J Euk. Microbiol.. 43(1), 1996. pp. 26-31 0 1996 by the Society of Prolozoologirls Identification of a Microsporidian Polar Tube Protein Reactive Monoclonal Antibody ELAINE M. KEOHANE,* PETER M. TAKVORIAN,* ANN CALI,* HERBERT B. TANOWITZ,**~*** MURRAY WITTNER** and LOUIS M. WEISS**,***' *Department of Biological Sciences. Rutgers University, Newark, New Jersey 071 02, and **Department of Pathology. Division of Parasitology, and *** Department of Medicine, Division of Infectious Diseases, Albert Einstein College of Medicine, Bronx, New York. 10461 ABSTRACT. The microsporidia are characterized by spores containing a single polar tube that coils around the sporoplasm. When triggered by appropriate stimuli, the polar tube rapidly discharges out of the spore forming a hollow tube. The sporoplasm passes out of the spore through this tube serving as a unique vehicle of infection. Due to the unusual hnctional and solubility properties of the polar tube, the proteins comprising it are likely to be members of a protein family with a highly conserved amino acid composition among the various microsporidia. Polar tube proteins were separated from the majority of other proteins in glass bead disrupted spores of Glugea americanus using sequential 1 % sodium dodecyl sulfate (SDS) and 9M urea extractions. The resultant spore pellet demonstrated broken, empty spore coats and numerous polar tubes in straight and twisted formations by negative stain transmission electron microscopy. After subsequent incubation of the pellet with 2% dithiothreitol(DTT), empty spore coats were still observed but the polar tubes were no longer present in the pellet. The DTT supernatant demonstrated four major protein bands by SDS-PAGE 23, 27, 34 and 43 kDa. Monoclonal antibodies were produced to these proteins using Hunter's Titermax adjuvant. Mab 3C8.23.1 which cross- reacted with a 43-kDa antigen by immunoblot analyis, demonstrated strong reactivity with the polar tube of G. americanus spores by immunogold electron microscopy. This antibody will be useful in further characterization of polar tube proteins and may lead to novel diagnostic and therapeutic reagents. Supplementary key words. Glugea arnericanus, immunogold electron microscopy, Microspora, microsporidia, polar filament. ICROSPORIDIA are protozoa classified in the phylum M Microspora that are eukaryotic, obligate intracellular, spore-forming parasites. They are ubiquitous in the animal king- dom with hundreds of species capable of parasitizing a wide range of invertebrate and vertebrate hosts, including humans [2, 41. Five microsporidian genera have been associated with human disease: Nosema. generally found in insects associated with keratitis; Pleistophora, a pathogen of fish and insects as- sociated with myositis; Encephalitoroon, found in many mam- mals associated with disseminated disease, keratitis and sinus- itis; Enterocytozoon associated with diarrhea and reported from AIDS patients and several species of fish; and Septata reported from AIDS patients and associated with diarrhea and dissem- inated disease [l-6, 11, 13, 151. The small spores (1-12 pm) produced by microsporidia are characteristic in that they con- tain a single polar tube (also called the polar filament [ 191) that coils around the sporoplasm and is attached to the inside of the spore by an anchoring disc [2,4]. When triggered by appropriate stimuli, the polar tube rapidly discharges from the anterior pole of the spore forming a hollow tube that remains attached to the spore [8, 9, 171. After complete discharge of the polar tube, the sporoplasm flows through the tube and appears as a droplet at its distal end [8, 9, 171. This process serves as an unique mech- anism of infection in that the polar tube can pierce an adjacent cell resulting in sporoplasm transfer directly into the host cell cytoplasm [9, 171. Polar tubes range from 50 to 100 pm in length and 0.1 to 0.1 5 pm in diameter [8, 181. The polar tube has some flexibility in that it shows variation in diameter from 0.1 to 0.25 pm during discharge, its diameter can increase to 0.4 pm during sporoplasm passage, and its length shortens by 5%10% after sporoplasm passage [8, 12, 17, 181. Inside the spore the polar tube is filled with dense material (believed to be unassembled polar tube protein) and is often referred to as the polar filament, while the discharged tube appears as a hollow cylinder and is usually referred to as the polar tube [ 18, 191. Incompletely discharged tubes appear as a cylinder within a cylinder at their distal ends [ 191. Discharged tubes appear to be two to three times as long ' To whom correspondence should be addressed. as the coiled tube inside the spore, and it has been suggested that the internal contents of the tube are incorporated at its growing tip during discharge [8, 191. This theory is supported by observations using pulse labelling with latex particles [ 191 and by video enhanced contrast microscopy [8]. The polar tube has unusual solubility properties in that it resists dissociation in 1% to 3% sodium dodecyl sulfate (SDS), 1% Triton X-100, 10% H,02, 8N H,SO,, 2N HCl, chloroform, 1% guanidine HCI, 0.1 M proteinase K and 10 M urea [17-191. The polar tube, however, dissociates in various concentrations of 2-mercapto- ethanol (2-ME) or 1% dithiothreitol (DTT) [18, 191. Weidner reported isolation of a polar tube protein from the microspo- ridian, Ameson michaelis, by treatment of discharged spores with 3% SDS which removed sporoplasm and soluble spore coat proteins, but left the resistant spore coats with attached polar tubes intact [ 181. Subsequent treatment with 1% DTT or 50% 2-ME solubilized the polar tubes, but not the spore coat, into the supernatant. SDS-polyacrylamide gel electrophoresis (SDS-PAGE) of the DTT supernatant showed a single 23-kDa protein, presumably representing polar tube protein [ 181. In the present study, a monoclonal antibody was produced to a polar tube protein (PTP) of a microsporidium, which lo- calized to the polar tube by immunogold electron microscopy. The presence of a single polar tube is a defining characteristic of microsporidia, and study of PTP may suggest novel thera- peutic approaches to human infections or result in new diag- nostic reagents. MATERIALS AND METHODS Isolation of polar tube protein. Spores of Glugea americanus (formerly Spraguea lophii) were obtained from cysts found in the cranial and spinal ganglia of the angler fish, Lophius amer- icanus, and were stored at 4" C in sea water [14]. Polar tube proteins were separated from other proteins using a modification of a published protocol [ 181. A 3 x lo7 spore suspension in 1% SDS was disrupted with 0.5 p acid-washed glass beads (Sigma, St. Louis, MO) for 4 min on a Mini Beadbeater (Biospec Prod- ucts, Bartlesville, OK). The disrupted spores were centrifuged and the pellet extracted five times with 1% SDS and then with 9 M urea to remove sporoplasm released from broken spores and soluble spore coat proteins. After centrifugation, an aliquot 26

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

J Euk. Microbiol.. 43(1), 1996. pp. 26-31 0 1996 by the Society of Prolozoologirls
Identification of a Microsporidian Polar Tube Protein Reactive Monoclonal Antibody
ELAINE M. KEOHANE,* PETER M. TAKVORIAN,* ANN CALI,* HERBERT B. TANOWITZ,**~*** MURRAY WITTNER** and LOUIS M. WEISS**,***'
*Department of Biological Sciences. Rutgers University, Newark, New Jersey 071 02, and **Department of Pathology. Division of Parasitology, and *** Department of Medicine, Division of Infectious Diseases,
Albert Einstein College of Medicine, Bronx, New York. 10461
ABSTRACT. The microsporidia are characterized by spores containing a single polar tube that coils around the sporoplasm. When triggered by appropriate stimuli, the polar tube rapidly discharges out of the spore forming a hollow tube. The sporoplasm passes out of the spore through this tube serving as a unique vehicle of infection. Due to the unusual hnctional and solubility properties of the polar tube, the proteins comprising it are likely to be members of a protein family with a highly conserved amino acid composition among the various microsporidia. Polar tube proteins were separated from the majority of other proteins in glass bead disrupted spores of Glugea americanus using sequential 1 % sodium dodecyl sulfate (SDS) and 9M urea extractions. The resultant spore pellet demonstrated broken, empty spore coats and numerous polar tubes in straight and twisted formations by negative stain transmission electron microscopy. After subsequent incubation of the pellet with 2% dithiothreitol (DTT), empty spore coats were still observed but the polar tubes were no longer present in the pellet. The DTT supernatant demonstrated four major protein bands by SDS-PAGE 23, 27, 34 and 43 kDa. Monoclonal antibodies were produced to these proteins using Hunter's Titermax adjuvant. Mab 3C8.23.1 which cross- reacted with a 43-kDa antigen by immunoblot analyis, demonstrated strong reactivity with the polar tube of G. americanus spores by immunogold electron microscopy. This antibody will be useful in further characterization of polar tube proteins and may lead to novel diagnostic and therapeutic reagents.
Supplementary key words. Glugea arnericanus, immunogold electron microscopy, Microspora, microsporidia, polar filament.
ICROSPORIDIA are protozoa classified in the phylum M Microspora that are eukaryotic, obligate intracellular, spore-forming parasites. They are ubiquitous in the animal king- dom with hundreds of species capable of parasitizing a wide range of invertebrate and vertebrate hosts, including humans [2, 41. Five microsporidian genera have been associated with human disease: Nosema. generally found in insects associated with keratitis; Pleistophora, a pathogen of fish and insects as- sociated with myositis; Encephalitoroon, found in many mam- mals associated with disseminated disease, keratitis and sinus- itis; Enterocytozoon associated with diarrhea and reported from AIDS patients and several species of fish; and Septata reported from AIDS patients and associated with diarrhea and dissem- inated disease [l-6, 11, 13, 151. The small spores (1-12 pm) produced by microsporidia are characteristic in that they con- tain a single polar tube (also called the polar filament [ 191) that coils around the sporoplasm and is attached to the inside of the spore by an anchoring disc [2,4]. When triggered by appropriate stimuli, the polar tube rapidly discharges from the anterior pole of the spore forming a hollow tube that remains attached to the spore [8, 9, 171. After complete discharge of the polar tube, the sporoplasm flows through the tube and appears as a droplet at its distal end [8, 9, 171. This process serves as an unique mech- anism of infection in that the polar tube can pierce an adjacent cell resulting in sporoplasm transfer directly into the host cell cytoplasm [9, 171.
Polar tubes range from 50 to 100 pm in length and 0.1 to 0.1 5 pm in diameter [8, 181. The polar tube has some flexibility in that it shows variation in diameter from 0.1 to 0.25 pm during discharge, its diameter can increase to 0.4 p m during sporoplasm passage, and its length shortens by 5%10% after sporoplasm passage [8, 12, 17, 181. Inside the spore the polar tube is filled with dense material (believed to be unassembled polar tube protein) and is often referred to as the polar filament, while the discharged tube appears as a hollow cylinder and is usually referred to as the polar tube [ 18, 191. Incompletely discharged tubes appear as a cylinder within a cylinder at their distal ends [ 191. Discharged tubes appear to be two to three times as long
' To whom correspondence should be addressed.
as the coiled tube inside the spore, and it has been suggested that the internal contents of the tube are incorporated at its growing tip during discharge [8, 191. This theory is supported by observations using pulse labelling with latex particles [ 191 and by video enhanced contrast microscopy [8]. The polar tube has unusual solubility properties in that it resists dissociation in 1% to 3% sodium dodecyl sulfate (SDS), 1% Triton X-100, 10% H,02, 8N H,SO,, 2N HCl, chloroform, 1% guanidine HCI, 0.1 M proteinase K and 10 M urea [17-191. The polar tube, however, dissociates in various concentrations of 2-mercapto- ethanol (2-ME) or 1% dithiothreitol (DTT) [18, 191. Weidner reported isolation of a polar tube protein from the microspo- ridian, Ameson michaelis, by treatment of discharged spores with 3% SDS which removed sporoplasm and soluble spore coat proteins, but left the resistant spore coats with attached polar tubes intact [ 181. Subsequent treatment with 1% DTT or 50% 2-ME solubilized the polar tubes, but not the spore coat, into the supernatant. SDS-polyacrylamide gel electrophoresis (SDS-PAGE) of the DTT supernatant showed a single 23-kDa protein, presumably representing polar tube protein [ 181.
In the present study, a monoclonal antibody was produced to a polar tube protein (PTP) of a microsporidium, which lo- calized to the polar tube by immunogold electron microscopy. The presence of a single polar tube is a defining characteristic of microsporidia, and study of PTP may suggest novel thera- peutic approaches to human infections or result in new diag- nostic reagents.
MATERIALS AND METHODS Isolation of polar tube protein. Spores of Glugea americanus
(formerly Spraguea lophii) were obtained from cysts found in the cranial and spinal ganglia of the angler fish, Lophius amer- icanus, and were stored at 4" C in sea water [14]. Polar tube proteins were separated from other proteins using a modification of a published protocol [ 181. A 3 x lo7 spore suspension in 1% SDS was disrupted with 0.5 p acid-washed glass beads (Sigma, St. Louis, MO) for 4 min on a Mini Beadbeater (Biospec Prod- ucts, Bartlesville, OK). The disrupted spores were centrifuged and the pellet extracted five times with 1% SDS and then with 9 M urea to remove sporoplasm released from broken spores and soluble spore coat proteins. After centrifugation, an aliquot
26

KEOHANE ET AL.-A MICROSPORIDIAN POLAR TUBE PROTEIN REACTIVE MONOCLONAL ANTIBODY 27
Fig. 1-3. Negative stain transmission electron microscopy of glass bead disrupted spores of G. umericunus. 1-2. Disrupted spores after extracting five times with 1% SDS and once with 9 M urea. Note broken spores (S) and straight and twisted polar tubes (PT, closed arrows). 3. Disrupted spores after washing five times with 1% SDS, once with 9 M urea, and incubated 2 h with 2% DTT. Note broken spores (S), lack of spore contents (open arrows), and absence of polar tubes.
of the resultant pellet was examined by negative stain trans- mission electron microscopy (TEM) using Formvar and carbon coated grids stained with 0.5% uranyl acetate or 1% phospho- tungstic acid. The pellet was then incubated with 0.5 ml of 2% DTT, 10 &ml each of aprotinin and leupeptin (Sigma), 5 mM EGTA and 25 mM Tris (pH 7.4) at 20" C for 2 h to solubilize the PTP. After DTT incubation and centrifugation, an aliquot of the resultant pellet was again examined by negative stain TEM.
Polyacrylamide gel electrophoresis (PAGE). The protein concentration in the DTT supernatant was determined by a Bradford dye binding procedure (Bio-Rad, Richmond, CA). Proteins in the DTT supernatant were visualized using SDS- PAGE according to the procedure described by Laemmli [lo]. A mid-range, prestained molecular weight marker (Diversified Biotech, Newton Centre, MA) was used for estimation of mo-
lecular weight. Gels were stained for 12 h with 0.0 1% Coomassie Brilliant Blue R-250 (Bio-Rad, Richmond, CA).
Production of monoclonal antibodies. After removal of the DTT by ultrafiltration (Centricon 10, Amicon, Beverly, MA) the soluble PTP were mixed in a 1:1 dilution with Hunter's TiterMax adjuvant (CytRx, Norcross, GA). BALB/c mice were immunized with a subcutaneous injection at the base of the tail. Fusion of mouse spleen cells with myeloma cells was performed at 12 wk post immunization at a 4: 1 ratio in 50% polyethylene glycol [7]. Supernatants from wells with hybridomas were screened by immunoblotting using spore lysate of G. americanus as the antigen. Hybridomas in immunoblot positive wells were subcloned twice into soft agarose.
Immunoblotting. Spore lysate proteins were prepared by glass bead disruption for 4 min (Mini-beadbeater) of 3 x 10' spores in 1 ml of solubilization buffer (1 50 mM NaC1, 1% Nonidet

28 J. EUK. MICROBIOL., VOL. 43, NO. 1, JANUARY-FEBRUARY 1996
Fig. 4. SDS-PAGE of G. americanus, Coomassie Blue. Lanes: 1, DTT supernatant showing major bands of 22, 27, 34 and 43 kDa, and 2, spore pellet after DTT incubation. The DTT washed pellet had bands of different molecular weight than the DTT supernatant.
P-40, 0.5% deoxycholate, 0.1% SDS, 50 mM Tris at pH 7.5). DTT solubilized PTP were prepared in the manner described above. Both preparations were mixed (2: 1) with sample buffer and subjected to SDS-PAGE as described above. Proteins were transferred to a nitrocellulose membrane by the method of Tow- bin et al. and blocked with BLOTTO buffer as previously de- scribed [ 16, 201. Hybridoma culture supernatants were incu- bated with the nitrocellulose membranes for 12 h at 20" C in a Miniblotter-45 apparatus (Immunometics, Cambridge, MA), followed by incubation with a 1:200 dilution of peroxidase con- jugated goat anti-mouse (IgG, IgM, IgA) antisera (Cappel, Or- ganon Teknika, West Chester, PA) for 3 h at 20" C. Immune complexes were visualized with 4-chloro- 1 -naphthol [20].
Immunogold electron microscopy. Spore suspensions were fixed in 2.5% (v/v) glutaraldehyde/O. 1 M sodium cacodylate (pH 7.2) in 1% paraformaldehyde, dehydrated in graded ethanols, and imbedded in LR White. Sections were placed on 300 mesh nickel grids coated with Formvar and carbon. Grids were in- cubated in blocking buffer (1 O/o bovine serum albumin-Sigma Fraction V, 0.02 O/o sodium azide in phosphate buffered saline, pH 7.35, with 5% goat serum) at 4" C for 12 h, followed by incubation with a 1:5 dilution of antibody in ascites for 2 h at 20" C, then a 1:20 dilution of goat anti-mouse IgG conjugated with 12 nm colloidal gold (Jackson Research, West Grove, PA) at 20" C for 90 min. Grids were stained with 1% uranyl acetate for 45 min and examined by TEM.
Epitope characterization. The DTT supernatant was incu- bated at 37" C for 12 h with each of the following: lysozyme (Boehringer Mannheim, Indianapolis, IN), 0.02 pglpg protein
Table 1. Monoclonal antibodies produced after immunization with G. americanus PTP.
Mab Isotype Immunoblot reactivity
7E6.29.29 2B12.8.7 8C10.4.32 3C8.23.1 683.20.1 2B8.6.24 2G2.5.1 2G6.16.1
23 & 42 kDa 27 kDa 34 kDa 43 kDa Multiple HMWa bands Single HMW band Single HMW band 12, 42, 80 kDa
a HMW, high molecular weight.
Fig. 5 . Immunoblot reactivity of Mab 3C8.23.1 with spore lysate of G. americanus. Lanes: 1, molecular weight marker; and 2, spore lysate reacted with a 1: lO dilution of Mab 3C8.23.1, a 1:200 dilution ofper- oxidase conjugated secondary antibody, and developed with 4-chloro- 1 -naphthol.
in 20 mM Tris, pH 6.5; neuraminidase (Boehringer Mannheim), 1.2 mU/pg protein in 20 mM Tris, pH 5.5; trypsin (Sigma), 0.02 pdpg protein in 20 mM Tris, pH 7.4; chymotrypsin (Boeh- ringer Mannheim), 0.01 pg/pg protein in 20 mM Tris, pH 7.4. After the incubation period, an identical amount of enzyme was again added, and incubated for an additional 2 h at 37" C. The mixtures were subjected to SDS-PAGE and immunoblotted with Mab 3C8.23.1 as described above.
RESULTS Glass bead disrupted spore pellets of G. americanus sequen-
tially extracted with 1 O/o SDS and 9 M urea demonstrated broken spores, empty spore coats and numerous polar tubes in straight or twisted formations by negative stain TEM (Fig. 1, 2). After subsequent incubation With 2% DTT, empty spore coats were still observed but the polar tubes were no longer present (Fig. 3). Since the polar tubes were soluble in DTT and spore coat was resistant to the treatment, it was assumed that the DTT supernatant contained one or more of the proteins comprising the polar tube.
The 2% DTT supernatant produced from the spores of G. americanus had four major protein bands by SDS-PAGE: 22, 27, 34 and 43 kDa (Fig. 4). The DTT supernatant had protein bands of different molecular weights than the DTT washed pel- let. The PTP in the DTT supernatant had no immunoblot reac- tivity with anti-actin C4 (Boehringer Mannheim), anti-actin W43A (Sigma), anti-0 tubulin KMX- 1 (Boehringer Mann- heim), anti-0 tubulin TUB 2.1 (Sigma) or anti-a tubulin DMlA (Sigma). These antibodies are reactive to actin and tubulins across multiple phyla.
Twelve weeks after BALB/c mice were immunized with the DTT supernatant, mouse serum showed multiple bands of reac- tivity against spore lysate antigen by immunoblot, including reactivity with each of the major protein bands identified in the DTT supernatant. After fusion and hybridoma screening, eight monoclonal antibodies were identified which reacted to Glugea proteins (Table 1). Mab 3C8.23.1 was reactive against a 43-kDa Glugea protein by immunoblot (Fig. 5). The immunoblot reac- tivity of Mab 3C8.23.1 was not affected by treatment of the DTT supernatant with lysozyme or neuraminidase but was abol- ished by treatment with chymotrypsin suggesting a protein epi- tope was recognized by the antibody. Mab 3C8.23.1 did not react with actins from rabbit heart or from Dictyostelium.
Longitudinal sections of G. americanus spores reveal six to eight polar tube cross sections in the coiled portion of the tube (Fig. 6). Spore cross sections (Fig. 7) contain a concentric coil of the polar tube winding around the spore. The outer edge of polar tube is limited by an electron-dense "sheath" that is visible
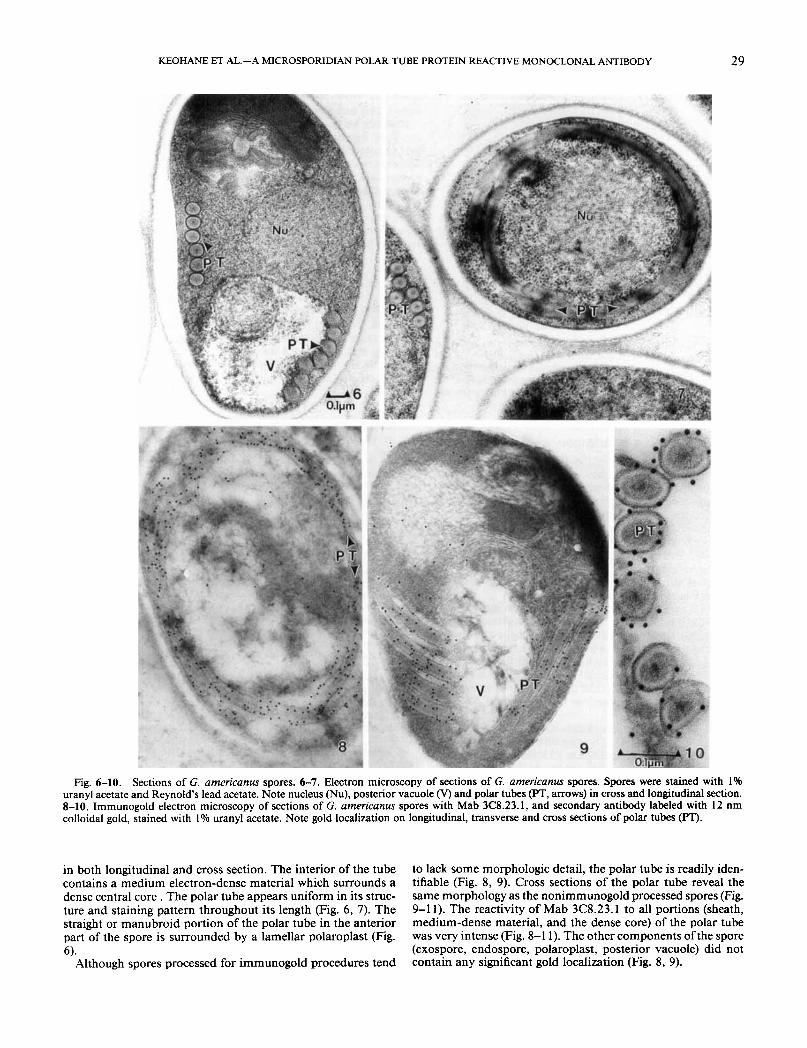
KEOHANE ET AL.-A MICROSPORIDIAN POLAR TUBE PROTEIN REACTIVE MONOCLONAL ANTIBODY 29
Fig. 6-10. Sections of G. americanus spores. 6-7. Electron microscopy of sections of G. americanus spores. Spores were stained with 1% uranyl acetate and Reynold's lead acetate. Note nucleus (Nu), posterior vacuole (V) and polar tubes (PT, arrows) in cross and longitudinal section. 8-10. Irnrnunogold electron microscopy of sections of G. americanus spores with Mab 3C8.23.1, and secondary antibody labeled with 12 nm colloidal gold, stained with I % uranyl acetate. Note gold localization on longitudinal, transverse and cross sections of polar tubes (PT).
in both longitudinal and cross section. The interior of the tube contains a medium electron-dense material which surrounds a dense central core . The polar tube appears uniform in its struc- ture and staining pattern throughout its length (Fig. 6, 7). The straight or manubroid portion of the polar tube in the anterior part of the spore is surrounded by a lamellar polaroplast (Fig. 6).
Although spores processed for immunogold procedures tend
to lack some morphologic detail, the polar tube is readily iden- tifiable (Fig. 8, 9). Cross sections of the polar tube reveal the same morphology as the nonimmunogold processed spores (Fig. 9-1 1). The reactivity of Mab 3C8.23.1 to all portions (sheath, medium-dense material, and the dense core) of the polar tube was very intense (Fig. 8-1 1). The other components of the spore (exospore, endospore, polaroplast, posterior vacuole) did not contain any significant gold localization (Fig. 8, 9).

30 J. EUK. MICROBIOL.. VOL. 43, NO. I , JANUARY-FEBRUARY 1996
Fig. 11. Section through a G. americanus spore containing four polar tube (PT) cross sections, a portion of the lamellar polaroplast (PL), and a sagital cut through the anterior straight portion (manubroid) of the polar tube (MPT) indicated by arrowheads. Note the localization of the gold on the “sheath” or outer portion, the “dense” core or center, and the medium-dense material of the polar tube.
DISCUSSION A single coiled polar tube is the structure which uniquely
identifies microsporidia. It is present, in similar structural and functional form, in all microsporidia regardless of species, host or geographic location. Because of its uniqueness, it is likely that this structure evolved prior to the divergence of micro- sporidia into various genera, and is not the result ofconvergence of an independently evolved polar tube structure in individual microsporidia. Therefore, the proteins comprising the polar tube are likely to be members of a unique protein family evolved from the same ancestral gene, with a highly conserved amino acid sequence among the various microsporidia.
In our PTP isolation protocol, glass bead disrupted spores, extracted with 1% SDS and 9 M urea, demonstrated broken spore coats and numerous polar tubes in straight and twisted formations. After incubation with DTT, and subsequent cen- trifugation, polar tubes were no longer seen in the spore pellet preparation, and it was presumed that the DTT supernatant contained one or more polar tube proteins. This implied that disulfide linkages are important for stabilization of the polar tube structure.
SDS-PAGE of the DTT supernatant showed four major pro- tein bands: 23,27,34 and 43 kDa. This differs from a previously reported isolation of a single 23-kDa PTP from the microspo- ridian A. michaelis [ 181. Using this material as the antigen source, monoclonal antibodies were raised to proteins of the same mo- lecular weight as the four major proteins found in the DTT supernatant. After reacting these antibodies with G. americanus spores by immunogold electron microscopy, Mab 3C8.23.1, which reacted to a 43-kDa protein by immunoblot, showed strong localization to the polar tube. Immunogold labeling by Mab 3C8.23.1 was very intense on all aspects of the polar tube (cross, longitudinal, and sagittal section) and it appears that Mab 3C8.23.1 recognizes a PTP present in the dense core, medium dense material, and the “sheath” of the polar tube. There was no reaction of Mab 3C8.23.1 with other spore structures nor was their significant nonspecific background reactivity.
Although similar in molecular weight to actin, the PTP rec- ognized by Mab 3C8.23.1 had different solubility properties than actin and did not react with anti-actin antibodies of broad
specificity. Furthermore, Mab 3C8.23.1 did not react with actin from rabbit heart or Dictyostelium.
Recently we have been able to purify the 43-kDa PTP from the DTT supernatant of G. americanus by reverse phase high performance liquid chromatography (HPLC) (unpubl. data). Since the microsporidian polar tube is a unique structure serving to facilitate transmission of this parasite into a new host cell, further characterization of this 43-kDa PTP may reveal it to be a member of a unique protein family. Study of polar tube pro- teins may furthermore yield new therapeutic strategies for the treatment of microsporidian infections as well as new diagnostic reagents for these parasites.
ACKNOWLEDGMENTS This work was supported by National Institutes of Health
grants A13 1788 and CA13330. We wish to thank Dr. Earl Weid- ner (Louisiana State University, Baton Rouge, LA) for his advice and John Ziskowski (NMFS, NOAA Laboratory, Milford, CT) for collection of spores. A presentation of part of the data con- tained in this manuscript was made at the June 1994 Proto- zoology Society meeting in Cleveland, and was published as an abstract for that meeting.
LITERATURE CITED 1. Cali, A. 199 1. General microsporidian features and recent find-
ings on AIDS isolates. J. Protozool., 38:625-630. 2. Cali, A. & Owen, R. L. 1988. Microsporidiosis. In: Ballows, A,,
Hausler, W. J., Jr., Ohashi, M. & Turano, H. (ed), Laboratory Diagnosis of Infectious Diseases: Principles and Practice, Vol. 1, Bacterial, My- cotic, and Parasitic Diseases. Springer-Verlag, New York, New York.
3. Cali, A., Kotler, D. P. & Orenstein, J. M. 1993. Seprata intes- tinalis N. G., N. Sp., an intestinal microsporidian associated with chron- ic diarrhea and dissemination in AIDS patients. J. Euk. Microbiol., 40: 101-1 12.
4. Canning, E. U. & Lom, J. 1986. The Microsporidia of Verte- brates. Academic Press, New York, New York.
5. Chilmonczyk, S., Cox, W. T. & Hedrick, R. P. 1991. Entero- cytozoon salmonis n. sp.: an intranuclear microsporidium from sal- monid fish. J. Protozool., 38;264-269.
6. Desportes, I., Le Charpentier, Y., Galian, A., Bernard, F., Co- chand-Priollet, B., Lavergne, A., Ravisse, P. & Modigliani, R. 1985. Occurrence of a new microsporidian: Enterocytozoon bieneusi n. g., n. sp., in the enterocytes of a human patient with AIDS. J. Protozool., 32: 250-254.
7. Fazekas de St. Groth, S. & Scheidegger, D. 1980. Production of monoclonal antibodies. Strategy and tactics. J. Imm. Meth., 35: 1-2 1.
8. Frixione, E., Ruiz, L., Santillan, M., de Vargas, L. V., Tejero, J. M. & Undeen, A. H. 1992. Dynamics of polar filament discharge and sporoplasm expulsion by microsporidian spores. Cell Motil. Cytoskl., 22:38-50.
9. Ishihara, R. 1968. Some observations on the fine structure of sporoplasm discharged from spores of a microsporidian, Nosema bom- bycis. J. Invert. Pathol., 12:245-258.
10. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227:680-685.
1 1 . Ledford, D. K., Overman, M. D., Gonzolvo, A., Cali, A., Mester, S. W. & Lockey, R. F. 1985. Microsporidiosis myositis in a patient with the acquired immunodeficiency syndrome. Ann. Intern. Med., 102: 628-630.
12. Scarborough-Bull, A. & Weidner, E. 1985. Some properties of discharged Glugea hertwigi (Microsporida) sporoplasms. J. Protozool., 32~284-289.
13. Strano, A. J., Cali, A. & Neafie, R. C. 1976. Microsporiodiosis. In: Binford, C. H. & Connor, D. H. (ed.), Pathology of Tropical and Extraordinary Diseases. Vol. 1. Armed Forces Institute of Pathology, Washington, D.C. Pp.336-339.
14. Takvorian, P. M. & Cali, A. 1986. The ultrastructure of spores (Protozoa:Microsporida) from Lophius americanus, the angler fish. J. Prorozool.. 33: 57&5 75.
Pp. 929-95 0.

KEOHANE ET AL.- A MICROSPORIDIAN POLAR TUBE PROTEIN REACTIVE MONOCLONAL ANTIBODY 31
15. Terada, S. , Reddy, K. R., Jeffers, L. J., Cali, A. & Schiff, E. R. 1987. Microsporidian hepatitis in the acquired immunodeficiency syn- drome. Ann. Intern. Med., 107:61-62.
16. Towbin, H., Staehelin, T. & Gordon, J. 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc. Natl. Acad. Sci. (USA), 76: 4350-4354. Hyg., 47:456462.
17. Weidner, E. 1972. Ultrastructural study of microsporidian in- vasion into cells. Z. Parasitenk., 40:227-242.
18. Weidner, E. 1976. The microsporidian spore invasion tube. The ultrastructure, isolation, and characterization of the protein com- prising the tube. J. Cell Biol., 71:23-34.
19. Weidner, E. 1982. The microsporidian spore invasion tube. 111. Tube extrusion and assembly. J. Cell Biol., 93:976-979.
20. Weiss, L. M., Cali, A., Levee, E., LaPlace, D., Tanowitz, H., Simon, D. & Wittner, M. 1992. Diagnosis ofEncephalitozoon cuniculi infection by western blot and the use of cross-reactive antigens for the possible detection of microsporidiosis in humans. Am. J. Trop. Med.
Received 5- 15- 95, 8-2-95; accepted 8- 17- 95
J. Euk. Microbiol., 43(1), 1996, pp. 31-34 0 1996 by the Society of Protozoologists
Entamoeba histolytica Encodes a Highly Divergent @-Tubulin SANTOSH K. KATIYAR and THOMAS D. EDLIND’
Department of Microbiology and Immunology, Medical College of Pennsylvania and Hahnemann University, Philadelphia. Pennsylvania I9129
ABSTRACT. The microtubules of the amitochondrial parasite Entamoeba histolytica are atypical in certain respects. Consistent with this, we report that E. histolyrica encodes the most divergent p-tubulin identified to date, with only 54% to 58% identity to 0-tubulins from various species. A similarly divergent o-tubulin is encoded by the related Entamoeba invadens; single gene copies appear to be present in both organisms. The Entamoeba sequences were compared with a database of 101 8-tubulins, including the highly divergent sequence from another amitochondrial protozoan, Trichomonas vaginalis. A total of 8 1 residues were universally conserved, and 76 residues varied only once. Correlations with previous studies indicate that microtubule function is altered when most, but not all, conserved residues are mutated.
Supplementary key words. Archezoa, microtubule, Trichomonas vaginalis, tubulin.
ICROTUBULES are a characteristic feature of eukaryotic M cells. They are major components of the mitotic spindle and, in many cells, the cytoskeleton and axonemes. Microtu- bules form by polymerization of tubulin dimers, consisting of a- and P-tubulin subunits. A third subunit, y-tubulin, is asso- ciated with the centrosome. A large number of tubulin sequences have been determined, although primarily from higher eukary- otes. Analysis of these sequences reveals a remarkable degree of evolutionary conservation. For example, comparing slime mold [3] and human [ 151 0-tubulin, there is 8 1% identity and no insertions or deletions from amino acids 1 to 430. The re- maining (typically 15 to 20) carboxy terminal residues are more variable. Analysis of about 90 different P-tubulin sequences available in 199 1 identified 136 residues which were universally conserved [4].
With the exception of Giardia lamblia j3-tubulin [14], until recently no tubulin sequences have been available from the amitochondrial protozoa, a group of organisms (Archezoa) which may be directly descended from early, pre-mitochondria1 eu- karyotes [5, 61. Analysis of such sequences would help identify the minimal requirements for tubulin structure and function; furthermore, it might shed light on the evolutionary origins of microtubules. Consequently, we examined P-tubulin genes from Trichomonas vaginalis which, like G. lamblia, possesses abun- dant microtubules and is highly sensitive to the microtubule- targeted benzimidazole drugs [ 1 11. Indeed, T. vaginah p-tu- bulin is relatively divergent, with 76% to 80% identity to other protozoa1 P-tubulins. Here, we present the complete sequence of a @-tubulin gene from Entamoeba histolytica. This important intestinal parasite lacks cytoplasmic microtubules, and even spindle microtubules were thought to be absent until techniques for synchronizing cultures were developed [ 191. Furthermore, this organism is largely insensitive to known microtubule-tar-
To whom correspondence should be addressed.
geted drugs [ 121. The E. histolytica gene encodes the most di- vergent @-tubulin identified to date. A partial sequence of En- tamoeba invadens P-tubulin and an analysis of an updated @-tu- bulin database for conserved residues are also presented.
MATERIALS AND METHODS Entamoeba histolytica strain 200:NIH was cultured as de-
scribed [lo]; E. invadens 30994 was obtained from the American Type Culture Collection and cultured as recommended. To am- plify P-tubulin gene fragments, primers BT107 [7] and
CGTTGTC-3’; K = G+T, PstI site italicized) were employed. Genomic DNA preparation and amplification conditions were as described [ I 11. To obtain the flanking E. histolytica DNA, inverse PCR [ 181 was performed with circularized HindIII-di- gested genomic DNA. The 3’ terminus ofE. histolytica @-tubulin mRNA was characterized by reverse transcription of total RNA [ 1 11 with an oligo(dT) primer, followed by amplification [9] with the same primer and an upstream primer. All amplification products were cloned into M13 and sequenced on both strands. Sequences have been deposited in the GenBank database with accession numbers L3 1809 and L3 18 10.
TUB203PR (5’-GTTCTGCAGCMTGTCGTAKAGKGCTT-
RESULTS AND DISCUSSION Cloning complete E. hisiolytica and partial E. invadens &tu-
bulin genes. With DNA from both E. histolytica and the related reptilian parasite E. invadens, a primer pair representing con- served @-tubulin residues 100 to 107 and 203 to 208 generated a DNA amplification product of the expected size. Cloning and sequence analysis confirmed that these fragments encoded @-tu- bulin, although highly divergent (Fig. 1, and data not shown). Hybridization of the amplified DNA under conditions of mod- erate stringency to Southern blots of digested E. histolylica and E. invadens genomic DNAs identified single bands (not shown), consistent with single gene copies.
Related Documents











