System. Appl. Microbiol. 20, 232-237 (1997) © Gustav Fischer Verlag Identification of Bacillus kaustophilus, Bacillus thermocatenulatus and Bacillus Strain HSR as Members of Bacillus thermoleovorans ANWAR SUNNAl, SIMA TOKA]IAN 2 , JUTTA BURGHARDT3, FRED RAINEy 3 , GARABED ANTRANIKIANI and FUAD HASHWA 2 I Technical University Hamburg-Harburg, Institute of Biotechnology, Department of Technical Microbiology, Hamburg, Germany 2 Lebanese American University, Byblos, Lebanon 3 DSMZ - Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany Received November 25,1996 Summary An aerobic, endospore forming thermophilic microorganism was isolated from a soil sample in Jordan. The cells stained Gram-positive and the cylindrical spores are terminal and wider than the mother cell. Growth occurs at temperature values between 45°C and 70 °e, with an optimum at 55°C. The opti- mum pH for growth is 7.0. The G + e content of the DNA is 50.4 mol% and the sequence analysis of the 16S rDNA shows that the new isolate is phylogenetically closely related to the members of the Bacil- lus rRNA Group 5. DNA-DNA hybridization studies revealed a high similarity between the new isolate and the thermophilic species of the above mentioned group. A number of thermophilic Bacillus species are combined on the basis of DNA-DNA hybridization data. Key words: Bacillus - DNA-DNA hybridization - rDNAIrRNA analysis - Thermophile Introduction The increasing number of new extremophilic micro- organisms isolated in recent years has prompted studies in the utilization of thermophilic bacteria and hyper- thermophilic archaea, and the use of their thermostable enzymes, in several industrial processes (ZAMOST et aI., 1991; COWAN, 1992). The majority of the investi- gated thermophilic bacteria belong to the genus Bacillus and they have been isolated from thermo- philic and mesophilic environments (SHARP et aI., 1992). The inadequacy of Bacillus classification on the basis of morphology, physiology and ecological features has been emphasized by studies at the molecular level (ASH et aI., 1991; RAINEY et aI., 1994). The diversity of the genus Bacillus is indicated by the wide range of DNA base composition (33 to 68 mol% G + C) for the species (FHAMY et aI., 1985). Using the 16S rRNA sequences from 51 different Bacillus species, ASH et aI., (1991), proposed the division of the genus Bacillus in five phylo- genetically different clusters. According to this division group 5 was composed of three thermophilic species, namely B. stearothermophilus, B. kaustophilus and B. thermoglucosidasius. Recently RAINEY et aI., (1994), in- vestigated 16 thermophilic Bacillus strains by compara- tive 16S rDNA sequence analysis. Seven strains were found to cluster with B. stearothermophilus, B. kausto- phi/us and B. thermoglucosidasius in the rRNA group 5. In this study we describe the isolation and characteriza- tion of a thermophilic Bacillus strain. On the basis of mor- phological, physiological and phylogenetic studies, this new isolate is found to be closely related to the members of the Bacillus rRNA group 5. Furthermore, a DNA-DNA hybridization study of this group is presented. Materials and Methods Strains: The following Bacillus strains were obtained from the DSMZ - Deutsche Sammlung von Mikroorganismen und Zellkulturen, Braunschweig, Germany: "B. caldolyticus" (DSM 405), "B. caldotenax" (DSM 406), "B. caldovelox" (DSM 411), B. kaustophilus (DSM 7263), B. thermoleovorans (DSM 5366) and B. thermocatenulatus (DSM 730). Morphological studies: Gram-staining was done by the HUCKER method and spore-staining by the method of SCHAEF- FER and FULTON as described by SMIBERT and KRIEG (1994). Samples for photomicrographs under the light microscope were

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

System. Appl. Microbiol. 20, 232-237 (1997) © Gustav Fischer Verlag
Identification of Bacillus kaustophilus, Bacillus thermocatenulatus and Bacillus Strain HSR as Members of Bacillus thermoleovorans
ANWAR SUNNAl, SIMA TOKA]IAN2, JUTTA BURGHARDT3, FRED RAINEy3, GARABED ANTRANIKIANI and FUAD HASHWA2
I Technical University Hamburg-Harburg, Institute of Biotechnology, Department of Technical Microbiology, Hamburg, Germany 2 Lebanese American University, Byblos, Lebanon 3 DSMZ - Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany
Received November 25,1996
Summary
An aerobic, endospore forming thermophilic microorganism was isolated from a soil sample in Jordan. The cells stained Gram-positive and the cylindrical spores are terminal and wider than the mother cell. Growth occurs at temperature values between 45°C and 70 °e, with an optimum at 55°C. The optimum pH for growth is 7.0. The G + e content of the DNA is 50.4 mol% and the sequence analysis of the 16S rDNA shows that the new isolate is phylogenetically closely related to the members of the Bacillus rRNA Group 5. DNA-DNA hybridization studies revealed a high similarity between the new isolate and the thermophilic species of the above mentioned group. A number of thermophilic Bacillus species are combined on the basis of DNA-DNA hybridization data.
Key words: Bacillus - DNA-DNA hybridization - rDNAIrRNA analysis - Thermophile
Introduction
The increasing number of new extremophilic microorganisms isolated in recent years has prompted studies in the utilization of thermophilic bacteria and hyperthermophilic archaea, and the use of their thermostable enzymes, in several industrial processes (ZAMOST et aI., 1991; COWAN, 1992). The majority of the investigated thermophilic bacteria belong to the genus Bacillus and they have been isolated from thermophilic and mesophilic environments (SHARP et aI., 1992).
The inadequacy of Bacillus classification on the basis of morphology, physiology and ecological features has been emphasized by studies at the molecular level (ASH et aI., 1991; RAINEY et aI., 1994). The diversity of the genus Bacillus is indicated by the wide range of DNA base composition (33 to 68 mol% G + C) for the species (FHAMY et aI., 1985). Using the 16S rRNA sequences from 51 different Bacillus species, ASH et aI., (1991), proposed the division of the genus Bacillus in five phylogenetically different clusters. According to this division group 5 was composed of three thermophilic species, namely B. stearothermophilus, B. kaustophilus and B. thermoglucosidasius. Recently RAINEY et aI., (1994), in-
vestigated 16 thermophilic Bacillus strains by comparative 16S rDNA sequence analysis. Seven strains were found to cluster with B. stearothermophilus, B. kaustophi/us and B. thermoglucosidasius in the rRNA group 5.
In this study we describe the isolation and characterization of a thermophilic Bacillus strain. On the basis of morphological, physiological and phylogenetic studies, this new isolate is found to be closely related to the members of the Bacillus rRNA group 5. Furthermore, a DNA-DNA hybridization study of this group is presented.
Materials and Methods
Strains: The following Bacillus strains were obtained from the DSMZ - Deutsche Sammlung von Mikroorganismen und Zellkulturen, Braunschweig, Germany: "B. caldolyticus" (DSM 405), "B. caldotenax" (DSM 406), "B. caldovelox" (DSM 411), B. kaustophilus (DSM 7263), B. thermoleovorans (DSM 5366) and B. thermocatenulatus (DSM 730).
Morphological studies: Gram-staining was done by the HUCKER method and spore-staining by the method of SCHAEFFER and FULTON as described by SMIBERT and KRIEG (1994). Samples for photomicrographs under the light microscope were
Identification of Bacillus as members of Bacillus thermoleovorans 233
prepared by adding one drop of bacterial suspension on agarcoated slides. The preparation of transmission electron micrographs of whole cells was done as follows; one drop of bacterial suspension was placed on a copper grid covered with formvar film. The bacteria were negatively stained with an aqueous solution of 2% (w/v) phosphotungestic acid adjusted to pH 7.0 with NaOH. The grids were examined and electron micrographs were taken with a Zeiss EM lOB transmission electron microscope.
Physiological tests: Tests were performed as described by MAC FADDING (1980), and SMIBERT and KRIEG (1994). Cellulose and xylan hydrolysis were determined by using nutrient agar (Oxoid, Wesel, Germany) supplemented with 0.5% (w/v) CMC-celluloseRBB and CMC-xylan-RBB (Loewe Biochemica, Miinchen, Germany), respectively. Optimum temperature and pH for growth were determined using flasks containing 100 ml nutrient broth (Oxoid) prepared in the universal buffer of BRITTON and ROBINSON (1931). The effect of different concentration of NaCI was determined in the same medium. In all experiments the cultures (in triplicates) were incubated in a shaking incubator (New Brunswick Scientific, USA) with an agitation rate of 150 rpm. Samples were taken at different time intervals and growth was determined spectrophotometrically at 550 nm.
Membrane fatty acids: Strain HSR was grown on trypticase soy broth agar (Becton Dickinson, Cockeysville, USA) at 55°C. Fatty acids were obtained from biomass by saponification, methylation and extraction using minor modification (KUYKENDALL et al., 1988) of the method of MILLER (1982). The fatty acid methylesters mixtures were separated using a model 5898A microbial identification system (Microbial ID Inc., Newark, Delaware, USA) as described by VAINSHTEIN et al. 1992.
Determination of G + C content of DNA: The DNA was isolated as described by CASHION et al. (1977). The G +C content of the DNA was determined by HPLC according to M ESBAH et al. (1989).
16S rDNA sequence determination and analysis: Genomic DNA extraction, PCR mediated amplification of the 16S rDNA
and sequencing of the purified PCR products were carried out as described previously (RAINEY et al., 1996). Sequence reaction mixtures were electrophoresed using a model 373A automated DNA sequencer (Applied Biosystem, Foster City, California, USA). The 16S rDNA sequence obtained in this study were manually aligned with reference sequences for members of the genus Bacillus using the ae2 editor (MAlDAK et al., 1994).
DNA-DNA hybridization method: DNA was isolated by chromatography on hydroxyapatite by the procedure of CASHION et al. (1977). DNA-DNA hybridization was carried out as described by DE LEY et al. (1970), with the modifications described by Huss et al. (1983), using a Gilford System model 2600 spectrophotometer equipped with a Gilford model 2527-R thermoprogrammer and plotter. Renaturation rates were computed with the TRANSFER.BAS programm (JANKE, 1992).
Results and Discussion
Isolation and growth conditions
The strain HSR was isolated from a soil sample in Jordan on a medium containing (w/v): 0.5% peptone (Difco), 0.4% beef extract (Oxoid), 0.2% yeast extract (Difco), 0.5% NaCI and 2% agar. The pH of the medium was adjusted to 7.0. Plates were incubated at 55°C for 24 h, and purity of the colonies was checked microscopically.
Morphology and physiological properties of the isolate
After 24 h of growth on nutrient agar strain HSR colonies were small, flat, irregular to circular in form, with smooth shiny surface and undulate edges. The colonies were 1 to 2 mm in diameter. Strain HSR cells were Gram-positive straight rods with peritrichous flag-
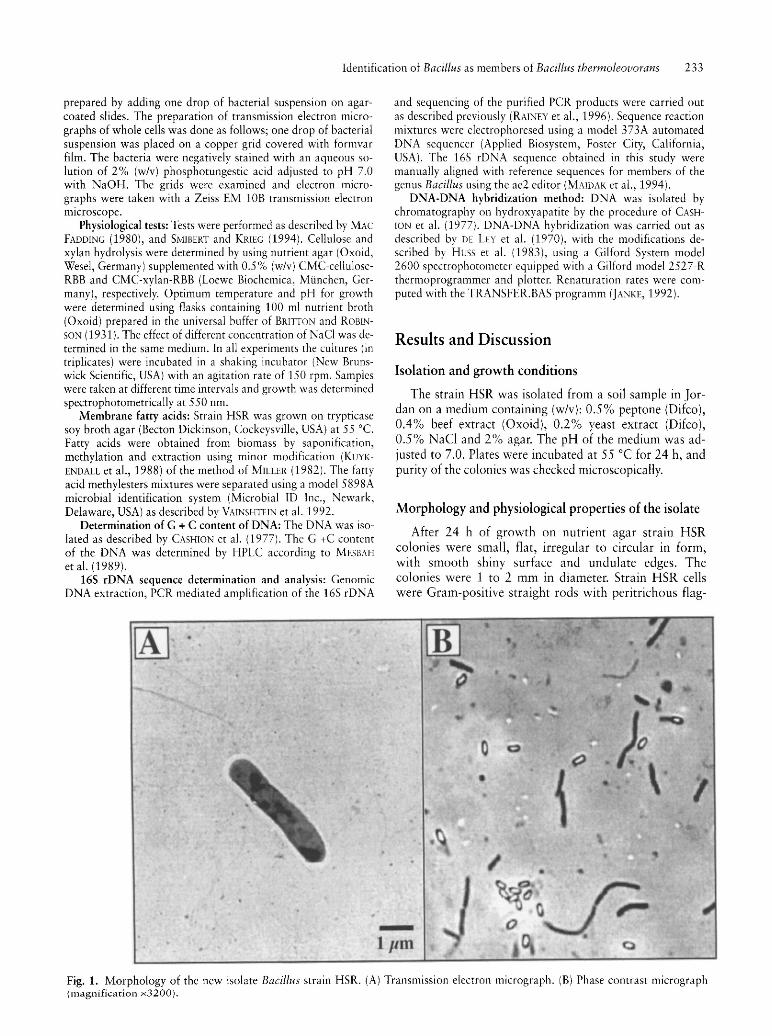
-1 Jim
Fig. 1. Morphology of the new isolate Bacillus strain HSR. (A) Transmission electron micrograph. (B) Phase contrast micrograph (magnification x3200).

234 A. SUNNA et al.
ellation that occurred either singly or in chains of two to four cells. The cells were 0.8-1.0 }lm wide and 3.0-6.0 }lm long (Fig. 1A). The cylindrical endospores were terminal and wider than the mother cell (Fig. 1B). When strain HSR was cultivated on nutrient broth elongated structures were observed even after 48 h of incubation. Electron micrographs revealed that these structures were actually chains composed of several unseparated cells. Formation of spheroplast is an important morphological feature of strain HSR. Spheroplast were mainly produced in the stationary and decline growth phase upon cultivation in liquid media. Spores were rarely formed in liquid cultures but they are observed after cultivation on solid media.
Optimal growth occurred at 55°C with the lower and upper growth limit at 45 °C and 70°C, respectively. The optimal pH for growth was 7.0 and no growth was observed at pH 5.0 or 9.0, respectively. The main physiological properties of strain HSR are presented in Table 1.
Fatty acid composition
The main fatty acids of strain HSR are the iso-branched pentadecanoic acid (iso-C15), hexadecanoic acid (isoC16) and heptadecanoic acid (iso-C17) (data not shown). They account for 84% of the total fatty acids. In particular, iso-C15 is the most abundant representing 51 % of the total fatty acids. Similar to strain HSR, in "B. caldolyticus", "B. caldove/ox", "B. caldotenax" and "B. thermocloacae" the branched iso-C15, iso-C16 and iso-C17 acids have been reported to represent about 80% of the total membrane fatty acids (HEINEN and HEINEN, 1972; WEERKAMP and HEINEN, 1972; DEMHARTER and HENSEL, 1989). Adaptation to high growth temperatures resulted in an increase in the relative amount of the high melting point iso-fatty acids and a decrease of the low melting point anteiso-fatty acids. This biochemical adaptation seems to be necessary to mantain membrane fluidity at elevated growth temperatures (KANEDA, 1977; KANEDA, 1991). Recently, NICOLAUS et al. (1995) isolated seven thermophilic strains from the Antartic. Growth of the new isolates at temperatures between 40°C and 71 °C showed that at higher temperatures in all strains the synthesis of the higher melting point (iso-C17) rather than that of lower melting point acids was preferred.
Table 1. Physiological properties of Bacillus strain HSR.
Catalase
Oxidase
Anaerobic growth
Growth in the presence of 0.5% NaCI 2.0% NaCl 5.0% NaCI
Optimum temperature of growth
Optimum pH of growth
Spore formation
Hydrolysis of starch casem gelatin urea cellulose xylan tween 20 tween 80
Citrate utilization
Acid from glucose
Formation of acetoin indole H2S
Nitrate reduction
Denitrification
+ positive; - negative
Phylogenetic position
+
+
+
+
+
+ + + +
+
+
+
The G + C content of the DNA of strain HSR was determined to be 50.4%. The results of the 16S rDNA sequencing indicate that Bacillus strain HSR share more than 99 % sequence similarity to the type strains" B. caldovelox", "B. caldotenax", "B. caldolyticus", B. kaustophilus, B. thermoleovorans and B. thermocatenulatus (Table 2). This indicates clearly that Bacillus strain HSR is phylogenetically closely related to the members of the Bacillus rRNA group 5 (ASH et aI., 1991; RAINEY et aI., 1994). DNA-DNA hybridization experiments between Bacillus strain HSR and those thermophilic bacilli strains showing more than 99% 16S rDNA similarity values, re-
Table 2. 16S rDNA similarity values for Bacillus strain HSR an the closely related thermophilic species of Bacillus rRNA group 5.
Strains 1. 2. 3. 4. 5. 6. 7. 8.
1. Bacillus strain HSR 2. B. kaustophilus 99.9 3. "B. caldolyticus" 99.9 100.0 4. B. thermoleovorans 99.8 99.9 99.9 5. "B. caldotenax" 99.8 99.9 99.9 99.9 6. "B. caldovelox" 99.6 99.8 99.8 99.7 99.7 7. B. thermocatenulatus 99.1 99.3 99.3 99.2 99.2 99.2 8. B. stearothermophilus 99.0 99.1 99.1 99.1 99.1 98.9 98.8
Identification of Bacillus as members of Bacillus thermoleovorans 235
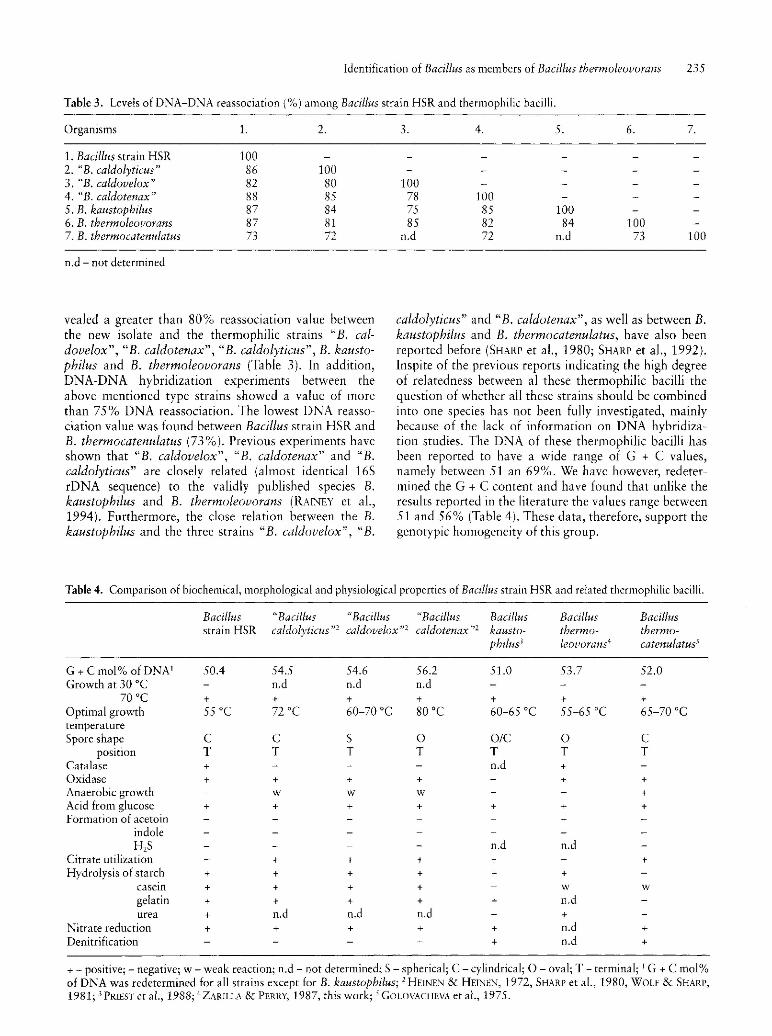
Table 3. Levels of DNA-DNA reassociation (%) among Bacillus strain HSR and thermophilic bacilli.
Organisms 1. 2.
1. Bacillus strain HSR 100 2. "B. caldolyticus" 86 100 3. "B. caldovelox" 82 80 4. "B. caldotenax" 88 85 5. B. kaustophilus 87 84 6. B. thermoleovorans 87 81 7. B. thermocatenulatus 73 72
n.d - not determined
vealed a greater than 80% reassociation value between the new isolate and the thermophilic strains "B. caldovelox", "B. caldotenax", "B. caldolyticus", B. kaustophilus and B. thermoleovorans (Table 3). In addition, DNA-DNA hybridization experiments between the above mentioned type strains showed a value of more than 75% DNA reassociation. The lowest DNA reassociation value was found between Bacillus strain HSR and B. thermocatenulatus (73%). Previous experiments have shown that "B. caldovelox", "B. caldotenax" and "B. caldolyticus" are closely related (almost identical 16S rDNA sequence) to the validly published species B. kaustophilus and B. thermoleovorans (RAINEY et aI., 1994). Furthermore, the close relation between the B. kaustophilus and the three strains" B. caldovelox", "B.
3.
100 78 75 85
n.d
4.
100 85 82 72
5.
100 84
n.d
6.
100 73
7.
100
caldolyticus" and "B. caldotenax", as well as between B. kaustophilus and B. thermocatenulatus, have also been reported before (SHARP et aI., 1980; SHARP et aI., 1992). Inspite of the previous reports indicating the high degree of relatedness between al these thermophilic bacilli the question of whether all these strains should be combined into one species has not been fully investigated, mainly because of the lack of information on DNA hybridization studies. The DNA of these thermophilic bacilli has been reported to have a wide range of G + C values, namely between 51 an 69%. We have however, redetermined the G + C content and have found that unlike the results reported in the literature the values range between 51 and 56% (Table 4). These data, therefore, support the genotypic homogeneity of this group.
Table 4. Comparison of biochemical, morphological and physiological properties of Bacillus strain HSR and related thermophilic bacilli.
Bacillus "Bacillus "Bacillus "Bacillus Bacillus Bacillus Bacillus strain HSR caldolyticus"2 caldovelox "2 caldotenax "2 kausto- thermo- thermo-
philus3 leovorans4 catenulatus5
G + C mol% ofDNN 50.4 54.5 54.6 56.2 51.0 53.7 52.0 Growth at 30°C n.d n.d n.d
70°C + + + + + + + Optimal growth 55 °C 72 °C 60-70°C 80°C 60-65°C 55-65°C 65-70 °C temperature Spore shape C C S 0 OIC 0 C
position T T T T T T T Catalase + n.d + Oxidase + + + + + + Anaerobic growth w w w + Acid from glucose + + + + + + + Formation of acetoin
indole H 2S n.d n.d
Citrate utilization + + + + + Hydrolysis of starch + + + + + +
casein + + + + w w gelatin + + + + + n.d urea + n.d n.d n.d +
Nitrate reduction + + + + + n.d + Denitrification + n.d +
+ - positive; - negative; w - weak reaction; n.d - not determined; S - spherical; C - cylindrical; 0 - oval; T - terminal; 1 G + C mol % of DNA was redetermined for all strains except for B. kaustophilus; 2 HEINEN & HEINEN, 1972, SHARP et ai., 1980, WOLF & SHARP, 1981; 3 PRIEST et aI., 1988; 4 ZARILLA & PERRY, 1987, this work; 5 GOLOVACHEVA et ai., 1975.

236 A. SUt>;NA et al.
The level of DNA-DNA similarity for species boundaries was suggested to be around 70% (WAYNE et aI., 1987). Accordingly, B. thermoleovorans, B. kaustophilus and B. thermocatenulatus should be combined into one species, namely B. thermoleovorans, which also includes the strains Bacillus strain HSR, "B. caldolyticus", "B. caldovelox" and "B caldotenax". Thus, the emended Bacillus thermoleovorans species is characterized by its phenotypic heterogeneity (Table 4) and genotypic homogeneity.
Description of Bacillus thermoleovorans
[Bacillus thermoleovorans (ZARILLA and PERRY, 1987; emend SUNNA et aI., 1997)]
therm'o. Ie. o. vo'rans. Gr. n. therme heat; L n. oleum oil; L v. vorare to devour; M. L pres. part. thermoleovorans indicating heat-requiring bacteria capable of utilizing oil (hydrocarbons).
Cells are rod-shaped, I-811m long and 0.2-9 11m wide, occurring single or in long unbranched chains. The Gram reaction can be negative or positive depending on the strain. The terminal spores are oval to cylindrical. Yellowish pigmented as well as non pigmented colonies have been described. Growth is aerobic, however, one strain is able to grow under facultative anaerobic conditions. Growth occurs at temperatures between 35 and 78 °C with optimum growth between 55-70°C and pH 6.0-7.5. Catalase and oxidase are negative or positive depending on the strain. Growth factors are not required. Substrates that support growth include: glucose, maltose, xylose, mannose, saccharose, cellobiose, galactose, mannitol, glycerol. Growth in the presence of nalkanes (C 13-C20 ) is the main characteristic of some strains. No growth is observed with arabinose, lactose or rhamnose. Citrate is utilized. Starch is hydrolyzed but not urea. Casein is weakly hydrolyzed while gelatine and Tween 80 can be hydrolyzed by some strains. Indole and acetoin are not formed. Nitrate is reduced. The G + C content of the DNA ranges between 51 and 56 mol%.
Cells can be isolated from mud, soil, pasteurized milk, deteriorated canned food as well as, thermal and nonthermal environments.
The type strain of B. thermoleovorans is strain DSM 5366 (ATCC 43513). The G + C of the DNA of this strain is 53.7 mol%.
Acknowledgements We thank Prof. M. DA COSTA, from the University of Coim
bra, Portugal, for determining the fatty acid composition.
References
ASH, e., FARROW, J.A.E., WALLBANKS, S., COLLINS, M.D.: Phylogenetic heterogeneity of the genus Bacillus revealed by comparative analysis of sma ll-subunit-ribosomal RNA. Lett. Appl. Microbiol. 13,202-206 (1991).
BRITTON, H .T.S., ROBINSON, R.A.: Universal buffer solutions and the dissociation constant of verona 1. J. Chern. Soc. 1456-1462 (1931).
CASHION, P., HOLDER-FRANKLIN, M.A., MCCULLY,]., FRANKLIN, M.: A rapid method for the base ratio determination of bacterial DNA. Anal. Biochem. 81,461-466 (1977).
COWAN, D.A.: Biotechnology of the archaea . TIBTECH 10, 315-323 (1992).
DE LEY, ]., CATTOIR, H., RAYNAERTS, A.: The quantitative measurement of DNA hybridization from renaturation rates . Eur. J. Biochem. 12, 133-142 (1970).
DEMHARTER, W., HENSEL, R.: Bacillus thermocloacae sp. nov., a new thermophilic species from sewage sludge. System. Appl. Microbiol. 11,272-276 (1989) .
FAHMY, E, FLOSSDORF, ]., CLAUS, D.: The DNA base composition of the type strains of the genus Bacillus. System. Appl. Microbiol. 6, 60-65 (1985).
GOLOVACHEVA, R.S., LOGINOVA, L.G., SALIKHOV, T.A., KOLESNIK OV, A.A., ZAITSEVA, G.N.: A new thermophilic species, Bacillus thermocatenulatus nov. sp. Microbiology 44, 230-233 (1975).
HEINEN, U.]., HEINEN, W.: Characteristics and properties of a caldo-active bacterium producing extracellular enzymes and two related strains. Arch. Mikrobiol. 82, 1-23 (1972).
Huss, VA.R., FESTL, H., SCHLEIFER, K.H.: Studies on the spectrophotometric determination of DNA hybridization from renaturation rates. System. Appl. Microbiol. 4, 184-192 (1983 ).
JANKE, K.-D.: BASIC computer program for evaluation of spectroscopic DNA renaturation data from GILFORD SYSTEM 2600 spectrophotometer on a Pc/XT/AT type personal computer. J. Microbiol. M ethods 15, 61-73 (1992).
KANEDA, T.: Fatty acids in the genus Bacillus: an example of branched-chain preference. Bacteriol. Rev. 41, 391-418 (1977).
KANEDA, T.: Iso- and anteiso-fatty acids in bacteria : biosynthesis, function, and taxonomic significance. Microbiol. Rev. 55,288-302 (1991).
KUYKENDALL, L.D., ROY, M.D., A'NEILL, J.J., DEVIt>;E, T.E.: Fatty acids, antibiotic resistance, and deoxyribonucleic acid homology groups of Bradyrhizobium japonicum. Int. J. Syst. Bacteriol. 38, 358-361 (1988).
MAC FAD DI N, J.: Biochemical tests for the identification of medical bacteria, 2nd ed., Baltimore, Waverly Press 1980.
MAlDAK, B.L., LARSEN, N., MCCAUGHEY, M.J., OVERBEEK, R., OLSEN, G.]., FOGEL, K., BLANDY, ]., WOESE, e.R.: The Ribosomal Data Base Project. Nucleic Acids Res. 22, 3485-3487 (1994).
MESBAH, M., PREMACHANDRAN, U., WHITMAN, W.B.: Precise measurement of the G + C content of deoxyribonucleic acid by high-performance liquid chromatography. Int. J. Syst. Bacteriol. 39, 159-167 (1989).
MILLER, L.T.: A single derivatization method for bacterial fatty acid methyl esters including hydroxy acids. J. Clin. Microbi-01. 16,584-586 (1982).
NICOLAUS, B., MANCA, M.e., LAMA, L., ESPOSITO, E., GAMBACORTA, A.: Effects of growth temperature on the polar lipid pattern and fatty acid composition of seven thermophilic isolates from the Antarctic Continent. System. Appl. Microbiol. 18,32-36 (1995).
PRIEST, EG., GOODFELLOW, M., TODD, e.: A numerical classification of the genus Bacillus. J. Gen. Microbiol. 134, 1847-1882 (1988).
RAINEY, EA., FRITZE, D. , STACKEBRANDT, E.: The phylogenetic diversity of thermophilic members of the genus Bacillus as revealed by 16S rDNA analysis. FEMS Microbiol. Lett. 115, 205-212 (1994).
RAINEY, EA., WARD-RAINEY, N., KROPPENSTEDT, R.M., STACKEBRANDT, E.: The genus Nocardiopsis represents a phylogenet-

Identification of Bacillus as members of Bacillus thermoleovorans 237
ically coherent taxon and a distinct actinomycete lineage: proposal of Nocardiopsaceae fam. nov. Int.]. Syst. Bacteriol. 46,1088-1092(1996).
SHARP, R.]., BOWN, K.]., ATKINSON, A.: Phenotypic and genotypic characterization of some thermophilic species of BacilIus.]. Gen. Microbiol. 117,201-210 (1980).
SHARP, R.J., RILEY, P.W., WHITE, D.: Heterotrophic thermophilic Bacilli, pp. 19-50. In: Thermophilic bacteria U.K. KRISTJANSSON, ed.) Boca Raton, CRC Press 1992.
SMIBERT, R.M., KRIEG, N.R.: Phenotypic characterization, pp. 607-654. In: Methods for general and molecular bacteriology (P. GERHARDT, R.G.E. MURRAY, W.A. WOOD, N.R. KRIEG, eds.) Washington, American Society for Microbiology 1994.
VAINSHTEIN, M., HIPPE, H., KROPPENSTEDT, R.M.: Cellular fatty acid composition of Desulfovibrio species and its use in classification of sulfate-reducing bacteria. System. Appl. MicrobioI. 15,554-566 (1992).
WAYNE, L.G., BRENNER, D.]., COLWELL, R.R., GRIMONT, P.A.D., KANDLER, 0., KRICHEVSKY, M.l., MOORE, L.H., MOORE, W.E.C., MURRAY, R.G.E., STACKEBRANDT, E., STARR, M.P., TROPER, H.G.: Report the Ad Hoc Committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Bacteriol. 37,463-464 (1987).
WOLF, ]., SHARP, R.].: Taxonomic and related aspects of thermophiles within the genus Bacillus, pp. 251-296. In: The aerobic endospore-forming bacteria: classification and identification (R.C.W. BERKELEY, M. GOODFELLOW, eds.) London, Academic Press 1981.
WEERKAMP, A., HEINEN, W.: Effect of temperature on the fatty acid composition of the extreme thermophiles B. caldolyticus and B. caldotenax. J. Bacteriol. 109,443-446 (1972).
ZAMOST, B.L., NIELSEN, H.K., STARNES, R.L.: Thermostable enzymes for industrial applications. J. Ind. Microbiol. 8, 71-82 (1992).
ZARILLA, K.A., PERRY, J.J.: Bacillus thermoleovorans, sp. nov., a species of obligately thermophilic hydrocarbon utilizing endospore-forming bacteria. System. Appl. Microbiol. 9, 258-264 (1987).
Corresponding author: G. ANTRANIKIAN, Technical University Hamburg-Harburg, Institute of Biotechnology, Department of Technical Microbiology, Denickestr. 15, D-21071 Hamburg, Germany Phone: +4940-7718-3117; Fax: +4940-7718-2909; E-mail: antranikian @tu-harburg.d400.de
Related Documents











