Title Identification and Molecular Characterization of Bioactive Molecules Expressed in Salivary Glands of the Hard Tick,Haemaphysalis longicornis( 本文(FULLTEXT) ) Author(s) HARNNOI, Thasaneeya Report No.(Doctoral Degree) 博士(獣医学) 乙第084号 Issue Date 2007-09-14 Type 博士論文 Version author URL http://hdl.handle.net/20.500.12099/23174 ※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

TitleIdentification and Molecular Characterization of BioactiveMolecules Expressed in Salivary Glands of the HardTick,Haemaphysalis longicornis( 本文(FULLTEXT) )
Author(s) HARNNOI, Thasaneeya
Report No.(DoctoralDegree) 博士(獣医学) 乙第084号
Issue Date 2007-09-14
Type 博士論文
Version author
URL http://hdl.handle.net/20.500.12099/23174
※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。

Identification and Molecular Characterization of Bioactive Molecules
Expressed in Salivary Glands of the Hard Tick, Haemaphysalis longicornis
(フタトゲチマダニの唾液腺に発現する生理活性分子の同定
および機能解明に関する研究)
2007
The United Graduate School of Veterinary Sciences, Gifu University
(Obihiro University of Agriculture and Veterinary Medicine)
HARNNOI, Thasaneeya

Identification and Molecular Characterization of Bioactive Molecules
Expressed in Salivary Glands of the Hard Tick, Haemaphysalis longicornis
(フタトゲチマダニの唾液腺に発現する生理活性分子の同定
および機能解明に関する研究)
HARNNOI, Thasaneeya

I
Contents
Contents I-II
Abbreviations III-V
General introduction 1-5
Chapter 1
Exploring bioactive molecules from salivary gland cDNA library of Haemaphysalis
longicornis
1-1. Introduction 6-8
1-2. Materials and Methods 8-11
1-3. Results 11-14
1-4. Discussion 14-17
1-5. Summary 17-18
Chapter 2
Molecular characterization and comparative study of 6 salivary gland
metalloproteases from the hard tick, Haemaphysalis longicornis
2-1. Introduction 26-28
2-2. Materials and Methods 28-35
2-3. Results 35-38
2-4. Discussion 38-42
2-5. Summary 42-43

II
Chapter 3
Identification of genes encoding cement-like antigens expressed in the salivary
glands of Haemaphysalis longicornis
3-1. Introduction 55-56
3-2. Materials and Methods 56-58
3-3. Results 58-60
3-4. Discussion 60-63
3-5. Summary 63-64
Chapter 4
Characterization of Haemaphysalis longicornis recombinant cement-like antigens
and evaluation of their vaccination effects
4-1. Introduction 69-71
4-2. Materials and Methods 71-76
4-3. Results 76-79
4-4. Discussion 80-83
4-5. Summary 83
General discussion 95-100
Conclusion 101-104
Acknowledgements 105-106
References 107-125

III
Abbreviations
A ADAM: a disintegrin and metalloprotease
B BCIP: 5-Bromo-4-chloro-3-indolyl Phosphate
C cDNA: complementary DNA
CIAP: calf intestinal phosphatase
C-terminal: carboxy terminal
D DAB: 3, 3’-Diaminobenzidine
DMSO: dimethylsulfoxide
DNA: deoxyribonucleic acid
E EDTA: ethylenediaminetetraacetic acid
EST: expressed sequence tag
G GST: glutathione S-transferase
H hlMP: H. longicornis metalloprotease
HRP: horseradish peroxidase
I IFAT: indirect fluorescent antibody test
Ig: immunoglobulin
IPTG: isopropyl-β-D(-)-thiogalactopyranoside
M MP: metalloprotease
Mr: marker
mRNA: messenger RNA
N NBT: nitroblue tetrazolium
N-terminal: amino terminal
NZY agar

IV
O ORF: open reading frame
P PBS: phosphate buffered saline
PCR: polymerase chain reaction
pfu: plaque forming unit
PI: propidium iodide
poly(A)+ RNA: polyadenylated RNA
PVDF: polyvinylidine difluoride
R RGD: Arginine-Glycine-Aspartate
RIM 36: Rhipicephalus immunodominant molecule 36
RNA: ribonucleic acid
RT: room temperature
RT-PCR: reverse-transcription-polymerase chain reaction
S SDS: sodium dodecyl sulfate
SDS-PAGE: SDS-polyacrylamide gel electrophoresis
SE: standard error
SVMP: snake venom metalloprotease
T TFPI: tissue factor pathway inhibitor
TAE: Tris-acetic acid-EDTA
TBE: Tris-Borate EDTA
TBS: Tris-buffered saline
X X-gal: 5-Bromo-4-chloro-3-indolyl-β-D-galactopyranoside

V
Unit abbreviation
D oC: degree
H hr: hour
K kDa: kilo Dalton
kbp: kilo base pair
M M: mol/liter
μg: microgram
mg: milligram
min: minute
ml: milliliter
mmol: millimol
mM: milliM
N ng: nanogram
nm: nanometer
S s: second
μ μg: microgram
μl: microliter

1
General introduction
Ticks are blood feeding ectoparasite of mammals, birds and reptiles throughout
the world. Approximately 900 species have been described worldwide (9, 53). There
are two well established families of ticks, the Ixodidae (hard ticks), and Argasidae (soft
ticks). Both are important vectors of disease causing agents to humans and animals
throughout the world. A third family, the Nuttalielidae, contains only a single species.
Public health importance of ticks resides in their ability to transmit a greater variety of
infectious agents to humans and other animal species than any other blood-feeding
arthropods. They are also directly responsible for damage to hides and lost production
in livestock. The economic impact of ticks together with costs of control measures has
been estimated at between 14-18.7 billion USD globally in the livestock sector (23, 58).
Emerging tick-borne infectious agents include arboviruses, spotted fever group
rickettsias, ehrlichias, anaplasmas, and spirochetes (110, 124, 125). In addition, the
zoonotic potential of tick-transmitted disease of companion animals, livestock and
wildlife cannot be ignored (22).
Haemaphysalis longicornis Neumann, 1901 belonging to the Metastriata is
characterized by having both the parthenogenetic and the bisexual races (36, 52, 66).
As well as other ixodid ticks, H. longicornis has 4 stages including embryonated egg,
larva, nymph and adult. H. longicornis is distributed mainly in northeastern areas of
both Russia and China, the Korean peninsula, Japan, Australia, New Zealand, New
Caledonia and the Fiji Islands (52). This species is known as a vector of the rickettsia
causing Q-fever, viruses causing Russian spring-summer encephalistis and protozoa
causing theileriosis and babesiosis (10, 52). In Japan, H. longicornis is the most

2
abundant species in Japanese pastures that transmits bovine theileriosis caused by
Theileria bufferi/orientalis, bovine babesiosis caused by Babesia ovata among grazing
cattle (37, 38, 56) and canine babesiosis caused by Babesia gibsoni (51). Rodriguez
and coworkers also inferred the potential role of H. longicornis in the transmission of
equine babesiosis caused by Babesia cabali (99).
Ticks are pool feeder, ripping and tearing the delicate membranes and tiny
blood vessels of the dermis and sucking the fluids that are exuded into the wound.
Within 5-30 min, cement is secreted into the wound (109). This material harden quickly
into a latex-like covering around the mouth parts, which enable ticks to remain attached
to the host during the prolonged feeding period of 4-8 days and prevent host immune
response molecules from coming into contact with the tick proboscis (14). Cement
cone proteins represent candidates for inclusion in vaccine against ticks, or pathogen
they transmit, since the formation of the cone is essential for tick attachment and
feeding.
Tick feeding success is dependent upon the parasite’s ability to overcome three
components of the host defense system, namely (I) hemostasis, (II) inflammatory
response and (III) cell mediated immunity, especially the cell mediated cutaneous
basophil hypersensitivity (109). During probing for host blood, ticks cause physical
trauma to skin and small blood vessels will suffer extent of damage varying with the
tick species. The normal physiological response to disruption of blood vessels is
hemostasis, which involves coordinated interaction among platelets, coagulation
pathway proteins and endothelial cells (39). Ticks are able to prevent blood coagulation
at their feeding sites, ensuring continuous blood flow during the feeding period. Several
anticoagulant proteins from saliva have been described (71), some inhibit the extrinsic

3
and intrinsic pathway of blood coagulation (45, 70) and others inhibit platelet
aggregation (60, 62, 126). To deal with the inflammatory response, ticks secret several
anti-inflammatory and immunomodulatory components through their saliva such as
anticomplement or protease inhibitors (13, 69, 116). It is well known that ticks can
modulate both innate and acquired host immunity (129). Ticks are thought to have
evolved saliva that inhibits the cutaneous immune responses of their most common
host/s (92). Based on studies involving a variety of ixodid tick species and vertebrate
host species, elements of host immune defenses modulated by ticks include: inactivation
of complement components; diminished killing activity of NK cells; reduced antibody
responses to heterologous immunogens; inhibition of T-lymphocyte in vitro
proliferation induced by mitogens; suppression of production of pro-inflammatory
cytokines by macrophages; and suppression of Th1 cytokines concomitant with up-
regulation of Th2 cytokines (103).
Besides the essential role in blood feeding, tick saliva is also important in the
transmission of tick-borne pathogens (131). Some research has demonstrated that
components secreted from salivary glands can facilitate pathogen transmission to
vertebrate hosts (57). There is some evidence suggesting that animals exposed to tick
saliva were protected against parasite infections (11, 79). Factors that might affect
pathogen transmission and establishment in host repeatedly infested with pathogen-free
tick include alteration in the cutaneous environment at the tick attachment site, which
would interrupt feeding and/or be deleterious to introduced pathogens, reduction of
duration of attachment to the host (130). These evidences indicated that salivary protein
may be used as an immunogen to control pathogen transmission.

4
Control of tick infestations has been difficult because ticks have few natural
enemies. A major control component of integrated tick control methods is the use of
acaricides. However, application of acaricides has had limited efficacy in reducing tick
infestations and is often accompanied by serious drawbacks, including the selection of
acaricide-resistant ticks, environmental contamination and contamination of milk and
meat products with drug residues (46). Alternative approaches for tick control involve
the use of hosts with natural resistance to tick. Vaccines have been developed that
induce immunological protection of vertebrate hosts against tick infestation. Two main
approaches have been considered for tick vaccine development: the use of exposed
antigens and the use of concealed antigens (134, 140). Exposed antigens are tick
proteins injected into the host, which interact with the host defense system during the
course of tick feeding. Concealed antigens are hidden antigens that not exposed to the
host immune system. A vaccine for tick population control based on hidden gut antigen
has already been commercialized, but it is not boosted by tick challenge in the field
(135). Thus there is interest in the evaluation of salivary gland proteins that are exposed
to the host during tick feeding. Novel strategies are being sought to control tick and
tick-borne diseases including transmission-blocking vaccines that target tick saliva
proteins essential for pathogen transmission and/or establishment.
Success of host vaccination will depend on the identification, cloning and
expression of key physiological tick molecules. Recent advances in genomics and
proteomics studies on the transcriptmoes and proteomes of blood feeding arthropods
have provided important new insights into the complex pharmacology of vector saliva.
The discovery of tick salivary proteins has been greatly increased by novel molecular
biology techniques and bioinformatics analysis (95). Identification of the transcripts

5
and proteins present in the salivary gland of hard ticks such as Ixodes scapularis (97,
118), Rhipicephalus (Boophilus) microplus (101), Ixodes ricinus (117), Ixodes pacificus
(34), Amblyoma americanum (15) and Dermacentor andersoni (2, 15) have been
identified recently. Host-vector-pathogen interaction has been studied through the
identification of genes differentially expressed in salivary glands of female
Rhipicephalus appendiculatus infected with Theileria parva (81) and in I. ricinus and I.
scapularis salivary glands in response to the blood feeding (69, 143).
The identification of tick-protective antigens remains the limiting step in the
development of effective tick vaccines. Despite recent advances in the identification of
tick-protective antigens, few antigens have been tested as recombinant vaccines against
tick infestation (26). The aim of this study focused on identification, molecular cloning
and characterization of salivary gland proteins of H. longicornis for development of tick
control strategies. The objectives of this study are summarized as follows: (I) to
identify bioactive molecules or protective antigen that were expressed during blood
feeding by random sequencing of cDNA library from salivary glands of H. longicornis,
(II) to further characterize one selected bioactive molecule, metalloprotease that was
identified from the library, (III) to identify immunodominant antigens from cDNA
library by immunoscreening technique and (IV) to evaluate the vaccine potency of
immunoscreening positive clones.

6
Chapter 1
Exploring bioactive molecules from salivary gland cDNA library of
hard tick, Haemaphysalis longicornis
1-1. Introduction
Ticks are medically important vectors of human and animal diseases. They are
best known because of their ability to transmit a variety of pathogenic organisms,
including fungi, viruses, rickettsia, bacteria, and protozoa. Unlike most hematophagous
arthropods, which engorge rapidly, ticks remain on the host for a long time to complete
their blood feeding. Tick saliva contains a mixture of peptides and proteins serving a
variety of functions that are essential for creating and maintaining the blood pool or the
feeding lesion in host skin. Several bioactive molecules in tick saliva that affect the
host’s hemostatic, inflammatory, and immune system have been extensively studied
(93-95, 128). The proteins possessed anti-hemostatic activity including vasodilators,
inhibitors of platelet aggregation and blood-coagulation cascade have been identified
and characterized (30, 91, 116). Currently, post-transcriptional gene silencing using
RNA interference (RNAi) is a useful tool to elucidate the gene function. The first
application of RNAi in tick research has been demonstrated in a histamine binding
protein of A. americanum (3). By using RNAi for silencing gene encoding for salivary
anticoagulant, disruption of I. scapularis anticoagulation was demonstrated (78).
Ticks have also evolved a strategy to deal with the inflammatory response of the
host following tissue injury using several components such as anticomplement or
protease inhibitors (13, 69, 116). It is well known that ticks can modulate both innate
and acquired host immunity (129). Molecules in tick saliva, which possess

7
immunomodulatory activity that is important for survival and transmission of
pathogenic infectious agents (129) were identified (1). The characterization of a
recombinant immunomodulatory protein from the salivary glands of D. andersoni
suggested that p36 recombinant protein expressed in insect cells could suppress T-
lymphocyte-mitogen-driven in vitro proliferation of splenocytes from tick-naïve mice.
Saliva is also important in transmission of tick-borne pathogens, as it may enhance
pathogen transmission (131).
Currently, EST databases from salivary glands libraries of R. appendiculatus
uninfected and infected with T. parva were established (81). No major differences were
observed when these 2 libraries were compared, but there was evidence of up-regulation
of some proteins, such as a glycine-rich protein, in infected salivary glands. These data
provided information for the construction of a DNA microarray, which would be a
useful tool for probing vector-pathogen interactions.
The development of new technologies and approaches in the field of molecular
biology has shed light on the study of salivary molecules. The discovery of novel
proteins and genes expressed in salivary glands was accomplished by massive
sequencing of full-length cDNA libraries together with approaches involving
proteomics and functional genomics. A catalog of the transcripts and proteins from the
salivary glands of the malaria vector Anopheles gambiae has been published (31),
becoming an invaluable tool to study parasite-host and -vector interactions. The same
approach has been applied to identify a sialome of other vectors of disease (96, 119,
120). The same authors also studied the salivary compounds of I. scapularis by mass
sequencing of clones from salivary glands in the cDNA library (118). Random
sequencing of 735 clones yielded 410 cDNA clusters. One hundred clusters have been

8
identified as probable secretory products. Several novel sequences may have biological
activity and possibly serve as vaccine targets.
The hard tick, H. longicornis, which commonly infests cattle and dogs, is a
major vector that transmits pathogens such as Babesia, Theileria, and Borrelia to
domestic and wild animals in Japan and other East Asian countries (44). In this
research, the bioactive molecules produced from the salivary glands of the ticks are of
particular interest. A cDNA library from the salivary glands of H. longicornis was
constructed. Random sequencing of cDNA clones was used as a tool to recruit
biologically important key molecules that might be useful for the development of tick
control strategies.
1-2. Materials and Methods
Ticks
The parthenogenetic Okayama strain of H. longicornis (36) has been maintained
by feeding on rabbits and mice for several generations in our laboratory since 1997.
Extraction of total RNA and poly (A)+ RNA
Salivary glands from 160 adult ticks fed on rabbits for 3 days were homogenized
using a homogenizer (Ultra Turrax T8; IKA Labortechnik, Hohenstaufen, Germany) in a
Tri reagent (Sigma, St Louis, MO, U.S.A). The total RNA was subsequently isolated
according to the manufacturer’s instructions. Approximately 600 μg of total RNA was
obtained from 320 pairs of salivary glands. The poly (A)+ RNA was purified using an
Oligotex-dT30 (Super) mRNA isolation kit (Takara, Tokyo, Japan). The yield of
purified mRNA was 1% of the total RNA.

9
Construction of a cDNA library
The construction of the cDNA library was performed with 5 μg of mRNA using
a ZAP-cDNA® synthesis kit (Stratagene, CA, USA) according to the manufacturer’s
instructions. After size fractionation, the cDNA was ligated into an EcoRI/XhoI-ended
UniZap XR® vector. The ligation product was packaged using a ZAP-cDNA®
Gigapack® III packaging extract (Stratagene, CA, USA). Plating and titering of the
primary library was performed to determine the titer and ratio of the recombinant/non-
recombinant clones. The construction of the cDNA library yielded 1.65 X 105 pfu of the
primary library. The determination of the ratio of recombinant to non-recombinant
clones with the use of 5-Bromo-4-chloro-3-indolyl-β-D-galactopyranoside (X-gal) and
Isopropyl-1-thio-β-D-thiogalactopyranoside (IPTG) showed that the library contained
1.61 X 105 recombinant clones and 3,500 non-recombinant clones. Half the amount of
the primary library was used for further amplification. The amplified library with 4.0
X109 pfu/ml was aliquoted and kept at –85°C in 7% dimethylsulfoxide (DMSO).
Random sequencing of the H. longicornis cDNA library
The recombinant phage clones from the primary library were randomly selected
for sequencing. Four hundred and thirty-two white plaques were randomly selected for
PCR amplification. Two microliters of the phage suspension was used as a template for
PCR using T3 and T7 vector-specific primers flanking the cDNA inserts. The condition
of PCR was 30 cycles of denaturation at 94°C for 1 min, annealing at 50°C for 1 min,
and extension at 72°C for 2 min. The PCR products were analyzed by 1.5% Tris-Borate
EDTA (TBE) agarose gel electrophoresis. The clones containing large inserts were
selected for sequencing. After an in vivo excision experiment was performed, in order

10
to transfer the cDNA inserts from the lambda phage system to pBluescript phagemid,
plasmid DNA was further purified using a Qiagen mini kit for DNA sequencing.
Nucleotide sequences of cDNA fragments were determined using a BigDye terminator
cycle sequencing kit (PerkinElmer Life Sciences) and an automated sequencer (ABI
PRISM 310 Genetic Analyzer).
DNA sequence analysis
The sequences obtained by using vector-specific T3 and RPV primers were
analyzed. Primer and vector sequences were removed from raw sequences. The
nucleotide sequences were translated into amino acid sequences using an ExPASy
server (40) (http://www.expasy.org). The deduced amino acid sequences were
submitted to a similarity search against a non-redundant protein database at the National
Center for Biotechnology Information (NCBI) (at http://www.ncbi.nlm.nih.gov) using
the BLASTP program (7) and searched against the Conserved Domains Database
(CDD) (72) (http://www.ncbi.nlm.nih.gov/structure/cdd) DNA Databank of Japan
(DDBJ) (http://www.ddbj.nig.ac.jp/E-mail/homology.html). The sequences were next
analyzed for protein domain by using Pfam 12.0 (47) (Saint Louis) (at
http://pfam.wustl.edu). The FASTA files of the translated amino acid sequences were
also submitted to the SIGNALP server (82) (at http://www.cbs.dtu.dk/services/SignalP/)
to determine the presence of signal peptides. Multiple sequence alignment of nucleic
and amino acids was performed using ClustalW (85) at the EBML-EBI server
(http://www.ebi.ac.uk/clustalw/#) and edited using the Genedoc program (available at
http://www.psc.edu/biomed/genedoc). Conserved protein motifs were analyzed by
using a motif scan (28) (at http://myhits.isb-sib.ch/cgi-bin/motif_scan).

11
Nucleotide accession number
The nucleotide sequences reported in this thesis will appear in the
DDBJ/EMBL/GenBank nucleotide sequence databases with the accession numbers
AB205028, AB205029, AB205030, and AB205031
1-3. Results
Random sequencing of the H. longicornis cDNA library
From 432 white plaques that have been randomly selected for PCR
amplification, 238 clones contained cDNA inserts with sizes ranging from 400 bp to 1.8
kbp. From 238 sequences studied, 80 had no possible open reading frame. These
clones were considered as junk sequences and were, therefore, excluded from further
analysis. The FASTA-formatted of all nucleotide sequences are available upon request
via email ([email protected]). Out of 158 sequences, 106 full-length cDNA
clones, which accounted for 67%, were obtained, as indicated by the presence of the
initiation codon. Sixty-five sequences were found to possibly contain the secretion
signal, whereas the secretion signal of 38 clones could not be determined since those
sequences were truncated.
A schematic display of cDNA sequence clusters from salivary gland proteins of
H. longicornis is shown in Fig. 1 and Fig. 2. Fig. 1 shows that, of the 158 cDNA
sequences studied, 43% match those related to Drosophila melanogastor and other
organisms. No significant similarity was found for 41% of the clusters, whereas only
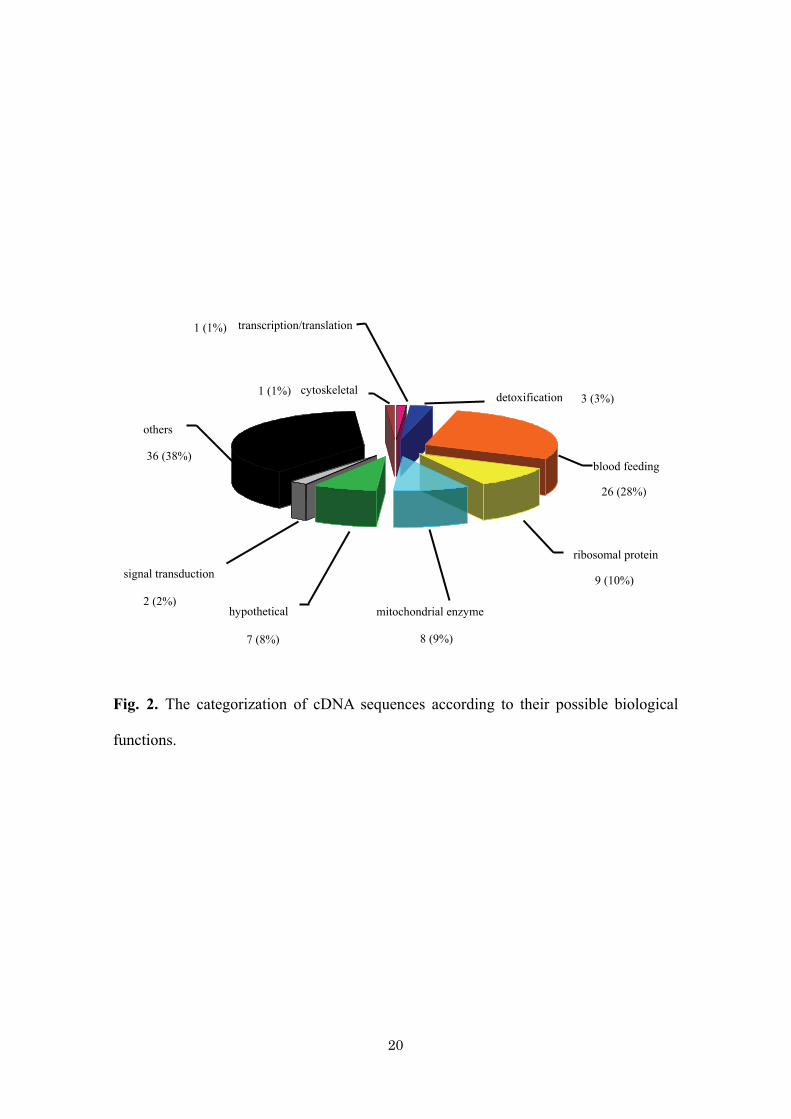
16% gave significant hits with tick sequences. Fig. 2 shows that, of the 93 known
sequences, among the most common categories were housekeeping proteins including
ribosomal proteins (10%), mitochondrial enzymes (9%), hypothetical proteins (8%),

12
detoxification-associated proteins (3%), signal transduction-associated proteins (2%),
transcription/translation-associated proteins (1%), cytoskeletal proteins (1%), and
proteins that have been classified as “others,” which accounted for 38% of the total
sequences studied. Interestingly, 28% were coded for proteins that were probably
expressed during blood feeding.
There are 26 cDNA clones (28%) that possibly play an important role in the
blood-feeding mechanism. Twenty-three of them matched previously reported
sequences from ticks, and the remaining 3 sequences matched another organism. There
are 7 sequences that share similarity with the cDNA sequence isolated from H.
longicornis (accession number AB014612) (75). This protein, designated p29, was
shown to induce protective immunity against tick infestation in an experimental animal.
As shown in Fig. 3, the amino acid sequence of the p29 extracellular matrix of H.
longicornis was compared to 7 deduced amino acid sequences isolated from this cDNA
library. There were 3 sequences that gave the best match with H. longicornis HL35
antigen U protein (accession number AY550980) (115). The other 13 clones gave a
match to bioactive molecules involved in antihemostatic, immunosuppressant, and
secretory proteins with unknown functions. Three sequences matched other organisms
but possibly have similar functions to proteins in ticks, including metalloprotease and
dipeptidyl peptidase IV in Apis mellifera (honeybee), and the tissue factor pathway
inhibitor in Gallus gallus (red jungle fowl chicken). Multiple sequence alignment of
some of the deduced amino acid sequences of these cDNA was performed as shown in
Fig. 4 to 7. Mucins are large glycoproteins that generally have a high prolin, threonine,
and serine content. They are normally associated with mucus layers at the interface
between epithelia and their respective environment, particularly the lumens of hollow

13
organs of the vertebrate respiratory, digestive, and urogenital tracts (122). One cDNA
clone, designated H50 (accession number AB205029), gave the best match to the
mucin-like glycoprotein in agarisid tick, Ornithodoros moubata. The prediction of the
conserved domain using the Pfam server suggested that this protein contains a chitin-
binding domain. The clustal alignment of H50 (HLmucin) and similar proteins isolated
from other organisms is shown in Fig. 4. During the blood feeding stage, ticks secrete
some cement-like components to assist attachment to the host skin. Fig. 5 shows the
sequence alignment of the putative cement protein of R. appendiculatus, Rhipicephalus
immunodominant molecule 36 (RIM36) (17), which shares similarity with 2 cDNA
sequences isolated in this study and designated H184 and H416.
The tissue factor pathway inhibitor (TFPI) is a key molecule in the blood
coagulation pathway. In the present study, one cDNA sequence matched the TFPI
family. The alignment of H. longicornis TFPI and the first 2 best-matched species is
shown in Fig. 6 with the predicted Kunitz conserved domain.
Metalloproteases are other bioactive molecules of interest that possess anti-
hemostatic activity. The multiple sequence alignment of the metalloprotease enzymes is
shown in Fig. 7. The first sequence, called H135 (accession number AB205030),
matched I. scapularis MP1, which is similar to the hemorrhagic snake venom
metalloprotease of the reprolysin family, whereas the other sequence, called H292
(accession number AB205031), shared low similarity with the disintegrin-like protease
in A. mellifera (accession number XP_396339). Two other sequences, designated
H364 (accession number AB205028) and H410 matched the metalloprotease in I.
ricinus (69) and I. scapularis (32), respectively. The deduced amino acid sequence
showed that all sequences except clone H292 contained catalytic domain of reprolysin

14
family including catalytic domain (zinc-binding motif) metzincin (20)
(HEXXHXXGXXH), triad methionine (MSY) and a distally located methionine from
the first H of the zinc binding motif. On the contrary, H292 contains an ADAM
Cysteine-rich domain (ACR) located at the C-terminal of the protein (data not shown).
No significant similarity was found for 65 sequences. Multiple sequence
alignment of deduced amino acid sequences was performed via the ClustalW server
(data not shown). From the alignment score, 55% of the sequences appeared to be
multiple copies, whereas the remainders were single copies. From the prediction of
secretory signal peptide, it was found that 33 out of 65 possibly secreted. The
determination of the isoelectric point of these secretory proteins using an EMBL server
indicated that all of these proteins were basic in nature, with predicted pIs ranging from
8.1 to 12.2. The 12 most abundantly expressed sequences were further searched for a
conserved motif by using a motif-scanning server. It was found that most of the proteins
contained similar patterns of post-translational modification, such as the presence of a
N-myristolation site, a protein kinase C phosphorylation site, and a glycosaminoglycan-
attachment site. There were 6 cDNA sequences that encoded for the glycine-proline rich
protein, 5 sequences that were glycine-rich, and 3 sequences that were glycine-serine
rich. These 3 sequences also contained a multicopper oxidase signature and 20 copies of
a collagen triple helix repeat at the N-terminal part.
1-4. Discussion
A high-throughput approach designed to identify a large number of cDNAs was
used to obtain information on a variety of genes that are expressed in the salivary glands
of arthropods of medical importance (31, 95, 118-120). In the present study, a cDNA

15
expression library of salivary glands from the adult female tick H. longicornis was
constructed and randomly sequenced. The cDNA sequences can be categorized into 3
groups according to the results of blast search, namely, novel sequences for which no
significant similarity was found, sequences that matched previously reported proteins
from ticks, and sequences that matched other organisms.
Twenty-six clones possibly play important roles during blood feeding, and 7 of
these are identical to each other. Clustal alignment of these sequences revealed 39%
identity, suggesting that these cDNA sequences are similar but not identical to the
previously reported p29 sequences and can therefore be considered as a novel protein.
This p29 protein was described as a collagen-like protein that may be associated with
the formation of tick cement (75). Collagenous proteins belong to a multi-family, which
indicates that these proteins might share similar properties and functions to the p29
protein. The other 3 cDNA clones matched the H. longicornis HL35 antigen U protein.
Two sequences matched an unknown protein of Rhipicephalus haemaphysaloides, and 2
sequences matched the cement protein RIM 36 of R. appendiculatus (17). This protein
was shown to be present in salivary glands and the cement cone material secreted by the
tick. The data indicates that this cement component can induce strong antibody
responses in cattle and can therefore be a diagnostic marker for the detection of cattle
that have been exposed to R. appendiculatus ticks.
Calreticulin, a major calcium-binding protein of the endoplasmic reticulum,
appeared to be secreted in the saliva of A. americanum and Dermacentor variabilis.
Some evidence showed that an antibody raised against recombinant tick calreticulin
may be a biological marker of tick exposure (100). The cDNA sequence H186 was
shown to be identical to previously reported calreticulin in H. longicornis (142). The

16
biological function of this protein has not been clarified, although calreticulin may have
a role in angiogenesis in a feeding lesion.
Tick saliva contains pharmacologically active molecules that inhibit host
hemostasis, reduce pain, and modulate host inflammatory and immune responses. A
recombinant immunomodulatory protein from the salivary glands of D. andersoni has
been characterized (1) and shown to have immunosuppressive activity. One cDNA
sequence found in this library (H318) matched this protein, indicating a possible
function in counteraction with the host immune response. In the present study, 4 cDNAs
encoding for metalloprotease were isolated from a salivary gland cDNA library. Further
study of this cDNA sequence should be performed in order to elucidate the function and
biological activity of this bioactive molecule. In addition to its antihemostatic activities,
the metalloprotease enzyme also participates in angiogenesis, which is interesting for its
potential in wound healing and tissue remodeling. In this salivary gland library, one
cDNA sequence, which matched dipeptidyl peptidase IV (dPPIV), was found. This
molecule possesses angiogenesis activity. Characterization of these molecules may lead
to a better understanding of the biological process of tissue repair upon blood feeding.
The clone H50 matched with the mucin-like glycoprotein of O. moubata.
Mucins have been investigated as vaccine antigens. The protein was shown to induce
protective immunity in experimental animals (136). Though mostly isolated from
intestine, mucin-like glycoprotein was also reported from salivary glands of An.
gambiae (31). It is still not known whether the protein modulates parasite infectivity.
The evidence shows that this protein family is a promising candidate for a vaccine.
In this library, 41% of the sequence studied did not match any sequence reported
in the database. Analysis of the 12 most abundantly expressed sequences by using the

17
motif-scanning method suggested that these proteins contained some phosphorylation
sites. This indicated a possible involvement in signal transduction of some biological
process. Though the gene information or biological function still unknown, these
proteins secreted during blood feeding stage indicating some important role for tick to
take blood meal.
Herein, the results of the random sequencing of cDNAs from salivary glands
cDNA library of H. longicornis were reported. According to the characteristics and
possible functions of these proteins, including antihemostatic and immunodominant
ones or some that are involved in host-parasite interaction, further study will be required
to clarify their roles.
1-5. Summary
In tick salivary glands, numerous genes were induced during blood feeding in
order to counteract with host defense mechanism. The newly synthesized protein might
be important for the blood meal or be involved in host immune evasion and pathogen
transmission. In this study, cDNA library from salivary glands of partially fed adult
female H. longicornis was constructed and randomly sequenced, in an attempt to
identify genes that were expressed during blood feeding. The open reading frames of
158 cDNA clones were submitted to similarity search. A number of novel sequences
that gave no match to any sequence reported in the database were identified and
approximately 50% of these sequences are possibly secretory product. The majority of
known sequences are housekeeping genes. Several coding sequences possessed various
degree of homology to previously described proteins from other tick species. Several
genes encoding putative antihemostatic, immunomodulatory proteins and cement

18
proteins were found. This work provides information into the diversity of messages
expressed in the salivary glands of H. longicornis, describes novel sequences that may
be responsible for known biological activities, and identifies novel vaccine targets that
may be used in development of tick control.

19
Fig. 1. The BLASTP search results of cDNA sequences assigned as having no match to
the database or having hits to sequences obtained from ticks or other organisms.
Tick 16% (25)
Others 43% (68)
Novel 41% (65)
20
Fig. 2. The categorization of cDNA sequences according to their possible biological
functions.
signal transduction
transcription/translation
cytoskeletal detoxification
ribosomal protein
hypothetical mitochondrial enzyme
blood feeding
others
8 (9%)
26 (28%)
36 (38%)
7 (8%)
1 (1%) 3 (3%)
1 (1%)
9 (10%)
2 (2%)

21
Fig. 3. Clustal alignment of novel H. longicornis extracellular matrix proteins and
previously reported p29 protein (AB014612-1). Identical amino acids are shown in
black, whereas similar residues are shown in grey.

22
Fig. 4. Clustal alignment of novel H. longicornis mucin-like glycoprotein (H50) and
orthologous proteins from other organisms, including cabbage looper, Trichoplusia ni
(AAC47557), Aedes aegypti (AAM94146), Diamond-back moth, Plutella xylostella
(AAN63949), Anophleles gambiae (AAL68783), and O. moubata (AAS01023). The
predicted chitin-binding domain of HL mucin from the Pfam server is shown as a bar,
ranging from amino acid residue 83 to 138.

23
Fig. 5. Clustal alignment of the novel H. longicornis putative cement protein (H184,
H416) and the R. appendiculatus cement protein (RIM 36, AAK98794).

24
Fig. 6. Clustal aligment of the novel H. longicornis tissue factor pathway inhibitor
(H371), G. gallus TFPI (XP_421849), and Bos taurus TFPI (AAF61248). The Kunitz
conserved domain of HLTFPI, as predict by the Pfam server, is shown as a bar, ranging
from amino acid residue 122 to 174.

25
Fig. 7. Clustal alignment of the H. longicornis metalloprotease and previously reported
metalloprotease from I. scapularis MP1 (JC7969), MP2 (AAM93652), and I. ricinus.
MP (CAB55817). The conserved zinc-binding motif is shown in a box
(HEXXHXXGXXH). The methionine triad is shown as thick line.
HEXXHXXGXXH

26
Chapter 2
Molecular characterization and comparative study of 6 salivary gland
metalloproteases from the hard tick, Haemaphysalis longicornis
2-1. Introduction
As blood-pool feeders, ticks utilize a unique strategy to complete their long-
period blood meal (109). Their feeding mechanism is orchestrated initially by
penetrating their mouthparts into the host skin and damaging blood vessels for the
release of blood. During subsequent feeding, ticks secrete molecules through saliva,
which greatly contribute to establish and maintain the blood pool by interfering with the
host’s hemostatic, inflammatory, and immune systems (93-95, 128). Since the saliva
molecules are in direct contact with the hosts, a thorough understanding of their
functions is crucially important to the development of a tick vaccine.
Metalloproteases comprise a family of Zn2+-dependent enzymes that degrade
most components of the extracellular matrix (16). They are secreted in a proenzyme
latent form that requires activation for proteolytic activity. The ADAM (a disintegrin
and metalloprotease) family is characterized by the presence of disintegrin-like and
metalloprotease-like domains (141). Some ADAMs also displays additional domains
such as transmembrane and cytoplasmic domains and thrombospondin domains
characteristic for the ADAMTS (a disintegrin and metalloprotease with
thrombospondin) (29). The metalloprotease domain has the active site that contains zinc
atom and water molecules that are necessary for the hydrolytic processing of protein
substrates. Both the disintegrin-like domain and cysteine-rich domains have been
implicated in adhesive events such as binding to integrins and interacting with haeparan

27
sulfate proteoglycans, respectively (127). ADAMs are involved in diverse processes,
such as extracellular matrix degradation, development, cell-cell interaction, and cell
migration (104).
In ticks, genes encoding metalloproteases were identified from the saliva of I.
scapularis (118). These sequences are similar to snake venom metalloproteases
(SVMPs). SVMPs are members of the ADAMs family, whose main toxic effects are
due to the disruption of the hemostatic system. SVMPs are classified according to their
multidomain organization into class PI to PIV (48). The PI-class has only pre-
prodomain and metalloprotease domain, class PII has subsequence disintegrin domain,
class PIII has additional cysteine-rich domain and class PIV has the PIII domain
structure plus lectin-like domain connected by disulfide bonds. Tick metalloproteases
have molecular weight compatible with the PII-class (30 to 60 kDa) of SVMP, with
additional cysteine-rich domain (32). The tick metalloproteases have no Arginine-
Glycine-Aspartate (RGD) triplet typical for the disintegrin (118). The cysteine residue
spacing follows neither pattern found in disintegrins nor that found in the cysteine-rich
domain of the PIII-class (118). Such metalloproteases are similar to atrolysins, found in
snake venoms, which have fibrinogenolytic and gelatinase activities (48, 108). In
general, SVMPs display proteolytic activity against the basement membrane proteins,
which result in diapedesis of white blood cells (8, 18, 19, 105). As well as SVMPs, the
saliva of I. scapularis also possesses these metal-dependent anti-coagulant activities
(32). In addition, some types of snake venom also inhibit angiogenesis by the induction
of endothelial cell apoptosis (114). Similarly, it has been shown that late host response
to injury, such as endothelial cell-dependent wound healing in the blood feeding lesion,
is negatively modulated by tick saliva, and some evidence supports the notion that

28
metalloprotease is the enzyme that most likely accounts for this activity (33). In
conclusion, these studies suggest that metalloprotease in ticks is possibly essential for
inhibiting blood clotting and degrading extracellular matrix proteins for the preparation
of the feeding site and inhibiting host tissue repair in the late feeding phase via its anti-
angiogenic activity.
However, the biological characteristics of tick metalloproteases remain elusive,
although SVMPs have been extensively characterized due to their pathological
relevance and potential for therapeutic applications (88). In the present study, 6
metalloprotease genes from a salivary gland cDNA library of the hard tick, H.
longicornis were isolated and characterized. This is the first study to evaluate the
dynamic expressions in response to the blood feeding and the expression of the
recombinant protein and proenzyme activation mechanism of metalloproteases from the
salivary glands of ticks.
2-2. Materials and Methods
Ticks
The parthenogenetic Okayama strain of the H. longicornis tick has been
maintained by feeding on rabbits and mice for several generations in our laboratory
since 1997 (36).
Identification of metalloprotease genes in the salivary glands of H. longicornis
The cDNA library from the salivary glands of an adult female H. longicornis
was constructed using the vector-capping method (61, 147). The cDNA was

29
synthesized from 5 μg of total RNA by the G-C-capping method and ligated into the
plasmid vector pGCAP1. The EST database was constructed by randomly sequencing
10,000 recombinants, and 2 full-length metalloproteases designated as hlESTMP1 and
hlESTMP2 were isolated. On the other hand, 4 other metalloprotease cDNA sequences
herein referred to as hlMP1, hlMP2, hlMP3 and hlMP4 (referred to as H135, H292,
H364 and H410, respectively, in chapter 1) were isolated by random sequencing of 158
recombinants from a salivary gland cDNA library of the adult H. longicornis as
described in chapter 1. Since hlMP1, hlMP2, and hlMP4 appeared to be truncated,
identification of the 5’end terminus was performed by PCR amplification of the phage
library using T3 and gene-specific reverse primers (hlMP1:
5’GCCGAATTCGGACATTCC3’, hlMP2: 5’CTCAGGGCCAGTGGCTACT3’,
hlMP4: 5’ GATTTGGTCCAACTACACCT3’). The PCR reaction conditions were as
follows: 94°C for 10 min, 30 cycles of 94°C for 1 min, 50°C for 1 min, 72°C for 2 min,
and 1 cycle of 72°C for 7 min. DNA fragments with the expected size based on
multiple sequences alignment were excised from the gel, purified using Geneclean II kit
(Q-BIO gene®, Bio101 system, MP Biomedicals, CA, USA), and cloned into pGEM-T
Easy vector (Promega, Madison, WI, USA). Colony screening by PCR was performed
using gene-specific primers with 20 white colonies, and positive clones were selected
for sequencing. The analyses of the nucleotide and deduced amino acid sequences were
performed as previously described in chapter 1.
Phylogenetic analysis
A phylogenetic tree was constructed with the topology algorithms using
Genebee-Net (available at http://www.genebee.msu.su/services/phtree_reduced.html

30
server). The default method was used with PHYLIP and unrooted parameters. A
distance matrix method was used to create the tree with boostrap values of the full-
length metalloproteases.
Nucleotides accession numbers
The nucleotide sequences reported in this thesis will appear in the
DDBJ/EMBL/GenBank nucleotide sequence databases with the accession numbers
AB264113, AB264114 and AB265817
Analysis of metalloprotease mRNA expression during blood feeding
To study the expression of H. longicornis metalloprotease (hlMP) genes during
blood feeding, 60 adult ticks were infested on a Japanese white rabbit, and 8 ticks were
collected every day until engorged (7 days). Ticks were dissected, and salivary glands
were pooled and kept in TRI® reagent (Sigma, St. Louis, MO, USA) at –80°C for
further RNA extraction. The expression patterns were studied by reverse transcription
PCR (RT-PCR) using gene specific primers (Table 1) and compared with those of
salivary glands from unfed ticks, as previously described in chapter 1. The RT-PCR
reaction conditions were as follows: 50°C for 30 min, 94°C for 2 min, 30 cycles of
94°C for 30 s, 50°C for 30 s, 72°C for 2 min, and 1 cycle of 72°C for 7 min. The H.
longicornis actin primers (accession number AY254898) were used as an internal
control for the RT-PCR reaction. To exclude the possibility of genomic DNA
contamination, negative-control reactions were performed without adding reverse-
transcriptase. The intensity of each RT-PCR product was quantified with the Macintosh

31
Luminous Imager version 2.0, and the values were expressed as a relative expression in
comparison to the actin gene. Data are expressed as means ± standard error of three
replicate experiments.
Expression of hlESTMP1 in E. coli
The PCR amplification of the pGCAP1 harboring the hlESTMP1 cDNA
sequence for the expression was performed using primers flanking the coding region of
that gene. A full-length hlESTMP1 containing a prodomain plus a metalloprotease
domain and a mature hlESTMP1 that contains only a metalloprotease domain were
subcloned into a BamHI restriction site of the pRSET-B expression vector (Invitrogen,
Carlsbad, CA). The full-length hlESTMP1 was amplified using the forward primer
(5’GAGGATCCGATGATTCTTCTTCTC3’) and the hlESTMP1 reverse primer
(5’GAGGATCCGTCATTTGTTTTGCCA3’). The start codon (ATG) and stop codon
(TCA) are indicated in bold. The BamHI restriction sites are indicated in italics. The
mature hlESTMP1 was amplified using the forward primer fused in-frame with the N-
terminal histidine tag gene (5’GAGGATCCGCGGCCAAATGTTCAA3’) and the same
reverse primer. The resulting plasmids were transformed into E. coli BL21 cell for the
expression of the recombinant hlESTMP1.
Purification of the recombinant hlESTMP1 fused with the histidine tag
The expression and refolding of the recombinant hlESTMP1 in E. coli were
performed as follows. Briefly, a single colony of E. coli BL21 harboring the
recombinant full-length or mature hlESTMP1 plasmids was grown overnight in 5 ml of

32
an LB-ampicillin broth. The culture diluted to 1:50 in 200 ml of a fresh LB-ampicillin
medium was grown until O.D600 reached 0.4. The isopropyl-beta-D-
thiogalactopyranoside (IPTG) induction was then performed with a final concentration
of 0.5 mM at 25°C overnight. For the purification of both recombinant proteins, the
inclusion bodies were resuspended with 10 ml of 6 M urea in an equilibration buffer
(100 mM NaH2PO4, 10 mM Tris-HCl pH 8.0). The mixture was incubated at 4°C for 2
hr and centrifuged at 15,000 X g for 20 min at 4°C. The supernatant was collected and
purified under a denaturation condition using Ni-NTA resin (Qiagen, Tokyo, Japan)
according to the manufacturer’s instructions. The proteins were eluted using the pH
gradient method. The refolding of the purified proteins was performed as described by
Ramos et al (87).
Production of an anti-hlESTMP1 serum
The full-length hlESTMP1 inclusion body was used to immunize ddy mice
(female, 6 weeks old) to obtain antiserum against hlESTMP1. The inclusion body was
washed for 6 times to remove the cellular proteins by sonicating in 1% triton-X-PBS
and one time in PBS. Immunization was performed for totally 4 times. First,
immunization was performed with an intraperitoneal injection of 100 μg protein in
Freund’s complete adjuvant (Sigma, St. Louis, MO, USA). The boosters were given at
2-week intervals with the same amount of protein in Freund’s incomplete adjuvant.
Blood samples were collected via a heart puncture after the third booster. The non-
specific antibodies were absorbed prior to use for Wester blotting and IFAT by
incubating antisera with lysate prepared from E. coli BL21 overnight at 4°C (100 μl
antisera: 50 μl E. coli lysate: 900 μl 5% skimmed milk in PBS).

33
Analysis of native hlMP1 in salivary glands by Western blotting
The salivary glands of unfed and 4-day fed female ticks were dissected out and
washed with cold PBS. The tissue was homogenized using a tissue homogenizer,
sonicated for 15 s, 3 times, and then centrifuged at 15,000 X g for 20 min at 4°C. The
supernatant was collected, and the protein concentration was measured using a Bio-Rad
Protein assay kit (Bio-Rad, Hercules, CA, USA). Ten micrograms of the soluble
fraction from the salivary gland was used as an antigen for sodium dodecyl sulfate poly
acrylamide gel electrophoresis (SDS-PAGE) and Western blotting. The protein was
transferred onto a polyvinylidine difluoride (PVDF) membrane. The membrane was
reacted with mouse sera followed by goat anti-mouse-conjugated horseradish
peroxidase. The signal was detected by using a chemiluminescent substrate (Perkin
Elmer Life Sciences, Boston, MA, USA) following the manufacturer’s protocol.
Indirect immunofluorescent assay
The localization of native hlESTMP1 in salivary glands was detected by the
indirect immunofluorescent assay (IFAT). The whole salivary glands of unfed and 4-
day fed adults were placed on 14-well slides, air-dried, and fixed in acetone for 30 min.
The slides were further blocked with 0.5% skimmed milk in PBS for 1 hr at 37°C,
washed with phosphate-buffered saline (PBS), and incubated with anti-hlESTMP1
mouse serum diluted at 1:100 with 0.5% skimmed milk under the same conditions.
After washing, the slides were incubated with goat anti-mouse-conjugated alexa 488
diluted at 1:1,000 (Invitrogen). Nuclei staining were performed by incubating the slides
with 25 μg/ml propidium iodide (PI; Molecular Probe, Eugene, OR, USA) for 5 min at

34
room temperature and washing twice with PBS. The fluorescent signal was examined
with a confocal laser scanning microscope (TCS NT; Leica, Heiderberg, Germany).
Expression of hlESTMP1 in the baculovirus
The full-length hlESTMP1 with the signal peptide was amplified using primers
flanking the coding sequence (forward primer:
5’GCGGGATCCATGATTCTTCTTCTC3’, reverse primer:
5’GCGGGATCCTCAGTGATGGTGATGGTGATGTTTGTTTTGCCACCGATG3’).
The start codon (ATG) and stop codon (TCA) are indicated in bold. The BamHI
restriction sites are indicated in italics. The 6 histidine tag was added to a C-terminal to
aid purification. The PCR product was cloned into the BamHI restriction site of the
pFastBac Dual vector (Invitrogen). The recombinant donor plasmid was transposed to
the target site on the bacmid present in E. coli DH10BAC. Recombinant bacmid DNA
was prepared and used for transfection into Spodoptera frugiperda cells. The
recombinant baculovirus was amplified by multiple infections of insect cells. The viral
supernatant was used for the infection of High Five™ cells. After 5 days of infection,
the cells were harvested and washed with PBS. The cell pellet was resuspended in PBS
and lysed by sonication. The suspension was centrifuged at 10,000 X g, 10 min at 4°C
to separate the soluble fraction and cell debris fraction was utilized for SDS-PAGE and
Western blot analysis.
N-terminal sequencing of baculovirus-expressed hlESTMP1
The insoluble fraction of High Five™ infected cells was resuspended in a lysis

35
buffer (1% Nonidet P40 in PBS) and used for SDS-PAGE. Three major bands
corresponding to hlESTMP1, as proven by Western blotting using anti-hist tag
monoclonal antibody, were selected for sequencing. The protein was transferred onto a
PVDF membrane, and N terminal sequencing was performed by Edman degradation
(Hokkaido System Science Co., Ltd.).
2-3. Results
cDNAs cloning and sequences analyses of hlMPs
The PCR amplification of the 5’ terminus of 3 metalloproteases cDNA using a
phage library gave the multiple band products observed. The DNA fragments with
expected sizes were selected for sequencing. The full-length sequence of hlMP1 was
successfully obtained, whereas that of hlMP2 was unsuccessful due to the non-specific
amplification of the primers used. Identification of the full-length of the remaining
truncated cDNAs by using plaque hybridization and 5’ RACE failed to obtain the
missing 5’ terminus. The additional sequence of hlMP4 was obtained, but the starting
methionine was not identified.
It was found that the 6 hlMP proteins shared 13-41% identity to each other and
shared 16-42% identity with metalloproteases from other tick species. The multiple
sequence alignment of hlMPs with metalloproteases from other ticks is shown in Fig. 8.
Although the primary sequences shared a low percent identity, essential conserved
domains and critical residues were observed. These proteins were synthesized as a
proenzyme containing signal peptides, a prodomain, and a metalloprotease/cysteine-rich
domain. The amino acid sequences and conserved domain structures suggested that the
6 genes belong to the SVMP M12 family (EC 3.24.21) (90), with the catalytic site of the

36
metzincin subgroup (20). The phylogenetic analysis shown in Fig. 9 reveals the distinct
evolutionary patterns of these proteins. The hlESTMP1 is closely related to
Rhipicephalinae MPs. Except for hlMP1 that branch independently, the remaining
sequences, including hlESTMP2, hlMP2, hlMP3, and hlMP4, are close to each other.
Besides hlMP2, all the sequences studied possess a prodomain and a mature
metalloprotease domain. The structural features of each sequence are summarized in
Table 2. The hlMP3 contained an additional fragilysin domain that is similar to that of
the Bacteroides fragilis toxin (74), whereas hlMP2 contained only the ADAM/cysteine-
rich domain. All of the full-length sequences (hlESTMP1, hlESTMP2, and hlMP3)
contained a secretory signal peptide, as predicted by the SignalP server. The size of the
proteins and conserved domains suggested that hlMPs share common features with
snake venom class P-II except for the absence of the RGD triplet sequence of the
disintegrin domain.
The hlMPs have the zinc binding motif domain of the reprolysin family
(HELGHNLGXXHD), with some substitutions within the active site. The conserved
residues and motifs are summarized in Table 3. Besides hlMP2, the remaining 5
sequences possess all common characteristics of SVMPs, as described in I. scapularis
MPs, except for several amino acid substitutions of the reprolysin conserved domain.
The putative methionine turn of the triad MSY following the zinc-binding motif is
present in all 5 hlMPs. Three active site sequences, namely, hlESTMP1, hlESTMP2,
and hlMP3, have a G to A substitution as I. scapularis MPs, whereas hlMP4 remains
unchanged. The majority of substitutions are conservative except for hlMP3, which
changes from a hydrophobic amino acid to a polar one (L→T) and vice versa (N→V).

37
The expression of hlMPs mRNA transcripts during blood feeding
The expression profile of hlMPs during tick blood feeding is shown in Fig. 10
(a). The relative expression of hlMPs in comparison to the internal control actin based
on the band densitometry of RT-PCR products is shown in Fig. 10 (b). Accordingly,
hlMPs were strongly upregulated in salivary glands during blood feeding. Three
patterns of expression were observed (Fig. 10 a, b). Three genes, namely, hlESTMP2,
hlMP1, and hlMP2, were markedly induced when ticks started taking their blood meal,
gradually increased upon blood feeding, and decreased after ticks had fed near repletion.
In contrast, the expression of hlESTMP1 and hlMP4 was still relatively high even in
engorged ticks. Moreover, hlMP3 expression was apparently different from that of
other genes since the level of the transcript was markedly downregulated after the third
day of feeding.
Analysis of native protein and localization of hlESTMP1 in salivary glands
The heterologous expression of the full-length and mature hlESTMP1 in E. coli
was performed. The recombinant protein fused with a polyhistidine tag protein with an
expected size of 54 kDa and 36 kDa was obtained (data not shown). Western blotting of
salivary gland extracts showed that the antiserum against hlESTMP1 reacted strongly
with a protein of approximately 34 kDa from salivary gland extracts of partially fed
ticks and weakly reacted with that of unfed ticks (Fig. 11). The localization of
hlESTMP1 expression in salivary glands was shown by IFAT (Fig. 12). The fluorescent
signal was clearly detected in the cytoplasm of acini (mainly, type III acini) of the
glands of partially fed ticks reacted with antiserum against hlESTMP1 (Fig. 12 a).

38
Non-specific autofluorescence was observed in the chitinous basement membrane of the
gland acini reacted with normal mouse serum (Fig. 12 b).
Proenzyme activation and partial amino acid sequences analysis of baculuvirus-
expressed hlESTMP1
The full-length hlESTMP1 was expressed as a C-terminal hexahistidine tag
using a baculovirus expression system with the calculated size of 53 kDa (Fig. 13). The
majority of the recombinant protein was presented in an insoluble fraction. Western
blotting using a mouse anti-hist tag monoclonal antibody suggested that no recombinant
protein was secreted into the culture supernatant (data not shown). The recombinant
hlESTMP1 appeared to be processed into 2 smaller fragments with sizes of
approximately 38 and 26 kDa. These 3 bands are the processed forms of the
recombinant protein, as confirmed by N-terminal sequencing and Western blotting with
an anti-hist tag monoclonal antibody. The first band with molecular mass 53 kDa is the
full-length hlESTMP1 without signal peptide. The amino acid sequence of this
fragment, KEVSV (Fig. 13), corresponded to the signal peptidase cleavage site, as
predicted by SignalP (Fig. 13). The amino acid terminal sequences of two smaller bands
with predicted sizes of 38 and 26 kDa are APQVR and TYYSQ (Fig. 13), respectively.
The position of these cleavage sites are highlighted as red boxes in Fig.8.
2-4. Discussion
Identification and characterization of 6 metalloprotease genes in salivary
glands of H. longicornis were performed on the basis of sequence similarity and mRNA

39
expression patterns during blood feeding. The diverse functions of enzymes belonging
to this family, which included extracellular matrix degradation, inhibition of blood
coagulation (88), and inhibition of angiogenesis (145), permit a speculation on the
similar functions of these metalloproteases in ticks for the facilitation of the blood
feeding. Remarkably, the sequences of hlMPs showed unique and different features
when compared with those of SVMPs and I. scapularis MPs. Firstly, unlike SVMPs,
which contain a highly conserved prodomain that possibly resulted from gene
duplication (88), this region of hlMPs is quite diverse, indicating a different
evolutionary process. Secondly, several substitutions of active site residues were
observed in hlMPs, which apparently differ from I. scapularis MPs. The functional
significance of these substitutions remains unknown but possibly resulted in a high
degree of substrate preference. Since ticks feed on a variety of animals, the synthesis of
various metalloproteases may provide a large repertoire of tools to interact with animal
target proteins. Interestingly, these hlMPs also displayed different mRNA expression
patterns during feeding. Most hlMPs were constitutively expressed to different extents
before the ticks took blood meal. The subsequent feeding strongly triggered the
expression of all 6 genes. Remarkably, except for hlMP2 that showed slight expression
in ovaries, all of these genes were expressed only in salivary glands (data not shown).
This evidence may infer to the role of these metalloproteases around the host feeding
lesion, which might be involved in maintaining a feeding cavity and dealing with a late
host response to tissue injury, such as endothelial-cell-dependent wound healing.
Residue changes in the active site of hlESTMP1 are the subtlest among those
observed in other hlMPs. Therefore, hlESTMP1 possibly maintains its catalytic activity
or substrate specificity. Moreover, this gene is constitutively expressed and expected to

40
be involved in functions other than feeding. For these reasons, this gene was selected
for further characterization by expression as a recombinant protein. The recombinant
hlESTMP1 was used for the production of a specific antibody in mouse. The anti-
hlESTMP1 antibody reacted with the 34-kDa protein in the soluble fraction of the
salivary gland extract, which corresponded to the predicted size of the mature protein.
These results are in agreement with those obtained in I. scapularis MPs, in which the
mature form could be detected in the tick saliva (118). The full-length protein cannot be
detected even in unfed glands, indicating that the prodomain was intracellularly cleaved
after synthesis. As shown by IFAT, this protein is mainly localized in the salivary gland
cytoplasm of acini III, which is the main production site of several bioactive molecules
(95, 128), indicating that native hlESTMP1 might be synthesized and processed there
and released as the soluble mature metalloprotease into the acini lumen. The
simultaneous activation of the proenzyme suggested the existence of a natural specific
inhibitor of tick metalloproteases that regulate the activity of these mature enzymes.
The synthesis of proteases as proenzyme is one way to prevent the damage
caused by their actions in undesired locations on unintended substrates (68).
Understanding the proenzyme activation mechanisms would provide the necessary
information for the design of a selective inhibitor. In the case of ADAMs, it is likely
that the mechanism of maturation occurs via a pro-protein convertase-dependent
pathway. However, there are some cases in which ADAMs may undergo autocatalytic
activation (19). The autolysis mechanism was not observed in E. coli-expressed
hlESTMP1. The in vivo processing of the multidomain of hlESTMP1 was successfully
demonstrated using a baculovirus expression system. The full-length hlESTMP1 was
post-translationally modified into the mature domain in insect cells. Furthermore, the

41
N-terminal sequence of the recombinant hlESTMP1 suggests that the recombinant
enzyme was directed into the secretory pathway, where the signal peptide could be
cleaved by signal peptidase and the proenzyme was further processed into 2 smaller
fragments. The presence of 53 kDa fragment suggested the inefficient cleavage of the
prodomain by insect cells. It is still unknown why the third band with molecular mass
26 kDa is presented in the sample. This unexplainable phenomenon possibly happens
by the post-translational processing of insect cells that might not occur under
physiological condition of tick salivary glands. The most probable active form of the
baculovirus-expressed hlESTMP1 is the 38-kDa protein, since the amino acid APQVR
is in the position following the residue R or paired basic residue KK, reported cleavage
sites of furin-type proprotein convertase (12), indicating that the activation of this
proenzyme possibly follows this pathway. Post-transcriptional gene silencing, such as
the RNA interference technique targeting the enzyme responsible for the proenzyme
activation, would be a valuable tool to clarify this biological process.
Attempts to express both a full-length and truncated form of hlMPs as a GST
fusion protein failed to produce a soluble protein (data not shown). The insolubility of
recombinant hlESTMP1 hinders the characterization of its enzymatic activity. As in the
case of SVMPs expression, there have only been a few papers report the production of
this enzyme in a recombinant active form due to the large number of cysteine residues
in the disintegrin/cysteine rich domains (88). Searching for a proper expression or
refolding system will pave the way to clarify the role of these metalloprotease in ticks.
While most studies of tick-host interactions have focused on the mechanism of
anti-hemostasis, immune evasion and pathogen transmission, few studies have
concentrated on the effects of tick salivary gland molecules on wound healing (33, 60).

42
Most expression profiles of tick salivary glands have revealed a number of
metalloproteases (2, 32, 60, 118) with homology to those involved in extracellular
matrix (ECM) remodeling. The proteolytic environment created by tick-secreted
metalloproteases might be generating inhibitory peptides from ECM degradation that
contribute not only to delayed bite site healing and inhibition of blood coagulation but
also to immune evasion (2).
To this end, the integrated data obtained from this study could provide a novel
target for the development of a tick control strategy. This could be achieved by
disrupting the proenzyme activation mechanism to inhibit the production of an active
mature enzyme or by exploring metalloprotease regulators, such as inhibitors or
blockers of enzyme function, by vaccinating host animals with this bioactive molecule.
2-4. Summary
Genes encoding metalloproteases were identified from salivary gland cDNA
library of H. longicornis. The proteins are similar to snake venom metalloprotease of
the reprolysin family. The H. longicornis metalloproteases are proteins containing pre-
and prodomains, the zinc binding motif HEXXHXXGXXH common to the
metalloprotease and a cystein-rich region. Among these genes, apparent differences in
evolution and gene regulation during blood feeding were observed. Interestingly,
several amino acid substitutions within the active site of catalytic metalloprotease
domain were also observed. Molecular cloning and expression as a recombinant protein
of one selected gene was performed. This provided interesting data concerning tissue
localization and native protein size of this molecule in salivary gland. Moreover,
mechanism of proenzyme activation to produce a mature active enzyme was speculated.

43
These integrated information provided a better understanding of the role of this enzyme
in tick feeding mechanism.
Table 1. The gene specific primers used for RT-PCR in this study.

44
Primer Nucleotide (5’-3’)
hlESTMP1 forward GCGGAACTCATTGCTTATATGG
hlESTMP1 reverse GCAGTACACCCCCATGTAGGAC
hlESTMP2 forward GCGGGATCCGTTGAAGTATTTATA
hlESTMP2 reverse GCGGGATCCTCTCATTGTCTCCGACG
hlMP1 forward GGCACTGAA GCAGTAGCACTT3’
hlMP1 reverse GCGGGATCCCTATTCTTGTACACA
hlMP2 forward GCTTCTGAAAGGAGTGGAGC
hlMP2 reverse GTTCCAACTCTGACTGCAATTTCTCC
hlMP3 forward CGCCACAAGTTCACCAAGAA
hlMP3 reverse CATTTCATGTAGTCCACGGATT
hlMP4 forward CCGGCTCACGCTTGTCTT
hlMP4 reverse GATTTGGTCCAACTACACCT
hlactin forward GGTTGCCGCCCTGGTGGTTGA
hlactin reverse GCCGCACGATTCCATACCCAGG

45
Tabl
e 2.
Stru
ctur
al fe
atur
es o
f hlM
Ps
Sequ
ence
Id
Nuc
leot
ide
acce
ssio
n
num
ber
Nuc
leot
ide
(bp)
Am
ino
acid
aa
(kD
a)
Sign
al
pept
ide
aa (k
Da)
Prod
omai
n
aa (k
Da)
Met
allo
prot
ease
/cys
tein
e-ri
ch
dom
ain
aa (k
Da)
AD
AM
/
cyst
eine
-ric
h
dom
ain
aa (k
Da)
Frag
ilysi
n
dom
ain
aa (k
Da)
hlES
TMP1
A
B26
4113
15
93
483
(54.
8)
17 (1
.7)
185
(20.
8)
298
(34.
0)
- -
hlES
TMP2
A
B26
4114
17
71
512
(57.
5)
16 (1
.6)
174
(19.
8)
376
(41.
9)
- -
hlM
P1
AB
2050
30
1575
48
2
(55.
1)
25 (2
.7)
180
(20.
6)
303
(34.
6)
- -
hlM
P2
AB
2050
31
1090
27
4
(30.
7)
- -
- 27
4 (3
0.7)
-
hlM
P3
AB
2050
28
1708
50
6
(56.
8)
16 (1
.8)
171
(19.
3)
335
(37.
5)
- 10
2 (1
1.0)
hlM
P4
AB
2658
17
1193
39
7
(45.
9)
- -
250
(29.
0)
- -

46
Table 3. Conserved residues and motifs of hlMPs
Sequence Id Metzincin HELGHNLGXXHD MSY No. of
cysteine
hlESTMP1 3 HELAHSLGAEHD 3 13
hlESTMP2 3 HEVAHTLGATHD 3 15
hlMP1 3 HEIAHSFGCVHD 3 15
hlMP2 5 LQVALLLNASRD 5 12
hlMP3 3 HETAHVLGSFHD 3 11
hlMP4 3 HEMGHTLGCSHD 3 7
Metzincin is characterized by the presence of the triad MSY and conserved Zn2+
binding motif (HEXXHXXDXXH). The amino acid sequences of the reprolysin family
are shown (HELGHNLGXXHD). The substitutions of residues in the active site of
hlMPs are shown in bold.

47
Metalloprotease-cysteine rich domain
Prodomain

48
Fig. 8. Multiple sequence alignments of hlMPs with other tick MPs.
The CLUSTAL alignments of 6 hlMPs including hlESTMP1 (AB264113), hlESTMP2
(AB264114), hlMP1 (AB205030), hlMP2 (AB205031), hlMP3 (AB205028), hlMP4
with R. haemaphyloides (ABD66751), R. (B.) microplus, (AAZ39657) I. ricinus
(CAB55817), I. scapularis MP1 (JC7969), I. scapularis MP2 (AAM93652) and I.
scapularis MP3 (AAM93653) are shown. The predicted signal peptides are underlined.
The arrows indicate above the aligned sequences are prodomain and metalloprotease-
cysteine rich domain of hlMPs, as predicted by conserved domain database. The black
box indicates active site. The closed triangles denote cysteine residues. The triad MSY
is underlined by thick line. The identity (black shadow), highly conserved (dark grey
shadow) and conserved residues (light grey shadow) are marked. The red boxes
indicate N-terminal sequences of processed baculovirus expressed hlESTMP1, KEVSV,
APQVR and TYYSQ.

49
Fig. 9. Phylogenetic analysis of hlMPs with other tick MPs.
The phlylogenic analysis of hlMPs with R. haemaphyloides, R. (B.) microplus, I. ricinus,
I. scapularis MP1, I. scapularis MP2, I. scapularis MP3. The bootstrap values are
shown on the lineage of the tree.
R. (B.) microplus
hlESTMP1
I. scapularis MP2
R. haemaphysaloides
I. scapularis MP3
I. ricinus
I. scapularis MP1
100
97
100 hlESTMP2
100
hlMP3
100
100
hlMP4
hlMP2
100
100
100
92
hlMP1

50
Fig. 10. The expression analyses of hlMPs mRNA in pooled salivary glands of female
ticks fed on rabbits.
a) The mRNA expression using RT-PCR of hlESTMP1, hlESTMP2, hlMP1, hlMP2,
hlMP3 and hlMP4 in salivary glands from unfed, 1 day fed (day1), 3 day fed (day3), 5
days fed (day5) and engorged ticks. The amplification of actin gene (hlactin) was use as
an internal control.
hlESTMP1 0.4
0.3
hlMP1 0.5 0.4
hlMP2 0.1 0.2
hlMP3 0.3 0.2
hlMP4 0.4 0.3
hlactin0.5 0.4
Unfed Day1 Day3 Day5 engorged
hlESTMP2 1.0 1.5
Kbp
a)

51
b) The relative expression of hlMPs in comparison to actin gene.
hlESTMP2
hlESTMP1
hlMP2
hlMP1
hlMP3
hlMP4
b)
0.000
0.200
0.400
0.600
0.800
1.000
1.200
unfe
d
Day1
Day3
Day5
engo
rged
rela
tive e
xp
ress
ion
0.0000.2000.4000.6000.8001.0001.200
unfe
d
Day1
Day3
Day5
engo
rged
rela
tive e
xp
ress
ion
0.000
0.200
0.400
0.600
0.800
1.000
1.200
unfe
d
Day1
Day3
Day5
engo
rged
rela
tive e
xp
ress
ion
0.0000.2000.4000.6000.8001.0001.200
unfe
d
Day1
Day3
Day5
engo
rged
rela
tive e
xp
ress
ion
0.0000.2000.4000.6000.8001.0001.200
unfe
d
Day1
Day3
Day5
engo
rged
rela
tive e
xp
ress
ion
0.0000.2000.4000.6000.8001.0001.200
unfe
d
Day1
Day3
Day5
engo
rged
rela
tive e
xp
ress
ion

52
Fig. 11. Western blot analysis of salivary gland antigen extract with anti-hlESTMP1.
Western blotting of salivary gland extract of unfed (lanes 1 and 3) and 4 days fed ticks
(lanes 2 and 4) reacted with non-immune mouse serum (lanes 1 and 2), mouse
antiserum against hlESTMP1 (lanes 3 and 4). Mr, pre-stained marker. Arrow indicates
the size of native mature hlESTMP1.
kDa Mr 1 2 3 4
98
22 16
64
50
36
148
34 kDa

53
Fig. 12. The localization of hlESTMP1 in salivary glands of partially fed ticks by IFAT.
The cytoplasm of salivary glands acini, mainly acini III, from partially fed ticks
specifically reacted with anti-hlESTMP1 sera (green), but not with granules (a). Non-
specific autofluorescence was observed in the chitinous basement membrane of the
gland acini reacted with non-immune mouse serum (b). PI (red nuclei staining by
propidium iodide).
Anti-hlESTMP1 PI Merge
Non-immune mouse serum PI Merge
(a)
(b)

54
Fig. 13. The baculovirus-expressed hlESTMP1 and amino terminal sequences of the
cleavage sites. The amido black staining of hlESTMP1 expressed by baculovirus on
PVDF membrane (lane 1) and Western blotting of the same sample reacted with mouse
anti-HIS tag monoclonal antibody (lane 2). Mr, pre-stained marker. The corresponding
amino terminal sequences are shown as KEVSV, APQVR and TYYSQ.
Mr 1 2
98
22
16
64
50
36
148
KEVSV
APQVR
TYYSQ
N-terminal sequence

55
Chapter 3
Identification of genes encoding cement-like antigens expressed in the
salivary glands of Haemaphysalis longicornis
3-1. Introduction
Ticks are medically important vectors of human and animal diseases. In
addition to transmitting pathogens to mankind, tick infestations in animals produce
worldwide economic losses (23). Although tick control strategies rely extensively on
the use of chemical acaricides, this type of control is becoming less sustainable due to
the development of resistance against acaricides among ticks and the undesirable
contamination of the environment and animal food products (137). Therefore, the
development of alternative control strategies is necessary, in which vaccination against
ticks is a candidate method. A concealed antigen based-vaccine against R. (B.)
microplus (54) infestation was successfully commercialized (135). A concealed antigen
in the tick gut, however, is unlikely for the boost of the immune response by natural tick
exposure compared with salivary glands antigens.
As a vaccine target, salivary glands are a rich source of exposed antigens that
can stimulate host immune responses following natural infestation. Tick saliva contains
a mixture of peptides and proteins serving various functions that are essential for the
creation and maintenance of the blood pool or the lesion in the host skin for feeding.
Several bioactive molecules in tick saliva that affect the host’s hemostatic, inflammatory,
and immune systems have been extensively studied (93-95, 128). The discovery of
novel proteins and genes expressed in salivary glands has been accomplished by
massive sequencing of full-length cDNA libraries together with approaches involving

56
proteomics and functional genomics. This information assists in the exploration of
protective antigens for vaccine development.
Several vaccine candidates have been studied and tested for vaccine potency
against H. longicornis infestation (55, 75, 115). However, the animals vaccinated with
these molecules conferred partial protection against not all stages of tick. The
identification of other candidates for use in a cocktail vaccine is still required.
In this research, of particular interest are the immunogens produced from the
salivary glands of ticks. A cDNA library from the salivary glands of H. longicornis
would provide valuable material, as it would contain a wide variety of genes encoding
putative vaccine candidates. The immunoscreening method was used to recruit
immunodominant key molecules that might be useful for the tick control strategy.
3-2. Materials And Methods
Ticks
The adults and nymphs of parthenogenetic Okayama strain of H. longicornis
maintained by feeding on rabbits and mice for several generations in our laboratory
since 1997 were used in this experiment (36).
Animals
All animal experiments were conducted in accordance with the Standards
Relating to the Care and Management of Experimental Animals promulgated by
Obihiro University of Agriculture and Veterinary Medicine, Hokkaido, Japan
(Allowance No. 17-77).

57
Immunoscreening of the cDNA library
The cDNA library of salivary glands from adult H. longicornis was obtained, as
previously described in chapter 1. Approximately 20,000 pfu per plate of the amplified
library was used for plaque lifting. The library was grown on NZY agar for 3.5-4.0 hrs
at 42°C. Then, IPTG induction was performed at 37°C for 3.5 hrs by placing
nitrocellulose membranes soaked with 10 mM IPTG onto the surface of the agar. For
secondary and tertiary screening, 600 and 200 pfu were used for plating, respectively.
The total of 5 X 105 pfu of the cDNA library was screened using a polyclonal rabbit
anti-H. longicornis tick immune serum prepared as previously described (144).
Immunoscreening was performed using a Picoblue TM immunoscreening kit (Stratagene,
CA, U.S.A), according to the manufacturer’s instructions. After blocking a non-specific
binding site with 1% bovine serum albumin (BSA) for 1 hr, the membranes were
allowed to react with polyclonal rabbit anti-tick saliva serum diluted 1:200 in 1% BSA
in Tris-Buffered Saline (TBS) for 3 hrs. After washing 3 times with Tris-Buffered
Saline and 0.05% Tween 20 (TBS-T), the membranes were further incubated with
alkaline phosphatase-conjugated goat anti-rabbit IgG at a dilution of 1:10,000. The
Nitroblue Tetrazolium (NBT) and 5-Bromo-4-chloro-3-indolyl Phosphate (BCIP)
substrates were used for the visualization of the positive signals. The positive plaques
were subjected to further secondary and tertiary screening until the isolated clones were
obtained. The cDNA inserts of the positive clone were then excised from the lambda
phages to obtain the plasmid DNA for sequencing.
Nucleotides accession numbers
The DNA sequences analysis was performed as described in chapter 1. The

58
nucleotide sequences reported in this thesis will appear in the DDBJ/EMBL/GenBank
nucleotide sequence databases with the accession numbers AB252633, AB259292,
AB259293, AB259294, AB259295, AB259296, AB259297, AB259298 and AB259299.
RT-PCR analysis of mRNA expression in salivary glands
The expression of immunodominant mRNA transcripts in the salivary glands of
H. longicornis was studied. RT-PCR was performed using a TAKARA one-step RNA
PCR kit (Takara, Tokyo, Japan) according to the manufacturer’s instructions. For the
PCR amplification of the total RNA from the salivary glands of unfed and partially fed
ticks, the specific primers of each cDNA clone were used. The primers specific for the
H. longicornis actin gene were used as an internal control for the RT-PCR reaction. A
negative-control reaction was simultaneously performed under the condition without
adding reverse-transcriptase to exclude some contamination of genomic DNA.
3-3. Results
The amplified salivary gland cDNA library was screened with polyclonal rabbit
anti-H. longicornis tick immune serum. Screening of 5 X 105 pfu yielded 17 positive
clones. PCR amplification showed that positive clones contained inserts with sizes
ranging from 0.5 to 1.7 kbp. As shown in Table 4, the BLASTP search indicated that 8
amino acid sequences were novel proteins; 8 matched with hypothetical proteins in H.
longicornis; and one with the H. longicornis HL35 antigen U (115). Ten of 17
sequences were found to possibly contain a secretory signal peptide. The remaining 7
sequences appeared to be truncated since starting methionine or 5’UTR were not found
(marked as “T”). Multiple amino acid sequence alignments of 8 novel proteins are

59
shown in Fig. 14. These sequences shared 23-97% identity to each other. Three
sequences, designated hlim10, hlim11, and hlim20, shared greater than 90% identity
were, therefore, considered to be the same. The amino acid sequence alignments
suggested that these proteins would have the unique features of cement proteins
contained in tick saliva (17), as indicated by the presence of several GL[G/Y/S/L/F]
tripeptide repeats.
One sequence, designated hlim3 (accession number AB252633), matched with
the previously reported HL35 (115). This sequence also possessed the characteristics of
a cement component (17). The multiple sequence alignments of hlim3, HL35
(accession number BAB20588), and its homologue, HL34 (accession number
BAB1376), are shown in Fig. 15. Even though the amino acid sequence of hlim3
shared greater than 80% identity with HL35, several amino acid substitutions and
insertions were observed, suggesting the presence of sequence polymorphism.
The remaining identical 8 sequences shared weak similarity with the
hypothetical protein of H. longicornis (77). The calculated molecular weight was
approximately 12 kDa and acidic in nature. The prediction from the SIGNALP server
suggested that the sequence contained a secretory signal peptide.
The expression of the mRNA transcripts of immunoscreening-positive clones
in the salivary glands of unfed and partially fed ticks was studied using RT-PCR (Fig.
16). Based on the amino acid sequence identity, the clones can be arbitrarily grouped
into 8 categories, namely, 6 novel sequences (5 singletrons and one tripletron), the H.
longicornis hypothetical protein, and the HL35 homologue. The results suggested that
all 8 sequences studied were expressed in the salivary glands of partially fed ticks and 7
sequences, namely hlim1, hlim2, hlim4 (faint band), hlim13, hlim14 (faint band), hlim20,

60
and hlim21, were also expressed in unfed ticks. No obvious difference in the expression
level of 4 transcripts (hlim2, hlim13, hlim20, and hlim21) between RNA of unfed and
fed ticks could be observed, whereas the remaining 4 (hlim1, hlim3, hlim4, and hlim14)
appeared to be upregulated during blood feeding.
3-4. Discussion
The cloning and expression of several key molecules for use as tick vaccine
candidates are ongoing worldwide. The study on antigens that elicit an antibody
response is a useful strategy to identify vaccine antigens. For this purpose, the cDNA
expression library of salivary glands from the adult female tick H. longicornis was
constructed in the present study. The genes encoding for immunodominant proteins
were identified by immunoscreening using rabbit sera after repeated infestation with H.
longicornis (144). The BLASTP search of 17 positive clones revealed that 8 clones did
not yield any similarities with known proteins, 8 clones gave matches with hypothetical
proteins of H. longicornis and one clone, hlim3, shared similarity with the HL35
antigen U. The vaccination effect of HL34, the homologue of HL 35, has been
evaluated, suggesting its suitability as a tick vaccine candidate (115). The clone hlim3
shared 83% identity with the HL35 protein; however, the substitution and insertion of
several amino acids were observed from the sequence alignments, indicating the
presence of gene polymorphism.
Eight cDNA clones gave best matches with the hypothetical protein from H.
longicornis, indicating that this gene was abundantly expressed. The RT-PCR results
showed that the mRNA transcript was upregulated in salivary glands during blood
feeding. The peptides are possibly related to a secretory product, as they display a

61
secretion signal. These findings implied that this protein plays an important role during
blood feeding, although its function is still unknown.
The deduced amino acid sequences of the 8 novel protein possessed striking
feature of cement substances even though no hits to any sequence reported in the
database were obtained (17). Further localization study by immuno-staining
experiments using specific antiserum that detect the proteins at the feeding site would
confirm the possible function of these proteins. The presence of glycine triplet repeats
and the high content of glycine are characteristic features of the vertebrate extracellular
matrix protein (17). One clone, hlim4, contained a signal peptide and an 18 GLX repeat,
a representative feature of cement substances. A cement protein 36 (RIM36) of the tick
R. appendiculatus is regarded as an immunodominant molecule and a promising target
of eliciting antibodies in vaccinated cattle (17). The cement components are initially
secreted to assist in the attachment to the host tissue. The process by which cement
hardens are currently unknown and the individual protein comprising tick cement are
poorly characterized (17). Recently, transcriptome of the salivary glands of D. andrsoni
was analyzed by random sequencing of 1440 cDNA clones from the salivary glands of
adult female ticks collected during the early stages of feeding (18-24 hr) (2).
Interestingly, among all the categories, some of the highest percentage of potentially
secreted proteins was observed for the glycine-rich genes (67.7%). The relatively
abundance of messages potentially encoding cement-like protein in D. andersoni
salivary glands is consistent with the amount of attachment cement usually secreted by
ixodid ticks. Ticks from this group, which include Haemaphysalis, Dermacentor,
Rhipicephalus and Boophilus ticks, penetrate only the superficial epidermis of the host’s
skin and mechanical support is entirely dependent on the cement secreted (2).

62
With regard to vaccine targeting, no consensus agreement is obtained as to
whether an exposed or a concealed antigen is the best choice (67, 76, 140). The use of a
concealed antigen, as the commercial vaccines in cattle, can successfully induce
protective immunity against tick feeding. However, immunity to concealed antigens
may not prevent the attachment and feeding of the ticks or the transmission of tick-
borne infections. In addition, immunity from this vaccine is short-lived because natural
infestation does not stimulate a response to the concealed antigen (35, 138). Salivary
gland proteins are exposed antigens that are injected into the host during blood feeding.
Resistance to salivary gland proteins is naturally acquired by multiple tick infestation or
immunization with crude tick extracts (128, 132). The positive aspects of salivary gland
antigens as potential candidates for the regulation of tick attachment and feeding (92)
and transmission blocking vaccines (27, 84, 131) are superior beyond the drawbacks of
the reduction of their antigenicity under the pressure of host immunity (111) or their
inferior immunity compared to that induced by concealed antigens (133). In particular,
the immune response induced by the exposed antigen is expected to be continually
boosted by the natural exposure of ticks to livestock in the field.
The salivary glands of ticks produce a cocktail of molecules having different
functions (93-95, 129). One of the categories is the cement components that are
initially secreted from the salivary glands and enable the tick to attach to the host.
Several studies have shown the attractiveness of the cement protein as vaccine
candidates. Vaccination with the recombinant p29, the glycine-rich extracellular matrix
protein from H. longicornis, led to a significant reduction of the engorged body weight
in adult ticks and the mortality of both larvae and nymphs (75). Another candidate,
HL34, could induce morbidity and mortality in adult and nymphal ticks of the same tick

63
species (115). The recombinant cement cone protein RIM36 has been characterized
from R. appendiculatus, in which it was indicated that this cement component could
potentially elicit strong antibody responses in cattle exposed to feeding ticks (17).
When a putative tick cement protein (64P) from R. appendiculatus was examined as a
vaccine target, a cross-reactivity of anti-sera raised against several truncated forms of
recombinant 64P with several tissue extracts, suggesting that this cement protein was a
promising candidate targeting both exposed and concealed antigens (112). Moreover, it
was shown that vaccination with the recombinant 64P cement protein induced
protective humoral and cell-mediated immune responses in addition to antigenic cross-
reactivity with the other tick species, suggesting its potential for use as a broad-
spectrum anti-tick vaccine (113).
In the present study, I identified immunodominant antigens from the salivary
glands of H. longicornis and several novel putative cement proteins were obtained.
Further study on the recombinant expressed individual protein will clarify the role of
these proteins for tick attachment and their possible use in a recombinant cocktail
vaccine.
3-5. Summary
The components of tick saliva have many effects that may aid the vector during
engorgement and could be important in evading host responses. Animals acquire
resistant to tick bites, a phenomenon known as tick immunity is partially mediated by
antibody. The present study showed the results of immunoscreening of cDNA
expression library from salivary glands of H. longicornis using tick sensitized rabbit
serum. Screening of 500,000 library clones yielded 17 positive clones. As a result of

64
BLASTP search, 8 genes gave best match to hypothetical protein from H. longicornis
and one clone matched with HL35 antigen U from the same tick species. Eight novel
proteins possess common characteristic of adhesive cement substance were obtained.
Expression of these genes was confirmed by RT-PCR. From this study, it is of great
interest in further evaluating the vaccine potency of these immunodominant antigens.

65
Table 4. Categorization of the positive clones from immunoscreening. Sequence
Id Accession
number Genbank match* Species of NR
match
E-value Signal
peptide
hlim1 AB259293 No matches found - - T hlim2 AB259292 No matches found - - T hlim3 AB252633 >gi|12060354|dbj|BAB20558.1|
HL35 antigen U
H. longicornis 1e-119 S
hlim4 AB259294 No matches found - - S hlim6 - >gi|68051227|dbj|BAE02556.1|
hypothetical protein
H. longicornis 0.055 S
hlim7 - >gi|68051227|dbj|BAE02556.1|
hypothetical protein
H. longicornis 0.055 S
hlim8 - >gi|68051227|dbj|BAE02556.1|
hypothetical protein
H. longicornis 0.055 S
hlim9 - >gi|68051227|dbj|BAE02556.1|
hypothetical protein
H. longicornis 0.055 S
hlim10 AB259299 No matches found - - T hlim11 - No matches found - - T hlim12 - >gi|68051227|dbj|BAE02556.1|
hypothetical protein
H. longicornis 0.055 T
hlim13 AB259296 No matches found - - S hlim14 AB259298 >gi|68051227|dbj|BAE02556.1|
hypothetical protein
H. longicornis 0.055 S
hlim17 - >gi|68051227|dbj|BAE02556.1|
hypothetical protein
H. longicornis 0.055 S
hlim18 - >gi|68051227|dbj|BAE02556.1|
hypothetical protein
H. longicornis 0.055 S
hlim20 AB259297 No matches found - - T hlim21 AB259295 No matches found - - T
*BLASTP performed with the Blosum62 matrix against the non-redundant (NR) protein database of NCBI; S, a secretory signal sequence was found; T, truncated sequence. Note; hlim10 and hlim11 are identical sequences (AB259299), 8 hypothetical proteins are the same sequences (AB259298)

66
Fig. 14. Multiple amino acid sequence alignment of novel proteins. The CLUSTAL
alignments of 8 novel proteins obtained by immunoscreening are shown (hlim1, hlimi2,
hlim4, hlim10, hlim13, hlim20 and hlim21; accession number AB2529293,
AB2529292, AB2529294, AB2529299, AB2529296, AB2529297 and AB2529295,
respectively). The unique features of cement protein, GLX repeats, are highlighted by
boxes. Residues conserved in all sequences are marked with black shadow. Similarity
is marked with a dark grey and light grey shadow. Note; hlim10 and hlim11 are
identical sequences (AB2529299).

67
Fig. 15. Multiple sequence alignments of the HL35 homologues. Amino acid sequence
alignments of hlim3 (AB252633), previously reported HL35 (BAB20588), and its
homologue, HL34 (BAB1376), are shown. The GLX repeats are underlined. Residues
conserved in all sequences are marked with black shadow. Similarity is marked with a
dark grey shadow.

68
Fig. 16. Analysis of mRNA encoding for immunodominant antigens in salivary glands
of unfed and partially fed female H. longicornis. Total RNA was isolated from the
salivary glands of unfed and partially fed female ticks. The RT-PCR reaction with
specific primers of each gene was performed. Primers specific for the H. longicornis
actin gene were used as an internal control for the RT-PCR reaction.
Unfed fed
hlim3
hlim1
hlim2
hlactin
hlim13
hlim14
hlim20
hlim21
hlim4

69
Chapter 4
Characterization of Haemaphysalis longicornis recombinant cement-
like antigens and evaluation of their vaccination effects
4-1. Introduction
Ticks are ectoparasites found in almost all parts of the world and surpass all
other arthropods in the number of diseases they transmit to animals and humans. Tick-
borne diseases in domestic animals are major constraints in livestock production,
especially in developing countries. Although tick control strategies rely extensively on
the use of chemical acaricides, this type of control is becoming less sustainable due to
the development of resistance against acaricides among ticks and the undesirable
contamination of the environment and animal food products (137). The limitation in the
current control measures against ticks and tick borne diseases have stimulated research
in alternative method. Anti-tick vaccination may be an ideal, affordable, and
sustainable way of controlling tick-borne diseases. There is currently only one
commercially available tick vaccine against R. (B.) microplus (54, 135). This midgut
membrane-bound protein antigen (BM86) can successfully induce protective immunity
against tick feeding. Antibodies bind to epitopes on the midgut cells of the feeding tick
causing damage and leakage of blood into the body cavity, killing the tick or reducing
fecundity. Concealed antigens are those found on the tick gut wall and normally not
presented to the host. Therefore, immunity to concealed antigens may not prevent the
attachment of the ticks or the transmission of tick-borne infections. In addition,
immunity from this vaccine is short-lived because natural infestation does not stimulate
a response to the concealed antigen (35, 138).

70
Salivary gland proteins are exposed antigens that are injected into the host
during blood feeding. Resistance to salivary gland proteins is naturally acquired by
multiple tick infestation or immunization with crude tick extracts (128, 132). As such,
salivary gland antigens are potential candidates for a transmission blocking vaccine (84,
131) and the regulation of tick attachment and feeding (92). Immune responses induced
by exposed antigens are expected to be continually boosted by natural exposure of
livestock to ticks in the field. Several exposed antigens have been identified, expressed
as recombinant proteins, and evaluated as vaccine candidates (75, 112, 113, 115). One
of such promising target molecule is the cement protein (112, 113). The cement
components are initially secreted to assist in the attachment to the host tissue. A cement
protein, RIM36, of the tick R. appendiculatus, is regarded as a strong antigenic
molecule target for eliciting antibodies in cattle (17). Furthermore, a putative tick
cement protein (64P) from R. appendiculatus is another vaccine target of interest (112).
A cross-reactivity study of anti-sera raised against several truncated forms of
recombinant 64P with several tissue extracts suggested that this cement protein may be
a promising candidate targeting both exposed and concealed antigens for use as a broad-
spectrum anti-tick vaccine (113).
From chapter 3, I identified immunodominant antigens from the salivary
glands of H. longicornis by immunoscreening. Several sequences that code for putative
cement proteins were obtained. Further studies of recombinant expressed individual
proteins were performed to clarify the role of these proteins for tick attachment and
their possible use as a recombinant cocktail vaccine. In the present study, 2 genes
encoding cement-like antigens, hlim2 and hlim3, obtained from the salivary gland
cDNA library of H. longicornis, were selected for expression as recombinant proteins.

71
Their potential as anti-tick vaccine candidates is discussed herein.
4-2. Materials And Methods
Ticks
The parthenogenetic Okayama strain of H. longicornis has been maintained by
feeding on rabbits and mice for several generations in our laboratory since 1997. The
adults and nymphs were used in this experiment (36).
Animals
All animal experiments were conducted in accordance with the Standards
Relating to the Care and Management of Experimental Animals promulgated by
Obihiro University of Agriculture and Veterinary Medicine, Hokkaido, Japan
(Allowance No. 17-77).
Nucleotide accession numbers
The nucleotide sequences reported in this thesis will appear in the
DDBJ/EMBL/GenBank nucleotide sequence databases with the accession numbers
AB252633 and AB259292.
Analysis of hlim2 and hlim3 mRNA expression in different tick tissues
Total RNA was prepared from salivary glands, midguts, ovaries, synganglions,
and carcasses (remnants of ticks after the removal of salivary glands, midguts, ovaries,
and synganglions) of partially fed ticks using the TRI® reagent, as previously described
in chapter 1. RT-PCR was performed using the Takara one-step RNA PCR kit (Takara,

72
Tokyo, Japan) according to the manufacturer’s instructions. Primers specific for the H.
longicornis actin gene were used as an internal control for the RT-PCR reaction. A
negative-control reaction was performed without adding reverse-transcriptase to
exclude some contamination of genomic DNA.
Subcloning of hlim2 and hlim3 cDNA sequences into the pGEX-4T-3 expression
vector
The PCR amplification of the pBluescript phagemid harboring the hlim2 and
hlim3 cDNA sequence for expression was performed using primers flanking the coding
sequences of each gene. A truncated hlim2 was amplified using the hlim2 forward
primer fused in-frame with the GST gene (5’AGGAATTCGGCACGAGGGGGA3’) and
the hlim2 reverse primer (5’GCGAATTCTTATGACCGTGCA3’). The full-length
hlim3 was amplified using the hlim3 forward primer
(5’GCGAATTCTCATGACGCCTGT3’) and the hlim3 reverse primer
(5’GCGAATTCCATCATGAAGATA3’). The start codon (ATG) and stop codon (TTA,
TCA) are indicated in bold. The EcoRI restriction sites are indicated in italics. The
PCR reaction conditions were as follows: 94°C for 10 min, 30 cycles of 94°C for 1 min,
50°C for 1 min, 72°C for 2 min, and 1 cycle of 72°C for 7 min. The expression vector
pGEX-4T-3 (Amersham Pharmacia Biotech, Tokyo, Japan) was digested with EcoRI,
treated with calf intestinal alkaline phosphatase (CIAP), and purified using the
Geneclean II kit (Q-BIO gene®, Bio101 system, MP Biomedicals, CA, USA). The
insert DNA was digested with EcoRI, purified using Geneclean, and ligated into the
compatible ends of pGEX-4T-3 using the DNA ligation kit version 2.0 (Takara, Tokyo,
Japan) according to the manufacturer’s instructions. The ligation product was

73
transformed into E. coli DH5∝. Transformants grown on the plate were selected for
plasmid DNA mini-preparation using alkaline lysis methods. After restriction digestion
analysis, positive clones with correct orientation were selected for sequencing. The
recombinant constructs were transformed into E. coli BL21 cell for expression.
Expression and purification of the GST-hlim2 fusion protein
A single colony of E. coli BL21 haboring the recombinant pGEX-hlim2 plasmid
was grown overnight in 5 ml LB-ampicillin broth. The culture was diluted 1:50 in 200
ml of a fresh LB-ampicillin medium and grown until O.D600 reached 0.4. Then,
isopropyl-beta-D-thiogalactopyranoside (IPTG) induction was performed with a final
concentration of 0.5 mM at 25°C overnight. The cells were pelleted by centrifugation at
4,000 ×g for 20 min at 4°C. The pellet was resuspended in a 10 ml TNE buffer (50
mM Tris-HCl pH 7.5, 2 mM EDTA, 0.1 M NaCl) and further incubated at room
temperature for 10 min in the presence of a 20 μg/ml final concentration of lysozyme.
The cell suspension was lysed by sonication 3 times, 2 min each time, and incubated on
ice for 1 hr with 1% Triton-X 100 in PBS. After repeat sonications, the suspension was
centrifuged at 15,000 ×g for 20 min at 4°C. The supernatant containing soluble GST-
hlim2 was collected for further purification by glutathione sepharose 4B according to
the manufacturer’s instructions (Amersham Pharmacia Biotech, Tokyo, Japan).
Expression of GST-hlim3 fusion protein
The expression of E. coli BL21 harboring the recombinant pGEX-hlim3 plasmid
was performed in a 1 L culture at 37°C for 4 hrs with a 1 mM final concentration of

74
IPTG. After the cells were lysed by sonication, the suspension was centrifuged at
15,000 ×g for 10 min at 4°C, and the supernatant was discarded. The pellet was
washed 3 times by sonication in 1% triton-X-PBS and one time in PBS. The pellet was
resuspended with 10 ml of 8 M urea in a urea-free buffer (50 mM Tris-HCl pH 8.0, 1
mM DTT, 1 mM EDTA). The mixture was incubated at room temperature for 1 hr and
centrifuged at 15,000 ×g for 20 min at 4°C. The supernatant was transferred to a
dialysis tube, and slow removal of urea was performed by stepwise dialysis against 1 L
of a 4 M and 2 M urea solution, one hour each, and with a urea-free buffer 3 times, 30
min each time. The supernatant was further dialyzed against 2 L of a urea-free buffer
overnight at 4°C. After centrifugation at 15,000 ×g for 20 min at 4°C, the supernatant
was collected. Determination of the fusion protein concentration was performed by
SDS-PAGE in comparison to the known concentration of the standard BSA protein.
Immunization of mice using GST-hlim2 and the GST-hlim3 fusion protein
Nine ddy mice (female, 6-weeks old) were divided into 3 groups for the
immunization and challenge infestation experiment. The animals were immunized for
totally 4 times with the control GST protein, GST-hlim2, and the GST-hlim3 fusion
protein. First, immunization was performed by injecting 100 μg of the protein mixed
with equal volume of Freund’s complete adjuvant intraperitoneally. The booster was
given at 2-week intervals with the same amount of protein mixed with Freund’s
incomplete adjuvant. Blood samples were collected from the mice tail vein after the
third booster. The host immune response to the immunization was analyzed by Western
blot analysis of immunized mouse sera with the immunoblots of recombinant proteins.

75
Immunization of rabbits using GST-hlim2 and the GST-hlim3 fusion protein
Six Japanese white rabbits were immunized for totally 4 times with the control
GST protein, GST-hlim2, and the GST-hlim3 fusion protein. First, immunization was
performed by subcutaneous injection of 500 μg of the protein mixed with Freund’s
complete adjuvant. The booster was given at 2-week intervals with the same amount of
protein mixed with Freund’s incomplete adjuvant. Blood samples were collected from
the marginal ear vein after the third booster.
Analysis of native hlim2 and hlim3 in salivary glands by Western blotting
Salivary glands of 4-day fed H. longicornis were dissected, washed with ice-
cold PBS, and kept at -80°C until used. The tissue was homogenized using a tissue
homogenizer, sonicated for 15 s 3 times, and centrifuged at 15,000 ×g for 20 min at
4°C. The supernatant was collected, and the protein concentration was measured using
a BCA kit (Pierce, Rockford, IL, USA). Salivary gland extract was used as an antigen
for Western blotting by electrophoresing on a 12% polyacrylamide gel. The protein was
transferred onto a polyvinylidine difluoride (PVDF) membrane. The membrane was
incubated in 3% skim milk in PBS for 1 hr at room temperature and further incubated
with mouse sera (1:100) and goat anti-mouse-conjugated horseradish peroxidase (Dako
Cytomation, Tokyo, Japan, 1:2,000). The signal was detected by 3,3-diaminobenzidine
tetrahydrochloride (DAB) staining.
Challenge infestation

76
The antibody titer of animal immunized with GST-hlim2 and GST-hlim3 was
evaluated using ELISA and Western blot, respectively. When the antibody titer reached
1:5,000 to 1:8,000 dilution, the animal were challenged with ticks. Three ddy mice per
group were used for the challenge infestation. Ten days after the third booster, 12 H.
longicornis nymphs per mouse were applied to the backs of the mice with the help of a
polypropylene cap fixed with cement mass as described elsewhere (59). Sixty unfed
adult ticks were applied on shaved ears of immunized rabbits as described elsewhere
(36). The vaccine effect was determined 24 hrs later by visual examination of the
attachment rate (number of tick attach on the host) and duration of feeding (the time
since attachment was observed until engorgement was completed). The engorged body
weights, molting rate, egg weight, and mortality rates were determined post-
engorgement.
Statistical analysis
All data are presented as a mean ± standard error or percentages where
applicable. Differences are considered to be statistically significant if the P value was
less than 0.05 in the Student’s t test.
4-3. Results
Sequence analysis of hlim2 and hlim3
The nucleotide sequences and deduced amino acid sequences of hlim2 and hlim3
are shown in Fig. 17. The hlim2-truncated sequence (Fig. 17 a) (accession number
AB259292) is 696 bp in length, with an open reading frame (ORF) of 539 bp. The
deduced amino acid is 179 aa with an expected molecular mass of 16.5 KDa and a pI of

77
8.85. This protein has no similarity with any sequence reported in the database. The
hlim2 protein has high glycine (34.6%) and serine (14.53%) contents. Several glycine-
rich repeats (GGXG, GXGG) were observed, as reported for the R. appendiculatus
glycine-rich protein (67). The presence of GLX repeats, the common features of
cement component were shown.
The hlim3 full-length sequence (Fig. 17 b) (accession number AB252633) is
1,089 bp, has an ORF extending from position 40 to position 1,003, and codes 321
amino acid residues with a predicted molecular mass of about 33 kDa and a pI of 9.1.
This deduced amino acid has a secretory signal peptide signature. The hlim3 sequence
shared 41% and 83% identities with previously reported H. longicornis salivary gland
proteins, HL34 and HL35, respectively (115). The common features of the adhesive
protein, YPG motifs, within the tyrosine-rich domain or PXP motifs in the c-terminal
proline-rich domain were observed as described by Tsuda et al (115).
The expression of hlim2 and hlim3 mRNA transcripts in different tick tissues
The expression of the mRNA transcripts of hlim2 and hlim3 was studied using
RT-PCR (Fig. 18). Both mRNA transcripts were expressed predominantly in salivary
glands. The hlim2 transcript was expressed in every organ except the midgut. For hlim3,
the mRNA expression was observed in the salivary gland, synganglion, and carcass. The
RT-PCR internal control was performed using the actin gene.
Expression and purification of recombinant hlim2 and hlim3
The heterologous expression of recombinant hlim2 in E. coli was performed.

78
The GST fusion protein with an expected size of 42 kDa was obtained (Fig. 19 a). The
recombinant hlim3 was expressed with the expected size of 59 kDa. Although the
expression conditions were optimized with various concentrations of IPTG at different
temperatures, the protein did not become soluble (data not shown). For this reason,
hlim3 was expressed as the inclusion body, solubilized using urea, and refolded by
stepwise dialysis as described in materials and methods (Fig. 19 b). The purification of
the refolded hlim3 gave a low yield (less than 300 μg from 1 L culture). Therefore, the
hlim3 inclusion body was used for immunization.
Analysis of native hlim2 and hlim3 in salivary glands by Western blotting
The Western blotting of a salivary gland extract and mouse anti-hlim2 and anti-
hlim3 is shown in Fig. 20. The mouse anti-hlim2 reacted with 64 kDa and 26 kDa
proteins, whereas anti-hlim3 reacted with a 33 kDa protein from a salivary gland extract.
The reaction with the 26 kDa protein possibly resulted from a cross-reaction between
the anti-GST antibody and the GST protein in the salivary glands of ticks (50, 112, 113)
It is not known whether the molecular mass of the native hlim2 is 64 kDa, since the full-
length sequence of this clone is not yet available.
Challenge infestation in mice
The challenge infestation was performed as described in materials and methods.
The attachment rate of ticks fed on GST-hlim3-immunized mice (34.2 ± 17.6%) was
significantly lower (p < 0.05) than that in the control group immunized with GST
(100%) after the first day of tick application (Fig. 21). The feeding duration was

79
prolonged, as indicated by the significantly lower engorgement rate (p < 0.05) on day 3
(4.8 ± 3.9%) and day 4 (23.8 ± 3.8%, p < 0.01) of feeding (Fig. 22). Most of ticks
(95%) were engorged after 6 days of feeding. In contrast, GST-hlim2-immunized mice
exhibited no apparent differences in attachment and feeding periods from those
observed in the control group. However, the body weights of engorged ticks fed on
GST-hlim2-vaccinated mice (3.93 ± 3.9 mg) were significantly lower (p < 0.01) than
that of the control group (4.37 ± 0.05 mg) as shown in Fig. 23. The vaccination was
found to have no effect on tick molting, since all ticks were able to progress to the adult
stage.
Challenge infestation in rabbits
Challenge infestation in rabbits immunized with recombinant proteins was
performed using adult female ticks. There were no significant difference in the
attachment rate or feeding period of ticks fed on GST-hlim2 or GST-hlim3 immunized
rabbits in comparison to the control group. However, significant reduction of engorged
body weight of tick following feeding on GST-hlim2 (147 ± 45 mg, p < 0.01) and GST-
hlim3 (163 ± 37 mg, p < 0.05) in comparison to the control GST immunized group (184
± 32 mg) was observed (Fig. 24). An apparent reduction in egg weight was observed
for adult ticks fed on GST-hlim2 immunized rabbit (120 ± 13 mg, p < 0.05) compared
to the control value (153 ± 15 mg) (Fig. 25). However, this affect cannot be observed in
tick fed on GST-hlim3 immunized rabbit (150 ± 29 mg).
4-4. Discussion
Several studies have demonstrated that vaccination with defined protein

80
antigens is able to induce significant immunity to tick infestation (140). As a vaccine
target, salivary glands are a rich source of exposed antigens that can stimulate the host
immune response following natural infestation. As reported in chapter 3, I identified 17
genes encoding immunodominant antigens from the salivary glands of H. longicornis.
These genes were obtained by immunoscreening of a salivary gland cDNA library using
a tick-sensitized rabbit serum. Of the 17 positive clones obtained, 8 sequences did not
yield any similarities with known proteins, in addition that one sequence shared
similarity with the HL35 antigen U. These 9 sequences were found to possess
characteristics in common with cement proteins that may assist ticks in their attachment
to the host during blood feeding. To test whether or not this immune response was
protective, individual genes were selected for expression as recombinant proteins and
the vaccine potency was evaluated. In this study, 2 genes that gave a strong positive
reaction with a rabbit serum, hlim2 and hlim3, were selected for further study. The
hlim2 mRNA transcript was equally expressed in the salivary gland of unfed and fed
ticks, whereas hlim3 was upregulated during blood feeding. The analysis of mRNA
expression in various tick organs suggested that both genes were also expressed in
tissues other than the salivary glands. These 2 genes were subcloned and expressed as a
recombinant GST fusion protein using a bacterial expression system. The recombinant
GST-hlim2 was expressed as a soluble protein at 25°C, whereas GST-hlim3 was
expressed as an inclusion body. The recombinant GST-hlim2 and GST-hlim3 were used
for the immunization of mice. Challenge infestation was performed using H.
longicornis nymphs and adult ticks fed on vaccinated mice and rabbits, respectively. In
comparison to control GST, the ticks that fed on GST-hlim2-immunized mice had
significantly lower engorged body weight, although the attachment and engorgement

81
process was unaltered. On the other hand, ticks fed on mice vaccinated with GST-hlim3
showed a significantly lower attachment rate 24 h post-infestation, which prolonged the
engorgement rate on day 3 and day 4. However, those ticks were later attached and fed
until repletion with no significant difference in their engorged body weight.
Vaccination of rabbits using GST-hlim2 and GST-hlim3 separately could reduce
engorged body weight of adult ticks. However, apparent difference in oviposition can
be observed only in ticks fed on GST-hlim2 immunized rabbits. These results suggested
that antibodies against immunodominant cement-like antigens in vaccinated animals
could interfere with tick feeding in some way.
Both hlim2 and hlim3 proteins contain glycine repeats and a glycine-rich
region. The glycine-rich proteins have been found to be expressed in the salivary
glands of R. appendiculatus (17, 81, 112) and A. variegatum (80) and are believed to be
a component of tick cement, the proteins of which are rich in glycine, serine, leucine,
tyrosine, and proline (65). A tick attachment site usually consists of a cement cone that
protects the tick hypostome during feeding and secures the site of skin penetration (65).
Since cement proteins are essential for tick attachment and feeding on the host, there is
interest in using cement proteins or their derivative as anti-tick vaccine components.
Immunization of rabbits with purified cement proteins elicit delayed-type
hypersensitivity to the antigen (107). The hypersensitive rabbit demonstrated resistance
to feeding of the R. appendiculatus tick but a slightly enhanced feeding of
Rhiphicephalus pulchellus ticks. Vaccination with recombinant p29, glycine-rich
extracellular matrix protein from H. longicornis led to a significant reduction of the
engorged body weight in adult ticks and the mortality of both larvae and nymphs (75).
HL34, another candidate, was reported to induce morbidity and mortality in adult and

82
nymphal stages of the same tick species (115). The recombinant hlim3 studied in this
work shared 41% amino acid identity with HL34. However, no mortality of ticks fed on
vaccinated mice was observed. Instead, this recombinant protein seemed to interfere
with tick attachment with an unknown mechanism. Visual observation indicated that
ticks fed on vaccinated animals accumulated on the polypropylene cap instead of
seeking a host.
A putative tick cement protein (64P) from R. appendiculatus is currently a
promising vaccine candidate that was used for vaccine targeting for both exposed and
concealed antigens (112). As reported with regard to the vaccination effects of cement
protein 64TRP, most adult R. sanguineus on vaccinated guinea pigs were unattached
and observed to be crawling on the retaining gauze as if attempting to leave the host 24
hr post-infestation (113) As discussed by those authors, if the ticks had not been
restrained within the feeding chambers, they would most likely have abandoned the
immunized animals and sought alternative hosts.
Cement is among the initial chemical compounds that ixodid ticks secreted and
inject into the feeding site after proboscis penetrate host skin (17). Cement is critical for
tick attachment and prevention of host immune response molecules for interacting with
proboscis. Since the 9 sequences including hlim2 and hlim3 identified in the present
study possessed common characteristic with cement protein, it is conceivable to
speculate the mice immunized with hlim3 mounted a strong immunity against cement
proteins that prevented tick attachment and subsequent feeding.
In the present study, the recombinant putative cement proteins from H.
longicornis were expressed, characterized, and preliminarily evaluated their vaccine
potency. The complementary effects of hlim2 on tick engorged body weight and hlim3

83
on tick attachment indicate the suitability of the 2 recombinant proteins for use in a
cocktail vaccine.
4-5. Summary
Several immunodominat antigens were identified from salivary gland cDNA
library of H. longicornis by immunoscreening. Two genes encoding cement-like
antigens were selected for expression as recombinant proteins in E. coli. Immunization
of animals with individual recombinant protein affected blood feeding and oviposition
of immature and mature ticks. Cement protein is one of the promising vaccine
candidates for several tick species. The result obtained from this study suggested the
possibility to use these recombinant proteins for future development of cocktail vaccine
against tick infestation.
1 GGCACGAGGGGGAGCTGGACTTGGAGCCCCTGGCGCTAGCTTGCCAGGAGCCTCCGGAAG
1 GGCACGAGGGGGAGCTGGACTTGGAGCCCCTGGCGCTAGCTTGCCAGGAGCCTCCGGAAG
a)

84
A R G G A G L G A P G A S L P G A S G S 20
61 CGGCAGCCCTGCTCTAAGCGGTCAACCAGGATTCGGCTTCGGAGGCTTCTCCGTCGCACC
G S P A L S G Q P G F G F G G F S V A P 40
121 CTATGGCGGTCAATGGGGTGTCCCGCAAGGCTTCTATGGAGTGTACCCAGGATTCAGCTA
Y G G Q W G V P Q G F Y G V Y P G F S Y 60
181 CGGATTCCCCGGTTACGGTAACGGTTATGATGCCTTCGGATTGGGTGGTGGATACGGCGG
G F P G Y G N G Y D A F G L G G G Y G G 80
241 CTACGGACCCTTCGGTGTCAACGGCTTCTACTCTCCACTCGCCGGCTACGGCTTCGGCGG
Y G P F G V N G F Y S P L A G Y G F G G 100
301 CTATGGCCTGGGCGGCTACCCTGCCCTCGGCGGCTACGGCTTCGGTGGCTATGGCTACCC
Y G L G G Y P A L G G Y G F G G Y G Y P 120
361 ATTCTTCGGTGGATTGGGAGGTGCTTACGGCTTTGGCCCCTCAGGTTTCTCCGGTAGCAG
F F G G L G G A Y G F G P S G F S G S S 140
421 CGCAAGCCAGAGTTCCCTGGGTGGTGCAGGTCTAGGTGCATCCGGCGCTTCTGGAGCGGC
A S Q S S L G G A G L G A S G A S G A A 160
481 AGCAGGCTCCAGCGCCAGGTCCGGATCCAGCTCCAGCAGCGGCGCCGCTGCACGGTCATA
A G S S A R S G S S S S S G A A A R S *
541 AGCGTAACTTCTGCCCTAACTAAGTTACTACCAACAAATTTCTTGACGGGCTTTCAAGGC
601 TGTTCTATATTTGGGCTGACCAACCTACAGGTTACAACGAGTCTAAAAACAATAAATTA
661 TGATTGTTCAATTTGAAAAAAAAAAAAAAAAAAAA
Fig. 17. Nucleotide sequences of genes encoding hlim2, hlim3 and their deduced amino
acid sequences.
a) Truncated nucleotide sequence and deduced amino acid sequence of hlim2. The stop
codon (TAA) is underlined with a thick line. Several glycine-rich repeats (GGXG,
GXGG), as reported for the R. appendiculatus glycine-rich protein motifs are
underlined. The GLX repeats, the common features of cement component are
highlighted by boxes.

85
1 GGCACGAGGCCGACCAGCATCACAGGAGAACGCGTAATCATGAAGATATCCGCTGCGCTC
M K I S A A L 7
61 ACACTTCTTTCCATGGTAGCCTATGCGGCAGGAGCCGTGAAGACCTTCTATAAAGCGGCA
T L L S M V A Y A A G A V K T F Y K A A 27
121 AAGCTCAAGAGCACTGTTGCTGCCGCTGTTCCCGTAGCTGTGTCACCCACTGAGGCTTTT
K L K S T V A A A V P V A V S P T E A F 47
181 CCTGAGCTCCTTGGTTCGACCTTGGTTAATGGTGGCACGGGTACGGTAGCAGTTTCGTCC
P E L L G S T L V N G G T G T V A V S S 67
241 ACCAGCAATCCTGTTGGTACCAGCGTCAGTCATGGAACCACTACTCACCAGGTCACTCGA
T S N P V G T S V S H G T T T H Q V T R 87
301 ACGAACATATATCCTGGCATCCCTGGCGATGCTTACTACAACCCCTATTTCCCAGCAGGC
T N I Y P G I P G D A Y Y N P Y F P A G 107
361 TATCCTGGATACGGTACAGGGCTGTACCCTGGCTTCCACAGCGGAGCTTATAACAGCTAC
Y P G Y G T G L Y P G F H S G A Y N S Y 127
421 CTCCGTTGGGCTTACCCTGGATTAAGTGCAGGTTTTTATCCCGGCTTCTTTCACTCAACT
L R W A Y P G L S A G F Y P G F F H S T 147
481 CGTCATGGATTCTCTAGCCCACTTTACCCTGGCCTTCTGGAAGCATCTCAGCACGGTCTT
R H G F S S P L Y P G L L E A S Q H G L 167
541 TTTGGAGGAGCCTACTCCAGATACCTGAGCGGAGTGTATCCTGGACTCTTCGGAGGGGAA
F G G A Y S R Y L S G V Y P G L F G G E 187
601 TACGGAAGTCTCTATGGCTCTCGCTTCTTCGGCAACGGTGCTTTGGAGAAGGTCCGAACG
Y G S L Y G S R F F G N G A L E K V R T 207
661 CATTTCGCAGGATACCAGAGAGAAGTTACTGTCACCGTCGACCCACTGACTGGGCTACCA
H F A G Y Q R E V T V T V D P L T G L P 227
721 GCGCCAGTTACGATTCCTGTTGCCAACCGCGTCGTGACGATCGAGCGCTTCGTACCATTC
A P V T I P V A N R V V T I E R F V P F 247
781 CCATCTCCGTATCCAGTACCGGTCCCAGCTCCTCACCCAGCCCCGTTCCCGTTGCCGCAC
P S P Y P V P V P A P H P A P F P L P H 267
841 CCTGGAGTCTCCAACCCAGTTTCGTCAACTACTCAAATGGGATCTACCACCACAGTAAGC
P G V S N P V S S T T Q M G S T T T V S 287
901 CAAGGTGGACTGGTCACTCCAGTCTCGGGTGGTGCAGATCTCGTTTCCGGCCAACAGGAA
Q G G L V T P V S G G A D L V S G Q Q E 307
961 GCTGTTGCGCAGACCCAGGCCCATGTTAATTTACAGGCGTCATGAAGACGGGTAATGTCA
A V A Q T Q A H V N L Q A S *
1021 CGCGACAAGCTTCGCAAAACAGTTAAATAAAATGGAATGTGAACAAAAATTAAAAAAAAA
1081 AAAAAAAAA
b)

86
b) Full-length nucleotide and deduced amino acid sequence of hlim3. The signal
peptides are underlined with a double line. The start (ATG) and stop (TGA) codons are
underlined with a thick line. The common features of the adhesive protein, YPG motifs,
within the tyrosine-rich domain or PXP motifs in the c-terminal proline-rich domain are
shown. The tyrosine-rich and proline-rich regions are underlined with a broken line. The
YPG motifs are indicated in bold, and the PXP motifs are in italics. The GLX repeats
within the deduced amino acid are highlighted by boxes.

87
Fig. 18. Analysis of hlim2 and hlim3 mRNA expression in different tick tissues.
The mRNA expression of hlim2 (a), hlim3 (b) and internal control actin (c) in different
tick organs was studied by RT-PCR. Lane 1, salivary glands. Lane 2, midgut. Lane 3,
ovary. Lane 4, synganglion. Lane 5, remnants of carcass after removal of the salivary
glands, midgut, ovary, and synganglion.
bp Mr 1 2 3 4 5
600 400
bp Mr 1 2 3 4 5
600 400
bp Mr 1 2 3 4 5
400 200
(a)
(c)
(b)

88
Fig. 19. The SDS-PAGE of purified recombinant GST/hlim2 and GST/hlim3. (a) Mr =
Low range molecular weight marker, hlim2 = purified GST/hlim2. (b) Mr = Low range
molecular weight marker, hlim3 = GST/hlim3 inclusion body.
kDa Mr hlim3
97
30
22
66
45
97
30
22
66
45
kDa Mr hlim2
a) b)

89
Fig. 20. Analysis of native hlim2 and hlim3 in salivary glands by Western blotting
Western blotting of salivary gland extracts from female ticks reacted with mouse sera.
Mr, pre-stained marker. Lane 1, anti-GST mouse serum. Lane 2, anti-hlim3 mouse
serum. Lane 3, anti-hlim2 mouse serum.
Mr 1 2 3
98
22
16
64
50
36
kDa
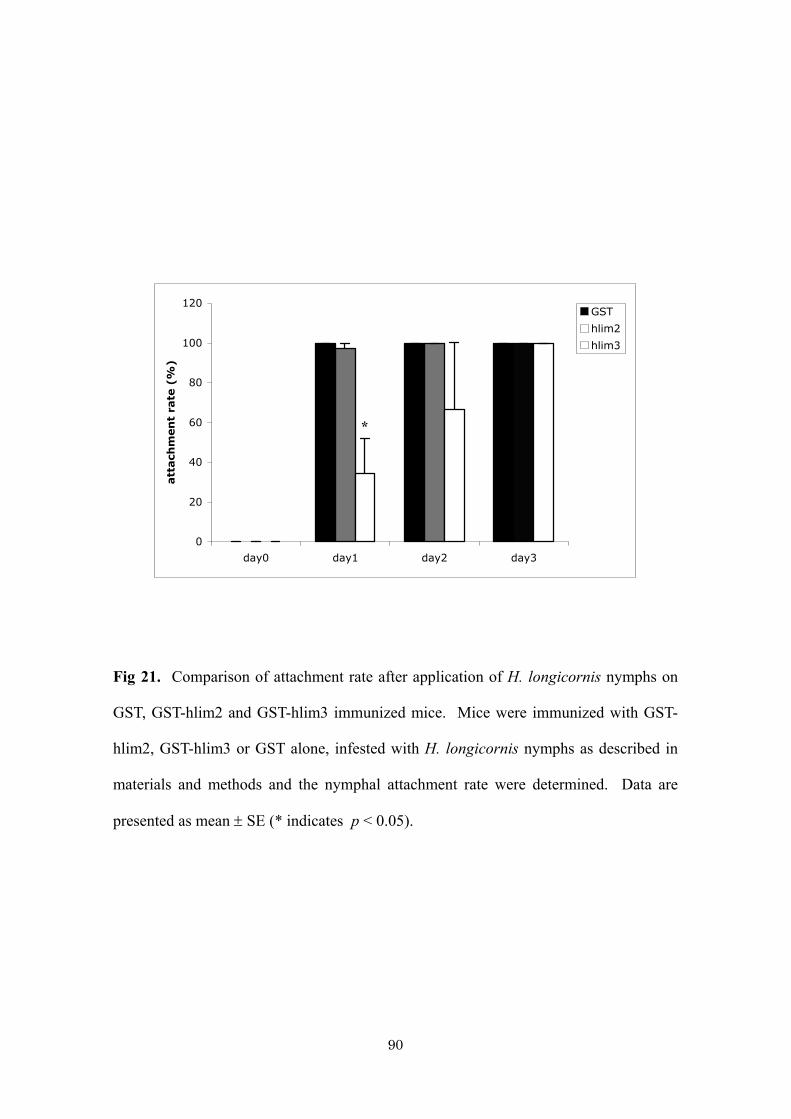
90
0
20
40
60
80
100
120
day0 day1 day2 day3
att
ach
men
t ra
te (
%)
GST
hlim2
hlim3
Fig 21. Comparison of attachment rate after application of H. longicornis nymphs on
GST, GST-hlim2 and GST-hlim3 immunized mice. Mice were immunized with GST-
hlim2, GST-hlim3 or GST alone, infested with H. longicornis nymphs as described in
materials and methods and the nymphal attachment rate were determined. Data are
presented as mean ± SE (* indicates p < 0.05).
*
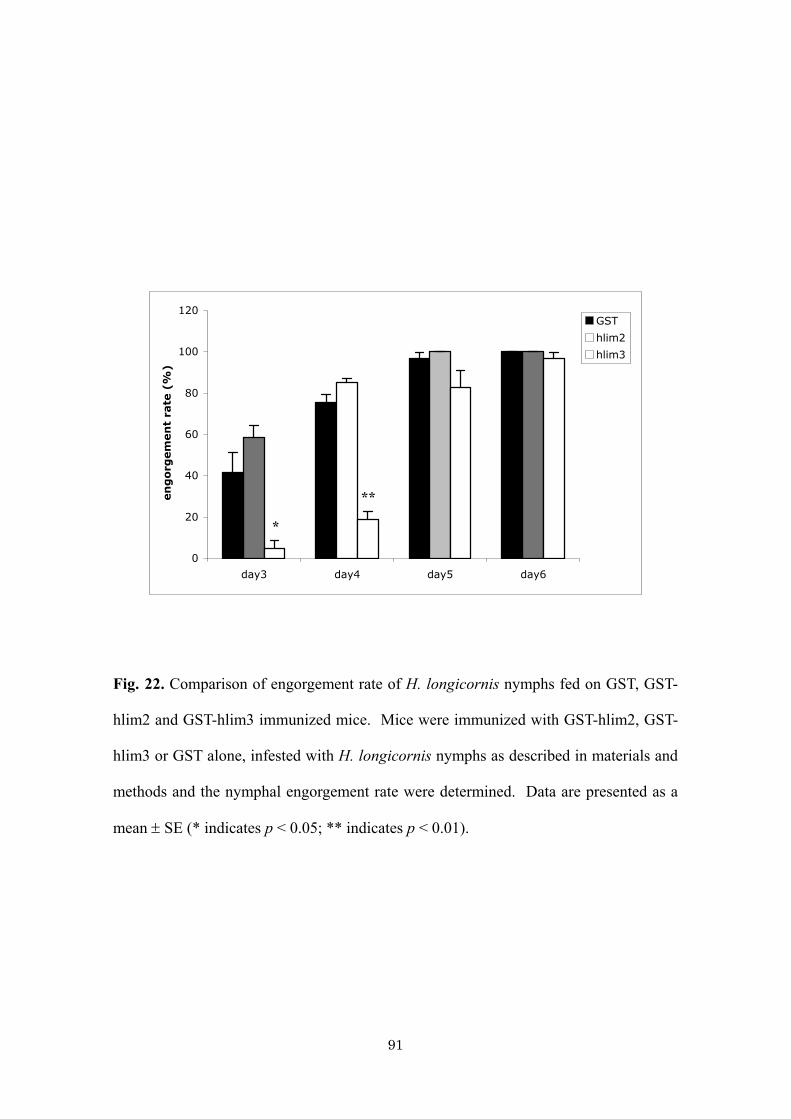
91
0
20
40
60
80
100
120
day3 day4 day5 day6
en
go
rgem
en
t ra
te (
%)
GST
hlim2
hlim3
Fig. 22. Comparison of engorgement rate of H. longicornis nymphs fed on GST, GST-
hlim2 and GST-hlim3 immunized mice. Mice were immunized with GST-hlim2, GST-
hlim3 or GST alone, infested with H. longicornis nymphs as described in materials and
methods and the nymphal engorgement rate were determined. Data are presented as a
mean ± SE (* indicates p < 0.05; ** indicates p < 0.01).
**
*

92
3.50
3.70
3.90
4.10
4.30
4.50
4.70
GST hlim2 hlim3
engo
rged
body
weig
ht
(mg) GST
hlim2
hlim3
Fig. 23. Comparison of engorged body weights of nymphal ticks fed on GST, GST-
hlim2 and GST-hlim3 immunized mice. Mice were immunized with GST-hlim2, GST-
hlim3 or GST alone, infested with H. longicornis nymphs as described in materials and
methods and the nymphal engorged body weights was determined (** indicates p <
0.01).
**

93
140
145
150
155
160
165
170
175
180
185
190
GST hlim2 hlim3
engo
rged
body
weig
ht
(mg)
GST
hlim2
hlim3
Fig. 24 Comparison of engorged body weights of adult ticks fed on GST, GST-hlim2
and GST-hlim3 immunized rabbits. Rabbit were immunized with GST-hlim2, GST-
hlim3 or GST alone, infested with H. longicornis adults as described in materials and
methods and the engorged body weights was determined (* indicates p < 0.05, **
indicates p < 0.01).
**
*

94
^
100
110
120
130
140
150
160
170
GST hlim2 hlim3
egg
weig
ht
(mg)
GST
hlim2
hlim3
Fig. 25 Comparison of egg weights of adult ticks 3 weeks after drop-off from GST,
GST-hlim2 and GST-hlim3 immunized rabbits. Rabbit were immunized with GST-
hlim2, GST-hlim3 or GST alone, infested with H. longicornis adults as described in
materials and methods and the egg weights was determined. (* indicates p < 0.05).
*

95
General discussion
Control of ticks and tick-borne diseases of livestock depends almost exclusively
on the use of acaricides (41, 89). Widespread occurrence of tick resistance to acaricides
and environmental concerns prompted research into alternative methods for controlling
ticks (41, 140). Vaccines designed to reduce tick infestation, which are deployed as part
of integrated control measures involving reduced acaricide usage, are therefore being
actively explored. Development of new tick control measures requires the identification
of candidate genes that affect immune evasion and pathogen transmission. Functional
genomics has proved a valuable tool that can accomplish this task.
Successful tick feeding and pathogen transmission are both facilitated by
pharmacologically active molecules secreted in tick saliva (21, 121). Tick salivary
gland molecules inhibit host hemostasis, reduce pain and itch responses, and modulate
host innate and specific acquired immune defenses (43, 73, 94, 103, 131). Some
components of tick saliva are highly immunogenic and vaccination with salivary gland
proteins can interfere with the tick life cycle (75, 139). The composition of tick saliva
is interesting in the study of biology of host-parasite relationship, discovery of
biologically active components and identification of novel vaccine targets. For this
reason, the cDNA library from salivary glands of H. longicornis was constructed in this
study. Besides providing basic information on the genes expressed in tick salivary
glands, I have interested in defining genes encoding components that are secreted in
saliva as these play a critical role in the biology of tick feeding (102) and pathogen
transmission (84, 131). Since the function of many sequences described still unknown,
cloning and expression of selected cDNA sequences would give a better understanding

96
of tick blood feeding mechanism. RNA interference has emerged as one of the most
promising technologies for rapid systematic functional genomic analyses. The
application of RNAi to ticks has opened the possibility of manipulating gene expression
in these organisms (25, 26, 49). RNAi has been successfully used for the
characterization of tick genes and for the screening of tick-protective antigens (24). The
application of this technology to ticks could impact on tick vaccine development
through the identification and characterization of tick-protective antigens.
As an antihemostatic molecule, tick metalloprotease(s) appears to cleave
fibrinogen and fibrin, therefore inhibiting clot formation (32, 33). Metalloprotease also
may affect endothelial cell function and angiogenesis (33). Based on the information
obtained from the random sequencing of salivary gland cDNA library of H. longicornis,
4 genes encoding metalloproteases were selected for further study. Full-length cDNA
from 2 out of the 4 genes were obtained. At the mean time, another cDNA library was
constructed using vector-capping method, which has several advantages compared to
classical lambda Zap library. By using this method, approximately 95% of cDNA
clones are full-length. The EST database was constructed by random sequencing and
annotating of 10,000 cDNA sequences. The other 2 full-length metalloproteases were
got from this EST library. Phylogenetic analysis and mRNA expression profiles during
blood feeding of each gene indicated an evolutionary distinct and high degree of
diversity of this protein. The presence of these diverse enzymes might contribute to the
greater functional complexity of this molecule in tick saliva to facilitate blood feeding.
Tick metalloproteases contain N-terminal prodomain, which, as in most zymogens or
proenzymes, is believed to maintain enzyme latency and to be proteolytically excised to
permit catalytic activity, a process termed “activation”. Since to my best knowledge

97
there is no report on the processing mechanism of tick metalloproteases, one of the goal
of this work was the expression of the proenzymes, subsequently, we can ascertain
whether the removal of its prodomain by a pro-protein convertase, as in the case of the
activation of some matrix proteases and the ADAMs (86) or by autolysis, as in the case
of snake venom metalloproteases (88). Several evidences from this study indicated that
proenzyme activation of this protein possibly occurs through furin type pro-protein
convertase dependent pathway. In order to confirm this finding, further experiments
have to be done such as site-directed mutagenesis to confirm the furin cleavage site or
baculovirus expression of the proenzme in the presence of furin inhibitor. It is also of
interest to identify furin or other enzymes responsible for activating proenzymes in tick
salivary gland. Accordingly, understanding the pathway for metalloprotease processing
may provide novel targets for the development of tick control since disruption of
proenzyme activation may inhibit the production of active enzyme, as a consequence,
interfering tick biological processes including blood feeding mechanism.
Skin lacerated by feeding ticks shows defective healing responses since these
ectoparasites are able to remain attached and fed on their host for a long period of time.
Therefore, knowledge of the host-parasite interface in terms of tissue repair can
contribute to understanding of the feeding process, transmission of infectious agents,
and potentially development of new vector-blocking vaccine candidates that could
interfere with tick attachment and at the same time prevent pathogen transmission (2).
Vaccines have been shown to be a feasible tick control method that offers a cost
effective, environmental safety alternative to chemical control. However, identification
of tick protective antigens remains the limiting step in vaccine development. Herein,
immunoscreening was used as a tool to identify 17 genes encoding immunodominant

98
salivary gland antigens that elicit antibodies in the hosts. Interestingly, several of these
proteins are rich in glycine, serine and leucine, which are prominent in the overall
amino acid content previously determined for the cement substance of the related ixodid
tick R. (B.) microplus (6). Cement is among the initial chemical compounds ixodid
ticks secreted and inject into feeding site after proboscis penetrated host skin (17) and
critical for tick attachment and prevention of host immune response molecules for
interacting with proboscis. The cement material is in contact with Langerhans cells in
the host’s skin; these dendritic cells present tick antigens to the host immune system (5,
83). Since cement proteins are essential for tick attachment and feeding on the host,
there is interest in using cement proteins or their derivative as anti-tick vaccine
components (75, 112, 113, 115, 146). Hence, recombinant expression and evaluation of
vaccine potency of these cement-like proteins was performed in this study. Initially, 2
sequences that gave strong positive reactions from immunoscreening, hlim2 and hlim3
were selected for further study. Immunization and challenge infestation of nymphal
ticks was performed in mice and that of adult ticks was performed in rabbit. The adult-
on mice model was not investigated here due to the limit number of adult tick that can
apply onto mice and difficulties in attachment of adult tick on small animal such as
mice. Immunization of GST-hlim2 affected immature and mature ticks to the same
degree as evidenced by significant reduction of engorged body weight of nymphs fed on
mice, adult fed on rabbit and also reduction of oviposition. On the other hand,
immunization of mice with GST-hlim3 showed significant lower attachment rate,
prolong feeding period but no affect on engorged body weight of nymphs. In contrast,
adult ticks fed on GST-hlim3 immunized rabbits had no apparent difference in
attachment rate but showed significant reduction in engorged body weight. Data from

99
several studies provide evidence which may support the hypothesis that immature and
mature ticks have different sensitivities to host acquired resistance against tick
molecules (63, 64, 75, 146). Finding from my finding are therefore in agreement with
their studies of different vaccine effects on nymphs and adult ticks fed on GST-hlim3
immunized animals.
Cellular immunity is an important mechanism of resistance to ticks.
Characterizing the inflammatory reactions at tick feeding sties, particularly cell
migration, will enhance understanding of the anti-tick effect of vaccines. Hosts that
develop resistance following tick infestations show a cutaneous basophil
hypersensitivity reaction, with skin infiltration by basophils in hyperplastic epidermis in
guinea pigs (4), and infiltration of neutrophils, macrophages, eosinophils and basophils
in rabbits and cattle (42, 123). From the recent study of truncated constructs of 64P, a
secreted cement protein from salivary glands of the tick R. appendiculatous suggested
that tick feeding on immunized animals stimulated local inflammatory immune
response (involving basophils, eosinophils, lymphocytes, mast cells, macrophages and
dendritic-like cells) that boosted the immune status of vaccinated animals (113).
Though the vaccination effects of cement-like antigens in my study was not conclusive,
protective response may be similar to 64P protein. Mechanism by which the
immunization affect ticks infestation may be further investigated by histo-pathological
examination at the site of infestation. The composition of the surrounding host skin
epidermal/dermal proteins would indicates a strategy whereby the tick avoids rejection
at the tick-skin attachment site by host immune defense mechanism.
Taken together, the results from this study suggested that animal immunized
with recombinant cement-like antigens could confer partial protection against ticks

100
feeding. This data, in consistent with several studies, have emphasized the fact that a
single tick molecule may not confer an inclusive protective immunity. An effective tick
vaccine will require a cocktail of target antigens each mediating a physiological
function either independently or synergistically (106). Several evidences suggested that
anti-tick immunity induced by a cocktail antigen vaccine is more effective compared to
a single antigen vaccine (98, 136). The results from my study reveal the possibility to
use cement-like antigens as a candidate for cocktail vaccine. It is of crucial interest to
evaluate the individual vaccine potency of the remaining positive clones obtained from
immunoscreening and later, in combination as a cement-based cocktail vaccine.

101
Conclusion
Since tick saliva contains a mixture of pharmacologically active compounds that
are essential for creating and maintaining the blood pool or the feeding lesion in host
skin. As a vaccine target, salivary glands would be a rich source of antigens.
Identification and characterization of tick salivary gland proteins is of considerably
interest since such information might contribute to the improvement of methods for the
control of tick and tick-born diseases. Therefore, a cDNA library from the salivary
glands of hard ticks, Haemaphysalis longicornis, was constructed in this study. Random
sequencing and immunoscreening of cDNA clones was used as a tool to recruit key
molecules that might be useful for the development of tick control strategies.
In chapter 1, a cDNA expression library of salivary glands from the adult female
H. longicornis tick was constructed and randomly sequenced. The aim of this study
was to identify salivary protein profiles of tick during blood feeding stage. Based on a
similarity search of deduced amino acid sequences using the BLASTP program, it was
found that 65 (41%) of 158 sequences are novel proteins. The remaining 93 (57%)
sequences matched sequences reported in the GenBank database. Of these 93 sequences,
25 matched those previously reported from ticks. These 93 cDNA clones were
classified according to their possible functions. The majority of the studied sequences
were housekeeping ones. Several bioactive molecules, which were expressed during
blood feeding, were found in this library; they included antihemostatic,
immunomodulatory, or cement formation proteins. Therefore, it is of great interest in
further characterization of the biological activities or evaluation of vaccine potency of
these proteins.

102
In chapter 2, among the bioactive molecules obtained in the cDNA library, 6
genes encoding metalloproteases were studied. Since this enzyme is capable of
hydrolyzing various components of the extracellular matrix and blood coagulation
related proteins, molecular cloning and characterization of these molecules was
performed in order to clarify their roles in tick feeding mechanism. Comparative
analyses have shown the evolutionary distinct and different mRNA expression patterns
of each gene during blood feeding. The proteins are synthesized in a proenzyme latent
form that requires activation for proteolytic activity. This proenzyme contains a signal
peptide, prodomain and a metalloprotease/cysteine-rich domain. The recombinant
Escherichia coli expression of one gene, hlESTMP1 was performed and used to obtain
anti-hlESTMP1 antibody. The immunoblot analysis and indirect fluorescent assay
suggested that this protein is mainly expressed in the cytoplasm of the salivary gland
cells and the mature form of 34 KDa was detectable. Since understanding the
proenzyme activation mechanisms would provide the necessary information for the
design of a selective inhibitor, the processing of the proenzyme of hlESTMP1 was
demonstrated using a baculovirus expression system. Accordingly, the signal peptide of
full-length hlESTMP1 was cleaved by signal peptidase and the proenzyme was further
processed into 2 smaller fragments. The sequences of the fragments suggested that the
enzyme activation mechanism occurs via pro-protein covertase-dependent pathway.
Thus, to identify a novel target for the development of a tick control approach, the
methods by disrupting the proenzyme activation mechanism to inhibit the production of
an active mature enzyme or by exploring metalloprotease regulators, such as inhibitors
or blockers of enzyme function, by vaccinating host animals with this bioactive
molecule, are considered.

103
In chapter 3, to identify the salivary gland antigens that elicit antibodies in the
host, immunoscreening of salivary gland cDNA library was performed using sera of
rabbit repeatedly infested with ticks. A BLASTP search suggested that from 17 positive
clones obtained, 8 sequences matched with that of hypothetical H. longicornis sequence
and one clone encoded HL35 antigen U from the same tick species. Eight of 17 gave no
match to any sequence reported in the database. Interestingly, the deduced amino acid
sequences of these novel genes possess common characteristics with cement proteins,
which assist ticks in their attachment to the host during blood feeding. The expression
of these genes in salivary glands was confirmed by RT-PCR. Four of the 8 sequences
showed to be upregulated upon blood feeding. These immunodominant antigens are of
particular interest as candidates for future cement protein based-tick vaccine. For this
reason, 2 genes encoding cement-like antigens, hlim2 and hlim3, were selected for
further elucidation of vaccine potency. In chapter 4, the results were discussed. The
recombinant proteins were expressed in E. coli as the GST fusion protein and used for
immunization. It was observed that the attachment rate of nymphal ticks fed on mice
immunized with GST-hlim3 was significantly lower than that in the control group
during the initial days of feeding. However, immunization with GST-hlim3 did not
affect the engorgement rate of the adult ticks. In sharp contrast, GST-hlim2 did not
influence the attachment rate and feeding period of ticks but had a significant reduction
in the engorgement body weight. Vaccination of rabbits with GST-hlim2 resulted in
significant reduction of engorgement body weight and oviposition of adult ticks
whereas vaccination with GST-hlim3 affected only engorgement body weight. These
data highlight the suitability of the 2 recombinant cement-like proteins for use in a
cocktail vaccine. Based on several studies, anti-tick immunity induced by cocktail

104
vaccine is likely to be more effective than single vaccine antigen. For this reason,
further studies are required to evaluate the remaining novel cement-like antigens and
compare the efficacy of the single and cocktail antigens.
In conclusion, this study reports result from sequence analysis of salivary gland
cDNA library. Several bioactive molecules that possibly play important role in
countering host defense systems were identified. In this report, 2 unprecedented
molecules including metalloproteases and cement-like proteins were selected for further
studied. This is the first report to evaluate the expression profiles upon blood feeding,
expression of the recombinant protein and proenzyme activation mechanism of
metalloproteases from the salivary glands of ticks. Another molecule, cement-like
antigens is one of the interesting vaccine candidates. Vaccination of animal using the 2
recombinant cement-like antigens can affect the tick feeding. The information obtained
from this study would assist in the development of tick control strategy. Further studies
with reverse-genetic approaches will provide further insight into the functions and
significances of these molecules in tick biology and parasitology.

105
Acknowledgements
This research work was carried out at the National Research Center for
Protozoan Diseases (NRCPD), Obihiro University of Agriculture and Veterinary
Medicine (Obihiro Univ. Agric. Vet. Med.). This study was supported by Bio-orientated
Technology Research Advancement Institution (BRAIN) and the 21th century COE
Program (A-1) Ministry of Education, Culture, Science and Technology, Japan.
I would like to acknowledge Dr. Naoyoshi Suzuki, a president of Obihiro Univ.
Agric. Vet. Med. for providing me an opportunity to carry out research in this university.
I would like to express my deep and sincere gratitude to my previous
supervisor, Prof. Kozo Fujisaki (Obihiro Univ. Agric. Vet. Med. and Kagoshima Univ.)
for his intellectual instruction, detailed and constructive comments, invaluable
suggestions, patiently revising this dissertation and tireless supports throughout this
work.
I am deeply grateful to my present supervisor, Prof. Hiroshi Suzuki (Obihiro
Univ. Agric. Vet. Med.) for his critical comments, suggestions and supporting me to
complete this thesis.
I wish to express my warm and sincere thanks to Prof. Makoto Sugiyama (Gifu
University), Prof. Tadashi Itagaki (Iwate University), Prof. Yoshikazu Hirota (Tokyo
University of Agriculture and Technology) and Prof. Shin-ichiro Kawazu (Obihiro Univ.
Agric. Vet. Med.) for constitutive and invaluable suggestions, and patiently revising this
dissertation.
I would like to express my special gratitude to carry out my research under
excellent supervision of Prof. Xuenan Xuan, Assoc. Prof. Yoshifumi Nishikawa at

106
NRCPD, Assoc. Prof. Tomohide Matsuo at Kyorin University School of Medicine and
Dr. Naotoshi Tsuji (Natl. Inst. Anim. Hlt. Q). I also would like to acknowledge Prof.
Ikuo Igarashi, director of NRCPD, Assoc. Prof. Noboru Inoue, Assoc. Prof. Naoaki
Yokoyama, Assoc. Prof. Makoto Igarashi, Prof. Hirotaka Kanuka and Dr. Shinya
Fukumoto for their meaningful and helpful suggestion.
I warmly thank all my colleagues in research unit of Molecular and Vector
Arthropodology in NRCPD for their hospitality, technical and mental supports. I owe
special thanks to Mr. Takeshi Sakaguchi, the previous undergraduate student who
greatly contributed to my work, Dr. Takeharu Miyoshi (Natl. Inst. Anim. Hlt. Q) for his
technical supports and Dr. Namangala Boniface at the University of Zambia for his
critical review for manuscripts preparation.
A special appreciation to all the scientific- and non-scientific staff, as well as
students at NRCPD for their kind co-operation and help in many ways.
Lastly, I owe my loving thanks to my husband, Dr. Noritaka Kuboki, for his
help in Japanese translation, documents preparation and mental supports. Without his
understanding and encouragement, it would have been impossible for me to complete
this work. Special thanks are due to my father, mother and family for their loving
supports.

107
References
1) Alarcon-Chaidez, F. J., Muller-Doblies, U. U. and Wikel, S. (2003).
Characterization of a recombinant immunomodulatory protein from the salivary
glands of Dermacentor andersoni. Parasite Immunol. 25, 69-77.
2) Alarcon-Chaidez, F. J., Sun, J. and Wikel, S. K. (2007). Transcriptome analysis
of the salivary glands of Dermacentor andersoni Stiles (Acari: Ixodidae). Insect
Biochem. Mol. Biol. 37, 48-71.
3) Aljamali, M. N., Bior, A. D., Sauer, J. R. and Essenberg, R. C. (2003). RNA
interference in ticks: a study using histamine binding protein dsRNA in the
female tick Amblyomma americanum. Insect Mol. Biol. 12, 299-305.
4) Allen, J. R. (1973). Tick resistance: basophils in skin reactions of resistant
guinea pigs. Int. J. Parasitol. 3, 195-200.
5) Allen, J. R., Khalil, H. M. and Wikel, S. K. (1979). Langerhans cells trap tick
salivary gland antigens in tick-resistant guinea pigs. J. Immunol. 122, 563-565.
6) Allen, J. R. and Kemp, D. H. (1982). Observations on the behaviour of
Dermacentor andersoni larvae infesting normal and tick resistant guinea-pigs.
Parasitology 84, 195-204.
7) Altschul, S. F., Gish, W., Miller, W., Myers, E. W. and Lipman, D. J. (1990).
Basic local alignment search tool. J. Mol. Biol. 215, 403-410.
8) Baramova, E. N., Shannon, J. D., Bjarnason, J. B. and Fox, J. W. (1989).
Degradation of extracellular matrix proteins by hemorrhagic metalloproteinases.
Arch. Biochem. Biophys. 275, 63-71.

108
9) Barker, S. C. and Murrell, A. (2004). Systematics and evolution of ticks with a
list of valid genus and species names. Parasitology 129, S15-36.
10) Battsetseg, B., Lucero, S., Xuan, X., Claveria, F., Byambaa, B., Battur, B.,
Boldbaatar, D., Batsukh, Z., Khaliunaa, T., Battsetseg, G., Igarashi, I., Nagasawa,
H. and Fujisaki, K. (2002). Detection of equine Babesia spp. gene fragments in
Dermacentor nuttalli Olenev 1929 infesting mongolian horses, and their
amplification in egg and larval progenies. J. Vet. Med. Sci. 64, 727-730.
11) Bell, J. F., Stewart, S. J. and Wikel, S. K. (1979). Resistance to tick-borne
Francisella tularensis by tick-sensitized rabbits: allergic klendusity. Am. J. Trop.
Med. Hyg. 28, 876-880.
12) Bergeron, F., Leduc, R. and Day, R. (2000). Subtilase-like pro-protein
convertases: from molecular specificity to therapeutic applications. J. Mol.
Endocrinol. 24, 1-22.
13) Bergman, D. K., Ramachandra, R. N. and Wikel, S. K. (1998). Characterization
of an immunosuppressant protein from Dermacentor andersoni (Acari:
Ixodidae) salivary glands. J. Med. Entomol. 35, 505-509.
14) Binnington, K. C. and Kemp, D. H. (1980). Role of tick salivary glands in
feeding and disease transmission. Adv. Parasitol. 18, 315-339.
15) Bior, A. D., Essenberg, R. C. and Sauer, J. R. (2002). Comparison of
differentially expressed genes in the salivary glands of male ticks, Amblyomma
americanum and Dermacentor andersoni. Insect Biochem. Mol. Biol. 32, 645-
655.
16) Birkedal-Hansen, H. (1995). Proteolytic remodeling of extracellular matrix. Curr.
Opin. Cell Biol. 7, 728-735.

109
17) Bishop, R., Lambson, B., Wells, C., Pandit, P., Osaso, J., Nkonge, C., Morzaria,
S., Musoke, A. and Nene, V. (2002). A cement protein of the tick Rhipicephalus
appendiculatus, located in the secretory e cell granules of the type III salivary
gland acini, induces strong antibody responses in cattle. Int. J. Parasitol. 32, 833-
842.
18) Bjarnason, J. B., Hamilton, D. and Fox, J. W. (1988). Studies on the mechanism
of hemorrhage production by five proteolytic hemorrhagic toxins from Crotalus
atrox venom. Biol. Chem. Hoppe Seyler 369, 121-129.
19) Bjarnason, J. B. and Fox, J. W. (1994). Hemorrhagic metalloproteinases from
snake venoms. Pharmacol. Ther. 62, 325-372.
20) Bode, W., Gomis-Ruth, F. X. and Stockler, W. (1993). Astacins, serralysins,
snake venom and matrix metalloproteinases exhibit identical zinc-binding
environments (HEXXHXXGXXH and Met-turn) and topologies and should be
grouped into a common family, the 'metzincins'. FEBS Lett. 331, 134-140.
21) Brossard, M. and Wikel, S. K. (2004). Tick immunobiology. Parasitology 129,
S161-176.
22) Daszak, P., Cunningham, A. A. and Hyatt, A. D. (2000). Emerging infectious
diseases of wildlife-threats to biodiversity and human health. Science 287, 443-
449.
23) de Castro, J. J. (1997). Sustainable tick and tickborne disease control in
livestock improvement in developing countries. Vet. Parasitol. 71, 77-97.
24) de la Fuente, J., Almazan, C., Blouin, E. F., Naranjo, V. and Kocan, K. M.
(2005). RNA interference screening in ticks for identification of protective
antigens. Parasitol. Res. 96, 137-141.

110
25) de la Fuente, J., Almazan, C., Naranjo, V., Blouin, E. F., Meyer, J. M. and Kocan,
K. M. (2006). Autocidal control of ticks by silencing of a single gene by RNA
interference. Biochem. Biophys. Res. Commun. 344, 332-338.
26) de la Fuente, J. and Kocan, K. M. (2006). Strategies for development of vaccines
for control of ixodid tick species. Parasite Immunol. 28, 275-283.
27) Dinglasan, R. R., Valenzuela, J. G. and Azad, A. F. (2005). Sugar epitopes as
potential universal disease transmission blocking targets. Insect Biochem. Mol.
Biol. 35, 1-10.
28) Falquet, L., Pagni, M., Bucher, P., Hulo, N., Sigrist, C. J., Hofmann, K. and
Bairoch, A. (2002). The PROSITE database, its status in 2002. Nucleic Acids
Res. 30, 235-238.
29) Fox, J. W. and Serrano, S. M. (2005). Structural considerations of the snake
venom metalloproteinases, key members of the M12 reprolysin family of
metalloproteinases. Toxicon 45, 969-985.
30) Francischetti, I. M., Valenzuela, J. G., Andersen, J. F., Mather, T. N. and Ribeiro,
J. M. (2002). Ixolaris, a novel recombinant tissue factor pathway inhibitor
(TFPI) from the salivary gland of the tick, Ixodes scapularis: identification of
factor X and factor Xa as scaffolds for the inhibition of factor VIIa/tissue factor
complex. Blood 99, 3602-3612.
31) Francischetti, I. M., Valenzuela, J. G., Pham, V. M., Garfield, M. K. and Ribeiro,
J. M. (2002). Toward a catalog for the transcripts and proteins (sialome) from
the salivary gland of the malaria vector Anopheles gambiae. J. Exp. Biol. 205,
2429-2451.
32) Francischetti, I. M., Mather, T. N. and Ribeiro, J. M. (2003). Cloning of a

111
salivary gland metalloprotease and characterization of gelatinase and
fibrin(ogen)lytic activities in the saliva of the Lyme disease tick vector Ixodes
scapularis. Biochem. Biophys. Res. Commun. 305, 869-875.
33) Francischetti, I. M., Mather, T. N. and Ribeiro, J. M. (2005). Tick saliva is a
potent inhibitor of endothelial cell proliferation and angiogenesis. Thromb.
Haemost. 94, 167-174.
34) Francischetti, I. M., My Pham, V., Mans, B. J., Andersen, J. F., Mather, T. N.,
Lane, R. S. and Ribeiro, J. M. (2005). The transcriptome of the salivary glands
of the female western black-legged tick Ixodes pacificus (Acari: Ixodidae).
Insect Biochem. Mol. Biol. 35, 1142-1161.
35) Frisch, J. E. (1999). Towards a permanent solution for controlling cattle ticks.
Int. J. Parasitol. 29, 57-71.
36) Fujisaki, K. (1978). Development of acquired resistance precipitating antibody
in rabbits experimentally infested with females of Haemaphysalis longicornis
(Ixodoidea: Ixodidae). Natl. Inst. Anim. Health Q (Tokyo) 18, 27-38.
37) Fujisaki, K. (1992). A review of the taxonomy of Theileria sergenti/buffeli/
orientalis group parasites in cattle. J. Protozool. Res. 2, 87-96.
38) Fujisaki, K., Kawazu, S. and Kamio, T. (1994). The taxonomy of the bovine
Theileria spp. . Parasitol. Today 10, 31-33.
39) Furie, B. and Furie, B. C. (2005). Thrombus formation in vivo. J Clin Invest 115,
3355-3362.
40) Gasteiger, E., Gattiker, A., Hoogland, C., Ivanyi, I., Appel, R. D. and Bairoch, A.
(2003). ExPASy: The proteomics server for in-depth protein knowledge and
analysis. Nucleic Acids Res. 31, 3784-3788.

112
41) George, J. E., Pound, J. M. and Davey, R. B. (2004). Chemical control of ticks
on cattle and the resistance of these parasites to acaricides. Parasitology 129,
S353-366.
42) Gill, H. S. (1986). Kinetics of mast cell, basophil and eosinophil populations at
Hyalomma anatolicum anatolicum feeding sites on cattle and the acquisition of
resistance. Parasitology 93 ( Pt 2), 305-315.
43) Gillespie, R. D., Mbow, M. L. and Titus, R. G. (2000). The immunomodulatory
factors of bloodfeeding arthropod saliva. Parasite Immunol. 22, 319-331.
44) Goodman, J. T., Dennis, D. T. and Sonenshine, D. E. (2005). Tick-borne diseases
of humans. In:Gulland, P. J., Cranston, P. S., [eds] The Insects: An Outline of
Entomology. pp. 422-424. ASM Press, Washington.
45) Gordon, J. R. and Allen, J. R. (1991). Factors V and VII anticoagulant activities
in the salivary glands of feeding Dermacentor andersoni ticks. J. Parasitol. 77,
167-170.
46) Graf, J. F., Gogolewski, R., Leach-Bing, N., Sabatini, G. A., Molento, M. B.,
Bordin, E. L. and Arantes, G. J. (2004). Tick control: an industry point of view.
Parasitology 129, S427-442.
47) Guy, E. C., Bateman, D. E., Martyn, C. N., Heckels, J. E. and Lawton, N. F.
(1989). Lyme disease: prevalence and clinical importance of Borrelia
burgdorferi specific IgG in forestry workers. Lancet 1, 484-486.
48) Hati, R., Mitra, P., Sarker, S. and Bhattacharyya, K. K. (1999). Snake venom
hemorrhagins. Crit. Rev. Toxicol. 29, 1-19.
49) Hatta, T., Umemiya, R., Liao, M., Gong, H., Harnnoi, T., Tanaka, M., Miyoshi,
T., Boldbaatar, D., Battsetseg, B., Zhou, J., Xuan, X., Tsuji, N., Taylor, D. and

113
Fujisaki, K. (2007). RNA interference of cytosolic leucine aminopeptidase
reduces fecundity in the hard tick, Haemaphysalis longicornis. Parasitol. Res.
100, 847-854.
50) He, H., Chen, A. C., Davey, R. B., Ivie, G. W. and George, J. E. (1999).
Characterization and molecular cloning of a glutathione S-transferase gene from
the tick, Boophilus microplus (Acari: Ixodidae). Insect Biochem. Mol. Biol. 29,
737-743.
51) Higuchi, S., Simomura, S., Yoshida, H., Hoshi, F., Kawamura, S. and Yasuda, Y.
(1991). Development of Babesia gibsoni in the hemolymph of the vector tick,
Haemaphysalis longicornis. J. Vet. Med. Sci. 53, 491-493.
52) Hoogstraal, H., Roberts, F. H., Kohls, G. M. and Tipton, V. J. (1968). Review of
Haemaphysalis (Kaiseriana) longicornis Neumann (resurrected) of Australia,
New Zealand, New Caledonia, Fiji, Japan, Korea, and Northeastern China and
USSR, and its parthenogenetic and bisexual populations (Ixodoidea, Ixodidae). J.
Parasitol. 54, 1197-1213.
53) Hoogstraal, H. (1985). Argasid and nuttalliellid ticks as parasites and vectors.
Adv. Parasitol. 24, 135-238.
54) Horak, I. G., Camicas, J. L. and Keirans, J. E. (2002). The Argasidae, Ixodidae
and Nuttalliellidae (Acari: Ixodida): a world list of valid tick names. Exp. Appl.
Acarol. 28, 27-54.
55) Imamura, S., da Silva Vaz Junior, I., Sugino, M., Ohashi, K. and Onuma, M.
(2005). A serine protease inhibitor (serpin) from Haemaphysalis longicornis as
an anti-tick vaccine. Vaccine 23, 1301-1311.
56) Ishihara, T. (1968). Bovine piroplasmosis in Japan. Jpn. Arg. Res. quart 3, 23-31.

114
57) Jones, L. D., Matthewson, M. and Nuttall, P. A. (1992). Saliva-activated
transmission (SAT) of Thogoto virus: dynamics of SAT factor activity in the
salivary glands of Rhipicephalus appendiculatus, Amblyomma variegatum, and
Boophilus microplus ticks. Exp. Appl. Acarol. 13, 241-248.
58) Jongejan, F. and Uilenberg, G. (2004). The global importance of ticks.
Parasitology 129, S3-14.
59) Kabamoto, S., Bhagat, N. A., Suzuki, H., You, M., Kamio, T., Tsuji, N., Claveria,
F. G., Nagasawa, H., Igarashi, I., Toyoda, Y. and Fujisaki, K. (2002).
Comparison of macrophage scavenger receptor-A knockout mice with wild type
ones in the immune response against repeated infestation with Haemaphysalis
longicornis. J. Vet. Med. Sci. 64, 355-359.
60) Karczewski, J., Waxman, L., Endris, R. G. and Connolly, T. M. (1995). An
inhibitor from the argasid tick Ornithodoros moubata of cell adhesion to
collagen. Biochem. Biophys. Res. Commun. 208, 532-541.
61) Kato, S., Ohtoko, K., Ohtake, H. and Kimura, T. (2005). Vector-capping: a
simple method for preparing a high-quality full-length cDNA library. DNA Res.
12, 53-62.
62) Keller, P. M., Waxman, L., Arnold, B. A., Schultz, L. D., Condra, C. and
Connolly, T. M. (1993). Cloning of the cDNA and expression of moubatin, an
inhibitor of platelet aggregation. J. Biol. Chem. 268, 5450-5456.
63) Kemp, D. H., Agbede, R. I., Johnston, L. A. and Gough, J. M. (1986).
Immunization of cattle against Boophilus microplus using extracts derived from
adult female ticks: feeding and survival of the parasite on vaccinated cattle. Int.
J. Parasitol. 16, 115-120.

115
64) Kemp, D. H., Pearson, R. D., Gough, J. M. and Willadsen, P. (1989).
Vaccination against Boophilus microplus: localization of antigens on tick gut
cells and their interaction with the host immune system. Exp. Appl. Acarol. 7,
43-58.
65) Kemp, D. H., Stone, B. F. and Binnington, K. C. (1982). Tick attachment and
feeding: role of the mouthparts, feeding apparatus, salivary gland secretions and
host response. In:Obenchain, F. D. and Galun R, [eds] Physiology of Ticks. pp.
119-168. Pergamon Press, Oxford.
66) Kitaoka, S. (1961). Physiological and ecological studies on some ticks. VII.
Parthenogenetic and bisexual races of Haemaphysalis longicornis in Japan and
experimental crossing between them. Natl. Inst. Anim. Health Q (Tokyo) 1, 142-
149.
67) Lambson, B., Nene, V., Obura, M., Shah, T., Pandit, P., Ole-Moiyoi, O., Delroux,
K., Welburn, S., Skilton, R., de Villiers, E. and Bishop, R. (2005). Identification
of candidate sialome components expressed in ixodid tick salivary glands using
secretion signal complementation in mammalian cells. Insect Mol. Biol. 14, 403-
414.
68) Lazure, C. (2002). The peptidase zymogen proregions: nature's way of
preventing undesired activation and proteolysis. Curr. Pharm. Des. 8, 511-531.
69) Leboulle, G., Rochez, C., Louahed, J., Ruti, B., Brossard, M., Bollen, A. and
Godfroid, E. (2002). Isolation of Ixodes ricinus salivary gland mRNA encoding
factors induced during blood feeding. Am. J. Trop. Med. Hyg. 66, 225-233.
70) Limo, M. K., Voigt, W. P., Tumbo-Oeri, A. G., Njogu, R. M. and ole-MoiYoi, O.
K. (1991). Purification and characterization of an anticoagulant from the

116
salivary glands of the ixodid tick Rhipicephalus appendiculatus. Exp. Parasitol.
72, 418-429.
71) Mans, B. J. and Neitz, A. W. (2004). Adaptation of ticks to a blood-feeding
environment: evolution from a functional perspective. Insect Biochem. Mol.
Biol. 34, 1-17.
72) Marchler-Bauer, A., Anderson, J. B., DeWeese-Scott, C., Fedorova, N. D., Geer,
L. Y., He, S., Hurwitz, D. I., Jackson, J. D., Jacobs, A. R., Lanczycki, C. J.,
Liebert, C. A., Liu, C., Madej, T., Marchler, G. H., Mazumder, R., Nikolskaya, A.
N., Panchenko, A. R., Rao, B. S., Shoemaker, B. A., Simonyan, V., Song, J. S.,
Thiessen, P. A., Vasudevan, S., Wang, Y., Yamashita, R. A., Yin, J. J. and Bryant,
S. H. (2003). CDD: a curated Entrez database of conserved domain alignments.
Nucleic Acids Res. 31, 383-387.
73) Mejri, N., Rutti, B. and Brossard, M. (2002). Immunosuppressive effects of
Ixodes ricinus tick saliva or salivary gland extracts on innate and acquired
immune response of BALB/c mice. Parasitol. Res. 88, 192-197.
74) Moncrief, J. S., Obiso, R., Jr., Barroso, L. A., Kling, J. J., Wright, R. L., Van
Tassell, R. L., Lyerly, D. M. and Wilkins, T. D. (1995). The enterotoxin of
Bacteroides fragilis is a metalloprotease. Infect. Immun. 63, 175-181.
75) Mulenga, A., Sugimoto, C., Sako, Y., Ohashi, K., Musoke, A., Shubash, M. and
Onuma, M. (1999). Molecular characterization of a Haemaphysalis longicornis
tick salivary gland-associated 29-kilodalton protein and its effect as a vaccine
against tick infestation in rabbits. Infect. Immun. 67, 1652-1658.
76) Mulenga, A., Sugimoto, C. and Onuma, M. (2000). Issues in tick vaccine
development: identification and characterization of potential candidate vaccine

117
antigens. Microbes Infect. 2, 1353-1361.
77) Nakajima, C., da Silva Vaz, I., Jr., Imamura, S., Konnai, S., Ohashi, K. and
Onuma, M. (2005). Random sequencing of cDNA library derived from partially-
fed adult female Haemaphysalis longicornis salivary gland. J. Vet. Med. Sci. 67,
1127-1131.
78) Narasimhan, S., Montgomery, R. R., DePonte, K., Tschudi, C., Marcantonio, N.,
Anderson, J. F., Sauer, J. R., Cappello, M., Kantor, F. S. and Fikrig, E. (2004).
Disruption of Ixodes scapularis anticoagulation by using RNA interference. Proc.
Natl. Acad. Sci. USA 101, 1141-1146.
79) Nazario, S., Das, S., de Silva, A. M., Deponte, K., Marcantonio, N., Anderson, J.
F., Fish, D., Fikrig, E. and Kantor, F. S. (1998). Prevention of Borrelia
burgdorferi transmission in guinea pigs by tick immunity. Am. J. Trop. Med.
Hyg. 58, 780-785.
80) Nene, V., Lee, D., Quackenbush, J., Skilton, R., Mwaura, S., Gardner, M. J. and
Bishop, R. (2002). AvGI, an index of genes transcribed in the salivary glands of
the ixodid tick Amblyomma variegatum. Int. J. Parasitol. 32, 1447-1456.
81) Nene, V., Lee, D., Kang'a, S., Skilton, R., Shah, T., de Villiers, E., Mwaura, S.,
Taylor, D., Quackenbush, J. and Bishop, R. (2004). Genes transcribed in the
salivary glands of female Rhipicephalus appendiculatus ticks infected with
Theileria parva. Insect Biochem. Mol. Biol. 34, 1117-1128.
82) Nielsen, H., Engelbrecht, J., Brunak, S. and von Heijne, G. (1997). A neural
network method for identification of prokaryotic and eukaryotic signal peptides
and prediction of their cleavage sites. Int. J. Neural. Syst. 8, 581-599.
83) Nithiuthai, S. and Allen, J. R. (1985). Langerhans cells present tick antigens to

118
lymph node cells from tick-sensitized guinea-pigs. Immunology 55, 157-163.
84) Nuttall, P. A., Paesen, G. C., Lawrie, C. H. and Wang, H. (2000). Vector-host
interactions in disease transmission. J. Mol. Microbiol. Biotechnol. 2, 381-386.
85) Pearson, W. R. and Lipman, D. J. (1988). Improved tools for biological
sequence comparison. Proc. Natl. Acad. Sci. USA 85, 2444-2448.
86) Primakoff, P. and Myles, D. G. (2000). The ADAM gene family: surface proteins
with adhesion and protease activity. Trends Genet. 16, 83-87.
87) Ramos, O. H., Carmona, A. K. and Selistre-de-Araujo, H. S. (2003). Expression,
refolding, and in vitro activation of a recombinant snake venom pro-
metalloprotease. Protein Expr. Purif. 28, 34-41.
88) Ramos, O. H. and Selistre-de-Araujo, H. S. (2006). Snake venom
metalloproteases-structure and function of catalytic and disintegrin domains.
Comp. Biochem. Physiol. C Toxicol. Pharmacol. 142, 328-346.
89) Randolph, S. E. (2001). The shifting landscape of tick-borne zoonoses: tick-
borne encephalitis and Lyme borreliosis in Europe. Philos. Trans. R. Soc. Lond.
B Biol. Sci. 356, 1045-1056.
90) Rawlings, N. D. and Barrett, A. J. (1993). Evolutionary families of peptidases.
Biochem. J. 290 ( Pt 1), 205-218.
91) Ribeiro, J. M., Makoul, G. T., Levine, J., Robinson, D. R. and Spielman, A.
(1985). Antihemostatic, antiinflammatory, and immunosuppressive properties of
the saliva of a tick, Ixodes dammini. J. Exp. Med. 161, 332-344.
92) Ribeiro, J. M. (1987). Role of saliva in blood-feeding by arthropods. Annu. Rev.
Entomol. 32, 463-478.
93) Ribeiro, J. M. (1989). Role of saliva in tick/host interactions. Exp. Appl. Acarol.

119
7, 15-20.
94) Ribeiro, J. M. (1995). How ticks make a living. Parasitol Today 11, 91-93.
95) Ribeiro, J. M. and Francischetti, I. M. (2003). Role of arthropod saliva in blood
feeding: sialome and post-sialome perspectives. Annu. Rev. Entomol. 48, 73-88.
96) Ribeiro, J. M., Andersen, J., Silva-Neto, M. A., Pham, V. M., Garfield, M. K.
and Valenzuela, J. G. (2004). Exploring the sialome of the blood-sucking bug
Rhodnius prolixus. Insect Biochem. Mol. Biol. 34, 61-79.
97) Ribeiro, J. M., Alarcon-Chaidez, F., Francischetti, I. M., Mans, B. J., Mather, T.
N., Valenzuela, J. G. and Wikel, S. K. (2006). An annotated catalog of salivary
gland transcripts from Ixodes scapularis ticks. Insect Biochem. Mol. Biol. 36,
111-129.
98) Riding, G. A., Jarmey, J., McKenna, R. V., Pearson, R., Cobon, G. S. and
Willadsen, P. (1994). A protective "concealed" antigen from Boophilus
microplus. Purification, localization, and possible function. J. Immunol. 153,
5158-5166.
99) Rodriguez Bautista, J. L., Ikadai, H., You, M., Battsetseg, B., Igarashi, I.,
Nagasawa, H. and Fujisaki, K. (2001). Molecular evidence of Babesia caballi
(Nuttall and Strickland, 1910) parasite transmission from experimentally-
infected SCID mice to the ixodid tick, Haemaphysalis longicornis (Neuman,
1901). Vet. Parasitol. 102, 185-191.
100) Sanders, M. L., Jaworski, D. C., Sanchez, J. L., DeFraites, R. F., Glass, G. E.,
Scott, A. L., Raha, S., Ritchie, B. C., Needham, G. R. and Schwartz, B. S. (1998).
Antibody to a cDNA-derived calreticulin protein from Amblyomma americanum
as a biomarker of tick exposure in humans. Am. J. Trop. Med. Hyg. 59, 279-285.

120
101) Santos, I. K., Valenzuela, J. G., Ribeiro, J. M., de Castro, M., Costa, J. N., Costa,
A. M., da Silva, E. R., Neto, O. B., Rocha, C., Daffre, S., Ferreira, B. R., da
Silva, J. S., Szabo, M. P. and Bechara, G. H. (2004). Gene discovery in
Boophilus microplus, the cattle tick: the transcriptomes of ovaries, salivary
glands, and hemocytes. Ann. N. Y. Acad. Sci. 1026, 242-246.
102) Sauer, J. R., Essenberg, R. C. and Bowman, A. S. (2000). Salivary glands in
ixodid ticks: control and mechanism of secretion. J. Insect Physiol. 46, 1069-
1078.
103) Schoeler, G. B. and Wikel, S. K. (2001). Modulation of host immunity by
haematophagous arthropods. Ann. Trop. Med. Parasitol. 95, 755-771.
104) Seals, D. F. and Courtneidge, S. A. (2003). The ADAMs family of
metalloproteases: multidomain proteins with multiple functions. Genes Dev. 17,
7-30.
105) Shannon, J. D., Baramova, E. N., Bjarnason, J. B. and Fox, J. W. (1989). Amino
acid sequence of a Crotalus atrox venom metalloproteinase which cleaves type
IV collagen and gelatin. J. Biol. Chem. 264, 11575-11583.
106) Shapiro, S. Z., Voigt, W. P. and Fujisaki, K. (1986). Tick antigens recognized by
serum from a guinea pig resistant to infestation with the tick Rhipicephalus
appendiculatus. J. Parasitol. 72, 454-463.
107) Shapiro, S. Z., Voigt, W. P. and Ellis, J. A. (1989). Acquired resistance to ixodid
ticks induced by tick cement antigen. Exp. Appl. Acarol. 7, 33-41.
108) Shimokawa, K., Jia, L. G., Wang, X. M. and Fox, J. W. (1996). Expression,
activation, and processing of the recombinant snake venom metalloproteinase,
pro-atrolysin E. Arch. Biochem. Biophys. 335, 283-294.

121
109) Sonenshine, D. E. (1991). Biology of Ticks, Vol 1. Oxford University
Press\ New York.
110) Telford, S. R., 3rd and Goethert, H. K. (2004). Emerging tick-borne infections:
rediscovered and better characterized, or truly 'new' ? Parasitology 129, S301-
327.
111) Tellam, R., Smith, D., Kemp, D. H. and Willadsen, P. (1992). Vaccination
against ticks, In:Wong, W.K. [eds], Animal Parasite Control Utilising
Biotechnology. pp. 303-331. CRC press Inc., Roca Raton.
112) Trimnell, A. R., Hails, R. S. and Nuttall, P. A. (2002). Dual action ectoparasite
vaccine targeting 'exposed' and 'concealed' antigens. Vaccine 20, 3560-3568.
113) Trimnell, A. R., Davies, G. M., Lissina, O., Hails, R. S. and Nuttall, P. A. (2005).
A cross-reactive tick cement antigen is a candidate broad-spectrum tick vaccine.
Vaccine 23, 4329-4341.
114) Trummal, K., Tonismagi, K., Siigur, E., Aaspollu, A., Lopp, A., Sillat, T., Saat,
R., Kasak, L., Tammiste, I., Kogerman, P., Kalkkinen, N. and Siigur, J. (2005). A
novel metalloprotease from Vipera lebetina venom induces human endothelial
cell apoptosis. Toxicon 46, 46-61.
115) Tsuda, A., Mulenga, A., Sugimoto, C., Nakajima, M., Ohashi, K. and Onuma, M.
(2001). cDNA cloning, characterization and vaccine effect analysis of
Haemaphysalis longicornis tick saliva proteins. Vaccine 19, 4287-4296.
116) Valenzuela, J. G., Charlab, R., Mather, T. N. and Ribeiro, J. M. (2000).
Purification, cloning, and expression of a novel salivary anticomplement protein
from the tick, Ixodes scapularis. J. Biol. Chem. 275, 18717-18723.
117) Valenzuela, J. G. (2002). Exploring the messages of the salivary glands of Ixodes

122
ricinus. Am. J. Trop. Med. Hyg. 66, 223-224.
118) Valenzuela, J. G., Francischetti, I. M., Pham, V. M., Garfield, M. K., Mather, T.
N. and Ribeiro, J. M. (2002). Exploring the sialome of the tick Ixodes scapularis.
J. Exp. Biol. 205, 2843-2864.
119) Valenzuela, J. G., Pham, V. M., Garfield, M. K., Francischetti, I. M. and Ribeiro,
J. M. (2002). Toward a description of the sialome of the adult female mosquito
Aedes aegypti. Insect Biochem. Mol. Biol. 32, 1101-1122.
120) Valenzuela, J. G., Francischetti, I. M., Pham, V. M., Garfield, M. K. and Ribeiro,
J. M. (2003). Exploring the salivary gland transcriptome and proteome of the
Anopheles stephensi mosquito. Insect Biochem. Mol. Biol. 33, 717-732.
121) Valenzuela, J. G. (2004). Exploring tick saliva: from biochemistry to 'sialomes'
and functional genomics. Parasitology 129, S83-94.
122) Van Klinken, B. J., Dekker, J., Buller, H. A. and Einerhand, A. W. (1995). Mucin
gene structure and expression: protection vs. adhesion. Am. J. Physiol. 269,
G613-627.
123) Walker, A. R. and Fletcher, J. D. (1986). Histological study of the attachment
sites of adult Rhipicephalus appendiculatus on rabbits and cattle. Int. J. Parasitol.
16, 399-413.
124) Walker, D. H. (1998). Tick-transmitted infectious diseases in the United States.
Annu. Rev. Public Health 19, 237-269.
125) Walker, D.H. (2005). Ehrlichia under our noses and no one notices. Arch. Virol.,
147-156.
126) Wang, X., Coons, L. B., Taylor, D. B., Stevens, S. E., Jr. and Gartner, T. K.
(1996). Variabilin, a novel RGD-containing antagonist of glycoprotein IIb-IIIa

123
and platelet aggregation inhibitor from the hard tick Dermacentor variabilis. J.
Biol. Chem. 271, 17785-17790.
127) White, J. M. (2003). ADAMs: modulators of cell-cell and cell-matrix
interactions. Curr. Opin. Cell Biol. 15, 598-606.
128) Wikel, S. K. (1996). Host immunity to ticks. Annu. Rev. Entomol. 41, 1-22.
129) Wikel, S. K. (1996). Tick modulation of host cytokines. Exp. Parasitol. 84, 304-
309.
130) Wikel, S. K., Ramachandra, R. N., Bergman, D. K., Burkot, T. R. and Piesman, J.
(1997). Infestation with pathogen-free nymphs of the tick Ixodes scapularis
induces host resistance to transmission of Borrelia burgdorferi by ticks. Infect.
Immun. 65, 335-338.
131) Wikel, S. K. (1999). Tick modulation of host immunity: an important factor in
pathogen transmission. Int. J. Parasitol. 29, 851-859.
132) Willadsen, P. (1980). Immunity to ticks. Adv Parasitol 18, 293-311.
133) Willadsen, P. (1987). Immunological approaches to the control of ticks. Int. J.
Parasitol. 17, 671-677.
134) Willadsen, P. and Kemp, D. H. (1988). Vaccination with 'concealed' antigens for
tick control. Parasitol. Today 4, 196-198.
135) Willadsen, P., Bird, P., Cobon, G. S. and Hungerford, J. (1995).
Commercialisation of a recombinant vaccine against Boophilus microplus.
Parasitology 110, S43-50.
136) Willadsen, P., Smith, D., Cobon, G. and McKenna, R. V. (1996). Comparative
vaccination of cattle against Boophilus microplus with recombinant antigen
Bm86 alone or in combination with recombinant Bm91. Parasite Immunol. 18,

124
241-246.
137) Willadsen, P. (1997). Vaccines, genetics and chemicals in tick control: the
Australian experience. Trop. Anim. Health Prod. 29, 91S-94S.
138) Willadsen, P. and Jongejan, F. (1999). Immunology of the tick-host interaction
and the control of ticks and tick-borne diseases. Parasitol. Today 15, 258-262.
139) Willadsen, P. (2001). The molecular revolution in the development of vaccines
against ectoparasites. Vet. Parasitol. 101, 353-368.
140) Willadsen, P. (2004). Anti-tick vaccines. Parasitology 129, S367-387.
141) Wolfsberg, T. G., Straight, P. D., Gerena, R. L., Huovila, A. P., Primakoff, P.,
Myles, D. G. and White, J. M. (1995). ADAM, a widely distributed and
developmentally regulated gene family encoding membrane proteins with a
disintegrin and metalloprotease domain. Dev. Biol. 169, 378-383.
142) Xu, G., Fang, Q. Q., Keirans, J. E. and Durden, L. A. (2004). Cloning and
sequencing of putative calreticulin complementary DNAs from four hard tick
species. J. Parasitol. 90, 73-78.
143) Xu, Y., Bruno, J. F. and Luft, B. J. (2005). Identification of novel tick salivary
gland proteins for vaccine development. Biochem. Biophys. Res. Commun. 326,
901-904.
144) You, M., Xuan, X., Tsuji, N., Kamio, T., Igarashi, I., Nagasawa, H., Mikami, T.
and Fujisaki, K. (2001). Molecular characterization of a troponin I-like protein
from the hard tick Haemaphysalis longicornis. Insect Biochem. Mol. Biol. 32,
67-73.
145) You, W. K., Seo, H. J., Chung, K. H. and Kim, D. S. (2003). A novel
metalloprotease from Gloydius halys venom induces endothelial cell apoptosis

125
through its protease and disintegrin-like domains. J. Biochem. (Tokyo) 134, 739-
749.
146) Zhou, J., Gong, H., Zhou, Y., Xuan, X. and Fujisaki, K. (2006). Identification of
a glycine-rich protein from the tick Rhipicephalus haemaphysaloides and
evaluation of its vaccine potential against tick feeding. Parasitol. Res. 100, 77-
84.
147) Zhou, J., Liao, M., Hatta, T., Tanaka, M., Xuan, X. and Fujisaki, K. (2006).
Identification of a follistatin-related protein from the tick Haemaphysalis
longicornis and its effect on tick oviposition. Gene. 372, 191-198.

126

127
Related Documents











