Published Ahead of Print 18 March 2013. 2013, 81(5):1852. DOI: 10.1128/IAI.00199-13. Infect. Immun. Palmer M. Noh, Kelly A. Brayton, Joseph T. Agnes and Guy H. David R. Herndon, Massaro W. Ueti, Kathryn E. Reif, Susan following Tick-Borne Transmission subsp. centrale and Its Restriction Heterogeneity in Anaplasma marginale Identification of Multilocus Genetic http://iai.asm.org/content/81/5/1852 Updated information and services can be found at: These include: REFERENCES http://iai.asm.org/content/81/5/1852#ref-list-1 at: This article cites 23 articles, 13 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on June 10, 2014 by guest http://iai.asm.org/ Downloaded from on June 10, 2014 by guest http://iai.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Published Ahead of Print 18 March 2013. 2013, 81(5):1852. DOI: 10.1128/IAI.00199-13. Infect. Immun.
PalmerM. Noh, Kelly A. Brayton, Joseph T. Agnes and Guy H. David R. Herndon, Massaro W. Ueti, Kathryn E. Reif, Susan following Tick-Borne Transmissionsubsp. centrale and Its RestrictionHeterogeneity in Anaplasma marginale Identification of Multilocus Genetic
http://iai.asm.org/content/81/5/1852Updated information and services can be found at:
These include:
REFERENCEShttp://iai.asm.org/content/81/5/1852#ref-list-1at:
This article cites 23 articles, 13 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
on June 10, 2014 by guest
http://iai.asm.org/
Dow
nloaded from

Identification of Multilocus Genetic Heterogeneity in Anaplasmamarginale subsp. centrale and Its Restriction following Tick-BorneTransmission
David R. Herndon,a Massaro W. Ueti,a Kathryn E. Reif,b Susan M. Noh,a,b Kelly A. Brayton,b Joseph T. Agnes,b Guy H. Palmerb
Animal Diseases Research Unit, Agricultural Research Service, U.S. Department of Agriculture, Pullman, Washington, USAa; Department of Veterinary Microbiology andPathology and Paul G. Allen School for Global Animal Health, Washington State University, Pullman, Washington, USAb
Anaplasma marginale subsp. centrale was the first vaccine used to protect against a rickettsial disease and is still in widespreaduse a century later. As its use preceded development of either cryopreservation or cell culture, the vaccine strain was maintainedfor decades by sequential passage among donor animals, excluding the natural tick-borne transmission cycle that provides a se-lective pressure or population “bottleneck.” We demonstrated that the vaccine strain is genetically heterogeneous at 46 chromo-somal loci and that heterogeneity was maintained upon inoculation into recipient animals. The number of variants per siteranged from 2 to 11 with a mean of 2.8/locus and a mode and median of 2/locus; variants included single-nucleotide polymor-phisms, insertions/deletions, polynucleotide tracts, and different numbers of perfect repeats. The genetic heterogeneity is highlyunlikely to be a result of strain contamination based on analysis using a panel of eight gene markers with a high power for straindiscrimination. In contrast, heterogeneity appears to be a result of genetic drift in the absence of the restriction of tick passage.Heterogeneity could be reduced following tick passage, and the reduced heterogeneity could be maintained in sequential intrave-nous and tick-borne passages. The reduction in vaccine strain heterogeneity following tick passage did not confer an enhancedtransmission phenotype, indicating that a stochastically determined population bottleneck was likely responsible as opposed toa positive selective pressure. These findings demonstrate the plasticity of an otherwise highly constrained genome and highlightthe role of natural transmission cycles in shaping and maintaining the bacterial genome.
“Heifer 906, about 18 months old, arrived in Pretoria on the28th October, 1909, and was immediately placed in a clean
stable and kept free from ticks. The temperature was taken twicedaily and a detailed record kept; no deviations from normal wereobserved. On the 7th December, 1909 —thirty-eight days afterarrival—she was tapped and 20 c.c. defibrinated blood were in-jected into calf 881 (1).” These precise notes by Arnold Theiler(later Sir Arnold Theiler) describe the isolation and first expansionof Anaplasma marginale subsp. centrale (originally described byTheiler as A. marginale “variety” centrale; it has been renamed A.marginale subsp. centrale in keeping with current taxonomic con-vention). As shown in a series of experiments conducted early in1910, A. marginale subsp. centrale was both attenuated relative toA. marginale sensu stricto and capable of inducing protectionagainst virulent A. marginale (1). These results led directly to theuse of this organism as a live vaccine to prevent severe morbidityand mortality due to tick-borne anaplasmosis, representing thefirst rickettsial vaccine (1, 2). Initially deployed in South Africa,use of this live vaccine spread during the 20th century to tropicaland subtropical countries throughout Africa and in Asia, Austra-lia, and Latin America and remains in use today (3). In Australiaalone, nearly a million doses of vaccine have historically been usedannually (3).
While this extensive record is by itself rare among live bacterialvaccines, the propagation history of the vaccine provides a uniqueopportunity to examine genetic drift over time. Following its ini-tial isolation described above, the vaccine strain was maintainedby sequential passage among cattle until the 1970s when cryo-preservation was developed and then implemented (3, 4). Impor-tantly, passage over this approximately 70-year period was only byintravenous inoculation, eliminating any selective pressure or
population “bottleneck” that may occur during the natural trans-mission cycle in which the bacteria colonize and replicate in themidguts and salivary glands of ixodid ticks prior to transmission(5). As positive selection has been identified to occur early in col-onization of the tick midgut by sensu stricto A. marginale strains(6, 7), we hypothesized that long-term propagation without tickpassage has resulted in genetic heterogeneity in the vaccine strain.If this is correct, we would further predict that passage through anatural tick vector can restrict this heterogeneity. In the presentwork, we test these hypotheses and report the results in the contextof bacterial genetic drift.
MATERIALS AND METHODSIdentification of variable loci within the Anaplasma marginale subsp.centrale vaccine strain. The 1.2-Mb vaccine strain was sequenced usingthe whole-genome shotgun method with approximately 6.5� coverage(8). During the closure phase of the project, multiple sequence gaps werenoted to be refractory to closure: sequencing across adjacent contigs re-sulted in observed single-nucleotide polymorphisms (SNPs) and nucleo-tide insertions/deletions (indels). In order to exclude the possibility thatthe variations found were the result of chimeric cloning artifacts and/orareas of misassembled sequence, locus-encompassing subclones were se-
Received 12 February 2013 Returned for modification 25 February 2013Accepted 7 March 2013
Published ahead of print 18 March 2013
Editor: R. P. Morrison
Address correspondence to Guy H. Palmer, [email protected].
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/IAI.00199-13
1852 iai.asm.org Infection and Immunity p. 1852–1858 May 2013 Volume 81 Number 5
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

quenced to completion by primer walking, and their inserts were sized byrestriction digestion (8). Individual subclone sizes were then compared totheir predicted sizes following alignment of their complete sequence to theadjacent areas of nonvariant sequence. In addition, a BLAST searchabledatabase was constructed for all of the initial assembly contigs and non-incorporated singleton sequences. For each described locus, subclone se-quences were searched for their respective locations, with particular at-tention paid to subclone ends. The A. marginale subsp. centrale genomesequence available from GenBank (accession number CP001759) and re-ported previously (8) reflects the most frequently observed subclone orPCR product sequence in the observed variable loci.
Screening for the presence of multiple strains. Individual A. mar-ginale strains can be distinguished by their complement of outer mem-brane proteins (Omps), shown to be capable of inducing protective im-munity (9–11). A panel of eight omp genes as strain markers wasdeveloped based on either completed genome sequences (St. Maries andFlorida strains) (12, 13) or by searching the whole-genome shotgun contigdatabase at NCBI and constructing the gene sequences from the availablecontigs (Oklahoma, South Idaho, and Washington-Okanogan strains)(13). As the omp10 sequences were incomplete in the available contigs forthe Oklahoma, South Idaho, and Washington-Okanogan strains, a com-bination of PCR, cloning, and sequencing using primers in the knownflanking regions was used to complete the sequences. Pairwise Clustalalignment was used to enumerate sequence differences among the Omp-encoding genes.
Locus variation within the bacterial population following live-strain vaccination and tick transmission of the vaccine strain. Popula-tion variation was measured at four loci identified as being variable in theoriginal sequencing of the vaccine strain by sequencing fosmid librariesand cloned PCR products derived from either (i) calves vaccinated withthe live strain, (ii) infected ticks that fed on vaccinates, or (iii) calvesinfected with the vaccine strain following transmission. The animals andpassages are shown in Fig. 1. Seronegative (14) calves were intravenouslyinoculated with 108 organisms of the A. marginale subsp. centrale vaccinestrain (animals V1 to V3). Blood samples were collected during acutebacteremia for DNA isolation. During acute bacteremia, adult male Der-macentor andersoni ticks were allowed to attach and feed for 7 days (15).Following incubation to allow digestion of the blood meal, cohorts of 500ticks were allowed to transmission feed on uninfected calves; the ticks thatinitially acquired infection by feeding on vaccinate V1 were allowed to feedon calf TT1, ticks that fed on V2 were allowed to feed on TT2, and ticks thatfed on V3 were allowed to feed on TT3. DNA was isolated from individualtick midguts and salivary glands following transmission feeding, from ticksaliva samples induced following transmission feeding, and from bloodsamples from calves TT1 to TT3 following successful tick transmission ofthe vaccine strain (16). Fosmid libraries of approximately 40 kb wereconstructed with the CopyControl fosmid library production kit (Epicen-tre Biotechnologies). The resulting clones were selected, transferred to96-well plates, and arrayed on Hybond N� membranes using an auto-mated colony picker (Genetix QPixII). Digoxigenin-labeled probes wereused to screen for the loci of interest, and DNA was isolated from positiveclones with the PerfectPrep bacterial artificial chromosome (BAC) kit (5Prime). PCR and cloning were performed with Pfx Supermix and the ZeroBlunt cloning kit (Life Technologies), and all sequencing was carried outon an ABI Prism 3130xl genetic analyzer. Sequence analysis was per-formed using a combination of Sequencher (GeneCodes) and Vector NTI(Life Technologies).
Locus variation and transmission phenotype of the vaccine strainfollowing selection by tick passage. Following the observation that locusvariation was reduced following successful tick transmission of the livevaccine strain to animal TT2, this “tick passage-selected” bacterial popu-lation was transmitted by intravenous inoculation to an uninfected calf(TP1). Using the same procedures described above for the live vaccinestrain in animals V1 to V3, male D. andersoni ticks were allowed to acqui-sition feed on TP1 and then subsequently transmission fed on an unin-
fected calf (TP2) using 500 ticks (Fig. 1). DNA was isolated from theindividual tick midguts and salivary glands following transmission feed-ing, from tick saliva induced following transmission feeding, and fromblood samples from both TP1 and TP2 during acute bacteremia. For locusvariation, the same sequencing procedures were used as described abovefor V1 to V3 and TT1 to TT3. For phenotypic variation, the tick infectionrate (number of fed ticks that acquired infection) and the bacterial loadper midgut, per salivary gland pair, or per microliter of saliva were deter-mined using quantitative real-time msp5 PCR, as previously described indetail (16). In addition, a second cohort of ticks acquisition fed on TP1 wasused to test whether the threshold for transmission of the vaccine strainhad been decreased by feeding 100 ticks on an uninfected recipient calf(TP3). For comparison to the initial, unselected vaccine strain, ticks fromthe same cohort were acquisition fed on V3 and then transmission to anuninfected recipient calf (TT4) was attempted. Following attemptedtransmission, both calves, TP3 and TT4, were screened daily for evidenceof infection by microscopic examination of Giemsa stained blood filmsand by nested msp5 PCR for 90 days (17).
RESULTSIdentification of variable loci within the Anaplasma marginalesubsp. centrale vaccine strain. Based on whole-genome shotgunsequencing, 46 variable loci were identified in the vaccine straingenome (Table 1). The possibility that these variable loci were theresult of chimeric cloning artifacts was excluded by determiningthe actual size of subclones that encompassed the complete locusby restriction enzyme digestion, followed by comparison to thepredicted in silico size after joining the complete subclone se-quence to the adjacent nonvariant sequences. Similarly, the pos-
FIG 1 Infection and transmission of the Anaplasma marginale subsp. centralevaccine strain. The experiments shown in panels A, B, C, and D represent fourindependent transmission experiments. Animals V1 to V3 were inoculatedintravenously with the vaccine strain and then used as the source for acquisi-tion feeding of ticks for transmission to TT1 to TT4. TP1 was inoculated intra-venously with the tick-passaged bacteria from TT2 and then used as the sourcefor acquisition feeding of ticks for transmission to TP2 and TP3. Cohorts of 500ticks were used for transmission to TT1-TT3 and TP2; cohorts of 100 ticks wereused for transmission to TT4 and TP3. Gray shading indicates animals infectedby tick transmission.
Genetic Heterogeneity in Anaplasma
May 2013 Volume 81 Number 5 iai.asm.org 1853
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
sibility of misassembly was minimized utilizing the BLAST data-base of contigs and singletons. Subclones that satisfied the size-matching criterion described above were included in the analysisonly if they were found to be anchored to a unique region within
the genome assembly at one or both termini. The number of vari-ants per site ranged from 2 to 11 with a mean of 2.8 per locus anda mode and median of 2 per locus. The majority of the changesbetween variants at a given locus were SNPs (25/46), seven of thesealso were accompanied by an indel, and one by variation in apolynucleotide tract. There was one locus in which variation wascharacterized by differences in the number of perfect repeats andseven in which the differences were only in the length of polynu-cleotide tracts. Locus variation was present in both intergenic (13/46) and coding regions (33/46). Of the SNP and indel variantswithin coding regions, none were predicted to result in frameshiftsor premature truncation of encoded proteins.
Screening for multiple strains. Genetic heterogeneity couldreflect contamination with an additional strain or strains either inthe initial 1909 isolation, in which the strain was isolated directlyfrom blood from an infected animal, or subsequently during thesequential in vivo passages of the vaccine strain that were requiredto maintain the vaccine stock prior to development of cryopreser-vation. To screen for multiple strains, a panel of eight Omp-en-coding genes was developed and tested using five A. marginalesensu stricto strains with subsequent comparison to the A. mar-ginale subsp. centrale vaccine strain (Table 2). Most sensu strictostrains could be distinguished using any individual omp nucleo-tide sequence; all strains could be distinguished by sequence dif-ferences when multiple genes were examined. The comparisonbetween the Florida and Washington-Okanogan strains illustratesthe discriminating power of the panel. While three omp genes,omp11, omp12, and omp13, are identical between the two strains,there are marked differences in the other five omp genes and apanel-wide cumulative total of 311 nucleotide differences betweenthe two (Table 2). Comparison of the vaccine strain to the sensustricto strains revealed even more dramatic differences at eachomp gene: the vaccine strain could be clearly distinguished usingany individual omp sequence (Table 2). The vaccine strain was asignificant outlier compared to the sensu stricto strains; there weresignificantly more sequence differences between the vaccine strainand sensu stricto strains than among the sensu stricto strains (Stu-dent’s t test with correction for multiple comparisons). This heldboth for any of the individual omp genes (P values ranged fromP � 1 � 10�7 for omp7 to P � 3 � 10�19 for omp12) or the fullpanel (P � 2 � 10�7). There was no observed sequence heteroge-neity at any of the eight loci within the vaccine strain, consistentwith the lack of any contamination with additional strains.
Locus variation within the bacterial population followinglive-strain vaccination and tick transmission of the vaccinestrain. Locus 56 was selected for initial tracking of heterogeneityfollowing live vaccination of individual calves and subsequent ticktransmission of the vaccine strain due to the magnitude of differ-ence (261 bp, reflecting either one iteration of the sequence or asecond perfect repeat within the locus) between the two variants atthis site. The presence of both variants could be detected in allthree live-strain vaccinates, V1 to V3 (Fig. 2A), consistent with itsdetection in the sequenced genome and maintenance by intrave-nous passage. Interestingly, following tick transmission, three dif-ferent patterns of locus 56 variant predominance were observedduring acute bacteremia: the populations in TT1 and TT2 wereskewed toward the two-repeat variant and one-repeat variant, re-spectively, while the population in TT3 was clearly represented byboth variants (Fig. 2B). To test whether this reflected the presenceof only a single variant in the transmitting tick population, salivary
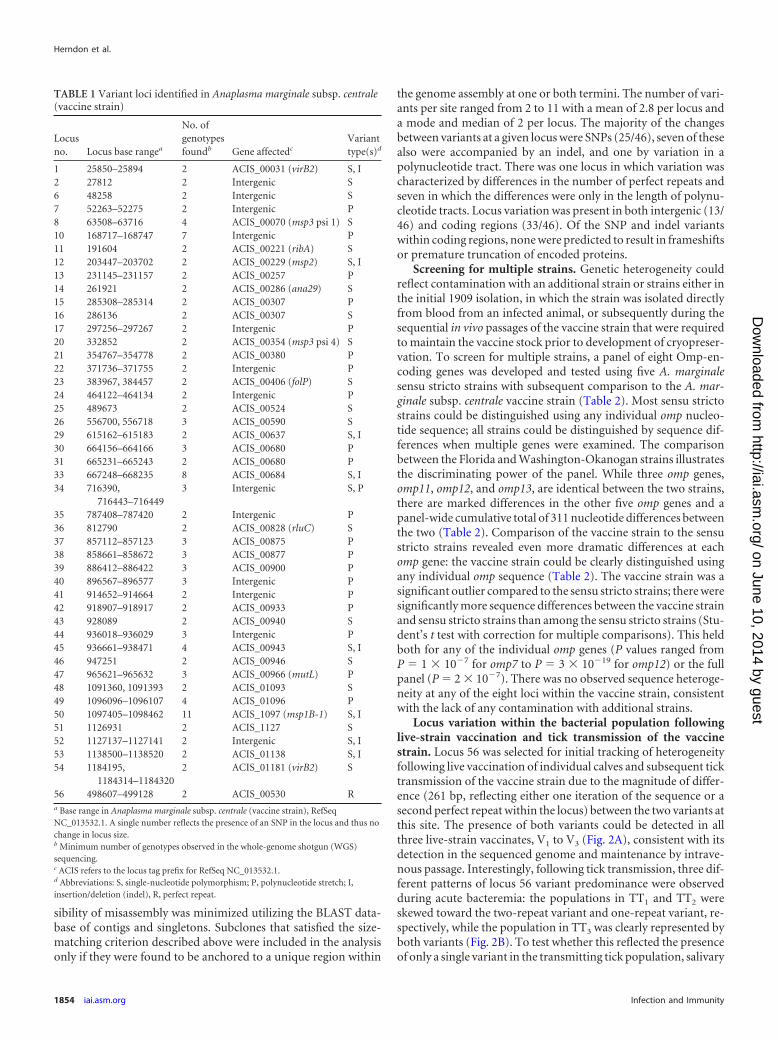
TABLE 1 Variant loci identified in Anaplasma marginale subsp. centrale(vaccine strain)
Locusno. Locus base rangea
No. ofgenotypesfoundb Gene affectedc
Varianttype(s)d
1 25850–25894 2 ACIS_00031 (virB2) S, I2 27812 2 Intergenic S6 48258 2 Intergenic S7 52263–52275 2 Intergenic P8 63508–63716 4 ACIS_00070 (msp3 psi 1) S10 168717–168747 7 Intergenic P11 191604 2 ACIS_00221 (ribA) S12 203447–203702 2 ACIS_00229 (msp2) S, I13 231145–231157 2 ACIS_00257 P14 261921 2 ACIS_00286 (ana29) S15 285308–285314 2 ACIS_00307 P16 286136 2 ACIS_00307 S17 297256–297267 2 Intergenic P20 332852 2 ACIS_00354 (msp3 psi 4) S21 354767–354778 2 ACIS_00380 P22 371736–371755 2 Intergenic P23 383967, 384457 2 ACIS_00406 (folP) S24 464122–464134 2 Intergenic P25 489673 2 ACIS_00524 S26 556700, 556718 3 ACIS_00590 S29 615162–615183 2 ACIS_00637 S, I30 664156–664166 3 ACIS_00680 P31 665231–665243 2 ACIS_00680 P33 667248–668235 8 ACIS_00684 S, I34 716390,
716443–7164493 Intergenic S, P
35 787408–787420 2 Intergenic P36 812790 2 ACIS_00828 (rluC) S37 857112–857123 3 ACIS_00875 P38 858661–858672 3 ACIS_00877 P39 886412–886422 3 ACIS_00900 P40 896567–896577 3 Intergenic P41 914652–914664 2 Intergenic P42 918907–918917 2 ACIS_00933 P43 928089 2 ACIS_00940 S44 936018–936029 3 Intergenic P45 936661–938471 4 ACIS_00943 S, I46 947251 2 ACIS_00946 S47 965621–965632 3 ACIS_00966 (mutL) P48 1091360, 1091393 2 ACIS_01093 S49 1096096–1096107 4 ACIS_01096 P50 1097405–1098462 11 ACIS_1097 (msp1B-1) S, I51 1126931 2 ACIS_1127 S52 1127137–1127141 2 Intergenic S, I53 1138500–1138520 2 ACIS_01138 S, I54 1184195,
1184314–11843202 ACIS_01181 (virB2) S
56 498607–499128 2 ACIS_00530 Ra Base range in Anaplasma marginale subsp. centrale (vaccine strain), RefSeqNC_013532.1. A single number reflects the presence of an SNP in the locus and thus nochange in locus size.b Minimum number of genotypes observed in the whole-genome shotgun (WGS)sequencing.c ACIS refers to the locus tag prefix for RefSeq NC_013532.1.d Abbreviations: S, single-nucleotide polymorphism; P, polynucleotide stretch; I,insertion/deletion (indel), R, perfect repeat.
Herndon et al.
1854 iai.asm.org Infection and Immunity
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
glands from individual ticks that transmission fed on TT1, TT2, orTT3 were screened. Both locus 56 variants could be identified inindividual ticks (Fig. 2C), regardless of whether both variants wereequally represented following transmission, e.g., both variantswere identified in individual ticks that transmitted to TT1 (thebacteremia in TT1 had a population skew toward the two-repeatvariant at locus 56), TT2 (the bacteremia in TT2 which had a pop-ulation skew toward the one-repeat variant) and those that trans-mitted to TT3 (which had no apparent skew in the bacteremia).Testing the cohorts of ticks revealed that there was no significantdifference (P � 0.59 by Fisher’s exact test) in the proportion ofeach locus 56 variant type between the ticks that transmitted toTT1 and those that transmitted to TT2.
Locus variation following selection by tick passage. On thebasis of the observation that locus variation can be reduced fol-lowing successful tick transmission of the live vaccine strain asevidenced by animal TT2, we tested whether reduced locus 56variation was maintained with subsequent passage. The “tick pas-sage-selected” bacterial population was transmitted by intrave-nous inoculation of blood from TT2 into a seronegative, unin-fected calf (TP1). Ticks were subsequently allowed to acquisitionfeed on TP1 and then transmission feed on TP2. The locus 56variants were then examined in TP1 during acute bacteremia, the
TABLE 2 Pairwise nucleotide comparison among five Anaplasma marginale sensu stricto strains and the vaccine strain
Strain comparison
No. of nucleotide differencesa
Total no. ofnucleotidedifferencesb
omp5(AM_1139)
omp7(AM_1220)
omp9(AM_1222)
omp10(AM_1223)
omp11(AM_1255)
omp12(AM_1257)
omp13(AM_1258)
omp14(AM_075)
St. Maries vs Florida 74 160 12 17 22 6 8 18 317St. Maries vs
Oklahoma54 176 15 15 16 7 3 20 306
St. Maries vs SouthIdaho
4 61 62 4 27 11 14 25 208
St. Maries vsWashington-Okanogan
0 1 0 21 5 15 22 43 107
Florida vs Oklahoma 26 128 5 4 36 3 11 4 217Florida vs South Idaho 70 127 62 13 5 15 22 39 353Florida vs Washington
-Okanogan74 127 72 30 0 0 0 8 311
Oklahoma vs SouthIdaho
50 165 59 15 31 14 15 39 388
Oklahoma vsWashington-Okanogan
54 165 69 32 36 3 11 10 380
South Idaho vsWashington-Okanogan
4 1 0 21 5 15 22 43 111
Vaccine strain vs St.Maries
247 337 391 381 317 197 286 296 2,452
Vaccine strain vs SouthIdaho
246 344 388 381 315 200 277 316 2,467
Vaccine strain vsWashington-Okanogan
247 343 388 369 315 194 269 290 2,415
Vaccine strain vsOklahoma
231 363 390 390 316 193 285 297 2,465
Vaccine strain vsFlorida
232 370 390 39 313 194 280 295 2,113
a The gene identified and locus tag (AM number) refer to the St. Maries strain reference genome (NC004842).b The total numbers of nucleotide differences between the two strains being compared are shown in boldface type.
FIG 2 Genetic heterogeneity in locus 56 in vaccinates and following tick trans-mission. The locus 56 variants differed by 261 bp, representing either oneiteration of this sequence or two iterations, a perfect repeat. (A to C) Presenceof the one-repeat variant (total amplicon size of 819 bp) and the two-repeatvariant (amplicon size of 1,080 bp) of locus 56 in animals inoculated with thelive vaccine strain (A), in animals following tick transmission (B), and in thesalivary glands of transmitting ticks (C). Each lane in panel C shows ampliconsderived from the salivary glands of an individual tick; results from two repre-sentative individual ticks are shown for each of the three transmissionsV1¡TT1, V2¡TT2, and V3¡TT3.
Genetic Heterogeneity in Anaplasma
May 2013 Volume 81 Number 5 iai.asm.org 1855
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

salivary glands of ticks that transmitted to TP2, and the acute bac-teremia in TP2. The reduced heterogeneity originally observed inTT2 was maintained in TP1, in the salivary glands of individualtransmission-fed ticks, and in TP2 (Fig. 3A). Direct comparisonbetween the locus variant populations in TP1 and in V3 followinginitial infection with the live vaccine strain confirmed that the lossof heterogeneity was statistically significant (Fig. 3B) (P � 0.0001by chi-square test). We then examined whether loss of heteroge-neity occurred at independent loci elsewhere in the genome. Vari-ants in loci 45, 51, and 52 have both SNPs and indels; locus 52 isintergenic, the other two are within coding regions. Comparisonof variants in all three loci revealed a progressive and significantloss of heterogeneity following tick transmission in TT2, TP1, andTP2 compared to the initial vaccine strain, represented by both V2
and V3 (Fig. 4).Transmission phenotype following selection by tick passage.
To test whether passage by tick transmission and the reduced lo-cus heterogeneity reflected phenotypic selection for enhanced col-onization and replication within the tick vector, we compared thetick infection rate and bacterial load in the midguts, salivaryglands, and saliva of adult male D. andersoni that had acquisitionfed on either V3 or TP1 and were then transmission fed. Therewere no statistically significant differences in the infection ratesfor midguts, salivary glands, and saliva; there were also no signif-
icant differences in the bacterial loads in the midguts, salivaryglands, and saliva (Table 3). Consistent with this lack of differencein colonization and replication, there was no marked reduction inthe number of ticks required to successfully transmit the vaccinestrain following the reduction in locus heterogeneity by tick pas-sage: 500 adult D. andersoni that were acquisition fed on either V3
or TP1 successfully transmitted the bacteria, while 100 ticks acqui-sition fed at the same time failed to transmit regardless of whetherticks were acquisition fed on V3 or TP1. These results are consis-tent with prior data indicating that the live vaccine strain could besuccessfully transmitted only with �250 ticks (18).
DISCUSSION
We accept the hypothesis that the A. marginale subsp. centralevaccine strain is genetically heterogeneous, characterized by thepresence of variant sequences at 46 loci distributed throughout thechromosome. That these locus variants represent true genotypicdiversity within the vaccine strain, rather than artifact, is sup-ported by the following five lines of evidence. (i) The diversity wasinitially detected by sequencing a library of mechanically gener-ated fragments, and the size of an individual variant could bedefinitively mapped to the genome. (ii) The identical divergentsequence was detected in multiple independent clones. (iii) Geno-typic variation was consistent with retention of function—none ofthe SNP or indel changes resulted in premature truncation orframeshifts. (iv) For the four loci examined in depth, diversity wasobserved in at least four infected animals. (v) Using locus 56 as amarker, diversity could be detected at each step of the bacterial lifecycle, in blood during acquisition feeding, in colonized ticks, andin the acute bacteremia following transmission.
The genotypic diversity is highly unlikely to result from eitheroriginal isolation of two or more strains or from subsequent con-tamination with a different strain. Using the panel developed forthe current study, A. marginale sensu stricto strains isolated withinthe same geographical region (e.g., the St. Maries, Washington-Okanogan, and South Idaho strains) could be unambiguously dis-criminated. The omp sequences of the vaccine strain are even moredivergent (Table 2), suggesting that contamination with a sensustricto strain would be readily detectable. The caveat to this con-clusion is that very little is known of the diversity among A. mar-ginale strains in southern Africa, where the original isolation oc-curred. If the interstrain diversity, including that between A.marginale subsp. centrale and A. marginale sensu stricto, was re-markably lower in southern Africa than in the Americas, there is apossibility that the presence of a second strain could be missed bythe multi-omp panel approach used here.
In contrast, the presence of differing numbers of variants at
FIG 4 Maintenance of reduced genetic heterogeneity in independent loci. The proportions of the most predominant variants, designated A and B, at locus 45and at loci 51 and 52 are shown. Loci 51 and 52 are separated by less than 300 bp and thus were analyzed together.
FIG 3 Maintenance of reduced genetic heterogeneity in locus 56. (A) Thepredominance of the one-repeat variant observed in TT2 was maintained fol-lowing inoculation into animal TP1, in the salivary glands (SG) of ticks acqui-sition fed on TP1 and transmission fed on TP2, and during the acute bactere-mia of TP2. Results from two representative individual ticks are shown. (B)Quantitation of locus 56 variants in the bacteremia following initial vaccina-tion (represented by V3) versus the reduction in heterogeneity following tickpassage and maintained in TP1. 1 and 2, The proportions of variants with oneversus two repeats are indicated in white and black, respectively.
Herndon et al.
1856 iai.asm.org Infection and Immunity
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

individual loci, ranging from 2 to 11, suggests that strain contam-ination is highly unlikely versus genetic drift over time at specificloci sensitive to variation, including the detected variation in poly-nucleotide tracts. This heterogeneity would appear most likely tohave been generated over the long period when the vaccine strainwas maintained by serial animal passage, prior to development ofcryopreservation (4). These mutations appear to have a minimalimpact on microbial fitness in terms of bacteremia: there was noapparent difference in the bacteremia levels in animals TT2, TP1,and TP2 with reduced heterogeneity compared to the levels in V1,V2, and V3 in which there was clear population heterogeneity.Thus, in the absence of any additional selective pressure, the mu-tations could be maintained within the vaccine strain.
We accept the hypothesis that tick passage can restrict the het-erogeneity observed in the vaccine strain based on the multilocusrestriction observed in TT2 and then maintained in TP1 and TP2.This reduction in heterogeneity does not appear to reflect a strongselective pressure for colonization, replication, or transmissionfitness in the tick. There were no significant differences in infec-tion rate or bacterial numbers in any tick tissue between ticks thathad acquisition fed on V3, which had been vaccinated with theheterogeneous vaccine strain, and those that had fed on TP1, inwhich heterogeneity was significantly reduced. Similarly, therewas no evident reduction in the threshold for tick transmission ofthe vaccine strain: 500 ticks successfully transmitted from both V3
and TP1, while 100 ticks fed at the same time failed to transmit.While these transmission studies would not have detected a minorreduction in the threshold for transmission, they are consistentwith both the quantitative data showing no difference in tick in-fection rates, colonization, and replication as well as prior dataindicating that �250 ticks were required for transmission of thevaccine strain (18).
The observed restriction in heterogeneity appears to be sto-chastic rather than strong positive selection. Although transmis-sion to TT2 resulted in restriction of heterogeneity, this was nei-ther consistent (e.g., there was no loss of heterogeneity in TT3) norlinked to a clear restriction in the salivary glands of the transmit-ting ticks (e.g., both variants were represented in the population,and individual ticks could carry more than one locus variant). Aparsimonious explanation for this observation is that very fewticks, and perhaps a single tick, is responsible for transmission. Ifthe salivary gland from the tick contains the locus 56 variant withone 261-bp repeat, the pattern exemplified by TT2 would occur.Similarly, the pattern shown by TT1 or TT3 would occur when asingle tick was colonized with either the two-repeat locus 56 vari-
ant or both variants, respectively. Consistent with this explanationwas the identification of individual ticks colonized with the one-repeat locus 56 variant, the two-repeat variant, or both variants. Asingle tick colonized with a highly infectious A. marginale sensustricto strain has been shown to transmit infection (19). The re-quirement for numerous ticks to transmit the attenuated vaccinestrain may be related more to the effects of tick feeding to dampenthe host inflammatory response and lower the threshold for trans-mission than to the number of ticks that actually transmit infec-tion (20–22). In contrast to intravenous inoculation of the vaccinestrain, when �108 bacteria are introduced (3, 23), an individualtick secretes a small inoculum, roughly 100 bacteria per �l ofsaliva, thus dramatically reducing the effective population sizewith a corresponding stochastically determined loss in heteroge-neity. This has been reported in Trypanosoma brucei developmentwithin the tsetse fly in which heterogeneity is dramatically re-duced, as the first mutant to invade the salivary gland then pre-dominates (24). The bottleneck for A. marginale subsp. centrale inthe tick appears more likely to occur at transmission, either by theneed for only a few ticks to transmit or a bottleneck at the point ofsalivary secretion. The natural cycle of A. marginale transmission,tick acquisition by feeding on a bacteremic mammal, followed bytransmission to initiate a new mammalian infection, would beexpected to dramatically reduce or eliminate heterogeneity withinseveral cycles.
In summary, we have identified remarkable multilocus geneticheterogeneity in a widely used live bacterial vaccine. These find-ings demonstrate the plasticity of an otherwise highly constrainedgenome (8, 13) and highlight the role of natural transmission cy-cles in shaping and maintaining the bacterial genome.
ACKNOWLEDGMENTS
This work was supported by NIH AI044005, Wellcome TrustGR075800M, BARD US4187-09C, and USDA ARS 5348-32000-033-00D/-01S. J. T. Agnes was supported in part by a NIH Fellowship inProtein Biotechnology (GM008336), and K. E. Reif was supported by aNIH Fellowship in Infectious Diseases (AI07025).
The technical assistance of James Allison, Ralph Horn, and BeverlyHunter is appreciated.
REFERENCES1. Theiler A. 1911. Further investigations into anaplasmosis of South Afri-
can cattle. First report of the Director of Veterinary Research. Union ofSouth Africa, Johannesburg, South Africa.
2. Theiler A. 1912. Gallsickness of imported cattle and the protective inoc-ulation against this disease. Agric. J. Union South Africa 3:7– 46.
TABLE 3 Infection within Dermacentor andersoni ticks prior to and following tick passage
Characteristic
Value for ticksa
Calf V3 given vaccine strain Calf TP1 given tick passage strain
TT3 TT4 TP2 TP3
Midgut infection rate, % (no. ticks infected/total no. of ticks) 90 (45/50) 90 (45/50) 76 (38/50) 78 (39/50)Bacteria per midgut, mean no. (�SEM) 104.3 (�100.82) 104.3 (�100.69) 103.9 (�100.68) 104.2 (�100.8)Salivary gland infection rate, % (no. ticks infected/total no. of ticks) 80 (40/50) 82 (41/50) 78 (39/50) 84 (42/50)Bacteria per salivary gland, mean no. (�SEM) 105.9 (�100.9) 105.8 (�100.8) 105.8 (�100.9) 105.7 (�100.8)Saliva containing Anaplasma, % (no. ticks infected/total no. of ticks) 18 (9/50) 20 (10/50) 8 (4/50) 20 (10/50)Bacteria per �l of saliva, mean no. (�SEM) 102.7 (�100.7) 102.3 (�100.5) 102.7 (�100.4) 101.8 (�100.4)a The values for ticks from calves V3 and TP1 exposed to a vaccine strain or tick passage strain. Calves V3 and TP1 were acquisition-fed calves. Calves TT3, TT4, TP2, and TP3 weretransmission-fed calves. Calves TT3 and TP2 were exposed to 500 ticks, while calves TT4 and TP3 were exposed to 100 ticks.
Genetic Heterogeneity in Anaplasma
May 2013 Volume 81 Number 5 iai.asm.org 1857
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

3. Bock R, De Vos A. 2001. Immunity following use of Australian tick fevervaccine: a review of the evidence. Aust. Vet. J. 79:832– 839.
4. Barnett SF. 1964. The preservation of Babesia bigemina, Anaplasma cen-trale, and Anaplasma marginale by deep freezing. Vet. Rec. 76:4 – 8.
5. Kocan KM, de la Fuente J, Gugliemone AA, Melendez RD. 2003.Antigens and alternatives for control of Anaplasma marginale in cattle.Clin. Microbiol. Rev. 16:698 –712.
6. Löhr CV, Rurangirwa FR, McElwain TF, Stiller D, Palmer GH. 2002.Specific expression of Anaplasma marginale major surface protein 2 sali-vary gland variants occurs in the midgut and is an early event during ticktransmission. Infect. Immun. 70:114 –120.
7. Rurangirwa FR, Stiller DS, French DM, Palmer GH. 1999. Restriction ofmajor surface protein 2 (MSP2) variants during tick transmission of theehrlichiae Anaplasma marginale. Proc. Natl. Acad. Sci. U. S. A. 96:3171–3176.
8. Herndon DR, Palmer GH, Shkap V, Knowles DP, Brayton KA. 2010.Complete genome sequence of Anaplasma marginale subsp. centrale. J.Bacteriol. 192:379 –380.
9. Noh SM, Brayton KA, Knowles DP, Agnes JT, Dark MJ, Brown WC,Baszler TV, Palmer GH. 2006. Differential expression and sequenceconservation of the Anaplasma marginale msp2 gene superfamily outermembrane proteins. Infect. Immun. 74:3471–3479.
10. Noh SM, Brayton KA, Brown WC, Norimine J, Munske GR, Davitt CM,Palmer GH. 2008. Composition of the surface proteome of Anaplasmamarginale and its role in protective immunity induced by outer membraneimmunization. Infect. Immun. 76:2219 –2226.
11. Brown WC, Shkap V, Zhu D, McGuire TC, Tuo W, McElwain TF,Palmer GH. 1998. CD4� T-lymphocyte and immunoglobulin G2 re-sponses in calves immunized with Anaplasma marginale outer membranesand protected against homologous challenge. Infect. Immun. 66:5406 –5413.
12. Brayton KA, Kappmeyer LS, Herndon DR, Dark MJ, Tibbals DL,Palmer GH, McGuire TC, Knowles DP. 2005. Complete genome se-quencing of Anaplasma marginale reveals that the surface is skewed to twosuperfamilies of outer membrane proteins. Proc. Natl. Acad. Sci. U. S. A.102:844 – 849.
13. Dark M, Herndon D, Kappmeyer L, Gonzales M, Nordeen E, Palmer G,Knowles D, Brayton K. 2009. Conservation in the face of diversity: mul-tistrain analysis of an intracellular bacterium. BMC Genomics 10:16. doi:10.1186/1471-2164-10-16.
14. Torioni de Echaide S, Knowles DP, McGuire TC, Palmer GH, SuarezCE, McElwain TF. 1998. Detection of cattle naturally infected with Ana-plasma marginale in a region of endemicity by nested PCR and a compet-itive enzyme-linked immunosorbent assay using recombinant major sur-face protein 5. J. Clin. Microbiol. 36:777–782.
15. Scoles GA, Broce AB, Lysyk TJ, Palmer GH. 2005. Relative efficiency ofbiological transmission of Anaplasma marginale (Rickettsiales: Anaplas-mataceae) by Dermacentor andersoni Stiles (Acari: Ixodidae) compared tomechanical transmission by the stable fly, Stomoxys calcitrans (L.) (Dip-tera: Muscidae). J. Med. Entomol. 42:668 – 675.
16. Ueti MW, Knowles DP, Davitt CM, Scoles GA, Baszler TV, Palmer GH.2009. Quantitative differences in salivary pathogen load during tick trans-mission underlie strain-specific variation in transmission efficiency ofAnaplasma marginale. Infect. Immun. 77:70 –75.
17. Scoles GA, Ueti MW, Palmer GH. 2005. Variation among geographicallyseparated populations of Dermacentor andersoni (Acari: Ixodidae) inmidgut susceptibility to Anaplasma marginale (Rickettsiales: Anaplas-mataceae). J. Med. Entomol. 42:153–162.
18. Agnes JA, Herndon D, Ueti MW, Ramabu S, Evans M, Brayton KA,Palmer GH. 2010. Association of pathogen strain-specific gene transcrip-tion and transmission efficiency phenotype of Anaplasma marginale. In-fect. Immun. 78:2446 –2453.
19. Ueti MW, Reagan JO, Knowles DP, Scoles GA, Shkap V, Palmer GH.2007. Identification of midgut and salivary gland as specific and distinctbarriers to efficient tick-borne transmission of Anaplasma marginale. In-fect. Immun. 75:2959 –2964.
20. de Silva AM, Tyson KR, Pal U. 2009. Molecular characterization of thetick-Borrelia interface. Front. Biosci. 14:3051–3063.
21. Francischetti IM, Sa-Nunes A, Mans BJ, Santos IM, Ribeiro JM. 2009.The role of saliva in tick feeding. Front. Biosci. 14:2051–2088.
22. Juncadella IJ, Anguita J. 2009. The immunosuppressive tick salivaryprotein, Salp15. Adv. Exp. Med. Biol. 666:121–131.
23. Galletti MF, Ueti MW, Knowles DP, Jr, Brayton KA, Palmer GH. 2009.Independence of Anaplasma marginale strains with high and low trans-mission efficiencies in the tick vector following simultaneous acquisitionby feeding on a superinfected mammalian reservoir host. Infect. Immun.77:1459 –1464.
24. Oberle M, Balmer O, Brun R, Roditi I. 2010. Bottlenecks and themaintenance of minor genotypes during the life cycle of Trypanosomabrucei. PLoS Pathog. 6:31001023. doi:10.1371/journal.ppat.1001023.
Herndon et al.
1858 iai.asm.org Infection and Immunity
on June 10, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
Related Documents











