Identification of plasmids by PCR-based replicon typing Alessandra Carattoli a, * , Alessia Bertini a , Laura Villa a , Vincenzo Falbo b , Katie L. Hopkins c , E. John Threlfall c a Department of Infectious, Parasitic and Immune-mediated Diseases, Istituto Superiore di Sanita `, Viale Regina Elena 299, 00161 Rome, Italy b Department of Cell Biology and Neurosciences, Istituto Superiore di Sanita ` , Rome, Italy c Laboratory of Enteric Pathogens, Health Protection Agency Centre for Infections, 61 Colindale Avenue, London NW9 5HT, United Kingdom Received 2 February 2005; received in revised form 15 March 2005; accepted 17 March 2005 Available online 2 June 2005 Abstract The epidemiological importance of tracing plasmids conferring drug resistance prompted us to develop a PCR method based on replicons (inc/rep PCR) of the major plasmid incompatibility groups among Enterobacteriaceae . Eighteen pairs of primers were designed to perform 5 multiplex- and 3 simplex-PCRs, recognizing FIA, FIB, FIC, HI1, HI2, I1-Ig, L/M, N, P, W,T, A/C, K, B/O, X, Y, F, and FIIA. The specificity of the method was tested on a collection of 61 reference plasmids and on 20 Salmonella enterica strains of different serotypes isolated in Italy. Results indicated that the inc/rep PCR method demonstrates high specificity and sensitivity in detecting replicons on reference plasmids and also revealed the presence of recurrent and common plasmids in epidemiologically unrelated Salmonella isolates of different serotypes. These results suggest that the method is potentially applicable to a large number of strains to trace the diffusion of specific multi-drug resistance plasmids in different environments. D 2005 Elsevier B.V. All rights reserved. Keywords: Plasmids; PCR; Replicon 1. Introduction Plasmids are extra-chromosomal circular fragments of DNA that replicate autonomously in a host cell. They are present in nearly all bacterial species and vary in size from a few to more than several hundred kilobase pairs (kb) (Waters, 1999). Plasmids appear to increase bacterial genetic diversity, acquiring and los- ing genes, and can be horizontally exchanged among bacterial populations by conjugation or mobilization (Francia et al., 2004). They contain genes essential for initiation and control of replication and accessory genes that may be useful to their bacterial host such as antimicrobial resistance or virulence genes (Ama- bile-Cuevas and Chicurel, 1992; Bergstrom et al., 2000; Thomas, 1973). 0167-7012/$ - see front matter D 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.mimet.2005.03.018 * Corresponding author. Tel.: +39 6 4990 3128; fax: +39 6 4938 7112. E-mail address: [email protected] (A. Carattoli). Journal of Microbiological Methods 63 (2005) 219 – 228 www.elsevier.com/locate/jmicmeth

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/jmicmeth
Journal of Microbiological Methods 6
Identification of plasmids by PCR-based replicon typing
Alessandra Carattolia,*, Alessia Bertinia, Laura Villaa, Vincenzo Falbob,
Katie L. Hopkinsc, E. John Threlfallc
aDepartment of Infectious, Parasitic and Immune-mediated Diseases, Istituto Superiore di Sanita, Viale Regina Elena 299, 00161 Rome, ItalybDepartment of Cell Biology and Neurosciences, Istituto Superiore di Sanita, Rome, Italy
cLaboratory of Enteric Pathogens, Health Protection Agency Centre for Infections, 61 Colindale Avenue, London NW9 5HT, United Kingdom
Received 2 February 2005; received in revised form 15 March 2005; accepted 17 March 2005
Available online 2 June 2005
Abstract
The epidemiological importance of tracing plasmids conferring drug resistance prompted us to develop a PCR method based
on replicons (inc/rep PCR) of the major plasmid incompatibility groups among Enterobacteriaceae. Eighteen pairs of primers
were designed to perform 5 multiplex- and 3 simplex-PCRs, recognizing FIA, FIB, FIC, HI1, HI2, I1-Ig, L/M, N, P, W, T, A/C,
K, B/O, X, Y, F, and FIIA. The specificity of the method was tested on a collection of 61 reference plasmids and on 20
Salmonella enterica strains of different serotypes isolated in Italy. Results indicated that the inc/rep PCR method demonstrates
high specificity and sensitivity in detecting replicons on reference plasmids and also revealed the presence of recurrent and
common plasmids in epidemiologically unrelated Salmonella isolates of different serotypes. These results suggest that the
method is potentially applicable to a large number of strains to trace the diffusion of specific multi-drug resistance plasmids in
different environments.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Plasmids; PCR; Replicon
1. Introduction
Plasmids are extra-chromosomal circular fragments
of DNA that replicate autonomously in a host cell.
They are present in nearly all bacterial species and
vary in size from a few to more than several hundred
0167-7012/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.mimet.2005.03.018
* Corresponding author. Tel.: +39 6 4990 3128; fax: +39 6 4938
7112.
E-mail address: [email protected] (A. Carattoli).
kilobase pairs (kb) (Waters, 1999). Plasmids appear to
increase bacterial genetic diversity, acquiring and los-
ing genes, and can be horizontally exchanged among
bacterial populations by conjugation or mobilization
(Francia et al., 2004). They contain genes essential for
initiation and control of replication and accessory
genes that may be useful to their bacterial host such
as antimicrobial resistance or virulence genes (Ama-
bile-Cuevas and Chicurel, 1992; Bergstrom et al.,
2000; Thomas, 1973).
3 (2005) 219–228

A. Carattoli et al. / Journal of Microbiological Methods 63 (2005) 219–228220
A formal scheme of plasmid classification is based
on incompatibility (Inc) groups (Novick, 1987). The
procedure for incompatibility grouping is based on the
introduction, by conjugation or transformation, of a
plasmid of an bunknownQ Inc group into a strain
carrying a plasmid of a known Inc group. If the
resident plasmid is eliminated in the progeny,
the incoming plasmid is assigned to its same Inc
group (Datta and Hedges, 1971). Plasmids with the
same replication control are bincompatibleQ, whereasplasmids with different replication controls are
bcompatibleQ. On this basis two plasmids belonging
to the same Inc group cannot be propagated in the
same cell line (Datta and Hughes, 1983; Couturier et
al., 1988). Inc group identification has been frequently
used to classify plasmids. The method has been an
important tool to trace the diffusion of plasmids con-
ferring antimicrobial resistance and also to follow
the evolution and spread of emerging plasmids
(Anderson et al., 1977).
In 1988 Couturier et al. developed a new method
for the identification of the major replicons of plas-
mids circulating among the Enterobacteriaceae.This
method was based on hybridization with 19 DNA
probes that recognize different basic replicons (Cou-
turier et al., 1988). The conjugation- and hybridiza-
tion-based methodologies cannot be easily applied to
a large number of strains and their application has
been limited by the laborious and time-consuming
work required. A PCR-based detection of plasmids
was previously devised on the basis of published
sequences but it was limited to the IncP, IncN, IncW
and IncQ plasmids (Gotz et al., 1996).
The necessity of tracing plasmids conferring drug
resistance prompted us to develop an inc/rep PCR-
based typing method. In this method, 18 pairs of
primers were designed to perform 5 multiplex- and
3 simplex-PCRs, recognizing the FIA, FIB, FIC, HI1,
HI2, I1-Ig, L/M, N, P, W, T, A/C, K, B/O, X, Y, F, and
FIIA replicons, representative of the major plasmid
incompatibility groups circulating among the Entero-
bacteriaceae (Couturier et al., 1988).
The specificity of the method was tested on 61
reference plasmids. Plasmids in a collection of 20
previously characterized multi-drug resistant Salmo-
nella enterica isolates of different serotypes (Pezzella
et al., 2004) have then been typed by PCR-based inc/
rep typing.
2. Materials and methods
2.1. Bacterial strains and plasmids
Sixty-one Escherichia coli and S. enterica strains
carrying plasmids belonging to 22 different incompat-
ibility (Inc) groups, were available from the collec-
tions of the Istituto Superiore di Sanita in Rome and
the UK Health Protection Agency. Thirty-nine plas-
mids analyzed in this study were reference plasmids
of known Inc groups (Anderson et al., 1977; Carattoli
et al., 2001; Couturier et al., 1988; Frydman and
Meynell, 1969; Grindley et al., 1972; Hedges and
Datta, 1971;Villa et al., 2000); 22 plasmids were
assigned to Inc groups by conjugation with reference
plasmids (Datta and Hedges, 1971), or by hybridiza-
tion with replicon probes (Couturier et al., 1988).
Twenty S. enterica among the 58 isolates previously
characterized (Pezzella et al., 2004), isolated in Italy
from animals and food of animal origin at the Istituto
Zooprofilattico Sperimentale delle Venezie and at the
Istituto Zooprofilattico Sperimentale dell’Abruzzo e
Molise in 2000–2001, were also analysed by the PCR-
based inc/rep typing method.
2.2. PCR-based inc/rep typing method
Template DNA was prepared by one of two me-
thods. Method 1 consisted of resuspending a single
colony directly in the 25 Al of PCR reaction mixture;
this method was used during the initial testing of
primers on positive controls but it is not recommended
for the screening of replicons on uncharacterized plas-
mids, because the sensitivity of the PCR using this
template DNA is quite low. Method 2 generated total
DNA of higher purity and followed the Wizard Geno-
mic DNA purification System (Promega, Madison,
WI), starting from 2 ml of LB-broth cultures contai-
ning a suitable concentration of antibiotic for selection;
this method was used in the inc/rep typing of reference
plasmids and on the collection of Salmonella strains,
using no more than 200 ng total DNA per reaction.
Specific primer pairs were designed for eighteen
different basic replicons (Table 1).
Primers for HI1, HI2, I1, X, L/M, FIA, FIB, W, Y,
P, K and B/O replicons were designed based on
sequences obtained by DNA sequencing of the inserts
cloned in the respective pULB inc/rep constructs,
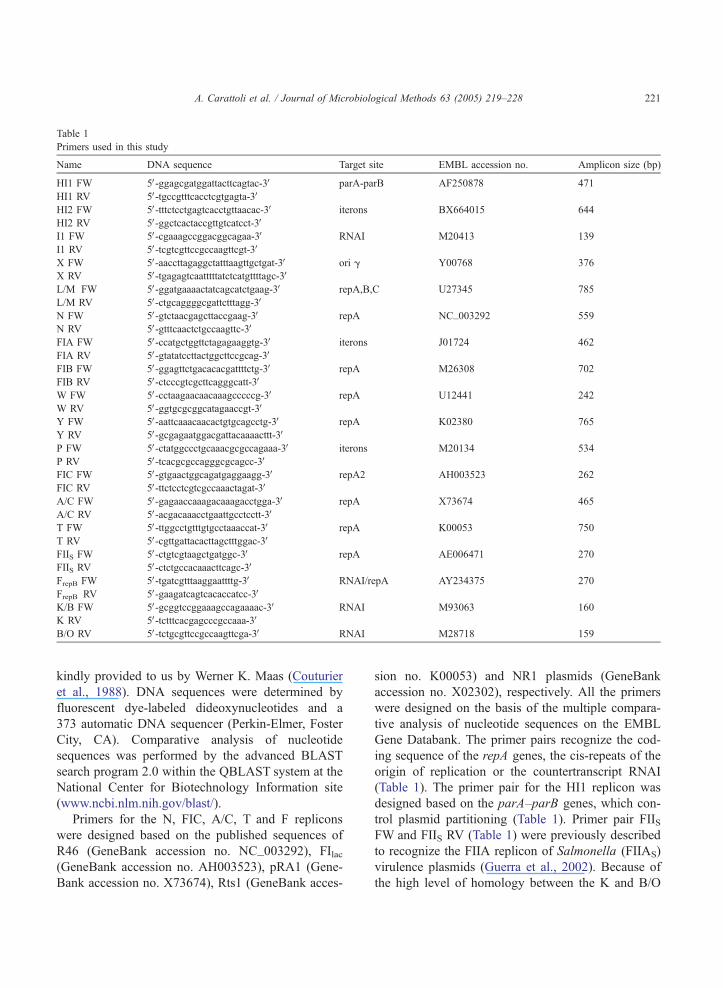
Table 1
Primers used in this study
Name DNA sequence Target site EMBL accession no. Amplicon size (bp)
HI1 FW 5V-ggagcgatggattacttcagtac-3V parA-parB AF250878 471
HI1 RV 5V-tgccgtttcacctcgtgagta-3VHI2 FW 5V-tttctcctgagtcacctgttaacac-3V iterons BX664015 644
HI2 RV 5V-ggctcactaccgttgtcatcct-3VI1 FW 5V-cgaaagccggacggcagaa-3V RNAI M20413 139
I1 RV 5V-tcgtcgttccgccaagttcgt-3VX FW 5V-aaccttagaggctatttaagttgctgat-3V ori g Y00768 376
X RV 5V-tgagagtcaatttttatctcatgttttagc-3VL/M FW 5V-ggatgaaaactatcagcatctgaag-3V repA,B,C U27345 785
L/M RV 5V-ctgcaggggcgattctttagg-3VN FW 5V-gtctaacgagcttaccgaag-3V repA NC_003292 559
N RV 5V-gtttcaactctgccaagttc-3VFIA FW 5V-ccatgctggttctagagaaggtg-3V iterons J01724 462
FIA RV 5V-gtatatccttactggcttccgcag-3VFIB FW 5V-ggagttctgacacacgattttctg-3V repA M26308 702
FIB RV 5V-ctcccgtcgcttcagggcatt-3VW FW 5V-cctaagaacaacaaagcccccg-3V repA U12441 242
W RV 5V-ggtgcgcggcatagaaccgt-3VY FW 5V-aattcaaacaacactgtgcagcctg-3V repA K02380 765
Y RV 5V-gcgagaatggacgattacaaaacttt-3VP FW 5V-ctatggccctgcaaacgcgccagaaa-3V iterons M20134 534
P RV 5V-tcacgcgccagggcgcagcc-3VFIC FW 5V-gtgaactggcagatgaggaagg-3V repA2 AH003523 262
FIC RV 5V-ttctcctcgtcgccaaactagat-3VA/C FW 5V-gagaaccaaagacaaagacctgga-3V repA X73674 465
A/C RV 5V-acgacaaacctgaattgcctcctt-3VT FW 5V-ttggcctgtttgtgcctaaaccat-3V repA K00053 750
T RV 5V-cgttgattacacttagctttggac-3VFIIS FW 5V-ctgtcgtaagctgatggc-3V repA AE006471 270
FIIS RV 5V-ctctgccacaaacttcagc-3VFrepB FW 5V-tgatcgtttaaggaattttg-3V RNAI/repA AY234375 270
FrepB RV 5V-gaagatcagtcacaccatcc-3VK/B FW 5V-gcggtccggaaagccagaaaac-3V RNAI M93063 160
K RV 5V-tctttcacgagcccgccaaa-3VB/O RV 5V-tctgcgttccgccaagttcga-3V RNAI M28718 159
A. Carattoli et al. / Journal of Microbiological Methods 63 (2005) 219–228 221
kindly provided to us by Werner K. Maas (Couturier
et al., 1988). DNA sequences were determined by
fluorescent dye-labeled dideoxynucleotides and a
373 automatic DNA sequencer (Perkin-Elmer, Foster
City, CA). Comparative analysis of nucleotide
sequences was performed by the advanced BLAST
search program 2.0 within the QBLAST system at the
National Center for Biotechnology Information site
(www.ncbi.nlm.nih.gov/blast/).
Primers for the N, FIC, A/C, T and F replicons
were designed based on the published sequences of
R46 (GeneBank accession no. NC_003292), FIlac(GeneBank accession no. AH003523), pRA1 (Gene-
Bank accession no. X73674), Rts1 (GeneBank acces-
sion no. K00053) and NR1 plasmids (GeneBank
accession no. X02302), respectively. All the primers
were designed on the basis of the multiple compara-
tive analysis of nucleotide sequences on the EMBL
Gene Databank. The primer pairs recognize the cod-
ing sequence of the repA genes, the cis-repeats of the
origin of replication or the countertranscript RNAI
(Table 1). The primer pair for the HI1 replicon was
designed based on the parA–parB genes, which con-
trol plasmid partitioning (Table 1). Primer pair FIISFW and FIIS RV (Table 1) were previously described
to recognize the FIIA replicon of Salmonella (FIIAS)
virulence plasmids (Guerra et al., 2002). Because of
the high level of homology between the K and B/O
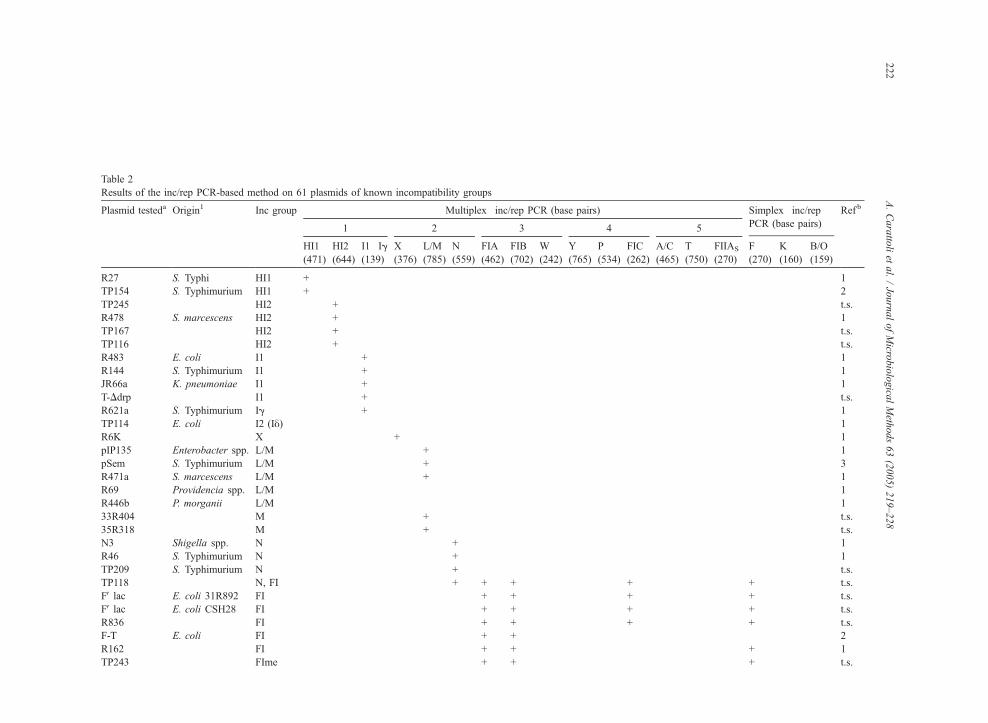
Table 2
Results of the inc/rep PCR-based method on 61 plasmids of known incompatibility groups
Plasmid testeda Origin1 Inc group Multiplex�inc/rep PCR (base pairs) Simplex�inc/rep
PCR (base pairs)
Ref b
1 2 3 4 5
HI1
(471)
HI2
(644)
I1�Ig
(139)
X
(376)
L/M
(785)
N
(559)
FIA
(462)
FIB
(702)
W
(242)
Y
(765)
P
(534)
FIC
(262)
A/C
(465)
T
(750)
FIIAS
(270)
F
(270)
K
(160)
B/O
(159)
R27 S. Typhi HI1 + � � � � � � � � � � � � � � � � � 1
TP154 S. Typhimurium HI1 + � � � � � � � � � � � � � � � � � 2
TP245 HI2 � + � � � � � � � � � � � � � � � � t.s.
R478 S. marcescens HI2 � + � � � � � � � � � � � � � � � � 1
TP167 HI2 � + � � � � � � � � � � � � � � � � t.s.
TP116 HI2 � + � � � � � � � � � � � � � � � � t.s.
R483 E. coli I1 � � + � � � � � � � � � � � � � � � 1
R144 S. Typhimurium I1 � � + � � � � � � � � � � � � � � � 1
JR66a K. pneumoniae I1 � � + � � � � � � � � � � � � � � � 1
T-Ddrp I1 � � + � � � � � � � � � � � � � � � t.s.
R621a S. Typhimurium Ig � � + � � � � � � � � � � � � � � � 1
TP114 E. coli I2 (Iy) � � � � � � � � � � � � � � � � � � 1
R6K X � � � + � � � � � � � � � � � � � � 1
pIP135 Enterobacter spp. L/M � � � � + � � � � � � � � � � � � � 1
pSem S. Typhimurium L/M � � � � + � � � � � � � � � � � � � 3
R471a S. marcescens L/M � � � � + � � � � � � � � � � � � � 1
R69 Providencia spp. L/M � � � � � � � � � � � � � � � � � � 1
R446b P. morganii L/M � � � � � � � � � � � � � � � � � � 1
33R404 M � � � � + � � � � � � � � � � � � � t.s.
35R318 M � � � � + � � � � � � � � � � � � � t.s.
N3 Shigella spp. N � � � � � + � � � � � � � � � � � � 1
R46 S. Typhimurium N � � � � � + � � � � � � � � � � � � 1
TP209 S. Typhimurium N � � � � � + � � � � � � � � � � � � t.s.
TP118 N, FI � � � � � + + + � � � + � � � + � � t.s.
FV lac E. coli 31R892 FI � � � � � � + + � � � + � � � + � � t.s.
FV lac E. coli CSH28 FI � � � � � � + + � � � + � � � + � � t.s.
R836 FI � � � � � � + + � � � + � � � + � � t.s.
F-T E. coli FI � � � � � � + + � � � � � � � � � � 2
R162 FI � � � � � � + + � � � � � � � + � � 1
TP243 FIme � � � � � � + + � � � � � � � + � � t.s.
A.Caratto
liet
al./JournalofMicro
biologica
lMeth
ods63(2005)219–228
222
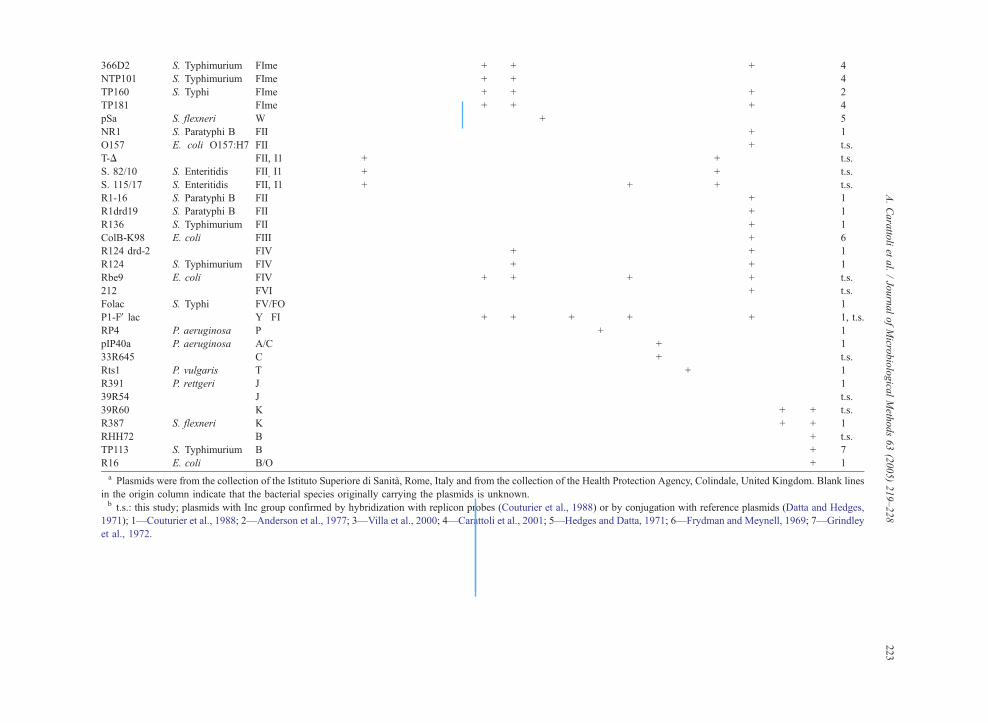
366D2 S. Typhimurium FIme � � � � � � + + � � � � � � � + � � 4
NTP101 S. Typhimurium FIme � � � � � � + + � � � � � � � � � � 4
TP160 S. Typhi FIme � � � � � � + + � � � � � � � + � � 2
TP181 FIme � � � � � � + + � � � � � � � + � � 4
pSa S. flexneri W � � � � � � � � + � � � � � � � � � 5
NR1 S. Paratyphi B FII � � � � � � � � � � � � � � � + � � 1
O157 E. coli O157:H7 FII � � � � � � � � � � � � � � � + � � t.s.
T-D FII, I1 � � + � � � � � � � � � � � + � � � t.s.
S. 82/10 S. Enteritidis FII, I1 � � + � � � � � � � � � � � + � � � t.s.
S. 115/17 S. Enteritidis FII, I1 � � + � � � � � � � � + � � + � � � t.s.
R1-16 S. Paratyphi B FII � � � � � � � � � � � � � � � + � � 1
R1drd19 S. Paratyphi B FII � � � � � � � � � � � � � � � + � � 1
R136 S. Typhimurium FII � � � � � � � � � � � � � � � + � � 1
ColB-K98 E. coli FIII � � � � � � � � � � � � � � � + � � 6
R124 drd-2 FIV � � � � � � � + � � � � � � � + � � 1
R124 S. Typhimurium FIV � � � � � � � + � � � � � � � + � � 1
Rbe9 E. coli FIV � � � � � � + + � � � + � � � + � � t.s.
212 FVI � � � � � � � � � � � � � � � + � � t.s.
Folac S. Typhi FV/FO � � � � � � � � � � � � � � � � � � 1
P1-FV lac Y�FI � � � � � � + + � + � + � � � + � � 1, t.s.
RP4 P. aeruginosa P � � � � � � � � � � + � � � � � � � 1
pIP40a P. aeruginosa A/C � � � � � � � � � � � � + � � � � � 1
33R645 C � � � � � � � � � � � � + � � � � � t.s.
Rts1 P. vulgaris T � � � � � � � � � � � � � + � � � � 1
R391 P. rettgeri J � � � � � � � � � � � � � � � � � � 1
39R54 J � � � � � � � � � � � � � � � � � � t.s.
39R60 K � � � � � � � � � � � � � � � � + + t.s.
R387 S. flexneri K � � � � � � � � � � � � � � � � + + 1
RHH72 B � � � � � � � � � � � � � � � � � + t.s.
TP113 S. Typhimurium B � � � � � � � � � � � � � � � � � + 7
R16 E. coli B/O � � � � � � � � � � � � � � � � � + 1
a Plasmids were from the collection of the Istituto Superiore di Sanita, Rome, Italy and from the collection of the Health Protection gency, Colindale, United Kingdom. Blank lines
in the origin column indicate that the bacterial species originally carrying the plasmids is unknown.b t.s.: this study; plasmids with Inc group confirmed by hybridization with replicon probes (Couturier et al., 1988) or by conjuga n with reference plasmids (Datta and Hedges,
1971); 1—Couturier et al., 1988; 2—Anderson et al., 1977; 3—Villa et al., 2000; 4—Carattoli et al., 2001; 5—Hedges and Datta, 197 ; 6—Frydman and Meynell, 1969; 7—Grindley
et al., 1972.
A.Caratto
liet
al./JournalofMicro
biologica
lMeth
ods63(2005)219–228
223
A
tio
1

A. Carattoli et al. / Journal of Microbiological Methods 63 (2005) 219–228224
replicons the same forward primer was used in both
these simplex PCRs (Table 1).
The PCR-based inc/rep typing method consists of
five different multiplex-PCRs recognizing three dif-
ferent replicon types, and three simplex-PCRs for F, K
and B/O (Table 2). All PCR amplifications, except the
F-simplex, were performed with the following ampli-
fication scheme: 1 cycle of denaturation at 94 8C for 5
min, followed by 30 cycles of denaturation at 94 8Cfor 1 min, annealing at 60 8C for 30 s and elongation
at 72 8C for 1 min. The amplification was concluded
with an extension program of 1 cycle at 72 8C for 5
min. The F-simplex PCR was performed with the
same amplification program but at an annealing tem-
perature of 52 8C.
2.3. Positive controls
Plasmids pULB-2436, -2433, -2428, -2405, -2423,
-2154, -2404, -2426, -2410, -2420, -2439, and -2406
from the inc/rep plasmid bank were initially used as
positive controls for the respective replicons (Coutur-
ier et al., 1988). The R46, FlacV from E. coli CSH28,
pIP40a, Rts1, S. 82/10 and NR1 plasmids were used
as positive controls to test the N, FIC, A/C, T, FIIAS
and F primer pairs, respectively. All the amplicons
obtained were cloned into a TA-cloning vector (Invi-
trogen-Life Technologies, Milano, Italy) and
sequenced. The cloned amplicons were then used as
positive controls for the multiplex and simplex PCRs.
3. Results and discussion
3.1. Specificity of the PCR-based inc/rep typing
method on reference plasmids
Eighteen specific primer pairs were designed, on
the basis of the multiple comparative analysis of
nucleotide sequence on the EMBL Gene Databank,
for HI1, HI2, I1-Ig, X, L/M, N, FIA, FIB, W, Y, P,
FIC, A/C, T, FIIAS, F, K and B/O replicons. The
specificity of each primer pair, listed in Table 1, was
initially performed on bacterial genomic DNA
extracted from the plasmid-free E. coli DH5a strain
(Invitrogen-Life Technologies). None of the primer
pairs gave positive amplification results on this tem-
plate. For each PCR amplification positive controls
were cloned and sequenced as described in Materials
and methods.
Fifty-eight plasmids belonging to 20 Inc groups
(Table 2), whose replicons are recognized by the set
of primers listed in Table 1, were chosen among our
plasmid collections. Two IncJ plasmids and one IncI2
(Iy) plasmid were also included as negative controls;
since there were no DNA sequence information avail-
able for these replicons they were not included in the
method.
The specificity of the PCR-based inc/rep typing
was then tested on 61 reference plasmids (58 positives
and 3 negatives) originally isolated from 16 different
bacterial species (Table 2).
3.1.1. Multiplex 1
This reaction was designed to recognize HI1-, HI2-,
I1/Ig-carrying plasmids, producing 471 bp, 644 bp
and 139 bp amplicons, respectively. Results obtained
on the 61 plasmids revealed that all the IncHI1,
IncHI2 and IncI1/Ig (also called Ia) plasmids tested
were correctly recognized by Multiplex 1. No false
positive results were observed on the collection of
plasmids. Several IncHI2 plasmids produced the
expected amplicon plus higher molecular mass ampli-
cons (data not shown). These extra bands may be
explained by the presence of multiple iterons located
upstream of the HI2 replicon that can be recognized
by the HI2 RV primer (Gilmour et al., 2004). This
reaction is very specific and positive PCR results,
even in presence of multiple amplicons, should be
interpreted as positive for the presence of the HI2
replicon.
The IncI2 (Iy) plasmid included in the collection
was negative in all the PCR amplifications. This was
as expected because this plasmid is classified into a
different subgroup in respect to the IncI1/Ig plasmids
and also exhibits a different pilus serotype (Couturier
et al., 1988).
3.1.2. Multiplex 2
This reaction was designed to recognize the X, L/
M and N replicons by amplification of 376 bp, 785 bp
and 559 bp amplicons, respectively. When used on the
61 reference plasmids, this reaction correctly detected
all the IncX and IncN plasmids and gave no false
positive results with other replicons. Multiplex 2
failed in detecting two of the seven plasmids belong-

A. Carattoli et al. / Journal of Microbiological Methods 63 (2005) 219–228 225
ing to the IncL/M group (R69 and R446b in Table 2).
R69 and R446b were positively hybridized by South-
ern blot hybridization using the L/M cloned amplicon
as a probe (data not shown). These results suggest that
the L/M replicons may diverge in sequence and occa-
sionally give false negative results by PCR. Since
only a few DNA sequences are available for these
replicons, the higher sensitivity hybridization-based
method can be applied to recognize them.
3.1.3. Multiplex 3
This reaction recognizes the FIA, FIB and W
replicons, producing 462 bp, 702 bp and 242 bp
amplicons, respectively. This amplification success-
fully recognized all the IncFI plasmids, including
IncFIme plasmids (Anderson et al., 1977), as well as
the IncW plasmid pSa (Okumura and Kado, 1992),
producing the respective amplicon (Table 2). The
IncFIV R124drd-2, R124 and Rbe9 plasmids were
positive for the FIB replicon (Table 2), and Rbe9
plasmid was also positive for FIA and FIC. These
results reproduce previously described replicon typing
results obtained by inc/rep hybridization. (Couturier et
al., 1988; Campbell and Mee, 1989).
3.1.4. Multiplex 4
This reaction was designed to recognize IncY and
IncP plasmids and also included a primer pair spe-
cific for the FIC replicon (262 bp) of the E. coli FVlac fertility plasmid. The IncY P1 and the IncP RP4
(Smith and Thomas, 1987) plasmids were positive
for Y (765 bp) and P (534 bp) replicons, respec-
tively. The FIC PCR was designed to recognize the
repA2 gene located on the E. coli FV lac plasmid and
does not recognize the FIC of the IncFIme plasmids.
This is recognized by the simplex F-PCR as dis-
cussed below.
The IncY P1 plasmid was also unexpectedly posi-
tive for FIA, FIB and FIC (Table 2). Further charac-
terization of this plasmid revealed the presence of a
multireplicon fusion of the plasmid P1 with the FV lacplasmid (data not shown), confirming the specificity
of the inc/rep PCR-based method in detecting both FI
and Y replicons.
3.1.5. Multiplex 5
This reaction was designed to recognize the IncA/
C, IncT plasmids and the repFIIA replicon of the
virulence plasmids of S. enterica serovars Enteriditis,
Dublin, Cholerae-suis and Typhimurium (Guerra et
al., 2002; Llanes et al., 1996), producing 465 bp,
750 bp and 270 bp PCR products, respectively. The
reference plasmids of the A/C and T groups were
correctly identified by the PCR-based method. Three
S. enterica serovar Enteritidis strains (82/10, 115/17
and T-D) carrying both the virulence and the IncI1
plasmids, were positively recognized by the FIIs and
I1/Ig reactions as expected. This experiment demon-
strated that multiple plasmids simultaneously present
in the same cell can be easily detected by this PCR-
based method.
3.2. The three simplex-PCRs for the F, K and B/O
replicons
Particular attention has been paid to the design of
primer pairs recognizing the I1, K and B/O repli-
cons. The I1, K and B/O replicons were not discri-
minated by the hybridization-based inc/rep typing,
sharing a DNA sequence homology higher than
90% (Couturier et al., 1988). The incompatibility
phenotype of this class of plasmids is an expression
of the evolutionary changes in the DNA sequence of
the RNA inhibitor (RNAI). Few base substitutions
within the major stem-and-loop structures predicted
for analogous RNAI of different plasmids lead to
compatible phenotypes. The specificity of our assay
relies on the specific I1/Ig PCR in Multiplex 1 and
in two simplex PCRs designed to detect the K and
B/O replicons. In the conditions used, the B/O-sim-
plex specifically detects the B/O replicon while the
K-simplex recognizes both the K and B/O (Table 2),
allowing the identification and discrimination of both
replicons.
The F amplification reaction was designed to
recognize, by a single PCR, all the plasmids belong-
ing to the heterogeneous group of the IncF-plasmids.
This reaction was based on the highly conserved
region of homology showed by the FIC and FII
replicons (Saadi et al., 1987). This PCR amplifica-
tion recognized 20 of the 26 FI, FII, FIII, FIV, FV
and FVI plasmids tested, including the FII replicon
of the E. coli O157 virulence plasmid. Among the
six F-plasmids not recognized by the F-simplex,
three were Salmonella virulence plasmids, whose
FII replicon was specifically recognized in the Multi-
A. Carattoli et al. / Journal of Microbiological Methods 63 (2005) 219–228226
plex 5 reaction and three were IncFIme plasmids that
probably lack the FIC replicon but were positive for
the FIA and FIB replicons by Multiplex 3 (Couturier
et al., 1988). The Folac plasmid assigned to the
IncFV/FO group was also negative for the F-sim-
plex. This plasmid was previously reported to give
negative results by hybridization with the inc/rep
probes (Couturier et al., 1988), and its replication
controls have never been analyzed at a DNA sequence
level.
In conclusion, the PCR-based method demon-
strates high specificity and sensitivity in detecting
replicons on reference plasmids. This method is
more specific than the hybridization-based method.
This was particularly evident when applied to FII,
FIC, I1-Ig, K and B/O replicons that exhibit consider-
able cross-reactions by hybridization-based inc/rep
typing, thus limiting the use of this method in the
classification of plasmids (Couturier et al., 1988).
Although these eighteen replicons are representative
of the major plasmid incompatibility groups among
the family Enterobacteriaceae, several other replicons
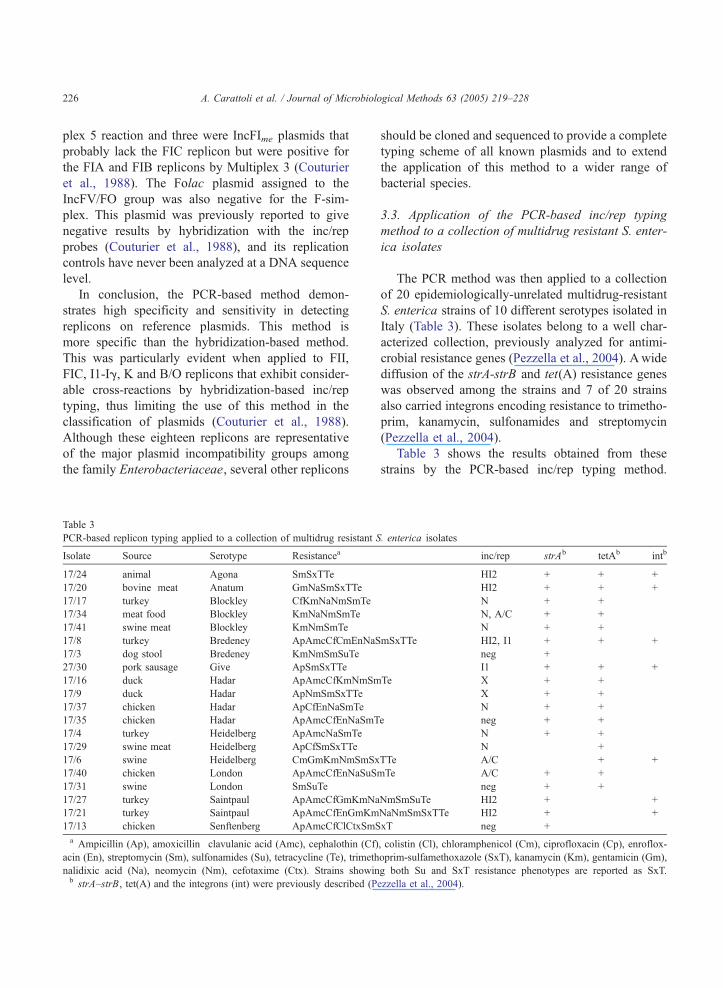
Table 3
PCR-based replicon typing applied to a collection of multidrug resistant S
Isolate Source Serotype Resistancea
17/24 animal Agona SmSxTTe
17/20 bovine meat Anatum GmNaSmSxTTe
17/17 turkey Blockley CfKmNaNmSmTe
17/34 meat food Blockley KmNaNmSmTe
17/41 swine meat Blockley KmNmSmTe
17/8 turkey Bredeney ApAmcCfCmEnNaS
17/3 dog stool Bredeney KmNmSmSuTe
27/30 pork sausage Give ApSmSxTTe
17/16 duck Hadar ApAmcCfKmNmSm
17/9 duck Hadar ApNmSmSxTTe
17/37 chicken Hadar ApCfEnNaSmTe
17/35 chicken Hadar ApAmcCfEnNaSmT
17/4 turkey Heidelberg ApAmcNaSmTe
17/29 swine meat Heidelberg ApCfSmSxTTe
17/6 swine Heidelberg CmGmKmNmSmSx
17/40 chicken London ApAmcCfEnNaSuS
17/31 swine London SmSuTe
17/27 turkey Saintpaul ApAmcCfGmKmNa
17/21 turkey Saintpaul ApAmcCfEnGmKm
17/13 chicken Senftenberg ApAmcCfClCtxSmS
a Ampicillin (Ap), amoxicillin�clavulanic acid (Amc), cephalothin (Cf)
acin (En), streptomycin (Sm), sulfonamides (Su), tetracycline (Te), trimeth
nalidixic acid (Na), neomycin (Nm), cefotaxime (Ctx). Strains showinb strA–strB, tet(A) and the integrons (int) were previously described (P
should be cloned and sequenced to provide a complete
typing scheme of all known plasmids and to extend
the application of this method to a wider range of
bacterial species.
3.3. Application of the PCR-based inc/rep typing
method to a collection of multidrug resistant S. enter-
ica isolates
The PCR method was then applied to a collection
of 20 epidemiologically-unrelated multidrug-resistant
S. enterica strains of 10 different serotypes isolated in
Italy (Table 3). These isolates belong to a well char-
acterized collection, previously analyzed for antimi-
crobial resistance genes (Pezzella et al., 2004). Awide
diffusion of the strA-strB and tet(A) resistance genes
was observed among the strains and 7 of 20 strains
also carried integrons encoding resistance to trimetho-
prim, kanamycin, sulfonamides and streptomycin
(Pezzella et al., 2004).
Table 3 shows the results obtained from these
strains by the PCR-based inc/rep typing method.
. enterica isolates
inc/rep strAb tetAb intb
HI2 + + +
HI2 + + +
N + + �N, A/C + + �N + + �
mSxTTe HI2, I1 + + +
neg + � �I1 + + +
Te X + + �X + + �N + + �
e neg + + �N + + �N � + �
TTe A/C � + +
mTe A/C + + �neg + + �
NmSmSuTe HI2 + � +
NaNmSmSxTTe HI2 + � +
xT neg + � �, colistin (Cl), chloramphenicol (Cm), ciprofloxacin (Cp), enroflox-
oprim-sulfamethoxazole (SxT), kanamycin (Km), gentamicin (Gm),
g both Su and SxT resistance phenotypes are reported as SxT.
ezzella et al., 2004).

A. Carattoli et al. / Journal of Microbiological Methods 63 (2005) 219–228 227
The inc/rep typing detected the presence of N, I1,
HI2, A/C and X replicons in several isolates of this
collection. Four strains were negative for all the
replicons tested. The N and the HI2 replicons were
detected in six and four strains, respectively, belong-
ing to different serotypes (Agona, Anatum, Blockley,
Bredeney, Hadar, Heidelberg and Saintpaul). Three
strains were positive for the A/C replicon and the X
replicon was identified in two Hadar isolates. Inte-
grons were associated with I1-, HI2-and A/C-positive
strains, but not with the N-positive strains (Table 3).
These results suggest the presence of recurrent and
common plasmids in epidemiological unrelated Sal-
monella isolates of different serotypes isolated from
distant geographical areas in Italy, suggesting the
successful spread of these genetic determinants in
zoonotic pathogens.
This analysis demonstrates that the PCR-based
method can be applied directly on collections of
strains, suggesting the potential use of the PCR-
based replicon typing method in tracking the spread
of plasmids conferring drug resistance. The method
could be applied to monitor the circulation of plas-
mids within strains from different environments or to
follow the horizontal transmission of antimicrobial
resistance genes among the Enterobacteriaceae.
Acknowledgments
We are grateful to V. Miriagou, L.S. Tzouvelekis,
E. Tzelepi and G.M. Rossolini for helpful discussion
and continuous encouragement. We thank Fabio Ric-
cobono for the DNA sequencing. This work was
supported by grants from the Istituto Superiore di
Sanita (Art. 502, project no. 2012/RI; Art. 524, project
no. C3MD).
References
Amabile-Cuevas, C.F., Chicurel, M.E., 1992. Bacterial plasmids
and gene flux. Cell 70, 189–199.
Anderson, E.S., Threlfall, E.J., Carr, J.M., Mc Connell, M.M.,
Smith, H.R., 1977. Clonal distribution of resistance plasmid-
carrying Salmonella typhimurium, mainly in the Middle East. J.
Hyg. (Lond) 79, 425–448.
Bergstrom, C.T., Lipsitch, M., Levin, B.R., 2000. Natural selection,
infectious transfer and the existence conditions for bacterial
plasmids. Genetics 155, 1505–1519.
Campbell, I.G., Mee, B.J., 1989. Distribution of plasmid mainte-
nance regions among IncFIV plasmids. FEMS Microbiol. Lett.
48, 45–49.
Carattoli, A., Villa, L., Pezzella, C., Bordi, E., Visca, P., 2001.
Expanding drug resistance through integron acquisition by
IncFI plasmids of salmonella enterica typhimurium. Emerg.
Infect. Dis. 7, 444–447.
Couturier, M., Bex, F., Bergquist, P.L., Maas, W.K., 1988. Identi-
fication and classification of bacterial plasmids. Microbiol. Rev.
52, 375–395.
Datta, N., Hedges, R.W., 1971. Compatibility groups among fi-R
factors. Nature (London) 234, 222–223.
Datta, N., Hughes, V.M., 1983. Plasmids of the same Inc groups in
Enterobacteria before and after the medical use of antibiotics.
Nature 306, 616–617.
Frydman, A., Meynell, E., 1969. Interactions between de-
repressed F-like R factors and wild type colicin B factors:
superinfection immunity and repressor susceptibility. Genet.
Res. 14, 315–322.
Francia, M.V., Varsaki, A., Garcillan-Barcia, M.P., Latorre, A.,
Drainas, C., de la Cruz, F.A., 2004. Classification scheme for
mobilization regions of bacterial plasmids. FEMS Microbiol.
Rev. 28, 79–100.
Gilmour, M.W., Thomson, N.R., Sanders, M., Parkhill, J., Taylor,
D.E., 2004. The complete nucleotide sequence of the resistance
plasmid R478: defining the backbone components of incompat-
ibility group H conjugative plasmids through comparative geno-
mics. Plasmid 52, 182–202.
Gotz, A., Pukall, R., Smit, E., Tietze, E., Prager, R., Tschape, H.,
van Elsas, J.D., Smalla, K., 1996. Detection and characterization
of broad-host-range plasmids in environmental bacteria by PCR.
Appl. Environ. Microbiol. 62, 2621–2628.
Grindley, N.D., Grindley, J.N., Anderson, E.S., 1972. R factor
compatibility groups. Mol. Gen. Genet. 119, 287–297.
Guerra, B., Soto, S., Helmuth, R., Mendoza, M.C., 2002. Character-
ization of a self-transferable plasmid from Salmonella enterica
serotype Typhimurium clinical isolates carrying two integron-
borne gene cassettes together with virulence and drug resistance
genes. Antimicrob. Agents Chemother. 46, 2977–2981.
Hedges, R.W., Datta, N., 1971. fi- R-factors giving chloramphenicol
resistance. Nature New Biol. 234, 220–221.
Llanes, C., Gabant, P., Couturier, M., Bayer, L., Plesiat, P., 1996.
Molecular analysis of the replication elements of the broad-host-
range RepA/C replicon. Plasmid 36, 26–35.
Novick, R.P., 1987. Plasmid incompatibility. Microbiol. Rev. 51,
381–395.
Okumura, M.S., Kado, C.I., 1992. The region essential for efficient
autonomous replication of pSa in Escherichia coli. Mol. Gen.
Genet. 235, 55–63.
Pezzella, C., Ricci, A., Di Giannatale, E., Luzzi, I., Carattoli, A.,
2004. Tetracycline and streptomycin resistance genes, transpo-
sons, and plasmids in Salmonella enterica isolates from animals
in Italy. Antimicrob. Agents Chemother. 48, 903–908.
Saadi, S., Maas, W.K., Hill, D.F., Bergquist, P.L., 1987. Nucleotide
sequence analysis of RepFIC, a basic replicon present in IncFI
plasmids P307 and F, and its relation to the RepA replicon of
IncFII plasmids. J. Bacteriol. 169, 1836–1846.

A. Carattoli et al. / Journal of Microbiological Methods 63 (2005) 219–228228
Smith, C.A., Thomas, C.M., 1987. Comparison of the organisation
of the genomes of phenotypically diverse plasmids of incompat-
ibility group P: members of the IncP beta sub-group are closely
related. Mol. Gen. Genet. 206, 419–427.
Thomas, C.M., 1973. Paradigms of plasmid organization. Mol.
Microbiol. 74, 335–337.
Villa, L., Pezzella, C., Tosini, F., Visca, P., Petrucca, A., Carattoli,
A., 2000. Multiple-antibiotic resistance mediated by structu-
rally-related IncL/M plasmids carrying an extended-spectrum
h-lactamase gene and a class 1 integron. Antimicrob. Agents
Chemother. 44, 2911–2914.
Waters, V.L., 1999. Conjugative transfer in the dissemination of
beta-lactam and aminoglycoside resistance. Front. Biosci. 4,
416–439.
Related Documents










