Identification of a novel cell cycle regulated gene, HURP, overexpressed in human hepatocellular carcinoma Ann-Ping Tsou 1,11 , Chu-Wen Yang 2,11 , Chi-Ying F Huang 3,11 , Ricky Chang-Tze Yu 3 , Yuan-Chii G Lee 3 , Cha-Wei Chang 4 , Bo-Rue Chen 5 , Yu-Fang Chung 5 , Ming-Ji Fann 6 , Chin-Wen Chi 7,8 , Jen-Hwey Chiu 9 and Chen-Kung Chou* ,3,7,10 1 Institute of Biotechnology in Medicine, National Yang-Ming University, Taipei 112, Taiwan, ROC; 2 Institute of Biochemistry, National Yang-Ming University, Taipei 112, Taiwan, ROC; 3 Division of Molecular and Genomic Medicine, National Health Research Institutes, Nankang, Taipei 115, Taiwan, ROC; 4 Institute of Microbiology and Immunology, National Yang-Ming University, Taipei 112, Taiwan, ROC; 5 Institute of Genetics, National Yang-Ming University, Taipei 112, Taiwan, ROC; 6 Institute of Neuroscience, National Yang-Ming University, Taipei 112, Taiwan, ROC; 7 Department of Medical Research and Education, Veterans General Hospital-Taipei 112, Taiwan, ROC; 8 Institute of Pharamacology, National Yang-Ming University, Taipei 112, Taiwan, ROC; 9 Institute of Traditional Medicine, National Yang-Ming University, Taipei 112, Taiwan, ROC; 10 Department of Life Science, National Yang-Ming University, Taipei 112, Taiwan, ROC An analytic strategy was followed to identify putative regulatory genes during the development of human hepatocellular carcinoma (HCC). This strategy employed a bioinformatics analysis that used a database search to identify genes, which are differentially expressed in human HCC and are also under cell cycle regulation. A novel cell cycle regulated gene (HURP) that is overexpressed in HCC was identified. Full-length cDNAs encoding the human and mouse HURP genes were isolated. They share 72 and 61% identity at the nucleotide level and amino- acid level, respectively. Endogenous levels of HURP mRNA were found to be tightly regulated during cell cycle progression as illustrated by its elevated expression in the G 2 /M phase of synchronized HeLa cells and in regener- ating mouse liver after partial hepatectomy. Immuno- fluorescence studies revealed that hepatoma up-regulated protein (HURP) localizes to the spindle poles during mitosis. Overexpression of HURP in 293T cells resulted in an enhanced cell growth at low serum levels and at polyhema-based, anchorage-independent growth assay. Taken together, these results strongly suggest that HURP is a potential novel cell cycle regulator that may play a role in the carcinogenesis of human cancer cells. Oncogene (2003) 22, 298–307. doi:10.1038/sj.onc.1206129 Keywords: HURP; heptocellular carcinoma; liver regeneration; cell cycle regulator; bioinformatics Introduction Hepatocellular carcinoma (HCC) is one of the most frequent malignant neoplasms in humans and is especially prevalent among Asian populations. Chronic infection with hepatitis B virus or hepatitis C virus, the ingestion of food contaminated with chemical carcino- gens and the consumption of alcoholic beverages are considered to be the major risk factors for HCC development (Feitelson and Duan, 1997; Bosch et al., 1999; Boyer and Marcellin, 2000). Molecular analyses have shown that the pathogenesis of HCC is a multi- factorial process, in which the occurrence of somatic mutations, the loss of function of p53 or similar tumor suppressor gene(s), and activation or overexpression of certain proto-oncogenes may all play a role in the various stages (Su et al., 1994; Okuda, 2000; Tsou et al., 1998). Furthermore, the presence of cirrhosis and other types of lesions add further complexity to the task of dissecting the molecular mechanisms of hepatocarcino- genesis. In the postgenomic era, the emerging technologies of cDNA microarrays (Ferea and Brown, 1999; Okabe et al., 2001) and bioinformatics analysis of cDNA sequence databases make it possible to analyze global gene expression in different human cancer cells or tissues. Microarray studies have revealed that genes pivotal in cell cycle regulation may have important roles in tumorigenesis. Study of HCC gene expression profiles showed a group of mitosis-associated genes to be upregulated in the majority of the liver tumors examined (Okabe et al., 2001). Independently, identifi- cation of mutations in the mitotic checkpoint genes BUB1 (Cahill et al., 1998), Mad1 (Tsukasaki et al., 2001), hsMAD2, and p55CDC (Takahashi et al., 1999) in human cancers strongly suggests that there is a significant correlation between M-phase control and tumor progression. Received 25 June 2002; revised 4 October 2002; accepted 8 October 2002 *Correspondence: C-K Chou, Department of Life Science, National Yang-Ming University, Shih-Pai, Taipei 112, Taiwan, ROC; Email: [email protected] 11 These authors contributed equally to this work. Oncogene (2003) 22, 298–307 & 2003 Nature Publishing Group All rights reserved 0950-9232/03 $25.00 www.nature.com/onc ONCOGENOMICS

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Identification of a novel cell cycle regulated gene, HURP, overexpressed in
human hepatocellular carcinoma
Ann-Ping Tsou1,11, Chu-Wen Yang2,11, Chi-Ying F Huang3,11, Ricky Chang-Tze Yu3, Yuan-ChiiG Lee3, Cha-Wei Chang4, Bo-Rue Chen5, Yu-Fang Chung5, Ming-Ji Fann6, Chin-Wen Chi7,8,Jen-Hwey Chiu9 and Chen-Kung Chou*,3,7,10
1Institute of Biotechnology in Medicine, National Yang-Ming University, Taipei 112, Taiwan, ROC; 2Institute of Biochemistry,National Yang-Ming University, Taipei 112, Taiwan, ROC; 3Division of Molecular and Genomic Medicine, National Health ResearchInstitutes, Nankang, Taipei 115, Taiwan, ROC; 4Institute of Microbiology and Immunology, National Yang-Ming University, Taipei112, Taiwan, ROC; 5Institute of Genetics, National Yang-Ming University, Taipei 112, Taiwan, ROC; 6Institute of Neuroscience,National Yang-Ming University, Taipei 112, Taiwan, ROC; 7Department of Medical Research and Education, Veterans GeneralHospital-Taipei 112, Taiwan, ROC; 8Institute of Pharamacology, National Yang-Ming University, Taipei 112, Taiwan, ROC;9Institute of Traditional Medicine, National Yang-Ming University, Taipei 112, Taiwan, ROC; 10Department of Life Science,National Yang-Ming University, Taipei 112, Taiwan, ROC
An analytic strategy was followed to identify putativeregulatory genes during the development of humanhepatocellular carcinoma (HCC). This strategy employeda bioinformatics analysis that used a database search toidentify genes, which are differentially expressed in humanHCC and are also under cell cycle regulation. A novel cellcycle regulated gene (HURP) that is overexpressed inHCC was identified. Full-length cDNAs encoding thehuman and mouse HURP genes were isolated. They share72 and 61% identity at the nucleotide level and amino-acid level, respectively. Endogenous levels of HURPmRNA were found to be tightly regulated during cell cycleprogression as illustrated by its elevated expression in theG2/M phase of synchronized HeLa cells and in regener-ating mouse liver after partial hepatectomy. Immuno-fluorescence studies revealed that hepatoma up-regulatedprotein (HURP) localizes to the spindle poles duringmitosis. Overexpression of HURP in 293T cells resultedin an enhanced cell growth at low serum levels and atpolyhema-based, anchorage-independent growth assay.Taken together, these results strongly suggest that HURPis a potential novel cell cycle regulator that may play arole in the carcinogenesis of human cancer cells.Oncogene (2003) 22, 298–307. doi:10.1038/sj.onc.1206129
Keywords: HURP; heptocellular carcinoma; liverregeneration; cell cycle regulator; bioinformatics
Introduction
Hepatocellular carcinoma (HCC) is one of the mostfrequent malignant neoplasms in humans and isespecially prevalent among Asian populations. Chronicinfection with hepatitis B virus or hepatitis C virus, theingestion of food contaminated with chemical carcino-gens and the consumption of alcoholic beverages areconsidered to be the major risk factors for HCCdevelopment (Feitelson and Duan, 1997; Bosch et al.,1999; Boyer and Marcellin, 2000). Molecular analyseshave shown that the pathogenesis of HCC is a multi-factorial process, in which the occurrence of somaticmutations, the loss of function of p53 or similar tumorsuppressor gene(s), and activation or overexpression ofcertain proto-oncogenes may all play a role in thevarious stages (Su et al., 1994; Okuda, 2000; Tsou et al.,1998). Furthermore, the presence of cirrhosis and othertypes of lesions add further complexity to the task ofdissecting the molecular mechanisms of hepatocarcino-genesis.
In the postgenomic era, the emerging technologies ofcDNA microarrays (Ferea and Brown, 1999; Okabeet al., 2001) and bioinformatics analysis of cDNAsequence databases make it possible to analyze globalgene expression in different human cancer cells ortissues. Microarray studies have revealed that genespivotal in cell cycle regulation may have importantroles in tumorigenesis. Study of HCC gene expressionprofiles showed a group of mitosis-associated genes tobe upregulated in the majority of the liver tumorsexamined (Okabe et al., 2001). Independently, identifi-cation of mutations in the mitotic checkpoint genesBUB1 (Cahill et al., 1998), Mad1 (Tsukasaki et al.,2001), hsMAD2, and p55CDC (Takahashi et al., 1999)in human cancers strongly suggests that there is asignificant correlation between M-phase control andtumor progression.
Received 25 June 2002; revised 4 October 2002; accepted 8October 2002
*Correspondence: C-K Chou, Department of Life Science, NationalYang-Ming University, Shih-Pai, Taipei 112, Taiwan, ROC; Email:[email protected] authors contributed equally to this work.
Oncogene (2003) 22, 298–307& 2003 Nature Publishing Group All rights reserved 0950-9232/03 $25.00
www.nature.com/onc
ON
CO
GE
NO
MIC
S

Over 9 million expressed sequences from diverseorgan-, species-, and disease-oriented cDNA librarieshave been deposited in the expressed sequence tag(EST) database, dbEST (http://www.ncbi.nlm.nih.gov/dbEST/). The development of bioinformatics tools toanalyze the EST database has become an importantapproach in the identification of candidate genes thatmay be preferentially expressed in cancer tissues.Combined data mining from both EST and microarraydatabases provides a unique opportunity to identifyputative diagnostic and therapeutic targets for humancancer. Recently, we have collected over 180 000 ESTsderived from cDNA libraries of various human livertissues including fetal liver, normal adult liver, pre-cancerous liver, HCC, and cultured hepatoma cells. Inthis study, we searched for novel genes specificallypresent in the EST collection derived from human HCCtissues and also shown to have a cell cycle-dependentexpression profile in microarray databases (Iyer et al.,1999). Search results led to the identification andcharacterization of one novel cell cycle regulated gene,HURP.
Materials and methods
Database analysis
An NCBI UniGene database search for ESTs expressed inhuman liver-related cDNA libraries resulted in a set of 137 142sequences. In addition, seven cDNA libraries were constructedfrom one normal adult liver tissue specimen and three pairs ofspecimens of HCC tumor tissues and their adjacent tissues. Atotal of 5615 sequences were collected from these sevenlibraries (http://lestdb.nhri.org.tw). According to the tissueorigin of the cDNA libraries, these 142 757 ESTs were furthergrouped into five categories: fetal liver/spleen tissues (116 698),normal adult liver tissues (19 944), HCC-tumor (hepatoma)tissues (2694), tumor-adjacent normal tissues (2457), and HCCcell lines (964). The identities of these ESTs were searchedagainst the UniGene Build#79 and Genbank (version on May16, 1999) using NCBI blast software version 2.0. An EST wasassigned to its UniGene group when it shared 85% sequenceidentity with a stretch of 100 base pairs (bp) from a knowngene. We then searched for the cell cycle expression profile ofeach UniGene group in the database established by Iyer et al.(1999). Iyer’s database was compiled using a microarraysystem, which contains the expression profile of 8600 genes ofnormal human fibroblasts after serum stimulation.
Cloning of human and mouse HURP genes
Cloning of the full-length HURP cDNAs of human and mouseorigin was accomplished by assembling EST clones usingSequencher software, PCR cloning, cDNA library screening,and RACE (Rapid Amplification of cDNA Ends). Humanfull-length cDNA of HURP was isolated from Hep3B, acell line derived from HCC. A total of 13 mouse ESTclones (AI592008, C88298, AI510131, AA162837, AI1550612,AA511899, AI552952, C78700, AA212615, AI605993,AI482307, AI427201, and AI563636) showed homology withhuman HURP sequences. Assembly of these ESTs generated aDNA fragment of 1759 bp with an overall similarity of 55%with human HURP. The missing 30 fragment (approximately600 bp) was subsequently cloned from a mouse cDNA library.
50-RACE generated clones with a longer 50 end, with themouse hepatoma up-regulated protein (HURP) containing 808amino acids. The results of investigation of the characteristicsof human and mouse HURP are described in this paper.
Cell cultures, transfection and cell cycle synchronization
HeLa cells and human 293T cells from the American TypeCulture Collection were maintained at 371C in a 5% CO2
incubator and grown in DMEM medium supplemented with10% heat-inactivated fetal bovine serum and 100 mg/mlpenicillin/streptomycin (BRL). Stable transfectants of HURPwere generated by transfecting 293T cells with pCMV2-FLAG-HURP and pTK-Hygo at a 9 : 1 ratio. Several stablytransfected cell lines were isolated and cultured in the selectiveculture medium with hygromycin B (0.5mg/ml).Cells were synchronized at the G1/S boundary by thymidine
and aphidicolin. Briefly, HeLa cells were cultured for 14 h inthe presence of 2mm thymidine (Sigma), released for 12 h infresh medium, and finally arrested for 14 h in the presence of1.6 mg/ml aphidicolin (Sigma). Cells were released from theaphidicolin block by changing to fresh culture medium in theabsence of aphidicolin. The percentage of cells at differentphases of the cell cycle was determined by flow cytometry(FACS analysis, Becton Dickinson FACStar Plus flowcytometer).
Liver regeneration
Experiments involving the regeneration of liver tissue werecarried out in BALB/C mice. Mice were anesthetized andsubjected to partial hepatectomy by the removal of left lateraland median lobes (70% liver resection) as previously described(Higgins and Anderson, 1931). After the surgery, the animalswere allowed to recover for times ranging from 15min to 72 h.Animals were killed and liver samples were immediately frozenin liquid nitrogen and stored at �801C.
RNA preparation and real-time RT–PCR
Total RNA was isolated from cultured cells and the frozentissues using an RNeasy kit (Qiagen) and a guanidiumisothiocyanate/CsCl method, respectively. RNA was quanti-fied by spectrophotometry at 260 nm. Complementary DNA(cDNA) was prepared from the total RNA of liver tissues,HCC samples, HCC cell lines and HeLa cells with SuperScriptRNaseH�Reverse Transcriptase (BRL). Expressions ofHURP in liver tissues and HCC samples were examinedwith RT–PCR using the forward and reverse primers,50-CCCATCTTCCCTTGAGAAAG-30 and 50-AGGAGA-CATCAAGAACATGC-30. Expression of cyclin B2 wasexamined with real-time RT-PCR using the forward andreverse primers, 50-CATCGACTATGATATGGTGC-30 and50-CAAGGTCTTTGACGGC-30. Expressions of HURP innormal human tissues and synchronized HeLa cells wereanalysed by the TaqMan quantitative RT–PCR reactionaccording to the manufacturer’s protocol (PE AppliedBiosystems). The forward primer, the reverse primer, andthe FAM-labeled probe were 50-AAAGTTGTGCAGCCTG-TAATGC-30, 50-CTGTTCTGGGAACCTGCTTTG-30 and50-CACGTCGTTGAGAATGACTCGATCAGCTAC-30, res-pectively.
Northern analysis of HURP in regenerating mouse liver
The expression patterns of HURP mRNA in regeneratingmouse liver were studied using an RNA blot from regeneratingmouse liver. The hybridization probe was fragment of the
Overexpression of HURP gene in human hepatocellular carcinomaA-P Tsou et al
299
Oncogene

mouse (2014–2486 bp) HURP genes. Probe was labeled with[a-32P]dCTP using the rediprimetII random prime labelingsystem (Amersham-Pharmacia) and the hybridization signalwas quantified with a PhosphImager (Amersham-Pharmacia).Mouse b2-microglobulin, a housekeeping gene, was used as aloading control to normalize the hybridization signals fromregenerating mouse liver RNA (Cressman et al., 1996).
Antibodies
Human HURP cDNA was subcloned into the vector pET32(Novagen) and expressed in Escherichia coli as His-taggedfusion protein. The protein was expressed as an inclusion bodyand solubilized with 2m urea. The solubilized protein waspartially purified by nickel agarose under denaturing condi-tions. The purified protein was then dialyzed to remove thedenaturant as described in the manufacturer’s manual. Therecombinant protein was then injected into mice to raisepolyclonal antibodies, which were used for immunofluores-cence. An antibody to FLAG (M2 from Kodak) was used forthe detection of HURP protein in the stable transfectants.
Indirect immunofluorescence analysis of HURP protein
Cells were trypsinized and seeded onto coverslips. Afterreaching 75% confluence, cells were washed with PBS twiceand then fixed with methanol at �201C for 20min. After air-drying, cells were incubated with polyclonal antibodies againstHURP at room temperature for 2 h. In another experiment,HeLa cells were transfected with HURP-HA for 36 h andimmunofluoresence was performed using HA antibody (3F10from Roche). The coverslips were then washed three times withPBS and incubated with goat–anti-mouse-FITC or Texas Red(Jackson) and 1mg/ml DAPI (Sigma). After washing with PBScontaining 0.1% Tween 20, the samples were mounted. Thefluorescence images on the coverslips were analyzed using aZeiss microscope.
Nucleotide sequence accession numbers
The nucleotide sequence data of human and mouse HURPreported in this paper have been submitted to the DDBJ/EMBL/GenBank nucleotide sequence databases with theAccession AB 076695 and AB 076696, respectively.
Assay for low serum growth
HURP stable clones or 293T cells were seeded in 96-well platesin quadruplicate and cultured with DMEM containing 0, 0.5and 1% fetal bovine serum. After 1–6 days, MTT assay wasused to quantitate cell growth. Briefly, 20ml of 5mm MTT(Sigma) was added and incubated with the cells for 4 h at 371C.After that, the media were removed and 100 ml of DMSO wasadded for 5min at room temperature to extract the colorizedproduct catalyzed by living cells. The end products with purplecolor were quantitated with a spectrophotometer (OD570 nm).The OD values were normalized with the value on day 1.
Polyhema-based anchorage-independent growth (AIG) assay
The AIG assay was modified according to previous reports(Kawada et al., 1997). Briefly, 24-well plates were coated withvarious amounts of polyhema (Sigma) and left to dry at 371Cfor 3 days. The coated plate was irradiated with UV light inlaminar flow before use. HURP stable clones and 293T cellswere seeded in the polyhema-coated plates, and were photo-graphed microscopically after 5 days.
Results
Identification of HURP gene
A human liver EST database containing 142 757 ESTsequences was constructed by collecting EST sequencesgenerated from various human liver cDNA libraries inthe dbEST database. These ESTs were then groupedinto five categories according to the tissue source of thecDNA library: fetal liver/spleen tissues, normal adultliver tissues, HCC-tumor tissues, adjacent nontumoroustissues, and HCC cell lines. A total of 256 UniGenegroups that only were present in cDNA libraries ofhuman HCC tissue were identified. Furthermore, asearch of the gene expression profiles of these 256UniGene groups in the microarray database establishedby Iyer et al. (1999) was carried out. This microarraydatabase contains the expression profile of 8600 genes ofnormal human fibroblasts after serum stimulation. Only32 UniGene groups were found and 12 of them werehighly regulated during serum-induced cell cycle pro-gression. Subsequent examination of the expression ofthese eight ESTs in six pairs of HCC tissues using RT–PCR identified five genes, including N32765, as havingelevated expression in hepatoma tissues relative toadjacent nontumorous tissues (Table 1). Among thesefive genes, N32765 showed the most distinct pattern ofexpression (Figure 1) and we therefore designatedN32765 as HURP.
Table 1 Eight genes, identified from bioinformatics searches, showedupregulated transcription in six pairs of HCC samples.
Clone ID/gene name Level of expression in HCC(ratio of T>N in six
pairs of HCC)
N32765 (HURP) 6/6N69904 5/6AA019977 5/6Carboxypeptidase D 3/6Palmitoyl-protein thioesterase 3/6Amine oxide, copper-containing (AOC3) 3/6Presenilin 1 2/6Shc adaptor homolog 1/6
The expression level of the eight genes in six pairs of HCC samples wasdetected by RT–PCR. T>N means expression of the gene is higher inthe tumor tissue (T) than in adjacent nontumorous tissue (N)
T NT N T N T NT N T NHURP
G3PDH
729 746 747 803 808 815
NL G2 3B HNL-2
Figure 1 Overexpression of HURP in hepatoma. Expression ofHURP was examined by RT–PCR. The human liver tissuesexamined were two normal liver tissues (NL and HNL-2), twohepatoma-derived cell lines (HepG2 and Hep3B) and six HCCsamples. Primary liver tumor tissues (T) and the correspondingadjacent nontumorous liver tissues (N) were obtained from sixpatients (729, 746, 747, 803, 808, and 815). G3PDH served as aninternal control
Overexpression of HURP gene in human hepatocellular carcinomaA-P Tsou et al
300
Oncogene

hHURP
mHURP
1 60
1 59
M S S S H F A S R H I S T E M I R T K I A H R K S L S Q K E N R H K E Y E R N R H F G L K D V N I P T L E G R I LM V R T N L A H R K S L S Q K E N R H R V Y E R N R H F G L K D V N I P - L E G R E L
R K D
hHURP
mHURP
61 120
43 119
V E L D E T S Q E L V P E K T N V K P R A M K T I L G D Q R K Q M L Q K Y K E E K Q L Q K L K E Q R E K A K R G I F K VG N I H E T S Q D L S P E K A S S K T R S V K M V L S D Q R K Q L L Q K Y K E E K Q L Q K L K E Q R E K A K R G V F K V
hHURP
mHURP
121 180
103 179
G R Y R P D M P C F L L S N Q N A V K A E P K K A I P S S V R I T R S K A K D Q M E Q T K I D N E S D V R A I R P G P RG L Y R P A A P G F L V T D Q R G A K A E P E K A F P H T G R I T R S K T K E Y M E Q T K I G S R N V P K A T Q S D Q R
hHURP
mHURP
181 239
163 238
Q T S E K K V S D K E K K V V Q P V M P T S L R M T R - S A T Q A A K Q V P R T V S S T T A R K P V T R A A N E N E P EQ T S E K Q P L D R E R K V M Q P V L F T S G K G T E S A A T Q R A K L M A R T V S S T T - R K P V T R A T N E K G S E
hHURP
mHURP
240 299
222 298
G K V P S K G R P A K N V E T K P D K G I S C K V D S E E N T L N S Q T N A T S G M N P D G V L S K M E N L P E I N T AR M R P S G G R P A K K P E G K P D K V I P S K V E R D E K H L D S Q T R E T S E M G L L G V F R E V E S L P A T A P A
hHURP
mHURP
300 359
282 351
K I K G K N S F A P K D F M F Q P L D G L K T Y Q V T P M T P R S A N A F L T P S Y T WT P L K T E V D E S Q A T K E IQ G K E R K S F A P K H C V F Q P P C G L K S Y Q V A P L S P R S A N A F L T P N C D WN Q L R P E V F S - - - - - - -
hHURP
mHURP
360 419
335 403
L A Q K C K T Y S T K T I Q Q D S N K L P C P L G P L T V WH E E H V L N K N E A T T K N L N G L P I K E V P S L E R N- - - - - - - - T T T Q D K A N E I L V Q Q G L E S L T D R S K E H V L N Q K G A S T S D S N H A S V K G V P C S E G S
hHURP
mHURP
420 479
387 463
E G R I A Q P H H G V P Y F R N I L Q S E T E K L T S H C F E WD R K L E L D I P D D A K D L I R T A V G Q T R L L M KE G Q T S Q P P H D V P Y F R K I L Q S E T D R L T S H C L E WE G K L D L D I S D E A K G L I R T T V G Q T R L L I K
hHURP
mHURP
480 539
447 523
E R F K Q F E G L V D D C E Y K R G I K E T T C T D L D G F WD M V S F Q I E D V I H K F N N L I K L E E S G WQ V N NE R F R Q F E G L V D N C E Y K R G E K E T T C T D L D G F WD M V S F Q V D D V N Q K F N N L I K L E A S G WK D S N
hHURP
mHURP
540 599
507 579
N M N H N M N K N V F R K K V V S G I A S K P K Q D D A G R I A A R N R L A A I K N A M R E R I R Q E E C A E T A V S VN P - - - - S K K V L R K K I V P G R T S K A K Q D D D G R A A A R S R L A A I K N A M K G R P Q Q E V Q A H A A A P E
hHURP
mHURP
600 659
563 637
I P K E V D K I V F D A G F F R V E S P V K L F S G L S V S S E G P S Q R L G T P K S V N K A V S Q S R N E M G I P Q QT T K E V D K I V F D A G F F R I E S P V K S F S V L S S - - E R R S Q R F G T P L S A S K V V P E G R A A G D L L R Q
hHURP
mHURP
660 719
621 696
T T S P E N A G P Q N T K S E H V K K T L F L S I P E S R S S I E D A Q C P G L P D L I E E N H V V N K T D L K V D C LK M P L K K P D P Q S S K S E H V D R T F S D G L - E S R C H V E D T P C P G E Q D S S D I E H D V N K I N V K M D C F
hHURP
mHURP
720 779
680 748
S S E R M S L P L L A G G V A D D I N T N K K E G I S D V V E G M E L N S S I T S Q D V L M S S P E K N T A S Q N S I LS V E - T N L P L P A G - - - - D A N T N Q K E A I S A V E G - - - A S T A V T S Q D L L M S N P E T N T S S Q S N T S
hHURP
mHURP
780 839
732 806
E E G E T K I S Q S E L F D N K S L T T E C H L L D S P G L N C S N P F T Q L E R R H Q E H A R H I S F G G N L I T F SQ E - E A E A S Q S V L L H - K S L T S E C H L L E P P G L S C T S P C T R E E T R Q P D R S R Q F S F G G D L I L F S
hHURP
mHURP
840 846
790 808
P L Q P G E FP L
D-box KEN
Coiled-coil
NES
GH1
D-box
NLS
M L V S R F A S R F R KD S S T ER KDS F A S RHM
a
Figure 2 Structure of HURP gene and cDNAs. (a) The alignment of the deduced amino-acid sequences of human and mouse HURPis shown in a single-letter code. Identical residues or conservative replacements are boxed and dashes denote gaps in the alignment. Theopen boxes denote the motifs as indicated. The human and mouse HURP genes encode proteins of 846 and 808 amino acids,respectively. The putative motifs of the leucine-rich nuclear export signal (NES), the D-boxes, the KEN box, and a coiled-coil domainare indicated. (b) Organization of the human HURP gene on chromosome 14q22–23. In total, 19 exons are shown in boxes. Closedboxes designate the coding sequences. ATG is the translation start site and TGA is the translation stop site
Overexpression of HURP gene in human hepatocellular carcinomaA-P Tsou et al
301
Oncogene
Cloning of HURP cDNA of human and mouse origins
In order to examine the function of HURP, human andmurine HURP cDNAs were isolated. Cloning of a full-length HURP cDNA of human and mouse origin wasaccomplished by assembling the EST clones usingSequencher software, PCR cloning, cDNA libraryscreening, and 50 RACE. The human and mouse HURPcDNAs encoded 846 and 808 amino acids, respectively.They share 72% identity at the nucleotide sequence leveland 61% identity at the amino-acid sequence level(Figure 2a). The human HURP maps to chromosome14q22–23 (Figure 2b). Comparison of the cDNA andgenomic sequence of chromosome 14 showed thatHURP is organized into 19 exons displaying noncano-nical intron/exon and exon/intron borders. Motifanalysis revealed that HURP contains a putativeleucine-rich nuclear export signal (NES), two destruc-tion boxes (D-box), a KEN box, and a coiled-coildomain. In addition, a stretch of about 100 amino acids(432–537 in human HURP), termed the GH1 domain(Kim et al., 1997), showed significant sequence similarityto GKAP (guanylate kinase associated protein), Cae-norhabditis elegans U00058, and Drosophila Vulcan(Figure 2a).
Expression of HURP in normal human tissues
In order to determine whether HURP expressionexhibits any tissue specificity, quantitative RT–PCR
was used to examine a wide spectrum of human tissues.Among the tissues examined, the most abundantexpression of HURP transcript was detected in the fetalliver followed by bone marrow, testis, colon, and theplacenta (Figure 3). Interestingly, normal liver tissueshowed very little expression of the HURP transcript.
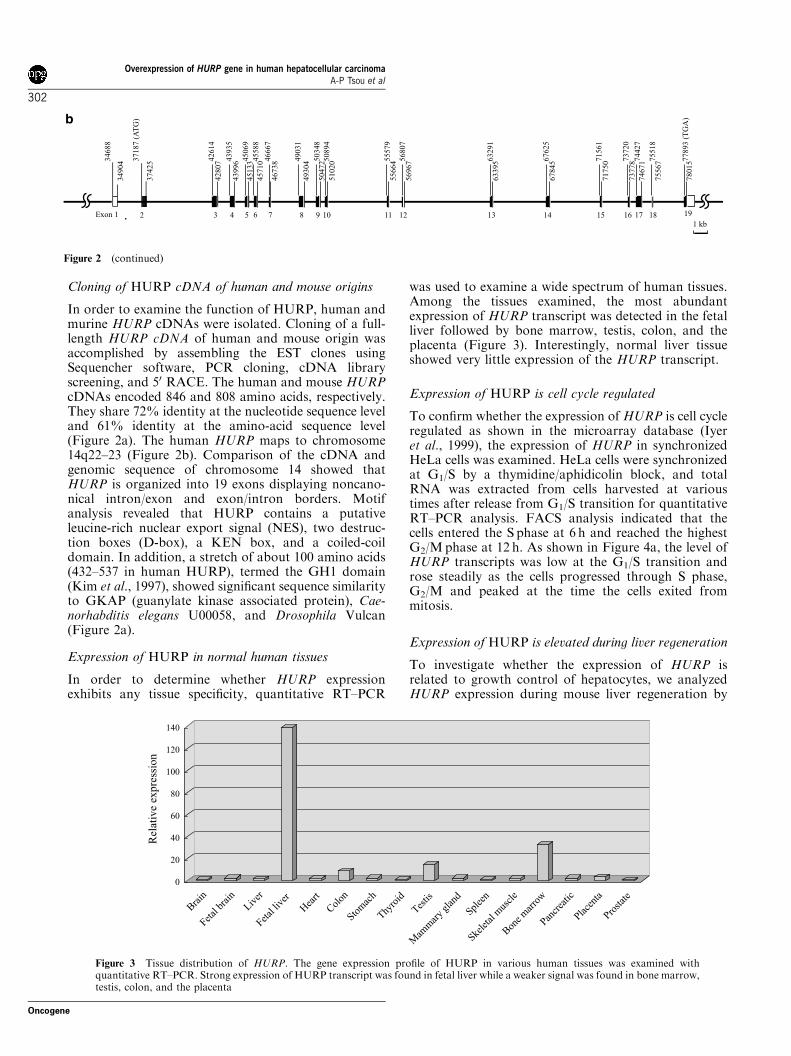
Expression of HURP is cell cycle regulated
To confirm whether the expression of HURP is cell cycleregulated as shown in the microarray database (Iyeret al., 1999), the expression of HURP in synchronizedHeLa cells was examined. HeLa cells were synchronizedat G1/S by a thymidine/aphidicolin block, and totalRNA was extracted from cells harvested at varioustimes after release from G1/S transition for quantitativeRT–PCR analysis. FACS analysis indicated that thecells entered the S phase at 6 h and reached the highestG2/Mphase at 12 h. As shown in Figure 4a, the level ofHURP transcripts was low at the G1/S transition androse steadily as the cells progressed through S phase,G2/M and peaked at the time the cells exited frommitosis.
Expression of HURP is elevated during liver regeneration
To investigate whether the expression of HURP isrelated to growth control of hepatocytes, we analyzedHURP expression during mouse liver regeneration by
Exon 1 19
37187 (
AT
G)
374253
4688
34904 4
2614
42807 4
3935
43996 4
5588
457104
5069
45133 4
6667
46738 4
9031
49304 5
0348
50472 5
0894
51020 5
5579
55664 5
6807
56967 6
3291
63395 6
7625
67845 7
1561
71750 7
3720
73778 7
4427
74671 7
5518
75567 7
7893 (
TG
A)
78015
b
Figure 2 (continued)
Rel
ativ
e ex
pre
ssio
n
Stom
ach
Thyro
id
Testis
Splee
n
Mam
mar
y gl
and
Skele
tal m
uscl
e
Bon
e m
arro
w
Pancr
eatic
Place
nta
Prostat
e
Bra
in
Fetal
bra
in
Liver
Fetal
live
r
Hea
rt
Col
on
140
120
100
80
60
40
20
0
Figure 3 Tissue distribution of HURP. The gene expression profile of HURP in various human tissues was examined withquantitative RT–PCR. Strong expression of HURP transcript was found in fetal liver while a weaker signal was found in bone marrow,testis, colon, and the placenta
Overexpression of HURP gene in human hepatocellular carcinomaA-P Tsou et al
302
Oncogene
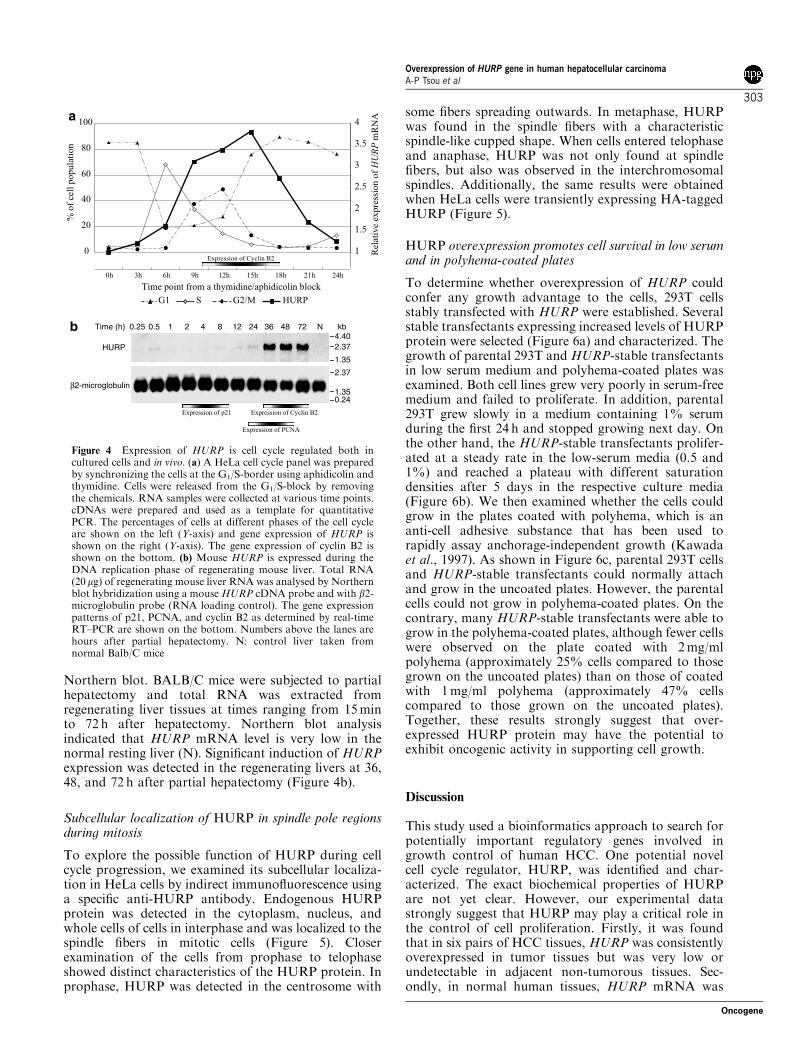
Northern blot. BALB/C mice were subjected to partialhepatectomy and total RNA was extracted fromregenerating liver tissues at times ranging from 15minto 72 h after hepatectomy. Northern blot analysisindicated that HURP mRNA level is very low in thenormal resting liver (N). Significant induction of HURPexpression was detected in the regenerating livers at 36,48, and 72 h after partial hepatectomy (Figure 4b).
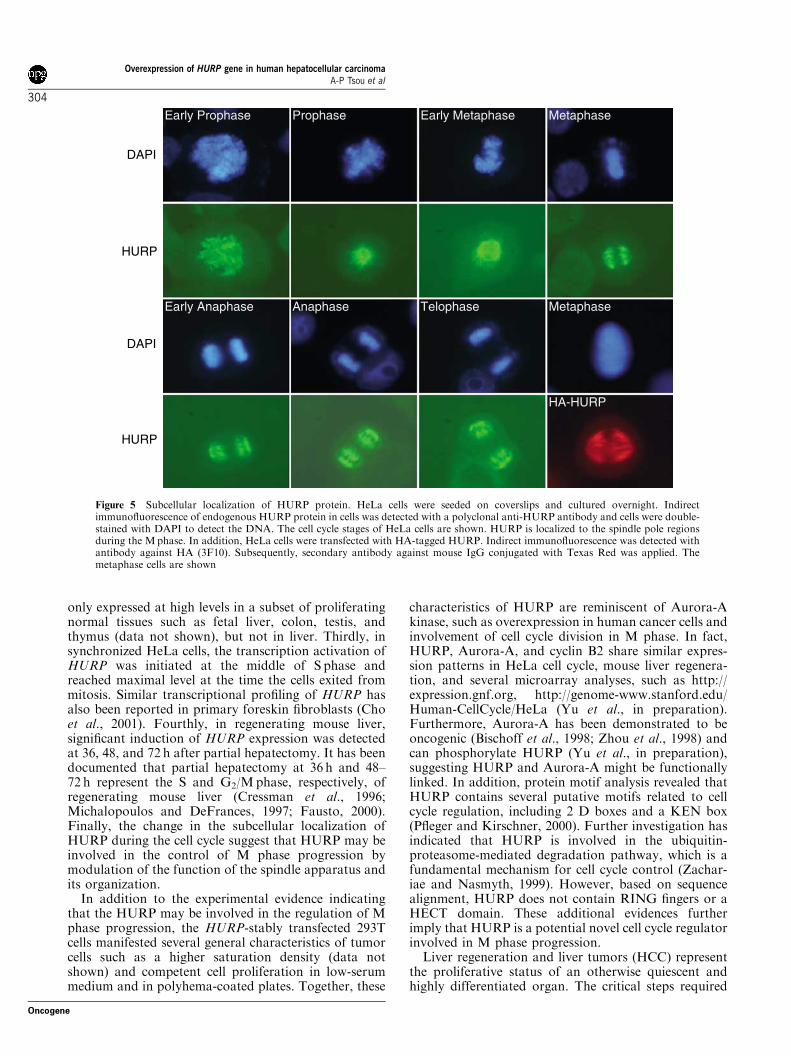
Subcellular localization of HURP in spindle pole regionsduring mitosis
To explore the possible function of HURP during cellcycle progression, we examined its subcellular localiza-tion in HeLa cells by indirect immunofluorescence usinga specific anti-HURP antibody. Endogenous HURPprotein was detected in the cytoplasm, nucleus, andwhole cells of cells in interphase and was localized to thespindle fibers in mitotic cells (Figure 5). Closerexamination of the cells from prophase to telophaseshowed distinct characteristics of the HURP protein. Inprophase, HURP was detected in the centrosome with
some fibers spreading outwards. In metaphase, HURPwas found in the spindle fibers with a characteristicspindle-like cupped shape. When cells entered telophaseand anaphase, HURP was not only found at spindlefibers, but also was observed in the interchromosomalspindles. Additionally, the same results were obtainedwhen HeLa cells were transiently expressing HA-taggedHURP (Figure 5).
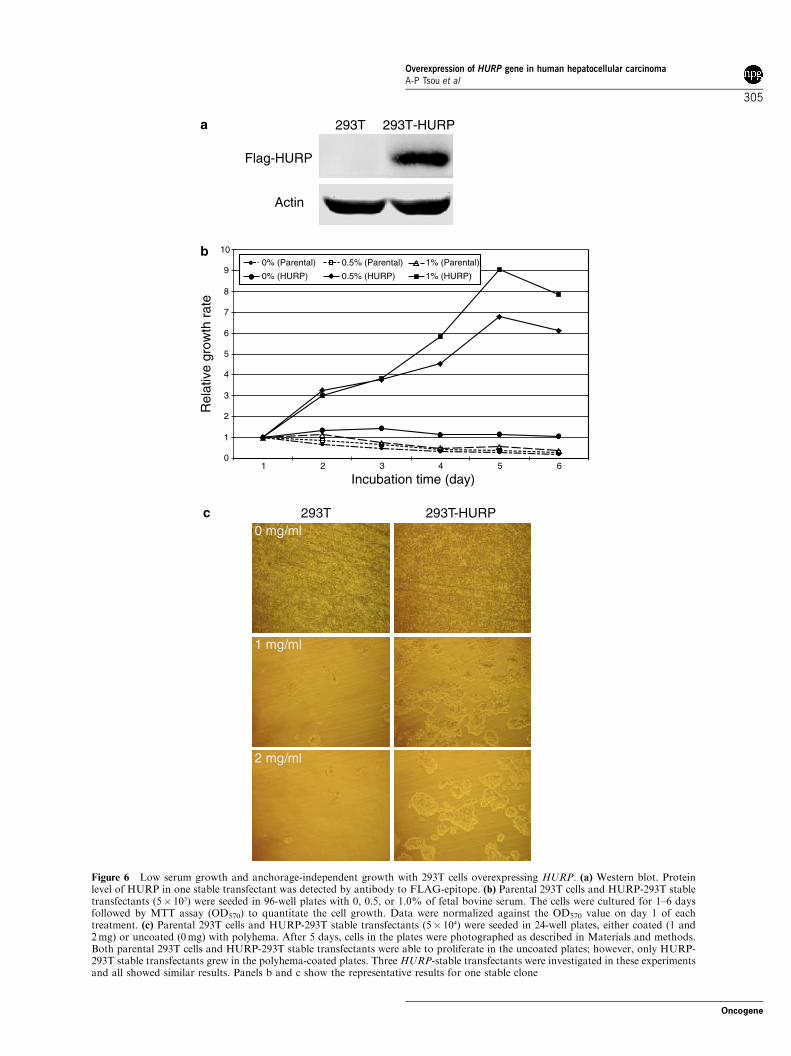
HURP overexpression promotes cell survival in low serumand in polyhema-coated plates
To determine whether overexpression of HURP couldconfer any growth advantage to the cells, 293T cellsstably transfected with HURP were established. Severalstable transfectants expressing increased levels of HURPprotein were selected (Figure 6a) and characterized. Thegrowth of parental 293T and HURP-stable transfectantsin low serum medium and polyhema-coated plates wasexamined. Both cell lines grew very poorly in serum-freemedium and failed to proliferate. In addition, parental293T grew slowly in a medium containing 1% serumduring the first 24 h and stopped growing next day. Onthe other hand, the HURP-stable transfectants prolifer-ated at a steady rate in the low-serum media (0.5 and1%) and reached a plateau with different saturationdensities after 5 days in the respective culture media(Figure 6b). We then examined whether the cells couldgrow in the plates coated with polyhema, which is ananti-cell adhesive substance that has been used torapidly assay anchorage-independent growth (Kawadaet al., 1997). As shown in Figure 6c, parental 293T cellsand HURP-stable transfectants could normally attachand grow in the uncoated plates. However, the parentalcells could not grow in polyhema-coated plates. On thecontrary, many HURP-stable transfectants were able togrow in the polyhema-coated plates, although fewer cellswere observed on the plate coated with 2mg/mlpolyhema (approximately 25% cells compared to thosegrown on the uncoated plates) than on those of coatedwith 1mg/ml polyhema (approximately 47% cellscompared to those grown on the uncoated plates).Together, these results strongly suggest that over-expressed HURP protein may have the potential toexhibit oncogenic activity in supporting cell growth.
Discussion
This study used a bioinformatics approach to search forpotentially important regulatory genes involved ingrowth control of human HCC. One potential novelcell cycle regulator, HURP, was identified and char-acterized. The exact biochemical properties of HURPare not yet clear. However, our experimental datastrongly suggest that HURP may play a critical role inthe control of cell proliferation. Firstly, it was foundthat in six pairs of HCC tissues, HURP was consistentlyoverexpressed in tumor tissues but was very low orundetectable in adjacent non-tumorous tissues. Sec-ondly, in normal human tissues, HURP mRNA was
% o
f ce
ll p
op
ula
tio
n
Rel
ativ
e ex
pre
ssio
n o
f HURP
mR
NA
0
20
40
60
80
100
0h 3h 6h 9h 12h 15h 18h 21h 24h
1
1.5
2
2.5
3
3.5
4
HURPG2/MSG1
Time point from a thymidine/aphidicolin block
a
Expression of Cyclin B2
4.402.37
1.35
724836241284210.50.25 N
2.37
1.350.24
kbTime (h)
HURP
β2-microglobulin
b
Expression of p21 Expression of Cyclin B2
Expression of PCNA
Figure 4 Expression of HURP is cell cycle regulated both incultured cells and in vivo. (a) A HeLa cell cycle panel was preparedby synchronizing the cells at the G1/S-border using aphidicolin andthymidine. Cells were released from the G1/S-block by removingthe chemicals. RNA samples were collected at various time points.cDNAs were prepared and used as a template for quantitativePCR. The percentages of cells at different phases of the cell cycleare shown on the left (Y-axis) and gene expression of HURP isshown on the right (Y-axis). The gene expression of cyclin B2 isshown on the bottom. (b) Mouse HURP is expressed during theDNA replication phase of regenerating mouse liver. Total RNA(20mg) of regenerating mouse liver RNA was analysed by Northernblot hybridization using a mouse HURP cDNA probe and with b2-microglobulin probe (RNA loading control). The gene expressionpatterns of p21, PCNA, and cyclin B2 as determined by real-timeRT–PCR are shown on the bottom. Numbers above the lanes arehours after partial hepatectomy. N: control liver taken fromnormal Balb/C mice
Overexpression of HURP gene in human hepatocellular carcinomaA-P Tsou et al
303
Oncogene
only expressed at high levels in a subset of proliferatingnormal tissues such as fetal liver, colon, testis, andthymus (data not shown), but not in liver. Thirdly, insynchronized HeLa cells, the transcription activation ofHURP was initiated at the middle of S phase andreached maximal level at the time the cells exited frommitosis. Similar transcriptional profiling of HURP hasalso been reported in primary foreskin fibroblasts (Choet al., 2001). Fourthly, in regenerating mouse liver,significant induction of HURP expression was detectedat 36, 48, and 72 h after partial hepatectomy. It has beendocumented that partial hepatectomy at 36 h and 48–72 h represent the S and G2/Mphase, respectively, ofregenerating mouse liver (Cressman et al., 1996;Michalopoulos and DeFrances, 1997; Fausto, 2000).Finally, the change in the subcellular localization ofHURP during the cell cycle suggest that HURP may beinvolved in the control of M phase progression bymodulation of the function of the spindle apparatus andits organization.
In addition to the experimental evidence indicatingthat the HURP may be involved in the regulation of Mphase progression, the HURP-stably transfected 293Tcells manifested several general characteristics of tumorcells such as a higher saturation density (data notshown) and competent cell proliferation in low-serummedium and in polyhema-coated plates. Together, these
characteristics of HURP are reminiscent of Aurora-Akinase, such as overexpression in human cancer cells andinvolvement of cell cycle division in M phase. In fact,HURP, Aurora-A, and cyclin B2 share similar expres-sion patterns in HeLa cell cycle, mouse liver regenera-tion, and several microarray analyses, such as http://expression.gnf.org, http://genome-www.stanford.edu/Human-CellCycle/HeLa (Yu et al., in preparation).Furthermore, Aurora-A has been demonstrated to beoncogenic (Bischoff et al., 1998; Zhou et al., 1998) andcan phosphorylate HURP (Yu et al., in preparation),suggesting HURP and Aurora-A might be functionallylinked. In addition, protein motif analysis revealed thatHURP contains several putative motifs related to cellcycle regulation, including 2 D boxes and a KEN box(Pfleger and Kirschner, 2000). Further investigation hasindicated that HURP is involved in the ubiquitin-proteasome-mediated degradation pathway, which is afundamental mechanism for cell cycle control (Zachar-iae and Nasmyth, 1999). However, based on sequencealignment, HURP does not contain RING fingers or aHECT domain. These additional evidences furtherimply that HURP is a potential novel cell cycle regulatorinvolved in M phase progression.
Liver regeneration and liver tumors (HCC) representthe proliferative status of an otherwise quiescent andhighly differentiated organ. The critical steps required
DAPI
HURP
DAPI
HURP
Early Prophase Prophase Early Metaphase Metaphase
Early Anaphase Anaphase Telophase Metaphase
HA-HURP
Figure 5 Subcellular localization of HURP protein. HeLa cells were seeded on coverslips and cultured overnight. Indirectimmunofluorescence of endogenous HURP protein in cells was detected with a polyclonal anti-HURP antibody and cells were double-stained with DAPI to detect the DNA. The cell cycle stages of HeLa cells are shown. HURP is localized to the spindle pole regionsduring the Mphase. In addition, HeLa cells were transfected with HA-tagged HURP. Indirect immunofluorescence was detected withantibody against HA (3F10). Subsequently, secondary antibody against mouse IgG conjugated with Texas Red was applied. Themetaphase cells are shown
Overexpression of HURP gene in human hepatocellular carcinomaA-P Tsou et al
304
Oncogene
Incubation time (day)
Rel
ativ
e gr
owth
rat
e
0
1
2
3
4
5
6
7
8
9
10
1 2 3 4 5 6
0% (Parental) 0.5% (Parental)
0.5% (HURP)
1% (Parental)
1% (HURP)0% (HURP)
a
293T 293T-HURP0 mg/ml
1 mg/ml
2 mg/ml
b
c
293T 293T-HURP
Flag-HURP
Actin
Figure 6 Low serum growth and anchorage-independent growth with 293T cells overexpressing HURP. (a) Western blot. Proteinlevel of HURP in one stable transfectant was detected by antibody to FLAG-epitope. (b) Parental 293T cells and HURP-293T stabletransfectants (5� 103) were seeded in 96-well plates with 0, 0.5, or 1.0% of fetal bovine serum. The cells were cultured for 1–6 daysfollowed by MTT assay (OD570) to quantitate the cell growth. Data were normalized against the OD570 value on day 1 of eachtreatment. (c) Parental 293T cells and HURP-293T stable transfectants (5� 104) were seeded in 24-well plates, either coated (1 and2mg) or uncoated (0mg) with polyhema. After 5 days, cells in the plates were photographed as described in Materials and methods.Both parental 293T cells and HURP-293T stable transfectants were able to proliferate in the uncoated plates; however, only HURP-293T stable transfectants grew in the polyhema-coated plates. Three HURP-stable transfectants were investigated in these experimentsand all showed similar results. Panels b and c show the representative results for one stable clone
Overexpression of HURP gene in human hepatocellular carcinomaA-P Tsou et al
305
Oncogene

for the transition from regulated to constitutivehepatocyte proliferation may set the stage for hepato-carcinogenesis. Various growth factors such as EGF,HGF, and IGF-1 have been shown to play an importantrole in proliferation of regenerating rodent hepatocytes(Michalopoulos and DeFrances, 1997; Fausto, 2000).Recently, accumulated evidence has also demonstratedthat modulation of cell cycle control genes downstreamof growth factors can affect hepatocyte proliferation(Menjo et al., 1998). Cyclin D1, in the absence of growthfactors, was sufficient to promote cell cycle progressionin cultured primary rat hepatocytes (Albrecht andHansen, 1999; Hansen and Albrecht, 1999). Moreover,cotransfection of cyclin E and skp2 synergisticallypromoted cell cycle progression in cultured hepatocytesand induced liver hyperplasia in BALB/C mice (Nelsenet al., 2001). In contrast, the blocking of cell cycleprogression with p21 overexpression in transgenic miceinhibited hepatocyte proliferation and liver regeneration(Wu et al., 1996). Most of these studies, however, havefocused on the regulation of G1 phase whereas regula-tion of M phase progression has not been extensivelyinvestigated in regenerating hepatocytes. The biologicalactivity of HURP presented here provides a new line ofevidence that supports the likelihood that highlyregulated compensatory growth can turn into uncon-trolled hyperplasia by loss of cell cycle regulation duringthe M phase.
In conclusion, using a combination of databasessearches and empirical data, we have identified apotential novel cell cycle regulator, HURP, whoseexpression is up-regulated during the M phase of thecell cycle. In addition, cells stably transfected withHURP appear to have the characteristics of tumor cellswith a reduced dependence upon extracellular growth
factors and the potential to exhibit anchorage-indepen-dent growth. Thus, overexpression of HURP in HCCmay represent a consequence of deregulation, which canlead to cancer by allowing cells to progress through thecell cycle. Inhibition of the expression or the activity ofHURP could theoretically revert cancer cells to a morenormal phenotype.
Note added in proof
During the preparation of this manuscript, a novelprotein KIAA0008 was reported (Bassal et al., 2001).KIAA0008 lacks the first two exons when compared tothe HURP gene sequence on chromosome 14 andhence resulted in a product shorter by 81 amino-acidresidues at the N-terminus of the coding region(Figure 2a).
Acknowledgments
We thank Mien-Chie Hung (The University of Texas MDAnderson Cancer Center) for critical reading of the manuscriptand Kuo-Ji Chen, Yu-Lun Liao, Min-Pey Ding, and Jung-Mao Hsu for their excellent technical assistance. Wealso thank Dr Anna F-Y Li for her assistance in patho-logical reading. This work was supported in part by grantsfrom ADImmune Corporation, Program for PromotingAcademic Excellence of Universities (89-B-FA22-2-4)and National Health Research Institutes (NHRI-EX90-9001-BL) to APT, MJF, and CKC, grant from the NationalHealth Research Institutes to CFH, and grant from NationalScience Counsel (NSC-91-2323-B075-001) and MolecularPathology core laboratory (National Yang Ming University)to CWC.
References
Albrecht JH and Hansen LK. (1999). Cell Growth Differ., 10,397–404.
Bassal S, Nomura N, Venter D, Brand K, McKay MJ and vander Spek PJ. (2001). Genomics, 77, 5–7.
Bischoff JR, Anderson L, Zhu Y, Mossie K, Ng L, Souza B,Schryver B, Flanagan P, Clairvoyant F, Ginther C, ChanCS, Novotny M, Slamon DJ and Plowman GD. (1998).Embo. J., 17, 3052–3065.
Bosch FX, Ribes J and Borras J. (1999). Semin. Liver. Dis., 19,271–285.
Boyer N and Marcellin P. (2000). J. Hepatol., 32, 98–112.Cahill DP, Lengauer C, Yu J, Riggins GJ, Willson JK,Markowitz SD, Kinzler KW and Vogelstein B. (1998).Nature, 392, 300–303.
Cho RJ, Huang M, Campbell MJ, Dong H, Steinmetz L,Sapinoso L, Hampton G, Elledge SJ, Davis RW andLockhart DJ. (2001). Nat. Genet., 27, 48–54.
Cressman DE, Greenbaum LE, DeAngelis RA, Ciliberto G,Furth EE, Poli V and Taub R. (1996). Science, 274,1379–1383.
Fausto N. (2000). J. Hepatol., 32, 19–31.Feitelson MA and Duan LX. (1997). Am. J. Pathol., 150,1141–1157.
Ferea TL and Brown PO. (1999). Curr. Opin. Genet. Dev., 9,715–722.
Hansen LK and Albrecht JH. (1999). J. Cell. Sci., 112, 2971–2981.
Higgins GM and Anderson RM. (1931). Arch. Pathol., 12,
186–202.Iyer VR, Eisen MB, Ross DT, Schuler G, Moore T, Lee JC,Trent JM, Staudt LM, Hudson Jr J, Boguski MS, LashkariD, Shalon D, Botstein D and Brown PO. (1999). Science,283, 83–87.
Kawada M, Fukazawa H, Mizuno S and Uehara Y. (1997).Biochem. Biophys. Res. Commun., 231, 735–737.
Kim E, Naisbitt S, Hsueh YP, Rao A, Rothschild A, CraigAM, Sheng M. (1997). J Cell Biol, 136, 669–678
Menjo M, Ikeda K and Nakanishi M. (1998). J. Gastroenterol.Hepatol., 13(Suppl), S100–S105.
Michalopoulos GK and DeFrances MC. (1997). Science, 276,60–66.
Nelsen CJ, Hansen LK, Rickheim DG, Chen C, Stanley MW,Krek W and Albrecht JH. (2001). Oncogene, 20, 1825–1831.
Okabe H, Satoh S, Kato T, Kitahara O, Yanagawa R,Yamaoka Y, Tsunoda T, Furukawa Y and Nakamura Y.(2001). Cancer Res., 61, 2129–2137.
Overexpression of HURP gene in human hepatocellular carcinomaA-P Tsou et al
306
Oncogene

Okuda K. (2000). J. Hepatol., 32, 225–237.Pfleger CM and Kirschner MW. (2000). Genes Dev., 14,655–665.
Su Q, Liu YF, Zhang JF, Zhang SX, Li DF and Yang JJ.(1994). Hepatology, 20, 788–799.
Takahashi T, Haruki N, Nomoto S, Masuda A, Saji S andOsada H. (1999). Oncogene, 18, 4295–4300.
Tsou AP, Wu KM, Tsen TY, Chi CW, Chiu JH, Lui WY, HuCP, Chang C, Chou CK and Tsai SF. (1998). Genomics, 50,331–340.
Tsukasaki K, Miller CW, Greenspun E, Eshaghian S,Kawabata H, Fujimoto T, Tomonaga M, Sawyers C,Said JW and Koeffler HP. (2001). Oncogene, 20,
3301–3305.Wu H, Wade M, Krall L, Grisham J, Xiong Y and Van DykeT. (1996). Genes Dev., 10, 245–260.
Zachariae W and Nasmyth K. (1999). Genes Dev., 13,
2039–2058.Zhou H, Kuang J, Zhong L, Kuo WL, Gray JW, Sahin A,Brinkley BR and Sen S. (1998). Nat. Genet., 20, 189–193.
Overexpression of HURP gene in human hepatocellular carcinomaA-P Tsou et al
307
Oncogene
Related Documents











