[CANCER RESEARCH 60, 1353–1360, March 1, 2000] Identification of a Human Glioma-associated Growth Factor Gene, granulin, Using Differential Immuno-absorption 1 Linda M. Liau, 2 Roger L. Lallone, Robert S. Seitz, Alexei Buznikov, Jeff P. Gregg, Harley I. Kornblum, Stanley F. Nelson, and Jeff M. Bronstein Division of Neurosurgery [L. M. L.], Departments of Neurology [A. B., J. M. B.], Pathology [J. P. G.], Pharmacology [H. I. K.], and Human Genetics [S. F. N.], and the Jonsson Comprehensive Cancer Center [L. M. L., S. F. N.], University of California at Los Angeles School of Medicine, Los Angeles, California 90095; Brookwood Biomedical, Birmingham, Alabama 35209 [R. L. L.]; and Research Genetics Inc., Huntsville, Alabama 35801 [R. S. S.] ABSTRACT Identification of the genes that are differentially expressed in brain tumor cells but not in normal brain cells is important for understanding the molecular basis of these neurological cancers and for defining possible targets for therapeutic intervention. In an effort to discover potentially antigenic proteins that may be involved in the malignant transformation and progression of human glioblastomas, a novel antibody-based ap- proach was developed to identify and isolate gene products that are expressed in brain tumors versus normal brain tissue. Using this method, whereby tumor-specific antibodies were isolated and used to screen a glioblastoma cDNA expression library, 28 gene products were identified. Nine of these clones had homology to known gene products, and 19 were novel. The expression of these genes in multiple different human gliomas was then evaluated by cDNA microarray hybridization. One of the iso- lated clones had consistently higher levels of expression (3–30-fold) in brain tumors compared with normal brain. Northern blot analysis and in situ hybridization confirmed this differential overexpression. cDNA se- quence analysis revealed that this gene was identical to a relatively new class of growth regulators known as granulins, which have tertiary struc- tures resembling the epidermal growth factor-like proteins. The 2.1-kb granulin mRNA was expressed predominantly in glial tumors, with lower levels in spleen, kidney, and testes, whereas expression was not detected in non-tumor brain tissues. Functional assays using [ 3 H]thymidine incorpo- ration indicated that granulin may be a glial mitogen, as addition of synthetic granulin peptide to primary rat astrocytes and three different early-passage human glioblastoma cultures increased cell proliferation in vitro, whereas increasing concentrations of granulin antibody inhibited cell growth in a dose-dependent manner. The differential expression pattern, tissue distribution, and implication of this glioma-associated mol- ecule in growth regulation suggest a potentially important role for granu- lin in the pathogenesis and/or malignant progression of primary brain neoplasms. INTRODUCTION Cancer is the result of cumulative multiple genetic mutations, which result in the activation of oncogenes and/or the inactivation of tumor suppressor genes. It is the differential expression of these critical genes and their downstream effectors that enables cells to override growth controls and undergo carcinogenesis (1, 2). The pathological changes that arise in cancer, whether caused by a single gene mutation or multiple genetic alterations, are essentially driven by changes in gene expression (1, 2). In the malignant progression of astrocytic cancers, it has been shown that accumulation of multiple genetic lesions underlies the neoplastic process. These lesions include mutations of the genes p53, p16, RB, and PTEN, as well as amplifi- cation of CDK4 and EGFR (3, 4). Although these known genetic abnormalities have been well documented in the formation of the most malignant brain tumor, glioblastoma, recent insight into the extent of gene expression differences underlying malignancy reveals that hun- dreds of gene transcripts may be expressed at significantly different levels between normal and neoplastic cells (5). Therefore, there is considerable room for the identification of novel genes that are differentially expressed in brain tumor cells to further our understand- ing of the complex molecular basis of these neurological cancers. Furthermore, this endeavor has direct clinical relevance if combined with the development of innovative rational therapies that specifically target these differentially expressed gene products. A variety of methods are currently used to isolate genes associated with particular differential phenotypes. Subtractive hybridization (6), differential display (7–10), representational difference analysis (11– 14), serial analysis of gene expression (5, 15), and suppression sub- tractive hybridization (16, 17) all allow for the cloning and identifi- cation of differentially expressed sequences. Although all these techniques identify tissue-enriched mRNAs, none select for tissue- specific proteins. Because we initially were interested in identifying glioma-associated antigens that may be potential targets for brain tumor immunotherapy, we set out to devise a differential screening technique that provided actual confirmation of the presence of a protein product, not just the capacity to synthesize a protein. Further- more, we wanted to select for proteins with antigenic determinants that may be potentially recognized by the immune system. Here we report an approach to identifying differentially expressed gene products that are actually translated from mRNA species, using antibody-based screening of a cDNA expression library. We further show that this method, which we termed DIA, 3 can be coupled to cDNA microarray hybridization and allowed the identification of a putative growth factor gene, granulin, which may play a role in the malignant progression of glioblastomas. MATERIALS AND METHODS DIA. GBM tumor tissue was immediately snap frozen in liquid nitrogen at the time of surgery. Non-tumor brain was obtained from a surgical resection for trauma and similarly frozen. Both tissue specimens were homogenized in PBS (pH 7.0) with a glass mortar and pestle. The soluble material was aspirated, and the insoluble material was re-extracted into a second fraction, using 0.1% SDS. Both fractions were used for affinity purification and immu- nizations. Affinity chromatography was carried out using an aliquot of ex- tracted material immobilized on CNBr-activated Sepharose 4B columns (Phar- macia) according to the manufacturer’s specifications. The immobilized GBM tumor tissue extract was loaded into a fritted column (Varian), blocked with 1 M glycine, precycled with 0.1 M HCl, and neutralized with 0.1 M BBS (pH 8.4). The extract of non-tumor brain tissue was similarly immobilized. Received 10/13/99; accepted 1/5/00. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1 Supported in part by grants from the National Cancer Institute (Grant NIH CA 82666-01), the American Cancer Society (Grant IN-131), the Howard Hughes Medical Institute (Grant 76296-549701), and the STOP Cancer Foundation (awarded to L. M. L.). We also thank the generous support of the Henry E. Singleton Brain Cancer Research Fund. 2 To whom requests for reprints should be addressed, at Division of Neurosurgery, UCLA School of Medicine, Center for Health Sciences, CHS 74-134, 10833 Le Conte Avenue, Box 956901, Los Angeles, CA 90095-6901. Phone: (310) 794-5664; Fax: (310) 825-7245; E-mail: [email protected]. 3 The abbreviations used are: DIA, differential immuno-absorption; GBM, glioblas- toma multiforme; BBS, borate-buffered saline; FBS, fetal bovine serum; EGF, epidermal growth factor; EGFR, EGF receptor. 1353 Research. on August 17, 2015. © 2000 American Association for Cancer cancerres.aacrjournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

[CANCER RESEARCH 60, 1353–1360, March 1, 2000]
Identification of a Human Glioma-associated Growth Factor Gene,granulin, UsingDifferential Immuno-absorption 1
Linda M. Liau, 2 Roger L. Lallone, Robert S. Seitz, Alexei Buznikov, Jeff P. Gregg, Harley I. Kornblum,Stanley F. Nelson, and Jeff M. BronsteinDivision of Neurosurgery [L. M. L.], Departments of Neurology [A. B., J. M. B.], Pathology [J. P. G.], Pharmacology [H. I. K.], and Human Genetics [S. F. N.], and the JonssonComprehensive Cancer Center [L. M. L., S. F. N.], University of California at Los Angeles School of Medicine, Los Angeles, California 90095; Brookwood Biomedical,Birmingham, Alabama 35209 [R. L. L.]; and Research Genetics Inc., Huntsville, Alabama 35801 [R. S. S.]
ABSTRACT
Identification of the genes that are differentially expressed in braintumor cells but not in normal brain cells is important for understandingthe molecular basis of these neurological cancers and for defining possibletargets for therapeutic intervention. In an effort to discover potentiallyantigenic proteins that may be involved in the malignant transformationand progression of human glioblastomas, a novel antibody-based ap-proach was developed to identify and isolate gene products that areexpressed in brain tumorsversusnormal brain tissue. Using this method,whereby tumor-specific antibodies were isolated and used to screen aglioblastoma cDNA expression library, 28 gene products were identified.Nine of these clones had homology to known gene products, and 19 werenovel. The expression of these genes in multiple different human gliomaswas then evaluated by cDNA microarray hybridization. One of the iso-lated clones had consistently higher levels of expression (3–30-fold) inbrain tumors compared with normal brain. Northern blot analysis and insitu hybridization confirmed this differential overexpression. cDNA se-quence analysis revealed that this gene was identical to a relatively newclass of growth regulators known asgranulins, which have tertiary struc-tures resembling the epidermal growth factor-like proteins. The 2.1-kbgranulin mRNA was expressed predominantly in glial tumors, with lowerlevels in spleen, kidney, and testes, whereas expression was not detected innon-tumor brain tissues. Functional assays using [3H]thymidine incorpo-ration indicated that granulin may be a glial mitogen, as addition ofsynthetic granulin peptide to primary rat astrocytes and three differentearly-passage human glioblastoma cultures increased cell proliferationinvitro, whereas increasing concentrations of granulin antibody inhibitedcell growth in a dose-dependent manner. The differential expressionpattern, tissue distribution, and implication of this glioma-associated mol-ecule in growth regulation suggest a potentially important role for granu-lin in the pathogenesis and/or malignant progression of primary brainneoplasms.
INTRODUCTION
Cancer is the result of cumulative multiple genetic mutations,which result in the activation of oncogenes and/or the inactivation oftumor suppressor genes. It is the differential expression of thesecritical genes and their downstream effectors that enables cells tooverride growth controls and undergo carcinogenesis (1, 2). Thepathological changes that arise in cancer, whether caused by a singlegene mutation or multiple genetic alterations, are essentially driven bychanges in gene expression (1, 2). In the malignant progression ofastrocytic cancers, it has been shown that accumulation of multiple
genetic lesions underlies the neoplastic process. These lesions includemutations of the genesp53,p16,RB, andPTEN, as well as amplifi-cation of CDK4 and EGFR (3, 4). Although these known geneticabnormalities have been well documented in the formation of the mostmalignant brain tumor, glioblastoma, recent insight into the extent ofgene expression differences underlying malignancy reveals that hun-dreds of gene transcripts may be expressed at significantly differentlevels between normal and neoplastic cells (5). Therefore, there isconsiderable room for the identification of novel genes that aredifferentially expressed in brain tumor cells to further our understand-ing of the complex molecular basis of these neurological cancers.Furthermore, this endeavor has direct clinical relevance if combinedwith the development of innovative rational therapies that specificallytarget these differentially expressed gene products.
A variety of methods are currently used to isolate genes associatedwith particular differential phenotypes. Subtractive hybridization (6),differential display (7–10), representational difference analysis (11–14), serial analysis of gene expression (5, 15), and suppression sub-tractive hybridization (16, 17) all allow for the cloning and identifi-cation of differentially expressed sequences. Although all thesetechniques identify tissue-enriched mRNAs, none select for tissue-specific proteins. Because we initially were interested in identifyingglioma-associated antigens that may be potential targets for braintumor immunotherapy, we set out to devise a differential screeningtechnique that provided actual confirmation of the presence of aprotein product, not just the capacity to synthesize a protein. Further-more, we wanted to select for proteins with antigenic determinantsthat may be potentially recognized by the immune system.
Here we report an approach to identifying differentially expressedgene products that are actually translated from mRNA species, usingantibody-based screening of a cDNA expression library. We furthershow that this method, which we termed DIA,3 can be coupled tocDNA microarray hybridization and allowed the identification of aputative growth factor gene,granulin, which may play a role in themalignant progression of glioblastomas.
MATERIALS AND METHODS
DIA. GBM tumor tissue was immediately snap frozen in liquid nitrogen atthe time of surgery. Non-tumor brain was obtained from a surgical resectionfor trauma and similarly frozen. Both tissue specimens were homogenized inPBS (pH 7.0) with a glass mortar and pestle. The soluble material wasaspirated, and the insoluble material was re-extracted into a second fraction,using 0.1% SDS. Both fractions were used for affinity purification and immu-nizations. Affinity chromatography was carried out using an aliquot of ex-tracted material immobilized on CNBr-activated Sepharose 4B columns (Phar-macia) according to the manufacturer’s specifications. The immobilized GBMtumor tissue extract was loaded into a fritted column (Varian), blocked with 1M glycine, precycled with 0.1M HCl, and neutralized with 0.1M BBS (pH 8.4).The extract of non-tumor brain tissue was similarly immobilized.
Received 10/13/99; accepted 1/5/00.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby markedadvertisementin accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1 Supported in part by grants from the National Cancer Institute (Grant NIH CA82666-01), the American Cancer Society (Grant IN-131), the Howard Hughes MedicalInstitute (Grant 76296-549701), and the STOP Cancer Foundation (awarded to L. M. L.).We also thank the generous support of the Henry E. Singleton Brain Cancer ResearchFund.
2 To whom requests for reprints should be addressed, at Division of Neurosurgery,UCLA School of Medicine, Center for Health Sciences, CHS 74-134, 10833 Le ConteAvenue, Box 956901, Los Angeles, CA 90095-6901. Phone: (310) 794-5664; Fax:(310) 825-7245; E-mail: [email protected].
3 The abbreviations used are: DIA, differential immuno-absorption; GBM, glioblas-toma multiforme; BBS, borate-buffered saline; FBS, fetal bovine serum; EGF, epidermalgrowth factor; EGFR, EGF receptor.
1353
Research. on August 17, 2015. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from

Antisera were raised against the GBM tumor homogenate by s.c. and i.m.immunization of New Zealand White rabbits, using complete and incompleteFreund’s adjuvants. Several bleeds were collected from two animals, pooled,and diluted 1:2 with 0.1M BBS (pH 8.4). The diluted antiserum was passedover the GBM affinity column, and unbound material was washed off withBBS. Bound material was eluted off using glycine buffers adjusted to pH 3, pH2, and then pH 1. The effluent and eluate were monitored at 280 nm (LKB),and the antiserum was passed repeatedly through the column until depleted.The eluate was then collected into BBS, checked for neutral pH, and cross-absorbed repeatedly (until depleted of cross-reactive antibodies) against thecolumn of non-tumor brain to select out antibodies that may bind normal brainantigens. The unbound material was further cross-absorbed against normalhuman plasma to select out nonspecific antibodies. The final product wasconcentrated using YM30 columns (Amicon) and dialyzed into carbonatebuffer (pH 9.5). The antibodies were biotinylated at a molar ratio of 15:1 usingNHS long-chain biotin (Sigma) and repurified using a column of G-25 (Phar-macia). These biotinylated antibodies were then used to screen a glioblastomaphagemid cDNA expression library.
Construction and Screening of cDNA Expression Library. For con-struction of the glioblastoma cDNA expression library, a human GBM tumorwas snap frozen in liquid nitrogen at the time of surgery and stored at280°C.Total RNA was extracted from 500 mg of fresh frozen tumor tissue, usingTrizol reagent according to the manufacturer’s protocol (Life Technologies,Inc.). mRNA from a total of 30mg of RNA was isolated using doublechromatography on oligo-dT cellulose columns (Life Technologies, Inc.).Double-stranded cDNA was synthesized from this mRNA, using a SuperscriptII cDNA synthesis kit (Life Technologies, Inc.), and the cDNAs were ligatedinto a l ZipLox phagemid vector (Life Technologies, Inc.). We obtained alibrary titer estimated at 5.03 106 plaque-forming units. Approximately2.0 3 106 plaque-forming units were plated and grown in the presence ofisopropyl-1-thio-b-D-galactoside, lifted onto nitrocellulose membranes, andincubated with biotinylated anti-GBM antibodies (1:1000 dilution). The mem-branes were then incubated with streptavidin-horseradish peroxidase and dia-minobenzidine tetrahydrochloride (Pierce). Positive clones were isolated, re-screened, and subcloned into the pZL1 plasmid vector (Life Technologies,Inc.) by in vivo excision. Inserts were verified by agarose gel electrophoresisand partially sequenced using a dsDNA cycle sequencing kit (Life Technolo-gies, Inc.) according to the manufacturer’s protocol.
Microarraying of Cloned DIA Products. After the subtractive productswere cloned into the pZL1 vector, plasmid inserts were PCR-amplified usingvector-specific primers. PCR was performed in 50-ml reactions containing 10mM Tris (pH 9.0), 50 mM KCl, 0.1% gelatin, 2.5 units of Taq DNA polym-erase, and 150mM deoxynucleotide triphosphates. Thermal cycling conditionsconsisted of an initial denaturation at 94°C for 2 min, followed by 35 cycles of94°C for 1 min, 68°C for 1 min, and 72°C for 1.5 min, with a final 72°Cextension for 10 min, in a PTC100 thermal cycler (MJ Research). Fivemicroliters of each PCR amplification product were examined by agarose gelelectrophoresis with ethidium bromide staining. A single band was detected in26 of the 28 PCR reactions performed. Each of the 26 successfully amplifiedPCR products (1–2mg) was recovered from the remaining 45ml of each PCRreaction by ethanol precipitation.
The PCR products were arrayed onto glass slides, following a protocolsimilar to that described previously (18). Briefly, the PCR products wereresuspended in 15ml of 13 SSC. A custom-built arraying robot picked up;600 nl of DNA solution and deposited 1–4 nl of DNA solution in triplicateonto a silanized glass slide surface (Sigma). After printing, the slide washydrated for 10 s over a 37°C water bath, snap dried for 2 s on a 100°C heatingblock, and then UV cross-linked with 4000 mJ short-wave irradiation (Strat-agene Stratalinker). The slide was then washed for 2 min sequentially in 0.2%SDS and distilled water. The bound DNA was denatured in distilled water at100°C, desiccated in an ice-cold bath of 95% ethanol, and air-dried.
Probe labeling, microarray hybridization, and washes were performed asdescribed previously (19). mRNA from a large batch of pooled tumor andnon-tumor brain specimens was used to make cDNA labeled with Cy5. TheCy5-labeled cDNA from this collective batch served as the common referenceprobe in all hybridizations. mRNA samples (2mg) from 10 individual tumorand non-tumor brain specimens (e.g.,8 gliomas and 2 normal brain tissues)were used to make cDNA labeled with Cy3.
After hybridization with the arrayed subtractive clones, Cy3 and Cy5
intensities were scanned using a custom-built two-color laser scanning fluo-rometer. The image files were analyzed with custom-written software thatperformed quantification similar to that published previously (20, 21). Therelative abundance of each of our 26 subtractive clones (L1–L26) in tumorversusnormal brain was calculated using the equation:
[(Cy3 signal2 Cy3 background)tumor/(Cy5 signal2 Cy5 background)]
[(Cy3 signal2 Cy3 background)normal/(Cy5 signal2 Cy5 background)]
Northern Blot Analysis of Granulin mRNA Expression. Tissue totalRNA was extracted using Trizol reagent (Life Technologies, Inc.) according tothe manufacturer’s instructions, and 10mg/lane were separated on 1.2%denaturing agarose gels, transferred overnight to Hybond membranes (Amer-sham) using 103SSC, and irreversibly fixed by UV cross-linking. Prehybrid-ization and hybridization were performed at 65°C in ExpressHyb solution(Clontech).32P-labeled cDNA probes were generated from our plasmid DNAcontaininggranulin cDNA using random primers according to the manufac-turer’s protocol (NEB). After hybridization, membranes were washed (23SSC containing 0.1% SDS at 37°C for 20 min, followed by 0.23SSCcontaining 0.1% SDS at 61°C for 20 min), and exposed to X-ray film (Kodak)at 80°C. Blots were then stripped with 0.1% SDS at 100°C for 15 min andreprobed with32P-labeled ribosomal 18S cDNA to control for gel loading andRNA integrity.
In Situ Hybridization of Granulin mRNA Expression. In situ hybridiza-tion was performed using35S-labeled riboprobes following previously pub-lished protocols (22). Briefly, surgically resected human brain tissues (tumorand non-tumor) were rapidly frozen in isopentane directly from the operatingroom. Frozen tissues were sectioned on a cryostat at 20-mm thickness, post-fixed in 4% paraformaldehyde, washed, and stored at275°C. Sections werewashed, acetylated, defatted, and incubated with35S-labeled sense or antisensegranulin cRNA probe (107 cpm/ml) at 60°C overnight (18–24 h). FollowingRNase A (20mg/ml) treatment at 45°C, sections were washed in descendingconcentrations of SSC, air dried, and dipped for emulsion autoradiography inKodak NTB2 (1:1 dilution). Following exposure to emulsion for 5 weeks, theslides were developed and counterstained with H&E.
Hybridization densities were measured from thein situ slides by countingsilver grains within representative cells, using an image analysis computer(Olympus microscope and MCID Imaging software; Imaging Research, Inc.;Ref. 23). Sections through several different tumor and non-tumor human brainspecimens that had been hybridized withgranulin cRNA were chosen forcounts. Briefly, two independent observers outlined labeled regions withineach slide, and the computer determined the absorbance and quantity of silvergrains within each outlined area. Ten measurements were performed for eachslide and averaged into single values per mm2 per specimen. These values werethen divided by the estimated number of cells per mm2 for each specimen toget the average units of silver grains per cell. The average quantity of silvergrains per cell for each tumor was compared to that of non-tumor brainspecimens using the Student’st-test.
Cell Cultures. Primary cultures of rat astrocytes from the brains of adultFischer 344 rats were isolated following a protocol described previously (24).Cultures were maintained in DMEM (Life Technologies, Inc.) supplementedwith 10% FBS,L-glutamine, and antibiotic drugs (100 units/ml penicillin and100 mg/ml streptomycin) at 37°C in 5% CO2.
Primary human glioblastoma cell cultures were established in our laboratoryusing a protocol similar to that published previously (25). Tumors were takendirectly from the operating room at the time of surgery. Tissues were finelyminced using sterile scissors, rinsed with PBS, and dispersed with trypsin-EDTA. Monolayer cells were plated in T75 flasks (Costar) and cultured inDMEM/Ham’s F12 (Irvine Scientific) supplemented with 10% FBS (LifeTechnologies, Inc.),L-glutamine, and antibiotics (100 units/ml penicillin and100 mg/ml streptomycin).
Measurement of Cell Proliferation. The effect of granulin D peptide andgranulin antibody on the proliferation of primary rat astrocytes and threeearly-passage human glioblastoma cell lines were examined. Synthetic peptide,consisting of the 55-amino acid sequence of granulin D (26), was provided byResearch Genetics. For the antibody studies, a polyclonal antibody was raisedagainst this 55-amino acid synthetic peptide conjugated to keyhole limpethemocyanin. The IgG fraction was isolated from sera, using protein A-Sepharose (Zymed), concentrated using a Centri-cell concentrator (Amicon),
1354
GRANULIN, A GLIAL MITOGEN IDENTIFIED BY DIA
Research. on August 17, 2015. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from

and stored in borate buffer consisting of 25 mM sodium borate, 100 mM boricacid, 75 mM NaCl, and 5 mM EDTA.
The biological effects of increasing concentrations of granulin D peptideand antibody onin vitro cell growth rates were assayed using [3H]thymidineincorporation. Cells were grown to;60% confluence in T75 flasks (Costar)and then plated in 12-well plates (Corning) at a density of 104 cells/well in 1ml of DMEM supplemented with 10% FBS. One day after plating, the mediumwas removed and replaced with medium containing increasing concentrationsof either synthetic granulin D peptide (0–1000 ng/ml) or granulin D antibody(1:1000 to 1:100) in triplicate. Three days later, the medium was againreplaced by fresh medium and supplemented with increasing amounts ofpeptide or antibody. After 3 days, 0.5mCi/well of [3H]thymidine was added forovernight incubation at 37°C. Wells were then washed twice with 1 ml ofice-cold PBS and collected by treatment with trypsin-EDTA. Cell suspensionswere transferred to scintillation vials, and radioactivity was counted with ascintillation counter. Six separate experiments were performed on each cellline, using triplicate wells per experiment (n5 18). The Student’st test wasused to interpret the significance of differences between groups.
RESULTS
Isolation of Glioblastoma-associated Gene Products by DIA.We hypothesized that tumor-specific antibodies could be generatedand used to screen a glioblastoma cDNA expression library to identifygene products specifically expressed in brain tumors. To test thispossibility, we used human GBM tumor homogenate to immunizerabbits to obtain anti-GBM antiserum. This antiserum was passed overa GBM affinity column, and anti-GBM antibodies were eluted off.Antibodies that were not specific for tumor antigens were absorbedout by subsequent passage through a normal brain affinity column andcross-absorption against normal human plasma. The tumor-associatedantibodies were then biotinylated and used to screen a glioblastomaphagemid cDNA expression library (Fig. 1). Using this approach,which we term DIA, positive reactions were found in 28 plaques.Positive clones were isolated, subcloned into the pZL1 plasmid vectorby in vivo excision, and partially sequenced.
BLAST analysis of these sequences revealed 19 novel clones and 9genes contained in the GenBank database. Of the nine known clonesthat were identified by our DIA technique, two are known to regulategrowth, one codes for a chemotherapy resistance protein, and tworegulate gene expression (Table 1). Of the 19 novel sequences thatwere identified, Northern blot analyses were performed on 9 of thesecDNAs and differential expression was confirmed in tumor tissuesversusnormal brain in all nine of the clones tested (data not shown).
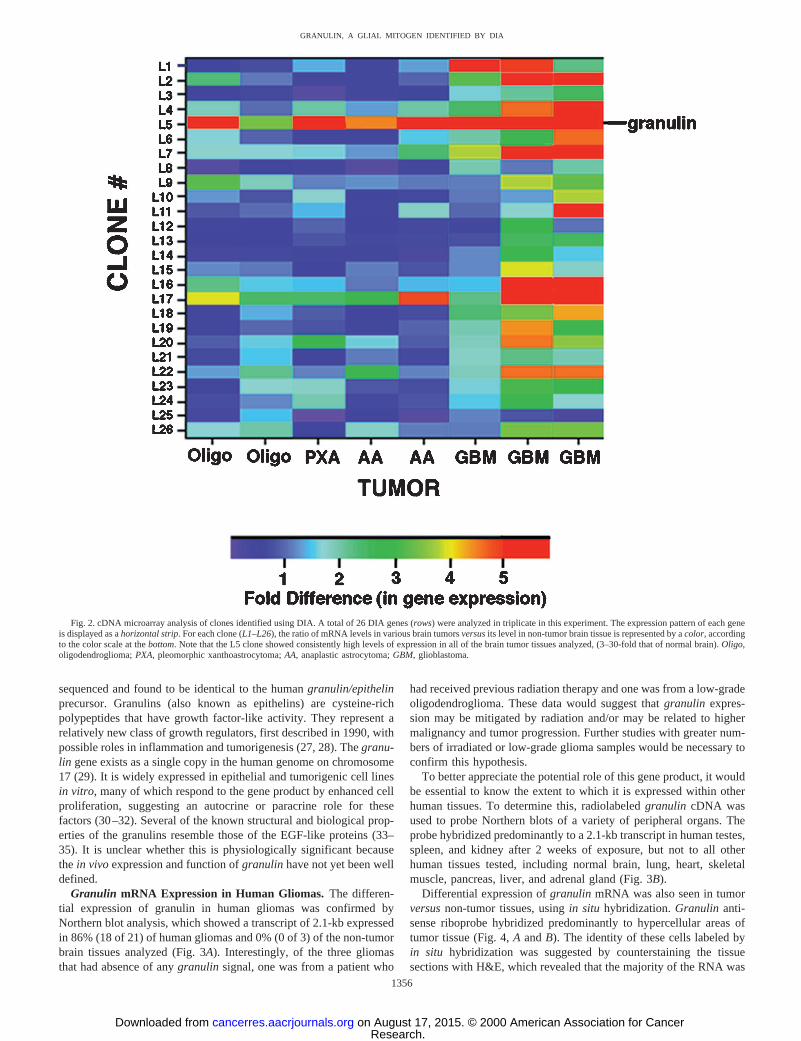
cDNA Microarray Analysis of DIA Products. To perform high-capacity screening of our DIA clones in multiple tumor and non-tumor brain tissues, we coupled our novel DIA approach with cDNAmicroarray analysis (18, 20, 21). Twenty-six of the 28 plasmid insertsisolated were successfully PCR-amplified using vector-specific prim-ers that amplify the inserted fragment from the pZL1 vector. EachPCR product was then arrayed onto glass slides and hybridized withtwo-color fluorophore-labeled probes in a manner similar to thatalready published (19). The cDNA made from each sample of tumoror normal brain mRNA was labeled with the fluorescent dye Cy3(green) and mixed with a common reference probe labeled with asecond fluorescent dye, Cy5 (red). Using this high throughput arrayer,we determined the relative abundance of each of our 26 DIA clones ineight different brain tumor samples compared with non-tumor braintissue (Fig. 2). As seen in Fig. 2, for the majority of the 26 genestested, the three glioblastoma samples analyzed appeared to havehigher levels of differential expression than the other tumor types.Furthermore, analysis of the expression of these candidate tumor-specific genes revealed that one of our isolated clones, L5, consis-tently had much higher levels of expression in gliomas compared withnormal brain (3–30-fold).
This differentially expressed clone, L5, was therefore chosen forfurther characterization. The 590-bp cDNA of clone L5 was manually
Fig. 1. Schematic diagram of strategy for DIA. GBM tumor and normal brain tissueswere each homogenized in saline, drawing off the soluble material. The insoluble materialwas then re-extracted with a small amount of 1% SDS. Aliquots of tumor homogenate andnormal brain homogenate were each linked to Sepharose, using CNBr activation (A).Theremainder of the saline-soluble GBM tumor homogenate was emulsified in completeFreund’s adjuvant and used to immunize rabbits; the detergent fraction was used to boostthe animals in incomplete Freund’s adjuvant to produce anti-GBM antiserum (B). Thisantiserum was passed through the GBM-Sepharose column, and anti-GBM antibodieswere eluted off (C). To select for anti-GBM antibodies that do not bind to normal brain,the eluate was then cross-absorbed against a normal brain affinity column (D), andanti-GBM antibodies that did not bind were collected. This final antibody preparation wasneutralized, concentrated, dialyzed, and biotinylated. These biotinylated antibodies werethen used to screen a GBM cDNA expression library transferred to nitrocellulose filterreplicas (E).
Table 1 Brain tumor-associated gene products identified by DIA
From the 28 cDNAs that were identified by this method, 9 represented genes with sequence homology to known genes in the GenBank database. Shown below are the clone numberand the known or putative function of each gene product.
Clone GenBank ID Function
L2, L6 Human GFAP Glial fibrillary acidic protein; expressed in astrocytes; used as marker for gliomasL5 Human granulin Peptide isolated from human granulocytes; autocrine growth factor for teratoma-
derived PC cells; mitogen for 3T3 fibroblastsL9 GAPDH Glyceraldehyde-3-phosphate dehydrogenase; glycolytic enzyme; increased in lung
cancer; putative role in apoptosis and neurodegenerative diseasesL10 Carbonyl reductase Enzyme involved in chemotherapy resistance and free radical modulationL19 DNA-dependent ATPase (putative) & X-linked nuclear protein Helicase involved in transcription initiation and brain differentiation (putative)L20 Homologous with tyrosine kinases ???a
L24 Hsa2 mitochondrion cytochrome oxidase ???a
L25 Osteopontin Ligand for integrin; involved in adhesion, migration, and osteoclastogenesisa Function not known.
1355
GRANULIN, A GLIAL MITOGEN IDENTIFIED BY DIA
Research. on August 17, 2015. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from
sequenced and found to be identical to the humangranulin/epithelinprecursor. Granulins (also known as epithelins) are cysteine-richpolypeptides that have growth factor-like activity. They represent arelatively new class of growth regulators, first described in 1990, withpossible roles in inflammation and tumorigenesis (27, 28). Thegranu-lin gene exists as a single copy in the human genome on chromosome17 (29). It is widely expressed in epithelial and tumorigenic cell linesin vitro, many of which respond to the gene product by enhanced cellproliferation, suggesting an autocrine or paracrine role for thesefactors (30–32). Several of the known structural and biological prop-erties of the granulins resemble those of the EGF-like proteins (33–35). It is unclear whether this is physiologically significant becausethe in vivoexpression and function ofgranulinhave not yet been welldefined.
Granulin mRNA Expression in Human Gliomas. The differen-tial expression of granulin in human gliomas was confirmed byNorthern blot analysis, which showed a transcript of 2.1-kb expressedin 86% (18 of 21) of human gliomas and 0% (0 of 3) of the non-tumorbrain tissues analyzed (Fig. 3A). Interestingly, of the three gliomasthat had absence of anygranulin signal, one was from a patient who
had received previous radiation therapy and one was from a low-gradeoligodendroglioma. These data would suggest thatgranulin expres-sion may be mitigated by radiation and/or may be related to highermalignancy and tumor progression. Further studies with greater num-bers of irradiated or low-grade glioma samples would be necessary toconfirm this hypothesis.
To better appreciate the potential role of this gene product, it wouldbe essential to know the extent to which it is expressed within otherhuman tissues. To determine this, radiolabeledgranulin cDNA wasused to probe Northern blots of a variety of peripheral organs. Theprobe hybridized predominantly to a 2.1-kb transcript in human testes,spleen, and kidney after 2 weeks of exposure, but not to all otherhuman tissues tested, including normal brain, lung, heart, skeletalmuscle, pancreas, liver, and adrenal gland (Fig. 3B).
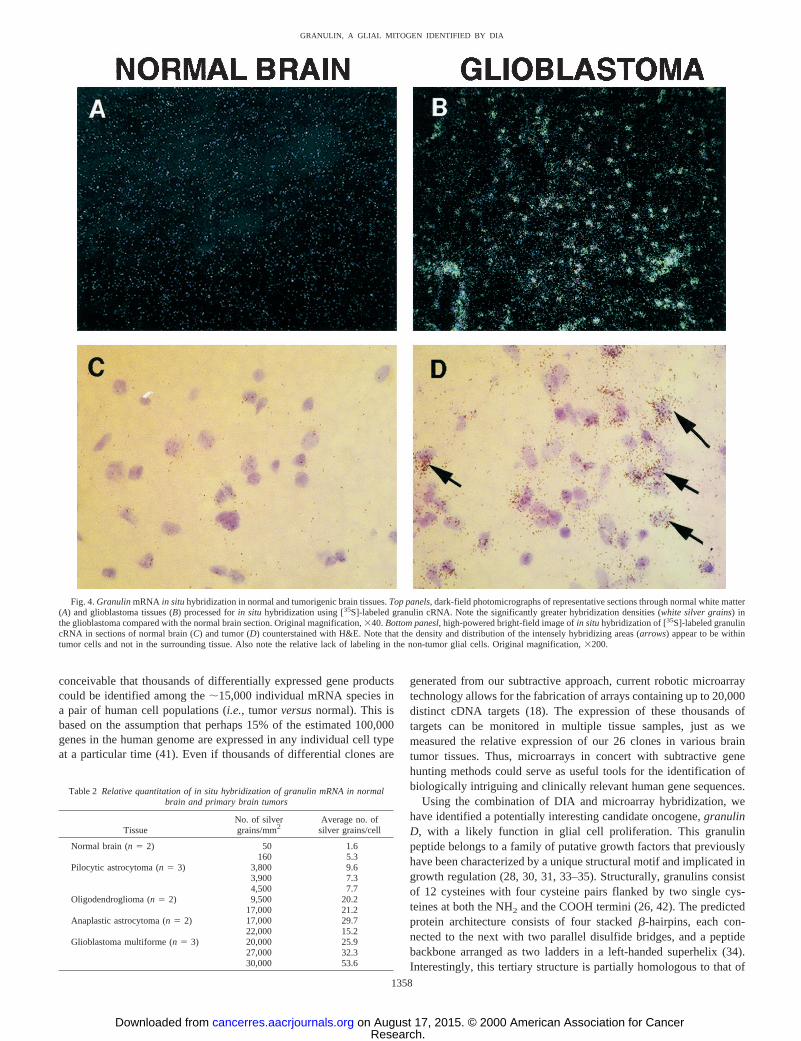
Differential expression ofgranulin mRNA was also seen in tumorversusnon-tumor tissues, usingin situ hybridization.Granulin anti-sense riboprobe hybridized predominantly to hypercellular areas oftumor tissue (Fig. 4,A andB). The identity of these cells labeled byin situ hybridization was suggested by counterstaining the tissuesections with H&E, which revealed that the majority of the RNA was
Fig. 2. cDNA microarray analysis of clones identified using DIA. A total of 26 DIA genes (rows) were analyzed in triplicate in this experiment. The expression pattern of each geneis displayed as ahorizontal strip. For each clone (L1–L26), the ratio of mRNA levels in various brain tumorsversusits level in non-tumor brain tissue is represented by acolor, accordingto the color scale at thebottom. Note that the L5 clone showed consistently high levels of expression in all of the brain tumor tissues analyzed, (3–30-fold that of normal brain).Oligo,oligodendroglioma;PXA, pleomorphic xanthoastrocytoma;AA, anaplastic astrocytoma;GBM, glioblastoma.
1356
GRANULIN, A GLIAL MITOGEN IDENTIFIED BY DIA
Research. on August 17, 2015. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from

within tumor cells and not in the tissue stroma (Fig. 4,C andD). Sensestrand riboprobe cDNA was used as a control and showed no specificlabeling (data not shown), indicating that the cellular hybridizationobtained with the antisense probe was specific for thegranulinmRNA. Quantitation ofgranulin hybridization densities was meas-ured from thein situ slides, using image analysis software. Thisanalysis revealed significantly greater numbers of silver grains withincells of the most malignant brain tumors (e.g.,anaplastic astrocytomasand GBM) compared with non-tumor glial cells (P 5 0.006), con-firming that elevated levels ofgranulin mRNA are expressed inhigh-grade primary brain tumors (Table 2).
In Vitro Growth Regulation of Glial Cells by Granulin D. Fourgranulins, A, B, C, and D, previously have been isolated from humaninflammatory cell exudates (28, 33, 36, 37). Each is a small protein of;6 kDa that is derived from a larger precursor of 593 amino acids,known asacrogranin(38–40). TheacrogranincDNA (clone L5) thatwe isolated from human glioblastomas contained the entire sequencefor granulin D (bp 1254–2099; Ref. 26).
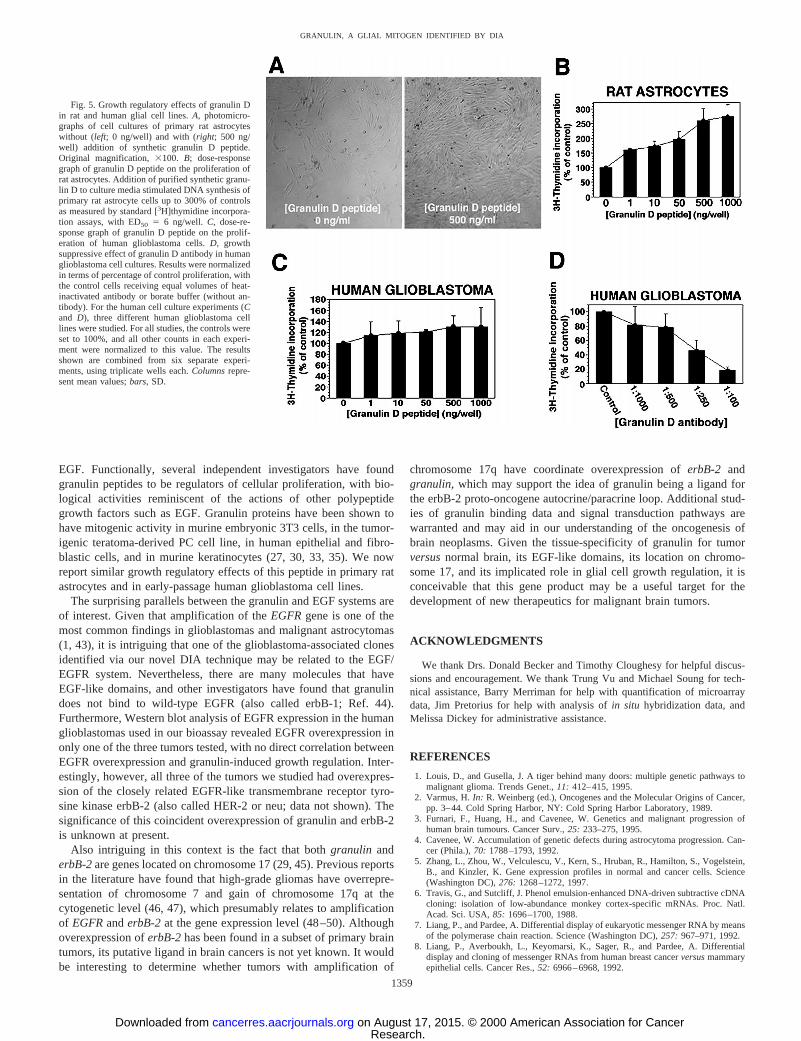
The implication of granulin molecules in growth regulation with atertiary structure reminiscent of that of EGF suggested a potentiallyimportant role for our L5/granulin D clone as a putative growth factor.To determine whether granulin D may modulate glial cell prolifera-tion, we synthesized a 55-amino acid peptide corresponding to theknown sequence of granulin D (26). We then studied the effect of thissynthetic peptide on proliferation rates of rat astrocytes in culture,using a standard [3H]thymidine incorporation assay. As shown in Fig.5, addition of synthetic granulin D peptide stimulated DNA synthesisof rat astrocytesin vitro up to 300% in a dose-dependent manner (Fig.5, A andB). Statistically significant increases in cell proliferation (upto 150% of controls) were seen with the addition of as little as 1 ng/ml(169 pM) of granulin D to cell culture (P5 0.025). Interestingly, thissynthetic peptide had a much more modest effect on the proliferationof primary human glioblastoma cells in culture, showing only a120–150% increase (P5 0.068) in growth with the addition of.1000 ng/ml (169 nM) granulin D (Fig. 5C). This may be explainedby the fact that these human cells were tumorous and already ex-pressed high levels of granulin (as shown by Northern blot andin situhybridization). Thus, the putative receptors of this potential autocrinegrowth factor may be saturated by endogenous granulin and thereby
preclude further growth stimulation by the addition of exogenouspeptide. To further evaluate the growth regulatory role of granulin Don human tumor cellsin vitro, a polyclonal antibody was raisedagainst the 55-amino acid granulin D peptide and assayed for itsability to inhibit cell proliferation in three primary human glioblas-toma cultures. As shown in Fig. 5D, the addition of increasingconcentrations of purified granulin D antibody to early-passage hu-man brain tumor cell cultures significantly inhibited cell growthinvitro. [3H]Thymidine incorporation was suppressed down to only18.6% of controls with the highest concentration of antibody tested(1:100 dilution;P 5 0.035).
DISCUSSION
The detection and characterization of a putative growth factordifferentially expressed in brain tumorsversusnon-tumor brain tissuedemonstrates the usefulness of our novel DIA technique for identifi-cation of subtractive tissue-specific gene products that may havesignificant biological activity. The DIA method described here is analternative to currently established methods for the purposes of iden-tifying differences in gene expression. It has the advantage of select-ing for gene products that are actually translated from mRNA species,which can readily be cloned and synthesized for use in functionalassays as we have described here. Furthermore, because this techniqueis based on the generation of subtractive antibodies used to screencDNA expression libraries, antibodies to clones of interest can begenerated for antibody-based studies. Because it is still speculativewhether the tumor-associated antigens we identified using this ap-proach can actually induce effective humoral or cytotoxic antitumorimmune responses, further studies are needed before such conclusionscan be made. Nevertheless, the results reported in this study demon-strate the potential of this technique to identify candidate glioma-associated peptides that readily allow study of expression patterns andbiological function.
With current microarray technology, it is feasible to screen rela-tively large numbers of tumor samples for the expression of subtrac-tive products. This allows easy discrimination of redundant clones andrapid confirmation of truly differentially expressed genes. Althoughwe isolated and screened only 26 clones from our DIA method, it is
Fig. 3. Granulin expression in human tissues.A, Northern blot analysis of granulin mRNA in normal and tumorigenic brain tissues.Lanes 1–3, nontumorigenic brain tissues takenfrom a surgical resection for epilepsy (Lane 1), surgical decompression for trauma (Lane 2), and autopsy normal brain (Lane 3).Lanes 4–24, surgically resected brain tumor tissuespathologically confirmed to be glioblastomas (WHO grade IV;Lanes 4–15), anaplastic astrocytomas (WHO grade III;Lanes 16–21), low-grade oligodendrogliomas (WHO grade II;Lane 22), or anaplastic oligodendrogliomas (WHO grade III;Lanes 23and24). All surgical specimens were snap frozen immediately in liquid nitrogen in the operating room. Thisblot was exposed for 48 h with intensifying screen.B, Northern blot analysis ofgranulin mRNA in various human peripheral organs. Tissue specimens were taken at autopsy fromnormal brain (Lane 1), lung (Lane 2), heart (Lane 3), skeletal muscle (Lane 4), pancreas (Lane 5), liver (Lane 6), testes (Lane 7), spleen (Lane 8), kidney (Lane 9), and adrenal gland(Lane 10). This blot was exposed for 2 weeks with intensifying screens. As a loading control, the same blots were reprobed with 18S cDNA and exposed for 1 h without a screen.
1357
GRANULIN, A GLIAL MITOGEN IDENTIFIED BY DIA
Research. on August 17, 2015. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from
conceivable that thousands of differentially expressed gene productscould be identified among the;15,000 individual mRNA species ina pair of human cell populations (i.e.,tumor versusnormal). This isbased on the assumption that perhaps 15% of the estimated 100,000genes in the human genome are expressed in any individual cell typeat a particular time (41). Even if thousands of differential clones are
generated from our subtractive approach, current robotic microarraytechnology allows for the fabrication of arrays containing up to 20,000distinct cDNA targets (18). The expression of these thousands oftargets can be monitored in multiple tissue samples, just as wemeasured the relative expression of our 26 clones in various braintumor tissues. Thus, microarrays in concert with subtractive genehunting methods could serve as useful tools for the identification ofbiologically intriguing and clinically relevant human gene sequences.
Using the combination of DIA and microarray hybridization, wehave identified a potentially interesting candidate oncogene,granulinD, with a likely function in glial cell proliferation. This granulinpeptide belongs to a family of putative growth factors that previouslyhave been characterized by a unique structural motif and implicated ingrowth regulation (28, 30, 31, 33–35). Structurally, granulins consistof 12 cysteines with four cysteine pairs flanked by two single cys-teines at both the NH2 and the COOH termini (26, 42). The predictedprotein architecture consists of four stackedb-hairpins, each con-nected to the next with two parallel disulfide bridges, and a peptidebackbone arranged as two ladders in a left-handed superhelix (34).Interestingly, this tertiary structure is partially homologous to that of
Fig. 4.GranulinmRNA in situhybridization in normal and tumorigenic brain tissues.Top panels, dark-field photomicrographs of representative sections through normal white matter(A) and glioblastoma tissues (B) processed forin situ hybridization using [35S]-labeled granulin cRNA. Note the significantly greater hybridization densities (white silver grains) inthe glioblastoma compared with the normal brain section. Original magnification,340.Bottom panesl, high-powered bright-field image ofin situhybridization of [35S]-labeled granulincRNA in sections of normal brain (C) and tumor (D) counterstained with H&E. Note that the density and distribution of the intensely hybridizing areas (arrows) appear to be withintumor cells and not in the surrounding tissue. Also note the relative lack of labeling in the non-tumor glial cells. Original magnification,3200.
Table 2 Relative quantitation of in situ hybridization of granulin mRNA in normalbrain and primary brain tumors
TissueNo. of silvergrains/mm2
Average no. ofsilver grains/cell
Normal brain (n5 2) 50 1.6160 5.3
Pilocytic astrocytoma (n5 3) 3,800 9.63,900 7.34,500 7.7
Oligodendroglioma (n5 2) 9,500 20.217,000 21.2
Anaplastic astrocytoma (n5 2) 17,000 29.722,000 15.2
Glioblastoma multiforme (n5 3) 20,000 25.927,000 32.330,000 53.6
1358
GRANULIN, A GLIAL MITOGEN IDENTIFIED BY DIA
Research. on August 17, 2015. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from
EGF. Functionally, several independent investigators have foundgranulin peptides to be regulators of cellular proliferation, with bio-logical activities reminiscent of the actions of other polypeptidegrowth factors such as EGF. Granulin proteins have been shown tohave mitogenic activity in murine embryonic 3T3 cells, in the tumor-igenic teratoma-derived PC cell line, in human epithelial and fibro-blastic cells, and in murine keratinocytes (27, 30, 33, 35). We nowreport similar growth regulatory effects of this peptide in primary ratastrocytes and in early-passage human glioblastoma cell lines.
The surprising parallels between the granulin and EGF systems areof interest. Given that amplification of theEGFRgene is one of themost common findings in glioblastomas and malignant astrocytomas(1, 43), it is intriguing that one of the glioblastoma-associated clonesidentified via our novel DIA technique may be related to the EGF/EGFR system. Nevertheless, there are many molecules that haveEGF-like domains, and other investigators have found that granulindoes not bind to wild-type EGFR (also called erbB-1; Ref. 44).Furthermore, Western blot analysis of EGFR expression in the humanglioblastomas used in our bioassay revealed EGFR overexpression inonly one of the three tumors tested, with no direct correlation betweenEGFR overexpression and granulin-induced growth regulation. Inter-estingly, however, all three of the tumors we studied had overexpres-sion of the closely related EGFR-like transmembrane receptor tyro-sine kinase erbB-2 (also called HER-2 or neu; data not shown).Thesignificance of this coincident overexpression of granulin and erbB-2is unknown at present.
Also intriguing in this context is the fact that bothgranulin anderbB-2are genes located on chromosome 17 (29, 45). Previous reportsin the literature have found that high-grade gliomas have overrepre-sentation of chromosome 7 and gain of chromosome 17q at thecytogenetic level (46, 47), which presumably relates to amplificationof EGFRanderbB-2at the gene expression level (48–50). Althoughoverexpression oferbB-2has been found in a subset of primary braintumors, its putative ligand in brain cancers is not yet known. It wouldbe interesting to determine whether tumors with amplification of
chromosome 17q have coordinate overexpression oferbB-2 andgranulin, which may support the idea of granulin being a ligand forthe erbB-2 proto-oncogene autocrine/paracrine loop. Additional stud-ies of granulin binding data and signal transduction pathways arewarranted and may aid in our understanding of the oncogenesis ofbrain neoplasms. Given the tissue-specificity of granulin for tumorversusnormal brain, its EGF-like domains, its location on chromo-some 17, and its implicated role in glial cell growth regulation, it isconceivable that this gene product may be a useful target for thedevelopment of new therapeutics for malignant brain tumors.
ACKNOWLEDGMENTS
We thank Drs. Donald Becker and Timothy Cloughesy for helpful discus-sions and encouragement. We thank Trung Vu and Michael Soung for tech-nical assistance, Barry Merriman for help with quantification of microarraydata, Jim Pretorius for help with analysis ofin situ hybridization data, andMelissa Dickey for administrative assistance.
REFERENCES
1. Louis, D., and Gusella, J. A tiger behind many doors: multiple genetic pathways tomalignant glioma. Trends Genet.,11: 412–415, 1995.
2. Varmus, H.In: R. Weinberg (ed.), Oncogenes and the Molecular Origins of Cancer,pp. 3–44. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory, 1989.
3. Furnari, F., Huang, H., and Cavenee, W. Genetics and malignant progression ofhuman brain tumours. Cancer Surv.,25: 233–275, 1995.
4. Cavenee, W. Accumulation of genetic defects during astrocytoma progression. Can-cer (Phila.),70: 1788–1793, 1992.
5. Zhang, L., Zhou, W., Velculescu, V., Kern, S., Hruban, R., Hamilton, S., Vogelstein,B., and Kinzler, K. Gene expression profiles in normal and cancer cells. Science(Washington DC),276: 1268–1272, 1997.
6. Travis, G., and Sutcliff, J. Phenol emulsion-enhanced DNA-driven subtractive cDNAcloning: isolation of low-abundance monkey cortex-specific mRNAs. Proc. Natl.Acad. Sci. USA,85: 1696–1700, 1988.
7. Liang, P., and Pardee, A. Differential display of eukaryotic messenger RNA by meansof the polymerase chain reaction. Science (Washington DC),257: 967–971, 1992.
8. Liang, P., Averboukh, L., Keyomarsi, K., Sager, R., and Pardee, A. Differentialdisplay and cloning of messenger RNAs from human breast cancerversusmammaryepithelial cells. Cancer Res.,52: 6966–6968, 1992.
Fig. 5. Growth regulatory effects of granulin Din rat and human glial cell lines.A, photomicro-graphs of cell cultures of primary rat astrocyteswithout (left; 0 ng/well) and with (right; 500 ng/well) addition of synthetic granulin D peptide.Original magnification,3100. B; dose-responsegraph of granulin D peptide on the proliferation ofrat astrocytes. Addition of purified synthetic granu-lin D to culture media stimulated DNA synthesis ofprimary rat astrocyte cells up to 300% of controlsas measured by standard [3H]thymidine incorpora-tion assays, with ED50 5 6 ng/well. C, dose-re-sponse graph of granulin D peptide on the prolif-eration of human glioblastoma cells.D, growthsuppressive effect of granulin D antibody in humanglioblastoma cell cultures. Results were normalizedin terms of percentage of control proliferation, withthe control cells receiving equal volumes of heat-inactivated antibody or borate buffer (without an-tibody). For the human cell culture experiments (Cand D), three different human glioblastoma celllines were studied. For all studies, the controls wereset to 100%, and all other counts in each experi-ment were normalized to this value. The resultsshown are combined from six separate experi-ments, using triplicate wells each.Columnsrepre-sent mean values;bars, SD.
1359
GRANULIN, A GLIAL MITOGEN IDENTIFIED BY DIA
Research. on August 17, 2015. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from

9. Linskens, M., Feng, J., Andrews, W., Enlow, B., Saati, S., Tonkin, L., Funk, W., andVilleponteau, B. Cataloging altered gene expression in young and senescent cellsusing enhanced differential display. Nucleic Acids Res.,23: 3244–3251, 1995.
10. Uchiyama, C., Zhu, J., Carroll, R., Leon, S., and Black, P. Differential display ofmessenger ribonucleic acid: a useful technique for analyzing differential gene ex-pression in human brain tumors. Neurosurgery,37: 464–469, 1995.
11. Braun, B., Frieden, R., Lessnick, S., May, W., and Denny, C. Identification of targetgenes for the Ewing’s sarcoma EWS/FL1 fusion protein by representational differ-ence analysis. Mol. Cell. Biol.,15: 4623–4630, 1995.
12. Hubank, M., and Schatz, D. Identifying differences in mRNA expression by repre-sentational difference analysis of cDNA. Nucleic Acids Res.,22: 5640–5648, 1994.
13. Lisitsyn, N., Leach, F., Vogelstein, B., and Wigler, M. Detection of genetic loss intumors by representational difference analysis. Cold Spring Harbor Symp. Quant.Biol., 59: 585–587, 1994.
14. Lucas, S., De Smet, C., Arden, K., Viars, C., Lethae, B., Lurquin, C., and Boon, T.Identification of a newMAGE gene with tumor-specific expression by representa-tional difference analysis. Cancer Res.,58: 743–752, 1998.
15. Velculescu, V., Zhang, L., Vogelstein, B., and Kinzler, K. Serial analysis of geneexpression. Science (Washington DC),270: 484–487, 1995.
16. Diatchenko, L., Lau, Y., Campbell, A., Chenchik, A., Moqadam, F., Huang, B.,Lukyanov, S., Lukyanov, K., Gurskaya, N., Sverdlov, E., and Siebert, P. Suppressionsubtractive hybridization: a method for generating differentially regulated or tissue-specific cDNA probes and libraries. Proc. Natl. Acad. Sci. USA,93: 6025–6030,1996.
17. Diatchenko, L., Lukyanov, S., Lau, Y., and Siebert, P. Suppression subtractivehybridization: a versatile method for identifying differentially expressed genes. Meth-ods Enzymol.,303: 349–380, 1999.
18. Schena, M., Shalon, D., Davis, R., and Brown, P. Quantitative monitoring of geneexpression patterns with a complementary DNA microarray. Science (WashingtonDC), 270: 467–470, 1995.
19. Welford, S., Gregg, J., Chen, E., Garrison, D., Sorensen, P., Denny, C., and Nelson,S. Detection of differentially expressed genes in primary tumor tissues using repre-sentational differences analysis coupled to microarray hybridization. Nucleic AcidsRes.,26: 3059–3065, 1998.
20. DeRisi, J., Iyer, V., and Brown, P. Exploring the metabolic and genetic control ofgene expression on a genomic scale. Science (Washington DC),278:680–686, 1997.
21. Iyer, V., Eisen, M., Ross, D., Schuler, G., Moore, T., Lee, J., Trent, J., Staudt, L.,Hudson, J., Boguski, M., Lashkari, D., Shalon, D., Botstein, D., and Brown, P. Thetranscriptional program in the response of human fibroblasts to serum. Science(Washington DC),283: 83–87, 1999.
22. Kornblum, H., Chugani, H., Tatsukawa, K., and Gall, C. Cerebral hemidecorticationalters expression of transforming growth factor-a mRNA in the neostriatum ofdeveloping rats. Mol. Brain Res.,21: 107–114, 1994.
23. Mathern, G., Pretorius, J., Mendoza, D., Lozada, A., and Kornblum, H. HippocampalAMPA and NMDA mRNA levels correlate with aberrant fascia dentata mossy fibersprouting in the pilocarpine model of spontaneous limbic epilepsy. J. Neurosci. Res.,54: 734–753, 1998.
24. Kumar, S., Holmes, E., Scully, S., Birren, B., Wilson, R., and de Vellis, J. Thehormonal regulation of gene expression of glial markers: glutamine synthetase andglycerol phosphate dehydrogenase in primary cultures of rat brain and in C6 cell line.J. Neurosci. Res.,16: 251–264, 1986.
25. Estes, M., Ransohoff, R., McMahon, J., Jacobs, B., and Barna, B. Characterization ofadult human astrocytes derived from explant culture. J. Neurosci. Res.,27: 697–705,1990.
26. Bhandari, V., Palfree, R., and Bateman, A. Isolation and sequence of the granulinprecursor cDNA from human bone marrow reveals tandem cysteine-rich granulindomains. Proc. Natl. Acad. Sci. USA,89: 1715–1719, 1992.
27. Shoyab, M., McDonald, V., Byles, C., Todaro, G., and Plowman, G. Epithelins 1 and2: isolation and characterization of two cysteine-rich growth-modulating proteins.Proc. Natl. Acad. Sci. USA,87: 7912–7916, 1990.
28. Bateman, A., Belcourt, D., Bennett, H., Lazure, C., and Solomon, S. Granulins, anovel class of peptide from leukocytes. Biochem. Biophys. Res. Commun.,173:1161–1168, 1990.
29. Bhandari, V., and Bateman, A. Structure and chromosomal location of the humangranulin gene. Biochem. Biophys. Res. Commun.,188: 57–63, 1992.
30. Zhou, J., Gao, G., Crabb, J., and Serrero, G. Purification of an autocrine growth factorhomologous with mouse epithelin from a highly tumorigenic cell line. J. Biol. Chem.,268: 10863–10869, 1993.
31. Zhang, H., and Serrero, G. Inhibition of tumorigenicity of the teratoma PC cell lineby transfection with antisense cDNA for PC cell-derived growth factor (PCDGF,epithelin/granulin precursor). Proc. Natl. Acad. Sci. USA,95: 14202–14207, 1998.
32. Bhandari, V., Giaid, A., and Bateman, A. The complementary deoxyribonucleic acidsequence, tissue distribution, and cellular localization of the rat granulin precursor.Endocrinology,133: 2683–2689, 1993.
33. Bateman, A., and Bennett, H. Granulins: the structure and function of an emergingfamily of growth factors. J. Endocrinol.,158: 145–151, 1998.
34. Hrabal, R., Chen, Z., James, S., Bennett, H., and Ni, F. The hairpin stack fold, a novelprotein architecture for a new family of protein growth factors. Nat. Struct. Biol.,3:747–752, 1996.
35. Xu, S., Tang, D., Chamberlain, S., Pronk, G., Masiarz, F., Kaur, S., Prisco, M.,Zanocco-Marani, T., and Baserga, R. The granulin/epithelin precursor abrogates therequirement for the insulin-like growth factor 1 receptor for growthin vitro. J. Biol.Chem.,273: 20078–20083, 1998.
36. Belcourt, D., Lazure, C., and Bennett, H. Isolation and primary structure of the threemajor forms of granulin-like peptides from hematopoietic tissues of a teleost fish(Cyprinus carpio). J. Biol. Chem.,268: 9230–9237, 1993.
37. Belcourt, D., Okawara, Y., Fryer, J., and Bennett, H. Immunocytochemical localiza-tion of granulin-1 to mononuclear phagocytic cells of the teleost fishCyprinus carpioandCarassius auratus. J. Leukoc. Biol.,57: 94–100, 1995.
38. Baba, T., Hoff, H., Nemoto, H., Lee, H., Orth, J., Arai, Y., and Gerton, G. Acrogranin,an acrosomal cysteine-rich glycoprotein, is the precursor of the growth-modulatingpeptides, granulins, and epithelins, and is expressed in somatic as well as male germcells. Mol. Reprod. Dev.,34: 233–243, 1993.
39. Baba, T., Nemoto, H., Watanabe, K., Arai, Y., and Gerton, G. Exon/intron organi-zation of the gene encoding the mouse epithelin/granulin precursor (acrogranin).FEBS Lett.,322: 89–94, 1993.
40. Trinh, D., Brown, K., and Jeang, K. Epithelin/granulin growth factors: extracellularcofactors for HIV-1 and HIV-2 Tat proteins. Biochem. Biophys. Res. Commun.,256:299–306, 1999.
41. Boguski, M., and Schuler, G. Establishing a human transcript map. Nat. Genet.,10:369–371, 1995.
42. Bhandari, V., Daniel, R., Lim, P., and Bateman, A. Structural and functional analysisof a promoter of the human granulin/epithelin gene. Biochem. J.,319: 441–447,1996.
43. Nagane, M., Huang, H-J., and Cavenee, W. Advances in the molecular genetics ofgliomas. Curr. Opin. Oncol.,9: 215–222, 1997.
44. Culouscou, J-M., Carlton, G., and Shoyab, M. Biochemical analysis of the epithelinreceptor. J. Biol. Chem.,268: 10458–10462, 1993.
45. Maguire, H. J., and Greene, M. Theneu (c-erbB-2) oncogene. Semin. Oncol.,16:148–155, 1989.
46. Kasai, H., Imahori, T., and Kawamoto, K. Detection of chromosomal numericalaberration in glioma by FISH. Hum. Cell. (Tokyo),6: 62–65, 1993.
47. Arnoldus, E., Wolters, L., Voormolen, J., van Duinen, S., Raap, A., van der Ploeg, M.,and Peters, A. Interphase cytogenetics: a new tool for the study of genetic changes inbrain tumors. J. Neurosurg.,76: 997–1003, 1992.
48. Dietzmann, K., and von Bossanyi, P. Coexpression of epidermal growth factorreceptor protein and c-erbB-2 oncoprotein in human astrocytic tumors. An immuno-histochemical study. Zentbl. Pathol.,140: 335–341, 1994.
49. Engelhard, H., Wolters, M., and Criswell, P. Analysis of c-erbB2 protein content inhuman glioma cells and tumor tissue. J. Neuro-oncol.,23: 31–40, 1995.
50. von Bossanyi, P., Sallaba, J., Dietzmann, K., Warich-Kirches, M., and Kirches, E.Correlation of TGF-a and EGF-receptor expression with proliferative activity inhuman astrocytic gliomas. Pathol. Res. Pract.,194: 141–147, 1998.
1360
GRANULIN, A GLIAL MITOGEN IDENTIFIED BY DIA
Research. on August 17, 2015. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from

2000;60:1353-1360. Cancer Res Linda M. Liau, Roger L. Lallone, Robert S. Seitz, et al.
, Using Differential Immuno-absorption granulinGene, Identification of a Human Glioma-associated Growth Factor
Updated version
http://cancerres.aacrjournals.org/content/60/5/1353
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/60/5/1353.full.html#ref-list-1
This article cites 46 articles, 24 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/60/5/1353.full.html#related-urls
This article has been cited by 25 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on August 17, 2015. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from
Related Documents











