Regular Article PHAGOCYTES, GRANULOCYTES, AND MYELOPOIESIS Hypoxia-inducible factor 2a regulates key neutrophil functions in humans, mice, and zebrafish A. A. Roger Thompson, 1 Philip M. Elks, 1,2 Helen M. Marriott, 1 Suttida Eamsamarng, 1,2 Kathryn R. Higgins, 1 Amy Lewis, 1 Lynne Williams, 1 Selina Parmar, 1 Gary Shaw, 1 Emmet E. McGrath, 1 Federico Formenti, 3 Fredericus J. Van Eeden, 2 Vuokko L. Kinnula, 4 Christopher W. Pugh, 5 Ian Sabroe, 1 David H. Dockrell, 1 Edwin R. Chilvers, 6 Peter A. Robbins, 3 Melanie J. Percy, 7 M. Celeste Simon, 8 Randall S. Johnson, 9 Stephen A. Renshaw, 1,2 Moira K. B. Whyte, 1,2 and Sarah R. Walmsley 1 1 Department of Infection and Immunity and 2 Medical Research Council Centre for Developmental and Biomedical Genetics, University of Sheffield, Sheffield, United Kingdom; 3 Department of Physiology, Anatomy and Genetics, University of Oxford, Oxford, United Kingdom; 4 Pulmonary Division, Department of Medicine, University of Helsinki and Helsinki University Central Hospital, Helsinki, Finland; 5 Nuffield Department of Clinical Medicine, Henry Wellcome Building for Molecular Physiology, University of Oxford, Oxford, United Kingdom; 6 Department of Medicine, University of Cambridge, Cambridge, United Kingdom; 7 Department of Haematology, Belfast City Hospital, Belfast, Northern Ireland, United Kingdom; 8 Department of Cell and Developmental Biology, University of Pennsylvania, Philadelphia, PA; and 9 Department of Physiology, Development and Neuroscience, University of Cambridge, Cambridge, United Kingdom Key Points • Neutrophil lifespan is extended in patients with gain-of-function HIF2A mutations. • HIF-2a regulates in vivo neutrophil longevity and thus tissue inflammation and repair. Neutrophil lifespan and function are regulated by hypoxia via components of the hypoxia inducible factor (HIF)/von Hippel Lindau/hydroxylase pathway, including specific roles for HIF-1a and prolyl hydroxylase-3. HIF-2a has both distinct and overlapping biological roles with HIF-1a and has not previously been studied in the context of neutrophil biology. We investigated the role of HIF-2a in regulating key neutrophil functions. Human and murine peripheral blood neutrophils expressed HIF-2a, with expression up- regulated by acute and chronic inflammatory stimuli and in disease-associated inflam- matory neutrophil. HIF2A gain-of-function mutations resulted in a reduction in neutrophil apoptosis both ex vivo, through the study of patient cells, and in vivo in a zebrafish tail injury model. In contrast, HIF-2a–deficient murine inflammatory neutrophils displayed increased sensitivity to nitrosative stress induced apoptosis ex vivo and increased neutrophil apoptosis in vivo, resulting in a reduction in neutrophilic inflammation and reduced tissue injury. Expression of HIF-2a was temporally dissociated from HIF-1a in vivo and predominated in the resolution phase of inflammation. These data support a critical and selective role for HIF-2a in persistence of neutrophilic inflammation and provide a platform to dissect the therapeutic utility of targeting HIF-2a in chronic inflammatory diseases. (Blood. 2014;123(3):366-376) Introduction Neutrophils are key mediators of tissue injury in acute and chronic inflammatory diseases. 1,2 Timely neutrophil apoptosis, with effective macrophage efferocytosis, ensures resolution of inflammation and protects against the cytotoxic effects of neutrophils. 3-5 As such, targeting neutrophil apoptosis represents an attractive therapeutic strategy. Neutrophil apoptosis is significantly inhibited by physiological hypoxia. 6-8 Adaptation to hypoxia is mediated by the heterodimeric nuclear transcription factor, hypoxia inducible factor (HIF), comprised of 1 of 3 known a subunits and a constitutively expressed b subunit. 9 In normoxia, HIF is inactive as a subunits are labeled for degradation by a family of 3 oxygen-sensitive prolyl hydroxylases (PHDs), and HIF transcription may be inactivated by factor inhibiting HIF. 10-12 Recent evidence reveals the importance of oxygen-sensing path- ways in innate immune biology. HIF-a subunits accumulate in myeloid cells in hypoxic conditions, as in other cell types, but also in response to bacteria and bacterial products irrespective of the ambient oxygen tension. These data demonstrate roles for the HIF pathway beyond the regulation of hypoxic signaling and implicate HIF in host responses to bacteria. 13-15 Indeed, myeloid-specific deficiency of HIF-1a not only abolishes the prolonged survival of neutrophils in hypoxia but also results in depletion of intracellular ATP levels and impairment of neutrophil granule protease pro- duction, macrophage motility and invasion, and bacterial killing. 6,13,16 These in vitro findings translated into reduced inflammatory cell infiltrates in murine models of inflammation and infection. 13,16 With such profound effects on innate immune cell function, HIF-1a itself is not an attractive therapeutic target for the many inflammatory diseases, eg, chronic obstructive pulmonary disease (COPD) and in- flammatory bowel disease, where inflammation and bacteria frequently coexist. In marked contrast to HIF-1a deficiency, we found that de- ficiency of a HIF hydroxylase, PHD3, had minimal consequences for Submitted May 1, 2013; accepted October 27, 2013. Prepublished online as Blood First Edition paper, November 6, 2013; DOI 10.1182/blood-2013-05- 500207. M.K.B.W. and S.R.W. contributed equally to the work. The online version of this article contains a data supplement. There is an Inside Blood commentary on this article in this issue. The publication costs of this article were defrayed in part by page charge payment. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734. © 2014 by The American Society of Hematology 366 BLOOD, 16 JANUARY 2014 x VOLUME 123, NUMBER 3

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Regular Article
PHAGOCYTES, GRANULOCYTES, AND MYELOPOIESIS
Hypoxia-inducible factor 2a regulates key neutrophil functions inhumans, mice, and zebrafishA. A. Roger Thompson,1 Philip M. Elks,1,2 Helen M. Marriott,1 Suttida Eamsamarng,1,2 Kathryn R. Higgins,1 Amy Lewis,1
Lynne Williams,1 Selina Parmar,1 Gary Shaw,1 Emmet E. McGrath,1 Federico Formenti,3 Fredericus J. Van Eeden,2
Vuokko L. Kinnula,4 Christopher W. Pugh,5 Ian Sabroe,1 David H. Dockrell,1 Edwin R. Chilvers,6 Peter A. Robbins,3
Melanie J. Percy,7 M. Celeste Simon,8 Randall S. Johnson,9 Stephen A. Renshaw,1,2 Moira K. B. Whyte,1,2 and
Sarah R. Walmsley1
1Department of Infection and Immunity and 2Medical Research Council Centre for Developmental and Biomedical Genetics, University of Sheffield,
Sheffield, United Kingdom; 3Department of Physiology, Anatomy and Genetics, University of Oxford, Oxford, United Kingdom; 4Pulmonary Division,
Department of Medicine, University of Helsinki and Helsinki University Central Hospital, Helsinki, Finland; 5Nuffield Department of Clinical Medicine, Henry
Wellcome Building for Molecular Physiology, University of Oxford, Oxford, United Kingdom; 6Department of Medicine, University of Cambridge, Cambridge,
United Kingdom; 7Department of Haematology, Belfast City Hospital, Belfast, Northern Ireland, United Kingdom; 8Department of Cell and Developmental
Biology, University of Pennsylvania, Philadelphia, PA; and 9Department of Physiology, Development and Neuroscience, University of Cambridge,
Cambridge, United Kingdom
Key Points
• Neutrophil lifespan is extendedin patients with gain-of-functionHIF2A mutations.
• HIF-2a regulates in vivoneutrophil longevity and thustissue inflammation and repair.
Neutrophil lifespan and function are regulated by hypoxia via components of the hypoxia
inducible factor (HIF)/von Hippel Lindau/hydroxylase pathway, including specific roles
for HIF-1a and prolyl hydroxylase-3. HIF-2a has both distinct and overlapping biological
roles with HIF-1a and has not previously been studied in the context of neutrophil
biology.We investigated the role of HIF-2a in regulating key neutrophil functions. Human
and murine peripheral blood neutrophils expressed HIF-2a, with expression up-
regulated by acute and chronic inflammatory stimuli and in disease-associated inflam-
matory neutrophil.HIF2Again-of-functionmutations resulted in a reduction in neutrophil
apoptosis both ex vivo, through the study of patient cells, and in vivo in a zebrafish tail
injury model. In contrast, HIF-2a–deficient murine inflammatory neutrophils displayed increased sensitivity to nitrosative stress
induced apoptosis ex vivo and increased neutrophil apoptosis in vivo, resulting in a reduction in neutrophilic inflammation and reduced
tissue injury. Expression of HIF-2a was temporally dissociated from HIF-1a in vivo and predominated in the resolution phase of
inflammation. These data support a critical and selective role for HIF-2a in persistence of neutrophilic inflammation and provide
a platform to dissect the therapeutic utility of targeting HIF-2a in chronic inflammatory diseases. (Blood. 2014;123(3):366-376)
Introduction
Neutrophils are key mediators of tissue injury in acute and chronicinflammatory diseases.1,2 Timely neutrophil apoptosis, with effectivemacrophage efferocytosis, ensures resolution of inflammation andprotects against the cytotoxic effects of neutrophils.3-5 As such,targeting neutrophil apoptosis represents an attractive therapeuticstrategy.
Neutrophil apoptosis is significantly inhibited by physiologicalhypoxia.6-8 Adaptation to hypoxia is mediated by the heterodimericnuclear transcription factor, hypoxia inducible factor (HIF), comprisedof 1 of 3 knowna subunits and a constitutively expressedb subunit.9 Innormoxia, HIF is inactive as a subunits are labeled for degradation bya family of 3 oxygen-sensitive prolyl hydroxylases (PHDs), and HIFtranscription may be inactivated by factor inhibiting HIF.10-12
Recent evidence reveals the importance of oxygen-sensing path-ways in innate immune biology. HIF-a subunits accumulate inmyeloid cells in hypoxic conditions, as in other cell types, but also
in response to bacteria and bacterial products irrespective of theambient oxygen tension. These data demonstrate roles for the HIFpathway beyond the regulation of hypoxic signaling and implicateHIF in host responses to bacteria.13-15 Indeed, myeloid-specificdeficiency of HIF-1a not only abolishes the prolonged survival ofneutrophils in hypoxia but also results in depletion of intracellularATP levels and impairment of neutrophil granule protease pro-duction, macrophage motility and invasion, and bacterial killing.6,13,16
These in vitro findings translated into reduced inflammatory cellinfiltrates in murine models of inflammation and infection.13,16 Withsuch profound effects on innate immune cell function, HIF-1a itselfis not an attractive therapeutic target for the many inflammatorydiseases, eg, chronic obstructive pulmonary disease (COPD) and in-flammatory bowel disease, where inflammation and bacteria frequentlycoexist. In marked contrast to HIF-1a deficiency, we found that de-ficiency of a HIF hydroxylase, PHD3, had minimal consequences for
Submitted May 1, 2013; accepted October 27, 2013. Prepublished online as
Blood First Edition paper, November 6, 2013; DOI 10.1182/blood-2013-05-
500207.
M.K.B.W. and S.R.W. contributed equally to the work.
The online version of this article contains a data supplement.
There is an Inside Blood commentary on this article in this issue.
The publication costs of this article were defrayed in part by page charge
payment. Therefore, and solely to indicate this fact, this article is hereby
marked “advertisement” in accordance with 18 USC section 1734.
© 2014 by The American Society of Hematology
366 BLOOD, 16 JANUARY 2014 x VOLUME 123, NUMBER 3

the functional status of neutrophils prior to their apoptosis.17 Specifictargeting of individual components of the HIF hydroxylase pathway,independent of HIF-1a itself, could therefore result in selectiveregulation of neutrophil survival pathways independent of key host-pathogen responses.
Distinct biological roles for HIF-1a and HIF-2a have recentlyemerged, with HIF-2a regulating a distinct but overlapping set oftarget genes to HIF-1a and, importantly, playing a less significantrole in regulating glycolytic enzyme expression.18,19 Furthermore,differential transcriptional activation of HIF-1a and HIF-2a canresult in coordinated cellular responses, dependent on the relativeabundance of each isoform, with HIF-1a and HIF-2a havingopposing effects on macrophage nitric oxide formation.20 HIF-2ahas been implicated in regulation of other macrophage functions,with myeloid-specific HIF-2a–deficient mice having reducedmacrophage-mediated inflammatory responses to endotoxemia andreduced tumor-associated macrophage infiltration associated withreduced tumor cell proliferation and progression.14 Given this evi-dence of differential functions for HIF-1a andHIF-2a in myeloid cellsand the known dominance of HIF-1a over HIF-2a in the regulation ofglycolysis and ATP generation, we hypothesized that HIF-2a defi-ciency may have a more selective immunomodulatory phenotype thanHIF-1a deficiency in neutrophils.
Materials and methods
Ethical approval
All participants gave written informed consent in accordance with the Dec-laration of Helsinki principles. South Sheffield Research Ethics Committeeapproved the study of healthy volunteers and individuals with inflammatoryarthritis, Oxfordshire Clinical Research Ethics Committee approved the study ofpatients with gain-of-function mutations in HIF2A, and Helsinki UniversityHospital, Helsinki, Finland, approved the use of human lung tissue.
Murine colonies
Lysozyme M-driven cre recombinase (LysMcre) was used to target Hif1a(Hif1aflox/flox;LysMcre1/2) or Hif2a (Hif2aflox/flox;LysMcre1/2) deletions inmyeloid lineage cells. Animals were back-crossed to a C57BL/6 back-ground.16,20 C57BL/6 mice or littermate LysMcre2/2
floxed mice were used ascontrols. All animal experiments were conducted in accordance with the HomeOffice Animals (Scientific Procedures) Act of 1986 with local ethics approval.
Isolation and culture of neutrophils from humans and mice
Human peripheral blood neutrophils were isolated from whole blood usingdextran sedimentation and discontinuous plasma-Percoll gradients.21 Ultra-purified human neutrophils were obtained using negative magnetic selectionas previously described.22 Murine peripheral blood neutrophils and bonemarrow–derived neutrophils were isolated by negative magnetic selection(Easysep; STEMCELL Technologies, Grenoble, France), and inflammatoryneutrophils were recovered from bronchoalveolar lavage (BAL) fluid 24hours following challenge with nebulized lipopolysaccharide (LPS). Cellculture is detailed in the supplemental Methods on the Blood Web site.
RNA isolation and relative quantification
Human neutrophils (103 106/condition) were lysed with 1 mL TRI Reagent(Sigma-Aldrich Ltd., Gillingham, United Kingdom), and RNAwas extractedusing chloroform phase partitioning and isopropanol. Murine peripheralblood, inflammatory (BAL), or bonemarrow neutrophils (13 106/condition)were lysed, and RNA was extracted using the mirVana total RNA isolationprotocol (Ambion, Austin, TX). Samples were treated with DNase (Ambion) andrandomhexamer cDNA synthesized by reverse transcription. Assays-on-demandgene expression TaqMan MGB 6FAM dye-labeled products (Applied
Biosystems) were used for relative quantification of cDNA and BigDyev3.1 sequencing kits for product verification (supplemental Methods).
Immunoblot detection of human and murine neutrophil protein
Whole cell human (hypotonic) and murine (sodium dodecyl sulfate [SDS])lysates were prepared as previously described.17 Immunoblotting was perfor-med with polyclonal anti-mouse HIF-1a (R&D), monoclonal anti-human HIF-1a (clone 54, BD,) or anti–HIF-2a (clone ep190b; Novus Biologicals) primaryantibodies. Sample loading was confirmed by p38 mitogen-activated proteinkinase (MAPK) expression (Cell Signaling Technology). All bands shownwere at the predicted molecular weight for the protein of interest.
Immunohistochemistry
Lung tissue sections were from nonsmokers and patients with mild (TheGlobal Initiative for Chronic Obstructive Lung Disease stage 2) andmoderate (The Global Initiative for Chronic Obstructive Lung Diseasestage 3/4) COPD undergoing resection for suspected lung tumor or lungtransplantation from the Department of Medicine and Pathology, HelsinkiUniversity Hospital. Slides were stained with anti–HIF-2a (clone ep190b;Novus Biologicals) or isotype control, developed using the ImmPRESSuniversal polymer detection kit (Vector Laboratories Ltd, Peterborough,United Kingdom) and visualized with diaminobenzidine.
Neutrophil functional assays
Phagocytosis. Uptake of Alexa Fluor 488 Escherichia coli (K-12 strain)BioParticles (Invitrogen, Paisley, United Kingdom) after 30 minutes of co-culture (multiplicity of infection of 1:1) was determined by flow cytometry(FACSCalibur; Becton Dickinson). Alternatively, neutrophils were in-cubated with opsonized zymosan (0.2-1 mg/mL) for 30 minutes, and thephagocytic index was calculated using microscopy.
Respiratory burst. Cells were cultured with 6 mM 29,79-dichlorofluor-escin diacetate (Sigma-Aldrich) for 30minutes, and then stimulated for a further30 minutes with N-formyl-Met-Leu-Phe (100 nM) or opsonized zymosan(0.2 mg/mL) before FL1 fluorescence was determined by flow cytometry andgeometric mean fluorescence calculated using FlowJo software (Tree Star Inc.).
Fish husbandry
The neutrophil specific fluorescent zebrafish line Tg(mpx:GFP)i114 wasused, subsequently referred to as mpx:GFP for simplicity.23 Zebrafish weremaintained according to standard protocols.24 Adult fish were maintainedon a 14-hour light and 10-hour dark cycle at 28°C in United KingdomHome Office–approved facilities in the Medical Research Council Centre forDevelopmental and Biomedical Genetics aquaria at the University of Sheffield.
Fish assays
Inflammatory responses were elicited in zebrafish larvae by tail transection aspreviously described, using the neutrophil specific line,mpx:GFP.23,25 Ratesof apoptosis were assessed using terminal deoxynucleotidyltransferase-mediated dUTP nick end labeling/tyramide signal amplification, by blindedassessors, and by anti-active caspase-3/tyramide signal amplification staining,as previously described.25 Collagen formation following tailfin injury wasassessed at 24, 48, or 72 hours postinjury (hpi). Wild-type zebrafish were fixedand stained with an anti-collagen1 primary antibody and Alexa Fluor488–conjugated secondary antibody. Fluorescence intensity was analyzed byVolocity 5 (Improvision; Perkin Elmer).
Wild-type and mutant hif2a cloning
Zebrafish 2dpf RNA purified using TRIzol (Invitrogen) was used for reversetranscriptase-polymerase chain reaction (PCR) cloning of HIF2A homologs,hif2aa (epas1a) and hif2ab (epas1b) (primer details in supplemental Table 1),using Pfusion polymerase (Finnzymes, Espoo, Finland). These were initiallycloned into the TOPOBlunt vector (Invitrogen) and subsequently subclonedinto the pCS21 vector (Invitrogen) for RNA synthesis. Dominant activeforms of hif2aa and hif2ab were generated by successive rounds of site-directed mutagenesis as previously described.25 The zebrafish amino acid
BLOOD, 16 JANUARY 2014 x VOLUME 123, NUMBER 3 HIF2A AND NEUTROPHIL FUNCTION 367

corresponding to human HIF-2a G537R and G537W was mutated in thesame fashion. RNA encoding dominant active isoforms were transcribed(mMessageMachine; Ambion, Life Technologies) and microinjected intozebrafish embryos at the 1-cell stage.25 To assess the function ofoverexpressed G487 mutant constructs, phd3 in situ hybridization wasperformed 24 hours following injection of RNA for G487R, G487W ordominant active hif2aa at the 1-cell stage.26 A dominant-negative form ofhif2aa was generated using primers amplifying DNA corresponding toamino acids 1-330 of human HIF-2a.25,27
Murine LPS acute lung injury model
Nebulized LPS (3 mg) was administered to awake mice. At specified timepoints, BAL sampling was performed and then analyzed for total(hemocytometer), differential, and apoptosis (cytospin) cell counts. BALsupernatant and plasma were stored for cytokine analysis. For histologicalsections, lungs were fixed with 10% formalin at 20 cm H2O and paraffin-embedded blocks were prepared. Following deparaffinization, serial sectionswere stained with anti–HIF-2a (clone ep190b).
Bone marrow transplantation
C57BL/6 recipient mice were irradiated with 3 fractions of 1 Gy each dayfor 4 days before injection with 1.5 3 106 bone marrow cells from wild-type Hif2aflox/flox;LysMcre2/2 or knockout Hif2aflox/flox;LysMcre1/2 mice.28
Acute lung injury experiments were performed 5 weeks following injection ofdonor marrow, and reconstitution was confirmed by genotyping of cellsrecovered from BAL (DNeasy; Qiagen, Crawley, United Kingdom).
Statistical analysis
Data were analyzed using Prism 5.0 software (GraphPad Software, SanDiego, CA) using unpaired, 2-tailed t-tests for comparisons between 2groups and 1-way or 2-way analysis of variance (ANOVA; with Bonferonnipost-test adjustment) for other data as appropriate. Significance was acceptedwhenP, .05. Data are expressed asmean6 standard error of themean (SEM).
Results
Human neutrophils express HIF-2a
We isolated human peripheral blood neutrophils and demonstratedHIF2AmRNA expression by nonquantitative PCR in highly purifiedneutrophil populations (Figure 1A), with sequencing of PCR products(data not shown). Real-time PCR assays showedHIF2A expression isnot altered over time in normoxic or hypoxic culture (Figure 1B) orfollowing stimulation by heat-killed Staphylococcus aureus or pep-tidoglycan (supplemental Figure 1). In contrast to HIF-1a, HIF-2aprotein was detected in freshly isolated neutrophils and neutrophilscultured in normoxia, with further induction of HIF-2a protein andexpression ofHIF-1a protein in hypoxia. HIF-2awas up-regulatedwith iron chelators, hydroxylase inhibitors, heat-killed bacteria,or LPS (Figure 1C-F).
Overexpression of HIF-2a delays neutrophil apoptosis but does
not affect function
Study of idiopathic cases of erythrocytosis with raised serum eryth-ropoietin has resulted in the identification of a rare group ofindividuals with gain-of-function mutations in the HIF2A gene.29
We obtained peripheral blood neutrophils from some of theseindividuals to determine the consequences of HIF-2a overexpres-sion for neutrophil survival and function. Subjects with gain-of-function HIF2A mutations had lower rates of neutrophil apoptosiscompared with controls but a preserved response to hydroxylaseinhibition by the pan-hydroxylase inhibitor dimethyloxalylglycine(DMOG) (Figure 2A). Neutrophils from these subjects also showedenhanced expression of the HIF2A target genes PAI1 and PHD3(Figure 2B), which were equivalent to changes previously described
Figure 1. Human neutrophils express HIF-2a, and
expression is up-regulated by hypoxia, hydroxy-
lase inhibition, and heat-killed bacteria. (A) Expres-
sion of HIF2A in freshly isolated neutrophils (t0) or cells
cultured for 4 hours in normoxia (N) or hypoxia (H) was
determined by PCR and agarose gel electrophoresis. A
representative gel image of n 5 3 is shown. (B) Fold
change in expression of HIF2A and HIF1A following
culture of human neutrophils in normoxia (filled bars) or
hypoxia (open bars) for 6 or 12 hours. TaqMan anal-
ysis of cDNA was performed with data normalized to
ACTB expression. Data show mean and SEM of fold
change with respect to normoxic samples at 6 hours,
n5 3, analyzed by ANOVA. (C-D) Expression of HIF-1a
and HIF-2a is differentially regulated by hydroxylase
inhibitors and up-regulated in response to heat-killed
bacteria. Neutrophils were cultured in normoxia (Nx) with
dimethyloxalylglycine (DMOG; 100 mM); deferoxamine
(DFO; 300 mM); heat-killed Streptococcus pneumo-
niae (Spn; multiplicity of infection 10:1); LPS (100 ng/mL);
or hypoxia (Hx) before being lysed. Proteins were sepa-
rated using SDS-polyacrylamide gel electrophore-
sis (PAGE), and blots were probed for (C) HIF-1a
and (D) HIF-2a. p38 MAPK was used as a loading
control. (E-F) Densitometry analysis was performed
on (E) HIF-1a and (F) HIF-2a blots using ImageJ
software and normalized to p38 MAPK expression.
Data are mean and SEM for minute n 5 4.
368 THOMPSON et al BLOOD, 16 JANUARY 2014 x VOLUME 123, NUMBER 3

following hypoxic culture.17 Functional assays showed phagocyto-sis (Figure 2C-D) and respiratory burst (Figure 2E) were equivalentbetween control and HIF2A mutant neutrophils, indicating HIF-2aoverexpression resulted in a selective prosurvival phenotype withoutalteration of key neutrophil functions.
Overexpression of hif2aa in zebrafish results in delayed
resolution of inflammation
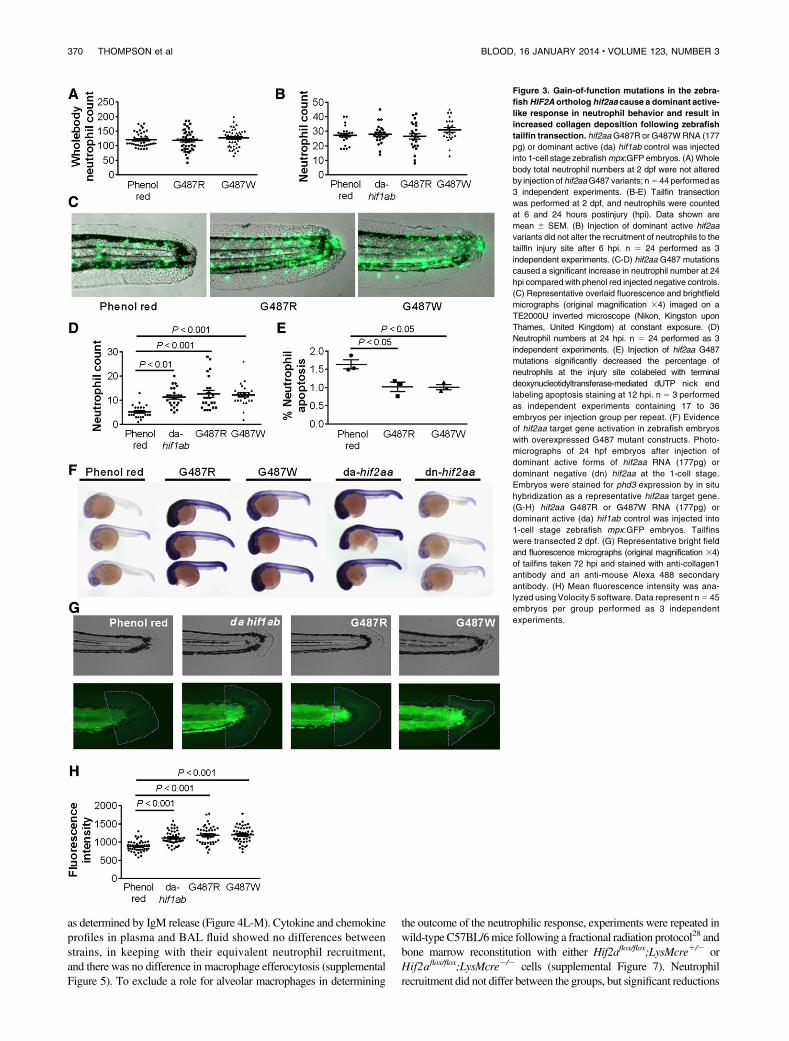
To explore the significance of the reduced neutrophil apoptosis seenin humans with gain-of-function mutations in HIF2A, we mutatedthe zebrafish ortholog, hif2aa, to produce a protein with an aminoacid substitution at the glycine site corresponding to the mutanthuman protein. We found that replacing the glycine with eitherarginine (G487R) or tryptophan (G487W), to replicate the HIF2Amutations observed in the patients, did not affect whole fishneutrophil numbers (Figure 3A). In a well-characterized tail injurymodel of neutrophilic inflammation,23 neutrophil recruitment did notdiffer between wild-type and mutants (Figure 3B), but the hif2aa-overexpressing fish showed delayed resolution of inflammation(Figure 3C-D). The magnitude of neutrophil persistence was equiv-alent to that seen with overexpression of hif1ab (Figure 3D) orpreviously reported with caspase inhibition.23 The increased neu-trophil numbers at the site of injury after 24 hours, when inflammationhas normally resolved,23 were associated with a significant reductionin neutrophil apoptosis (Figure 3E). Similar results were obtainedwhen we mutated the PHD target sites into nonhydroxylatable aminoacids to create a dominant active form of hif2aa (supplementalFigure 2). Importantly, all 3 hif2aa mutants displayed evidence ofup-regulation of recognizedHIF2A/hif2aa target genes (Figure 3F anddata not shown). Injection of a morpholino antisense oligonucleotideto achieve knockdown of the hif2aa binding partner, arnt,30 restoredresolution of inflammation in hif2aa overexpressing fish, confirmingthat the phenotype of delayed inflammation resolution required intacthif2aa signaling (supplemental Figure 2). To study the effects of in-creased hif2aa expression on wound healing responses, collagendepositionwas quantified in the tail transectionmodel, comparingwild
type with hif2aa (G487R and G487W) overexpressing and dominantactive hif1ab-expressing fish. Both hif2aa and hif1ab overexpressingfish had increased collagen deposition at the injury site 72 hpi(Figure 3G-H) after resolution of the acute neutrophilic response(supplemental Figure 2).
Loss of HIF-2a reduces neutrophilic inflammation and lung
injury during inflammation resolution
We isolated peripheral blood neutrophils from wild-type mice andconfirmed the presence of Hif2a by PCR (Figure 4A). In keepingwith previous data, Hif2a was not detected in bone marrow–derivedwild-type neutrophils.14 Consistent with our findings in humanneutrophils, and in contrast to HIF-1a (Figure 4B), murine neu-trophils showed basal expression of HIF-2a protein followingnormoxic culture, with expression also seen in hypoxia and withLPS stimulation or DMOG treatment (Figure 4C). To investigatethe consequences of HIF-2a deficiency, we used mice with myeloid-specific targeted deletion of Hif2a (Hif2aflox/flox;LysMcre1/2).20
HIF-2a–deficient neutrophils showed preserved rates of apoptosis innormoxia and, in contrast to HIF-1a–deficient neutrophils,17 adelay of apoptosis in hypoxia that was equivalent to wild-typecells (supplemental Figure 3). Functional assays on HIF-2a–deficientneutrophils revealed no deficits in respiratory burst, chemotaxis, orphagocytosis, and these cells also showed preserved changes inreceptor expression in response to stimulation with LPS (sup-plemental Figure 4).
A neutrophil-mediated LPS-induced model of acute lung injurywas used to determine the consequences of HIF-2a deficiency invivo. Mice with myeloid-specific deletion of Hif2a had very fewneutrophils in unstimulated airways (supplemental Figure 5) anddisplayed normal recruitment of neutrophils to the lung (Figure 4D,H),but thereafter, had significantly lower BAL total cell counts andneutrophil counts than controls at 48 (Figure 4E,I) and 72 hours(Figure 4F,J), and neutrophil clearance by 120 hours (Figure 4K).Importantly, the lower neutrophil counts were accompanied bya significant reduction in lung injury in the HIF-2a–deficient mice,
Figure 2. Neutrophils isolated from patients with gain-of-function HIF2A mutations have enhanced survival but normal function. (A) Apoptosis. Neutrophils from
patients with HIF2A mutations (closed squares) or healthy controls (open squares) were cultured for 20 hours with dimethyloxalylglycine (0-100 mM) and apoptosis was
determined by morphology. Data are mean 6 SEM for n 5 3, analyzed by 2-way ANOVA with Bonferroni’s multiple comparison post-test. Overall, there was a significant
difference between the control group and the patient group (P , .001), with multiple comparison testing showing statistical significance at the 1 mM concentration (*P , .05).
(B) Expression of the HIF targets VEGF, PAI1, and PHD3. TaqMan quantitative PCR analysis of cDNA prepared from freshly isolated neutrophils of patients with HIF2A
mutations (filled bars) or healthy controls (open bars). Data are mean and SEM for n 5 3. (C-D) Phagocytosis. (C) Phagocytic index was calculated from cytospin slides of
neutrophils from healthy controls (open bars) and HIF2A patients (filled bars) prepared after 30 minutes of culture with opsonized zymosan (0.2-1 mg/mL). (D) Flow cytometry
analysis of intracellular Alexa Fluor 488 E coli was performed after 30 minutes of culture of cells from healthy controls (open bars) and HIF2A patients (filled bars). (E)
Respiratory burst. Neutrophils from healthy controls (open bars) and HIF2A patients (filled bars) were cultured in the presence of dichlorofluorescin only or dichlorofluorescin
and N-formyl-Met-Leu-Phe (100 nM) or zymosan A (0.2 mg/ml) and analyzed by flow cytometry. Data show mean and SEM for n 5 3.
BLOOD, 16 JANUARY 2014 x VOLUME 123, NUMBER 3 HIF2A AND NEUTROPHIL FUNCTION 369
as determined by IgM release (Figure 4L-M). Cytokine and chemokineprofiles in plasma and BAL fluid showed no differences betweenstrains, in keeping with their equivalent neutrophil recruitment,and there was no difference in macrophage efferocytosis (supplementalFigure 5). To exclude a role for alveolar macrophages in determining
the outcome of the neutrophilic response, experiments were repeated inwild-type C57BL/6mice following a fractional radiation protocol28 andbone marrow reconstitution with either Hif2aflox/flox;LysMcre1/2 orHif2aflox/flox;LysMcre2/2 cells (supplemental Figure 7). Neutrophilrecruitment did not differ between the groups, but significant reductions
Figure 3. Gain-of-function mutations in the zebra-
fishHIF2A ortholog hif2aa cause a dominant active-
like response in neutrophil behavior and result in
increased collagen deposition following zebrafish
tailfin transection. hif2aaG487R or G487WRNA (177
pg) or dominant active (da) hif1ab control was injected
into 1-cell stage zebrafishmpx:GFP embryos. (A)Whole
body total neutrophil numbers at 2 dpf were not altered
by injection of hif2aaG487 variants; n5 44 performedas
3 independent experiments. (B-E) Tailfin transection
was performed at 2 dpf, and neutrophils were counted
at 6 and 24 hours postinjury (hpi). Data shown are
mean 6 SEM. (B) Injection of dominant active hif2aa
variants did not alter the recruitment of neutrophils to the
tailfin injury site after 6 hpi. n 5 24 performed as 3
independent experiments. (C-D) hif2aa G487 mutations
caused a significant increase in neutrophil number at 24
hpi compared with phenol red injected negative controls.
(C) Representative overlaid fluorescence and brightfield
micrographs (original magnification 34) imaged on a
TE2000U inverted microscope (Nikon, Kingston upon
Thames, United Kingdom) at constant exposure. (D)
Neutrophil numbers at 24 hpi. n 5 24 performed as 3
independent experiments. (E) Injection of hif2aa G487
mutations significantly decreased the percentage of
neutrophils at the injury site colabeled with terminal
deoxynucleotidyltransferase-mediated dUTP nick end
labeling apoptosis staining at 12 hpi. n 5 3 performed
as independent experiments containing 17 to 36
embryos per injection group per repeat. (F) Evidence
of hif2aa target gene activation in zebrafish embryos
with overexpressed G487 mutant constructs. Photo-
micrographs of 24 hpf embryos after injection of
dominant active forms of hif2aa RNA (177pg) or
dominant negative (dn) hif2aa at the 1-cell stage.
Embryos were stained for phd3 expression by in situ
hybridization as a representative hif2aa target gene.
(G-H) hif2aa G487R or G487W RNA (177pg) or
dominant active (da) hif1ab control was injected into
1-cell stage zebrafish mpx:GFP embryos. Tailfins
were transected 2 dpf. (G) Representative bright field
and fluorescence micrographs (original magnification 34)
of tailfins taken 72 hpi and stained with anti-collagen1
antibody and an anti-mouse Alexa 488 secondary
antibody. (H) Mean fluorescence intensity was ana-
lyzed using Volocity 5 software. Data represent n5 45
embryos per group performed as 3 independent
experiments.
370 THOMPSON et al BLOOD, 16 JANUARY 2014 x VOLUME 123, NUMBER 3
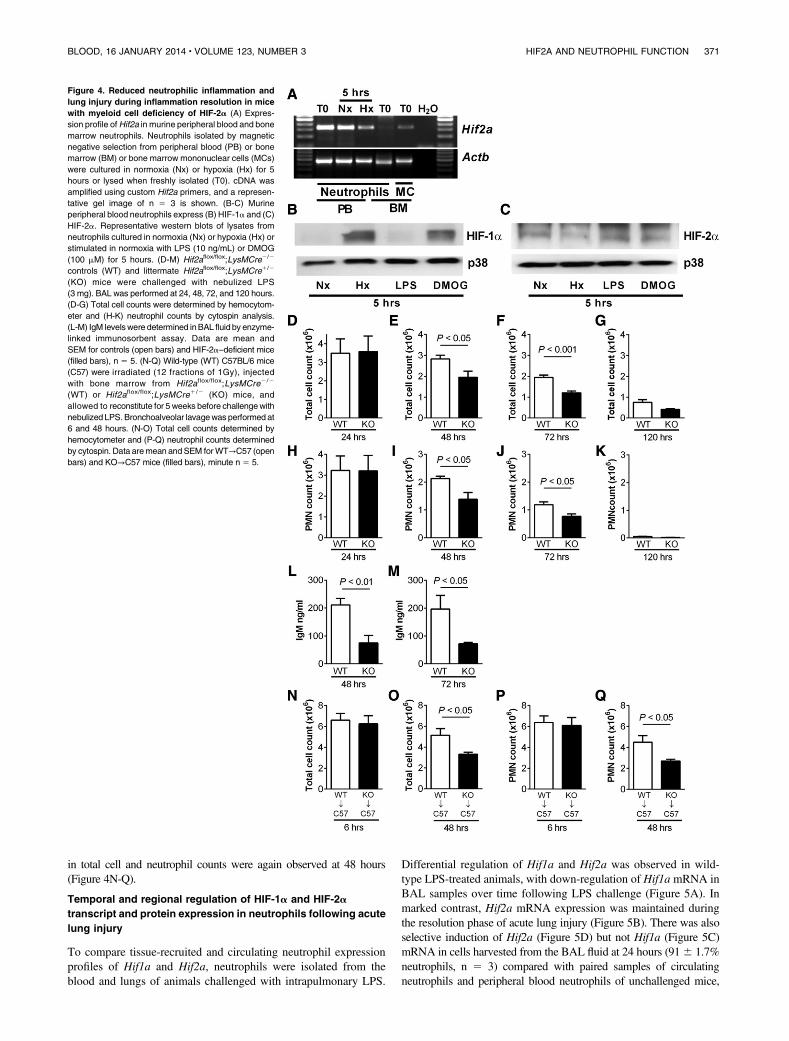
in total cell and neutrophil counts were again observed at 48 hours(Figure 4N-Q).
Temporal and regional regulation of HIF-1a and HIF-2a
transcript and protein expression in neutrophils following acute
lung injury
To compare tissue-recruited and circulating neutrophil expressionprofiles of Hif1a and Hif2a, neutrophils were isolated from theblood and lungs of animals challenged with intrapulmonary LPS.
Differential regulation of Hif1a and Hif2a was observed in wild-type LPS-treated animals, with down-regulation of Hif1a mRNA inBAL samples over time following LPS challenge (Figure 5A). Inmarked contrast, Hif2a mRNA expression was maintained duringthe resolution phase of acute lung injury (Figure 5B). There was alsoselective induction of Hif2a (Figure 5D) but not Hif1a (Figure 5C)mRNA in cells harvested from the BAL fluid at 24 hours (916 1.7%neutrophils, n 5 3) compared with paired samples of circulatingneutrophils and peripheral blood neutrophils of unchallenged mice,
Figure 4. Reduced neutrophilic inflammation and
lung injury during inflammation resolution in mice
with myeloid cell deficiency of HIF-2a (A) Expres-
sion profile ofHif2a in murine peripheral blood and bone
marrow neutrophils. Neutrophils isolated by magnetic
negative selection from peripheral blood (PB) or bone
marrow (BM) or bone marrow mononuclear cells (MCs)
were cultured in normoxia (Nx) or hypoxia (Hx) for 5
hours or lysed when freshly isolated (T0). cDNA was
amplified using custom Hif2a primers, and a represen-
tative gel image of n 5 3 is shown. (B-C) Murine
peripheral blood neutrophils express (B) HIF-1a and (C)
HIF-2a. Representative western blots of lysates from
neutrophils cultured in normoxia (Nx) or hypoxia (Hx) or
stimulated in normoxia with LPS (10 ng/mL) or DMOG
(100 mM) for 5 hours. (D-M) Hif2aflox/flox;LysMCre2/2
controls (WT) and littermate Hif2aflox/flox;LysMCre1/2
(KO) mice were challenged with nebulized LPS
(3 mg). BAL was performed at 24, 48, 72, and 120 hours.
(D-G) Total cell counts were determined by hemocytom-
eter and (H-K) neutrophil counts by cytospin analysis.
(L-M) IgM levelswere determined inBAL fluid by enzyme-
linked immunosorbent assay. Data are mean and
SEM for controls (open bars) and HIF-2a–deficient mice
(filled bars), n 5 5. (N-Q) Wild-type (WT) C57BL/6 mice
(C57) were irradiated (12 fractions of 1Gy), injected
with bone marrow from Hif2aflox/flox;LysMCre2 /2
(WT) or Hif2aflox/flox;LysMCre1/2 (KO) mice, and
allowed to reconstitute for 5 weeks before challengewith
nebulized LPS.Bronchoalveolar lavagewas performed at
6 and 48 hours. (N-O) Total cell counts determined by
hemocytometer and (P-Q) neutrophil counts determined
by cytospin. Data aremean andSEM forWT→C57 (open
bars) and KO→C57 mice (filled bars), minute n 5 5.
BLOOD, 16 JANUARY 2014 x VOLUME 123, NUMBER 3 HIF2A AND NEUTROPHIL FUNCTION 371
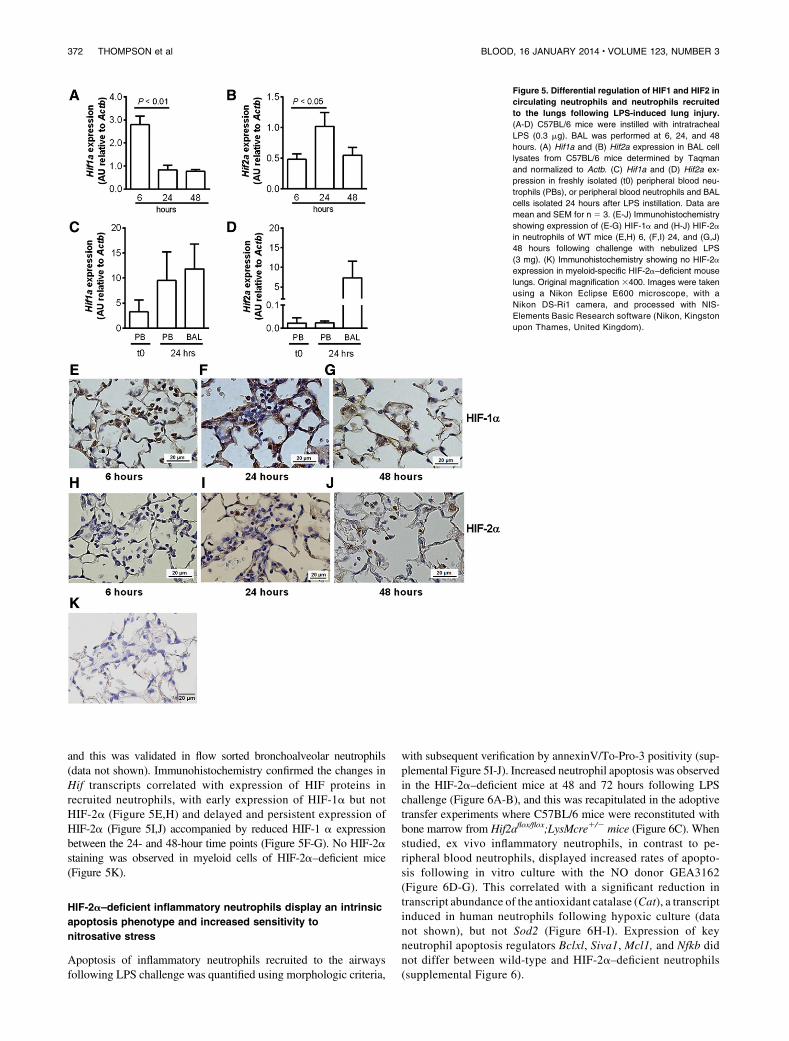
and this was validated in flow sorted bronchoalveolar neutrophils(data not shown). Immunohistochemistry confirmed the changes inHif transcripts correlated with expression of HIF proteins inrecruited neutrophils, with early expression of HIF-1a but notHIF-2a (Figure 5E,H) and delayed and persistent expression ofHIF-2a (Figure 5I,J) accompanied by reduced HIF-1 a expressionbetween the 24- and 48-hour time points (Figure 5F-G). No HIF-2astaining was observed in myeloid cells of HIF-2a–deficient mice(Figure 5K).
HIF-2a–deficient inflammatory neutrophils display an intrinsic
apoptosis phenotype and increased sensitivity to
nitrosative stress
Apoptosis of inflammatory neutrophils recruited to the airwaysfollowing LPS challenge was quantified using morphologic criteria,
with subsequent verification by annexinV/To-Pro-3 positivity (sup-plemental Figure 5I-J). Increased neutrophil apoptosis was observedin the HIF-2a–deficient mice at 48 and 72 hours following LPSchallenge (Figure 6A-B), and this was recapitulated in the adoptivetransfer experiments where C57BL/6 mice were reconstituted withbone marrow from Hif2aflox/flox;LysMcre1/2 mice (Figure 6C). Whenstudied, ex vivo inflammatory neutrophils, in contrast to pe-ripheral blood neutrophils, displayed increased rates of apopto-sis following in vitro culture with the NO donor GEA3162(Figure 6D-G). This correlated with a significant reduction intranscript abundance of the antioxidant catalase (Cat), a transcriptinduced in human neutrophils following hypoxic culture (datanot shown), but not Sod2 (Figure 6H-I). Expression of keyneutrophil apoptosis regulators Bclxl, Siva1, Mcl1, and Nfkb didnot differ between wild-type and HIF-2a–deficient neutrophils(supplemental Figure 6).
Figure 5. Differential regulation of HIF1 and HIF2 in
circulating neutrophils and neutrophils recruited
to the lungs following LPS-induced lung injury.
(A-D) C57BL/6 mice were instilled with intratracheal
LPS (0.3 mg). BAL was performed at 6, 24, and 48
hours. (A) Hif1a and (B) Hif2a expression in BAL cell
lysates from C57BL/6 mice determined by Taqman
and normalized to Actb. (C) Hif1a and (D) Hif2a ex-
pression in freshly isolated (t0) peripheral blood neu-
trophils (PBs), or peripheral blood neutrophils and BAL
cells isolated 24 hours after LPS instillation. Data are
mean and SEM for n 5 3. (E-J) Immunohistochemistry
showing expression of (E-G) HIF-1a and (H-J) HIF-2a
in neutrophils of WT mice (E,H) 6, (F,I) 24, and (G,J)
48 hours following challenge with nebulized LPS
(3 mg). (K) Immunohistochemistry showing no HIF-2a
expression in myeloid-specific HIF-2a–deficient mouse
lungs. Original magnification 3400. Images were taken
using a Nikon Eclipse E600 microscope, with a
Nikon DS-Ri1 camera, and processed with NIS-
Elements Basic Research software (Nikon, Kingston
upon Thames, United Kingdom).
372 THOMPSON et al BLOOD, 16 JANUARY 2014 x VOLUME 123, NUMBER 3
Neutrophils from patients with chronic inflammatory disease
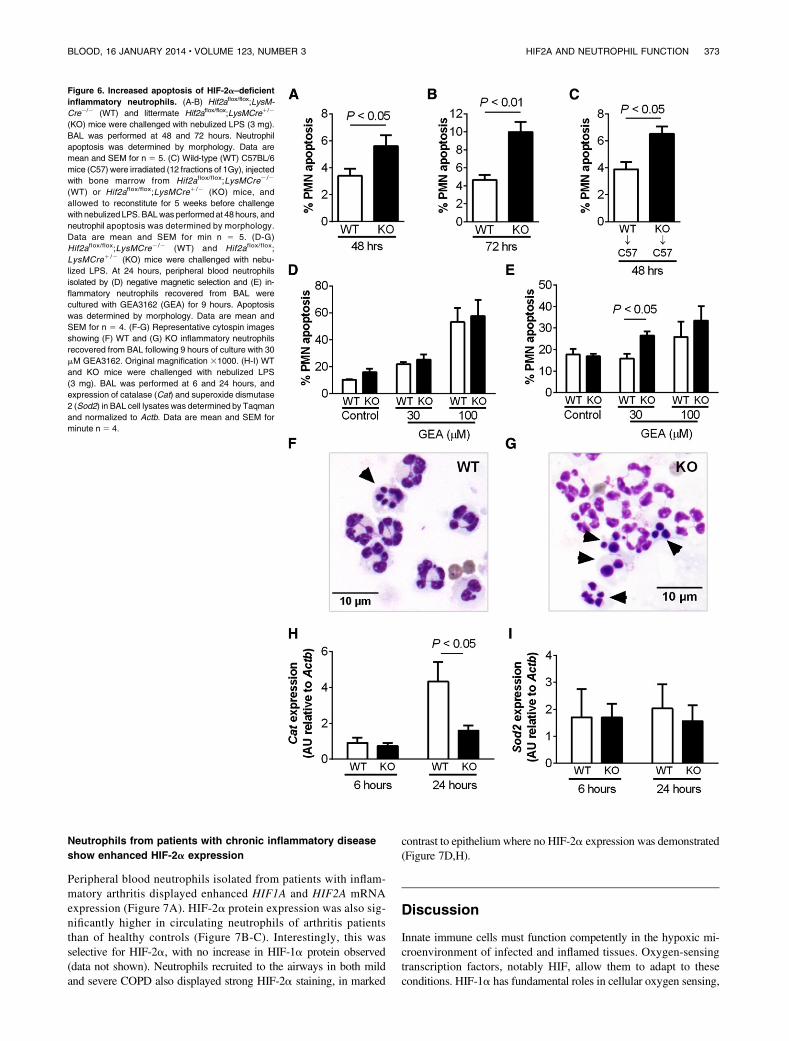
show enhanced HIF-2a expression
Peripheral blood neutrophils isolated from patients with inflam-matory arthritis displayed enhanced HIF1A and HIF2A mRNAexpression (Figure 7A). HIF-2a protein expression was also sig-nificantly higher in circulating neutrophils of arthritis patientsthan of healthy controls (Figure 7B-C). Interestingly, this wasselective for HIF-2a, with no increase in HIF-1a protein observed(data not shown). Neutrophils recruited to the airways in both mildand severe COPD also displayed strong HIF-2a staining, in marked
contrast to epithelium where no HIF-2a expression was demonstrated(Figure 7D,H).
Discussion
Innate immune cells must function competently in the hypoxic mi-croenvironment of infected and inflamed tissues. Oxygen-sensingtranscription factors, notably HIF, allow them to adapt to theseconditions. HIF-1a has fundamental roles in cellular oxygen sensing,
Figure 6. Increased apoptosis of HIF-2a–deficient
inflammatory neutrophils. (A-B) Hif2aflox/flox;LysM-
Cre2/2 (WT) and littermate Hif2aflox/flox;LysMCre1/2
(KO) mice were challenged with nebulized LPS (3 mg).
BAL was performed at 48 and 72 hours. Neutrophil
apoptosis was determined by morphology. Data are
mean and SEM for n 5 5. (C) Wild-type (WT) C57BL/6
mice (C57) were irradiated (12 fractions of 1Gy), injected
with bone marrow from Hif2aflox/flox;LysMCre2 /2
(WT) or Hif2aflox/flox;LysMCre1/2 (KO) mice, and
allowed to reconstitute for 5 weeks before challenge
with nebulized LPS.BALwas performed at 48 hours, and
neutrophil apoptosis was determined by morphology.
Data are mean and SEM for min n 5 5. (D-G)
Hif2aflox/flox;LysMCre2/2 (WT) and Hif2aflox/flox;
LysMCre1 /2 (KO) mice were challenged with nebu-
lized LPS. At 24 hours, peripheral blood neutrophils
isolated by (D) negative magnetic selection and (E) in-
flammatory neutrophils recovered from BAL were
cultured with GEA3162 (GEA) for 9 hours. Apoptosis
was determined by morphology. Data are mean and
SEM for n 5 4. (F-G) Representative cytospin images
showing (F) WT and (G) KO inflammatory neutrophils
recovered from BAL following 9 hours of culture with 30
mM GEA3162. Original magnification 31000. (H-I) WT
and KO mice were challenged with nebulized LPS
(3 mg). BAL was performed at 6 and 24 hours, and
expression of catalase (Cat) and superoxide dismutase
2 (Sod2) in BAL cell lysates was determined by Taqman
and normalized to Actb. Data are mean and SEM for
minute n 5 4.
BLOOD, 16 JANUARY 2014 x VOLUME 123, NUMBER 3 HIF2A AND NEUTROPHIL FUNCTION 373

and metabolic adaptation to hypoxia in many tissues is essential forneutrophil survival in hypoxia and regulates myeloid cell bacterialkilling.6,13,16 Myeloid-specific HIF-1a deficiency reduces inflam-mation,but increases susceptibility to bacterial infections bothlocally and systemically.13,16 This infection risk limits the utilityof direct targeting of HIF-1a as an anti-inflammatory approach,hence our search for more selective regulators of neutrophil functionand fate. With recent evidence suggesting a role for HIF-2a inmyeloid inflammatory responses,14 and a number of unique functionsalready ascribed to both HIF-1a and HIF-2a, we proposed HIF-2awould have a more selective role in neutrophil biology and the reg-ulation of inflammation resolution.
In keeping with previous findings from bone marrow–derivedneutrophils and murine neutrophil cell lines,14 we were unable todetect HIF-2a mRNA or protein in immature murine bone marrowneutrophils. Althoughe we were able to detect low levels of HIF2AmRNA in highly pure peripheral blood neutrophils, higher levels ofHIF-2 mRNA were seen following neutrophil activation in thecontext either of a systemic inflammatory condition (arthritis) orrecruitment to an injured site (LPS-mediated acute lung injury).
Transcriptional regulation of HIF mRNA expression has beendescribed by Takeda et al,20 who showed HIF1A and HIF2A mRNAare differentially expressed in M1- and M2-polarized macrophages,with labile HIF1A mRNA displaying a relatively short half-life andconversely HIF2A mRNA being much more stable with a lower rateof turnover. Of note, these changes in mRNA expression occurredindependent of oxygen availability and were themselves a strongpredictor of protein abundance. In keeping with this, we also observedmuch higher levels of HIF-2a protein in freshly isolated neutrophilsfrom patients with inflammatory arthritis compared with healthycontrol neutrophils and in neutrophils recruited to the lung both inpatients with mild and severe COPD and in a murine acute lung injurymodel.
The importance of HIF-2a accumulation with respect to diseaseoutcomes is unknown but, through access to 3 patients with knowngain-of-function mutations in theHIF2A gene, we were able to assessthe functional consequences of HIF-2a overexpression in neutrophils.Baseline rates of constitutive apoptosis were consistently lower inpatient neutrophils than controls, although further reductions in apo-ptosis were achieved by stimulation with DMOG. This is consistent
Figure 7. Expression of HIF-2a is up-regulated in
neutrophils from patients with active inflammatory
arthritis and is seen in neutrophils within lung
biopsies from patients with COPD. (A) Expression of
HIF1A and HIF2A in inflammatory arthritis patients
(filled bars) and controls (open bars) was determined
by TaqMan analysis of cDNA from freshly isolated
peripheral blood neutrophils with data normalized to
ACTB expression. Data are mean and SEM for n 5 4.
(B-C) HIF-2a protein expression is significantly higher
in neutrophils from patients with inflammatory arthritis
(IA) than healthy volunteers (HV). Freshly isolated
neutrophils were lysed and proteins separated by SDS-
PAGE. Blots were probed for HIF-2a, and densitom-
etry data were normalized to p38 MAPK. (B) Data are
mean and SEM for n 5 3. (C) HIF-2a blots of cir-
culating neutrophil lysates from 3 healthy volunteers
(HV) and 3 patients with inflammatory arthritis (IA).
(D-H) Immunohistochemistry showing HIF-2a expres-
sion in lung biopsies from a (E) nonsmoker and pa-
tients with (F) mild or (G-H) severe COPD. Images are
representative of n 5 2. D is a section stained with an
isotype control. Original magnification: (D-G) 3200;
(H) 31000.
374 THOMPSON et al BLOOD, 16 JANUARY 2014 x VOLUME 123, NUMBER 3

with experimental evidence showing only partial activation ofHIF-2ain these patients, so that hydroxylase inhibition could further stabilizeHIF-2a and stabilize HIF-1a.29 Neutrophils derived from patientswith gain-of-function mutations in HIF2A also showed increasedHIF-2a target gene expression (VEGF, PAI1, and PHD3) to a levelequivalent to that previously described for hypoxic culture.17
Delayed neutrophil apoptosis has been implicated in persistenceof inflammation in animal models,5,31 but whether the increase inHIF-2a expression and associated intrinsic delay of neutrophil apo-ptosis might predispose these patients to inflammatory disease is notknown, not least because of the rarity of the condition and thedominant clinical phenotype of erythrocytosis and its consequences.To directly address the importance of selective HIF-2a stabilizationfor regulation of neutrophil survival and resolution of inflammation,we replicated the human gain-of-function HIF2A mutation in thegenetically tractable zebrafish.23,25 The hydroxylation sites of HIF-asubunits are highly conserved across species,25 with an overall aminoacid homology of 58% between zebrafish hif2aa and human HIF-2a.Using mutated zebrafish RNA, we expressed hif2aa mutated at thesame glycine residues as are altered in the patients with gain-of-function HIF2A mutations. Expression of mutant hif2aa led to im-paired resolution of inflammation in a tail injury model that wasequivalent to overexpression of dominant active hif1ab and of asimilar magnitude to previous results using the pan-caspase inhibitor,zVD.fmk.23 These findings highlight that in vivo hif2aa can modulateneutrophil survival to the same order of magnitude as hif1ab. Impor-tantly, the impairment in inflammation resolution also has consequen-ces for wound healing responses, because overexpression of hif2aaresults in localized increases in collagen deposition 72 hpi.
To address the therapeutic potential of selectively targeting HIF-2a, we investigated the consequences of HIF-2a deficiency usingmice with myeloid-specific deletion of Hif2a, as previouslydescribed.20 In marked contrast to HIF-1a deficiency,16 we foundno impairment of neutrophil chemotaxis, phagocytosis, or respiratoryburst in HIF-2a–deficient neutrophils. In an in vivo model of LPS-mediated lung injury, HIF-2a deficiency was associated with reducedneutrophilic inflammation during resolution, with fewer neutrophilsin BAL samples, an increase in neutrophil apoptosis, and a reductionin lung damage and vascular leak. Given that the changes in neu-trophil accumulation could in part be a consequence of alteredmacrophage function, the lung injury model was repeated in wild-type mice following fractional radiation, which both preserves therecipient alveolar macrophage population and epithelium and enablesbone marrow reconstitution.28 Reduced neutrophilic inflammationwas again observed in the context of marrow reconstitution withHif2aflox/flox;LysMcre1/2 cells. An intrinsic neutrophil phenotypedependent on HIF-2a expression was subsequently confirmed throughthe ex vivo study of inflammatory neutrophils isolated from the airwaysfollowing LPS challenge. In this setting, HIF-2a–deficient neutrophilswere shown to have increased sensitivity to nitrosative stress-inducedapoptosis, matched with a significant reduction in catalase transcriptabundance, previously identified as aHif2a-regulated gene.32 Together,these data suggest that during inflammation resolution, the neutrophilicinflammatory process is regulated by HIF-2a rather than HIF-1a ex-pression through the intrinsic regulation of neutrophil apoptosis.This is further supported by HIF-1a and HIF-2a expression data.First, we observed that neutrophils recruited from the circulation to thelung following LPS challenge significantly up-regulate Hif2amRNAbut not Hif1a mRNA. Second, Hif2a mRNA expression persists inBAL cells during inflammation resolution after Hif1a mRNA ex-pression has substantially reduced.
The phenotype of reduced neutrophilic inflammation duringresolution of an acute lung injury contrasts with our findings inPHD3-deficient animals, in which we observed a specific role forPHD3 in regulating neutrophilic inflammation in the context bothof whole animal and localized tissue hypoxia.17 Although PHD3is essential for neutrophil survival and inflammatory responses inhypoxia, we propose that the effects of HIF-2a deficiency in infla-mmatory neutrophils occur independently of oxygen tension, andtargeting HIF-2a may therefore be of greater clinical utility ininflammation in tissues, such as the lung, where oxygen tensioncan vary widely.33 Through modulation of tumor cell apoptosis,HIF-2a has emerged as a potential therapeutic target in cancerbiology.34,35 Our data now implicate HIF-2a in neutrophilicinflammation and propose that selective inhibition of HIF-2a mayallow effective control of neutrophil-mediated inflammation andof inflammation resolution without compromising host defenses.
Acknowledgments
We thank Dr Rachael Kilding, Dr Mohammed Akil, and Dr JohnBoulton for help recruiting patients with inflammatory arthritis andperforming synovial fluid aspiration; Emma Connelly and YvonneStephenson for help with western blots and immunohistochemis-try; Dr Andrew Cowburn for help with in vivo murine models ofregional neutrophilic inflammation; Dr Colin Bingle for his helpobtaining lung biopsy specimens; and Prof Matthias Mack foruseful discussion of in vivo murine models.
This work was supported by a Medical Research Council (MRC)Clinical Training Fellowship award to A.A.R.T. (G0802255), aWellcome Trust Intermediate Clinical Fellowship award to S.R.W.(078244), a Wellcome Trust Senior Clinical Fellowship award toS.R.W. (098516), a Wellcome Trust Senior Clinical Fellowshipaward to D.H.D. (076945), a MRC Senior Clinical Fellowship awardto S.A.R. (G0701932), and a British Lung Foundation fellowshipto H.M.M. (F05/7). The National Institute for Health ResearchSheffield Biomedical Research Unit in Cardiovascular Diseaseand zebrafish work were supported by an MRC Centre Grant(G0700091).
Authorship
Contribution: A.A.R.T. performed the research and wrote themanuscript; P.M.E., H.M.M., S.E., K.R.H., A.L., L.W., S.P., G.S.,and E.E.M. performed the research; F.F., F.J.V.E., V.L.K., C.W.P.,P.A.R., M.J.P., M.C.S., R.S.J., and S.A.R. provided access tonovel analytical tools, transgenic organisms, and patient samples;S.A.R., I.S., D.H.D., and E.R.C. interpreted data; and M.K.B.W.and S.R.W. designed the research and wrote the manuscript.
Conflict-of-interest disclosure: C.W.P. is a scientific cofounderof, and holds equity in, ReOx Ltd. The remaining authors declareno competing financial interests.
Correspondence: Sarah Walmsley, Academic Unit of Respira-tory Medicine, The Medical School, University of Sheffield, SheffieldS10 2RX, United Kingdom; e-mail: [email protected]; andMoira Whyte, Academic Unit of Respiratory Medicine, The MedicalSchool, University of Sheffield, Sheffield S10 2RX,UnitedKingdom;e-mail: [email protected].
BLOOD, 16 JANUARY 2014 x VOLUME 123, NUMBER 3 HIF2A AND NEUTROPHIL FUNCTION 375

References
1. Barnes PJ. New molecular targets for thetreatment of neutrophilic diseases. J Allergy ClinImmunol. 2007;119(5):1055-1062, quiz 1063-1064.
2. Quint JK, Wedzicha JA. The neutrophil in chronicobstructive pulmonary disease. J Allergy ClinImmunol. 2007;119(5):1065-1071.
3. Whyte MK, Meagher LC, MacDermot J, Haslett C.Impairment of function in aging neutrophils isassociated with apoptosis. J Immunol. 1993;150(11):5124-5134.
4. Savill JS, Wyllie AH, Henson JE, Walport MJ,Henson PM, Haslett C. Macrophage phagocytosisof aging neutrophils in inflammation. Programmedcell death in the neutrophil leads to its recognitionby macrophages. J Clin Invest. 1989;83(3):865-875.
5. Rossi AG, Sawatzky DA, Walker A, et al.Cyclin-dependent kinase inhibitors enhancethe resolution of inflammation by promotinginflammatory cell apoptosis. Nat Med. 2006;12(9):1056-1064.
6. Walmsley SR, Print C, Farahi N, et al. Hypoxia-induced neutrophil survival is mediated by HIF-1alpha-dependent NF-kappaB activity. J ExpMed. 2005;201(1):105-115.
7. Hannah S, Mecklenburgh K, Rahman I, BellinganGJ, Greening A, Haslett C, Chilvers ER. Hypoxiaprolongs neutrophil survival in vitro. FEBS Lett.1995;372(2-3):233-237.
8. Mecklenburgh KI, Walmsley SR, Cowburn AS,et al. Involvement of a ferroprotein sensor inhypoxia-mediated inhibition of neutrophilapoptosis. Blood. 2002;100(8):3008-3016.
9. Wang GL, Jiang BH, Rue EA, Semenza GL.Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2tension. Proc Natl Acad Sci USA. 1995;92(12):5510-5514.
10. Lando D, Peet DJ, Whelan DA, Gorman JJ,Whitelaw ML. Asparagine hydroxylation of the HIFtransactivation domain a hypoxic switch. Science.2002;295(5556):858-861.
11. Epstein AC, Gleadle JM, McNeill LA, et al. C.elegans EGL-9 and mammalian homologs definea family of dioxygenases that regulate HIF byprolyl hydroxylation. Cell. 2001;107(1):43-54.
12. Bruick RK, McKnight SL. A conserved family ofprolyl-4-hydroxylases that modify HIF. Science.2001;294(5545):1337-1340.
13. Peyssonnaux C, Datta V, Cramer T, et al. HIF-1alpha expression regulates the bactericidalcapacity of phagocytes. J Clin Invest. 2005;115(7):1806-1815.
14. Imtiyaz HZ, Williams EP, Hickey MM, et al.Hypoxia-inducible factor 2alpha regulatesmacrophage function in mouse models of acuteand tumor inflammation. J Clin Invest. 2010;120(8):2699-2714.
15. Peyssonnaux C, Cejudo-Martin P, Doedens A,Zinkernagel AS, Johnson RS, Nizet V. Cuttingedge: Essential role of hypoxia inducible factor-1alpha in development of lipopolysaccharide-induced sepsis. J Immunol. 2007;178(12):7516-7519.
16. Cramer T, Yamanishi Y, Clausen BE, et al.HIF-1alpha is essential for myeloid cell-mediatedinflammation. Cell. 2003;112(5):645-657.
17. Walmsley SR, Chilvers ER, Thompson AA,et al. Prolyl hydroxylase 3 (PHD3) is essential forhypoxic regulation of neutrophilic inflammation inhumans and mice. J Clin Invest. 2011;121(3):1053-1063.
18. Hu CJ, Wang LY, Chodosh LA, Keith B, SimonMC. Differential roles of hypoxia-inducible factor1alpha (HIF-1alpha) and HIF-2alpha in hypoxicgene regulation. Mol Cell Biol. 2003;23(24):9361-9374.
19. Warnecke C, Weidemann A, Volke M, et al. Thespecific contribution of hypoxia-inducible factor-2alpha to hypoxic gene expression in vitro islimited and modulated by cell type-specific andexogenous factors. Exp Cell Res. 2008;314(10):2016-2027.
20. Takeda N, O’Dea EL, Doedens A, et al.Differential activation and antagonistic function ofHIF-alpha isoforms in macrophages are essentialfor NO homeostasis. Genes Dev. 2010;24(5):491-501.
21. Haslett C, Guthrie LA, Kopaniak MM, JohnstonRB Jr, Henson PM. Modulation of multipleneutrophil functions by preparative methodsor trace concentrations of bacteriallipopolysaccharide. Am J Pathol. 1985;119(1):101-110.
22. Sabroe I, Jones EC, Usher LR, Whyte MK, DowerSK. Toll-like receptor (TLR)2 and TLR4 in humanperipheral blood granulocytes: a critical role formonocytes in leukocyte lipopolysaccharideresponses. J Immunol. 2002;168(9):4701-4710.
23. Renshaw SA, Loynes CA, Trushell DM, ElworthyS, Ingham PW, Whyte MK. A transgenic zebrafishmodel of neutrophilic inflammation. Blood. 2006;108(13):3976-3978.
24. Nusslein-Volhard C. Zebrafish: A PracticalApproach, 1st Ed. Oxford: Oxford UniversityPress; 2002.
25. Elks PM, van Eeden FJ, Dixon G, et al. Activationof hypoxia-inducible factor-1a (Hif-1a) delaysinflammation resolution by reducing neutrophilapoptosis and reverse migration in a zebrafishinflammation model. Blood. 2011;118(3):712-722.
26. Thisse C, Thisse B. High-resolution in situhybridization to whole-mount zebrafish embryos.Nat Protoc. 2008;3(1):59-69.
27. Manotham K, Tanaka T, Ohse T, et al. A biologicrole of HIF-1 in the renal medulla. Kidney Int.2005;67(4):1428-1439.
28. Tarling JD, Lin HS, Hsu S. Self-renewal ofpulmonary alveolar macrophages: evidence fromradiation chimera studies. J Leukoc Biol. 1987;42(5):443-446.
29. Percy MJ, Furlow PW, Lucas GS, Li X, Lappin TR,McMullin MF, Lee FS. A gain-of-function mutationin the HIF2A gene in familial erythrocytosis.N Engl J Med. 2008;358(2):162-168.
30. Prasch AL, Tanguay RL, Mehta V, Heideman W,Peterson RE. Identification of zebrafish ARNT1homologs: 2,3,7,8-tetrachlorodibenzo-p-dioxintoxicity in the developing zebrafish requiresARNT1. Mol Pharmacol. 2006;69(3):776-787.
31. Jonsson H, Allen P, Peng SL. Inflammatoryarthritis requires Foxo3a to prevent Fas ligand-induced neutrophil apoptosis. Nat Med. 2005;11(6):666-671.
32. Scortegagna M, Ding K, Oktay Y, et al. Multipleorgan pathology, metabolic abnormalities andimpaired homeostasis of reactive oxygen speciesin Epas1-/- mice. Nat Genet. 2003;35(4):331-340.
33. Hamedani H, Kadlecek SJ, Emami K, et al. Amultislice single breath-hold scheme for imagingalveolar oxygen tension in humans. Magn ResonMed. 2012;67(5):1332-1345.
34. Bertout JA, Patel SA, Simon MC. The impact ofO2 availability on human cancer. Nat Rev Cancer.2008;8(12):967-975.
35. Raval RR, Lau KW, Tran MG, et al. Contrastingproperties of hypoxia-inducible factor 1 (HIF-1)and HIF-2 in von Hippel-Lindau-associated renalcell carcinoma. Mol Cell Biol. 2005;25(13):5675-5686.
376 THOMPSON et al BLOOD, 16 JANUARY 2014 x VOLUME 123, NUMBER 3
Related Documents











