The EMBO Journal vol.15 nO.22 pp.6069-6074, 1996 Myosin motors with artificial lever arms Michael Anson 1 , Michael A.Geeves 2 , Susanne E.Kurzawa 2 and Dietmar J.Manstein 1.3,4 INational Institute far Medical Research, Mill HilI. London W7 I AA. UK, 2Max-Planck-lnstitu( für Molekulare Physiologie. Postfach 102664. D-44026 Donmund and 3Max-Planck·lnstitut für Medizinische Forschung. Jahnstrasse 29. D-69120 Heidelberg. Germany 4Correspondillg aUlhor The myosin head consists of a globular catalytic domain and a light chain binding domain (LCBD). The coupling efficiency between ATP hydrolysis and myosin-induced actin movement is known to decline as the LCBD is truncated or destabilized. However, it was not clear whether the observed alteration in the production of force and movement reflects only the mechanical changes to the length of the LCBD or whether these changes also affect the kinetic properties of the catalytic domain. Here we show that replacement of the LCBD with genetically engineered domains of similar rigidity and dimensions produces functional molecular motors with unchanged kinetic properties. The resulting single- chain, single-headed motors were produced in Dictyo- stelilllll discoidelll/l and obtained after purification from a standard peptone-based growth medium at levels of up to 12 mgll. Their actin motility properties are similar or greater than those of native myosin. Rates of 2.5 and 3.3 were observed for motor domains fused to one or two of these domains, respectively. Their kinetic and functional similarity to the extens- ively studied myosin subfragment 1 (S 1) and their accessibility to molecular genetic approaches makes these simple constructs ideal models for the investig- ation of chemo-mechanical coupling in the myosin motor. Keywords: actin/u-actinin/Dictyostelium discoideuml molecular motorlspectrin repeat Introduction All myosins contain a highly conserved globular catalytic domain that contains the actin and nucleotide billCling sites and produces force and movement (Cheney et al., 1993; Goodson and Spudich, 1993). This 'generic' motor domain is attached via a neck region to a variely of functionally specialized tail domains. The neck region consists of an extended, u-helical region that contains binding sites for one or more calmodulin molecules, or calmodulin-like myosin light chains (Figure IA and B). The neck region of all conventional myosins binds one essential (ELC) and one regulatory light chain (RLC) and is generally referred to as the light chain binding domain (LCBD). In © Oxford University Press agreement with previous models (Vibert and Cohen, 1988) and based on the atomic structure of the myosin head (Rayment et al. I993a) and of the LCBD alone (Xie et al., 1994; Houdusse and Cohen, 1996), it was suggested that the LCBD acts as a rigid lever arm to amplify small conformational changes in the motor domain, thus produc- ing net movement of filamentous actin of -5-10 nm per ATP hydrolyzed (Rayment et al., 1993b; Schräder et al., 1993). In good agreement with this view, functional studies have shown that the coupling efficiency between ATP hydrolysis and movement declines as the neck region is truncated or destabilized by removal of myosin light chains (Pollenz et al., 1992; Itakura et al., 1993; Lowey et al., 1993; Uyeda and Spudich, 1993; VanBuren et al., 1994; Waller et al., 1995; Woodward el al., 1995). Recently, Uyecla and co-workers produced myosins with neck regions of variable length by adding or truncating light chain binding motifs (Uyeda el al., 1996). As predicted by the swinging neck-lever arm model, this study showed a linear relationship between the number of light chain binding sites and sliding velocity. Support for the lever arm hypothesis was also obtained by cryo- electron microscopy. Three-dimensional maps of myosin- decorated F-actin, obtained in the presence and absence of ADP, show linie difference in the region of the globular motor domain; but there is reorientation of the LCBD, compatible with a step of at least 3.5 nm (Jontes et al., 1995; Whittaker et al., 1995). While these studies c1early indicate that the neck region acts as alever, it remains controversial as to whether changes to the LCBD affect the interaction of the myosin motor with actin, binding of nucleotide, hydrolysis of ATP and communication between the actin and nucleotide binding sites. In order to get more direct insight into the functional role of the LCBD, we engineered myosin head fragments (MHFs) consisting of the globular catalytic domain of DicryosteliwlI discoideum myosin fused to a structural motif whose dimensions and rigidity are similar to the LCBD. The best site for fusing an artificial lever arm to the catalytic domain was determined by generating a variety of motor c10main constructs and screening their functional behavior using pre-steady-state kinetics. The shortest construct which behaved similarly 10 an S I-like construct was truncated at Arg761 of the D.discoideum myosin sequence (Geeves el al., 1996). The equivalent residue in adult chicken skeletal muscle myosin is Lys782 (see Figure IA). Dietyosleliul/l discoideum u-actinin repeats were used as artificial lever arms. Each u-actinin repeat (Figure IC) consists of -120 residues and is predicted to form a left-handed coiled-coil consisting of three u-helices, thus creating very compact and rigid structural modules of -6 nm length (Parry et al., 1992; Yan et al., 1993). Expression vectors for the production of two myosin motors with artificial lever arms were 6069

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The EMBO Journal vol.15 nO.22 pp.6069-6074, 1996
Myosin motors with artificial lever arms
Michael Anson1, Michael A.Geeves2,
Susanne E.Kurzawa2 andDietmar J.Manstein1.3,4
INational Institute far Medical Research, Mill HilI.London W7 IAA. UK, 2Max-Planck-lnstitu( für MolekularePhysiologie. Postfach 102664. D-44026 Donmund and3Max-Planck·lnstitut für Medizinische Forschung. Jahnstrasse 29.D-69120 Heidelberg. Germany
4Correspondillg aUlhor
The myosin head consists of a globular catalytic domainand a light chain binding domain (LCBD). The couplingefficiency between ATP hydrolysis and myosin-inducedactin movement is known to decline as the LCBD istruncated or destabilized. However, it was not clearwhether the observed alteration in the productionof force and movement reflects only the mechanicalchanges to the length of the LCBD or whether thesechanges also affect the kinetic properties of the catalyticdomain. Here we show that replacement of the LCBDwith genetically engineered domains of similar rigidityand dimensions produces functional molecular motorswith unchanged kinetic properties. The resulting singlechain, single-headed motors were produced in Dictyostelilllll discoidelll/l and obtained after purification froma standard peptone-based growth medium at levels ofup to 12 mgll. Their actin motility properties aresimilar or greater than those of native myosin. Ratesof 2.5 and 3.3 ~mJs were observed for motor domainsfused to one or two of these domains, respectively.Their kinetic and functional similarity to the extensively studied myosin subfragment 1 (S 1) and theiraccessibility to molecular genetic approaches makesthese simple constructs ideal models for the investigation of chemo-mechanical coupling in the myosinmotor.Keywords: actin/u-actinin/Dictyostelium discoideumlmolecular motorlspectrin repeat
Introduction
All myosins contain a highly conserved globular catalyticdomain that contains the actin and nucleotide billCling sitesand produces force and movement (Cheney et al., 1993;Goodson and Spudich, 1993). This 'generic' motor domainis attached via a neck region to a variely of functionallyspecialized tail domains. The neck region consists of anextended, u-helical region that contains binding sites forone or more calmodulin molecules, or calmodulin-likemyosin light chains (Figure IA and B). The neck regionof all conventional myosins binds one essential (ELC)and one regulatory light chain (RLC) and is generallyreferred to as the light chain binding domain (LCBD). In
© Oxford University Press
agreement with previous models (Vibert and Cohen, 1988)and based on the atomic structure of the myosin head(Rayment et al. I993a) and of the LCBD alone (Xie et al.,1994; Houdusse and Cohen, 1996), it was suggested thatthe LCBD acts as a rigid lever arm to amplify smallconformational changes in the motor domain, thus producing net movement of filamentous actin of -5-10 nm perATP hydrolyzed (Rayment et al., 1993b; Schräder et al.,1993). In good agreement with this view, functional studieshave shown that the coupling efficiency between ATPhydrolysis and movement declines as the neck region istruncated or destabilized by removal of myosin lightchains (Pollenz et al., 1992; Itakura et al., 1993; Loweyet al., 1993; Uyeda and Spudich, 1993; VanBuren et al.,1994; Waller et al., 1995; Woodward el al., 1995).Recently, Uyecla and co-workers produced myosins withneck regions of variable length by adding or truncatinglight chain binding motifs (Uyeda el al., 1996). Aspredicted by the swinging neck-lever arm model, thisstudy showed a linear relationship between the number oflight chain binding sites and sliding velocity. Support forthe lever arm hypothesis was also obtained by cryoelectron microscopy. Three-dimensional maps of myosindecorated F-actin, obtained in the presence and absenceof ADP, show linie difference in the region of the globularmotor domain; but there is reorientation of the LCBD,compatible with a step of at least 3.5 nm (Jontes et al.,1995; Whittaker et al., 1995). While these studies c1earlyindicate that the neck region acts as alever, it remainscontroversial as to whether changes to the LCBD affectthe interaction of the myosin motor with actin, binding ofnucleotide, hydrolysis of ATP and communication betweenthe actin and nucleotide binding sites.
In order to get more direct insight into the functionalrole of the LCBD, we engineered myosin head fragments(MHFs) consisting of the globular catalytic domain ofDicryosteliwlI discoideum myosin fused to a structuralmotif whose dimensions and rigidity are similar to theLCBD. The best site for fusing an artificial lever arm tothe catalytic domain was determined by generating avariety of motor c10main constructs and screening theirfunctional behavior using pre-steady-state kinetics. Theshortest construct which behaved similarly 10 an S I-likeconstruct was truncated at Arg761 of the D.discoideummyosin sequence (Geeves el al., 1996). The equivalentresidue in adult chicken skeletal muscle myosin is Lys782(see Figure IA). Dietyosleliul/l discoideum u-actininrepeats were used as artificial lever arms. Each u-actininrepeat (Figure IC) consists of -120 residues and ispredicted to form a left-handed coiled-coil consisting ofthree u-helices, thus creating very compact and rigidstructural modules of -6 nm length (Parry et al., 1992;Yan et al., 1993). Expression vectors for the productionof two myosin motors with artificial lever arms were
6069
M.Anson et sI.
created. The resulting proteins, M761-1 Rand M761-2R,consist of the catalytic domain fused to one or twoa-actinin repeats, respectively. The biochemical and functional competence of M761-1R and M761-2R was determined using pre-steady-state kinelics and in vitramotility assays.
This report addresses three questions raised by the leverarm hypothesis. Firstly, is it possible to replace the LCBDby a more simple structure of similar dimensions andrigidity and retain the motile properties? Secondly, doessubstitution of the LCBD alter the enzymatic propertiesof the myosin motor? Finally, do the motile properties
1500
6.6 min
1400
%
1300
""128~.O
'"1317.3 Retention time
'"1352.1
1200
...119<1.6
m/z
".115<1.6
11001000
A,100; 1027.6A97; 1059.~
" ..A,106 1093.1
"'.0 M.102658.3 ± 7.0
..'"969.~
900
100
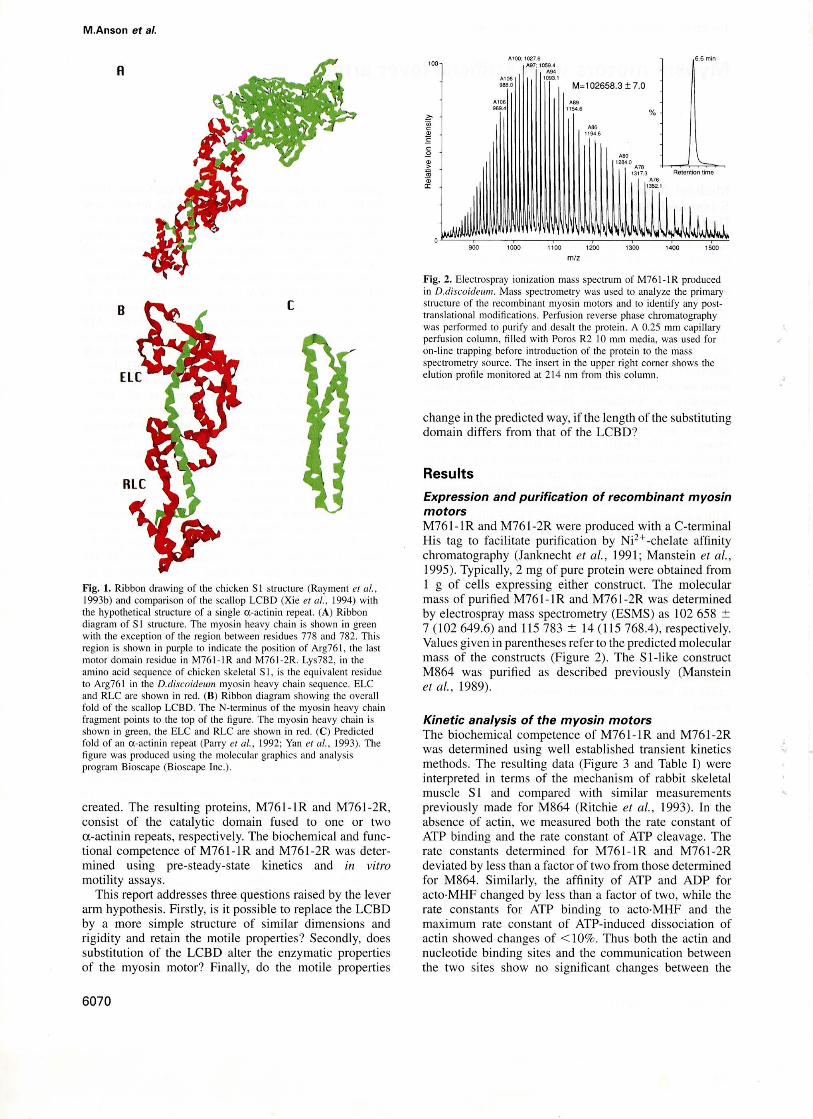
Fig. 2. Electrospray ionization mass spectrum of M761-IR producedin D.discoideu11l. Mass spectrometry was used to analyze the primarystructure of the recombinant myosin motors and 10 identify any posttranslational modifications. Perfusion reverse phase chromatographywas performed to purify and desalt the prolein. A 0.25 mm capillaryperfusion column, filled with Poros R2 10 mm media, was used foron-line trapping before introduction of the protein to the massspectrometry source. The insert in the upper right corner shows theelution profile monitored at 214 11m from this column.
change in the predicted way, if the length of the substitutingdomain differs from that of the LCBD?
Results
Expression and purification of recombinant myosinmotorsM76l-IR and M761-2R were produced with aC-terminalHis tag to facilitate purification by Ni2+ -chelate affinitychromatography (Janknecht et al., 1991; Manstein et al.,1995). Typically, 2 mg of pure protein were obtained from1 g of cells expressing either construct. The molecularmass of purified M761-IR and M761-2R was determinedby electrospray mass spectrometry (ESMS) as 102 658 :'::7 (102 649.6) and 115783 :':: 14 (115 768.4), respectively.Values given in parentheses refer to the predicted molecularmass of the constructs (Figure 2). The S l-like constructM864 was purified as described previously (Mansteinet al., 1989).
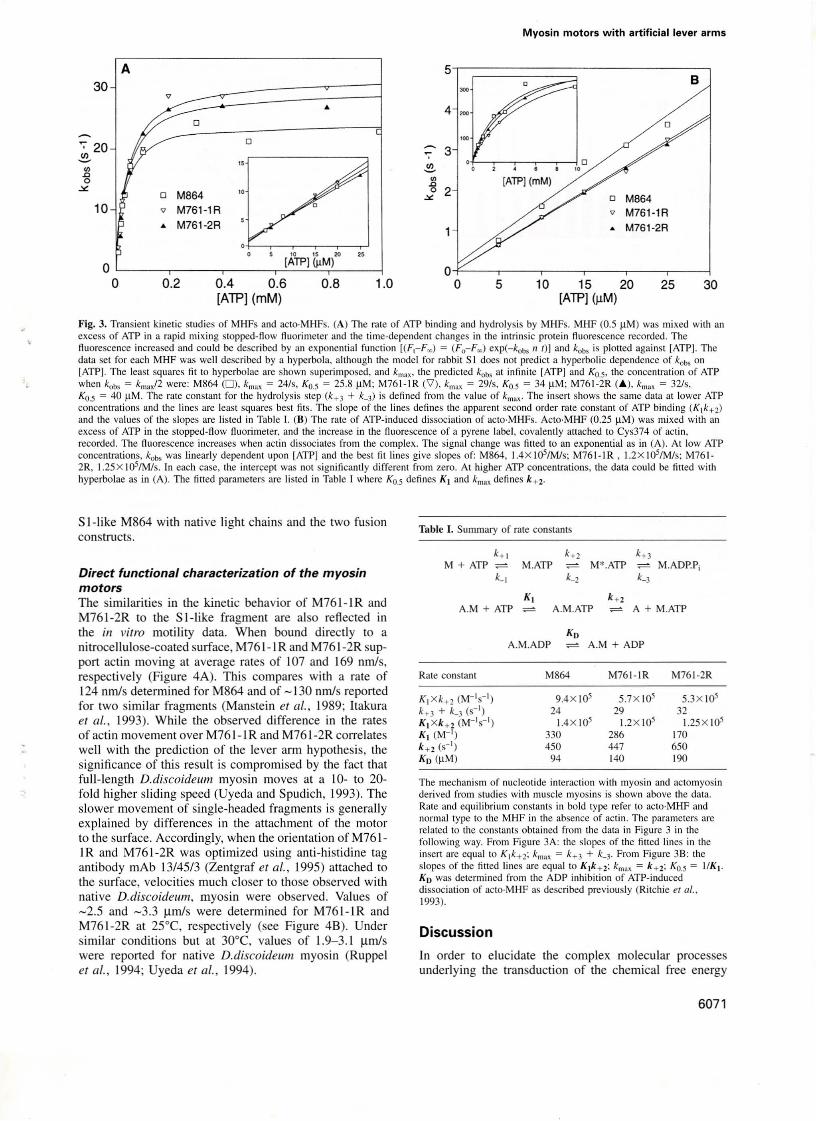
Kinetic analysis of the myosin motorsThe biochemical competence of M761-1 Rand M761-2Rwas determined using weil established transient kineticsmethods. The resulting data (Figure 3 and Table I) wereinterpreted in terms of the mechanism of rabbit skeletalmuscle SI and compared with similar measurementspreviously made for M864 (Ritchie et al., 1993). In theabsence of actin, we measured both the rate constant ofATP binding and the rate constant of ATP cleavage. Therate constants determined for M761-1R and M761-2Rdeviated by less than a factor of two from those determinedfor M864. Similarly, the affinity of ATP and ADP foracto·MHF changed by less than a factor of two, while therate constants for ATP binding to acto·MHF and themaximum rate constant of ATP-induced dissociation ofactin showed changes of < 10%. Thus both the actin andnucleotide binding sites and the communication betweenthe two sites show no significant changes between the
[
A
Fig. 1. Ribbon drawing of the chieken SI structure (Rayment er al.,1993b) and comparison of the scallop LCBD (Xie el al., 1994) withthe hypOlhelical structure of a single (X-actinin repeat. (A) Ribbondiagram of SI structure. The myosin heavy chain is shown in greenwirh the exception of the region between residues 778 and 782. Thisregion is shown in purple to indicate the position of Arg761, the laslmotor domain residue in M761-1R and M761-2R. Lys782, in theamino acid sequence of chicken skeletal SI, is the equivalent residue10 Arg761 in the D.discoideul1l myosin heavy chajn sequence. ELCand RLC are shown in red. (B) Ribbon diagram showing the overallfold of the scaUop LCBD. Thc N-terminus of the myosin heavy chainfragment points 10 the top of the figure. The myosin heavy chain isshown in green, the ELC and RLC are shown in red. (C) Predictedfold of an a-actinin repeat (Parry et al., 1992; Yan et al., 1993). Thefigure was produced using the molecular graphics and analysisprogram Bioscape (Bioscape lnc.).
6070
Myosin motors with artificial lever arms
302510 15 20[ATP] (~M)
5O-l"---,----,--,----,-------r----1
o
5
"'"4
~
3';"
.!!!..'"D0 2
""" 0 M864
" M761·1R. M761-2R
..o
o M864
" M761-1R.. M761-2R
10 15 20 25
o '--__r-__r-__r-'-[A_JP---'lc.;(I1c-M,.:.}__--1
o 0.2 0.4 0.6 0.8 1.0[ATP] (mM)
10
30A
Fig. 3. Transient kinelic studies of MHFs and aClo·MHFs. (A) The rale of ATP binding and hydrolysis by MHFs. MHF (0.5 11M) was mixed wilh anexcess of ATP in a rapid mixing stopped-flow fluorimclcr und the time-dependent changes in the intrinsic protein fluorescence recorded. Thefluorescence increased and could be described by an exponenlial funclion [(F,-F~) ; (Fo-F~) exp(-kobs 11 I)J and kobs is plolled against [ATP]. Thedala seI for each MHF was weil described by a hyperbola. although the model for rabbi I SI does nol predicI a hyperbolic dependence of kobs onlATPj. The leasl squares fil 10 hyperbolae are shown superimposed, and km", Ihe predicled kobs at infinile [ATP] and Ko.S' the concentralion of ATPwhen kob, ; k""xf2 were: M864 (0), k""" ; 24/s, Ko.s ; 25.8 11M; M761-IR ('1), km" ; 29/s, Ko.s ; 34 11M; M761-2R (A), km" ; 32/s,KO.5 = 40 11M. The rate constant for the hydrolysis step (k+ 3 + k_3) is defined from the value of kmax - The insert shows the same data at lower ATPconcentrations and the lioes are least squares best fils. The slape of the lines defines the apparent second order rate constant of ATP binding (K1k+ 2)
and Ihe values of the slopes are listed in Table I. (8) The rale of ATP-induced dissociation of aClo·MHFs. ACIO·MHF (0.25 11M) was mixed wilh anexcess of ATP in the slOpped-flow ftuorimeler. and the increase in the f1uorescence of a pyrene label, covalemly attached lO Cys374 of aClin,recorded. The fluorescence increases when aClin dissociates from Ihe complex. The signal change was fitted lO an exponential as in (A). At low ATPconcentrations, koo, was Iinearly dependenl upon [ATPJ and Ihe best fil Iines give slopes of: M864, 1.4X IOs/Mls; M761-IR, 1.2X 10s/M/s; M7612R, 1.25x 105/M/s. In each case, the inter~ept was not significantly different from zero. At higher ATP concentrations, lhe data could be fitted withhyperbolae as in (A). Tbe filled parameIers are lisled in Table I where Ko.s defines K I and km" defines k+2.
Discussion
In order to elucidate the complex molecular processesunderlying the transduction of the chemical free energy
Table I. Summary of rate constants
k+1 k+2 k+3M + ATP - M.ATP - M*.ATP - M.ADP,Pi
k_l k_2 k_3
KI k+2A.M + ATP - A.M.ATP - A + M.ATP
Rate constant M864 M761-IR M761-2R
K l xk+2 (M-ls- l) 9.4X 105 5.7X 105 5.3X 105k+ 3 + k_3 (s-') 24 29 32K l xk+ 2 (M-'s-l) 1.4X 105 1.2X 105 1.25X 105KI (M-') 330 286 170k+2 (S-I) 450 447 650Ko (11M) 94 140 190
A.M + ADPKo
A.M.ADP
The mechanism of nucleotide interaction with myosin and actomyosinderived from swdies with muscle myosins is shown above the data.Rate and equilibrium constants in bold type refer 10 ac1O·MHF andnormal lype lO the MHF in the absence of aClin. The parameters arerelated to the constams obtained from the data in Figure 3 in thefollowing way. From Figure 3A: Ihe slopes of Ihe fined lines in Iheinsert are equal to K1k+ 2; kmax = k+ 3 + k_3' From Figure 38: theslopes of Ihe fined lines are equallo Klk+2; k",,, = k+2: Ko.s; I/K I .
Ko was delermined from Ihe ADP inhibition of ATP-induceddissociation of aCIO·MHF as described previously (Ritchie er al.,1993).
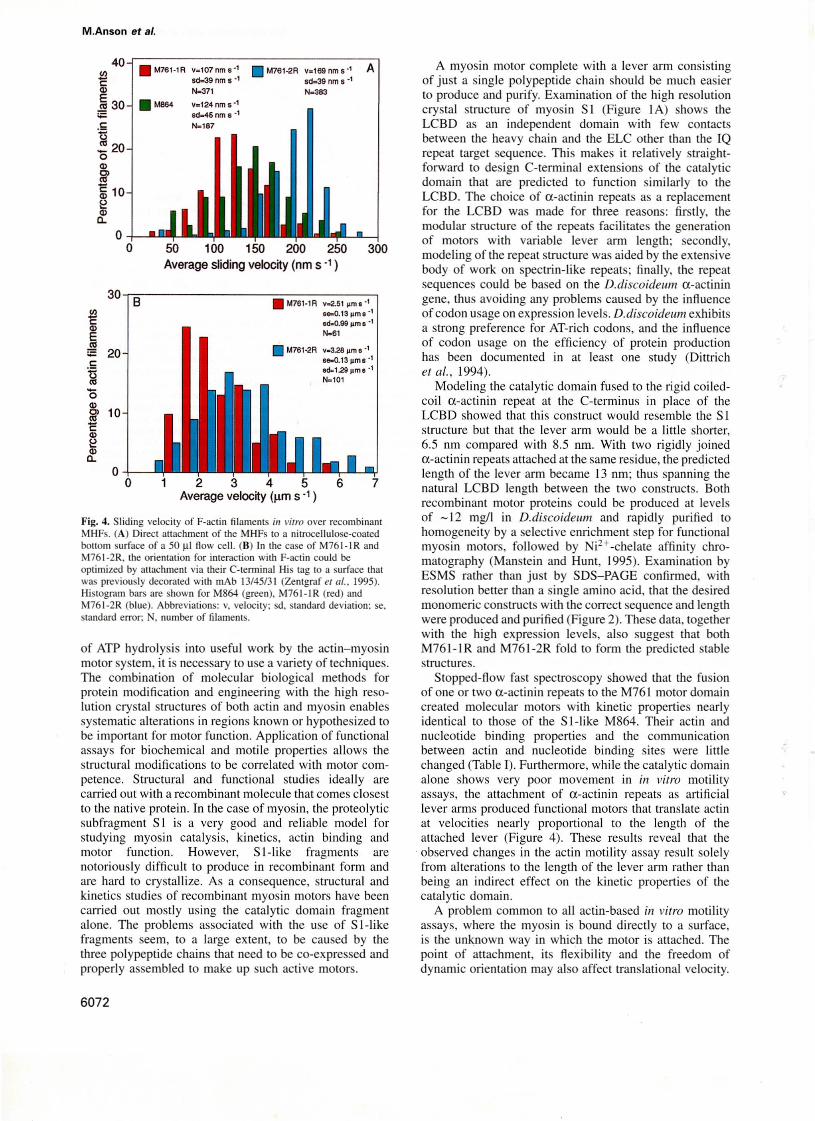
Direct functional characterization of the myosinmotorsThe similarities in the kinetic behavior of M761-1 RandM761-2R to the SI-like fragment are also reflected inthe in vitra motility data. When bound directly to anitrocellulose-coated surface, M761-IR and M761-2R support actin moving at average rates of 107 and 169 nm/s,respectively (Figure 4A). This compares with a rate of124 nm/s deterrnined for M864 and of - 130 nm/s reportedfor two similar fragments (Manstein et 01., 1989; ltakuraet 01., 1993). While the observed difference in the ratesof actin movement over M761- 1Rand M76 1-2R correlatesweil with the prediction of the lever arm hypothesis, thesignificance of this result is compromised by the fact thatfull-Iength D.discaideum myosin moves at a 10- to 20fold higher sliding speed (Uyeda and Spudich, 1993). Theslower movement of single-headed fragments is generallyexplained by differences in the attachment of the motorto the surface. Accordingly, when the orientation of M7611Rand M761-2R was optimized using anti-histidine tagantibody mAb 13/45/3 (Zentgraf et 01., 1995) attached tothe surface, velocities much cJoser to those observed withnative D.discaideum, myosin were observed. Values of-2.5 and -3.3 ~m/s were determined for M761-1R andM761 -2R at 2SOC, respectively (see Figure 48). Undersimilar conditions but at 30°C, values of 1.9-3.1 ~m/s
were reported for native D.discaideum myosin (Ruppelet 01., 1994; Uyeda et 01., 1994).
S I-like M864 with native light chains and the two fusionconstructs.
6071
M.Anson et si.
Fig. 4. Sliding velocily of F-aclin filaments in vitra Qver recombinantMHFs. (A) DirecI attachment uf the MHFs 10 a nitrocellulose·coatedbouom surface of a 50 fll flow cell. (8) In the case of M76 I-I RandM761-2R. the orientalion for interaction with F-aclin could beoptimized by allachment via their C-tenninal His lag 10 a surface thatwas previously decorated wilh mAb 13/45/3 I (Zentgraf el 111., 1995).Histogram bars are shown for M864 (green), M761-1 R (red) andM761-2R (blue). Abbrevialions: v, velocilY: sd, standard deviation: se,standard errar; N, "umber of filaments.
A myosin motor complete with a lever arm consistingof just a single polypeptide chain should be much easierto produce and purify. Examination of the high resolutioncrystal structure of myosin SI (Figure IA) shows theLCBD as an independent domain with few contactsbetween the heavy chain and the ELC other than the IQrepeat target sequence. This makes it relatively straightforward to design C-terminal extensions of the catalyticdomain that are predicted to function similarly 10 theLCBD. The choice of a-actinin repeats as areplacementfor the LCBD was made for three reasons: firstly, themodular structure of the repeats facilitates the generationof motors with variable lever arm length; secondly,modeling of the repeat structure was aided by the extensivebody of work on spectrin-like repeats; finally, the repeatsequences could be based on the D.discoideum a-actiningene, thus avoiding any problems caused by the influenceof codon usage on expression levels. D.discoideum exhibitsa strong preference for AT-rich codons, and the influenceof codon usage on the efficiency of protein productionhas been documented in at least one study (Dittrichel al., 1994).
Modeling the catalytic domain fused to the rigid coiledcoil a-actinin repeat at the C-terminus in place of theLCBD showed that this construct would resemble the SIstructure but that the lever arm would be a little shorter,6.5 nm compared with 8.5 nm. With two rigidly joineda-actinin repeats attached at the same residue, the predictedlength of the lever arm became 13 nm; thus spanning thenatural LCBD length between the two constructs. Bothrecombinant motor proteins could be produced at levelsof -12 mg/I in D.discoidewn and rapidly purified tohomogeneity by a selective enrichment step for functionalmyosin motors, followed by Ni2+-chelate affinity chromatography (Manstein and Hunt, 1995). Examination byESMS rather than just by SOS-PAGE confirmed, withresolution better than a single amino acid, that the desiredmonomeric constructs with the correct sequence and lengthwere produced and purified (Figure 2). These data, togetherwith the high expression levels, also suggest that bothM761-' Rand M761-2R fold to form the predicted stablestructures.
Stopped-flow fast spectroscopy showed that the fusionof one or two a-actinin repeats to the M761 motor domaincreated molecular motors with k..inetic properties nearlyidentical to those of the S I-Iike M864. Their actin andnucleotide binding properties and the communicationbetween actin and nucleotide binding sites were littlechanged (Table I). Furthermore, while the catalytic domainalone shows very poor movement in in vilro motilityassays, the attachment of a-actinin repeats as artificiallever arms produced functional motors that translate actinat velocities nearly proportional to the length of theattached lever (Figure 4). These results reveal that the
. observed changes in the actin motility assay result solelyfrom alterations to the length of the lever arm rather thanbeing an indirect effect on the kinetic properties of thecatalytic domain.
A problem common to all actin-based in vitro motilityassays, where the myosin is bound directly to a surface,is the unknown way in which the motor is attached. Thepoint of attachment, its flexibility and the freedom ofdynamic orientation mayaiso affect translational velocity.
7
300
6
• M761·1R v-2.51 )lms-1
se-O.13pms·1
ed_O.99 ",me·1
N.6'
• M761.2R v-3.28 jl.m B ·1se-O.13pms·1
ad-l.29 pm 8 ·1N.l01
12345Average velocity (1lßl S ., )
o
O-+-.....~.o 50 100 150 200 250
Average sliding velocity (nm s")
30 8
o-+----
E~~ 20c:
~ÖQlCl 10~Ql~
~
40-r-----------------,• M761-1A v.107nms" • M761·2R V.l69nms·' A
-Ql~ sd=39 nm s -1 sd.39 nm s .,~371 N.383
_._~_ 30 • Me64 v.124 nm s"sd....6nm 6"
.5 N-187ti~20oQl
~~ 10~Qlc..
of ATP hydrolysis into useful work by the actin-myosinmotor system, it is necessary to use a variety of techniques.The combination of molecular biological methods forprotein modifkation and engineering with the high resolution crystal structures of both actin and myosin enablessystematic alterations in regions known or hypothesized tobe important for motor function. Application of functionalassays for biochemical and motile properties allows thestructural modi/kations to be correlated with motor competence. Structural and functional studies ideally arecarried out with a recombinant molecule that comes closestto the native protein. In the case of myosin, the proteolyticsubfragment SI is a very good and reliable model forstudying myosin catalysis, kinetics, actin binding andmotor function. However, S l-Iike fragments arenotoriously difficult to produce in recombinant form andare hard to crystallize. As a consequence, structural andkinetics studies of recombinant myosin motors have beencarried out mostly using the catalytic domain fragmentalone. The problems associated with the use of S I-Iikefragments seem, to a large extent, to be caused by thethree polypeptide chains that need to be co-expressed andproperly assembled to make up such active motors.
6072

Further, molecules attached in an unfavorable orientationor not able to move freely may still interact with the actinpolymers and reduce their velocity. This may explain thevery low translational velocities seen for single-headedmyosin constructs «200 nmls) compared with those withfull-Iength wild-type D.discoideul1l double-headed myosin(2-3 ~m/s) which approach those of fast double-headedskeletal myosins (4-8 ~m/s). Thus the velocities whichwe observed with our constructs attached to nitrocellulose(Figure 4A) may ref1ect artifacts of the i/1 vitro assaysurface attachment. This condusion is supported by ourresults using the C-terminal His tag to orient the motorconstruct by binding this tag to antibody molecules adhering to the surface. In this way, the motor molecules wereattached at the extreme C-terminal end of the lever arm,raised above the nitrocellulose surface and given a largedegree of orientational f1exibility. As predicted, velocitiesthen obtained were 20-fold higher and similar to those offull-Iength wild-type myosin (Figure 4B). Indeed, thevelocities obtained (at 25°C) correlated with the estimatedlength of the lever arm from the atomic model : M7611R, 6.5 nm and 2.5 ~m/s , M761-2R, 13 nm and 3.3 ~m1s.
Uyeda and co-workers (1996) used a different approachto varying the length of the lever arm. They either deletedone or both of the light chain binding sites 6r added asecond binding site to the double-headed nativeD.discoideum myosin motor with full coiled-coil tai!. Inthis way, they generated constructs with predicted lengthsof 2, 6 and 13 nm from the putative fulcrum point ofattachment of the LCBD to the start of the 52 region, andcompared these with native wild-type myosin, to whichthey assigned alever length of 8.5 nm. They usedequilibrium ATPase measurements with or without 24 ~.t.M
actin as indications of biochemical competence and theill vitro motility assay to quantify motor function. Bindingthe full-Iength myosin motor proteins, presumably viatheir long coiled-coil tails, directly to the surface, theyfound (at 30°C) actin velocities between 0.6 and 4 ~m1s,
directly proportional to the predicted lever arm length.These are dose to the results presented here, and theirvelocity for wild-type myosin with LCBD length of8.5 nm, 3 ~m/s fits weil with our data.
Both from inspection of the chicken 5 I structure(Rayment et 01., 1993a) and from the functional propertiesobserved for point mutations in the interface between themyosin heavy chain and ELC, that are associated withvariarlts of human cardiac hypertrophy (Poetter et 01.,1996), one would expect a more critical role in mechanochemical coupling for this region than observed with ourconstructs in the i/1 vitro actin gliding assay. The mostlikely explanation for this discrepancy lies in the fact thatthis assay measures only one parameter of motor function.A motor has to produce force and movement, and theill vitra actin motility assay measures only the latterfunction against almost no load. The actin in vitro motilityassay thus gives little indication as to how severely acertain mutation may affect the force production by themotor and about its functional competence in vivo.
We have shown that it is possible to replace the LCBDof D.discoidelll1l myosin with rigid a-actinin constructsof variable length. The kinetics of such fusion proteinsare dosely similar to those of wild-type and they can beexpressed and purified in large quantities. When orientated
Myosin motors with artificial lever arms
by binding to antibody, they have in vitro propertiessimilar to those of the wild-type myosin, showing thatthey are fully functional motors, sustaining actin motilityat velocities related to the lever arm length. Their highlevel of expression, ease of purification and functionalcompetence should make such constructs extremely usefulfor further structural and functional studies of themyosin motor.
Materials and methods
Constructionl expression and purification of recombinantmyosin motorsThe expression vectors used for the production of the myosin-a-actininfusion constructs are based on pDXA-3H (Manstein er al., 1995). ThecQnSlructs wefe created by Iinking codan 761 of the D.discoideum ","cAgene (De Lozanne e1 al., 1985) to codon 264 of the D.discoidellllla-actinin gene ( oegel et al., 1987). Constructs fused to olle centealrepeat extended to codan 387 and conslrucls tagged with two repeats tocodan 505 of (he o.-aclinin gene. The peptide Leu-Gly-Ser separatesthe two sequences at 1he site of fusion, and both construc1S are 1aggedat their C-1erminus with the peptide Ala-Leu-{His)g. All oligonucleo1idelinkers were optimized for D.discoideum codon bias. Modeling of thea-actinin repeat slruClure was performed using the programs PHD (Rostand Sander, 1993) and Bioscape (Bioseape Ine., Palo Aho, CA), DNAmanipulations were done as described in Sambrook el al. (1989) andEgelhoff et al, (1991). M761-IR and M761-2R were overprodueed inAX3-0RF+ eells and purified by Ni2+-ehelate affinity ehromatographyas described previously (Manstein and Hunt, 1995; Manslein el al.,1995). M864 was purified as described by Ritchie et al, (1993).
Electrospray mass spectrometry was performed on a Fisons VGPlatform with on-line lrapping as described by Aitken er al. (1995).
Stopped-flow experimentsAll transient kine1ic measurements were made using a Hi-Tech SF-61slopped flow spectrofluorimeter. The details of this apparatus and its usefor measuring lhe transient kine1ics of rabbit SI and D.discoideum MHFshave been describcd in our previous sludies (Ritchie el al., 1993:Woodward el al., 1995). Actin was prepared from an ace10ne powder ofrabbi1 skelelal muscle and labeled Wi1h pyrene iodoacetamide as describedpreviously (Criddle el al., 1985). Experimental conditions for all measurements were: 20°C. 0, I M KCI. 5 mM MgCI2• 20 mM HEPES adjustedto pH 7.0.
Moti/ity assaysThe in vilro assay was perfonned in a manner similar to that describedpreviously (Anson. 1992) but with some modifications. Briefly, a I.3NA40x lens was used in epifluorescence with 546 nm excitation (Hg arc)and 580 nm emission band-pass (Omega filter set XF37). Filamentsweee imaged Wi1h a 4X lens on an intensified CCD TV camera (Darkstar800) and recorded, via an image processor (Hamamatsu Argus 20) andtime-date generator, by an S-VHS VCR (1VC BRS800·E). For slowmoving filaments (Figure 4A), the TV formal was converted from PALto NTSC by a CVR22 digital converter (SneIl and Wilcox). Thispermined aUlOmated filament tracking using a VPIIO digilizer atI frame/s utilizing Expert Vision software (Motion Analysis Inc.) runningon a 486DX2-66 Pe. For additional manual analysis of fast movingfilaments (Figure 4B), tapes were replayed from the VCR to the Argus20 and individual filaments tracked frame-by-frame using a mouse.Velocity was calculated from lhe posilional change and lhe time coderecorded Oll 1he tape. All solutions were kepl on ice and the microscopeslage was stabilized al 25°C.
Antibody-decorated surfaces were generated as folIows: 75 111 of mAb13/45131 (Dianova GmbH, Hamburg). 200 ~g/ml in phosphate-bufferedsaline, was infused inlO a 50 ,.11 f10w cell wilh a nitrocellulose-coatedbottom surface. The cell was placed in a humidifier at 4°C for 60 mill10 allow the anlibody to bind 10 the surface. It was flushed with 100 JlIof bovine serum albumin (BSA), 10 mg/mi in 100 mM KCI and 50 mMHEPES pH 7.2. and incubated for 15 min at 2SOe. This was repcated1O ensure blocking of vacant protein binding sites on the surface. It waswashed with 100 ~I of I mg/Oll BSA in 100 OlM KCI and 30 mMHEPES pH 7.2. Then 50 ~I of I mg/Oll M761-IR or M761·2R in HAB(25 mM KCI, 25 mM HEPES pH 7.5. 4 mM MgCI2 I OlM EGTA,5 mM di1hiothreitol) was infused from each end of the flow-cell channel
6073

M.Anson et 81.
and allowed to bind to the antibody~coated surface for 45 min in (hehumidifier at 4°C. After washing Wilh 100 111 of 0.5 mg/mi BSA inHAB. 100 ~I of 20 nM rabbit skeletal actin labeled with rhodaminephalloidin was added and incubated für 2 min. The cell was washedagain with 100 ~I of 0.5 mg/mi BSA in HAB. Then 100 ~I of AB (HABwith 100 ~g/ml of glucose oxidase. 20 ~g/ml of catalase. 5 mg/mi ofglucose and 0.5 mg/mi of BSA, pH 7.5) was infused and the celltransferred (0 an inverted microscope (Zeiss Axioven 35). After focusingand observation of the actin filaments in rigor, motion was initialcd byinfusing 100 ~I of AB containing 2 mM ATP.
AcknowledgementsWe would like to thank Or C.Cohcn for discussions and helpfulsuggestions, Or A.A.Noegel for her gift of the D.discoideulfI a-aclinincDNA, Dr H.Faulstich for his gift of rhodamine-phalloidin, DrJ.A.Spudich for critical rcarling of the manuscripI, Or A.Aitken andS.Howeli for the mass spectromelry, D.M.Hunl for expert 1echnicalassistance and N.Adamek fer the preparation of pyrene·actin. The workwas supported by the Medical Research Council (UK) and the MaxPlanck Society (Germany).
References
Aitken.A., Howell,S., Jones.D., Madrazo,!. and Patel,Y. (1995) 14-3-3a and 8 are the phosphorylated fonns of Raf activating 14-3-3 ß and~. 1. Biol. Chem., 270, 5706-5709.
Anson,M. (1992) Temperature dependence and Arrhenius activationenergy of F-actin velocily generated in vitra by skeletal myosin.J. Mol. Bio!., 224, 1029-1038.
Cheney,R.E., Riley,M.A. and Mooseker,M.S. (1993) Phylogeneticanalysis of the myosin superfamily. Ce!! Mord Cytoskeleton, 24,215-223.
Criddle,A.H., Geeves,M.A. and Jeffrries,T. (1985) The use of actinlabelIed with ·(l-pyrenyl) iodoacetamide 10 study the interaction ofactin with myosin sllbfragments and troponin/tropomyosin. Biochem.J., 232. 343-349.
De Lozanne,A.. Lewis,M., Spudich.J.A. and Leinwand,L.A. (1985)Cloning and characterization of a nonJ11l1scle myosin heavy chaincDNA. Proc. Na/! Acad. Sei. USA, 82, 6807-6810.
Dittrich,W.. Williams,K.L. and Slade,M.B. (1994) Production ofrecombinant proteins in DicJ)'osteliwIl discoideum. Bio/techno/agy,12,614-617.
Egelhoff,T.T., Titus,M.A., Manstein,DJ., Ruppel,K.M. and Spudich.J.A.(1991) Molecular genetic taols for study of the cytoskeletan inDictyosteliul11. Methods Enzymal., 196, 319-334.
Geeves,M.A., Kurzawa,S., Hunt,D.M. and Manstein,DJ. (1996) Kineticcharacterisation of different length fragments of the myosin headexpressed in Dictyosteliu11I. Biophys. 1.,70, A267.
Goodson.H.V. and Spudich,J.A. (1993) Molecular evolution of themyosin family: relationships derived from comparisons of amino acidsequenees. Proc. Neu! Acad. Sei. USA. 90, 659-663.
Houdusse,A. and Cohen,e. (1996) Structure of the regulatory domainof scallop myosin at 2 Aresolution: implication far regulation.Strucrure, 4, 21~32.
Itakura,S., Yamakawa,H., Toyoshima, YY, Ishijima,A., Kojima,T.,Harada,Y., Yanagida.T., Wakabayashi,T. and Sutoph.K. (1993) Forcegenerating domain of myosin motor. Biochem. Biophys. Res. COII/II/UII.,196.1504-1510.
Janknecht,R., De Martynoff,G., Lou,!., Hipskind,R.A., Nordheim,A. andStunnenberg,H.G. (1991) Rapid and efficient purification of nativehistidine-tagged protein expressed by recombinant vaccinia virus.Prac. Not! Acad. Sei. USA, 88, 8972-8976.
Jontes,JD., Wilson Kubalek,E.M. and Milligan,R.A. (1995) A 32 degreetail swing in brush borde I' myosin I on ADP release. Nature, 378,751-753.
Lowey,S., Waller,G.S. and Trybus,K.M. (1993) Skeletal musele myosinlight chains are essential for physiological speeds of shonening.Natllre, 365, 454-456.
Manstein,D.J. and Hunt,D.M. (1995) Overexpression of myosin motordomains in Dictyostelilll11: screening of transformants and purificationof the affinity tagged protein. 1. MIIscle Res. Cell Mo/i! .. 16,325-332.
Manstein,DJ .. Ruppel.K.M. and Splidieh,J.A. (1989) Expression andcharacterization of a functional myosin head fragment in Dictyostelil/IIIdiscoideum. Sciellce, 246, 656-658.
ManSlein,D.J., Schuster,H.-P., Morandini.P. and Hunt.D.M. (1995)
6074
Cloning vcctors for the production of proteins in DictyosteliulI/discoidelll/l. Gene, 162, 129-134.
Noegel,A., Witke,W. and Schleicher,M. (1987) Calcium-sensitive nOI1museIe alpha-actinin cOl1lains EF-hand structurcs and highly conservedregions. FEBS Lett.. 221. 391-396.
Parry,D.A., Dixon,T.W. and Cohen,C. (1992) Analysis of the threealpha-helix motif in the spectrin superfamily of proreills. Biophys. 1..61, 858-867.
Poetter,K. et a/. (1996) Mutations in eilher the essential or reglilatorylight chains of myosin are associated wilh a rare myopathy in humanheart and skelctal muscle. NafIlre Gellet., 13, 63-69.
Pollellz.R.S .. Chen,T.L.L., Trivinoslagos,L. and Chisholm,R.L. (1992)The Diclyostelium essential light chain is requircd for myosin function.Cell. 69, 951-962.
Rayment,I., Holden,H.M., Whitlaker,M., Yohn,e.B., Lorenz,M., Holmes,K.e. and Milligan,R.A. (1993a) Structure ofthe actin-myosin complexand its implications for muscle contraction. Sciellce, 261, 58-65.
Rayment,l., Rypniewski,W.R., Schmidtbase,K., Smith,R., TOlllChick,D.R., Benning,M.M., WinkelmanJ1,D.A., Wcscnberg,G. and Holden,H.M. (1993b) Three-dimensional structurc of myosin subfragment-I:a molecular motor. Science, 261, 50-58.
Ritchie,MD., Woodward,S.K.A., ManSlein.DJ. and Geeves,M.A. (1993)Characterization of the interac1ion of a DictyosteliwlI myosin headfragment with actin and nucleotide. Biophys. 1., 64, 359.
Rost,B. and SandeLe. (1993) Prediction of protein structure at bettel'than 70% accllracy. 1. Mol. Biai., 232. 584-599.
Ruppel,K.M .. Uyeda,T.Q.P. and Spudich,J.A. (1994) Role of highlyconserved lysine 130 of myosin motor domain: in vivo and in vitracharactcrization of site specifically mutated myosin. J. Biol. Chem.,269.18773-18780.
Sambrook,J., Fritsch,E.F. and Maniatis,T. (1989) Molecular Cloning: ALaborarory Manual. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, I Y
Schröder,R.R., Manstein,DJ., Jahn,W., Holden,H., Rayment,I., Holmes,K.e. and Spudich,J.A. (1993) Three-dimensional atOinic model ofF·actin decorated \Vitll Dictyoste/illfllillyosin S I. Nature, 364, 171~174.
Uyeda,T.Q.P. and Splidich,J.A. (1993) A functional recombinant myosin11 laeking a regulatory light chain-binding site. Sciellce, 262, 18671870
Uyeda,T.Q.P., Ruppel,K.M. and Spudich,J.A. (1994) Enzymatic activitiescorrelate with chimaeric substitutions at the actin-binding face 01'myosin. Nature, 368, 567-569.
Uyeda,T.Q.P., Abramson,PD. and Spudieh.J.A. (1996) The neck regionof the myosin motor domain acts as a lever arm to generale movemenl.Proc. Neu! Acad. Sei. USA, 93. 4459-4464.
VanBuren.P., Waller,G.S., Harris,D.E., Trybus,K.M., Warshaw,D.M. andLowey,S. (1994) The essential light ehain is required for fuH forceproduction by skeletal muscle myosin. Prac. Nml Acad. Sci. USA, 91,12403-12407.
Viben,P. and Cohen,C. (1988) Domains, molions and regulation in themyosin head. J. Mt/sele Res. CeU Mo/i!., 9, 296-305.
Waller.G.S., Ouyang,G., Swafford.J., Vibert,P. and Lowey,S. (1995) Aminimal motor domain from chicken skeletal museie myosin. J. Biol.Chem., 270, 15348-15352.
Whittaker.M .. Wilson Kubalek,E.M., Smith,J.E., FauSl.L., Milligan.R.A.and Sweeney,I-I.L. (1995) A 35-Ä movement of smooth museIe myosinon ADP release. Nawre, 378, 748-751.
Woodward,S.K.A., Geeves,M.A. and ManSlein,DJ. (1995) Kineticeharacteriza1ion of lhe ca1alytic domain of Dictyosteliwll di~icoidelllll
myosin. Biachelllis/ly, 34, 16056-16064.Xie,X., Harrison,D.H., Schlichting,I., Swee1,R.M., Kalabokis,VN.,
Szent-Gyorgyi,A.G. and Cohen.C. (1994) Strllcture 01' the regulatorydomain of seaUop myosin at 2.8 Aresolution. Natllre, 368, 306-312.
Yan, Y, Winograd,E., Viel,A., Cronin,T., Harrison,S.c. and Bramon,D.(1993) Crystal strllctllre of the repetitive segments 01' speetrin. Science,262, 2027-2030.
Zentgraf,l-l .. Frey,M., Schwinn.S., Tessmer,c., Willemann,B., Samstag, Yand Velhagen,I. (1995) Detection of histidine-1agged fusion proteinsby using a high-specific mouse monoclonat anti-histidine tag antibody.Nt/cleic Acids Res., 23. 3347-3348.
Received on June 4, /996; revised 0/1 July 3D, /996
.-
Related Documents











