Hypoxia and Expression of Hypoxia-Inducible Factor in the Aging Kidney Tetsuhiro Tanaka, 1 Hideki Kato, 1 Ichiro Kojima, 1 Takamoto Ohse, 1 Daisuke Son, 1 Takahisa Tawakami, 1 Toshiya Yatagawa, 2 Reiko Inagi, 1 Toshiro Fujita, 1 and Masaomi Nangaku 1 Divisions of 1 Nephrology and Endocrinology, and 2 Pediatrics, University of Tokyo School of Medicine, Japan. Renal senescence is characterized by interstitial fibrosis and loss of peritubular capillaries. In this study, we provided evidence of tubulointerstitial hypoxia and the operation of hypoxia-inducible factor (HIF) in the aging kidney. Using two distinct methods, pimonidazole immunostaining and the expression of the ‘‘hypoxia-responsive’’ reporter of the transgenic rats, we identified the age-related expansion of hypoxia in all areas of the kidney. Expansion was most prominent in the cortex. Clusters of hypoxic tubules were observed in the superficial cortical zones, areas adjacent to the outer nephrons and expanded in the medullary rays. The degree of hypoxia was positively correlated with the age-related tubulointerstitial injury (R 2 ¼ 0.88, p , .01), which was associated with the upregulation of HIF-regulated genes, such as vascular endothelial growth factor (VEGF) and glucose transporter-1 (GLUT1) (real-time polymerase chain reaction). These findings point to the involvement of hypoxia and highlight the pathological relevance of HIF and its target genes in the aging kidney. A GING in the kidney has been linked to a number of physiological as well as structural derangements (1,2). Functionally, glomerular filtration rate (GFR) starts to de- cline at 30 years of age by approximately 8 mL/min per 1.73 m 2 per decade (3,4). Renal blood flow (RBF) is also reduced, with redistribution of blood flow from the cortex to the medulla. As a result, renal cortex is likely to be exposed to an environment in which oxygen and nutrient supply is low. Structurally, various degrees of focal and segmental glomerular sclerosis develop with aging, as well as tubu- lointerstitial injury such as tubular basement membrane (TBM) thickening, tubular dilatation, cast formation, tubular atrophy, and interstitial fibrosis. In areas with severe tubulointerstitial injury, patchy loss of peritubular capillary networks has been documented (5). These previous observations point to the potential involvement of hypoxia during the pathogenesis of age- related tubulointerstitial injury. In fact, tubulointerstitial hypoxia has been recognized not only as a hallmark of, but as a common mediator to, progressive glomerular diseases (6–8). It remains unclear, however, whether these scenarios apply in the process of physiological aging; still less is it certain whether the aging kidney is exposed to hypoxia, thus hampering the relevance of hypoxia to the age-related renal pathology. At the cellular level, every intrinsic cell has an inborn mechanism to cope with hypoxia. On exposure to hypoxia, cells try to adapt to the environment by producing factors that promote for example, angiogenesis, erythropoiesis, and glycolysis. These adaptive mechanisms apply to all organs and tissues, including the kidney (9,10). Hypoxia-inducible factor (HIF) is a heterodimeric tran- scription factor that plays a central role in such cellular adaptation to hypoxia (11,12). It is composed of two basic helix–loop–helix proteins—HIF-a and HIF-b—of the PAS (Per-Arnt-Sim) family. In hypoxia, the a/b heterodimer binds to a core DNA motif in the hypoxia-responsive element (HRE) and transactivates its target genes, such as erythropoietin (EPO) and vascular endothelial growth factor (VEGF). The transcriptional activity of HIF is regulated through post-transcriptional hydroxylation catalyzed by a set of oxygen-dependent enzymes that belong to the 2- oxoglutarate-dependent oxygenase superfamily (13). In the presence of molecular oxygen, the a subunit of HIF undergoes hydroxylation at specific prolyl residues in the oxygen-dependent degradation domain (ODD; prolyl hydroxylation) (14,15). Besides, the HIF transcriptional activity is modulated through asparaginyl hydroxylation. Oxygen promotes hydroxylation at the specific asparaginyl residue and blocks the binding of transcriptional coactiva- tors such as p300 (16,17). Hence, the expression and the functional operation of HIF are under the tight regulation of hypoxia. On the basis of these previous observations, we postulated that the aging kidney might be exposed to hypoxia, investigated the activation of HIF, and character- ized the expression of HIF-regulated genes in the aging kidney. MATERIALS AND METHODS Experimental Rats Male, ‘‘hypoxia-sensing’’ transgenic rats of Wistar strain were used in this study (18). They are transgenic rats harboring a transgene composed of HRE (enhancer) and the FLAG-tagged luciferase reporter gene, allowing us to detect areas of hypoxia through HIF-mediated cellular hypoxic 795 Journal of Gerontology: BIOLOGICAL SCIENCES Copyright 2006 by The Gerontological Society of America 2006, Vol. 61A, No. 8, 795–805 by guest on August 11, 2015 http://biomedgerontology.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hypoxia and Expression of Hypoxia-InducibleFactor in the Aging Kidney
Tetsuhiro Tanaka,1 Hideki Kato,1 Ichiro Kojima,1 Takamoto Ohse,1 Daisuke Son,1
Takahisa Tawakami,1 Toshiya Yatagawa,2 Reiko Inagi,1 Toshiro Fujita,1 and Masaomi Nangaku1
Divisions of 1Nephrology and Endocrinology, and 2Pediatrics, University of Tokyo School of Medicine, Japan.
Renal senescence is characterized by interstitial fibrosis and loss of peritubular capillaries. In thisstudy, we provided evidence of tubulointerstitial hypoxia and the operation of hypoxia-induciblefactor (HIF) in the aging kidney. Using two distinct methods, pimonidazole immunostainingand the expression of the ‘‘hypoxia-responsive’’ reporter of the transgenic rats, we identified theage-related expansion of hypoxia in all areas of the kidney. Expansion was most prominentin the cortex. Clusters of hypoxic tubules were observed in the superficial cortical zones, areasadjacent to the outer nephrons and expanded in the medullary rays. The degree of hypoxia waspositively correlated with the age-related tubulointerstitial injury (R2¼ 0.88, p , .01), which wasassociated with the upregulation of HIF-regulated genes, such as vascular endothelial growthfactor (VEGF) and glucose transporter-1 (GLUT1) (real-time polymerase chain reaction). Thesefindings point to the involvement of hypoxia and highlight the pathological relevance of HIFand its target genes in the aging kidney.
AGING in the kidney has been linked to a number ofphysiological as well as structural derangements (1,2).
Functionally, glomerular filtration rate (GFR) starts to de-cline at 30 years of age by approximately 8 mL/min per1.73 m
2 per decade (3,4). Renal blood flow (RBF) is alsoreduced, with redistribution of blood flow from the cortex tothe medulla. As a result, renal cortex is likely to be exposedto an environment in which oxygen and nutrient supply islow. Structurally, various degrees of focal and segmentalglomerular sclerosis develop with aging, as well as tubu-lointerstitial injury such as tubular basement membrane(TBM) thickening, tubular dilatation, cast formation, tubularatrophy, and interstitial fibrosis. In areas with severetubulointerstitial injury, patchy loss of peritubular capillarynetworks has been documented (5).
These previous observations point to the potentialinvolvement of hypoxia during the pathogenesis of age-related tubulointerstitial injury. In fact, tubulointerstitialhypoxia has been recognized not only as a hallmark of, butas a common mediator to, progressive glomerular diseases(6–8). It remains unclear, however, whether these scenariosapply in the process of physiological aging; still less is itcertain whether the aging kidney is exposed to hypoxia, thushampering the relevance of hypoxia to the age-relatedrenal pathology.
At the cellular level, every intrinsic cell has an inbornmechanism to cope with hypoxia. On exposure to hypoxia,cells try to adapt to the environment by producing factorsthat promote for example, angiogenesis, erythropoiesis, andglycolysis. These adaptive mechanisms apply to all organsand tissues, including the kidney (9,10).
Hypoxia-inducible factor (HIF) is a heterodimeric tran-scription factor that plays a central role in such cellular
adaptation to hypoxia (11,12). It is composed of two basichelix–loop–helix proteins—HIF-a and HIF-b—of the PAS(Per-Arnt-Sim) family. In hypoxia, the a/b heterodimerbinds to a core DNA motif in the hypoxia-responsiveelement (HRE) and transactivates its target genes, such aserythropoietin (EPO) and vascular endothelial growth factor(VEGF). The transcriptional activity of HIF is regulatedthrough post-transcriptional hydroxylation catalyzed bya set of oxygen-dependent enzymes that belong to the 2-oxoglutarate-dependent oxygenase superfamily (13). In thepresence of molecular oxygen, the a subunit of HIFundergoes hydroxylation at specific prolyl residues inthe oxygen-dependent degradation domain (ODD; prolylhydroxylation) (14,15). Besides, the HIF transcriptionalactivity is modulated through asparaginyl hydroxylation.Oxygen promotes hydroxylation at the specific asparaginylresidue and blocks the binding of transcriptional coactiva-tors such as p300 (16,17). Hence, the expression and thefunctional operation of HIF are under the tight regulationof hypoxia. On the basis of these previous observations,we postulated that the aging kidney might be exposed tohypoxia, investigated the activation of HIF, and character-ized the expression of HIF-regulated genes in the agingkidney.
MATERIALS AND METHODS
Experimental RatsMale, ‘‘hypoxia-sensing’’ transgenic rats of Wistar strain
were used in this study (18). They are transgenic ratsharboring a transgene composed of HRE (enhancer) and theFLAG-tagged luciferase reporter gene, allowing us to detectareas of hypoxia through HIF-mediated cellular hypoxic
795
Journal of Gerontology: BIOLOGICAL SCIENCES Copyright 2006 by The Gerontological Society of America2006, Vol. 61A, No. 8, 795–805
by guest on August 11, 2015
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

response. Rats of two age groups (young rats [YO; 4 monthsold, n ¼ 8] and aging rats [AG; 20 months old, born inOctober 2003, n ¼ 7]) were housed in a light- and tem-perature-controlled environment and were allowed free ac-cess to standard animal chow and water.
Study ProtocolOne day prior to death, rats of both age groups were
placed in metabolic cages to collect urine. Twenty-fourhours later, they were anesthetized with ketamine (50 mg/kgi.p.), blood was taken via cardiac puncture, and the leftkidneys were removed with midperitoneal incision. Re-moved kidneys were cut transversely and fixed with methylCarnoy’s or buffered formalin fixatives. A fraction ofcortical tissues was set aside, snap-frozen in liquid nitrogen,and stored at �808C for subsequent RNA and proteinpreparation. Proteinuria and serum creatinine levels weremeasured using a standard laboratory method (Bio-Rad,Hercules, CA and Wako, Osaka, Japan). One rat in the AGgroup was found later to have suffered from lung cancerwith pleural invasion; the data on this rat were excludedfrom the study. All protocols conformed to the NationalInstitutes of Health Guide for the Care and Use ofLaboratory Animals and were approved by the committeeon ethical animal care at Tokyo University.
In another set of experiments, hypoxia in the aging kid-ney was corroborated with the use of pimonidazole, a 2-nitroimidazole (Hypoxyprobe-1 Kit; Chemicon, Temecula,CA). Both YO and AG rats received injections ofpimonidazole at 60 mg/kg via tail vein, followed 2 hourslater by removal of the kidneys for immunohistochemicaldetection of hypoxia.
Histological EvaluationParaffin-embedded sections (3 lm) were stained with
periodic acid-Schiff, and tubulointerstitial injury wasassessed semiquantitatively on the basis of morphologicalchanges such as tubular dilatation, cast formation, sloughingof tubular epithelial cells, and thickening of the TBM,as follows: Grade 1, ,10% of tubules involved; Grade2, ,25%; Grade 3, ,50%; Grade 4, ,75%; and Grade5, �75%. Twenty consecutive fields in the cortex were ex-amined at 3400 magnification and averaged per slide. Allquantification was made in a blinded manner.
ImmunohistochemistryHypoxic tubular cells were identified using a modified
indirect immunoperoxidase method, either on the basis of anincrease in the hypoxia-responsive transgene expression orby the positive staining for pimonidazole. Methyl Carnoy’s-fixed, paraffin-embedded sections were dewaxed andbrought to water through graded ethanols. After quenchingof endogenous peroxidase activity (0.3% H2O2 in methanol,20 minutes), sections were probed with anti-FLAG (M2)antibody (1:350; Sigma, St. Louis, MO) or Hypoxyprobe-1Mb1 (1:200; Chemicon) followed by incubation withcorresponding, biotinylated secondary antibodies (VectorLaboratories, Burlingame, CA). Specific signals weredetected with the tyramide signal amplification method(for FLAG; Perkin Elmer, Wellesley, MA) or the avidin-
biotinylated peroxidase complex method (for Hypoxyprobe)method, followed by color development with H2O2 anddiaminobenzidine.
For the immunodetection of HIF-1a and HIF-2a, spe-cific antibodies were used as follows: antihuman HIF-1aantibody (1:100; Novus Biologicals, Littleton, CO), anti-mouse HIF-2a antibody (1:3000, PM9; a gift from Dr.Michael S. Wiesener, University of Erlangen-Nuremberg,Germany). Sections were autoclaved (1218C, 20 minutes)in 10 mM citrate buffer (pH 6.0) for antigen retrieval. Toreduce run-to-run variations in the staining intensity, allstaining of the same antibody was performed in thesame run for all YO and AG rat kidneys, as previouslydescribed (19).
Quantification of the Transgene and HIF-RegulatedGene Messenger RNA
Messenger RNA (mRNA) expression of the transgene(HRE-Luc), HIF-a isoforms (HIF-1a and HIF-2a), andseveral of known HIF-regulated genes was quantified byreal-time polymerase chain reaction (PCR). RNA wasisolated from the renal cortex by using ISOGEN (NipponGene, Tokyo, Japan) and 1 lg of template was reverse-transcribed (ImProm-II Reverse Transcription System;Promega, Madison, WI). One-twentieth (vol/vol) of thecomplementary DNA product was used as a template forsubsequent quantification. Using iQ SYBR Green Supermix(Bio-Rad), PCRs were performed on an iCycler (Bio-Rad)under the following conditions: initial denaturation of thetemplate at 948C for 15 minutes, 40 cycles of amplifi-cation at 948C for 15 seconds, 558C for 30 seconds, and728C for 30 seconds. Data collection and analysis wereperformed using iCycler iQ Optical System software(version 3.0a; Bio-Rad), and the amount of mRNA ofinterest was corrected for that of b-actin. The sets ofprimers used are as follows: (i) Luciferase: forward (fw):59-CGTTTCCAAAAAGGGGTTGC-39, reverse (rv): 59-GAAGGACTCTGGCACAAAATCG-39 (accession No.U47295 [pGL3-Basic], expected PCR product length: 178bp); (ii) HIF-1a: fw: 59-GTTTACTAAAGGACAAGT-CACC-39, rv: 59-TTCTGTTTGTTGAAGGGAG-39 (acces-sion No. NM 024359, expected PCR product length: 193bp); (iii) HIF-2a: fw: 59-GTCACCAGAACTTGTGC-39,rv: 59-CAAAGATGCTGTTCATGG-39 (accession No. NM023090, expected PCR product length: 249 bp); (iv)EPO: fw: 59-TACGTAGCCTCACTTCACTGCTT-39, rv:59-GCAGAAAGTATCCGCTGTGAGTGTTC-39 (acces-sion No. NM 017001, expected PCR product length: 113bp); (v) VEGF (which recognizes all isoforms [VEGF120,VEGF144, VEGF164, VEGF188]): fw: 59-TTACTGCTG-TACCTCCAC -39, rv: 59-ACAGGACGGCTTGAAGATA-39 (accession No. NM 031836, expected PCR productlength: 189 bp); (vi) GLUT1: fw: 59-CAGTTCGGC-TATAACACCGGTGTC-39, rv: 59-ATAGCGGTGGTTC-CATGTTT-39 (accession No. NM 138827, expected PCRproduct length: 84 bp); (vii) heme oxygenase-1 (HO-1):fw: 59-TCTATCGTGCTCGCATGAAC-39, rv: 59-CAGCT-CCTCAAACAGCTCAA-39 (accession No. NM 012580,expected PCR product length: 110 bp); and (viii) b-actin: fw: 59-CTTTCTACAATGAGCTGCGTG-39, rv:
796 TANAKA ET AL.
by guest on August 11, 2015
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

59-TCATGAGGTAGTCTGTCAGG-39 (accession No. NM031144, expected PCR product length: 306 bp).
ImmunoblottingThe increase in the transgene, HIF-1a, VEGF, GLUT1,
and HO-1 in the aging kidney was corroborated by westernblotting. Whole cortical tissue was homogenized in 1%Nonidet P40 lysis buffer (150 mM sodium chloride, 1%Nonidet P40, 50 mM Tris–HCl, pH 8.0) containingleupeptin and pepstatin A (Sigma). Soluble lysates (50 lg)were loaded, resolved by 7.5% (HIF-1a), 10% (transgeneand GLUT1), and 12% (VEGF and HO-1) sodium dodecylsulfate–polyacrylamide gel electrophoresis (SDS–PAGE)under reducing conditions and transferred onto polyvinyli-dine difluoride (PVDF) membranes (Amersham, Piscat-away, NJ). After blocking with 5% skim milk, membraneswere probed with primary antibodies (above and antiratHO-1 antibody [1:1000; Stressgen, Victoria, BC, Canada])followed by corresponding alkaline phosphatase-conjugatedsecondary antibodies (Promega). Specific bands werevisualized with BCIP/NBT (Sigma). Coomassie brilliantblue (CBB) staining of the membrane confirmed equalloading and transfer.
Isolation of Inner Medullary Collecting Duct CellsTo test whether cells in the inner medulla activate HIF
under physiological hypoxia, inner medullary collectingduct (IMCD) cells were isolated from HRE-Luc–transgenicrats and exposed to 2% O2, as previously described (20,21).In brief, renal medulla were excised, cut into small (1–2mm3) pieces, and digested with 0.2% collagenase and 0.2%hyaluronidase (Sigma). After the enzymatic digestion, cellswere isolated by three short low-speed centrifugations (175g for 8 minutes, twice at 28 g for 2 minutes) and wereseeded on six-well culture plates (Techno Plastic Products,Trasadingen, Switzerland) precoated with gelatin. With thismethod, more than 90% of obtained cells were collectingduct cells, as confirmed by positive immunostaining foraquaporin-2 (Chemicon). Then, cells were cultured withDulbecco’s modified Eagle medium containing 5% fetalcalf serum (JRH Biosciences, Lenexa, KS) at 378C undera humidified atmosphere of 5%CO2/95% air, grown toconfluence, and exposed to hypoxia for 6 hours. Assumingthe physiological medullary partial pressure of oxygen tobe approximately 15 mmHg (22), we subjected IMCD cellsto 2% oxygen (APM-30D multi-gas incubator; Astec, Fu-kuoka, Japan). The HIF activity was estimated by measuringthe amount of the transgene (HRE-Luc) mRNA by real-timePCR, as described above. For practical reasons, only rats inthe YO group were used in this study.
Statistical AnalysisData were expressed as mean 6 standard deviation or
mean 6 standard error of the mean, as appropriate. Analyseswere carried out with StatView software (version 5.0; SASInstitute, Cary, NC). Differences among groups werecompared by unpaired Student t tests with the Bonferronicorrection or Dunn’s method. Values of p below .05 wereconsidered to be statistically significant.
RESULTS
Animal CharacteristicsPhysical and biochemical characteristics of the YO and
AG rats are summarized in Table 1. In the AG rats, therewas a large individual difference in the serum creatininelevel, which was yet significantly higher than in the YO rats(0.89 6 0.14 mg/dL vs 0.69 6 0.05 mg/dL, p , .01). Inaddition, the AG rats developed significant degrees ofproteinuria as compared to the YO rats (85.9 6 22.4 mg/dayvs 13.0 6 3.4 mg/day, p , .01), in keeping with theprevious description on proteinuria in the aged laboratoryrats (23).
Pathological Correlation of Hypoxia withTubulointerstitial Injury
Periodic acid-Schiff staining of the AG rat kidneysrevealed various degrees of glomerular and tubulointerstitialinjury. Some glomeruli were small and globally sclerotic,whereas the thickening of the glomerular basementmembrane (GBM) was more frequently observed. In thetubulointerstitium, TBMs appeared slightly thickened withpositive immunohistochemical staining for fibronectin andcollagen IV (not shown). In areas displaying moderate tosevere tubulointerstitial injury, structural changes such astubular dilatation, intratubular cast formation, tubularatrophy, and marked interstitial widening were observed(Figure 1A and B). Areas of fibrosis were also seen(Masson-Trichrome stain, not shown).
Pathological correlation between hypoxia and tubuloin-terstitial injury was obtained by measuring the transgeneexpression and the semiquantitative tubulointerstitial injuryscore in each individual AG rat (Figure 1C). A strongpositive correlation was observed (R2 ¼ 0.88, p , .01),suggesting the involvement of hypoxia in the developmentof age-related renal pathogenesis.
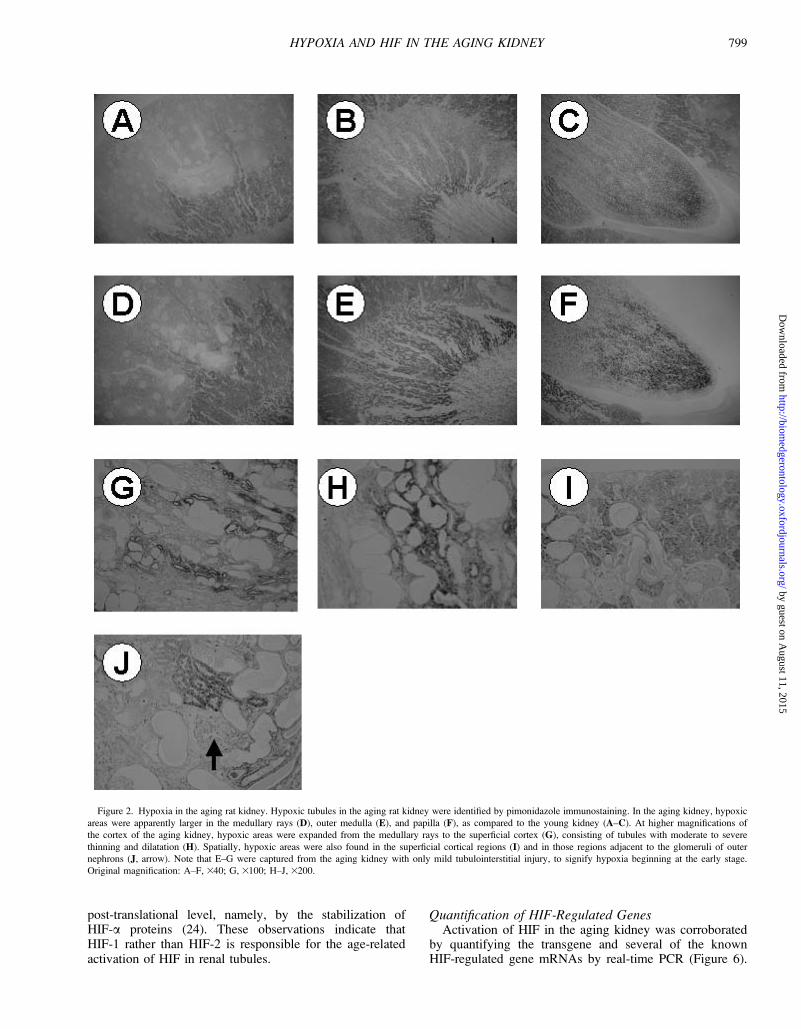
Identification of Hypoxic TubulesThen, areas of hypoxic tubules were identified by
pimonidazole, a chemical marker of hypoxia (Figure 2). Incontrast to the YO kidney (A–C), the AG kidney (D–F)showed marked expansion of hypoxic areas in the cortex (Aand D), medulla (B and E), and papilla (C and F), even inkidneys with mild to moderate tubulointerstitial injury.Overall, however, the age-related appearance of tubulointer-stitial hypoxia was most prominent in the cortex. In thecortex with mild to moderate injury, areas of positivestaining were broader in the medullary rays (D). On
Table 1. Animal Characteristics
Characteristic Young (YO) Rats Aged (AG) Rats
Age, mo 4 20
Body weight, g 441.3 6 53.8 450.0 6 45.4
Serum creatinine, mg/dL 0.69 6 0.05 0.89 6 0.14*
Urinary protein, mg/dL 13.0 6 3.4 85.9 6 22.4*
Notes: Data are expressed as mean 6 standard deviation.
N¼ 8 (YO) and n¼ 6 (AG), respectively.
*p , .01 versus the YO group.
797HYPOXIA AND HIF IN THE AGING KIDNEY
by guest on August 11, 2015
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

examination of the cortex with severe tubulointerstitialinjury, areas of hypoxic tubules obviously extended fromthe medullary rays to the superficial cortex (G), with sometubules markedly dilated and distorted (H), whereas othersappeared to reside in superficial cortical areas (I) and inthose areas adjacent to glomeruli of outer nephrons (J).
Activation of HIF in the Aging KidneyIdentification of hypoxic tubules in the aging kidney led
us to investigate whether there was any increase in HIF-activation (Figure 3). An age-related activation of HIF, asdetermined by the upregulation of the transgene, wasobserved in the cortex (A and B), outer medulla (C and D),and papilla (G and H), as could be envisaged from resultsof pimonidazole staining (Figure 2). Of interest, tubulesin the inner medulla appeared to express the transgeneto a similar degree in both groups, at levels higherthan in the cortex of the young kidney (E and F). The in-ner medullary expression of the transgene was reminiscentof the borderline hypoxia in these areas, and suggested thatHIF might be constitutively active in the physiologicconditions of the kidney, regardless of age.
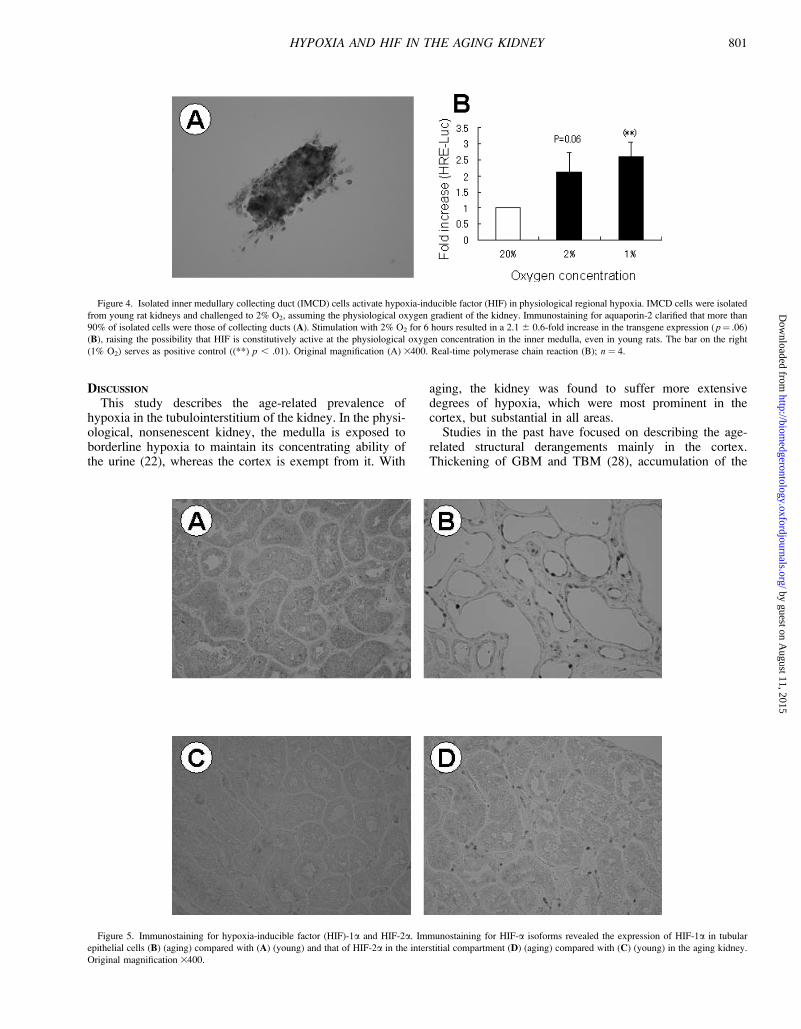
To address this question, IMCD cells were isolated fromthe HRE-Luc–transgenic rats and exposed to 2% O2,
assuming that the physiologic oxygenation in the deepmedulla is approximately 15 mmHg (Figure 4). Immunos-taining for aquaporin-2 confirmed that the isolated cellswere collecting ducts (A). When these cells were exposedto hypoxia for 6 hours, the transgene expression became2.1 6 0.6-fold as compared to normoxia (p ¼ .06) (B).These results further raise the possibility that HIF is con-stitutively active in the inner medulla, in the physiologicoxygenation status.
Expression of HIF-1a and HIF-2a IsoformsTo determine which of the major HIF-a isoforms was
responsible for the tubular upregulation of HIF in the agingkidney, immunostaining for HIF-1a and HIF-2a wasperformed (Figure 5). In the cortex of the AG kidney,HIF-1a was expressed in the nuclei of tubules dominatedby severe structural changes (B), whereas HIF-2a wasrestricted to the interstitial compartment (D). Both HIF-1aand HIF-2a were negative in the cortex of the YO group(A and C). Of note, the mRNA expression of HIF-1a andHIF-2a was similar between the YO and AG rat kidneys(Table 2), consistent with the notion that the activationof HIF by hypoxia is primarily determined at the
Figure 1. Progression of histological damage in the aging kidney. In the periodic acid-Schiff staining, various degrees of tubulointerstitial as well as glomerular
injury were noted in the aging kidney. Whereas some kidneys showed almost unremarkable injury (A), others exhibited striking histological damage, such as tubular
basement membrane thickening, tubular dilatation and atrophy, cast formation, brush border loss, and interstitial fibrosis (B). In the aging kidneys, a positive correla-
tion was observed between the degree of hypoxia (x axis: transgene expression in the cortex, relative to the average of the young kidney counterparts) and the
score of tubulointerstitial injury (y axis) (R2 ¼ 0.88, p , .01) (C). Original magnification for A and B, 3200.
798 TANAKA ET AL.
by guest on August 11, 2015
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from
post-translational level, namely, by the stabilization ofHIF-a proteins (24). These observations indicate thatHIF-1 rather than HIF-2 is responsible for the age-relatedactivation of HIF in renal tubules.
Quantification of HIF-Regulated GenesActivation of HIF in the aging kidney was corroborated
by quantifying the transgene and several of the knownHIF-regulated gene mRNAs by real-time PCR (Figure 6).
Figure 2. Hypoxia in the aging rat kidney. Hypoxic tubules in the aging rat kidney were identified by pimonidazole immunostaining. In the aging kidney, hypoxic
areas were apparently larger in the medullary rays (D), outer medulla (E), and papilla (F), as compared to the young kidney (A–C). At higher magnifications of
the cortex of the aging kidney, hypoxic areas were expanded from the medullary rays to the superficial cortex (G), consisting of tubules with moderate to severe
thinning and dilatation (H). Spatially, hypoxic areas were also found in the superficial cortical regions (I) and in those regions adjacent to the glomeruli of outer
nephrons (J, arrow). Note that E–G were captured from the aging kidney with only mild tubulointerstitial injury, to signify hypoxia beginning at the early stage.
Original magnification: A–F, 340; G, 3100; H–J, 3200.
799HYPOXIA AND HIF IN THE AGING KIDNEY
by guest on August 11, 2015
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

As already stated, the transgene mRNA, although diverseamong individuals, was significantly higher in the AG group(4.6 6 3.3-fold, p ¼ .03). The amount of EPO (25) (p ,.01), VEGF (26) (p , .001), and GLUT1 (p , .01) mRNAswere all significantly larger in the AG group, whereas thatof HO-1 (27) was comparable to the YO group, due tolarge individual variations (p¼ .08).
Quantitative changes in the transgene, HIF-1a, VEGF(p21 fragment), GLUT1, and HO-1 were analyzed at the
protein level as well. Western blotting revealed the over-all, coordinated upregulation of these HIF-associatedgenes in the AG group, although, once again, therewere quantitative variations among aging individuals(Figure 7 and quantified in Table 3). Unfortunately, how-ever, the age-related increase of HIF-1a and GLUT1 pro-teins was only marginal by this method, possibly due totechnical difficulties and their limited expression at thebasal level.
Figure 3. Expression of the ‘‘hypoxia-responsive’’ transgene. Tubular cell hypoxia was also identified by the upregulation of the hypoxia-responsive reporter
of the transgenic rats. In the aging kidney, the expression was obviously higher in the cortex (B), outer medulla (D), and papilla (H), as compared to the young kidney
(A, C, and G). In the inner medulla, however, the transgene appeared to be expressed at a higher level than in the cortex of the young kidney, regardless of age
(E and F). Original magnification 3200.
800 TANAKA ET AL.
by guest on August 11, 2015
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from
DISCUSSION
This study describes the age-related prevalence ofhypoxia in the tubulointerstitium of the kidney. In the physi-ological, nonsenescent kidney, the medulla is exposed toborderline hypoxia to maintain its concentrating ability ofthe urine (22), whereas the cortex is exempt from it. With
aging, the kidney was found to suffer more extensivedegrees of hypoxia, which were most prominent in thecortex, but substantial in all areas.
Studies in the past have focused on describing the age-related structural derangements mainly in the cortex.Thickening of GBM and TBM (28), accumulation of the
Figure 4. Isolated inner medullary collecting duct (IMCD) cells activate hypoxia-inducible factor (HIF) in physiological regional hypoxia. IMCD cells were isolated
from young rat kidneys and challenged to 2% O2, assuming the physiological oxygen gradient of the kidney. Immunostaining for aquaporin-2 clarified that more than
90% of isolated cells were those of collecting ducts (A). Stimulation with 2% O2 for 6 hours resulted in a 2.1 6 0.6-fold increase in the transgene expression ( p¼ .06)
(B), raising the possibility that HIF is constitutively active at the physiological oxygen concentration in the inner medulla, even in young rats. The bar on the right
(1% O2) serves as positive control ((**) p , .01). Original magnification (A) 3400. Real-time polymerase chain reaction (B); n ¼ 4.
Figure 5. Immunostaining for hypoxia-inducible factor (HIF)-1a and HIF-2a. Immunostaining for HIF-a isoforms revealed the expression of HIF-1a in tubular
epithelial cells (B) (aging) compared with (A) (young) and that of HIF-2a in the interstitial compartment (D) (aging) compared with (C) (young) in the aging kidney.
Original magnification 3400.
801HYPOXIA AND HIF IN THE AGING KIDNEY
by guest on August 11, 2015
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

extracellular matrix and focal areas of tubular atrophy (5)have been characterized, and the loss of peritubularcapillaries has been observed in the affected areas (29).These structural changes pointed to the potential relevanceof hypoxia to the age-related tubulointerstitial injury.However, no direct proof has been provided so far.
We confirmed the prevalence of hypoxia in the agingkidney by using two distinct methods, namely: (i) positivestaining for pimonidazole, and (ii) the upregulation of the‘‘hypoxia-responsive’’ transgene. The former is based onthe hypoxia (pO2 , 10 mmHg)-dependent binding of the2-nitroimidazole to form adducts that serve as immunogens,and allows us to detect hypoxia immunohistochemically.Using this method, we identified areas of hypoxic tubulesexpanding in the medullary rays, superficial cortical areasand those adjacent to glomeruli of outer nephrons, in theaging kidney. Although how these areas are exposed tohypoxia is unclear, it seems plausible that renal hemody-namic changes play important roles. This may be envisagedby the anatomical characteristics that the medullary rays,areas most severely affected by hypoxia with aging, arepartly vascularized by the ascending vasa recta draining theouter medulla until emptying into interlobular veins (30).In addition, the superficial area is known to be marginallysupplied with blood flow for anatomical reasons. Further-more, the perfusion of outer nephrons decreases more withage than does that of the corticomedullary nephrons (31).These previous observations are highly suggestive of theinvolvement of renal hemodynamics in the age-relateddevelopment of hypoxia.
Age-related hypoxia, while undoubtedly most prominentin the cortex, was also evident in the medulla. This findingmay be explained partly by the impaired oxygen diffu-sion, as can be envisaged by the accumulation of the extra-cellular matrix. In renal medulla, there is a significantincrease in connective tissue with age, which is reportedlymore striking than in the cortex (32). Once again, however,renal hemodynamics is also a candidate contributor, becausethe blood supply of the medulla is essentially postglomer-ular, except for a fraction of the ‘‘true medullary arterioles’’originating directly from arcuate or interlobular arteries thatcan be seen after degeneration of the corresponding glo-meruli and tubules.
Tubulointerstitial hypoxia was also evidenced by theactivation of HIF. Based on the upregulation of the hypoxia-responsive transgene, the hypoxic response was calculatedas 4.6 6 3.3-fold in the aging rat kidneys, as compared tothe young rat counterparts. Cortical immunodetection ofHIF-a isoforms allowed us to identify HIF-1a in tubularepithelial cells and HIF-2a in the interstitial compartment;
these identifications are consistent with previous findingsin ischemic young rat kidneys (9) and suggest that HIF-1plays a dominant role in the hypoxic response of the agingtubular cells. Furthermore, quantification of HIF-regulatedgenes, such as EPO, VEGF, GLUT1, and HO-1, revealedtheir coordinated upregulation in the cortex of the agingkidney, indicating the functional operation of the HIF-mediated cellular response to hypoxia. The pathologicalrelevance of HIF and its target genes merits further study.
The expression of HIF-1a with aging has been reported insuch systemic organs as brain, liver (33), and carotid body(34), but the background for the increase in HIF-1a proteinremained largely unknown, except for one study reportingthat the upregulation of HIF-1a in the aging liver wasassociated with an increase in the reactive oxygen species(35). However, an effect of reactive oxygen species on HIFgenerally remains elusive. Now that we identified hypoxicareas expanding in the aging kidney by pimonidazolestaining, we propose in this study that the increase in HIF-ain the aging kidney (HIF-1a in tubular cells and HIF-2a inthe interstitium) is associated with hypoxia.
Hypoxia in the aging kidney was found to have a positivecorrelation with the degree of tubulointerstitial injury. It hasbeen documented from a human pathological study that theloss of peritubular capillaries is closely related to the degreeof tubulointerstitial injury and the residual renal function(36). Considering that renal perfusion is a crucial compo-nent in determining regional oxygenation, it is tempting topropose that hypoxia in the tubulointerstitium serves asa hallmark of age-related renal injury as well.
SummaryThis study demonstrated that the aging kidney is ex-
posed to extensive degrees of hypoxia and that the intrinsictubular cells respond to it by upregulating a number ofHIF-regulated genes. The degree of hypoxia most likelyserves as a hallmark of the age-related tubulointerstitial in-jury, as has been proposed and reported in glomerular dis-eases (6,18). Our future perspectives include the researchfor the expression of HIF and the transcriptional regula-tion of HIF target genes in aging individuals, in the contextof additional ischemic insult. Although aging kidneys ex-press HIF at a higher level than do young kidney counter-parts, this finding does not necessarily mean that the formerhas enough potential to further upregulate HIF whenexposed to additional hypoxic stimulation. This is poten-tially of therapeutic significance because maneuvers toactivate HIF (37) have been suggested to work ina renoprotective manner in a number of acute (38,39) aswell as chronic (40–42) renal diseases.
ACKNOWLEDGMENT
We acknowledge Grants-in-Aid for Scientific Research from the JapanSociety for the Promotion of Science (17390246).
Address correspondence to Masaomi Nangaku, MD, PhD, Division ofNephrology and Endocrinology, University of Tokyo School of Medicine,7-3-1, Hongo, Bunkyo-ku, Tokyo, Japan. E-mail: [email protected]
Table 2. Quantification of HIF-1a and HIF-2a mRNA
mRNA Young (YO) Rats Aging (AG) Rats
HIF-1a 1.00 6 0.22 1.11 6 0.26
HIF-2a 1.00 6 0.34 0.98 6 0.17
Notes: Data are expressed as mean 6 standard deviation.
Average expression level in the YO group was arbitrarily set at 1.
N¼ 8 (YO) and n¼ 6 (AG), respectively.
mRNA¼ messenger RNA; HIF¼ hypoxia-inducible factor.
802 TANAKA ET AL.
by guest on August 11, 2015
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from
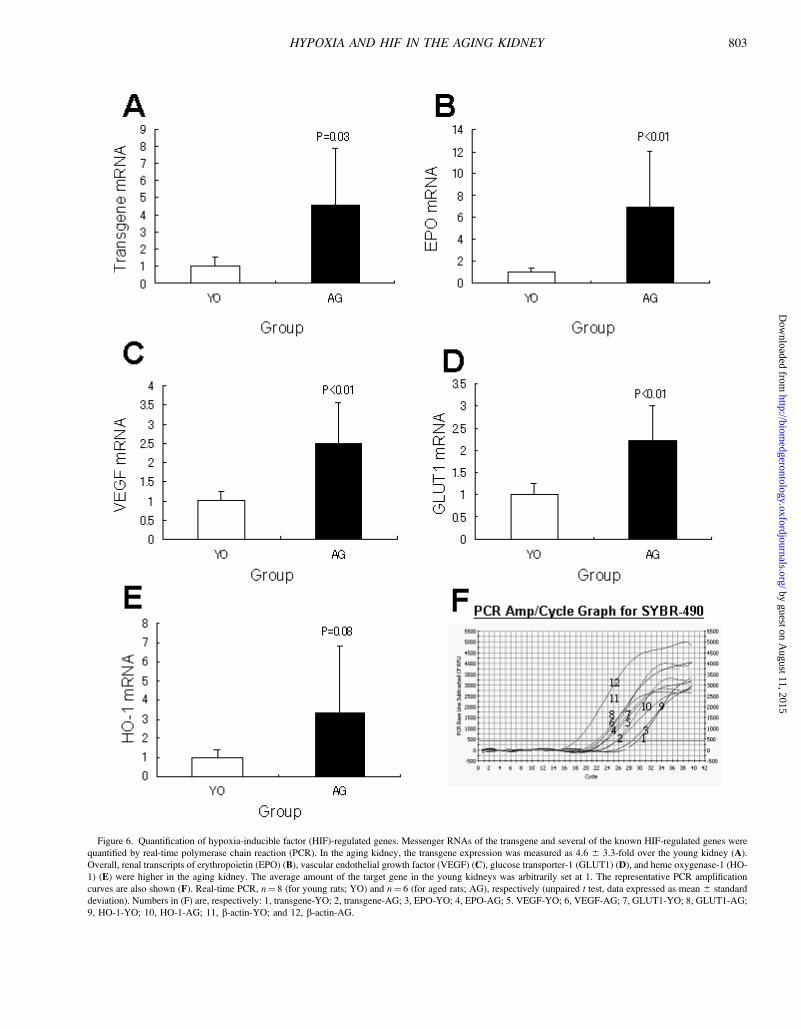
Figure 6. Quantification of hypoxia-inducible factor (HIF)-regulated genes. Messenger RNAs of the transgene and several of the known HIF-regulated genes were
quantified by real-time polymerase chain reaction (PCR). In the aging kidney, the transgene expression was measured as 4.6 6 3.3-fold over the young kidney (A).
Overall, renal transcripts of erythropoietin (EPO) (B), vascular endothelial growth factor (VEGF) (C), glucose transporter-1 (GLUT1) (D), and heme oxygenase-1 (HO-
1) (E) were higher in the aging kidney. The average amount of the target gene in the young kidneys was arbitrarily set at 1. The representative PCR amplification
curves are also shown (F). Real-time PCR, n¼ 8 (for young rats; YO) and n¼ 6 (for aged rats; AG), respectively (unpaired t test, data expressed as mean 6 standard
deviation). Numbers in (F) are, respectively: 1, transgene-YO; 2, transgene-AG; 3, EPO-YO; 4, EPO-AG; 5. VEGF-YO; 6, VEGF-AG; 7, GLUT1-YO; 8, GLUT1-AG;
9, HO-1-YO; 10, HO-1-AG; 11, b-actin-YO; and 12, b-actin-AG.
803HYPOXIA AND HIF IN THE AGING KIDNEY
by guest on August 11, 2015
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

REFERENCES
1. Anderson S, Brenner BM. Effects of aging on the renal glomerulus.
Am J Med. 1986;80:435–442.2. Lindeman RD, Goldman R. Anatomic and physiologic age changes
in the kidney. Exp Gerontol. 1986;21:379–406.3. Davies DF, Shock NW. Age changes in glomerular filtration rate,
effective renal plasma flow, and tubular excretory capacity in adult
males. J Clin Invest. 1950;29:496–507.4. Rowe JW, Andres R, Tobin JD, Norris AH, Shock NW. The effect of
age on creatinine clearance in men: a cross-sectional and longitudinal
study. J Gerontol. 1976;31:155–163.5. Thomas SE, Anderson S, Gordon KL, Oyama TT, Shankland SJ,
Johnson RJ. Tubulointerstitial disease in aging: evidence for underlying
peritubular capillary damage, a potential role for renal ischemia. J Am
Soc Nephrol. 1998;9:231–242.
6. Fine LG, Bandyopadhay D, Norman JT. Is there a common mechanism
for the progression of different types of renal diseases other than
proteinuria? Towards the unifying theme of chronic hypoxia. KidneyInt Suppl. 2000;75:S22–S26.
7. Eckardt KU, Rosenberger C, Jurgensen JS, Wiesener MS. Role of
hypoxia in the pathogenesis of renal disease. Blood Purif. 2003;21:
253–257.8. Nangaku M. Hypoxia and tubulointerstitial injury: a final common
pathway to end-stage renal failure. Nephron Exp Nephrol. 2004;98:
e8–e12.9. Rosenberger C, Mandriota S, Jurgensen JS, et al. Expression of
hypoxia-inducible factor-1alpha and -2alpha in hypoxic and ischemic
rat kidneys. J Am Soc Nephrol. 2002;13:1721–1732.10. Rosenberger C, Griethe W, Gruber G, et al. Cellular responses to
hypoxia after renal segmental infarction. Kidney Int. 2003;64:874–886.11. Wenger RH. Cellular adaptation to hypoxia: O2-sensing protein hy-
droxylases, hypoxia-inducible transcription factors, and O2-regulated
gene expression. FASEB J. 2002;16:1151–1162.12. Huang LE, Bunn HF. Hypoxia-inducible factor and its biomedical
relevance. J Biol Chem. 2003;278:19575–19578.13. Schofield CJ, Ratcliffe PJ. Oxygen sensing by HIF hydroxylases.
Nat Rev Mol Cell Biol. 2004;5:343–354.14. Ivan M, Kondo K, Yang H, et al. HIFalpha targeted for VHL-mediated
destruction by proline hydroxylation: implications for O2 sensing.
Science. 2001;292:464–468.15. Jaakkola P, Mole DR, Tian YM, et al. Targeting of HIF-alpha to the
von Hippel-Lindau ubiquitylation complex by O2-regulated prolyl
hydroxylation. Science. 2001;292:468–472.16. Lando D, Peet DJ, Whelan DA, Gorman JJ, Whitelaw ML. Asparagine
hydroxylation of the HIF transactivation domain a hypoxic switch.
Science. 2002;295:858–861.
Figure 7. Immunoblotting. The expression of hypoxia-inducible factor (HIF)-associated proteins was analyzed by western blotting. Cortical samples in the aged rat
(AG) group contained larger amounts of the transgene, vascular endothelial growth factor (VEGF; p21 fragment), and heme oxygenase-1 (HO-1) proteins, than did
those in the young rat (YO) group. Lanes 1–4: YO group; lanes 5–8: AG group.
Table 3. Quantification of the Transgene and HIF-Regulated Proteins
mRNA Young (YO) Rats Aged (AG) Rats
Transgene 36.8 6 8.2 88.5 6 35.9*
VEGF (p21) 38.0 6 3.8 79.8 6 12.4**
HO-1 29.8 6 1.0 78.3 6 35.1*
Notes: Expression levels of the transgene, vascular endothelial growth fac-
tor (VEGF) (p21 fragment), and heme oxygenase-1 (HO-1) were measured
densitometrically and expressed as mean 6 standard deviation. The expression
of hypoxia-inducible factor (HIF)-1a and glucose transporter-1 (GLUT-1) in the
YO group was below measurable limits, and was excluded from quantification.
*p , .05; **p , .01 versus the YO group, respectively.
804 TANAKA ET AL.
by guest on August 11, 2015
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

17. Mahon PC, Hirota K, Semenza GL. FIH-1: a novel protein that interactswith HIF-1alpha and VHL to mediate repression of HIF-1 transcrip-tional activity. Genes Dev. 2001;15:2675–2686.
18. Tanaka T, Miyata T, Inagi R, Fujita T, Nangaku M. Hypoxia in renaldisease with proteinuria and/or glomerular hypertension. Am J Pathol.2004;165:1979–1992.
19. Buzello M, Tornig J, Faulhaber J, Ehmke H, Ritz E, Amann K. Theapolipoprotein e knockout mouse: a model documenting acceleratedatherogenesis in uremia. J Am Soc Nephrol. 2003;14:311–316.
20. Stokes JB, Grupp C, Kinne RK. Purification of rat papillary collectingduct cells: functional and metabolic assessment. Am J Physiol. 1987;253:F251–F262.
21. Sandner P, Hofbauer KH, Tinel H, et al. Expression of adrenomedullin inhypoxic and ischemic rat kidneys and human kidneys with arterialstenosis. Am J Physiol Regul Integr Comp Physiol. 2004;286:R942–R951.
22. Brezis M, Rosen S. Hypoxia of the renal medulla–its implicationsfor disease. N Engl J Med. 1995;332:647–655.
23. Neuhaus OW, Flory W. Age-dependent changes in the excretion ofurinary proteins by the rat. Nephron. 1978;22:570–576.
24. Huang LE, Arany Z, Livingston DM, Bunn HF. Activation of hypoxia-inducible transcription factor depends primarily upon redox-sensitivestabilization of its alpha subunit. J Biol Chem. 1996;271:32253–32259.
25. Goldberg MA, Schneider TJ. Similarities between the oxygen-sensingmechanisms regulating the expression of vascular endothelial growthfactor and erythropoietin. J Biol Chem. 1994;269:4355–4359.
26. Levy AP, Levy NS, Wegner S, Goldberg MA. Transcriptional regu-lation of the rat vascular endothelial growth factor gene by hypoxia.J Biol Chem. 1995;270:13333–13340.
27. Lee PJ, Jiang BH, Chin BY, et al. Hypoxia-inducible factor-1 mediatestranscriptional activation of the heme oxygenase-1 gene in response tohypoxia. J Biol Chem. 1997;272:5375–5381.
28. Goldstein RS, Tarloff JB, Hook JB. Age-related nephropathy in labo-ratory rats. FASEB J. 1988;2:2241–2251.
29. Kang DH, Anderson S, Kim YG, et al. Impaired angiogenesis in theaging kidney: vascular endothelial growth factor and thrombospondin-1 in renal disease. Am J Kidney Dis. 2001;37:601–611.
30. Kriz W. Structural organization of the renal medulla: comparative andfunctional aspects. Am J Physiol. 1981;241:R3–R16.
31. Hollenberg NK, Adams DF, Solomon HS, Rashid A, Abrams HL,Merrill JP. Senescence and the renal vasculature in normal man. CircRes. 1974;34:309–316.
32. Darmady EM, Offer J, Woodhouse MA. The parameters of the ageing
kidney. J Pathol. 1973;109:195–207.33. Frenkel-Denkberg G, Gershon D, Levy AP. The function of hypoxia-
inducible factor 1 (HIF-1) is impaired in senescent mice. FEBS Lett.1999;462:341–344.
34. Di Giulio C, Bianchi G, Cacchio M, et al. Oxygen and life span:
chronic hypoxia as a model for studying HIF-1alpha, VEGF and NOS
during aging. Respir Physiol Neurobiol. 2005;147:31–38.35. Kang MJ, Kim HJ, Kim HK, et al. The effect of age and calorie
restriction on HIF-1-responsive genes in aged liver. Biogerontology.2005;6:27–37.
36. Bohle A, Mackensen-Haen S, Wehrmann M. Significance of post-
glomerular capillaries in the pathogenesis of chronic renal failure.
Kidney Blood Press Res. 1996;19:191–195.37. Warnecke C, Griethe W, Weidemann A, et al. Activation of the
hypoxia-inducible factor-pathway and stimulation of angiogenesis by
application of prolyl hydroxylase inhibitors. FASEB J. 2003;17:
1186–1188.38. Tanaka T, Kojima I, Ohse T, et al. Hypoxia-inducible factor modulates
tubular cell survival in cisplatin nephrotoxicity. Am J Physiol RenalPhysiol. 2005;289:F1123–F1133.
39. Matsumoto M, Makino Y, Tanaka T, et al. Induction of renoprotective
gene expression by cobalt ameliorates ischemic injury of the kidney in
rats. J Am Soc Nephrol. 2003;14:1825–1832.40. Kairaitis LK, Wang Y, Gassmann M, Tay YC, Harris DC. HIF-
1alpha expression follows microvascular loss in advanced murine
adriamycin nephrosis. Am J Physiol Renal Physiol. 2005;288:
F198–F206.41. Tanaka T, Kojima I, Ohse T, et al. Cobalt promotes angiogenesis via
hypoxia-inducible factor and protects tubulointerstitium in the remnant
kidney model. Lab Invest. 2005;85:1292–1307.42. Tanaka T, Matsumoto M, Inagi R, et al. Induction of protective genes
by cobalt ameliorates tubulointerstitial injury in the progressive Thy-1
nephritis. Kidney Int. 2005;68:2714–25.
Received September 30, 2005Accepted February 16, 2006Decision Editor: James R. Smith, PhD
805HYPOXIA AND HIF IN THE AGING KIDNEY
by guest on August 11, 2015
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from
Related Documents











