol. 19(12), 8412–
8421.
Stavreva, D.A., Kawasaki, M., Dundr, M. et al. (2006) Potential roles for
ubiquitin and the proteasome during ribosome biogenesis. Mol. Cell. Biol.
26(13), 5131–5145.
Thiry, M. and Lafontaine, D.L. (2005) Birth of a nucleolus: the evolution of
nucleolar compartments. Trends Cell Biol. 15(4), 194–199.
Tschochner, H. and Hurt, E. (2003) Pre-ribosomes on the road from the
nucleolus to the cytoplasm. Trends Cell Biol. 13(5), 255–263.
Turner, A.J., Knox, A.A., Prieto, J.L., McStay, B. and Watkins, N.J. (2009) A
novel SSU processome assembly intermediate that contains the U3
snoRNP, nucleolin, RRP5 and DBP4. Mol. Cell. Biol. 30, 30.
Valasek, L., Hasek, J., Nielsen, K.H. and Hinnebusch, A.G. (2001) Dual function
of eIF3j/Hcr1p in processing 20 S pre-rRNA and translation initiation.
J. Biol. Chem. 276(46), 43351–43360.
Venema, J. and Tollervey, D. (1995) Processing of pre-ribosomal RNA in
Saccharomyces cerevisiae. Yeast, 11(16), 1629–1650.
Wassenegger, M. and Krczal, G. (2006) Nomenclature and functions of RNA-
directed RNA polymerases. Trends Plant Sci. 10, 10.
Whittaker, E., Lossky, M. and Beggs, J.D. (1990) Affinity purification of
spliceosomes reveals that the precursor RNA processing protein PRP8, a
protein in the U5 small nuclear ribonucleoprotein particle, is a component
of yeast spliceosomes. Proc. Natl Acad. Sci. USA, 87(6), 2216–2219.
Zanetti, M.E., Chang, I.F., Gong, F., Galbraith, D.W. and Bailey-Serres, J.
(2005) Immunopurification of polyribosomal complexes of Arabidopsis for
global analysis of gene expression. Plant Physiol. 138(2), 624–635.
398 Hala Samaha et al.
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2010), 61, 383–398
Related Documents


![Ovissipour] Pasteurization Conditions (Spinacia Oleracea ...](https://static.cupdf.com/doc/110x72/620925e7e2850e2aa1004127/ovissipour-pasteurization-conditions-spinacia-oleracea-.jpg)








