1 23

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1 23
������������ �������������� ������������������������������������������������ ��!�"����#����$%�&�%%�'(����)���*+ ������������**�%
����������� ���� ������������������������� ������� ����������� ������ ������������ ���������� ����
����������� ���������������������������������������������������������������� !��������"����� ����������#$���������%���

1 23
Your article is protected by copyright andall rights are held exclusively by SpringerInternational Publishing Switzerland. This e-offprint is for personal use only and shall notbe self-archived in electronic repositories. Ifyou wish to self-archive your article, pleaseuse the accepted manuscript version forposting on your own website. You mayfurther deposit the accepted manuscriptversion in any repository, provided it is onlymade publicly available 12 months afterofficial publication or later and providedacknowledgement is given to the originalsource of publication and a link is insertedto the published article on Springer'swebsite. The link must be accompanied bythe following text: "The final publication isavailable at link.springer.com”.

Hydrological transitions drive dissolved organic matterquantity and composition in a temporary Mediterraneanstream
Daniel von Schiller • Daniel Graeber •
Miquel Ribot • Xisca Timoner • Vicenc Acuna •
Eugenia Martı • Sergi Sabater • Klement Tockner
Received: 1 September 2014 / Accepted: 29 January 2015 / Published online: 7 February 2015
� Springer International Publishing Switzerland 2015
Abstract The implications of stream flow intermitten-
cy for dissolved organic matter (DOM) are not well
understood despite its potential significance for water
quality and ecosystem integrity. We combined intensive
sampling with liquid chromatography and spectroscopic
techniques to follow changes in DOC and DON concen-
trations as well as in DOM size fractions and spectro-
scopic properties in a temporary stream during an entire
contraction–fragmentation–expansion hydrological cy-
cle. DOC and DON concentrations remained low
(range = 1.4–5.2 mg C L-1 and 0.05–0.15 mg N L-1)
during hydrological contraction and fragmentation, with
concomitant increases in the proportion of high mole-
cularweight substances (HMWS) during contraction and
of DOM aromaticity during fragmentation. DOC and
DON concentrations abruptly increased (up to
8.8 mg C L-1 and 0.37 mg N L-1) at the end of the
fragmentation phase, with a concomitant increase in the
non-humic, microbial and aquatic character of DOM.
Upon rewetting, the DOC and DON concentrations
reached their highest values (up to 12.7 mg C L-1 and
0.39 mg N L-1), with concomitant increases in the
proportion of HMWS and in the humic, aromatic and
terrestrial character of DOM. Subsequently, DOC and
DON concentrations recovered to values similar to those
at the contraction phase, while DOM composition
variables indicated the prevalence of a DOM of humic
and terrestrial character during the whole expansion
phase. Overall, our results emphasize the importance of
hydrological transitions forDOMdynamics in temporary
streams, and point to the potential response of perennial
streams under future water scarcity scenarios.
Responsible Editor: Sujay Kaushal.
D. von Schiller (&) � X. Timoner � V. Acuna �S. Sabater
Catalan Institute for Water Research, Emili Grahit 101,
17003 Girona, Spain
e-mail: [email protected]
D. Graeber
Department of Bioscience, Aarhus University, Vejlsøvej
25, 8600 Silkeborg, Denmark
M. Ribot � E. Martı
Integrative Freshwater Ecology Group, Centre for
Advanced Studies of Blanes (CEAB-CSIC), Acces a la
Cala St. Francesc 14, 17300 Blanes, Spain
X. Timoner � S. SabaterInstitute of Aquatic Ecology, University of Girona,
17071 Girona, Spain
K. Tockner
Leibniz-Institute of Freshwater Ecology and Inland
Fisheries, Muggelseedamm 301, 12587 Berlin, Germany
K. Tockner
Institute of Biology, Freie Universitat Berlin, Takustraße
3, 14195 Berlin, Germany
123
Biogeochemistry (2015) 123:429–446
DOI 10.1007/s10533-015-0077-4
Author's personal copy

Keywords Drying � Rewetting � Dissolved organic
matter � Dissolved organic carbon � Temporary
stream � Drought
Introduction
Temporary streams are waterways that cease to flow at
some points in space and time along their course
(Acuna et al. 2014). A major part of the world’s river
networks consists of temporary streams (Larned et al.
2010; Datry et al. 2014). For instance, more than half
of the total river length in South Africa (Uys and
O’Keefe 1997), USA (Nadeau and Rains 2007), and
Greece (Tzoraki et al. 2007) is subject to flow
intermittency, and most Alpine, Arctic, and Antarctic
rivers are temporary (Datry et al. 2014). A recent
global estimate shows that about two-thirds of the
first-order streams and one-third of the larger, fifth-
order rivers below 60� latitude experience flow
intermittency (Raymond et al. 2013). However, tem-
porary streams are widely neglected in policy imple-
mentation (Nikolaidis et al. 2013; Acuna et al. 2014).
At the same time, the biogeochemical implications of
stream flow intermittency are still not well understood,
despite of their potential relevance for stream ecosys-
tem functioning (Lake 2011; Steward et al. 2012).
Temporary streams are the dominant surface water
type in the Mediterranean Basin (Bonada and Resh
2013; Prat et al. 2014), and the duration and extent of
stream flow intermittency is expected to increase in
this region, as well as in other regions, in response to
climate and land use change (IPCC 2013). Mediter-
ranean temporary streams exhibit a highly dynamic
hydrological regime with a characteristic seasonal
drought period (Bernal et al. 2013). Typically, stream
flow gradually decreases in late spring to early
summer (contraction phase), followed by flow cessa-
tion and the formation of isolated pools (fragmentation
phase) before the stream completely dries up at the
surface (dry phase). At the end of summer, when
evapotranspiration in the catchment decreases and
intense precipitation events are frequent, stream flow
is gradually or abruptly re-established (expansion
phase). The drying and rewetting cycle controls both
the longitudinal hydrological connectivity along the
stream and the lateral hydrological connectivity
between the stream and its valley, with potentially
important but widely unexplored effects on in-stream
biogeochemical cycling (Dahm et al. 2003; von
Schiller et al. 2011; Vazquez et al. 2011).
Dissolved organic matter (DOM) is a complex
mixture of organic compounds, which represents the
largest pool of transported organic matter in streams
and plays an essential role in aquatic ecosystem
biogeochemistry (Findlay and Sinsabaugh 2003;
Prairie 2008; Tank et al. 2010). This role depends
not only on the quantity of DOM but also on its
composition, which result from a combination of
inputs from the catchment (i.e. allochthonous sources)
and from in-stream production (i.e. autochthonous
sources) (Webster and Meyer 1997). Among several
key functions, DOM supplies carbon and nitrogen for
heterotrophic production (Keil and Kirchman 1991;
Wetzel 1992), mediates the availability of metals
(Yamashita and Jaffe 2008), and modifies the optical
properties of waters (Rodrıguez-Zuniga et al. 2008).
Therefore, it is of fundamental interest to understand
the effect of flow intermittency on DOM quantity and
composition. Little is known, however, on the dy-
namics of DOM before and after flow cessation.
Hydrological contraction and fragmentation causes
the disruption of the longitudinal and lateral hydro-
logical pathways (Sabater and Tockner 2010), which
results in low-oxygen and acidic in-stream conditions
(Boulton and Lake 1990; von Schiller et al. 2011). In
addition, contraction and fragmentation influences the
availability of dissolved inorganic nutrients (Dent and
Grimm 1999; von Schiller et al. 2011) and promotes the
accumulation of particulate organicmatter (Boulton and
Lake 1992; Acuna et al. 2005). During these hydro-
logical phases, allochthonous DOM input from the
catchment is interrupted, which may lead to low
dissolved organic carbon (DOC) and nitrogen (DON)
concentrations when there is no relevant autochthonous
DOM source (Freeman et a. 1994; Dahm et al. 2003).
Hydrological contraction and fragmentation may sti-
mulate biotic processes such as algal blooms and
microbial uptake (Lake 2003; Fellman et al. 2011). In
addition, abiotic processes that affect in-stream DOM
such as evaporation and reduction of the dilution
capacity (Sabater and Tockner 2010), leaching of
particulate organic matter (McMaster and Bond 2008),
photodegradation (Rodrıguez-Zuniga et al. 2008) and
adsorption–desorption (Dahm 1981) may be enhanced.
In general, allocthonous DOM dominates the bulk pool
of DOM, but the proportion of DOM deriving from in-
430 Biogeochemistry (2015) 123:429–446
123
Author's personal copy

streammicrobial production andmineralization tends to
increase, while the proportions of organic nitrogen,
polysaccharides and amino acids decrease (Ylla et al.
2010; Fellman et al. 2011; Vazquez et al. 2011).
Moreover, stream fragmentation into isolated pools
tends to increase DOC concentrations and DOM
biodegradability, while enhancing the spatial hetero-
geneity of DOM quantity and composition (Fellman
et al. 2011; Vazquez et al. 2011).
Hydrological expansion reestablishes hydrological
connectivity, thereby increasing the concentration of
allochthonous DOC and DON derived from the
leaching of sediments/soils and particulate organic
matter accumulated on the streambed and the hill
slopes during the dry phase (Jacobson et al. 2000;
Inamdar et al. 2011; Catalan et al. 2013). The DOM
pool upon rewetting is thus typically characterized by
high concentrations of labile high molecular weight
substances (Romanı et al. 2006; Vazquez et al. 2007;
Inamdar et al. 2011), with a high proportion of proteins
(Inamdar et al. 2011; Singh et al. 2014) and polysac-
charides (Ylla et al. 2010), and a low proportion of
aromatic and basic amino acids (Ylla et al. 2011). In
general, changes in stream DOM quantity and com-
position following dry periods are more pronounced
than those observed for other events (e.g. storms)
during the rest of the year (Bernal et al. 2005; Vazquez
et al. 2007; Inamdar et al. 2011). The quantity and
composition of DOM during rewetting events in
temporary streams may thus differ significantly from
that measured more routinely during storm-events in
perennial streams (Fellman et al. 2009; Nguyen et al.
2010; Pellerin et al. 2012).
To date, there has been little work on the dynamics
of DOM in temporary streams that consider both DOM
quantity and composition. Some studies have focused
on DOM dynamics upon rewetting at the start of the
expansion phase (e.g. Bernal et al. 2005; Vazquez
et al. 2007; Inamdar et al. 2011; Catalan et al. 2013).
Much less is known on the dynamics of DOM during
hydrological contraction and, especially, during hy-
drological fragmentation (but see e.g. Ylla et al. 2010;
Vazquez et al. 2011: Fellman et al. 2011). Moreover,
because of lack of temporal sampling and/or limited
sampling frequency, most of these studies have missed
the short-term dynamics and the transitions between
hydrological phases (i.e. contraction–fragmentation
and fragmentation–dry), which may represent biogeo-
chemical ‘‘hot moments’’ (sensu McClain et al. 2003).
The objective of this study was to examine the
effect of flow intermittency on the short-term dynam-
ics of stream DOM quantity and composition, with
special emphasis on the transitions between hydro-
logical phases. We combined intensive sampling with
a variety of analytical techniques to follow changes in
DOC and DON concentrations as well as in DOM size
fractions and spectroscopic properties in a temporary
Mediterranean stream during an entire contraction–
fragmentation–expansion hydrological cycle. Our fi-
nal goal was to gain information that may help us
develop better conceptual and mechanistic models of
DOM biogeochemistry in temporary streams, and thus
facilitate the sustainable management of these ecosys-
tems. Furthermore, our results may serve as a template
for understanding and managing the potential biogeo-
chemical response of stream ecosystems under future
water scarcity scenarios.
Methods
Study site
The Fuirosos stream drains a 15.2-km2 granitic
catchment in the Montnegre-Corredor Natural Pro-
tected Area (NE Iberian Peninsula). The climate is
semiarid Mediterranean, with monthly mean air
temperature ranging from 5 �C (January) to 24 �C(August) and a seasonally and inter-annually highly
variable precipitation (annual mean = 750 mm)
(Ninyerola et al. 2000). The catchment is mostly
forested and human land use is restricted to disperse
agricultural fields (\2 % of the catchment area).
Evergreen forests dominate at the lower parts of the
catchment, whereas deciduous forests are common at
higher elevations. Riparian vegetation is well devel-
oped, with a leaf input peak in autumn, although
intense summer hydric stress causes additional leaf
fall during the dry phase (Acuna et al. 2007).
Stream flow is seasonally intermittent, with a dry
phase during summer of variable duration and spatial
extent among years (Vazquez et al. 2013). The
concentration of DOC (annual mean *6 mg C L-1)
peaks during the transition from dry to wet conditions
in autumn, whereas the concentration of DON (annual
mean *0.2 mg N L-1) shows no clear seasonal
pattern (Bernal et al. 2005). At storm flow conditions,
DOC and DON concentrations tend to increase and the
Biogeochemistry (2015) 123:429–446 431
123
Author's personal copy

highest concentrations occur in autumn (mean
*8 mg C/L and *0.6 mg N/L; Bernal et al. 2005).
DOM composition is also highly responsive to
seasonal hydrological changes in this stream (Bernal
et al. 2005; Romanı et al. 2006; Vazquez et al. 2007;
Ylla et al. 2010; Vazquez et al. 2011).
For this study, we selected a 300-m long reach,
representative of the third-order section of the
Fuirosos stream (UTM coordinates at the center of
the reach = 31T 464934E 4616159 N; eleva-
tion = 123 m a.s.l.). The selected reach had a mod-
erate slope (0.063 m m-1) with alternance of pools
and riffles. Substrate was composed of sand (60 %)
and boulders (30 %), intermixed with patches of
cobbles, pebbles, gravel, and bedrock. The streambed
was mostly covered by biofilms, with an algal
community dominated by diatoms and cyanobacteria
(Tornes and Sabater 2010).
Field sampling
We sampled surfacewater at 3 to 4-day intervals (totally
24 dates) during the contraction, fragmentation and
expansion phases (see Fig. 1 for representative photos
of the different hydrological phases). To account for
spatial heterogeneity, we took the samples from 4
locations along the study reach (80, 160, 240 and 280 m
from the top of the study reach). Surface water was
present at all locations during the contraction phase (8
June–4 July; 8 sampling dates) and the expansion phase
(23 October–19 November; 9 sampling dates), while it
was absent from the location at 80 m on the last two
sampling dates of the fragmentation phase (5 July–23
July; 7 sampling dates). Sampling was carried out
between0900 h and1200 h tominimize the influence of
potential diel changes inDOM(Wilson andXenopoulos
2013).
We measured dissolved oxygen concentration
(DO), pH and water temperature at the mid-channel
area of each sampling location on each date using
WTW (Weilheim, Germany) hand-held probes. At the
same spots, we collected surface water using 100-mL
syringes (flushed three times prior to use), and filtered
it in the field through ashed Albet (Barcelona, Spain)
GF55 glass fiber filters (0.7 lm pore size) into pre-
washed plastic containers (1 sample per location). We
chose glass fiber filters because they are inert, allow
rapid filtration in the field and can be ashed. We stored
Fig. 1 Representative photos of the hydrological phases in the
Fuirosos stream (NE Iberian Peninsula) during the study period
a contraction phase (29 June 2014), b fragmentation phase (6
July 2014), c dry phase (22 August 2014), and expansion phase
(27 October 2014). Photos were taken at the sampling location
180 m
432 Biogeochemistry (2015) 123:429–446
123
personal copy

the samples on ice in the dark, transported them
immediately to the laboratory and froze them at
-20 �C. At the end of the samplings, we transported
all samples frozen to the Leibniz-Institute of Fresh-
water Ecology and Inland Fisheries (Berlin, Germany)
and analyzed them within 1 month. To minimize the
effects of freezing and thawing, all samples were
subject to only one freezing-thawing cycle, and
thawing was conducted in a refrigerator at 4 �C(Fellman et al. 2008; Hudson et al. 2009; Spencer et al.
2010).
We estimated discharge from measurements of
water column depth at the bottom of the study reach at
10-min intervals using a YSI (Ohio, USA) 600-OMS-
V2 multiparameter sonde. We constructed an em-
pirical depth-discharge relationship based on several
additions of a conservative tracer (i.e. chloride) during
the sampling period (Gordon et al. 2004).
Laboratory analyses
Size exclusion chromatography
We measured total DOC and DON concentrations and
characterized the DOM size fractions on a DOC-Labor
Huber (Karlsruhe, Germany) liquid chromatography–
organic carbon–organic nitrogen detection system
(LC-OCD-OND) (Huber et al. 2011; Graeber et al.
2012a). The LC-OCD-OND system was driven by a
Knauer (Berlin, Germany) S-100 HPLC pump and
consisted of a MLE (Dresden, Germany) autosampler
and a YSK HW (Toso, Japan) chromatographic
column. Each sample was measured after passing
and bypassing the chromatographic column with a
Knauer (Berlin, Germany) S-200 UV detector. Sub-
sequently, flow was further divided into two streams.
One stream went to a UV reactor to measure nitrogen
at 220 nm after oxidation to nitrate. The second stream
went to a thin-film reactor where DOCwas oxidized to
CO2 and then to an infrared CO2 detector. The mobile
phase used was a phosphate buffer of pH 6.85 (2.5 g
KH2PO4 ? 1.5 g Na2HPO4 9 2H2O to 1 L).
We calibrated the LC-OCD-OND system according
to the manufacturer’s instructions (DOC-Labor Huber,
Karlsruhe, Germany), and analyzed the DOM size
fractions using the software FIFFIKUS (DOC-Labor
Huber, Karlsruhe, Germany). Following Attermeyer
et al. (2014), we classified the DOM into three
fractions: (i) high molecular weight substances
(HMWS), including polysaccharides, (ii) humic or
humic-like substances (HS), including buildings
blocks, and (iii) low molecular weight substances
(LMWS), which summarize low molecular weight
acids and low molecular weight neutral substances.
We expressed each fraction as the percent contribution
to total DOC and total DON.While DOCwas detected
in all DOM size fractions, DON was only detected in
the HMWS and HS fractions (Graeber et al. 2012a).
Thus, the percent contribution to total DON and the
DOC:DON molar ratios were only reported for these
two fractions. Noteworthy, the LC-OCD-OND system
allowed the direct determination of DON, which is less
prone to errors than indirect DON calculations at high
dissolved inorganic nitrogen concentrations (Graeber
et al. 2012a), such as those typically found during the
end of the fragmentation phase and the start of the
expansion phase (von Schiller et al. 2011).
Spectroscopic measurements
We determined the specific UV absorbance (SUVA; L
mg-1m-1), a surrogate forDOMaromaticity (Weishaar
et al. 2003), directly on the LC-OCD-OND system
(Huber et al. 2011). To determine other spectroscopic
indices, we conducted absorbance measurements on a
Shimadzu (Kyoto, Japan) UV-2401 UV/VIS spec-
trophotometer and fluorescence measurements on a
Perkin-Elmer (Waltham, MA, USA) LS-50b fluores-
cence spectrometer. We measured excitation from 240
to 450 nm (5 nm steps) and emission from 300 to
600 nm (2 nm steps) with a slit width of 5 nm to
produce excitation-emission-matrices (EEMs; Stedmon
andMarkager 2005).Wemeasured the samples at room
temperature and corrected the absorbance spectra
(800–190 nm) for instrument baseline offset (Green
and Blough 1994). Daily measurements of the area
under the Raman peak for MilliQ water (Millipore,
Schwalbach, Germany) indicated instrument stability
(Lawaetz and Stedmon 2009). Fluorescence and ab-
sorbance readings were within the linear range of the
spectrometers and we accounted for primary and
secondary inner-filter effects by inner-filter correction
(Lakowicz 2006). Moreover, we corrected the spectra
for excitation by using the correction provided by the
manufacturer and for emission by using the BAM
fluorescence calibration kit (Pfeifer et al. 2006), and
normalized them by the area under the Raman peak at
350 nm excitation wavelength (Lawaetz and Stedmon
Biogeochemistry (2015) 123:429–446 433
123
Author's personal copy
2009). These corrections provide the best possible
comparability to other DOM fluorescence studies
(Lawaetz and Stedmon 2009).
From absorbance spectra we calculated 4 indices: a)
the ratio of absorptions at 250 and 365 nm (E2:E3),
with values ranging *4–8 (De Haan and De Boer
1987), (b) the spectral slope for 275–295 nm
(S275–295), with values ranging *0.01–0.05 (Helms
et al. 2008) (c) the spectral slope for 350–400 nm
(S350–400), with a similar range of values as S275–295(Helms et al. 2008), and d) the ratio of these slopes
(SR), with values ranging*0.5–3 (Helms et al. 2008).
The indices E2:E3, S275–295, S350–400 and SR have been
found to be inversely related to DOM molecular size;
thus, the higher the index value, the lower the
molecular size (De Haan and De Boer 1987; Peu-
ravuori and Pihlaja 1997; Helms et al. 2008). In
addition, S275–295 can be used as a tracer of photo-
bleaching (Helms et al. 2008) and of terrestrial DOC
(Fichot and Benner 2012).
FromEEMswecalculated 3 indices: (a) humification
index (HIX), forwhich values*1–2 are associatedwith
non-humified material and values[10 are typical for
fulvic acid extracts (Zsolnay et al. 1999), (b) fluores-
cence index (FI), for which values *1.3 suggest the
dominance of terrestrial higher-plant DOM sources and
values*1.8 suggest the dominance of microbial DOM
sources (McKnight et al. 2001), and (c) freshness index
(b:a), for which values[1 correspond to DOM freshly
released into water, whereas values *0.6–0.7 corre-
spond to lower DOM production in natural waters and
higher input of terrestrial origin (Parlanti et al. 2000;
Wilson and Xenopoulos 2009).
To further characterize changes in DOM fluores-
cence, we created a parallel factor analysis (PAR-
AFAC) model with the EEMs using the DOM Fluor
Toolbox (version 1.7; Stedmon and Bro 2008) for
Matlab (version R2009b, MathWorks, Ismaning,
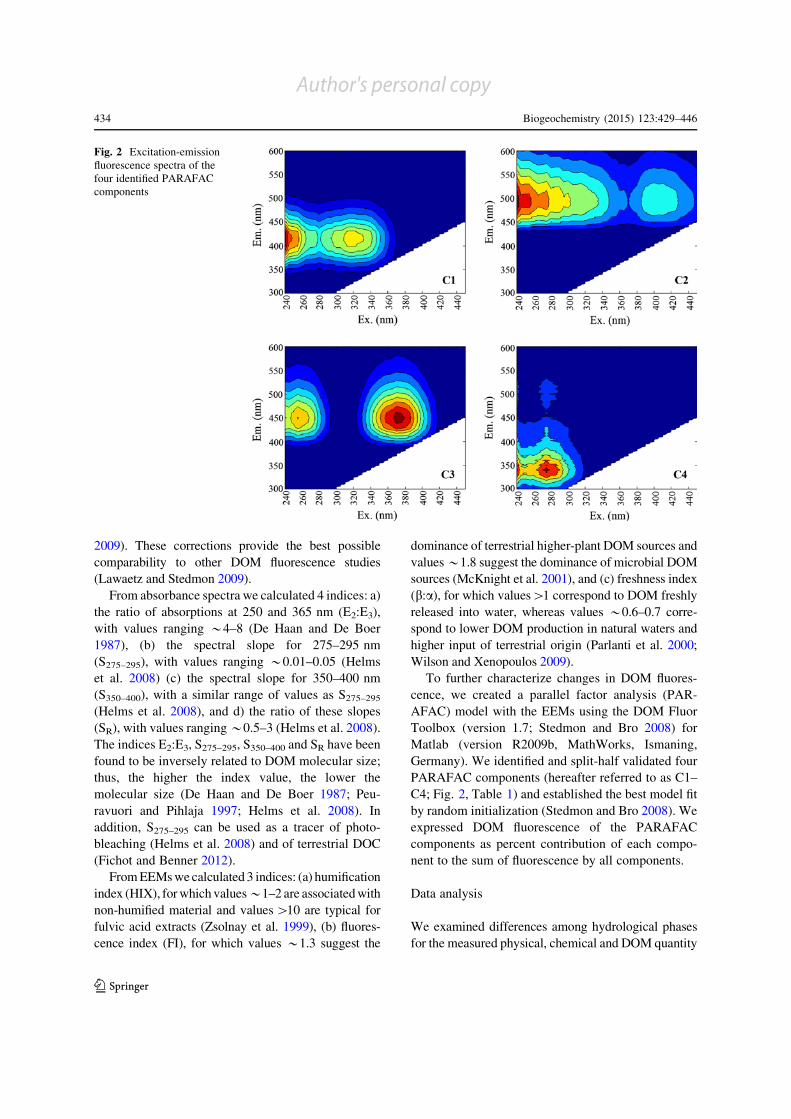
Germany). We identified and split-half validated four
PARAFAC components (hereafter referred to as C1–
C4; Fig. 2, Table 1) and established the best model fit
by random initialization (Stedmon and Bro 2008). We
expressed DOM fluorescence of the PARAFAC
components as percent contribution of each compo-
nent to the sum of fluorescence by all components.
Data analysis
We examined differences among hydrological phases
for the measured physical, chemical and DOM quantity
Fig. 2 Excitation-emission
fluorescence spectra of the
four identified PARAFAC
components
434 Biogeochemistry (2015) 123:429–446
123
Author's personal copy

and composition variables using non-parametric Krus-
kal–Wallis tests.Wegrouped themedianvalues for each
date by hydrological phase (i.e. contraction phase = 8
sampling dates, fragmentation phase = 7 sampling
dates, and expansion phase = 9 sampling dates). If the
testwas significant,we performed post hoc comparisons
of mean ranks of all pairs of groups using two-sided
significance levels with a Bonferroni adjustment. To
examine the temporal patterns of themeasured variables
under the different hydrological conditions,weexplored
the relationship between the median of these variables
along the reach and the sampling date using non-
parametric Spearman-rank correlations for each hydro-
logical phase separately. We ran all statistical analyses
with Statistica 6.1 (Statsoft, Tulsa, OK, USA). We
considered the statistical results significant if p\ 0.05.
Results
Physical and chemical variables
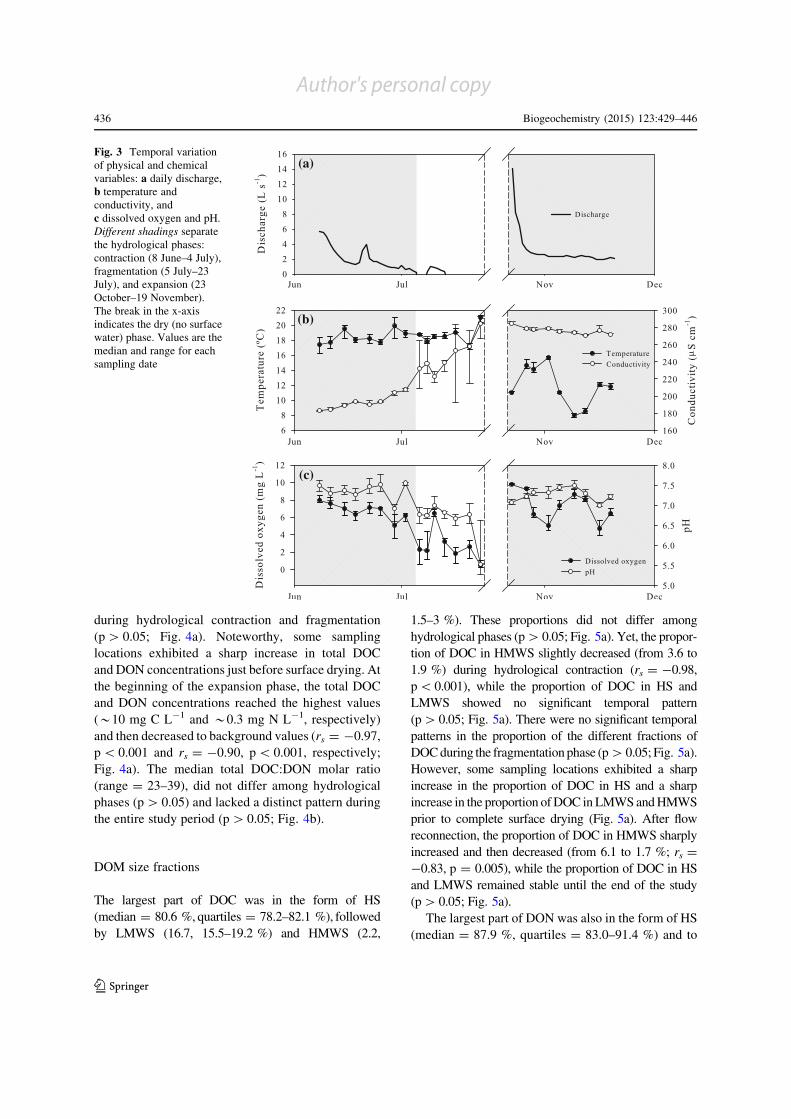
Stream discharge decreased during hydrological contrac-
tion (rs = -0.95, p\0.001; Fig. 3a). The subsequent
fragmentation phase was only interrupted by a short flow
pulse (10 July) that reconnected the isolated pools for
3 days. The study reach remained dry for 91 days until
stream flow reconnection occurred with a moderate
increase in discharge at the beginning of the expansion
phase (23 October). Subsequently, discharge decreased
until the endof the study (rs = -0.85, p\0.001; Fig. 3a).
Temperature, conductivity,DOandpHdifferedamong
hydrological phases (H[11.40, p\0.05; Fig. 3b, c).
Temperature and conductivity were lower and higher,
respectively, during hydrological expansion (post hoc,
p\0.01) while DO and pH were lower during the
fragmentation phase than during the other hydrological
phases (post hoc, p\0.01). Conductivity showed a clear
increase during hydrological contraction (rs = ? 0.93,
p\0.001), which continued during hydrological frag-
mentation (rs = ? 0.89, p = 0.007; Fig. 3b). Conversely,
temperature, DO and pH showed no clear temporal
patterns during these hydrological phases (p[0.05,
Fig. 3b, c). After flow reconnection, conductivity de-
creased (rs = -0.82, p = 0.007; Fig. 3b), while the rest
of physicochemical variables did not show a clear
temporal pattern (p[0.05; Fig. 3b, c).
Total DOM
Total DOC and DON concentrations did not differ
among hydrological phases (p[ 0.05; Fig. 4a). Total
DOC and DON concentrations remained low
(* 3 mg C L-1 and *0.1 mg N L-1, respectively)
Table 1 Emission (Em. max) and excitation (Ex. max) maxima, previous identifications and literature-based tentative interpretation
of the parallel factor analysis (PARAFAC) components
Component Em.
max.
(nm)
Ex. max. (nm) Similar components
identified in previous
studies
Tentative interpretation of components
C1 416 \240 (315) C1c, C3d, C2e Humic-acid fluorophore; aromatic, terrestrial originc; found in
marine DOM; positively related to forage crope, arable
farming and wetlands in small catchmentsd
C2 490 \240 (400) C2d Fulvic acid fluorophored; highly conjugated, terrestrial origind
C3 450 370 (255,\240) SQ2a, C4c, C4d Semi-quinone/fulvic acid fluorophorea,d; terrestrial origind,
microbially transformeda, high aromaticity, reduced statea,
positively related to arable farming in small catchmentsc
C4 340 \240 (275) C8a, C8b, C5c, C7d,
C5eTryptophan-like fluorophorea, b, d; probably derived from
aquatic microbial productiond, high bioavailability, not
humifiedb, positively related to forest in small catchmentsc
The values in the Ex. max column indicate primary and secondary (with brackets) peaksa Cory and McKnight (2005)b Fellman et al. (2009)c Graeber et al. (2012b)d Stedmon and Markager (2005)e Williams et al. (2010)
Biogeochemistry (2015) 123:429–446 435
123
Author's personal copy
during hydrological contraction and fragmentation
(p[ 0.05; Fig. 4a). Noteworthy, some sampling
locations exhibited a sharp increase in total DOC
and DON concentrations just before surface drying. At
the beginning of the expansion phase, the total DOC
and DON concentrations reached the highest values
(*10 mg C L-1 and *0.3 mg N L-1, respectively)
and then decreased to background values (rs = -0.97,
p\ 0.001 and rs = -0.90, p\ 0.001, respectively;
Fig. 4a). The median total DOC:DON molar ratio
(range = 23–39), did not differ among hydrological
phases (p[ 0.05) and lacked a distinct pattern during
the entire study period (p[ 0.05; Fig. 4b).
DOM size fractions
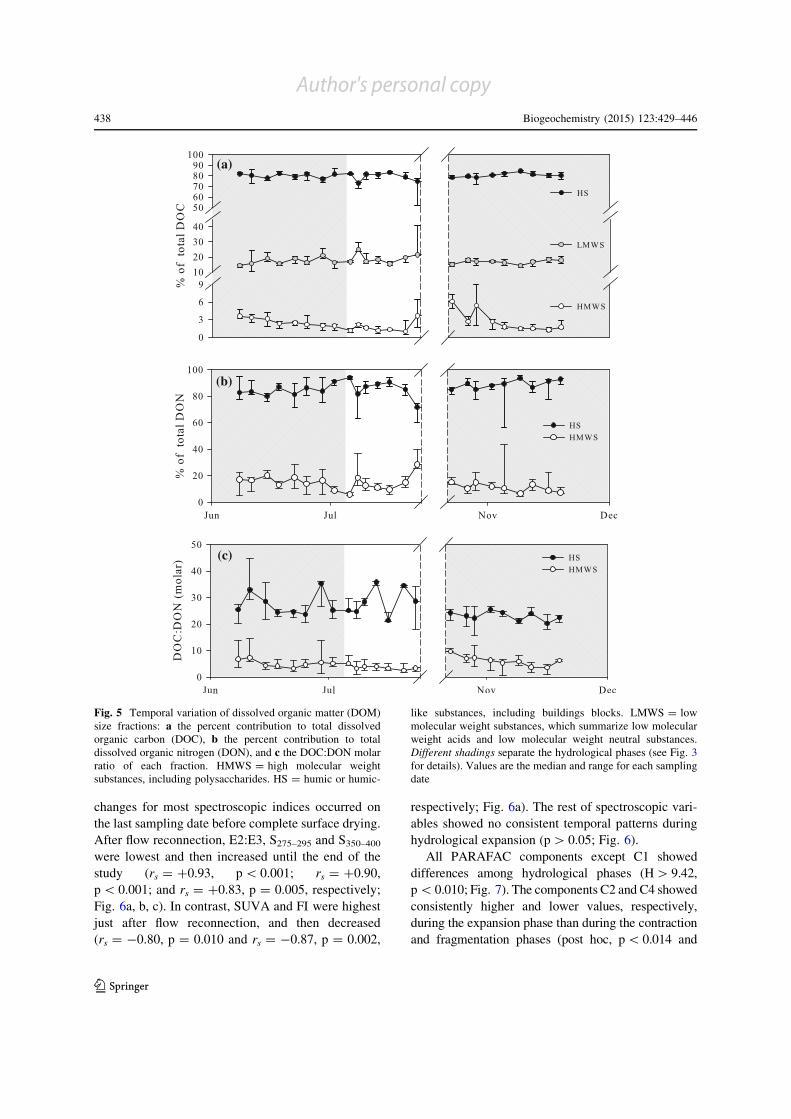
The largest part of DOC was in the form of HS
(median = 80.6 %, quartiles = 78.2–82.1 %), followed
by LMWS (16.7, 15.5–19.2 %) and HMWS (2.2,
1.5–3 %). These proportions did not differ among
hydrological phases (p[0.05; Fig. 5a). Yet, the propor-
tion of DOC in HMWS slightly decreased (from 3.6 to
1.9 %) during hydrological contraction (rs = -0.98,
p\0.001), while the proportion of DOC in HS and
LMWS showed no significant temporal pattern
(p[0.05; Fig. 5a). There were no significant temporal
patterns in the proportion of the different fractions of
DOCduring the fragmentationphase (p[0.05; Fig. 5a).
However, some sampling locations exhibited a sharp
increase in the proportion of DOC in HS and a sharp
increase in the proportion ofDOC inLMWSandHMWS
prior to complete surface drying (Fig. 5a). After flow
reconnection, the proportion of DOC in HMWS sharply
increased and then decreased (from 6.1 to 1.7 %; rs =
-0.83, p = 0.005), while the proportion of DOC in HS
and LMWS remained stable until the end of the study
(p[0.05; Fig. 5a).
The largest part of DON was also in the form of HS
(median = 87.9 %, quartiles = 83.0–91.4 %) and to
(a)
(b)
(c)
Fig. 3 Temporal variation
of physical and chemical
variables: a daily discharge,
b temperature and
conductivity, and
c dissolved oxygen and pH.
Different shadings separate
the hydrological phases:
contraction (8 June–4 July),
fragmentation (5 July–23
July), and expansion (23
October–19 November).
The break in the x-axis
indicates the dry (no surface
water) phase. Values are the
median and range for each
sampling date
436 Biogeochemistry (2015) 123:429–446
123
Author's personal copy

a lesser extent in the form of HMWS (12.1,
8.6–17.0 %). The proportion of DON in HS and
HMWS did not differ among hydrological phases
(p[ 0.05) and did not show any clear temporal pattern
during any of the investigated hydrological phases
(p[ 0.05). Noteworthy, in all sampling locations
there was a decrease in the proportion of DON in HS
and an increase in the proportion of DON in HMWS
before complete surface drying (Fig. 5b).
The DOC:DON ratio in HS was similar to the
DOC:DON ratio of total DOM and higher than the
DOC:DON ratio in HMWS (Fig. 5c). The DOC:DON
ratios in HS and HMWS differed among hydrological
phases (H = 9.11, p = 0.011 and H = 9.07,
p = 0.011, respectively). For HS, the DOC:DON
ratio was lower during the expansion phase than
during the other hydrological phases (post hoc,
p\ 0.037). For HMWS, the DOC:DON ratio was
higher during the expansion phase than during the
fragmentation phase (post hoc, p = 0.009), but there
were no differences between the contraction phase and
the other hydrological phases (post hoc, p[ 0.05).The
DOC:DON ratio in HS did not show any clear
temporal pattern throughout the study period
(p[ 0.05; Fig. 5c). Similarly, the DOC:DON ratio
in HMWS did not show any clear temporal pattern
during hydrological contraction and fragmentation
(p[ 0.05); however, it sharply increased and then
gradually decreased during hydrological expansion
(rs = -0.80, p = 0.010; Fig. 5c).
DOM spectroscopic properties
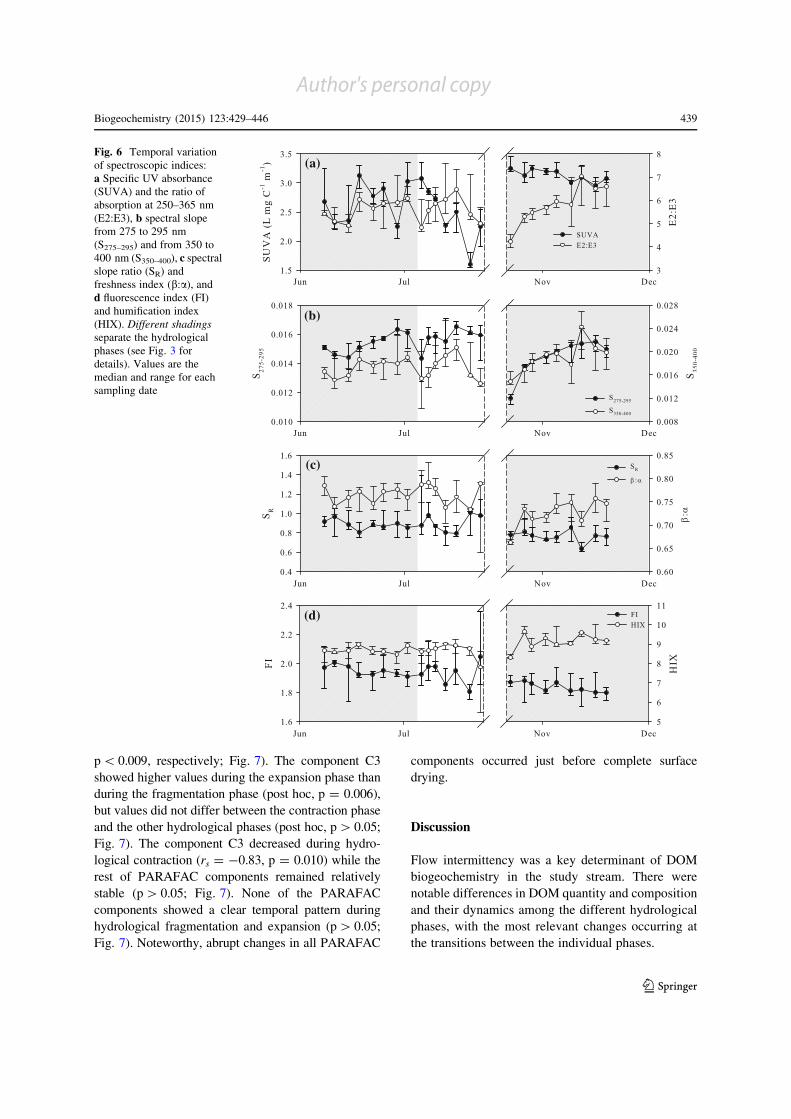
Most spectroscopic indices differed among hydro-
logical phases (H[ 7.90, p\ 0.020; Fig. 6). SUVA
was higher and SR, FI and b:a were lower during the
expansion phase than during the other hydrological
phases (post hoc, p\ 0.022, p\ 0.007, p\ 0.033,
p\ 0.042, respectively; Fig. 6). S275–295 was lower
during the expansion phase than during the fragmen-
tation phase (post hoc, p = 0.019), but there were no
differences between the contraction phase and the
other hydrological phases (post hoc, p[ 0.05;
Fig. 6b). HIX was higher during the expansion phase
than during the contraction phase (post hoc,
p = 0.019), but there were no differences between
the fragmentation phase and the other hydrological
phases (post hoc, p[ 0.05; Fig. 6d). During hydro-
logical contraction, E2:E3, S275–295 and S350–400increased (rs = ?0.76, p = 0.028; rs = ?0.88,
p = 0.004; and rs = ?0.76, p = 0.028, respectively;
Fig. 6 a, b). All other spectroscopic indices showed no
clear temporal pattern during this hydrological phase
(p[ 0.05; Fig. 6). Among all spectroscopic indices,
only SUVA decreased during hydrological fragmen-
tation (rs = -0.93, p = 0.003; Fig. 6a). Yet, abrupt
(a)
(b)
Fig. 4 Temporal variation
of total dissolved organic
matter (DOM)
concentrations: a Dissolved
organic carbon (DOC) and
dissolved organic nitrogen
(DON), b DOC:DON molar
ratio. Different shadings
separate the hydrological
phases (see Fig. 3 for
details). Values are the
median and range for each
sampling date
Biogeochemistry (2015) 123:429–446 437
123
Author's personal copy
changes for most spectroscopic indices occurred on
the last sampling date before complete surface drying.
After flow reconnection, E2:E3, S275–295 and S350–400were lowest and then increased until the end of the
study (rs = ?0.93, p\ 0.001; rs = ?0.90,
p\ 0.001; and rs = ?0.83, p = 0.005, respectively;
Fig. 6a, b, c). In contrast, SUVA and FI were highest
just after flow reconnection, and then decreased
(rs = -0.80, p = 0.010 and rs = -0.87, p = 0.002,
respectively; Fig. 6a). The rest of spectroscopic vari-
ables showed no consistent temporal patterns during
hydrological expansion (p[ 0.05; Fig. 6).
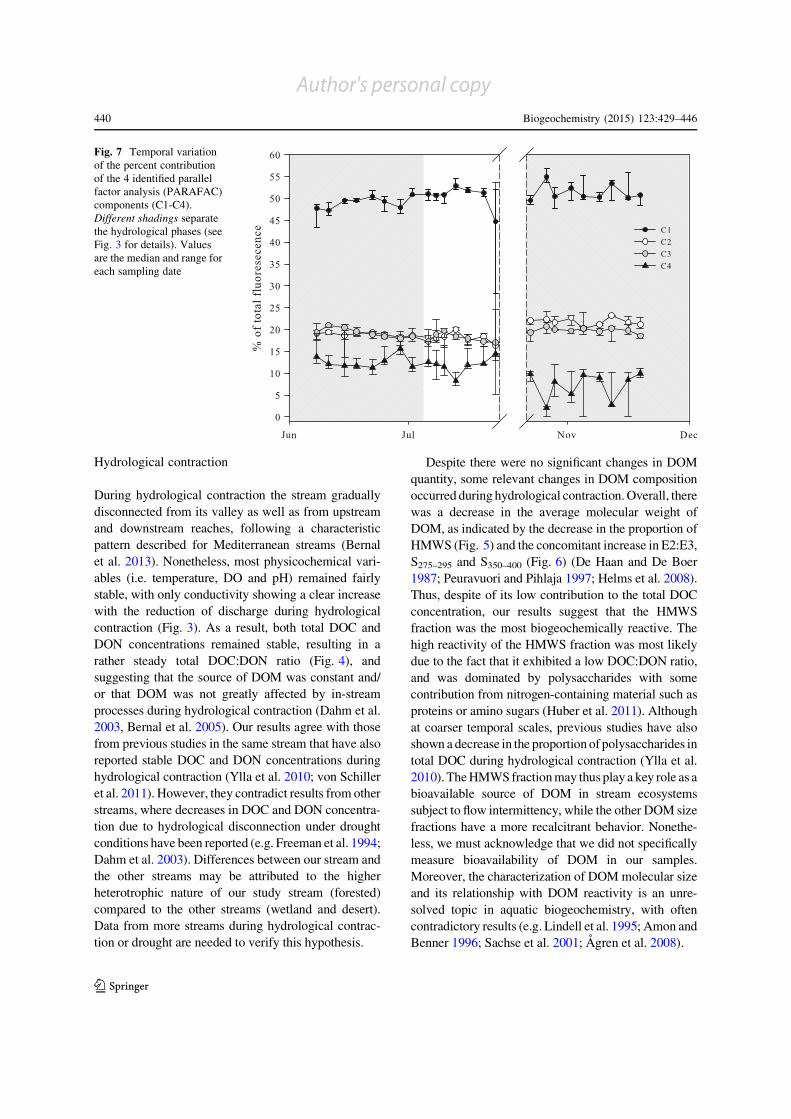
All PARAFAC components except C1 showed
differences among hydrological phases (H[ 9.42,
p\ 0.010; Fig. 7). The components C2 and C4 showed
consistently higher and lower values, respectively,
during the expansion phase than during the contraction
and fragmentation phases (post hoc, p\ 0.014 and
(a)
(b)
(c)
Fig. 5 Temporal variation of dissolved organic matter (DOM)
size fractions: a the percent contribution to total dissolved
organic carbon (DOC), b the percent contribution to total
dissolved organic nitrogen (DON), and c the DOC:DON molar
ratio of each fraction. HMWS = high molecular weight
substances, including polysaccharides. HS = humic or humic-
like substances, including buildings blocks. LMWS = low
molecular weight substances, which summarize low molecular
weight acids and low molecular weight neutral substances.
Different shadings separate the hydrological phases (see Fig. 3
for details). Values are the median and range for each sampling
date
438 Biogeochemistry (2015) 123:429–446
123
Author's personal copy
p\ 0.009, respectively; Fig. 7). The component C3
showed higher values during the expansion phase than
during the fragmentation phase (post hoc, p = 0.006),
but values did not differ between the contraction phase
and the other hydrological phases (post hoc, p[ 0.05;
Fig. 7). The component C3 decreased during hydro-
logical contraction (rs = -0.83, p = 0.010) while the
rest of PARAFAC components remained relatively
stable (p[ 0.05; Fig. 7). None of the PARAFAC
components showed a clear temporal pattern during
hydrological fragmentation and expansion (p[ 0.05;
Fig. 7). Noteworthy, abrupt changes in all PARAFAC
components occurred just before complete surface
drying.
Discussion
Flow intermittency was a key determinant of DOM
biogeochemistry in the study stream. There were
notable differences in DOM quantity and composition
and their dynamics among the different hydrological
phases, with the most relevant changes occurring at
the transitions between the individual phases.
(a)
(b)
(c)
(d)
Fig. 6 Temporal variation
of spectroscopic indices:
a Specific UV absorbance
(SUVA) and the ratio of
absorption at 250–365 nm
(E2:E3), b spectral slope
from 275 to 295 nm
(S275–295) and from 350 to
400 nm (S350–400), c spectralslope ratio (SR) and
freshness index (b:a), andd fluorescence index (FI)
and humification index
(HIX). Different shadings
separate the hydrological
phases (see Fig. 3 for
details). Values are the
median and range for each
sampling date
Biogeochemistry (2015) 123:429–446 439
123
Author's personal copy
Hydrological contraction
During hydrological contraction the stream gradually
disconnected from its valley as well as from upstream
and downstream reaches, following a characteristic
pattern described for Mediterranean streams (Bernal
et al. 2013). Nonetheless, most physicochemical vari-
ables (i.e. temperature, DO and pH) remained fairly
stable, with only conductivity showing a clear increase
with the reduction of discharge during hydrological
contraction (Fig. 3). As a result, both total DOC and
DON concentrations remained stable, resulting in a
rather steady total DOC:DON ratio (Fig. 4), and
suggesting that the source of DOM was constant and/
or that DOM was not greatly affected by in-stream
processes during hydrological contraction (Dahm et al.
2003, Bernal et al. 2005). Our results agree with those
from previous studies in the same stream that have also
reported stable DOC and DON concentrations during
hydrological contraction (Ylla et al. 2010; von Schiller
et al. 2011). However, they contradict results from other
streams, where decreases in DOC and DON concentra-
tion due to hydrological disconnection under drought
conditions have been reported (e.g. Freeman et al. 1994;
Dahm et al. 2003). Differences between our stream and
the other streams may be attributed to the higher
heterotrophic nature of our study stream (forested)
compared to the other streams (wetland and desert).
Data from more streams during hydrological contrac-
tion or drought are needed to verify this hypothesis.
Despite there were no significant changes in DOM
quantity, some relevant changes in DOM composition
occurred during hydrological contraction.Overall, there
was a decrease in the average molecular weight of
DOM, as indicated by the decrease in the proportion of
HMWS (Fig. 5) and the concomitant increase in E2:E3,
S275–295 and S350–400 (Fig. 6) (De Haan and De Boer
1987; Peuravuori and Pihlaja 1997; Helms et al. 2008).
Thus, despite of its low contribution to the total DOC
concentration, our results suggest that the HMWS
fraction was the most biogeochemically reactive. The
high reactivity of the HMWS fraction was most likely
due to the fact that it exhibited a low DOC:DON ratio,
and was dominated by polysaccharides with some
contribution from nitrogen-containing material such as
proteins or amino sugars (Huber et al. 2011). Although
at coarser temporal scales, previous studies have also
shown a decrease in the proportion of polysaccharides in
total DOC during hydrological contraction (Ylla et al.
2010). TheHMWSfractionmay thus play a key role as a
bioavailable source of DOM in stream ecosystems
subject to flow intermittency, while the other DOM size
fractions have a more recalcitrant behavior. Nonethe-
less, we must acknowledge that we did not specifically
measure bioavailability of DOM in our samples.
Moreover, the characterization of DOMmolecular size
and its relationship with DOM reactivity is an unre-
solved topic in aquatic biogeochemistry, with often
contradictory results (e.g. Lindell et al. 1995; Amon and
Benner 1996; Sachse et al. 2001; Agren et al. 2008).
Fig. 7 Temporal variation
of the percent contribution
of the 4 identified parallel
factor analysis (PARAFAC)
components (C1-C4).
Different shadings separate
the hydrological phases (see
Fig. 3 for details). Values
are the median and range for
each sampling date
440 Biogeochemistry (2015) 123:429–446
123
Author's personal copy

Hydrological fragmentation
In the hydrological transition between the contraction
and fragmentation phases, stream flowwas interrupted
and surface water got restricted to isolated pools
(Bernal et al. 2013). As a result, there was an abrupt
but predictable increase in acidic and low oxygen in-
stream conditions which remained for the rest of the
fragmentation phase (Boulton and Lake 1990; von
Schiller et al. 2011). Conductivity continued to
increase during the fragmentation phase, likely driven
by an increase in solute concentration through
evaporation (Sabater and Tockner 2010). Tem-
perature, however, remained rather constant probably
regulated by the shading effect of the well-developed
riparian canopy (Johnson 2004).
The significant shift in physicochemical conditions
between the contraction and the fragmentation phase
was not followed by relevant changes in DOM
quantity or composition. Total DOC and DON con-
centrations (Fig. 4), DOM size fractions (Fig. 5) and
spectroscopic properties (Fig. 6 and 7) remained
relatively constant with respect to the contraction
phase. Furthermore, among all DOM composition
variables, only the SUVA index showed a clear
temporal pattern during hydrological fragmentation
(Fig. 6). The negative relationship between SUVA
and sampling time indicated that stream DOM became
less aromatic with time since pool isolation (Weishaar
et al. 2003). These short-term temporal data support
the finding by Vazquez et al. (2011) in the same
stream, who found decreasing SUVA across a spatial
gradient of pools of increasing age. Vazquez et al.
(2011) attributed the decrease in SUVA to an increase
in the contribution of in-stream algal and microbial
processes to the total DOM pool with increasing pool
isolation time. We did not find correlations between
pool isolation time and any spectroscopic property
other than SUVA. Nonetheless, similar to Fellman
et al. (2011), who found higher protein-like fluores-
cence in highly evaporated pools in a subtropical
dryland river, we also found an abrupt increase in
protein-like fluorescence at the end of the fragmenta-
tion phase (see below).
Abrupt changes in DOM quantity and composition
occurred in most sampling locations at the end of the
fragmentation phase, just before the stream complete-
ly dried up at the surface. Both total DOC and DON
concentrations sharply increased (Fig. 4), while the
proportion of HS decreased and the proportion of
HMWS and LMWS increased (Fig. 5). These changes
indicate a major shift in the bulk composition of DOM
towards a higher proportion of non-humic substances
such as polysaccharides and low molecular weight
alcohols, aldehydes, ketones, sugars and amino acids
(Huber et al. 2011). In addition, the spectroscopic
indices indicated that the released DOM had a fresh
(high b:a; Wilson and Xenopoulos 2009), microbial
(high FI, McKnight et al. 2001), and non-humic (low
HIX; Zsolnay et al. 1999) character (Fig. 6). Further-
more, changes in the proportions of the PARAFAC
components (i.e. low C1 and high C4; Fig. 7) indicat-
ed that the DOM shifted from a more humic-acid like,
aromatic and terrestrial character to a more trypto-
phan-like, non-humified, bioavailable and aquatic
character (Stedmon and Markager 2005; Williams
et al. 2010; Graeber et al. 2012b). Together, these
results support the idea that the increase in DOC and
DON concentrations before complete surface drying
was most likely due to an abrupt microbial biofilm cell
lysis and/or DOM exudation under stress conditions
(e.g. high temperature, low DO, low pH), which could
not be mineralized (Humphries and Baldwin 2003;
Schimel et al. 2007; Timoner et al. 2012). This
biogeochemical hot moment (sensu McClain et al.
2003) at the end of the fragmentation phase has not
been captured by previous studies of DOM dynamics
during stream drying (e.g. Ylla et al. 2010; Fellman
et al. 2011; Vazquez et al. 2011), probably because
they covered coarser temporal scales than the present
study. Our results support the idea that hydrological
fragmentation, especially in its later stages, can
strongly enhance changes in stream DOM quantity
and composition, thereby increasing the spatial
heterogeneity of DOM and nutrient availability along
temporary stream networks (Dent and Grimm 1999;
Gomez et al. 2009; von Schiller et al. 2011; Vazquez
et al. 2011; Fellman et al. 2011).
Hydrological expansion
Upon rewetting, the lateral and longitudinal hydro-
logical connections along the stream channel were re-
established (Bernal et al. 2013). Flow reconnection
favored the release of high amounts of total DOC and
DON (Fig. 4), in line with previous studies in the same
stream (Bernal et al. 2005; Vazquez et al. 2007; Ylla
et al. 2010) and temporary streams elsewhere
Biogeochemistry (2015) 123:429–446 441
123
Author's personal copy

(Inamdar et al. 2011; Catalan et al. 2013). These
results are also consistent with other studies performed
in a wide range of perennial streams during storm
events (Fellman et al. 2009; Nguyen et al. 2010;
Pellerin et al. 2012). Yet, we must acknowledge that
even if we sampled the stream on the first day upon
rewetting, we did not capture the rising limb of the
hydrograph; thus, our samples may not be fully
representative of the DOM that gets released just after
flow reconnection (Nguyen et al. 2010; Inamdar et al.
2011). Nonetheless, the rewetting showed a relatively
low peak discharge with DOC values similar to those
reported in other investigated post-drought rewetting
events in the same stream (Bernal et al. 2005; Romanı
et al. 2006; Vazquez et al. 2007).
In parallel to the increase in DOC and DON
concentrations upon rewetting, there was an increase
in the proportion of the HMWS fraction of DOC and
DON (Fig. 5), supported by the observation of low
E2:E3 and S275–295 values (Fig. 6), indicative of high
average molecular weight (De Haan and De Boer
1987; Helms et al. 2008). This observation is in line
with previous studies in the same stream that have
reported an increase in the proportion of HMWS after
flow reconnection (Romanı et al. 2006; Vazquez et al.
2007), most likely associated with an increase in the
relative amount of polysaccharides at rewetting (Ylla
et al. 2010). Other studies using DOM spectroscopy
have reported similar increases in the proportion of
HMWS at rewetting for a temporary stream in North
America (Inamdar et al. 2011) and for perennial
streams from several bioclimatic regions during storm
events (Nguyen et al. 2010; Spencer et al. 2010).
As indicated by other spectroscopic indices
(Fig. 6), DOM just upon rewetting was also charac-
terized by high aromaticity (high SUVA; Weishaar
et al. 2003), terrestrial origin (low S275–295; Fichot and
Benner 2012) and old source (low b:a; Wilson and
Xenopoulos 2009). Furthermore, the higher and lower
values of the PARAFAC components C2 and C4,
respectively, indicated the preponderance of a more
humic-like DOM of terrestrial origin (Fig. 7; Table 1;
Cory and McKnight 2005; Stedmon and Markager
2005). Previous studies of storm-events during post-
drought periods in temporary streams (Catalan et al.
2013) and in perennial streams (Fellman et al. 2009;
Nguyen et al. 2010) have also reported increases of
aromatic and humic DOM of terrestrial origin,
supported primarily by spectroscopic data. In contrast,
an increase in protein-like fluorescence during the
first-flush event after drought was found in a tempo-
rary stream in North America (Inamdar et al. 2011).
The authors attributed this increase to the breakdown
and production of labile organic matter in the dry
stream sediments and riparian soils during drought
(Inamdar et al. 2011). In absence of a definitive
explanation, we hypothesize that the high accumula-
tion of leaves on the streambed and riparian soils of the
Fuirosos stream in summer due to hydric stress (Acuna
et al. 2007), which was most likely not as pronounced
in the North American stream (Inamdar et al. 2011),
may explain the observed difference between the two
streams. Therefore, the timing of flow reconnection
with respect to leaf fall may be an important factor in
determining the composition of the first-flush DOM in
temporary streams.
Overall, DOM composition results confirm previ-
ous observations in the same stream that attribute the
typical increase in DOC and DON concentrations
upon rewetting to leaching of accumulated detritus on
the streambed and hill slopes (Romanı et al. 2006;
Vazquez et al. 2007; Artigas et al. 2009). Nonetheless,
the leaching of dry sediments and near-stream soils
could represent an additional DOM source upon
rewetting (Inamdar et al. 2011). Interestingly, the
plant- and sediment/soil-derived DOM observed at the
beginning of the expansion phase is probably highly
labile (McDowell 1985; McArthur and Richardson
2002) and can be rapidly used by stream microorgan-
isms that show a rapid recovery of their activity after
rewetting (Romanı et al. 2006; Artigas et al. 2009;
Timoner et al. 2012).
After the initial increase at the beginning of the
expansion phase, the total DOC and DON concentra-
tions concurrently decreased and the total DOC:DON
molar ratio remained constant (Fig. 4), indicating no
major changes in the bulk DOM source (Bernal et al.
2005). Interestingly, this temporal pattern for DOC
and DON concentrations was similar to that observed
by Inamdar et al. (2011) but unlike previous observa-
tions in the same stream (Bernal et al. 2005; Ylla et al.
2010), where it was reported that DOC concentrations
followed a different pattern than DON concentrations
during hydrological expansion. We do not have a
definitive explanation for these differences in DOC
and DON patterns during hydrological expansion;
however, we suspect that it may be due to the
technique used to measure DON concentration. We
442 Biogeochemistry (2015) 123:429–446
123
Author's personal copy

determined the DON concentration directly with the
LC-OCD-OND system, which is more accurate than
the indirect DON determination technique used in
previous studies for measuring DON concentration
(Graeber et al. 2012a), especially at high dissolved
inorganic nitrogen concentrations such as those found
upon rewetting (Von Schiller et al. 2011).
In parallel to the decrease in total DOC and DON
concentrations, relevant changes in DOM composition
occurred during the expansion phase. Overall, there was
a decrease in the average molecular weight of DOM, as
indicated by the decrease in the proportion of HMWS
(Fig. 5) and the concomitant increase in E2:E3, S275–295and S350–400 (Fig. 6) (De Haan and De Boer 1987;
Peuravuori and Pihlaja 1997; Helms et al. 2008).
Simultaneously, there was a slight decrease of the
DOC:DONratio inHMWS(Fig. 5). These observations
support again the biogeochemical relevance of the
HMWS fraction, despite of its low contribution to the
total DOC concentration. The rest of DOMcomposition
variables did not show clear temporal patterns during
hydrological expansion, except for the decrease of
SUVA values (Fig. 6). This gradual reduction in stream
DOM aromaticity during hydrological expansion
(Weishaar et al. 2003)maybe at least partially explained
by a decrease in DOM leaching from the streambed and
hill slopes, but also by higher microbial processing of
DOM by recovering stream biofilms favored by
continuous flow conditions (Ylla et al. 2010; Timoner
et al. 2012). Interestingly, while some DOM composi-
tion variables recovered to values similar to those at the
contraction phase, other variables remained consistently
higher (e.g. SUVA,HIX,C2) or lower (e.g. SR, FI,HIX,
C4), indicating the prevalence of a humic-like DOM of
terrestrial origin during the whole expansion phase.
Conclusions
By applying an intensive sampling design, in combi-
nation with chromatographic and spectroscopic tech-
niques, this study shows how changes in hydrological
connectivity driven by seasonal stream flow intermit-
tency affect the dynamics of DOM quantity and
composition in a temporary Mediterranean stream.
The most relevant changes occurred at the transitions
between hydrological phases, thus highlighting the
importance of hydrological transitions for DOM
specifically, and biogeochemical processes generally,
in temporary streams. Despite the timely- and spatial-
ly-restricted nature of this study, our results suggest
that flow intermittency is a key determinant of DOM
dynamics in temporary streams, with relevant impli-
cations for the sustainable management and regulation
of these ecosystems. Changes in the quantity and
composition of DOM in surface waters have important
consequences for water quality, andmay thus affect the
ecological integrity of stream ecosystems and thewater
supply for human use. Results from this study will help
us develop better conceptual and mechanistic models
of DOM biogeochemistry in temporary streams, and
thus facilitate the sustainable management of these
ecosystems. Because DOM plays an essential role in
aquatic ecosystem biogeochemistry, while many per-
manent streams are increasingly becoming temporary;
our results may serve as a template for understanding
and managing the potential biogeochemical response
of stream ecosystems under future water scarcity
scenarios. Further research on the effects of flow
intermittency on DOM dynamics across different
geographical, hydrological and ecological settings is
needed to confirm the generality of our findings.
Acknowledgments We thank J. Rodrıguez, L. Proia, M.
Peipoch and A. Blesa for field assistance, and E. Zwirnmann, H.
J. Exner, A. Luder, S. Schell and H. Magnussen for laboratory
analyses. We are also grateful to the direction of the Montnegre-
Corredor Natural Park (Diputacio de Barcelona) for allowing
access to the sampling site. This studywas fundedby theEuropean
Union through the MIRAGE project (FP7 ENV 2007 1).
Additional funds were provided by the Spanish Ministry of
Economy and Competitiveness through the Consolider-Ingenio
projects SCARCE (CSD2009-00065) and GRACCIE (CSD2007-
00067). D. von Schiller was supported by a DAAD-‘‘laCaixa’’
fellowship and a ‘‘Juan de la Cierva’’ postdoctoral grant (JCI-
2010-06397).
References
Acuna V, Munoz I, Giorgi A et al (2005) Drought and post-
drought recovery cycles in an intermittent Mediterranean
stream: structural and functional aspects. J North Am
Benthol Soc 24:919–933
Acuna V, Giorgi A, Munoz I et al (2007) Meteorological and
riparian influences on organic matter dynamics in a
forested Mediterranean stream. J North Am Benthol Soc
26:54–69
Acuna V, Datry T, Marshall J et al (2014) Why should we care
about temporary waterways? Science 343:1080–1081
Agren A, Berggren M, Laudon H, Jansson M (2008) Terrestrial
export of highly bioavailable carbon from small boreal
catchments in spring floods. Freshw Biol 53:964–972
Biogeochemistry (2015) 123:429–446 443
123
Author's personal copy

Amon RMW, Benner R (1996) Bacterial utilization of different
size classes of dissolved organic matter. Limnol Oceanogr
41:41–51
Artigas J, Romani AM, Gaudes A et al (2009) Organic matter
availability structures microbial biomass and activity in a
Mediterranean stream. Freshw Biol 54:2025–2036
AttermeyerK,HornickT,KaylerZEet al (2014)Enhancedbacterial
decomposition with increasing addition of autochthonous to
allochthonous carbon without any effect on bacterial commu-
nity composition. Biogeosciences 11:1479–1489
Bernal S, Butturini A, Sabater F (2005) Seasonal variations of
dissolved nitrogen and DOC:DON ratios in an intermittent
Mediterranean stream. Biogeochemistry 75:351–372
Bernal S, von Schiller D, Sabater F, Martı E (2013) Hydro-
logical extremes modulate nutrient dynamics in mediter-
ranean climate streams across different spatial scales.
Hydrobiologia 719:31–42
Bonada N, Resh VH (2013) Mediterranean-climate streams and
rivers: geographically separated but ecologically compa-
rable freshwater systems. Hydrobiologia 719:1–29
Boulton AJ, Lake PS (1990) The ecology of two intermittent
streams in Victoria, Australia. I. Multivariate analyses of
physicochemical features. Freshw Biol 24:123–141
Boulton AJ, Lake PS (1992) Benthic organic matter and detri-
tivorous macroinvertebrates in two intermittent streams in
south-eastern Australia. Hydrobiologia 241:107–118
Catalan N, Obrador B, Alomar C, Pretus JL (2013) Seasonality
and landscape factors drive dissolved organic matter
properties in Mediterranean ephemeral washes. Biogeo-
chemistry 112:261–274
Cory RM, McKnight DM (2005) Fluorescence spectroscopy
reveals ubiquitous presence of oxidized and reduced qui-
nones in dissolved organic matter. Environ Sci Technol
39:8142–8149
Dahm CN (1981) Pathways and mechanisms for removal of
dissolved organic carbon from leaf leachate in streams.
Can J Fish Aquat Sci 38:68–76
Dahm CN, Baker MA, Moore DI, Thibault JR (2003) Coupled
biogeochemical and hydrological responses of streams and
rivers to drought. Freshw Biol 48:1219–1231
Datry T, Larned ST, Tockner K (2014) Intermittent rivers: a
challenge for freshwater ecology. Bioscience 64:229–235
De Haan H, De Boer T (1987) Applicability of light absorbance
and fluorescence as measures of concentration and mole-
cular size of dissolved organic carbon in humic Lake
Tjeukemeer. Water Res 21:731–734
Dent CL, Grimm NB (1999) Spatial heterogeneity of stream
water nutrient concentrations over successional time.
Ecology 80:2283–2298
Fellman JB, D’Amore DV, Hood E (2008) An evaluation of
freezing as a preservation technique for analyzing dis-
solved organic C, N and P in surface water samples. Sci
Total Environ 392:305–312
Fellman JB, D’Amore DV, Hood E et al (2009) Changes in the
concentration, biodegradability, and fluorescence proper-
ties of dissolved organic matter during storm flows in
coastal temperate watersheds. J Geophys Res
114:277–293. doi:10.1029/2008JG000790
Fellman JB, Dogramaci S, Skrzypek G et al (2011) Hydrologic
control of dissolved organic matter biogeochemistry in
pools of a subtropical dryland river. Water Resour Res.
doi:10.1029/2010WR010275
Fichot CG, Benner R (2012) The spectral slope coefficient of
chromophoric dissolved organic matter (S275–295) as a
tracer of terrigenous dissolved organic carbon in river-in-
fluenced ocean margins. Limnol Oceanogr 57:1453
Findlay S, Sinsabaugh RL (2003) Aquatic ecosystems: inter-
activity of dissolved organic matter. Academic Press, San
Diego
Freeman C, Gresswell R, Guasch H et al (1994) The role of
drought in the impact of climatic change on the microbiota
of peatland stream. Freshw Biol 32:223–230
Gomez R, Garcıa V, Vidal-Abarca R et al (2009) Effect of
intermittency on N spatial variability in an arid Mediter-
ranean stream. J N Am Benthol Soc 28:572–583
Gordon ND, McMahon TA, Finlayson BL (2004) Stream hy-
drology: an introduction for ecologists. Wiley, West
Sussex
Graeber D, Gelbrecht J, Kronvang B et al (2012a) Technical
Note: comparison between a direct and the standard,
indirect method for dissolved organic nitrogen determina-
tion in freshwater environments with high dissolved inor-
ganic nitrogen concentrations. Biogeosciences
9:4873–4884
Graeber D, Gelbrecht J, Pusch MT et al (2012b) Agriculture has
changed the amount and composition of dissolved organic
matter in Central European headwater streams. Sci Total
Environ 438:435–446
Green SA, Blough NV (1994) Optical absorption and fluores-
cence properties of chromophoric dissolved organic matter
in natural waters. Limnol Oceanogr 39:1903–1916
Helms JR, Stubbins A, Ritchie JD et al (2008) Absorption
spectral slopes and slope ratios as indicators of molecular
weight, source, and photobleaching of chromophoric dis-
solved organic matter. Limnol Oceanogr 53:955–969
Huber SA, Balz A, Abert M, Pronk W (2011) Characterisation
of aquatic humic and non-humic matter with size-exclusion
chromatography–organic carbon detection–organic nitro-
gen detection (LC-OCD-OND). Water Res 45:879–885
Hudson N, Baker A, Reynolds DM et al (2009) Changes in
freshwater organic matter fluorescence intensity with
freezing/thawing and dehydration/rehydration. J Geophys
Res. doi:10.1029/2008JG000915
Humphries P, Baldwin DS (2003) Drought and aquatic
ecosystems: an introduction. Freshw Biol 48:1141–1146
Inamdar S, Singh S, Dutta S et al (2011) Fluorescence charac-
teristics and sources of dissolved organic matter for stream
water during storm events in a forested mid-Atlantic wa-
tershed. J Geophys Res. doi:10.1029/2011JG001735
IPCC (2013) Climate Change 2013. The physical science basis.
Working group I contribution to the fifth assessment report
of the intergovernmental panel on climate change. Cam-
bridge University Press, New York
Jacobson PJ, Jacobson KM, Angermeier PL, Cherry DS (2000)
Variation in material transport and water chemistry along a
large ephemeral river in the Namib Desert. Freshw Biol
44:481–491
Johnson SL (2004) Factors influencing stream temperatures in
small streams: substrate effects and a shading experiment.
Can J Fish Aquat Sci 61:913–923
444 Biogeochemistry (2015) 123:429–446
123
Author's personal copy

Keil RG, Kirchman DL (1991) Contribution of dissolved amino
acids and ammonium to the nitrogen requirements of
heterotrophic bacterioplankton. Mar Ecol Prog Ser
73:1–10
Lake PS (2003) Ecological effects of perturbation by drought in
flowing waters. Freshwat Biol 48:1161–1172
Lake PS (2011) Drought and aquatic ecosystems: effects and
responses. Wiley, Oxford
Lakowicz JR (2006) Principles of fluorescence spectroscopy.
Springer, New York
Larned ST, Datry T, Arscott DB, Tockner K (2010) Emerg-
ing concepts in temporary-river ecology. Freshw Biol
55:717–738
Lawaetz AJ, Stedmon CA (2009) Fluorescence intensity
calibration using the Raman scatter peak of water. Appl
Spectrosc 63:936–940
Lindell MJ, Graneli W, Tranvik LJ (1995) Enhanced bacterial
growth in response to photochemical transformation of
dissolved organic matter. Limnol Oceanogr 40:195–199
McArthur MD, Richardson JS (2002) Microbial utilization of
dissolved organic carbon leached from riparian litterfall.
Can J Fish Aquat Sci 59:1668–1676
McClain ME, Boyer EW, Dent CL et al (2003) Biogeochemical
hot spots and hot moments at the interface of terrestrial and
aquatic ecosystems. Ecosystems 6:301–312
McDowell WH (1985) Kinetics and mechanisms of dissolved
organic carbon retention in a headwater stream. Biogeo-
chemistry 1:329–352
McKnight DM, Boyer EW, Westerhoff PK et al (2001) Spec-
trofluorometric characterization of dissolved organic mat-
ter for indication of precursor organic material and
aromaticity. Limnol Oceanogr 46:38–48
McMaster D, Bond NR (2008) A field and experimental study
on the tolerance of fish to Eucalyptus camaldulensis lea-
chate and low dissolved oxygen concentrations. Mar
Freshw Res 59:177–185
Nadeau TL, Rains MC (2007) Hydrological connectivity be-
tween headwater streams and downstream waters: how
science can inform policy. J Am Water Resour Assoc
43:118–133
Nguyen HVM, Hur J, Shin HS (2010) Changes in spectroscopic
and molecular weight characteristics of dissolved organic
matter in a river during a storm event. Water Air Soil Poll
212:395–406
Nikolaidis NP, Demetropoulou L, Froebrich J et al (2013)
Towards sustainable management of Mediterranean river
basins: policy recommendations on management aspects of
temporary streams. Water Policy 15:830–849
Ninyerola M, Pons X, Roure JM (2000) A methodological ap-
proach of climatological modelling of air temperature and
precipitation through GIS techniques. Int J Climatol
20:1823–1841
Parlanti E, Worz K, Geoffroy L, Lamotte M (2000) Dissolved
organic matter fluorescence spectroscopy as a tool to esti-
mate biological activity in a coastal zone submitted to
anthropogenic inputs. Org Geochem 31:1765–1781
Pellerin BA, Saraceno JF, Shanley JB et al (2012) Taking the
pulse of snowmelt: in situ sensors reveal seasonal, event
and diurnal patterns of nitrate and dissolved organic matter
variability in an upland forest stream. Biogeochemistry
108:183–198
Peuravuori J, Pihlaja K (1997) Molecular size distribution and
spectroscopic properties of aquatic humic substances. Anal
Chim Acta 337:133–149
Pfeifer D,HoffmannK,HoffmannA et al (2006) TheCalibration
Kit Spectral Fluorescence Standards—A simple and certi-
fied tool for the standardization of the spectral character-
istics of fluorescence instruments. J Fluoresc 16:581–587
Prairie YT (2008) Carbocentric limnology: looking back,
looking forward. Can J Fish Aquat Sci 65:543–548
Prat N, Gallart F, von Schiller D et al (2014) The MIRAGE
Toolbox: an integrated assessment tool for temporary
streams. River Res Appl 30:1318–1334
Raymond PA, Hartmann J, Lauerwald R et al (2013) Global
carbon dioxide emissions from inland waters. Nature
503:355–359
Rodrıguez-Zuniga UF, Milori DMBP, Da Silva WTL et al
(2008) Changes in optical properties caused by UV-irra-
diation of aquatic humic substances from the amazon river
basin: seasonal variability evaluation. Environ Sci Technol
42:1948–1953
Romanı AM, Vazquez E, Butturini A (2006) Microbial avail-
ability and size fractionation of dissolved organic carbon
after drought in an intermittent stream: biogeochemical
link across the stream-riparian interface. Microb Ecol
52:501–512
Sabater S, Tockner K (2010) Effects of hydrologic alterations on
the ecological quality of river ecosystems. In: Sabater S,
Barcelo D (eds) Water scarcity in the Mediterranean.
Perspectives under global change. Springer, New York,
pp 15–39
Sachse A, Babenzien D, Ginzel G et al (2001) Characterization
of dissolved organic carbon (DOC) in a dystrophic lake and
an adjacent fen. Biogeochemistry 54:279–296
Schimel JP, Balser TC, Wallenstein M (2007) Microbial stress-
response physiology and its implications for ecosystem
function. Ecology 88:1386–1394
Singh S, Inamdar S, Mitchell M, McHale P (2014) Seasonal
pattern of dissolved organic matter (DOM) in watershed
sources: influence of hydrologic flow paths and autumn
leaf fall. Biogeochemistry 118:321–333
Spencer RG, Hernes PJ, Ruf R, Baker A, Dyda RY, Stubbins A,
Six J (2010) Temporal controls on dissolved organic matter
and lignin biogeochemistry in a pristine tropical river,
Democratic Republic of Congo. J Geophys Res. doi:10.
1029/2009JG001180Stedmon CA, Bro R (2008) Characterizing dissolved organic
matter fluorescence with parallel factor analysis: a tutorial.
Limnol Ocean Methods 6:572–579
Stedmon CA, Markager S (2005) Tracing the production and
degradation of autochthonous fractions of dissolved or-
ganic matter by fluorescence analysis. Limnol Oceanogr
50:1415–1426
Steward AL, von Schiller D, Tockner K et al (2012) When the
river runs dry: human and ecological values of dry riv-
erbeds. Front Ecol Environ 10:202–209
Tank JL, Rosi-Marshall EJ, Griffiths NA et al (2010) A review
of allochthonous organic matter dynamics and metabolism
in streams. J North Am Benthol Soc 29:118–146
Timoner X, Acuna V, von Schiller D, Sabater S (2012) Func-
tional responses of stream biofilms to flow cessation, des-
iccation and rewetting. Freshw Biol 57:1565–1578
Biogeochemistry (2015) 123:429–446 445
123
Author's personal copy

Tornes E, Sabater S (2010) Variable discharge alters habitat
suitability for benthic algae and cyanobacteria in a forested
Mediterranean stream. Mar Freshw Res 61:441–450
Tzoraki O, Nikolaidis NP, Amaxidis Y, Skoulikidis NT (2007)
Instream biogeochemical processes of a temporary river.
Environ Sci Technol 41:1225–1231
Uys MC, O’Keefe JH (1997) Simple words and fuzzy zones:
early directions for temporary river research in South
Africa. Environ Manag 21:517–531
Vazquez E, Amalfitano S, Fazi S, Butturini A (2011) Dissolved
organic matter composition in a fragmented Mediterranean
fluvial system under severe drought conditions. Biogeo-
chemistry 102:59–72
Vazquez E, Acuna V, Artigas J et al (2013) Fourteen years of
hydro-biogeochemical monitoring in a Mediterranean
catchment. Die Bodenkult 13:3–4
Vazquez E, Romanı AM, Sabater F, Butturini A (2007) Effects
of the dry-wet hydrological shift on dissolved organic
carbon dynamics and fate across stream-riparian interface
in a Mediterranean catchment. Ecosystems 10:239–251
Von Schiller D, Acuna V, Graeber D et al (2011) Contraction,
fragmentation and expansion dynamics determine nutrient
availability in a Mediterranean forest stream. Aquat Sci
73:485–497
Webster JR, Meyer JL (1997) Organic matter budgets for
streams: a synthesis. J North Am Benthol Soc 16:141–161
Weishaar JL, Aiken GR, Bergamaschi BA et al (2003)
Evaluation of specific ultraviolet absorbance as an indi-
cator of the chemical composition and reactivity of dis-
solved organic carbon. Environ Sci Technol 37:4702–4708
Wetzel RG (1992) Gradient-dominated ecosystems: sources and
regulatory functions of dissolved organic matter in fresh-
water ecosystems. Hydrobiologia 229:181–198
Williams CJ, Yamashita Y, Wilson HF et al (2010) Unraveling
the role of land use and microbial activity in shaping dis-
solved organic matter characteristics in stream ecosystems.
Limnol Oceanogr 55:1159–1171
Wilson HF, Xenopoulos MA (2009) Effects of agricultural land
use on the composition of fluvial dissolved organic matter.
Nat Geosci 2:37–41
Wilson HF, Xenopoulos MA (2013) Diel changes of dissolved
organic matter in streams of varying watershed land use.
River Res Appl 29:1330–1339
Yamashita Y, Jaffe R (2008) Characterizing the interactions
between trace metals and dissolved organic matter using
excitation—emission matrix and parallel factor analysis.
Environ Sci Technol 42:7374–7379
Ylla I, Sanpera-Calbet I, Vazquez E et al (2010) Organic matter
availability during pre-and post-drought periods in a
Mediterranean stream. Hydrobiologia 657:217–232
Ylla I, Sanpera-Calbet I, Munoz I et al (2011) Organic matter
characteristics in a Mediterranean stream through amino
acid composition: changes driven by intermittency. Aquat
Sci 73:523–535
Zsolnay A, Baigar E, Jimenez M et al (1999) Differentiating
with fluorescence spectroscopy the sources of dissolved
organic matter in soils subjected to drying. Chemosphere
38:45–50
446 Biogeochemistry (2015) 123:429–446
123
Author's personal copy
Related Documents











