Steroids 69 (2004) 1–16 Identification of naturally occurring spirostenols preventing -amyloid-induced neurotoxicity Laurent Lecanu a,b , Wenguo Yao a,b , Gary L. Teper a,b , Zhi-Xing Yao a,b , Janet Greeson c , Vassilios Papadopoulos a,b,d,e,∗ a Department of Cell Biology, Division of Hormone Research, Georgetown University School of Medicine, Washington, DC 20057, USA b Samaritan Research Laboratories, Georgetown University School of Medicine, Washington, DC 20057, USA c Samaritan Pharmaceuticals, Las Vegas, NV 89109, USA d Department of Pharmacology, Georgetown University School of Medicine, Washington, DC 20057, USA e Department of Neurosciences, Georgetown University School of Medicine, Washington, DC 20057, USA Received 19 December 2002; received in revised form 3 July 2003; accepted 4 September 2003 Abstract 22R-Hydroxycholesterol is an intermediate in the steroid biosynthesis pathway shown to exhibit a neuroprotective property against -amyloid (1–42) (A) toxicity in rat PCl2 and human NT2N neuronal cells by binding and inactivating A. In search of potent 22R-hydroxycholesterol derivatives, we assessed the ability of a series of naturally occurring entities containing the 22R-hydroxycholesterol structure to protect PC12 cells against A-induced neurotoxicity, determined by measuring changes in membrane potential, mitochon- drial diaphorase activity, ATP levels and trypan blue uptake. 22R-Hydroxycholesterol derivatives sharing a common spirost-5-en-3-ol or a furost-5-en-3-ol structure were tested. Although some of these compounds were neuroprotective against 0.1 MA, only three pro- tected against the 1–10 MA-induced toxicity and, in contrast to 22R-hydroxycholesterol, all were devoid of steroidogenic activity. These entities shared a common structural feature, a long chain ester in position 3 and common stereochemistry. The neuroprotective property of these compounds was coupled to their ability to displace radiolabeled 22R-hydroxycholesterol from A, suggesting that the A-22R-hydroxycholesterol physicochemical interaction contributes to their beneficial effect. In addition, a 22R-hydroxycholesterol deriva- tive inhibited the formation of neurotoxic amyloid-derived diffusible ligands. Computational docking simulations of 22R-hydroxycholesterol and its derivatives on A identified two binding sites. Chemical entities, as 22R-hydroxycholesterol, seem to bind preferentially only to one site. In contrast, the presence of the ester chain seems to confer the ability to bind to both sites on A, leading to neuroprotection against high concentrations of A. In conclusion, these results suggest that spirost-5-en-3-ol naturally occurring derivatives of 22R-hydroxycholesterol might offer a new approach for Alzheimer’s disease therapy. © 2003 Elsevier Inc. All rights reserved. Keywords: Neurodegeneration; Neuroprotection; Alzheimer’s disease; Steroids; Cholesterol; Spirostenol; Amyloid-derived diffusible ligand (ADDL) 1. Introduction Excessive accumulation in the brain of the -amyloid peptide (A), due either to overproduction and/or decreased clearance and the formation of senile plaques, and neurofib- rillary tangles due to hyperphosphorylation of the Tau pro- tein, are the hallmarks of Alzheimer disease (AD) pathology [1]. These biochemical modifications have been shown to be associated with a slowly evolving cognitive deficit and memory impairment [2–4]. Recent advances in the understanding of the neuropathol- ogy of the AD have permitted the identification of potential ∗ Corresponding author. Tel.: +1-202-687-8991; fax: +1-202-687-7855. E-mail address: [email protected] (V. Papadopoulos). targets and the development of new therapeutic approaches. As circulating sexual steroid concentrations have been shown to decrease over time in life and to be lower in aging and Alzheimer’s disease patients [5], one such promising strategy is the utilization of steroids. Among the various steroids examined in vitro for their putative neuroprotective properties against A, estrogens showed the most promis- ing results [6–10]. In addition, ovariectomy was shown to impair spatial memory while estradiol replacement therapy allowed the recovery of memory functions in rhesus mon- keys [11]. In agreement with these findings, disruption of the estrogen receptor beta gene induced learning disabilities in mice [12]. In addition to the classical estrogens, phytoestro- gens, plant-derived compounds displaying estrogenic activ- ity, were also shown to exert protective activity against A 0039-128X/$ – see front matter © 2003 Elsevier Inc. All rights reserved. doi:10.1016/j.steroids.2003.09.007

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Steroids 69 (2004) 1–16
Identification of naturally occurring spirostenols preventing�-amyloid-induced neurotoxicity
Laurent Lecanua,b, Wenguo Yaoa,b, Gary L. Tepera,b, Zhi-Xing Yaoa,b,Janet Greesonc, Vassilios Papadopoulosa,b,d,e,∗
a Department of Cell Biology, Division of Hormone Research, Georgetown University School of Medicine, Washington, DC 20057, USAb Samaritan Research Laboratories, Georgetown University School of Medicine, Washington, DC 20057, USA
c Samaritan Pharmaceuticals, Las Vegas, NV 89109, USAd Department of Pharmacology, Georgetown University School of Medicine, Washington, DC 20057, USAe Department of Neurosciences, Georgetown University School of Medicine, Washington, DC 20057, USA
Received 19 December 2002; received in revised form 3 July 2003; accepted 4 September 2003
Abstract
22R-Hydroxycholesterol is an intermediate in the steroid biosynthesis pathway shown to exhibit a neuroprotective property against�-amyloid (1–42) (A�) toxicity in rat PCl2 and human NT2N neuronal cells by binding and inactivating A�. In search of potent22R-hydroxycholesterol derivatives, we assessed the ability of a series of naturally occurring entities containing the 22R-hydroxycholesterolstructure to protect PC12 cells against A�-induced neurotoxicity, determined by measuring changes in membrane potential, mitochon-drial diaphorase activity, ATP levels and trypan blue uptake. 22R-Hydroxycholesterol derivatives sharing a common spirost-5-en-3-ol ora furost-5-en-3-ol structure were tested. Although some of these compounds were neuroprotective against 0.1�M A�, only three pro-tected against the 1–10�M A�-induced toxicity and, in contrast to 22R-hydroxycholesterol, all were devoid of steroidogenic activity.These entities shared a common structural feature, a long chain ester in position 3 and common stereochemistry. The neuroprotectiveproperty of these compounds was coupled to their ability to displace radiolabeled22R-hydroxycholesterol from A�, suggesting that theA�-22R-hydroxycholesterol physicochemical interaction contributes to their beneficial effect. In addition, a 22R-hydroxycholesterol deriva-tive inhibited the formation of neurotoxic amyloid-derived diffusible ligands. Computational docking simulations of 22R-hydroxycholesteroland its derivatives on A� identified two binding sites. Chemical entities, as 22R-hydroxycholesterol, seem to bind preferentially only to onesite. In contrast, the presence of the ester chain seems to confer the ability to bind to both sites on A�, leading to neuroprotection against highconcentrations of A�. In conclusion, these results suggest that spirost-5-en-3-ol naturally occurring derivatives of 22R-hydroxycholesterolmight offer a new approach for Alzheimer’s disease therapy.© 2003 Elsevier Inc. All rights reserved.
Keywords: Neurodegeneration; Neuroprotection; Alzheimer’s disease; Steroids; Cholesterol; Spirostenol; Amyloid-derived diffusible ligand (ADDL)
1. Introduction
Excessive accumulation in the brain of the�-amyloidpeptide (A�), due either to overproduction and/or decreasedclearance and the formation of senile plaques, and neurofib-rillary tangles due to hyperphosphorylation of the Tau pro-tein, are the hallmarks of Alzheimer disease (AD) pathology[1]. These biochemical modifications have been shown tobe associated with a slowly evolving cognitive deficit andmemory impairment[2–4].
Recent advances in the understanding of the neuropathol-ogy of the AD have permitted the identification of potential
∗ Corresponding author. Tel.:+1-202-687-8991; fax:+1-202-687-7855.E-mail address: [email protected] (V. Papadopoulos).
targets and the development of new therapeutic approaches.As circulating sexual steroid concentrations have beenshown to decrease over time in life and to be lower in agingand Alzheimer’s disease patients[5], one such promisingstrategy is the utilization of steroids. Among the varioussteroids examined in vitro for their putative neuroprotectiveproperties against A�, estrogens showed the most promis-ing results[6–10]. In addition, ovariectomy was shown toimpair spatial memory while estradiol replacement therapyallowed the recovery of memory functions in rhesus mon-keys[11]. In agreement with these findings, disruption of theestrogen receptor beta gene induced learning disabilities inmice[12]. In addition to the classical estrogens, phytoestro-gens, plant-derived compounds displaying estrogenic activ-ity, were also shown to exert protective activity against A�
0039-128X/$ – see front matter © 2003 Elsevier Inc. All rights reserved.doi:10.1016/j.steroids.2003.09.007

2 L. Lecanu et al. / Steroids 69 (2004) 1–16
toxicity in neurons in vitro[13] and improve memory in ratsin vivo [14,15]. Moreover, the estrogen precursors DHEAand DHEAS were shown to improve memory in young andageing mice[16,17]. In humans, endogenous estradiol andtestosterone levels have been shown to correlate to cognitiveperformances in older women[18] and high dose estradioltherapy was shown to improve cognition in AD women[19,20]. Despite these findings, the use of estrogen as atherapeutic means in AD has been controversial because ofreports of absence of clear long-term effects[21–26] andthe potential side effects of long-term estrogen use[27,28].
We recently reported that the steroid 22R-hydroxychole-sterol, an intermediate in the cytochrome P450 C27 sidechain cleavage enzymatic pathway of pregnenolone for-mation from cholesterol[29], is present at lower levels inAD hippocampus and frontal cortex tissue specimens com-pared to age-matched controls[30]. 22R-hydroxycholesterolwas then found to protect, in a dose-dependent manner,against A�-induced rat sympathetic nerve pheochromocy-toma (PC12) and differentiated human NT2N neuronal celldeath. The effect of 22R-hydroxycholesterol was stereospe-cific because its enantiomer 22S-hydroxycholesterol failedto protect the neurons from A�-induced cell death. More-over the protective effect of this steroid was specific forA�-induced cell death because it did not protect againstglutamate-induced neurotoxicity. In search of the mecha-nism of action of 22R-hydroxycholesterol, we found that itbinds to A� and that the formed 22R-hydroxycholesterol/A�complex is not toxic to rodent and human neurons[30].Because 22R-hydroxycholesterol is a precursor of preg-nenolone, and thus of the final steroid products made, itwould be rapidly metabolized in steroid forming tissues bythe cytochrome P450 C27 side chain cleavage (P450scc);thus, we used 22R-hydroxycholesterol as the lead structureand searched for naturally occurring compounds contain-ing this structure and exhibiting neuroprotective propertiesagainst A� toxicity. We report herein the identificationof biologically active 22R-hydroxycholesterol derivativescontaining a common spirost-5-en-3-ol structure that mightoffer a new approach for AD therapy.
2. Experimental
2.1. Materials
A�1–42 peptide was purchased from American PeptideCo. (Sunnyvale, CA) and stored at−80◦C. Just before use,A�1–42 was reconstituted with fresh distilled water to reacha concentration of 500�M and then added directly to thecells at the desired concentration. 22R-Hydroxycholesterol(SP222) was purchased from Sigma (St. Louis, MO).[22-3H]R-hydroxycholesterol (specific activity 20 Ci/mmol)was synthesized by American Radiolabeled Chemical(St. Louis, MO). The 22R-hydroxycholesterol derivatives(SP223–238) were purchased from Interbioscreen (Moscow,
Russia). Cells culture supplies were purchased form GIBCO(Grand Island, NY) and cell culture plasticware was fromCorning (Corning, NY) and Packard BioSciences Co.(Meriden, CT).
2.2. In silico screening for 22R-hydroxycholesterolderivatives
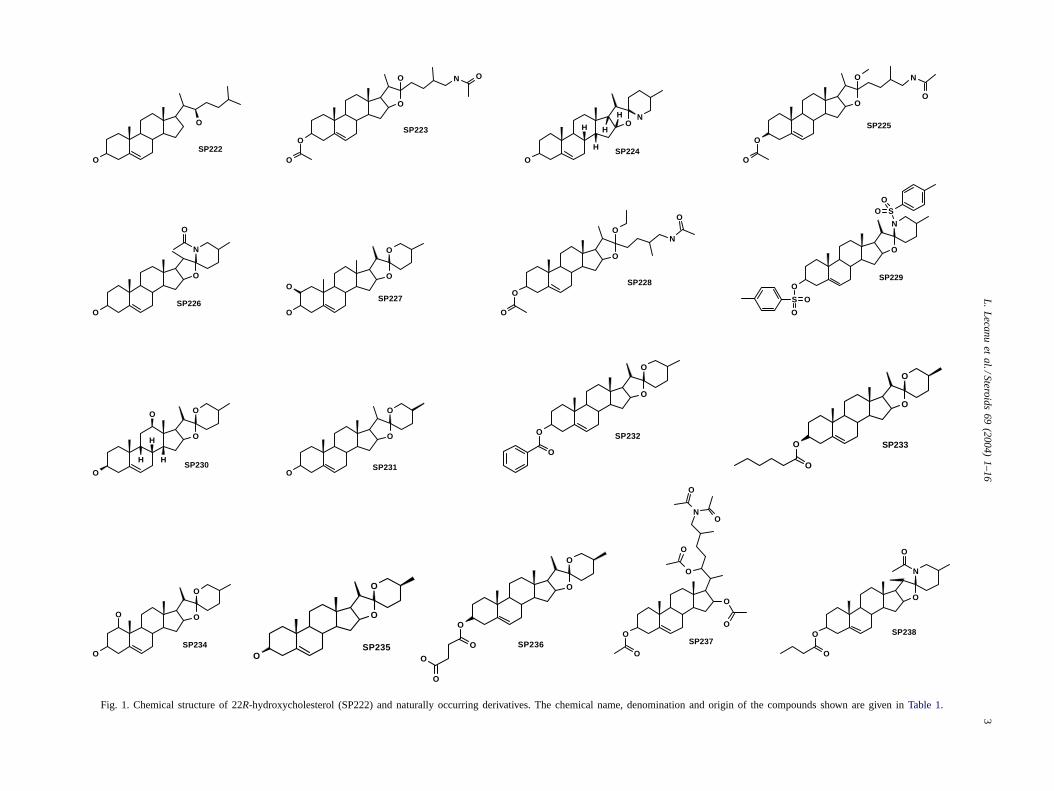
The Interbioscreen Database of naturally occurringentities was screened for compounds containing the22R-hydroxycholesterol structure using the ISIS software(Information Systems Inc., San Leandro, CA). The structureof the selected and tested 22R-hydroxycholesterol (SP222)and derivatives (SP223–238) are shown inFig. 1 and thedenomination, chemical name and origin for each of thesecompounds is shown inTable 1.
2.3. Cell culture and treatments
PC12 cells (rat pheochromocytoma neurons) from ATCC(Manassas, VA) were cultured at 37◦C and 5% CO2 in RPMI1640 medium devoid of glutamine and supplemented with10% fetal bovine serum and 5% horse serum[31]. Cellswere seeded in 96-well plates (8×104 cells per well). Afteran overnight period of incubation, increasing concentrationsof freshly solubilized A� (0.1, 1 and 10�M) were addedto the cells in the presence or absence of the indicated con-centrations of the SP compounds to be tested. After 72-hincubation time, various parameters markers of cell viabilitywere determined.
Mouse MA-10 tumor Leydig cells were maintained at37◦C in DMEM/Ham’s F12 (Biofluids, Rockville, MD)medium supplemented with 5% heat-inactivated fetal calfserum and 2.5% horse serum in 5% CO2. Cells were platedon 96-well plates at the density of 2.5×104 cells per well forovernight. The cells were stimulated with the indicated con-centrations of the SP compounds in 0.2 ml per well serum-free medium for 2 h. Culture media were collected andtested for progesterone production by radioimmunoassay.
2.4. MTT cytotoxicity assay
The cellular toxicity of A� was assessed using the3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bro-mide (MTT) assay (Trevigen, Gaithersburg, MD). Briefly,10�l of the MTT solution were added to the cells culturedin 100�l medium. After an incubation period of 4 h, 100�lof detergent were added and cells were incubated overnightat 37◦C. Formazan blue formation was quantified at 600 and690 nm using the Victor quantitative detection spectropho-tometer (EGG-Wallac, Gaithersburg, MD) and the resultsexpressed as (DO600 − DO690). Although the MTT assayhas been widely used to assess cytotoxicity in neuronalcells treated with A�, it has been suggested that the resultsobtain in the presence of various steroids might reflect theA�-dependent vesicle recycling leading to increased MTT
L.
Lecanu
etal./Steroids
69(2004)
1–163
O
O
O
NO O
SP223
NO
O
H
H
H
H
SP224
O
O
O
N
O
O
SP225
O
O
O
O
SP227
O
O
O
O
N
O
SP228
N
O
O
S O
O
SOO
SP229
O
O
O
O
H
H
HSP230
O
O
OSP231
O
O
O
O
SP232
O
O
O
O
SP234O
O
O
O
NO
O
O
O
SP237
N
O
O
O
O
SP238
O
O
SP222
N
O
O
O
SP226
O
O
O
O
O
O SP236
O
O
O
O
SP233
O
O
OSP235
Fig. 1. Chemical structure of 22R-hydroxycholesterol (SP222) and naturally occurring derivatives. The chemical name, denomination and origin of the compounds shown are given inTable 1.

4 L. Lecanu et al. / Steroids 69 (2004) 1–16
Table 1Chemical name, denomination and origin of naturally occurring compounds containing the 22R-hydroxycholesterol (SP222) lead structure
Denomination Chemical name Origin
SP222 22R-Hydroxycholesterol MammalianSP223 (20�)-26-Acetylamino-(22R)-hydroxyfurost-5-en-3�-yl acetate Gynura sp. (asteraceae)SP224 (20�)-25�-Methyl-(22R,26)-azacyclofurost-5-en-3�-ol Solanum asperum (solanaceae)SP225 (20�)-26-Acetylamino-(22R)-methoxyfurost-5-en-3�-yl acetate Gynura sp. (asteraceae)SP226 (20�)-25�-Methyl-N-acetyl-(22R,26)-azacyclofurost-5-en-3�-ol Solanum asperum (solanaceae)SP227 (22S,25�)-(20�)-Spirost-5-en-(2�,3�)-diol Gynura japonica (asteraceae)SP228 (20�)-26-Acetylamino-(22�)-ethoxyfurost-5-en-3�-yl acetate Gynura sp. (asteraceae)SP229 (20�)-25�-Methyl-N-paratoluenesulfonyl-(22S,26)-azacyclofurost-5-en-3�-yl
paratoluenesulfonateSolanum aviculare (solanaceae)
SP230 (22R,25�)-(20�)-(14�,20�)-Spirost-5-en-(3�,12�)-diol G. japonica (asteraceae)SP231 (22R,25S)-(20�)-Spirost-5-en-3�-ol G. japonica (asteraceae)SP232 (22R,25�)-(20�)-Spirost-5-en-3�-yl benzoate Gynura sp. (asteraceae)SP233 (22S,25S)-(20S)-Spirost-5-en-3�-yl hexanoate Gynura sp. (asteraceae)SP234 (22R,25�)-(20�)-Spirost-5-en-(1�,3�)-diol G. japonica (asteraceae)SP235 (22R,25S)-(20�)-Spirost-5-en-3�-ol G. japonica (asteraceae)SP236 (22R,25S)-(20�)-Spirost-5-en-3�-yl succinate Gynura sp. (asteraceae)SP237 26-Diacetylamino-(22�)-acetoxy-(16�)-acetoxy-cholest-5-en-yl acetate Achlya heterosexualis (saprolegniaceae)SP238 (20�)-25S-Methyl-N-acetyl-(22S,26)-azacyclofurost-5-en-3�-yl propanoate Solanum asperum (solanaceae)
formazan exocytosis and loss[6]. For that reason, additionalcytotoxicity and cell viability assays were used.
2.5. Trypan blue cell viability measurement
Cell viability was measured using the trypan blue exclu-sion method as we previously described[31]. In brief, cellswere treated for 72 h with SP compounds in the presenceor absence of increasing concentrations of A�. At the endof the incubation, cells were washed three times with PBSand incubated for 15 min with 0.1% trypan blue stain so-lution at room temperature. After washing three times withPBS, 0.1N NaOH was added to the cells and trypan bluewas quantified using the Victor quantitative detection spec-trophotometer at 450 nm.
2.6. Measurement of membrane potential
Cells viability was also assessed using the luminescence-based kit CytoLiteTM (Packard BioScience Co.) accord-ing to the recommendations of the manufacturer. Briefly,cells were cultured and treated in 96-well plates and af-ter 72-h incubation time, 25�l of Activator solution wasadded to the cells followed by 150�l of Amplifier solu-tion. Luminescence was measured on a TopCount NXTTM
counter (Packard BioSciences Co.) following a 5 min pre-count delay.
2.7. Determination of cellular ATP levels
Cellular ATP concentrations were measured using theATPLite-MTM luminescence assay (Packard BioSciencesCo.). For this assay, cells were cultured on black 96-wellViewPlateTM and the ATP concentrations were measured
on a TopCount NXTTM counter (Packard BioSciences Co.)following the recommendations of the manufacturer.
2.8. Radioimmunoassay
Progesterone production by MA-10 cells was measured byradioimmunoassay using anti-progesterone antisera (ICN,Costa Mesa, CA), following the conditions recommendedby the manufacturer. The progesterone production was nor-malized to the amount of protein in each well. Radioim-munoassay data was analyzed using the MultiCalc software(EG&G Wallac, Gaithersburg, MD).
2.9. 22R-hydroxycholesterol-protein binding blot assay(CPBBA)
Purified A� (50�M) and 3H-22R-hydroxycholesterol(SP222) were incubated either alone or in the presence of100�M of unlabeled 22R-hydroxycholesterol (SP222) orthe various 22R-hydroxycholesterol (SP222) derivatives in20�l volume for 8 or 24 h at 37◦C. At the end of the incu-bation time, samples were separated by 1.5% agarose (TypeI-B) gel electrophoresis under native conditions and trans-ferred to nitrocellulose membrane (Schleicher&Schuell,Keene, NH) in 10× SSC buffer. The membrane was exposedto tritium-sensitive screen and analyzed by phosphoimag-ing using the Cyclone Storage phosphor system (PackardBioScience). Image-densitometric analysis was performedusing the OptiQuant software (Packard BioScience). Thismethod allows for the separation, visualization and identifi-cation of A� complexes, which have incorporated radiola-beled cholesterol[32] and 22R-hydroxycholesterol[30] or22R-hydroxycholesterol derivatives under native conditions.Low molecular weight unincorporated 22R- hydroxycholes-
L. Lecanu et al. / Steroids 69 (2004) 1–16 5
terol and derivatives are separated and eliminated duringelectrophoresis.
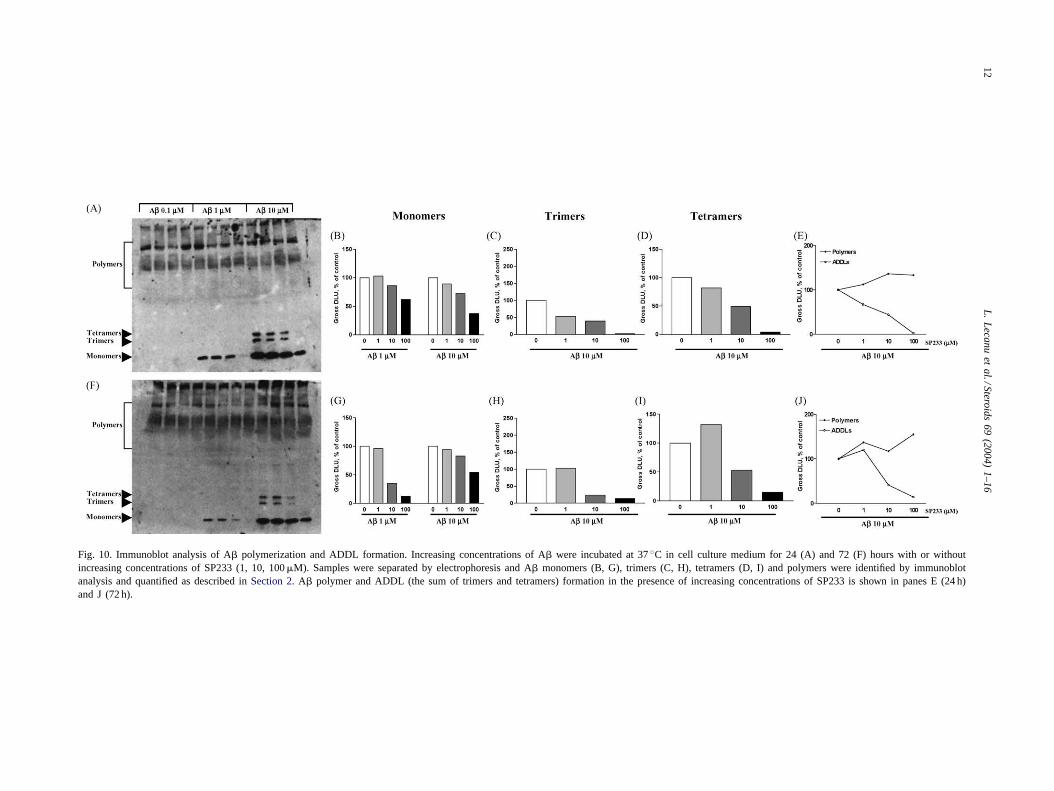
2.10. Western blot analysis of Aβ polymerization andamyloid-derived diffusible ligand (ADDL) formation
Increasing concentration of A� (0.1, 1 and 10�M) wereincubated in PC12 culture medium for 24 and 72 h at 37◦Cunder 5% CO2 with or without increasing concentrations ofSP233 (1, 10 and 100�M). At the end of the incubationtime, samples were separated by 4–20% Tris-Glycine gelelectrophoresis (Invitrogen, Carlsbad, CA) under native con-ditions at 125 V for 2 h and electrotrasferred to nitrocellu-lose membrane (HybondTM ECLTM, Amersham PharmaciaBiotech, Piscataway, NJ) at 130 A for 30 min. Non-specificadsorption of the antibodies was blocked by incubating thenitrocellulose in 5% milk. The blots were treated for im-munodetection of A� species using a polyclonal antibodyto A� that recognizes a 30 amino acid peptide of the A�protein (Zymed Laboratories, San Francisco, CA). Mem-branes were incubated in primary antibody for 1 h at roomtemperature at a dilution of 1:2000. Then, membranes wereincubated in the secondary antibody at a dilution of 1:1000for 1.5 h at room temperature. The blots were visualized us-ing the ECLTM Western Blotting Analysis System (Amer-sham Biosciences). Image-densitometric analysis was per-formed using the OptiQuant software (Packard BioScience,Meriden, CT). This method allows for the separation, visu-alization and identification of Aß complexes, polymers andADDLs.
2.11. Peptide modeling and docking simulations
Computer docking of 22R-hydroxycholesterol (SP222)and 16 of its derivatives with A�1–42 was accomplishedusing an A� structure initialized by the solution structure ofA�1–40 Met(O) (MMDB Id: 7993 PDB Id:1BA) resultingfrom data generated by CD and NMR spectroscopy[33].The Met(O) SME 35 residue was replaced by Met retainingthe adjacent backbone dihedral angles and the I41 and A42residues appended. The energy of the structure was thenminimized using the Alchemy 2000 program (Tripos, St.Louis, MO). The 22R-hydroxycholesterol derivative struc-tures were also generated using Alchemy 2000. Moleculardocking was accomplished using Monte Carlo simulatedannealing as previously described[34] implemented inmodified versions of Autogrid/Autodock[35]. For each ofthe compounds/A� pairs, approximately 108 conformationswere evaluated to obtain the selected one of minimum en-ergy. Three sessions consisting of 100 runs, each startingat a random initial relative location and orientation of theligand with respect to the target were executed. Each runwas comprised of 100 annealing cycles using about 2× 104
improvement steps. The average computation time for eachligand/target pair was about 21/2 h using a 1.7 GHz, 1 GBRAM PC.
2.12. Statistical analysis
Statistical analysis was performed by one-way analysis ofvariance (ANOVA) and unpaired Student’st-test using theINSTAT 3.00 (GraphPad, San Diego, CA).
3. Results
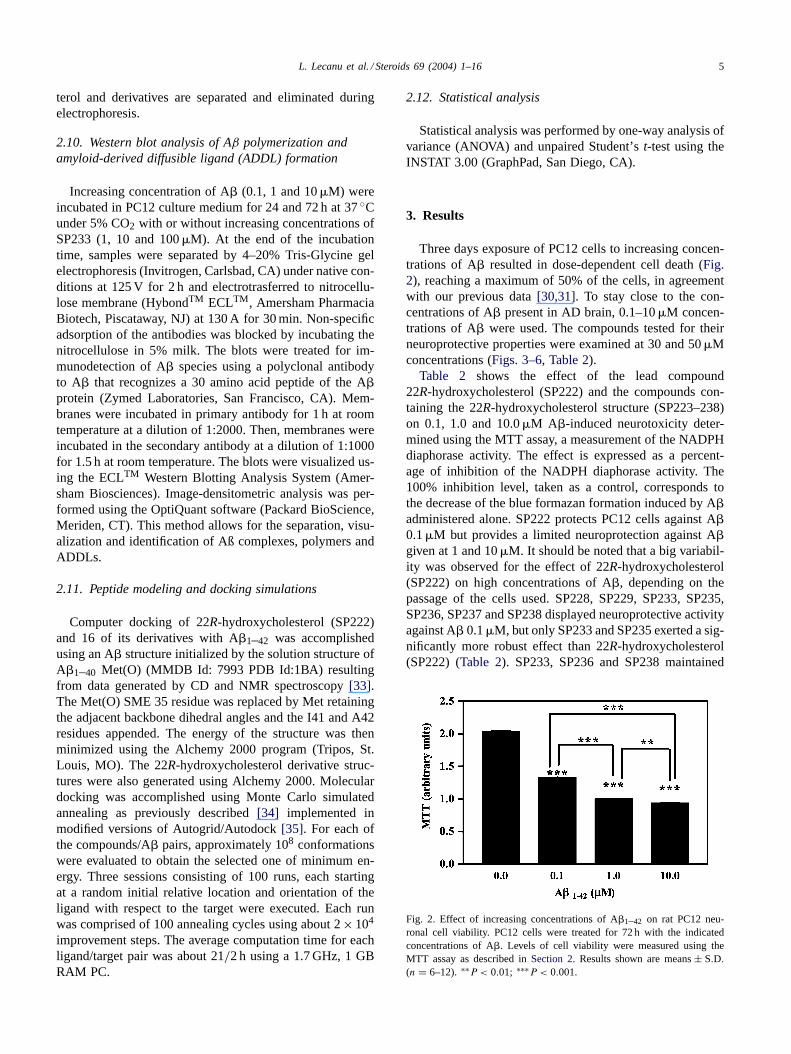
Three days exposure of PC12 cells to increasing concen-trations of A� resulted in dose-dependent cell death (Fig.2), reaching a maximum of 50% of the cells, in agreementwith our previous data[30,31]. To stay close to the con-centrations of A� present in AD brain, 0.1–10�M concen-trations of A� were used. The compounds tested for theirneuroprotective properties were examined at 30 and 50�Mconcentrations (Figs. 3–6, Table 2).
Table 2 shows the effect of the lead compound22R-hydroxycholesterol (SP222) and the compounds con-taining the 22R-hydroxycholesterol structure (SP223–238)on 0.1, 1.0 and 10.0�M A�-induced neurotoxicity deter-mined using the MTT assay, a measurement of the NADPHdiaphorase activity. The effect is expressed as a percent-age of inhibition of the NADPH diaphorase activity. The100% inhibition level, taken as a control, corresponds tothe decrease of the blue formazan formation induced by A�administered alone. SP222 protects PC12 cells against A�0.1�M but provides a limited neuroprotection against A�given at 1 and 10�M. It should be noted that a big variabil-ity was observed for the effect of 22R-hydroxycholesterol(SP222) on high concentrations of A�, depending on thepassage of the cells used. SP228, SP229, SP233, SP235,SP236, SP237 and SP238 displayed neuroprotective activityagainst A� 0.1�M, but only SP233 and SP235 exerted a sig-nificantly more robust effect than 22R-hydroxycholesterol(SP222) (Table 2). SP233, SP236 and SP238 maintained
Fig. 2. Effect of increasing concentrations of A�1–42 on rat PC12 neu-ronal cell viability. PC12 cells were treated for 72 h with the indicatedconcentrations of A�. Levels of cell viability were measured using theMTT assay as described inSection 2. Results shown are means± S.D.
(n = 6–12). ∗∗P < 0.01; ∗∗∗P < 0.001.
6 L. Lecanu et al. / Steroids 69 (2004) 1–16
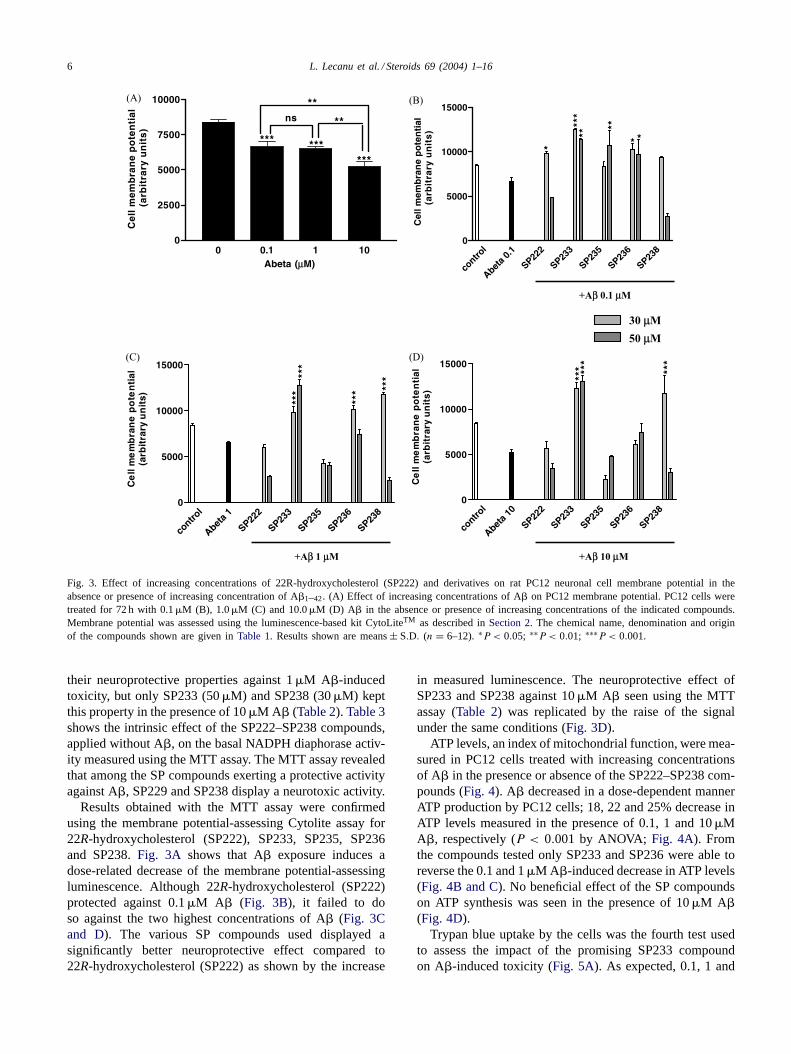
Fig. 3. Effect of increasing concentrations of 22R-hydroxycholesterol (SP222) and derivatives on rat PC12 neuronal cell membrane potential in theabsence or presence of increasing concentration of A�1–42. (A) Effect of increasing concentrations of A� on PC12 membrane potential. PC12 cells weretreated for 72 h with 0.1�M (B), 1.0�M (C) and 10.0�M (D) A� in the absence or presence of increasing concentrations of the indicated compounds.Membrane potential was assessed using the luminescence-based kit CytoLiteTM as described inSection 2. The chemical name, denomination and originof the compounds shown are given inTable 1. Results shown are means± S.D. (n = 6–12). ∗P < 0.05; ∗∗P < 0.01; ∗∗∗P < 0.001.
their neuroprotective properties against 1�M A�-inducedtoxicity, but only SP233 (50�M) and SP238 (30�M) keptthis property in the presence of 10�M A� (Table 2). Table 3shows the intrinsic effect of the SP222–SP238 compounds,applied without A�, on the basal NADPH diaphorase activ-ity measured using the MTT assay. The MTT assay revealedthat among the SP compounds exerting a protective activityagainst A�, SP229 and SP238 display a neurotoxic activity.
Results obtained with the MTT assay were confirmedusing the membrane potential-assessing Cytolite assay for22R-hydroxycholesterol (SP222), SP233, SP235, SP236and SP238.Fig. 3A shows that A� exposure induces adose-related decrease of the membrane potential-assessingluminescence. Although 22R-hydroxycholesterol (SP222)protected against 0.1�M A� (Fig. 3B), it failed to doso against the two highest concentrations of A� (Fig. 3Cand D). The various SP compounds used displayed asignificantly better neuroprotective effect compared to22R-hydroxycholesterol (SP222) as shown by the increase
in measured luminescence. The neuroprotective effect ofSP233 and SP238 against 10�M A� seen using the MTTassay (Table 2) was replicated by the raise of the signalunder the same conditions (Fig. 3D).
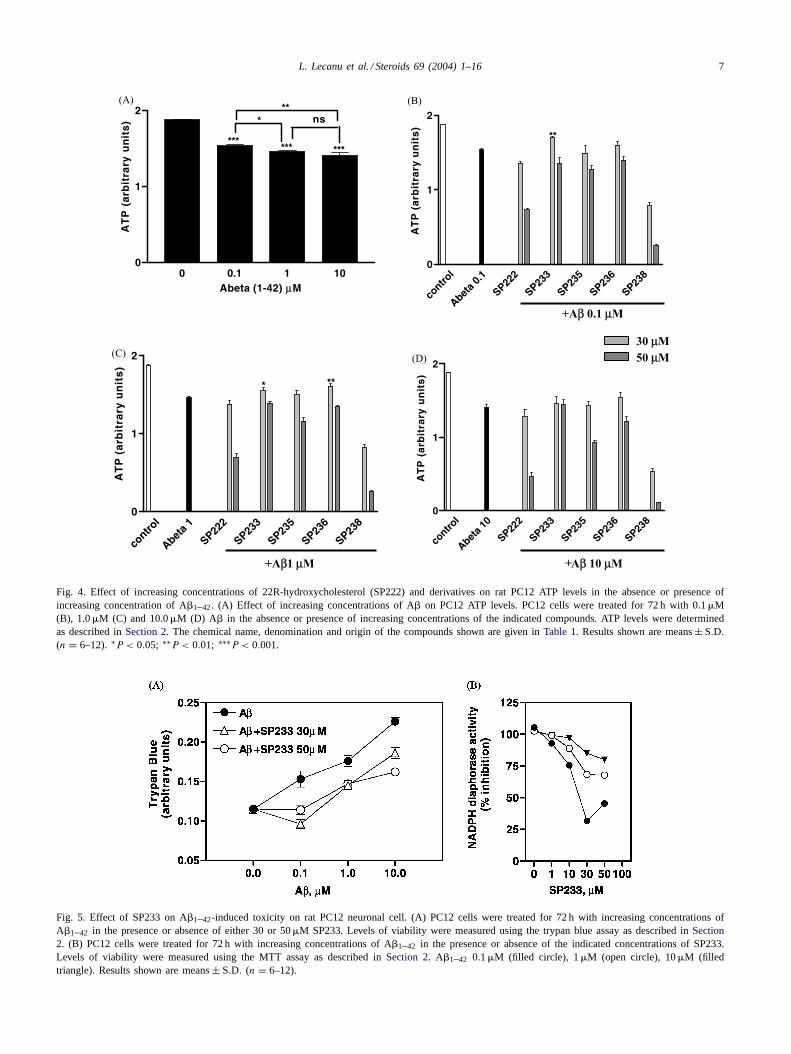
ATP levels, an index of mitochondrial function, were mea-sured in PC12 cells treated with increasing concentrationsof A� in the presence or absence of the SP222–SP238 com-pounds (Fig. 4). A� decreased in a dose-dependent mannerATP production by PC12 cells; 18, 22 and 25% decrease inATP levels measured in the presence of 0.1, 1 and 10�MA�, respectively (P < 0.001 by ANOVA; Fig. 4A). Fromthe compounds tested only SP233 and SP236 were able toreverse the 0.1 and 1�M A�-induced decrease in ATP levels(Fig. 4B and C). No beneficial effect of the SP compoundson ATP synthesis was seen in the presence of 10�M A�(Fig. 4D).
Trypan blue uptake by the cells was the fourth test usedto assess the impact of the promising SP233 compoundon A�-induced toxicity (Fig. 5A). As expected, 0.1, 1 and
L. Lecanu et al. / Steroids 69 (2004) 1–16 7
Fig. 4. Effect of increasing concentrations of 22R-hydroxycholesterol (SP222) and derivatives on rat PC12 ATP levels in the absence or presence ofincreasing concentration of A�1–42. (A) Effect of increasing concentrations of A� on PC12 ATP levels. PC12 cells were treated for 72 h with 0.1�M(B), 1.0�M (C) and 10.0�M (D) A� in the absence or presence of increasing concentrations of the indicated compounds. ATP levels were determinedas described inSection 2. The chemical name, denomination and origin of the compounds shown are given inTable 1. Results shown are means± S.D.
(n = 6–12). ∗P < 0.05; ∗∗P < 0.01; ∗∗∗P < 0.001.
Fig. 5. Effect of SP233 on A�1–42-induced toxicity on rat PC12 neuronal cell. (A) PC12 cells were treated for 72 h with increasing concentrations ofA�1–42 in the presence or absence of either 30 or 50�M SP233. Levels of viability were measured using the trypan blue assay as described inSection2. (B) PC12 cells were treated for 72 h with increasing concentrations of A�1–42 in the presence or absence of the indicated concentrations of SP233.Levels of viability were measured using the MTT assay as described inSection 2. A�1–42 0.1�M (filled circle), 1�M (open circle), 10�M (filledtriangle). Results shown are means± S.D. (n = 6–12).
8 L. Lecanu et al. / Steroids 69 (2004) 1–16
Table 2PC12 cells were treated for 72 h with 0.1, 1.0, and 10.0�M A� in the absence or presence of increasing concentrations of the indicated compounds
Compounds(�M)
A� (1–42) �M Compounds(�M)
A� (1–42) �M
0.1�M 1 �M 10�M 0.1�M 1 �M 10�M
Control 100± 6.7 100± 8.9 100± 5.1SP222 (30) 68± 9.3∗∗∗ 96.4 ± 4.5 109.5± 4.5SP222 (50) 98.1± 14.8 112± 9.9 131± 5.7SP223 (30) 104.8± 2.8 109± 1.9 111.1± 1.4 SP231 (30) 101.9± 11.2 97.9± 1.6 105.9± 3.0SP223 (50) 93.2± 7.5 100.0± 3 104± 3 SP231 (50) 97.1± 2.7 91.6± 0.3 103.0± 3.5SP224 (30) 130.5± 2.1 108.1± 1.2 105.3± 3.3 SP232 (30) 165.9± 10.1 108.1± 4.8 98.4± 2.4SP224 (50) 151.4± 2.6 117.6± 0.8 103.7± 3.9 SP232 (50) 162.9± 8.0 106.6± 2.7 97.7± 1.3SP225 (30) 294.3± 23.9 181.4± 7.3 151± 6.9 SP233 (30) 75.6± 1.2∗∗∗ 89.0 ± 3.6 ∗∗,++ 97.0 ± 4.6SP225 (50) 335.6± 9.3 198.6± 7.8 161.4± 3.8 SP233 (50) 31.6± 4.4 ∗∗∗,++ 68.2 ± 3.4+++,∗∗∗ 85.1 ± 4.1∗∗∗,+++SP226 (30) 183.4± 12.0 132.0± 6.6 120.3± 4.0 SP234 (30) 214.0± 3.2 127.0± 9.2 112.8± 10.3SP226 (50) 169.3± 11.9 124.8± 8.2 124.0± 7.7 SP234 (50) 371.6± 42.5 180.6± 14.3 129.8± 7.9SP227 (30) 109.2± 4.4 103.1± 3.2 108.8± 2.0 SP235 (30) 76.0± 2.2∗∗∗ 102.1± 5.7 103.0± 6.0SP227 (50) 113.7± 12.7 107.6± 8.3 112.6± 3.8 SP235 (50) 48.3± 8.3∗∗∗,++ 103.8± 8.6 123.8± 3.7SP228 (30) 106.1± 2.8 107.8± 0.3 107.1± 0.7 SP236 (30) 82.5± 1.9 ∗∗∗,++ 90.0 ± 1.0∗,+ 90.1 ± 20.4SP228 (50) 91.4± 2.3∗∗ 97.5 ± 3.1 101.2± 0.5 SP236 (50) 71.8± 3.1∗∗∗,++ 90.5 ± 1.9∗,++ 106.2± 5.5SP229 (30) 83.5± 4.7∗∗,++ 91.6 ± 4.6∗ 98.5 ± 15.9 SP237 (30) 83.6± 12.8∗,++ 100.1± 4.8 119.5± 2.9SP229 (50) 98.1± 4.5 128± 0.4 121.1± 5.4 SP237 (50) 369.2± 44.1 311.5± 1.0 282.4± 2.1SP230 (30) 101.0± 8.3 95.6± 8.3 101.7± 3.0 SP238 (30) 82.9± 2.6∗∗∗,++ 73.6 ± 1.0∗∗∗,++ 72.0 ± 1.8∗∗∗,+++SP230 (30) 154.1± 7.0 125.5± 2.4 116.2± 3.3 SP238 (50) 179.7± 3.5 155.8± 2.3 155.3± 1.4
Levels of cell viability were measured using the MTT assay as described inSection 2. The chemical name, denomination and origin of the compoundsshown are given inTable 1. The 100% inhibition level corresponds to the decrease of the blue formazan formation induced by A� administered alone.Results shown are means± S.D. (n = 6–12). ∗P < 0.05. ∗∗P < 0.01. ∗∗∗P < 0.001 compared to control and+P < 0.05. ++P < 0.01. +++P < 0.001compared to SP222-treated cells.
10�M A�-induced a dose-dependent (33, 36 and 97%,respectively;P < 0.001 by ANOVA) increase in trypanblue uptake by PC12 cells. SP233 at 30 and 50�M inhib-ited the A�-induced cell death (P < 0.001 by ANOVA).Fig. 5B shows that the neuroprotective effect of SP233 isdose-dependent and it is maintained in the presence of allthree concentrations of A�, although its efficacy decreasesin presence of high, supra-physiopathological, A� concen-trations.
One of the reasons in identifying 22R-hydroxycholesterol(SP222) derivatives is the need of biologically active (neu-roprotective) compounds that cannot be metabolized byP450scc to pregnenolone and then to tissue-specific finalsteroid products. To assess the metabolism of SP233 bysteroidogenic cells we examined their ability to form steroids
Table 3Effect of the SP compounds on the PC12 cells basal NADPH diaphorase activity measured using the MTT assay
SP222 SP223 SP224 SP225 SP226 SP227
30�M 109.3 ± 4.9 93.1± 2.1 47.0± 0.6 84.0± 1.5 38.3± 1.5 88.5± 2.050�M 91.7 ± 4.0 97.1± 8.3 32.9± 3.0 72.7± 0.9 38.0± 1.1 72.5± 8.4
SP228 SP229 SP230 SP231 SP232 SP233
30�M 89.9 ± 0.6 79.8± 1.6 97.1± 1.7 68.0± 1.3 49.0± 2.5 106.0± 0.850�M 97.0 ± 0.2 69.0± 2.6 52.3± 2.6 68.0± 0.2 50.1± 0.8 108.0± 0.7
SP234 SP235 SP236 SP237 SP238
30�M 102.8 ± 4.0 93.6± 2.1 94.6± 0.5 81.1± 3.0 62.0± 0.450�M 95.8 ± 2.0 95.9± 1.6 86.3± 2.1 5.37± 0.46 18.3± 2.0
The data revealed that among the SP compounds exerting a protective activity against A�, SP229 and SP238 display a neurotoxic activity. Results shownare means± S.D. (n = 6–12).
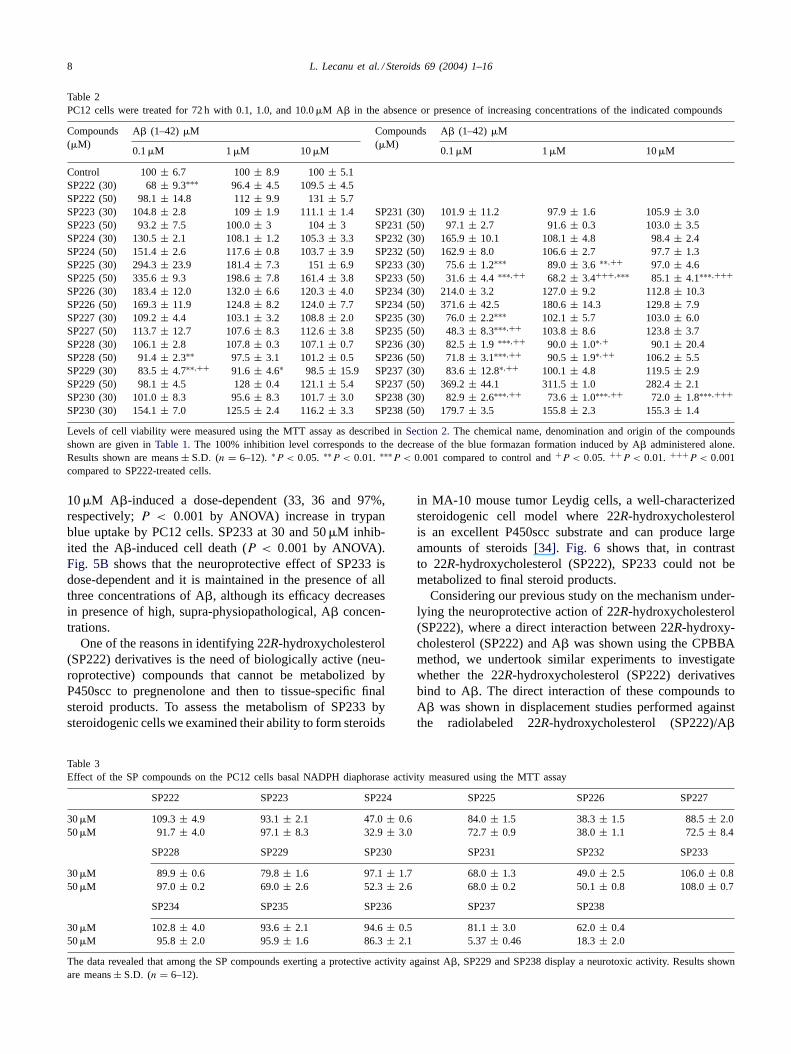
in MA-10 mouse tumor Leydig cells, a well-characterizedsteroidogenic cell model where 22R-hydroxycholesterolis an excellent P450scc substrate and can produce largeamounts of steroids[34]. Fig. 6 shows that, in contrastto 22R-hydroxycholesterol (SP222), SP233 could not bemetabolized to final steroid products.
Considering our previous study on the mechanism under-lying the neuroprotective action of 22R-hydroxycholesterol(SP222), where a direct interaction between 22R-hydroxy-cholesterol (SP222) and A� was shown using the CPBBAmethod, we undertook similar experiments to investigatewhether the 22R-hydroxycholesterol (SP222) derivativesbind to A�. The direct interaction of these compounds toA� was shown in displacement studies performed againstthe radiolabeled 22R-hydroxycholesterol (SP222)/A�
L. Lecanu et al. / Steroids 69 (2004) 1–16 9
0 10 20 300
250
500
750
SP222SP233SP235SP236SP238
Pro
ges
tero
ne
ng
/mg
pro
tein
Fig. 6. Effect of SP233 on MA-10 Leydig cell steroid formation.MA-10 cells were treated for 2 h in the presence or absence of either22R-hydroxycholesterol (SP222) or SP233 tested at the indicated con-centrations. Progesterone levels were determined by radioimmunoassay.Results shown are means± S.D. (n = 9).
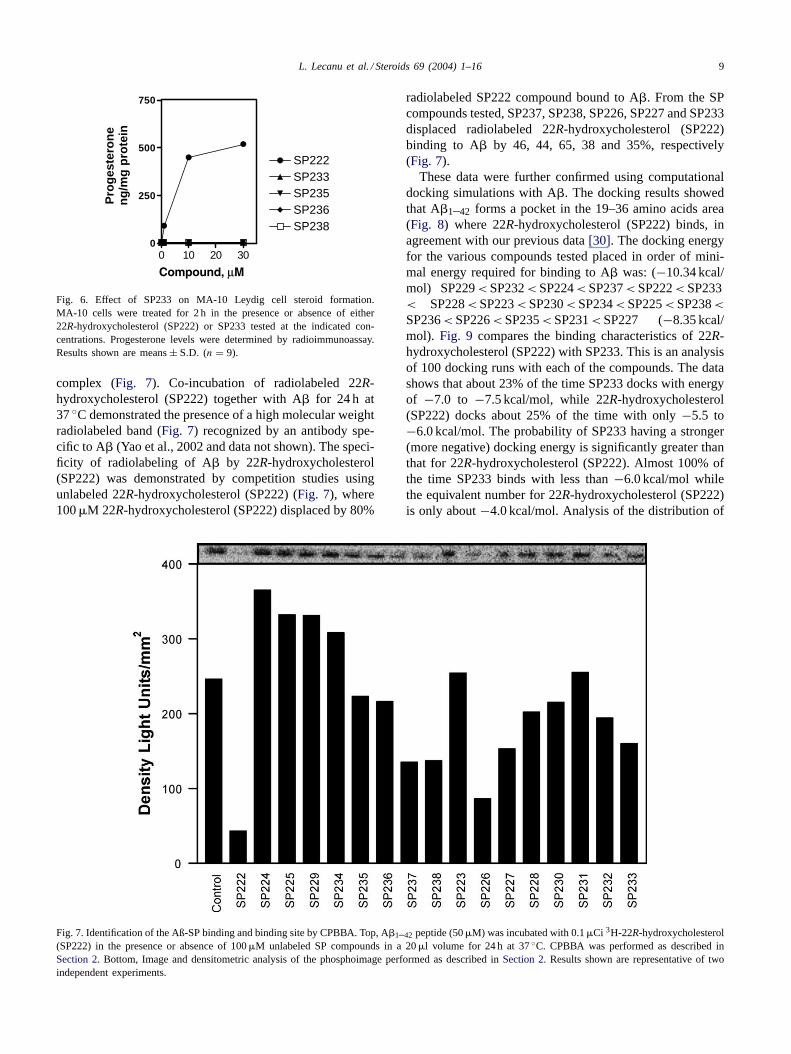
complex (Fig. 7). Co-incubation of radiolabeled 22R-hydroxycholesterol (SP222) together with A� for 24 h at37◦C demonstrated the presence of a high molecular weightradiolabeled band (Fig. 7) recognized by an antibody spe-cific to A� (Yao et al., 2002 and data not shown). The speci-ficity of radiolabeling of A� by 22R-hydroxycholesterol(SP222) was demonstrated by competition studies usingunlabeled 22R-hydroxycholesterol (SP222) (Fig. 7), where100�M 22R-hydroxycholesterol (SP222) displaced by 80%
Fig. 7. Identification of the Aß-SP binding and binding site by CPBBA. Top, A�1–42 peptide (50�M) was incubated with 0.1�Ci 3H-22R-hydroxycholesterol(SP222) in the presence or absence of 100�M unlabeled SP compounds in a 20�l volume for 24 h at 37◦C. CPBBA was performed as described inSection 2. Bottom, Image and densitometric analysis of the phosphoimage performed as described inSection 2. Results shown are representative of twoindependent experiments.
radiolabeled SP222 compound bound to A�. From the SPcompounds tested, SP237, SP238, SP226, SP227 and SP233displaced radiolabeled 22R-hydroxycholesterol (SP222)binding to A� by 46, 44, 65, 38 and 35%, respectively(Fig. 7).
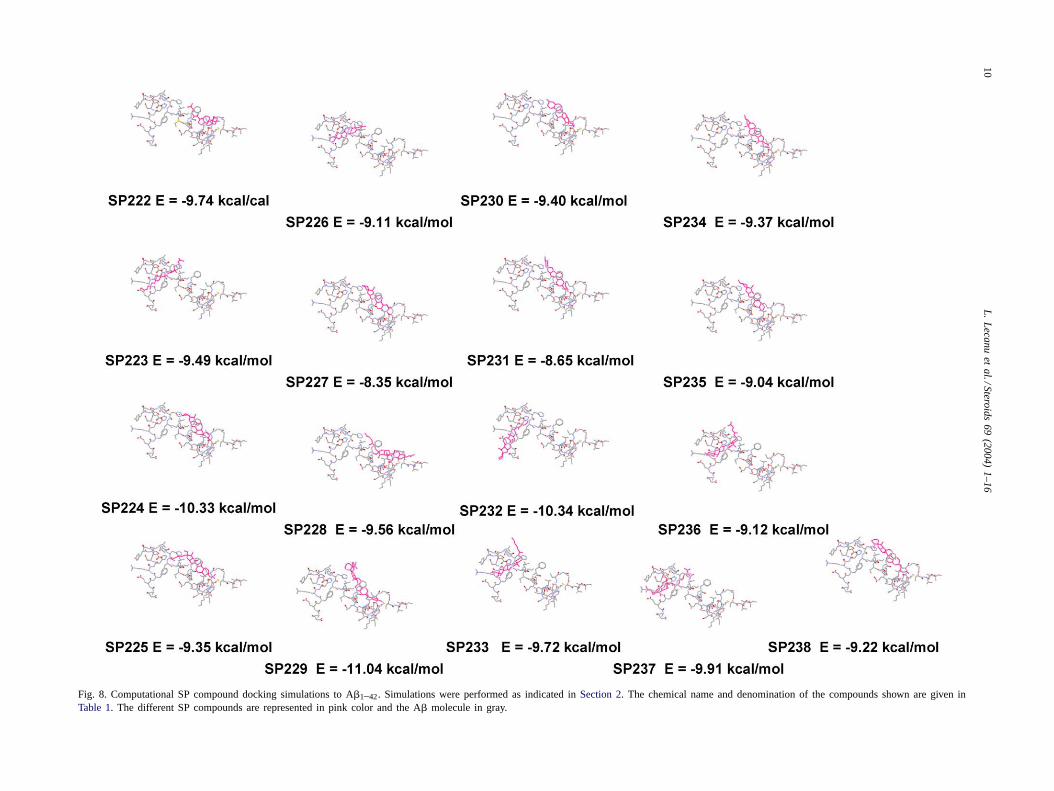
These data were further confirmed using computationaldocking simulations with A�. The docking results showedthat A�1–42 forms a pocket in the 19–36 amino acids area(Fig. 8) where 22R-hydroxycholesterol (SP222) binds, inagreement with our previous data[30]. The docking energyfor the various compounds tested placed in order of mini-mal energy required for binding to A� was: (−10.34 kcal/mol) SP229< SP232< SP224< SP237< SP222< SP233< SP228< SP223< SP230< SP234< SP225< SP238<SP236< SP226< SP235< SP231< SP227 (−8.35 kcal/mol). Fig. 9 compares the binding characteristics of 22R-hydroxycholesterol (SP222) with SP233. This is an analysisof 100 docking runs with each of the compounds. The datashows that about 23% of the time SP233 docks with energyof −7.0 to −7.5 kcal/mol, while 22R-hydroxycholesterol(SP222) docks about 25% of the time with only−5.5 to−6.0 kcal/mol. The probability of SP233 having a stronger(more negative) docking energy is significantly greater thanthat for 22R-hydroxycholesterol (SP222). Almost 100% ofthe time SP233 binds with less than−6.0 kcal/mol whilethe equivalent number for 22R-hydroxycholesterol (SP222)is only about−4.0 kcal/mol. Analysis of the distribution of
10L
.L
ecanuet
al./Steroids69
(2004)1–16
Fig. 8. Computational SP compound docking simulations to A�1–42. Simulations were performed as indicated inSection 2. The chemical name and denomination of the compounds shown are given inTable 1. The different SP compounds are represented in pink color and the A� molecule in gray.
L. Lecanu et al. / Steroids 69 (2004) 1–16 11
Binding Energy Frequencies
0
5
10
15
20
25
30
-10 -9 -8 -7 -6 -5 -4
Energy, KCal/mol
Fre
qu
ency
-%
SP222
SP233
Binding Energy Probabilities
.00%
20.00%
40.00%
60.00%
80.00%
100.00%
-10 -9 -8 -7 -6 -5 -4
Energy, Kcal/mol
Pro
bab
ility
SP222
SP233
(A)
(B)
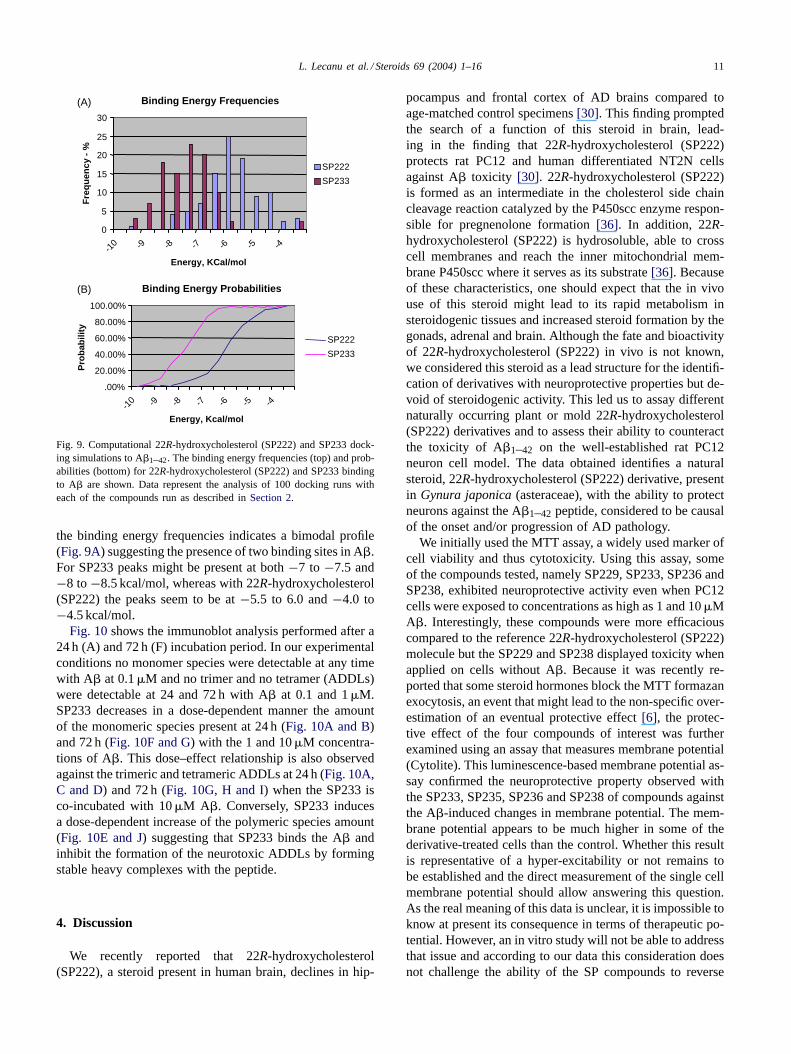
Fig. 9. Computational 22R-hydroxycholesterol (SP222) and SP233 dock-ing simulations to A�1–42. The binding energy frequencies (top) and prob-abilities (bottom) for 22R-hydroxycholesterol (SP222) and SP233 bindingto A� are shown. Data represent the analysis of 100 docking runs witheach of the compounds run as described inSection 2.
the binding energy frequencies indicates a bimodal profile(Fig. 9A) suggesting the presence of two binding sites in A�.For SP233 peaks might be present at both−7 to −7.5 and−8 to−8.5 kcal/mol, whereas with 22R-hydroxycholesterol(SP222) the peaks seem to be at−5.5 to 6.0 and−4.0 to−4.5 kcal/mol.
Fig. 10shows the immunoblot analysis performed after a24 h (A) and 72 h (F) incubation period. In our experimentalconditions no monomer species were detectable at any timewith A� at 0.1�M and no trimer and no tetramer (ADDLs)were detectable at 24 and 72 h with A� at 0.1 and 1�M.SP233 decreases in a dose-dependent manner the amountof the monomeric species present at 24 h (Fig. 10A and B)and 72 h (Fig. 10F and G) with the 1 and 10�M concentra-tions of A�. This dose–effect relationship is also observedagainst the trimeric and tetrameric ADDLs at 24 h (Fig. 10A,C and D) and 72 h (Fig. 10G, H and I) when the SP233 isco-incubated with 10�M A�. Conversely, SP233 inducesa dose-dependent increase of the polymeric species amount(Fig. 10E and J) suggesting that SP233 binds the A� andinhibit the formation of the neurotoxic ADDLs by formingstable heavy complexes with the peptide.
4. Discussion
We recently reported that 22R-hydroxycholesterol(SP222), a steroid present in human brain, declines in hip-
pocampus and frontal cortex of AD brains compared toage-matched control specimens[30]. This finding promptedthe search of a function of this steroid in brain, lead-ing in the finding that 22R-hydroxycholesterol (SP222)protects rat PC12 and human differentiated NT2N cellsagainst A� toxicity [30]. 22R-hydroxycholesterol (SP222)is formed as an intermediate in the cholesterol side chaincleavage reaction catalyzed by the P450scc enzyme respon-sible for pregnenolone formation[36]. In addition, 22R-hydroxycholesterol (SP222) is hydrosoluble, able to crosscell membranes and reach the inner mitochondrial mem-brane P450scc where it serves as its substrate[36]. Becauseof these characteristics, one should expect that the in vivouse of this steroid might lead to its rapid metabolism insteroidogenic tissues and increased steroid formation by thegonads, adrenal and brain. Although the fate and bioactivityof 22R-hydroxycholesterol (SP222) in vivo is not known,we considered this steroid as a lead structure for the identifi-cation of derivatives with neuroprotective properties but de-void of steroidogenic activity. This led us to assay differentnaturally occurring plant or mold 22R-hydroxycholesterol(SP222) derivatives and to assess their ability to counteractthe toxicity of A�1–42 on the well-established rat PC12neuron cell model. The data obtained identifies a naturalsteroid, 22R-hydroxycholesterol (SP222) derivative, presentin Gynura japonica (asteraceae), with the ability to protectneurons against the A�1–42 peptide, considered to be causalof the onset and/or progression of AD pathology.
We initially used the MTT assay, a widely used marker ofcell viability and thus cytotoxicity. Using this assay, someof the compounds tested, namely SP229, SP233, SP236 andSP238, exhibited neuroprotective activity even when PC12cells were exposed to concentrations as high as 1 and 10�MA�. Interestingly, these compounds were more efficaciouscompared to the reference 22R-hydroxycholesterol (SP222)molecule but the SP229 and SP238 displayed toxicity whenapplied on cells without A�. Because it was recently re-ported that some steroid hormones block the MTT formazanexocytosis, an event that might lead to the non-specific over-estimation of an eventual protective effect[6], the protec-tive effect of the four compounds of interest was furtherexamined using an assay that measures membrane potential(Cytolite). This luminescence-based membrane potential as-say confirmed the neuroprotective property observed withthe SP233, SP235, SP236 and SP238 of compounds againstthe A�-induced changes in membrane potential. The mem-brane potential appears to be much higher in some of thederivative-treated cells than the control. Whether this resultis representative of a hyper-excitability or not remains tobe established and the direct measurement of the single cellmembrane potential should allow answering this question.As the real meaning of this data is unclear, it is impossible toknow at present its consequence in terms of therapeutic po-tential. However, an in vitro study will not be able to addressthat issue and according to our data this consideration doesnot challenge the ability of the SP compounds to reverse
12L
.L
ecanuet
al./Steroids69
(2004)1–16
Fig. 10. Immunoblot analysis of A� polymerization and ADDL formation. Increasing concentrations of A� were incubated at 37◦C in cell culture medium for 24 (A) and 72 (F) hours with or withoutincreasing concentrations of SP233 (1, 10, 100�M). Samples were separated by electrophoresis and A� monomers (B, G), trimers (C, H), tetramers (D, I) and polymers were identified by immunoblotanalysis and quantified as described inSection 2. A� polymer and ADDL (the sum of trimers and tetramers) formation in the presence of increasing concentrations of SP233 is shown in panes E (24 h)and J (72 h).

L. Lecanu et al. / Steroids 69 (2004) 1–16 13
the effect of A� on the membrane potential and therefore tocounteract its neurotoxicity. Amyloid aggregates have beenshown to interact with the cell membrane and modify its flu-idity [37,38]. Decrease in plasma membrane fluidity couldhamper the function of cell surface receptors and ions chan-nel proteins with deleterious consequences for cell survival.Considering these observations, the results obtained with theSP compounds suggest that these compounds preserve theintegrity of the cytoplasmic membrane.
A late event in the mechanism of action of A� is thedirect or indirect disruption of the mitochondrial respiratorychain, leading to a decrease in ATP production that alonecould lead to cell death[39–42]. 22R-hydroxycholesterol(SP222), SP235, and SP238 compounds, which were ableto rescue the PC12 cells from A�-induced toxicity, did notblock the A�-induced changes in ATP synthesis. Althoughsuch an apparent discrepancy remains to be explained, itis possible that the MTT assay (mitochondrial diaphoraseactivity) and ATP synthesis do not reflect the status of thesame part of the respiratory chain. In contrast, SP233 andSP236 blocked, although in part, the A�-induced decreasein ATP production. The ability of SP233 to preserve ATPstocks could explain the potent neuroprotective effect of thiscompound, which was further confirmed by the trypan blueuptake cell viability assay. It should be noted that SP233, atconcentrations as low as 10�M, was found to be not onlythe most efficacious in all assays used but also the mostpotent, offering neuroprotection in vitro against A�.
The studies presented herein were performed using 0.1,1.0 and 10�M A�1–42. These concentrations are supra-physiopathological since the concentrations of A�1–42present in cerebrospinal fluid of AD patients and controlsrange from 500 to 1000 ng/l (0.1–0.2 nM)[43,44]. Even ifwe consider that A�1–42 might be present in AD brain at10 times higher concentration, the estimated pathophysio-logical concentrations of A�1–42 would be in the range of1–2 nM which is 100–10,000 times less than the concen-trations used in our experiments. With these considerationsin mind it is obvious that the 75% protection offered bySP233 against 0.1�M A� is pharmacologically relevant.
As noted above, one of the reasons of identifying bioac-tive 22R-hydroxycholesterol (SP222) derivatives was theneed of compounds that could not be metabolized to finalsteroid products in steroidogenic cells. Using the well-established MA-10 mouse Leydig cell model, we demon-strated that, unlike 22R-hydroxycholesterol (SP222), SP233failed to induce steroid formation suggesting that it is notmetabolized by P450scc.
The neuroprotective property of the SP compounds seemsto follow a structure/activity relationship (SAR). SP231and SP235 are stereoisomers of diosgenin (Fig. 1), but onlySP235 is protective against A�-induced neurotoxicity. Thestereochemistry of the SP235 is C3R, C10R, C13S, C20S,C22S, C25S, a motif shared by SP233 and SP236 (Figs. 1and 11). SP compounds exhibiting high neuroprotectiveactivity and being active in the presence of high concen-
Fig. 11. Model of the basic spirostenol structure present in the neuropro-tective SP compounds. The oxygen are represented in red, hydrogen inwhite and carbon in dark gray color. The stereochemistry of the commoncore to SP235, SP236 and SP233 is C3R, C10R, C13S, C20S, C22S, C25S.
trations of A� contained an ester, preferably a fatty acidor a fatty acid-like structure, on C3. Indeed, SP235 thatpossesses an unsubstituted hydroxyl group in C3 offeredlimited neuroprotection, acting only against 0.1�M A�. Incontrast, SP236 that is the succinic ester at C3 of SP235 isactive against higher A� concentrations, and SP233, whichis a hexanoic ester at C3 of SP235 was the most potentcompound. The finding that SP238 was able to protectPC12 cells against A�-induced toxicity, although it hadno effect on maintaining ATP levels, further supports thishypothesis, because its derivative without any side-chainon C3 (SP226) did not offer neuroprotection. The findingthat benzoic acid substitution, present on SP232, was noteffective in neuroprotection suggested that the presenceof an aliphatic chain at this level is more relevant that anaromatic structure. Although these data are indicative ofa SAR and highlights the importance of the presence of afatty acid chain at C3, further modeling and SAR studiesneed to be performed to optimize the SP233 structure forneuroprotection.
Yao et al.[30] recently demonstrated that the neuroprotec-tive effect of the 22R-hydroxycholesterol (SP222) lies in itsability to bind and inactivate A�1–42. Based on this obser-vation, we examined the ability of 22R-hydroxycholesterol(SP222) derivatives to offer neuroprotection by acting in asimilar manner. Our findings indicated that SP compoundsexhibiting neuroprotective properties against A�-inducedcell death displaced radiolabeled 22R-hydroxycholesterol(SP222) bound to the amyloid peptide. However, the inten-sity of the displacement could not be related to the level of

14 L. Lecanu et al. / Steroids 69 (2004) 1–16
protection exhibited by the different SP compounds. More-over, at present, we do not know whether this displacementcorresponds to a competitive or non-competitive behavior.To further examine the interaction of SP233 with A� weexamined the fate of A� in cell culture media incubatedwith and without SP233. A� aggregates and ADDLs wereseparated by electrophoresis and identified by immunoblotanalyses. Under our experimental conditions, the incuba-tion of 10�M A� resulted in the formation of trimers,tetramers and heavy polymers. The trimers and tetramersbelong to ADDLs, which are non fibrillar oligomers rang-ing approximately from 13 to 108 kDa[45], with potentneurotoxic properties at concentration as low as 5–10 nM[46,47]. A recent report described the ADDLs as baring theneurotoxic properties of A� [45]. Interestingly, we werenot able to detect any ADDLs formation in the presenceof 0.1 and 1�M A� whereas these concentrations induceda decrease of the cells viability raising the question of theADDLs as the only neurotoxic amyloid species. Whetherthe amounts of ADDLs formed were under the detectionlimit under these experimental conditions or ADDLs donot solely account for A� neurotoxicity remains to be es-tablished. However, SP233 decreased in a dose-dependentmanner the formation of the A� trimers and tetramersafter 24 and 72 h incubation, an event that may accountsfor its neuroprotective effect. Moreover, the SP233 wasable to decrease the amount of monomers available forADDL formation, further suggesting that SP233 binds toboth oligomers and monomers. The dose-dependent de-crease of the ADDL levels by SP233 was accompanied bya dose-dependent increase of high molecular weight poly-mer aggregation, confirming previous data obtained in ourlab with the 22R-hydroxycholesterol[31] and suggestingthat both the 22R-hydroxycholesterol and SP233 inactivateA� by binding to the peptide and forming stable non-toxicpolymers.
Computational docking simulations were used in a firstattempt to further characterize the SP-A� interaction. Thestudies revealed that two binding sites might be presenton A� for the bioactive SP compounds. One binding siteappears to be more specific for 22R-hydroxycholesterol(SP222), whereas the second binding site displays higheraffinity for compounds such as SP233 and SP236. Al-though SP226 is shown to bind to this second binding sitetoo, the calculated binding energy for this compound ismuch lower than the energy displayed by the neuroprotec-tive SP molecules. A subsequent computational dockingsimulation study indicated that the binding energies of 22R-hydroxycholesterol (SP222) and SP233 follow a bimodaldistribution, a finding that strongly supports the presenceof two binding sites on A�. Further calculation of bindingenergies indicated that 22R-hydroxycholesterol (SP222) hasless affinity for the second binding site compared to SP233and suggests that the presence of the ester chain might beresponsible for the ability of SP233 to bind to both siteson A�. Based on these observations, we hypothesize that
occupancy of the A� second binding site might be re-quired for a sustained inactivation of the amyloid peptide.This hypothesis goes along with the results obtained withthe 22R-hydroxycholesterol displacement study where weshowed that the SP compounds with the highest affinity forthe 22R-hydroxycholesterol binding site were not the bestneuroprotective agent because they did not bind or they didnot bind with a good affinity the SP233 site. We are now inthe process of testing this hypothesis in vitro and in silico.
Others mechanisms not related to a direct inactivation ofA� could also contribute to the neuroprotective activity ofSP233. A possible modulation of the steroid receptor familycannot be excluded, although little is known about the bind-ing of spirostenols on nuclear receptors. It has been shownthat A� inhibits the fusion of GLUT3-containing vesicles[42] leading to the disruption of mitochondrial homeosta-sis and, thus to neuronal death. On the other hand, the glu-cose absorption is enhanced in normal and streptozotocin-induced diabetic mice by spirostenol derivatives extractedfrom Polygonati rhizome[48]. Taken together, these resultssuggest that restoration of glucose transport inside the cellmight be a protective mechanism in our model activated bythe spirostenol SP233. Natural and synthetic derivatives ofdiosgenin have been also shown to lower cholesterol ab-sorption by the cell and to decrease cholesterol synthesisby inhibiting the key enzyme 3-hydroxy-3-methylglutarylcoenzyme A reductase[49,50]. It is also well known thatan increase of cellular cholesterol concentration induces theactivation of�- and�-secretase leading to A� production.Moreover, diosgenin derivatives have been shown to modifyintracellular cholesterol pools by inhibiting the cholesterylester transfer protein[51], an enzyme reported to positivelymodulate the generation of A� [52]. Although it is unlikelythat these protective mechanisms take place in our model,because A� is added in the culture medium, they could how-ever be part of the in vivo response to SP233.
Despite the tremendous efforts undertaken during the pastfew years to discover novel therapeutic modalities for thecure and/or slowing of the progression of AD, no majorclinical advances have been made since the introduction ofacetylcholine-esterase inhibitors, which produce modest im-provements in selected patients with mild or moderate AD[53,54]. Although many compounds are actually in clini-cal trials in an attempt to treat AD, for most of those, ADpathology is a target secondary to their primary action. Suchdrugs include antioxidants, COX-1 and COX-2 inhibitors,statins, and brain vessels vasodilators. Our results indicatethat naturally occurring spirostenol compounds may be ofinterest to protect neuronal cells against A�. The finding thatspirostenols were isolated from brain extracts of cows fedwith plants containing such compounds suggests that theycross the blood brain barrier[48,55], a property requiredfor in vivo activity. Although further in vitro and in vivostudies are required to establish the pharmacology of thesecompounds, the SP233 lead structure may constitute a newapproach for the treatment of AD.

L. Lecanu et al. / Steroids 69 (2004) 1–16 15
Acknowledgements
We would like to thank Drs. T. Reynolds and R. Hochberg(Yale University) for help with the steroid nomenclature, Dr.M. Ascoli (University of Iowa) for the MA-10 cells and Dr.M. Culty (Georgetown University) for critically reviewingthe manuscript.
References
[1] Alzheimer A. Uber eine eigenartige Erkrankung der Hirnrinde.Zeitschrift für Psychiatrie 1906;10:146–8.
[2] Selkoe DJ. Amyloid protein precursor and the pathogenesis ofAlzheimer’s disease. Cell 1989;58:611–2.
[3] Almqvist O. Neuropsychological features of early Alzheimer’sdisease: preclinical and clinical stages. Acta Neurol Scand1996;93(Suppl 185):63–71.
[4] Small BJ, Herlitz A, Bäckman L. Cognitive development inAlzheimer’s disease: charting the decline process. In: Edelstein B,editor. Comprehensive clinical psychology: clinical geropsychology,vol. 7. Oxford: Elsevier; 1998. p. 231–45.
[5] Berr C, Lafont S, Debuire B, Dartigues J-F, Baulieu E-E.Relationships of dehydroepiandrosterone sulfate in the elderly withfunctional, psychological, and mental status, and short-term mortality:a French community-based study. PNAS 1996;93:13410–5.
[6] Liu Y, Schubert D. Steroid hormones block amyloid fibril-induced3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)formazan exocytosis: relationship to neurotoxicity. J Neurochem1998;19:1639–62.
[7] Chae HS, Bach JH, Lee MW, Kim HS, Kim YS, Kim KY, et al.Estrogen attenuates cell death induced by carboxy-terminal fragmentof amyloid precursor protein in PC12 through a receptor-dependentpathway. J Neurosci Res 2001;65:403–7.
[8] Hosoda T, Nakajima H, Honjo H. Estrogens protect neuronalcells from amyloid beta-induced apoptotic cell death. Neuroreport2001;12(9):1965–70.
[9] Zhang L, Rubinow DR, Xaing G, Li BS, Chang YH, Maric D,et al. Estrogens protects against beta-amyloid-induced neurotoxicityin rat hippocampal neurons by activation of Akt. Neuroreport2001;12(9):1919–23.
[10] Greenfield JP, Leung LW, Cai D, Kaasik K, Gross RS, Rodriguez-Boulan E, et al. Estrogen lowers Alzheimer�-amyloid generationby stimulatingtrans-golgi network vesicle biogenesis. J Biol Chem2002;277(14):12136.
[11] Lacreuse A, Wilson ME, Herndon JG. Estradiol, but not raloxifene,improves aspects of spatial working memory in aged ovariectomizedrhesus monkeys. Neurobiol Aging 2002;23:589–600.
[12] Rissman EF, Heck AL, Leonard JE, Shupnik MA, Gustafsson JA.Disruption of receptor beta gene impairs spatial learning in femalemice. Proc Natl Acad Sci USA 2002;99(6):3996–4001.
[13] Zhao L, Chen Q, Diaz Brinton R. Neuroprotective and neuroefficacyof phytoestrogens in cultured hippocampal neurons. Exp Biol Med2002;227(7):509–19.
[14] Lund TD, West TW, Ly T, Bu LH, Simmons DL, Setchell KD, et al.Visual spatial memory is enhanced in female rats (but inhibited inmales) by dietary soy phytoestrogens. BMC Neurosci 2001;2(1):20.
[15] Lephart ED, West TW, Weber KS, Rhees RW, Setchell KD,Adlercreutz H, et al. Neurobehavorial effects of dietary soyphytoestrogens. Neurotoxicol Teratol 2002;24(1):5–16.
[16] Roberts E, Bologa L, Flood JF, Smith GE. Effects ofdehydroepiandrosterone and its sulfate on brain tissue in culture andon memory in mice. Brain Res 1987;406:357–62.
[17] Flood JF, Roberts E. Dehydroepiandrosterone sulfate improvesmemory in aging mice. Brain Res 1988;448:178–81.
[18] Wolf OT, Kirschaum C. Endogenous estradiol and testosterone levelsare associated with cognitive performance in older women and men.Horm Behav 2002;41(3):259–66.
[19] Simpkins JW, Singh M, Bishop J. The potential role for estrogenreplacement therapy in the treatment of the cognitive decline andneurodegeneration associated with Alzheimer’s disease. Neurology1994;15(Suppl 2):S195–7.
[20] Asthana S, Baker LD, Craft S, Stanczyk FZ, Veith RC, RaskindMA, et al. High-dose estradiol improves cognition for women withAD. Neurology 2001;57:605–12.
[21] Cholerton B, Gleason CE, Baker L, Asthana S. Estrogen andAlzheimer’s disease: the story so far. Drugs Aging 2002;19(6):405–27.
[22] Diaz-Brinton R. Cellular and molecular mechanisms of estrogenregulation of memory function and neuroprotection againstAlzheimer’s disease: recent insights and remaining challenges.Learning Memory 2001;8:121–33.
[23] Haskell SG, Richardson ED, Horwitz RI. The effect of estrogenreplacement therapy on cognitive function in women: a critical reviewof the literature. J Clin Epidemiol 1997;50(11):1249–64.
[24] Kesslak JP. Can estrogenplay a significant role in the prevention ofAlzheimer’s disease? J Neural Trans 2002;62:227–39.
[25] Marks SJ, Batra RR, Frishman H. Estrogen replacement therapy forcognitive benefits: viable treatment or forgettable “senior moment”?Heart Dis 2002;4:26–32.
[26] Van Duijn C. Hormone therapy replacement and Alzheimer’s disease.Maturitas 2001;1:201–5.
[27] Shetty K. Hormone replacement therapy. Logically, long termhormone replacement therapy cannot be recommended. Br Med J2002;235:1243.
[28] Voelker R. Questions about hormone replacement therapy. J Am MedAssoc 2002;288:2395–6.
[29] Dixon R, Furutachi T, Lieberman S. The isolation of crystalline22R-hydroxycholesterol and 20-alpha, 22R-hydroxycholesterol frombovine adrenals. Biochem Biophys Res Commun 1970;49:161–5.
[30] Yao ZX, Brown RC, Teper G, Greeson J, Papadopoulos V.22R-Hydroxycholesterol protects neuronal cells from�-amyloid-induced cytotoxicity by binding to�-amyloid peptide. J Neurochem2002;83:1110–9.
[31] Yao Z, Drieu K, Papadopoulos V. The Gingko biloba extract EGb 761rescues PC12 neuronal cells from�-amyloid-induced cell death byinhibiting the formation of�-amyloid-derived diffusible neurotoxicligands. Brain Res 2001;889:181–90.
[32] Yao Z, Papadopoulos V. Function of�-amyloid in cholesteroltransport: a lead to neurotoxicity. FASEB J 2002;16:1677–9.
[33] Watson AA, Fairlie DP, Craik DJ. Solution structure of methionine-oxidized amyloid beta-peptide (1-40). Does oxidation affectconformational switching? Biochemistry 1998;37:12700–6.
[34] Li H, Yao Z, Degenhardt B, Teper G, Papadopoulos V. Cholesterolbinding at the cholesterol recognition/interaction amino acidconsensus (CRAC) of the peripheral-type benzodiazepine receptorand inhibition of steroidogenesis by an HIV TAT-CRAC peptide.Proc Natl Acad Sci USA 2001;98:1267–72.
[35] Morris GM, Goodsell DS, Halliday RS, Huey R, Hart WE, BelewRK, et al. Distributed automated docking of flexible ligands toproteins: parallel applications of AutoDock 2.4.. J Comput Chem1998;19:1639–62.
[36] Hall PF. Cellular organization for steroidogenesis. Int Rev Cytol1984;86:53–95.
[37] Kremer JJ, Pallitto MM, Sklansky DJ, Murphy RM. Correlation of�-amyloid aggregate size and hydrophobicity with decreased bilayerfluidity of model membranes. Biochemistry 2000;39:10309–18.
[38] Kremer JJ, Sklansky DJ, Murphy RM. Profile of changes inlipid bilayer structure caused by�-amyloid peptide. Biochemistry2001;40:8563–71.
[39] Pereira C, Santos MS, Oliveira C. Involvement of oxidative stress onthe impairment of energy metabolism induced by Abeta peptides on

16 L. Lecanu et al. / Steroids 69 (2004) 1–16
PC12 cells: protection by antioxidants. Neurobiol Dis 1999;6(3):209–19.
[40] Eckert GP, Wood WG, Muller WE. Effects of aging and�-amyloidon the properties of brain synaptic and mitochondrial membranes. JNeural Transm 2001;108:1051–64.
[41] Kourie JI. Mechanisms of amyloid beta protein-induced modificationin ion transport systems: implications for neurodegenerative diseases.Cell Mol Neurobiol 2001;21(3):173–213.
[42] Prapong T, Buss J, Hsu WH, Heine P, West Greenlee H, UemuraE. Amyloid beta-peptide decreases neuronal glucose uptake despitecausing increase in GLUT3 mRNA transcription and GLUT3translocation to the plasma membrane. Exp Neurol 2002;174(2):253–8.
[43] Hulstaert F, Blennow K, Ivanoiu A, Schoonderwaldt HC,Riemenschneider M, De Deyn PP, et al. Improved discrimination ofAD patients using�-amyloid(1–42) and tau levels in CSF. Neurology1999;52:1555–62.
[44] Riemenschneider M, Schmolke M, Lautenschlager N, GuderWG, Vanderstichele H, Vanmechelen E, et al. Cerebrospinal�-amyloid(1–42) in early Alzheimer’s disease: association withapolipoprotein E genotype and cognitive decline. Neurosci Lett2000;284:85–8.
[45] Klein WL. A� toxicity in Alzheimer’s disease: globular oligomers(ADDLs) as new vaccine and drug targets. Neurochem Int2002;41:345–52.
[46] Lambert MP, Barlow AK, Chromy BA, Edwards C, Freed R, LiosatosM, et al. Diffusible, nonfibrillar ligands derived from A�1–42 arepotent central nervous system neurotoxins. Proc Natl Acad Sci USA1998;95:6448–53.
[47] Dahlgren KN, Manelli AM, Stine WB, Baker LK, KrafftGA, LaDu MJ. Oligomeric and fibrillar species of amyloid-� peptides differentially affect neuronal viability. J Biol Chem2002;277(35):32046–53.
[48] Juarez-Oropeza MA, Diaz-Zagoya JC, Rabinowitz JL. In vivo andin vitro studies of hypocholesterolemic effects of diosgenin in rats.Int J Biochem 1987;19(8):679–83.
[49] Kato A, Miura T. Hypoglycemic activity ofPolygonati rhizoma innormal and diabetic mice. Biol Pharm Bull 1993;16(11):1118–20.
[50] Kim HS, Wison WK, Needleman DH, Pinkerton FD, Wilson DK,Quiocho FA, et al. Inhibitors of sterol synthesis. Chemical synthesis,structure, and biological activities of (25R)-3�, 26-hydroxy-5�-cholest-8(14)-en-15-one, a metabolite of 3�-hydroxy-5�-cholest-8(14)-en-15-one. J Lipid Res 1989;30(2):247–61.
[51] Kim HS, Oh SH, Kim DI, Kim IC, Cho KH, Park YB. Chemicalsynthesis of 15-ketosterols and their inhibitions of cholesteryl estertransfer protein. Bioorg Med Chem 1995;3(4):367–74.
[52] Puglielli L, Konopka G, Pack-Chung E, MacKenzie Ingano LA,Berezovska O, Hyman BT, et al. Acyl-coenzyme A: cholesterolacyltransferase modulates the generation of the amyloid�-peptide.Nat Cell Biol 2001;3:905–12.
[53] Birks JS, Melzer D. Dopenezil for mild and moderate Alzheimer’sdisease. Cochrane Database Syst Rev 2000;2:CD001190.
[54] Grutzendler J, Moris JC. Cholinesterase inhibitors for Alzheimer’sdisease. Drugs 2001;61:41–52.
[55] Tsukatani H, Takauchi K, Yoshida J, Yamata S, Tokumura A,Hamaguchi C. Isolation of diosgenin [(25R)-spirost-5-en-3beta-ol]from a lysate of a new hypotensive phospholipid occurring in bovinebrain. J Neurochem 1985;44(20):658–61.
Related Documents











