Research Article Human Umbilical Cord Perivascular Cells Exhibited Enhanced Migration Capacity towards Hepatocellular Carcinoma in Comparison with Bone Marrow Mesenchymal Stromal Cells: A Role for Autocrine Motility Factor Receptor Juan Bayo, 1 Esteban Fiore, 1 Jorge B. Aquino, 1,2 Mariana Malvicini, 1 Manglio Rizzo, 1 Estanislao Peixoto, 1 Laura Alaniz, 1,2 Flavia Piccioni, 1 Marcela Bolontrade, 2,3 Osvaldo Podhajcer, 2,3 Mariana G. Garcia, 1,2 and Guillermo Mazzolini 1,2,4 1 Laboratorio de Terapia G´ enica, Facultad de Ciencias Biom´ edicas, Universidad Austral, B1629ODT Derqui-Pilar, Buenos Aires, Argentina 2 CONICET (Consejo Nacional de Investigaciones Cient´ ıficas y T´ ecnicas), C1033AAJ Buenos Aires, Argentina 3 Laboratorio de Terapia Molecular y Celular, Fundaci´ on Instituto Leloir, C1405BWE Buenos Aires, Argentina 4 Unidad de H´ ıgado, Hospital Universitario Austral, Universidad Austral, B1629ODT Derqui-Pilar, Argentina Correspondence should be addressed to Mariana G. Garcia; [email protected] and Guillermo Mazzolini; [email protected] Received 17 February 2014; Accepted 17 June 2014; Published 23 July 2014 Academic Editor: David Bernardo Ordiz Copyright © 2014 Juan Bayo et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Hepatocellular carcinoma (HCC) is the third cause of cancer-related death worldwide. Unfortunately, the incidence and mortality associated with HCC are increasing. erefore, new therapeutic strategies are urgently needed and the use of mesenchymal stromal cells (MSCs) as carrier of therapeutic genes is emerging as a promising option. Different sources of MSCs are being studied for cell therapy and bone marrow-derived cells are the most extensively explored; however, birth associated-tissues represent a very promising source. e aim of this work was to compare the in vitro and in vivo migration capacity between bone marrow MSCs (BM- MSCs) and human umbilical cord perivascular cells (HUCPVCs) towards HCC. We observed that HUCPVCs presented higher in vitro and in vivo migration towards factors released by HCC. e expression of autocrine motility factor (AMF) receptor, genes related with the availability of the receptor on the cell surface (caveolin-1 and -2) and metalloproteinase 3, induced by the receptor activation and important for cell migration, was increased in HUCPVCs. e chemotactic response towards recombinant AMF was increased in HUCPVCs compared to BM-MSCs, and its inhibition in the conditioned medium from HCC induced higher decrease in HUCPVC migration than in BM-MSC. Our results indicate that HUCPVCs could be a useful cellular source to deliver therapeutic genes to HCC. 1. Introduction Hepatocellular carcinoma (HCC) is one of the leading causes of cancer-related death worldwide. At early stages of the disease only a small fraction of patients are amenable to curative treatments that include surgical resection, liver transplantation, or radiofrequency ablation [1]. For patients with advanced HCC, the multikinase inhibitor sorafenib shows limited survival benefits in comparison with placebo [2]. us, the development of new therapeutic approaches is critical, and cellular therapy emerges as a new potential strategy to deliver therapeutic genes to HCC. Mesenchymal stromal cells (MSCs) constitute a heteroge- neous population of cells that include adult multipotent cells [3]. MSCs are present in adult tissues and their involvement in repair mechanisms has been demonstrated as the result of their ability to migrate towards sites of injury, their capacity to differentiate into tissues of mesodermal origin (adipocytes, Hindawi Publishing Corporation BioMed Research International Volume 2014, Article ID 837420, 9 pages http://dx.doi.org/10.1155/2014/837420

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Research ArticleHuman Umbilical Cord Perivascular Cells Exhibited EnhancedMigration Capacity towards Hepatocellular Carcinoma inComparison with Bone Marrow Mesenchymal Stromal CellsA Role for Autocrine Motility Factor Receptor
Juan Bayo1 Esteban Fiore1 Jorge B Aquino12 Mariana Malvicini1 Manglio Rizzo1
Estanislao Peixoto1 Laura Alaniz12 Flavia Piccioni1 Marcela Bolontrade23
Osvaldo Podhajcer23 Mariana G Garcia12 and Guillermo Mazzolini124
1 Laboratorio de Terapia Genica Facultad de Ciencias Biomedicas Universidad Austral B1629ODT Derqui-PilarBuenos Aires Argentina
2 CONICET (Consejo Nacional de Investigaciones Cientıficas y Tecnicas) C1033AAJ Buenos Aires Argentina3 Laboratorio de Terapia Molecular y Celular Fundacion Instituto Leloir C1405BWE Buenos Aires Argentina4Unidad de Hıgado Hospital Universitario Austral Universidad Austral B1629ODT Derqui-Pilar Argentina
Correspondence should be addressed to Mariana G Garcia mggarciaconicetgovar andGuillermo Mazzolini gmazzolicasaustraleduar
Received 17 February 2014 Accepted 17 June 2014 Published 23 July 2014
Academic Editor David Bernardo Ordiz
Copyright copy 2014 Juan Bayo et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Hepatocellular carcinoma (HCC) is the third cause of cancer-related death worldwide Unfortunately the incidence and mortalityassociated with HCC are increasingTherefore new therapeutic strategies are urgently needed and the use of mesenchymal stromalcells (MSCs) as carrier of therapeutic genes is emerging as a promising option Different sources of MSCs are being studied forcell therapy and bone marrow-derived cells are the most extensively explored however birth associated-tissues represent a verypromising sourceThe aimof thisworkwas to compare the in vitro and in vivomigration capacity between bonemarrowMSCs (BM-MSCs) and human umbilical cord perivascular cells (HUCPVCs) towards HCC We observed that HUCPVCs presented higher invitro and in vivo migration towards factors released by HCC The expression of autocrine motility factor (AMF) receptor genesrelated with the availability of the receptor on the cell surface (caveolin-1 and -2) and metalloproteinase 3 induced by the receptoractivation and important for cell migration was increased in HUCPVCs The chemotactic response towards recombinant AMFwas increased in HUCPVCs compared to BM-MSCs and its inhibition in the conditioned medium from HCC induced higherdecrease in HUCPVCmigration than in BM-MSC Our results indicate that HUCPVCs could be a useful cellular source to delivertherapeutic genes to HCC
1 Introduction
Hepatocellular carcinoma (HCC) is one of the leading causesof cancer-related death worldwide At early stages of thedisease only a small fraction of patients are amenable tocurative treatments that include surgical resection livertransplantation or radiofrequency ablation [1] For patientswith advanced HCC the multikinase inhibitor sorafenibshows limited survival benefits in comparison with placebo
[2] Thus the development of new therapeutic approachesis critical and cellular therapy emerges as a new potentialstrategy to deliver therapeutic genes to HCC
Mesenchymal stromal cells (MSCs) constitute a heteroge-neous population of cells that include adult multipotent cells[3] MSCs are present in adult tissues and their involvementin repair mechanisms has been demonstrated as the result oftheir ability to migrate towards sites of injury their capacityto differentiate into tissues ofmesodermal origin (adipocytes
Hindawi Publishing CorporationBioMed Research InternationalVolume 2014 Article ID 837420 9 pageshttpdxdoiorg1011552014837420
2 BioMed Research International
osteoblasts and chondroblasts) and their immunoregulatoryproperties [4] Moreover the possibility of easily expandingthese cells in vitro makes them useful tools for therapeuticuse in regenerative medicine immunomodulation purposesand also as cellular carriers for therapeutic genes [5]
MSCs can be isolated fromdifferent tissues Among thembone marrow- (BM-) derived stromal cells were the firstdescribed and are the most frequently used [6] However therequirement of invasive methods to obtain BM-MSCs is insupport of using alternative sources such as adipose tissueperipheral blood and dental pulp [7] In addition to theseadult tissues MSCs can be obtained from birth-associatedtissues including placenta amnion and umbilical cord withthe advantage of their availability avoiding the need ofinvasive procedures and eliminating other ethical concernsDifferent types of MSCs have been obtained from the umbil-ical cord depending on if they were isolated from the wholeumbilical cord (UC-MSCs) Whartonrsquos Jelly (WJ-MSCs) theumbilical cord blood (CB-MSCs) [8] or perivascular areas(human umbilical cord perivascular cells HUCPVCs) [9]In particular HUCPVCs may be advantageous candidatesfor cell therapy due to their lower donor variability fasterdoubling time and ready availability [10]
It has been demonstrated that MSCs from differentsources display similar phenotypic and cytological charac-teristics [11ndash13] however differences in their functional rolehave also been previously described For instance BM andWJ-derived MSCs have different proliferative capacity [11]and secretome and proteomic profiles [14 15]
The establishment and spread of a tumor is a processthat involves a complex cross-talk between cancer cells andthe tumor microenvironment HCC cells were found to bemodulated by the tumor milieu through the secretion ofseveral factors and the tumor cells modify their microen-vironment in order to promote their own growth [16] Par-ticularly MSCs express receptors for various cytokines andchemokines allowing them to migrate towards HCC tumors[17]Thus recruitment ofMSCs to cancer microenvironmentis likely mediated by the interaction of cytokineschemokinesand their specific receptors As an example it has beendemonstrated that CXCR1 is involved in UC-MSCmigrationtowards glioma [18] and that overexpression of CXCR1 [19] orCXCR4 in these cells [20] increases their homing into tumorsIn addition MSCs were engineered to express cytotoxiccytokines for treatment of lung tumors and non-Hodgkinrsquoslymphomas [21 22] Thus the possibility of increasingMSC migratory capacity and engraftment into tumors couldenhance beneficial effects of therapeutic genes
The aim of this work was to compare HUCPVCs andthe ldquogold standardrdquo BM-MSCs migratory capacity towardsfactors released from HCC and to analyze mechanismstherein involved
2 Materials and Methods
21 Cell Lines Human HCC cell line HuH7 was kindly pro-vided byProfessor Jesus Prieto (CIMAUniversity ofNavarraPamplona Spain) [23] LX-2 cell line (human hepatic stellatecells generated by spontaneous immortalization in low serum
conditions) was kindly provided by Dr Scott Friedman(Division of Liver Diseases Mount Sinai School of MedicineNew York NY USA) [24] Human microvascular endothe-lial cells (HMEC-1) were from CDC (Centers for DiseaseControl Atlanta GA USA) and WI-38 (human fibroblastcell line) from the American Type Culture Collection Celllines were cultured in complete DMEM (2 120583M glutamine100UmLpenicillin 100mgmL streptomycin) and 10heat-inactivated fetal bovine serum (FBS) Primary culture ofHCC cells (HC-PT-5) was previously generated in our lab-oratory [25] The collection of the sample and the projectwas approved by the Institutional Evaluation Committee(CIE) from School of Biomedical Sciences Austral University(Protocol no 11-007) and written informed consent wasobtained from the patient HC-PT-5 was cultured up to 8passages in 70 DMEM30 F12 (InvitrogenLife Technolo-gies) culture medium supplemented with 2 120583M glutamine100UmL penicillin 100mgmL streptomycin and 10FBS
22 Isolation of BM-MSCs and HUCPVCs BM-MSCs wereobtained from healthy donors (Hospital Naval Pedro MalloBuenos Aires Argentina) as described previously [25]HUCPVCs were isolated from umbilical cord obtainedfrom healthy donors at the Hospital Universitario Austral(Pilar Buenos Aires Argentina) using a protocol adaptedfrom Sarugaser et al [9] In brief umbilical cords weredissected and vessels with their surrounding WarthonrsquosJelly were pulled out The perivascular mesenchymal tissuewas removed from the vessels and mechanically disruptedMinced fragments were plated in complete DMEM lowglucose20FBS (Internegocios SA Argentina) After 7-dayincubation nonadherent cells and minced fragments wereremoved and adherentHUCPVCswere cultured and used fordifferent experiments at passages 4 to 6
MSCs were characterized according to the InternationalSociety for Cellular Therapy (ISCT) guidelines [26] Pheno-type characteristics ofMSCs were determined by flow cytom-etrywith anti-humanPE conjugated antibodies against CD14CD34 CD44 CD73 CD90 and CD105 (BD Biosciences)for 30min Samples were analyzed using a FACSCaliburflow cytometer (Becton Dickinson) and data acquired wereanalyzed using Cyflogic software (CyFlo Ltd)
23 Conditioned Medium To obtain tumor conditionedmedium (TCM) HuH7 cells (2 times 106) or HC-PT-5 cells (5times 106) were inoculated subcutaneously (sc) into the rightflank of nude mice When tumors reached 200mm3 in sizeapproximately tumors were dissected andminced into piecessmaller than 1mm3 and transferred to a 24-well tissue cultureplate (6 fragmentswell) with 500 120583L of complete DMEMwithout FBS Cell conditioned medium (CCM) was obtainedfrom cell lines cultured as described above Then cells thatreached a 90 of confluence were washed with PBS andcultured with complete DMEM without FBS In both cases18 hours later conditioned media were harvested and storedat minus80∘C until use
BioMed Research International 3
24 In Vitro Migration Assays In vitro migration was per-formed using a 48-Transwell microchemotaxis BoydenChamber unit (Neuroprobe Inc) as previously described[25] MSCs (12 times 103 cellswell) were placed in the upperchamber and DMEM CCM TCM or rAMF were appliedto the lower chamber of the transwell unit For blockingexperiments TCM were preincubated for 60min with anti-AMF antibody or isotype control IgG All systems were left at37∘C in a 5 CO
2humidified atmosphere for 4 hours except
for experiments involving rAMF that were maintained for18 hours Cells attached to the lower side of the membranewere fixed in 2 formaldehyde stained with 410158406-diamidino-2-phenylindole dihydrochloride (DAPI Sigma-Aldrich) andcounted using fluorescent-fieldmicroscopy at 100x Capturedimages from three representative visual fields were analyzedusing Cell Profiler software (httpwwwcellprofilercom)and the mean number of cellsfield plusmn SEM was calculated
25 Cell Adhesion Assays For analyses of MSC adhesionto endothelial cells 2 times 105 HMEC-1 were seeded in 96-well microplates and cultured for 1 day prior to the assayCoated wells were incubated for 5 minutes with 01mLof 5 times 104 cellsmL of Fast-DiO prelabelled MSCs Cellsuspension was discarded and attached cells were fixed with2 paraformaldehyde Cells were counted using fluorescent-field microscopy at 200x pictures taken from ten represen-tative visual fields were analyzed using Cell Profiler soft-ware (httpwwwcellprofilercom) and the mean number ofcellsfield plusmn SEM was calculated
26 Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Total RNA of MSCs was extracted using TrizolReagent (Sigma-Aldrich Co St Louis MO) Total RNA(4 120583g) was reverse-transcribed with 200U of SuperScriptII Reverse Transcriptase (Invitrogen Carlsbad CA) using500 ng of Oligo (dT) primers cDNAs were subjected toreal-time polymerase chain reaction (qPCR) (StratageneMx3005p Stratagene La Jolla CA USA) For qRT-PCRthe mRNA levels of CXCR1 CXCR2 CCR2 IL-6 receptor(IL-6R) IL-6 signal transducer (IL-6ST) AMF receptor(AMFR) metalloproteinase 3 (MMP3) insulin-like growthfactor-binding protein 3 (IGFBP3) caveolin-1 (CAV-1) andcaveolin-2 (CAV-2) were quantified by SYBR Green (Invitro-gen) using the following primers CXCR1 forward 51015840-TTT-TCCGCCAGGCTTACCAT-31015840 and reverse 51015840-AACACC-ATCCGCCATTTTGC-31015840 CXCR2 forward 51015840-TAAGTG-GAGCCCCGTGGGG-31015840 and reverse 51015840-TGGGCTCAG-GGGCAGGATG-31015840 CCR2 forward 51015840-CGAGAGCGG-TGAAGAAGTCA-31015840 and reverse 51015840-AGCATGTTGCCC-ACAAAACC-31015840 IL-6R forward 51015840-GCACTTGCTGGT-GGATGTTC-31015840 and reverse 51015840-AGCCTTTGTCGTCAG-GGATG-31015840 IL-6ST forward 51015840-CCCACCTCATGCACT-GTTGA-31015840 and reverse 51015840-TTATGTGGCGGATTCGGCTT-31015840 AMFR forward 51015840-ACAAGATGTGGGCCTTGCAAG-A-31015840 and reverse 51015840-AAAACGCAGTGCTCCCAGGATA-31015840 MMP3 forward 51015840-ACGCCAGCCAACTGTGATCCT-31015840 and reverse 51015840-ATATGCGGCATCCACGCCTGAA-31015840IGFBP3 forward 51015840-ACTGTGGCCATGACTGAG-31015840 and
reverse 51015840-AGAGTCTCCCTGAGCCTGA-31015840 CAV-1 for-ward 51015840-AATCCAAGCATCCCTTTGCCCA-31015840 and reverse51015840-ACCAGGCAGCTTTCTGTACGA-31015840 CAV-2 forward51015840-GAGAGACAGGGGAGTTGTCAACTT-31015840 and reverse51015840-GCCCGGCCCAGAAATAATGAGAT-31015840 PCR ampli-fications were carried out using a cycle of 95∘C for 10 minutesand 45 cycles under the following parameters 95∘C for 30seconds 58∘C for 60 seconds 72∘C for 30 seconds At the endof PCR reaction the temperature was increased from 60∘C to95∘C at a rate of 2∘Cmin and the fluorescence was measuredevery 15 seconds to construct the melting curve Values werenormalized to levels of glyceraldehyde-3-phosphate dehy-drogenase (GAPDH used as housekeeping) transcript (for-ward 51015840-CATCTCTGCCCCCTCTGCTG-31015840 and reverse 51015840-GCCTGCTTCACCACCTTCTTG-31015840) Data were processedby theΔΔCtmethodThe relative amount of the PCR productamplified fromBM-MSCswas set as 1 A nontemplate control(NTC) was run in every assay and all determinations wereperformed as triplicates in three separated experiments
27 Mice and In Vivo Experiments Six- to eight-week-old male nude mice (NuNu) were purchased from CNEA(Comision Nacional de Energıa Atomica Ezeiza BuenosAires Argentina) Subcutaneous HuH7 tumors (2 times 106cells)were established and 10 days later BM-MSCs or HUCPVCswere intravenously (iv) injected Tumor growthwas assessedby calliper measurement and tumor volume (mm3) wascalculated by the formula 1205876 times larger diameter times (smallerdiameter)2 For in vivo migration studies MSCs (5 times 105)were prelabeled with CM-DiI for histological analysis andDiR (Molecular Probes Invitrogen) for fluorescence imag-ing (FI) FI was performed using the Xenogen In VivoImaging System (IVIS Caliper Life Sciences HopkintonMA USA) 1 hour after MSC injection and every day untilexperimental end point At day 3 mice were sacrificedand isolated tissues were exposed to FI Images representthe radiant efficiency and were analyzed with IVIS LivingImage (Caliper Life Sciences) software Regions of inter-est (ROI) were automatically drawn around the isolatedorgans to assess the fluorescence signal emitted For thetotal signal present in mice results were expressed as totalradiant efficiency in units of photonssecond within theregion of interest [ps][120583Wcm2] Signal present in tumorliver spleen or lungs was expressed as percentage of totalsignal
28 Detection of BM-MSCs by Fluorescence To detect CM-DiI+ cells within tumors frozen sections were mounted inmounting media with DAPI (Vector Laboratories Inc) andobserved under a fluorescence microscope at 200x
29 Statistical Analyses Unpaired 2-sided Studentrsquos 119905-test andone-way analysis of variance followed by posttests orKruskal-Wallis and Dunnrsquos posttests (GraphPad Prism Software) wereused for statistical analyses 119875 values lower than 005 wereconsidered as statistically significant
4 BioMed Research International
HUCPVCs
BM-MSCs
CD73 CD90 CD105 CD44 CD14 CD34 CD799723 9290 9692 9772 066 074 090
9759 9833 9410 9621 095 073 096
FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H
FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H
(a)
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
BM-MSCsHUCPVCs
DMEM HuH7 HC-PT-5 LX-2 WI-38 HMEC-1
700
600
500
400
300
200
100
0
Num
ber o
f cel
lsfie
ld
(b)
lowastlowastlowast
BM-MSCsHUCPVCs
BM-MSCs HUCPVCs
Num
ber o
f cel
lsfie
ld
25
20
15
10
5
0
(c)
Figure 1 Characterization of BM-MSCs andHUCPVCs (a) Flow cytometry analysis of cell surfacemarkers of both types ofMSCs Grey areaindicates background fluorescence with IgG isotype control One representative experiment is shown (b) In vitro migration of BM-MSCs(black bars) or HUCPVCs (grey bars) towards CCM from HCC (HuH7 and HC-PT-5) hepatic stellate cells (LX-2) fibroblasts (WI-38)or endothelial cells (HMEC-1) Bars represent the average of MSCsfield (10x) plusmn SEM from three representative visual fields Results arerepresentative of 3 independent experiments 119875 lt 0001 versus DMEM lowastlowastlowast119875 lt 0001 versus BM-MSCs (c) Adhesion towards endothelialcells of BM-MSCs (black bars) or HUCPVCs (grey bars) Results are representative of 3 independent experiments lowastlowastlowast119875 lt 0001 versusBM-MSCs
3 Results
31 Characterization of BM-MSCs and HUCPVCs In accor-dance with the criteria for definingMSCs of the InternationalSociety for Cellular Therapy (ISCT) [26] surface markerexpression of BM-MSCs and HUCPVCs was evaluated byflow cytometry Both types of MSCs showed similar pheno-typic characteristics and were found to express CD73 CD90CD105 and CD44 but not to express the hematopoieticmarkersCD14CD34 orCD79 (Figure 1(a))Wenext decidedto evaluate the in vitro migration capacity of MSCs towardscell culture conditioned media (CCM) obtained from HCCcell lines (HuH7 and HC-PT-5) hepatic stellate cells (LX-2) fibroblasts (WI-38) and endothelial cells (HMEC-1)Interestingly a higher migratory capacity towards all theCCM was found for HUCPVCs when compared to BM-MSCs (Figure 1(b)) Moreover in contrast to our previousresults observed with BM-MCSs [25] HUCPVCs showedcapability to migrate to CCM derived from nontumoral
components (fibroblast and endothelial cells) Besides theircapacity to migrate toward factors secreted by HCC thearrest of MSCs within the microvasculature is considered acritical step for an efficient homing and anchorage to tumorsTherefore we next decided to evaluate adhesion ability ofMSCs and observed that HUCPVCs showed an increased invitro adhesion to HMEC-1 endothelial cells in comparisonwith BM-MSCs (Figure 1(c))
32 In Vivo Migration of BM-MSCs and HUCPVCs towardsHCC To further characterize MSC behavior in vivo non-invasive migration assay was performed CM-DiI and DiRprelabelled BM-MSCs or HUCPVCs were iv injected inHCC tumor-bearing mice in order to evaluate MSC recruit-ment Similar to our previous observation with BM-MSCs[25] at 3 days after cell transplantation a positive signal corre-sponding to HUCPVCs was found in liver lungs spleen andsc tumors (Figure 2(a)) Despite the fact that total signal was
BioMed Research International 5
50
40
30
20
10
times108
Radi
ant e
ffici
ency
(ps
cm
2s
r)(120583
Wc
m2)
(a)
lowastlowastlowast
BM-MSCsHUCPVCs
BM-MSCs HUCPVCsTota
l rad
iant
effici
ency
(ps
) (120583
Wc
m2) times1013
24
23
22
21
20
19
18
17
16
15
14
(b)
BM-MSCs HUCPVCs50
40
30
20
10
times108Ra
dian
t effi
cien
cy(p
sc
m2s
r)(120583
Wc
m2)
(c)
BM-MSCsHUCPVCs
Tota
l sig
nal (
) 90
8086420
Liver Lung Spleen Tumor
lowast
lowast
(d)
DAPI CM-DiI
(e)
BM-MSCsHUCPVCs
250
200
150
100
50
0HuH7 HC-PT-5
Num
ber o
f cel
lsfie
ld
lowastlowastlowast
lowastlowastlowast
(f)
Figure 2 In vivomigration of BM-MSCs and HUCPVCs CM-DiI and DiR prelabeled MSCs were iv injected in sc HuH7 tumor-bearingmice At day 3 mice were sacrificed and organs were removed lungs livers spleen (a) and tumors (c) were exposed to obtain FI Imagesrepresent the radiant efficiency Representative images are shown (b) Total FI for injected BM-MSCs or HUCPVCs was calculated bymeasuring the region of interest (ROI) for all the tissues isolated and results were expressed as total radiant efficiency [ps][120583Wcm2]lowastlowastlowast119875 lt 0001 (d) Signal present in the isolated liver spleen lungs and tumors was represented as percentage of total signal for BM-MSCs or
HUCPVCs-injected mice lowast119875 lt 005 versus BM-MSCs (e) Microscopic analysis of transplanted CM-DiI-labeled MSCs (red signal indicatedby arrows) andDAPI staining in frozen sections of tumors times200magnification (f) In vitromigration ofMSCs to TCMderived fromHuH7 orHC-PT-5 sc tumors Bars represent the average of MSCsfield (10x) plusmn SEM from three representative visual fields Results are representativeof 3 independent experiments lowastlowastlowast119875 lt 0001 versus BM-MSCs
6 BioMed Research International
BM-MSCsHUCPVCs
lowastlowastlowast
lowastlowastlowast
lowast20
15
10
05
00
Fold
chan
ge (2
ΔΔ
ct)
CXCR1 CXCR2 CCR2 IL-6R IL-6ST
(a)
BM-MSCsHUCPVCs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
300
2705
4
3
2
1
0
Fold
chan
ge (2
ΔΔ
ct)
AMFR MMP3 CAV-1 CAV-2 IGFBP3
(b)
Figure 3 Differential expression of cytokineschemokines receptors and AMFAMFR pathway in MSCs Expression of cytokines andchemokines receptors (a) and AMFAMFR axis proteins (b) was evaluated in BM-MSCs (black bars) or HUCPVCs (grey bars) by qPCRlowastlowastlowast119875 lt 0001 versus BM-MSCs
lower in mice injected with HUCPVCs compared to thoseinjected with BM-MSCs (Figure 2(b)) the percentage of totalsignal corresponding to sc tumor locations was increasedin mice administered with HUCPVCs in comparison withanimals that received BM-MSCs (Figures 2(c) and 2(d))indicating an enhanced engraftment of HUCPVCs into HCCtumors In the other evaluated tissues signal intensity wassimilar for BM-MSC or HUCPVCs in lung and liver andit was comparatively reduced in the spleen of HUCPVCs-injected mice (Figure 2(d)) Presence of MSCs in the sctumors was also confirmed by fluorescence microscopy(Figure 2(e)) Finally we evaluated whether MSCs mightpresent differential migratory capacity towards CM obtainedfrom sc tumors (TCM) Interestingly a greater in vitromigratory capacity towards TCM from HCC was observedfor HUCPVCs when compared to BM-MSCs (Figure 2(f))
33 AMFR Was Highly Expressed in HUCPVCs In orderto evaluate mechanisms partially explaining the differentialmigratory capacity of HUCPVCs compared to BM-MSCstowards tumor released factors we analyzed the expressionof some chemokinersquos receptor likely involved inMSC recruit-ment towards HCC Taking into account several reportsdemonstrating that interleukin- (IL-) 8 GRO chemokine(C-C motif) ligand (CCL)-2 and IL-6 are among the mostrelevant factors in HCC [17] we decided to evaluate by qPCRthe expression of CXCR1 CXCR2 CCR2 IL-6R and IL-6ST Interestingly constitutive CXCR1 and CXCR2 mRNAexpression was found to be lower and CCR2 slightly higherin HUCPVCs when compared to BM-MSCs while IL-6Rand IL-6ST expression was similar in both MSCs sources(Figure 3(a)) Next we decided to evaluate the axis of theautocrinemotility factor (AMF) a cytokine with chemotacticeffect on MSCs as recently observed by our group [27]By qPCR a significantly higher expression of the AMFreceptor (AMFR) was found in HUCPVCs when comparedto BM-MSCs Similarly genes known to be related to the
availability of the receptor in the cell surface such as caveolin-1 (CAV-1) and caveolin-2 (CAV-2) were also highly expressedin HUCPVCs as well as the metalloproteinase 3 (MMP3)necessary to the transmigration process In contrast expres-sion levels of insulin-like growth factor-binding protein 3(IGFBP3) a protein that negatively regulates AMFAMFRpathway were found to be reduced in HUCPVCs whencompared to BM-MSCs (Figure 3(b))
34 HUCPVCs Displayed Enhanced Migration towards AMFIn Vitro We have previously demonstrated that AMF acytokine produced by HCC cells plays a critical role inMSC migration [27] Due to the increased AMFR and AMF-AMFR-related proteins expression in HUCPVCs we decidedto test the in vitro migration response to the recombinantAMF (rAMF) of both types ofMSCsusing a chemotaxis assayA significantly higher migration to different doses of rAMF(05 and 075 120583gmL) was observed for HUCPVCs whencompared to BM-MSCs (Figure 4(a)) In spite of differenttypes of MSCs showing similar reduction in migration levels(50 of control) towards HuH7 TCM after the blockage withanti-AMF antibody (data not shown) preincubation of HC-PT-5 TCM with anti-AMF antibody (AMF-ab) resulted ina further reduction in HUCPVCs migration capacity (54of control) when compared to BM-MSCs (67 of control)(Figure 4(b))
4 Discussion
The aim of this study was to assess the capability of differentsources of MSCs to migrate towards HCC released factorsin order to select those showing higher tumor recruitmentcapacity for future therapeutic applications Our resultsindicated that although both types of MSCs share simi-lar phenotypic characteristics as MSCs HUCPVCs have ahigher potential to migrate in vitro towards the conditionedmedium derived not only from HCC cells but also from
BioMed Research International 7
BM-MSCsHUCPVCs
lowastlowastlowast
0 050 075
rAMF (120583gmL)
300
250
200
150
100
50
0
Con
trol (
)
(a)
BM-MSCsHUCPVCs
lowast
Con
trol (
)
110
90
100
70
50
30
10
80
60
40
20
0+ minus + minus
+minus +minus
IgG-abAMF-ab
(b)
Figure 4 HUCPVCs showed enhanced migration towards AMF in comparison with BM-MSCs (a) In vitromigration of BM-MSCs (blackbars) or HUCPVCs (grey bars) towards rAMF 119875 lt 005 versus DMEM (0 120583gmL rAMF) lowast119875 lt 005 versus BM-MSCs (b) In vitromigrationof BM-MSCs (black bars) or HUCPVCs (grey bars) towards HC-PT-5 TCM preincubated with anti-AMF antibody (AMF-ab) or controlisotype (IgG-ab) was evaluated 119875 lt 005 versus IgG-ab lowast119875 lt 005 versus BM-MSCs Bars represent the average of MSCsfield (10x) plusmn SEMfrom three representative visual fields Results are representative of 3 independent experiments
nontumoral components of the tumor microenvironmentlike activated hepatic stellate cells (LX-2) fibroblasts (WI-38) or tumor microendothelial cells (HMEC-1) MoreoverHUCPVCs also showed an enhanced adhesion capacityto HMEC-1 cells compared to BM-MSCs This enhancedadhesion to endothelial cells could allow a more efficientarrest in tumor microvasculature which is a required step forMSC recruitment into HCC In line with this we observedthat HUCPVCs also exhibited greater migration capacity invivo towards experimental HCC In contrast to that observedin the in vitro migration experiments HUCPVCs showed asimilar migration capability towards nontumoral tissues likeliver or lung and also showed a lower recruitment to thespleen AlthoughHUCPVCsmigrated better than BM-MSCstoward activated hepatic stellate cells in vitro no differenceswere observed in in vivo migration to normal liver wherehepatic stellate cells are quiescent In addition it should benoted that mice injected with HUCPVCs showed a lowertotal signal than those injected with BM-MSCs probablydue to differences in the DiR uptake for both kinds of cellsor in the number of inoculated cells These results are inagreement with previous works showing that MCSs derivedfrom umbilical cord are able tomigrate towards experimentaltumors such as glioma [28] breast cancer [29] and lung ade-nocarcinoma [30] Nonetheless to the best of our knowledgethis is the first report describing the comparative migrationcapacity of human umbilical cord perivascular cells towardshuman HCC
Tumor-homing is thought to be mediated by interactionsof cytokineschemokines with their specific receptors Anumber of cytokines growth factors and chemokines aresecreted by HCC cells and their microenvironment [17] Wehave recently found that AMF is critical for MSC migrationtoward HCC in vitro and in vivo Furthermore other factorsreleased by tumor stroma components like IL-8 GRO IL-6or CCL-2 could also be involved inMSC recruitment toHCCThus differences observed between HUCPVCs and BM-MSCs in their in vitro and in vivomigration capacity towardsHCC seem at least partially related to differences in theexpression of cytokinechemokine receptors Some studieshave screened for similarities or differences among BM-MSCs and UC-MSCs at the molecular level [31 32] In addi-tion a previous report demonstrated that migration of UCB-MSC towards glioma cells was higher when compared toBM-MSCs likely due to increased levels of the IL-8 receptorCXCR1 and CXCR2 in the former cell type [18] In contrastto these observations we found that HUCPVCs presentedlower levels of CXCR1 and CXCR2 expression compared toBM-MSCs and these differences could be explained partiallydue to differences in tissue source and culture conditions andmerit further investigationThe analysis of IL-6R and IL6-STexpression showed similar levels in both types ofMSCs whilelevels of MCP-1 receptor (CCR2) were slightly increased inHUCPVCs Interestingly we found that not only AMFR butalso CAV-1 and CAV-2 which are genes that regulate theavailability of AMFR on cell surface [33] were overexpressedin HUCPVCs compared to BM-MSCs In line with this a
8 BioMed Research International
reduced expression level of IGFBP3 a protein which canbind to AMF blocking its binding to AMFR [34] was foundin HUCPVCs when compared to BM-MSCs Moreover anincreased expression of MMP3 was observed in HUCPVCsThis matrix metalloproteinase is involved in AMF-inducedmigration and it is necessary for cell invasion and proteolysisof the extracellularmatrix [35] Finally and consistentwith anincrease in the expression of genes related to the AMFAMFRpathway not only HUCPVCs show an enhanced in vitromigration towards rAMF but also the specific blockageof AMF in HC-PT-5 TCM was able to reduce HUCPVCsmigration to a greater level when compared to BM-MSCs
The potential use of MSCs in cancer treatment as carriersof therapeutic genes has raised some concerns about thesafety of their use in the clinic There are some studiesthat indicated that MSCs have protumorigenic capacitydue to their immunosuppression properties modulation ofthe epithelial-to-mesenchymal transition and induction ofangiogenesis [36] However others have demonstrated thatMSCs decreased HCC growth [37 38] Similarly we havepreviously demonstrated that BM-MSCs did not modifytumor growth in vivo [25] Moreover we observed thatHUCPVCs did not induce tumor growth when injected ivin Huh7 sc tumor-bearing mice (data not shown) Theuse of HUCPVCs as cellular carriers could also have theadvantage of its allogeneic application because of its lowimmunogenicity [9] In this regard HUCPVCs could beuseful as carriers of therapeutic genes for cancer patientswhere isolation of BM or adipose tissue-derived MSCs couldbe less responsive to chemotactic factors and might dis-play higher immunosuppression capacities than MSCs fromhealthy donors [39] or even present genetic abnormalities
5 Conclusions
In summary our results demonstrate a greater migrationcapacity to HCC of HUCPVCs when compared to BM-MSCs not only in vitro but also in vivo likely due to anincrease in migratory response to AMF and to an enhancedadhesive capacity to tumor microvasculature Consideringtheir availability and that no invasive procedures are requiredto obtain HUCPVCs these cells have advantages over BM-MSCs as candidates for carriers of therapeutic genes for thetreatment of HCC
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors gratefully thank Guillermo Gaston MarcosCabrera and Vanina Ferreira for technical assistance andDr Mengshiao Meng (National Chung Hsing University(NCHU) Taichung Taiwan) for providing recombinantAMF This work was supported by Grants from Austral
University (for Juan Bayo I04-12 for Mariana G Garcia T13-12 and for Guillermo Mazzolini T13-11) and from AgenciaNacional de Promocion Cientıfica y Tecnologica (ANPCyT)PICTO 200800115 (Mariana G Garcia) PICT 200800123(Jorge B Aquino) PICTO 200800122 (Jorge B Aquino)and PICT 20102818 (Mariana G Garcia and GuillermoMazzolini)
References
[1] H B El-Serag J A Marrero L Rudolph and K R ReddyldquoDiagnosis and treatment of hepatocellular carcinomardquo Gas-troenterology vol 134 no 6 pp 1752ndash1763 2008
[2] J M Llovet S Ricci V Mazzaferro et al ldquoSorafenib inadvanced hepatocellular carcinomardquo The New England Journalof Medicine vol 359 no 4 pp 378ndash390 2008
[3] A J Friedenstein J F Gorskaja andN N Kulagina ldquoFibroblastprecursors in normal and irradiated mouse hematopoieticorgansrdquo Experimental Hematology vol 4 no 5 pp 267ndash2741976
[4] D J Prockop and J Y Oh ldquoMedical therapies with adultstemprogenitor cells (MSCs) a backward journey from dra-matic results in vivo to the cellular andmolecular explanationsrdquoJournal of Cellular Biochemistry vol 113 no 5 pp 1460ndash14692012
[5] J B Aquino M F Bolontrade M G Garcıa O L Podhajcerand GMazzolini ldquoMesenchymal stem cells as therapeutic toolsand gene carriers in liver fibrosis and hepatocellular carcinomardquoGene Therapy vol 17 no 6 pp 692ndash708 2010
[6] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[7] H Yoshimura T Muneta A Nimura A Yokoyama H Kogaand I Sekiya ldquoComparison of rat mesenchymal stem cellsderived from bone marrow synovium periosteum adiposetissue and musclerdquo Cell and Tissue Research vol 327 no 3 pp449ndash462 2007
[8] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popu-lations and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[9] R Sarugaser D Lickorish D Baksh M M Hosseini and J EDavies ldquoHuman umbilical cord perivascular (HUCPV) cells asource of mesenchymal progenitorsrdquo Stem Cells vol 23 no 2pp 220ndash229 2005
[10] D Baksh R Yao and R S Tuan ldquoComparison of proliferativeand multilineage differentiation potential of human mesenchy-mal stem cells derived from umbilical cord and bone marrowrdquoStem Cells vol 25 no 6 pp 1384ndash1392 2007
[11] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[12] O K Lee T K Kuo W M Chen K D Lee S L Hsieh andT H Chen ldquoIsolation of multipotent mesenchymal stem cellsfrom umbilical cord bloodrdquo Blood vol 103 no 5 pp 1669ndash16752004
[13] HWang S Hung S Peng et al ldquoMesenchymal stem cells in theWhartonrsquos jelly of the human umbilical cordrdquo StemCells vol 22no 7 pp 1330ndash1337 2004
BioMed Research International 9
[14] J Kim J M Shin Y J Jeon H M Chung and J Chae ldquoPro-teomic validation of multifunctional molecules in mesenchy-mal stem cells derived from human bone marrow umbilicalcord blood and peripheral bloodrdquo PLoS ONE vol 7 no 5Article ID e32350 2012
[15] J-Y Hsieh H-W Wang S-J Chang et al ldquoMesenchymalstem cells from human umbilical cord express preferentiallysecreted factors related to neuroprotection neurogenesis andangiogenesisrdquo PLoS ONE vol 8 no 8 Article ID e72604 2013
[16] V Hernandez-Gea S Toffanin S L Friedman and J MLlovet ldquoRole of the microenvironment in the pathogenesis andtreatment of hepatocellular carcinomardquo Gastroenterology vol144 no 3 pp 512ndash527 2013
[17] J Bayo M Marrodan J B Aquino M Silva M G Garciaand G Mazzolini ldquoThe therapeutic potential of bone marrow-derived mesenchymal stromal cells on hepatocellular carci-nomardquo Liver International vol 34 no 3 pp 330ndash334 2014
[18] D Kim J H Kim J Kwon Lee et al ldquoOverexpression ofCXC chemokine receptors is required for the superior glioma-tracking property of umbilical cord blood-derived mesenchy-mal stem cellsrdquo Stem Cells and Development vol 18 no 3 pp511ndash519 2009
[19] S M Kim D Kim C H Jeong et al ldquoCXC chemokinereceptor 1 enhances the ability of human umbilical cord blood-derived mesenchymal stem cells to migrate toward gliomasrdquoBiochemical and Biophysical Research Communications vol 407no 4 pp 741ndash746 2011
[20] S A Park C H Ryu S M Kim et al ldquoCXCR4-transfectedhuman umbilical cord blood-derived mesenchymal stem cellsexhibit enhanced migratory capacity toward gliomasrdquo Interna-tional Journal of Oncology vol 38 no 1 pp 97ndash103 2011
[21] R S Rachakatla F Marini M L Weiss M Tamura and DTroyer ldquoDevelopment of human umbilical cord matrix stemcell-based gene therapy for experimental lung tumorsrdquo CancerGene Therapy vol 14 no 10 pp 828ndash835 2007
[22] C Yan S Li Z Li et al ldquoHuman umbilical cord mesenchymalstem cells as vehicles of CD20-specific TRAIL fusion proteindelivery a double-target therapy against non-Hodgkinrsquos lym-phomardquo Molecular Pharmaceutics vol 10 no 1 pp 142ndash1512013
[23] J Hu A Dong V Fernandez-Ruiz et al ldquoBlockade of Wntsignaling inhibits angiogenesis and tumor growth in hepatocel-lular carcinomardquo Cancer Research vol 69 no 17 pp 6951ndash69592009
[24] L Xu A Y Hui E Albanis et al ldquoHuman hepatic stellate celllines LX-1 and LX-2 new tools for analysis of hepatic fibrosisrdquoGut vol 54 no 1 pp 142ndash151 2005
[25] M G Garcia J Bayo M F Bolontrade et al ldquoHepatocellularcarcinoma cells and their fibrotic microenvironment modulatebone marrow-derived mesenchymal stromal cell migration invitro and in vivordquo Molecular Pharmaceutics vol 8 no 5 pp1538ndash1548 2011
[26] M Dominici K le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells theInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[27] J Bayo E Fiore J B Aquino et al ldquoIncreased migration ofhumanmesenchymal stromal cells byAutocrineMotility Factor(AMF) resulted in enhanced recruitment towards hepatocellu-lar carcinomardquo PLoS ONE vol 9 no 4 Article ID e95171 2014
[28] M K Seong Y L Jung I P Sang et al ldquoGene therapyusing TRAIL-secreting human umbilical cord blood-derived
mesenchymal stem cells against intracranial gliomardquo CancerResearch vol 68 no 23 pp 9614ndash9623 2008
[29] R Ayuzawa C Doi R S Rachakatla et al ldquoNaıve humanumbilical cord matrix derived stem cells significantly attenuategrowth of human breast cancer cells in vitro and in vivordquoCancerLetters vol 280 no 1 pp 31ndash37 2009
[30] T Matsuzuka R S Rachakatla C Doi et al ldquoHuman umbilicalcord matrix-derived stem cells expressing interferon-120573 genesignificantly attenuate bronchioloalveolar carcinomaxenograftsin SCID micerdquo Lung Cancer vol 70 no 1 pp 28ndash36 2010
[31] G Li X Zhang H Wang et al ldquoComparative proteomicanalysis of mesenchymal stem cells derived from human bonemarrow umbilical cord and placenta implication in themigra-tionrdquo Proteomics vol 9 no 1 pp 20ndash30 2009
[32] H C Miranda R H Herai C H Thome et al ldquoA quan-titative proteomic and transcriptomic comparison of humanmesenchymal stem cells from bone marrow and umbilical cordveinrdquo Proteomics vol 12 no 17 pp 2607ndash2617 2012
[33] P U Le G Guay Y Altschuler and I R Nabi ldquoCaveolin-1 isa negative regulator of caveolae-mediated endocytosis to theendoplasmic reticulumrdquo The Journal of Biological Chemistryvol 277 no 5 pp 3371ndash3379 2002
[34] S Mishra A Raz and L J Murphy ldquoInsulin-like growthfactor binding protein-3 interacts with autocrine motility fac-torphosphoglucose isomerase (AMFPGI) and inhibits theAMFPGI functionrdquo Cancer Research vol 64 no 7 pp 2516ndash2522 2004
[35] F L Yu M H Liao J W Lee and W L Shih ldquoInductionof hepatoma cells migration by phosphoglucose isomeraseautocrine motility factor through the upregulation of matrixmetalloproteinase-3rdquo Biochemical and Biophysical ResearchCommunications vol 314 no 1 pp 76ndash82 2004
[36] A H Klopp A Gupta E Spaeth M Andreeff and F MariniIII ldquoConcise review dissecting a discrepancy in the literaturedomesenchymal stem cells support or suppress tumor growthrdquoStem Cells vol 29 no 1 pp 11ndash19 2011
[37] L Qiao Z Xu T Zhao et al ldquoSuppression of tumorigenesisby human mesenchymal stem cells in a hepatoma modelrdquo CellResearch vol 18 no 4 pp 500ndash507 2008
[38] Y R Lu Y Yuan X J Wang et al ldquoThe growth inhibitory effectof mesenchymal stem cells on tumor cells in vitro and in vivordquoCancer Biology andTherapy vol 7 no 2 pp 245ndash251 2008
[39] Z-G Zhao W Xu L Sun W-M Li Q-P Li and P Zou ldquoThecharacteristics and immunoregulatory functions of regulatorydendritic cells induced by mesenchymal stem cells derivedfrom bone marrow of patient with chronic myeloid leukaemiardquoEuropean Journal of Cancer vol 48 no 12 pp 1884ndash1895 2012

2 BioMed Research International
osteoblasts and chondroblasts) and their immunoregulatoryproperties [4] Moreover the possibility of easily expandingthese cells in vitro makes them useful tools for therapeuticuse in regenerative medicine immunomodulation purposesand also as cellular carriers for therapeutic genes [5]
MSCs can be isolated fromdifferent tissues Among thembone marrow- (BM-) derived stromal cells were the firstdescribed and are the most frequently used [6] However therequirement of invasive methods to obtain BM-MSCs is insupport of using alternative sources such as adipose tissueperipheral blood and dental pulp [7] In addition to theseadult tissues MSCs can be obtained from birth-associatedtissues including placenta amnion and umbilical cord withthe advantage of their availability avoiding the need ofinvasive procedures and eliminating other ethical concernsDifferent types of MSCs have been obtained from the umbil-ical cord depending on if they were isolated from the wholeumbilical cord (UC-MSCs) Whartonrsquos Jelly (WJ-MSCs) theumbilical cord blood (CB-MSCs) [8] or perivascular areas(human umbilical cord perivascular cells HUCPVCs) [9]In particular HUCPVCs may be advantageous candidatesfor cell therapy due to their lower donor variability fasterdoubling time and ready availability [10]
It has been demonstrated that MSCs from differentsources display similar phenotypic and cytological charac-teristics [11ndash13] however differences in their functional rolehave also been previously described For instance BM andWJ-derived MSCs have different proliferative capacity [11]and secretome and proteomic profiles [14 15]
The establishment and spread of a tumor is a processthat involves a complex cross-talk between cancer cells andthe tumor microenvironment HCC cells were found to bemodulated by the tumor milieu through the secretion ofseveral factors and the tumor cells modify their microen-vironment in order to promote their own growth [16] Par-ticularly MSCs express receptors for various cytokines andchemokines allowing them to migrate towards HCC tumors[17]Thus recruitment ofMSCs to cancer microenvironmentis likely mediated by the interaction of cytokineschemokinesand their specific receptors As an example it has beendemonstrated that CXCR1 is involved in UC-MSCmigrationtowards glioma [18] and that overexpression of CXCR1 [19] orCXCR4 in these cells [20] increases their homing into tumorsIn addition MSCs were engineered to express cytotoxiccytokines for treatment of lung tumors and non-Hodgkinrsquoslymphomas [21 22] Thus the possibility of increasingMSC migratory capacity and engraftment into tumors couldenhance beneficial effects of therapeutic genes
The aim of this work was to compare HUCPVCs andthe ldquogold standardrdquo BM-MSCs migratory capacity towardsfactors released from HCC and to analyze mechanismstherein involved
2 Materials and Methods
21 Cell Lines Human HCC cell line HuH7 was kindly pro-vided byProfessor Jesus Prieto (CIMAUniversity ofNavarraPamplona Spain) [23] LX-2 cell line (human hepatic stellatecells generated by spontaneous immortalization in low serum
conditions) was kindly provided by Dr Scott Friedman(Division of Liver Diseases Mount Sinai School of MedicineNew York NY USA) [24] Human microvascular endothe-lial cells (HMEC-1) were from CDC (Centers for DiseaseControl Atlanta GA USA) and WI-38 (human fibroblastcell line) from the American Type Culture Collection Celllines were cultured in complete DMEM (2 120583M glutamine100UmLpenicillin 100mgmL streptomycin) and 10heat-inactivated fetal bovine serum (FBS) Primary culture ofHCC cells (HC-PT-5) was previously generated in our lab-oratory [25] The collection of the sample and the projectwas approved by the Institutional Evaluation Committee(CIE) from School of Biomedical Sciences Austral University(Protocol no 11-007) and written informed consent wasobtained from the patient HC-PT-5 was cultured up to 8passages in 70 DMEM30 F12 (InvitrogenLife Technolo-gies) culture medium supplemented with 2 120583M glutamine100UmL penicillin 100mgmL streptomycin and 10FBS
22 Isolation of BM-MSCs and HUCPVCs BM-MSCs wereobtained from healthy donors (Hospital Naval Pedro MalloBuenos Aires Argentina) as described previously [25]HUCPVCs were isolated from umbilical cord obtainedfrom healthy donors at the Hospital Universitario Austral(Pilar Buenos Aires Argentina) using a protocol adaptedfrom Sarugaser et al [9] In brief umbilical cords weredissected and vessels with their surrounding WarthonrsquosJelly were pulled out The perivascular mesenchymal tissuewas removed from the vessels and mechanically disruptedMinced fragments were plated in complete DMEM lowglucose20FBS (Internegocios SA Argentina) After 7-dayincubation nonadherent cells and minced fragments wereremoved and adherentHUCPVCswere cultured and used fordifferent experiments at passages 4 to 6
MSCs were characterized according to the InternationalSociety for Cellular Therapy (ISCT) guidelines [26] Pheno-type characteristics ofMSCs were determined by flow cytom-etrywith anti-humanPE conjugated antibodies against CD14CD34 CD44 CD73 CD90 and CD105 (BD Biosciences)for 30min Samples were analyzed using a FACSCaliburflow cytometer (Becton Dickinson) and data acquired wereanalyzed using Cyflogic software (CyFlo Ltd)
23 Conditioned Medium To obtain tumor conditionedmedium (TCM) HuH7 cells (2 times 106) or HC-PT-5 cells (5times 106) were inoculated subcutaneously (sc) into the rightflank of nude mice When tumors reached 200mm3 in sizeapproximately tumors were dissected andminced into piecessmaller than 1mm3 and transferred to a 24-well tissue cultureplate (6 fragmentswell) with 500 120583L of complete DMEMwithout FBS Cell conditioned medium (CCM) was obtainedfrom cell lines cultured as described above Then cells thatreached a 90 of confluence were washed with PBS andcultured with complete DMEM without FBS In both cases18 hours later conditioned media were harvested and storedat minus80∘C until use
BioMed Research International 3
24 In Vitro Migration Assays In vitro migration was per-formed using a 48-Transwell microchemotaxis BoydenChamber unit (Neuroprobe Inc) as previously described[25] MSCs (12 times 103 cellswell) were placed in the upperchamber and DMEM CCM TCM or rAMF were appliedto the lower chamber of the transwell unit For blockingexperiments TCM were preincubated for 60min with anti-AMF antibody or isotype control IgG All systems were left at37∘C in a 5 CO
2humidified atmosphere for 4 hours except
for experiments involving rAMF that were maintained for18 hours Cells attached to the lower side of the membranewere fixed in 2 formaldehyde stained with 410158406-diamidino-2-phenylindole dihydrochloride (DAPI Sigma-Aldrich) andcounted using fluorescent-fieldmicroscopy at 100x Capturedimages from three representative visual fields were analyzedusing Cell Profiler software (httpwwwcellprofilercom)and the mean number of cellsfield plusmn SEM was calculated
25 Cell Adhesion Assays For analyses of MSC adhesionto endothelial cells 2 times 105 HMEC-1 were seeded in 96-well microplates and cultured for 1 day prior to the assayCoated wells were incubated for 5 minutes with 01mLof 5 times 104 cellsmL of Fast-DiO prelabelled MSCs Cellsuspension was discarded and attached cells were fixed with2 paraformaldehyde Cells were counted using fluorescent-field microscopy at 200x pictures taken from ten represen-tative visual fields were analyzed using Cell Profiler soft-ware (httpwwwcellprofilercom) and the mean number ofcellsfield plusmn SEM was calculated
26 Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Total RNA of MSCs was extracted using TrizolReagent (Sigma-Aldrich Co St Louis MO) Total RNA(4 120583g) was reverse-transcribed with 200U of SuperScriptII Reverse Transcriptase (Invitrogen Carlsbad CA) using500 ng of Oligo (dT) primers cDNAs were subjected toreal-time polymerase chain reaction (qPCR) (StratageneMx3005p Stratagene La Jolla CA USA) For qRT-PCRthe mRNA levels of CXCR1 CXCR2 CCR2 IL-6 receptor(IL-6R) IL-6 signal transducer (IL-6ST) AMF receptor(AMFR) metalloproteinase 3 (MMP3) insulin-like growthfactor-binding protein 3 (IGFBP3) caveolin-1 (CAV-1) andcaveolin-2 (CAV-2) were quantified by SYBR Green (Invitro-gen) using the following primers CXCR1 forward 51015840-TTT-TCCGCCAGGCTTACCAT-31015840 and reverse 51015840-AACACC-ATCCGCCATTTTGC-31015840 CXCR2 forward 51015840-TAAGTG-GAGCCCCGTGGGG-31015840 and reverse 51015840-TGGGCTCAG-GGGCAGGATG-31015840 CCR2 forward 51015840-CGAGAGCGG-TGAAGAAGTCA-31015840 and reverse 51015840-AGCATGTTGCCC-ACAAAACC-31015840 IL-6R forward 51015840-GCACTTGCTGGT-GGATGTTC-31015840 and reverse 51015840-AGCCTTTGTCGTCAG-GGATG-31015840 IL-6ST forward 51015840-CCCACCTCATGCACT-GTTGA-31015840 and reverse 51015840-TTATGTGGCGGATTCGGCTT-31015840 AMFR forward 51015840-ACAAGATGTGGGCCTTGCAAG-A-31015840 and reverse 51015840-AAAACGCAGTGCTCCCAGGATA-31015840 MMP3 forward 51015840-ACGCCAGCCAACTGTGATCCT-31015840 and reverse 51015840-ATATGCGGCATCCACGCCTGAA-31015840IGFBP3 forward 51015840-ACTGTGGCCATGACTGAG-31015840 and
reverse 51015840-AGAGTCTCCCTGAGCCTGA-31015840 CAV-1 for-ward 51015840-AATCCAAGCATCCCTTTGCCCA-31015840 and reverse51015840-ACCAGGCAGCTTTCTGTACGA-31015840 CAV-2 forward51015840-GAGAGACAGGGGAGTTGTCAACTT-31015840 and reverse51015840-GCCCGGCCCAGAAATAATGAGAT-31015840 PCR ampli-fications were carried out using a cycle of 95∘C for 10 minutesand 45 cycles under the following parameters 95∘C for 30seconds 58∘C for 60 seconds 72∘C for 30 seconds At the endof PCR reaction the temperature was increased from 60∘C to95∘C at a rate of 2∘Cmin and the fluorescence was measuredevery 15 seconds to construct the melting curve Values werenormalized to levels of glyceraldehyde-3-phosphate dehy-drogenase (GAPDH used as housekeeping) transcript (for-ward 51015840-CATCTCTGCCCCCTCTGCTG-31015840 and reverse 51015840-GCCTGCTTCACCACCTTCTTG-31015840) Data were processedby theΔΔCtmethodThe relative amount of the PCR productamplified fromBM-MSCswas set as 1 A nontemplate control(NTC) was run in every assay and all determinations wereperformed as triplicates in three separated experiments
27 Mice and In Vivo Experiments Six- to eight-week-old male nude mice (NuNu) were purchased from CNEA(Comision Nacional de Energıa Atomica Ezeiza BuenosAires Argentina) Subcutaneous HuH7 tumors (2 times 106cells)were established and 10 days later BM-MSCs or HUCPVCswere intravenously (iv) injected Tumor growthwas assessedby calliper measurement and tumor volume (mm3) wascalculated by the formula 1205876 times larger diameter times (smallerdiameter)2 For in vivo migration studies MSCs (5 times 105)were prelabeled with CM-DiI for histological analysis andDiR (Molecular Probes Invitrogen) for fluorescence imag-ing (FI) FI was performed using the Xenogen In VivoImaging System (IVIS Caliper Life Sciences HopkintonMA USA) 1 hour after MSC injection and every day untilexperimental end point At day 3 mice were sacrificedand isolated tissues were exposed to FI Images representthe radiant efficiency and were analyzed with IVIS LivingImage (Caliper Life Sciences) software Regions of inter-est (ROI) were automatically drawn around the isolatedorgans to assess the fluorescence signal emitted For thetotal signal present in mice results were expressed as totalradiant efficiency in units of photonssecond within theregion of interest [ps][120583Wcm2] Signal present in tumorliver spleen or lungs was expressed as percentage of totalsignal
28 Detection of BM-MSCs by Fluorescence To detect CM-DiI+ cells within tumors frozen sections were mounted inmounting media with DAPI (Vector Laboratories Inc) andobserved under a fluorescence microscope at 200x
29 Statistical Analyses Unpaired 2-sided Studentrsquos 119905-test andone-way analysis of variance followed by posttests orKruskal-Wallis and Dunnrsquos posttests (GraphPad Prism Software) wereused for statistical analyses 119875 values lower than 005 wereconsidered as statistically significant
4 BioMed Research International
HUCPVCs
BM-MSCs
CD73 CD90 CD105 CD44 CD14 CD34 CD799723 9290 9692 9772 066 074 090
9759 9833 9410 9621 095 073 096
FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H
FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H
(a)
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
BM-MSCsHUCPVCs
DMEM HuH7 HC-PT-5 LX-2 WI-38 HMEC-1
700
600
500
400
300
200
100
0
Num
ber o
f cel
lsfie
ld
(b)
lowastlowastlowast
BM-MSCsHUCPVCs
BM-MSCs HUCPVCs
Num
ber o
f cel
lsfie
ld
25
20
15
10
5
0
(c)
Figure 1 Characterization of BM-MSCs andHUCPVCs (a) Flow cytometry analysis of cell surfacemarkers of both types ofMSCs Grey areaindicates background fluorescence with IgG isotype control One representative experiment is shown (b) In vitro migration of BM-MSCs(black bars) or HUCPVCs (grey bars) towards CCM from HCC (HuH7 and HC-PT-5) hepatic stellate cells (LX-2) fibroblasts (WI-38)or endothelial cells (HMEC-1) Bars represent the average of MSCsfield (10x) plusmn SEM from three representative visual fields Results arerepresentative of 3 independent experiments 119875 lt 0001 versus DMEM lowastlowastlowast119875 lt 0001 versus BM-MSCs (c) Adhesion towards endothelialcells of BM-MSCs (black bars) or HUCPVCs (grey bars) Results are representative of 3 independent experiments lowastlowastlowast119875 lt 0001 versusBM-MSCs
3 Results
31 Characterization of BM-MSCs and HUCPVCs In accor-dance with the criteria for definingMSCs of the InternationalSociety for Cellular Therapy (ISCT) [26] surface markerexpression of BM-MSCs and HUCPVCs was evaluated byflow cytometry Both types of MSCs showed similar pheno-typic characteristics and were found to express CD73 CD90CD105 and CD44 but not to express the hematopoieticmarkersCD14CD34 orCD79 (Figure 1(a))Wenext decidedto evaluate the in vitro migration capacity of MSCs towardscell culture conditioned media (CCM) obtained from HCCcell lines (HuH7 and HC-PT-5) hepatic stellate cells (LX-2) fibroblasts (WI-38) and endothelial cells (HMEC-1)Interestingly a higher migratory capacity towards all theCCM was found for HUCPVCs when compared to BM-MSCs (Figure 1(b)) Moreover in contrast to our previousresults observed with BM-MCSs [25] HUCPVCs showedcapability to migrate to CCM derived from nontumoral
components (fibroblast and endothelial cells) Besides theircapacity to migrate toward factors secreted by HCC thearrest of MSCs within the microvasculature is considered acritical step for an efficient homing and anchorage to tumorsTherefore we next decided to evaluate adhesion ability ofMSCs and observed that HUCPVCs showed an increased invitro adhesion to HMEC-1 endothelial cells in comparisonwith BM-MSCs (Figure 1(c))
32 In Vivo Migration of BM-MSCs and HUCPVCs towardsHCC To further characterize MSC behavior in vivo non-invasive migration assay was performed CM-DiI and DiRprelabelled BM-MSCs or HUCPVCs were iv injected inHCC tumor-bearing mice in order to evaluate MSC recruit-ment Similar to our previous observation with BM-MSCs[25] at 3 days after cell transplantation a positive signal corre-sponding to HUCPVCs was found in liver lungs spleen andsc tumors (Figure 2(a)) Despite the fact that total signal was
BioMed Research International 5
50
40
30
20
10
times108
Radi
ant e
ffici
ency
(ps
cm
2s
r)(120583
Wc
m2)
(a)
lowastlowastlowast
BM-MSCsHUCPVCs
BM-MSCs HUCPVCsTota
l rad
iant
effici
ency
(ps
) (120583
Wc
m2) times1013
24
23
22
21
20
19
18
17
16
15
14
(b)
BM-MSCs HUCPVCs50
40
30
20
10
times108Ra
dian
t effi
cien
cy(p
sc
m2s
r)(120583
Wc
m2)
(c)
BM-MSCsHUCPVCs
Tota
l sig
nal (
) 90
8086420
Liver Lung Spleen Tumor
lowast
lowast
(d)
DAPI CM-DiI
(e)
BM-MSCsHUCPVCs
250
200
150
100
50
0HuH7 HC-PT-5
Num
ber o
f cel
lsfie
ld
lowastlowastlowast
lowastlowastlowast
(f)
Figure 2 In vivomigration of BM-MSCs and HUCPVCs CM-DiI and DiR prelabeled MSCs were iv injected in sc HuH7 tumor-bearingmice At day 3 mice were sacrificed and organs were removed lungs livers spleen (a) and tumors (c) were exposed to obtain FI Imagesrepresent the radiant efficiency Representative images are shown (b) Total FI for injected BM-MSCs or HUCPVCs was calculated bymeasuring the region of interest (ROI) for all the tissues isolated and results were expressed as total radiant efficiency [ps][120583Wcm2]lowastlowastlowast119875 lt 0001 (d) Signal present in the isolated liver spleen lungs and tumors was represented as percentage of total signal for BM-MSCs or
HUCPVCs-injected mice lowast119875 lt 005 versus BM-MSCs (e) Microscopic analysis of transplanted CM-DiI-labeled MSCs (red signal indicatedby arrows) andDAPI staining in frozen sections of tumors times200magnification (f) In vitromigration ofMSCs to TCMderived fromHuH7 orHC-PT-5 sc tumors Bars represent the average of MSCsfield (10x) plusmn SEM from three representative visual fields Results are representativeof 3 independent experiments lowastlowastlowast119875 lt 0001 versus BM-MSCs
6 BioMed Research International
BM-MSCsHUCPVCs
lowastlowastlowast
lowastlowastlowast
lowast20
15
10
05
00
Fold
chan
ge (2
ΔΔ
ct)
CXCR1 CXCR2 CCR2 IL-6R IL-6ST
(a)
BM-MSCsHUCPVCs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
300
2705
4
3
2
1
0
Fold
chan
ge (2
ΔΔ
ct)
AMFR MMP3 CAV-1 CAV-2 IGFBP3
(b)
Figure 3 Differential expression of cytokineschemokines receptors and AMFAMFR pathway in MSCs Expression of cytokines andchemokines receptors (a) and AMFAMFR axis proteins (b) was evaluated in BM-MSCs (black bars) or HUCPVCs (grey bars) by qPCRlowastlowastlowast119875 lt 0001 versus BM-MSCs
lower in mice injected with HUCPVCs compared to thoseinjected with BM-MSCs (Figure 2(b)) the percentage of totalsignal corresponding to sc tumor locations was increasedin mice administered with HUCPVCs in comparison withanimals that received BM-MSCs (Figures 2(c) and 2(d))indicating an enhanced engraftment of HUCPVCs into HCCtumors In the other evaluated tissues signal intensity wassimilar for BM-MSC or HUCPVCs in lung and liver andit was comparatively reduced in the spleen of HUCPVCs-injected mice (Figure 2(d)) Presence of MSCs in the sctumors was also confirmed by fluorescence microscopy(Figure 2(e)) Finally we evaluated whether MSCs mightpresent differential migratory capacity towards CM obtainedfrom sc tumors (TCM) Interestingly a greater in vitromigratory capacity towards TCM from HCC was observedfor HUCPVCs when compared to BM-MSCs (Figure 2(f))
33 AMFR Was Highly Expressed in HUCPVCs In orderto evaluate mechanisms partially explaining the differentialmigratory capacity of HUCPVCs compared to BM-MSCstowards tumor released factors we analyzed the expressionof some chemokinersquos receptor likely involved inMSC recruit-ment towards HCC Taking into account several reportsdemonstrating that interleukin- (IL-) 8 GRO chemokine(C-C motif) ligand (CCL)-2 and IL-6 are among the mostrelevant factors in HCC [17] we decided to evaluate by qPCRthe expression of CXCR1 CXCR2 CCR2 IL-6R and IL-6ST Interestingly constitutive CXCR1 and CXCR2 mRNAexpression was found to be lower and CCR2 slightly higherin HUCPVCs when compared to BM-MSCs while IL-6Rand IL-6ST expression was similar in both MSCs sources(Figure 3(a)) Next we decided to evaluate the axis of theautocrinemotility factor (AMF) a cytokine with chemotacticeffect on MSCs as recently observed by our group [27]By qPCR a significantly higher expression of the AMFreceptor (AMFR) was found in HUCPVCs when comparedto BM-MSCs Similarly genes known to be related to the
availability of the receptor in the cell surface such as caveolin-1 (CAV-1) and caveolin-2 (CAV-2) were also highly expressedin HUCPVCs as well as the metalloproteinase 3 (MMP3)necessary to the transmigration process In contrast expres-sion levels of insulin-like growth factor-binding protein 3(IGFBP3) a protein that negatively regulates AMFAMFRpathway were found to be reduced in HUCPVCs whencompared to BM-MSCs (Figure 3(b))
34 HUCPVCs Displayed Enhanced Migration towards AMFIn Vitro We have previously demonstrated that AMF acytokine produced by HCC cells plays a critical role inMSC migration [27] Due to the increased AMFR and AMF-AMFR-related proteins expression in HUCPVCs we decidedto test the in vitro migration response to the recombinantAMF (rAMF) of both types ofMSCsusing a chemotaxis assayA significantly higher migration to different doses of rAMF(05 and 075 120583gmL) was observed for HUCPVCs whencompared to BM-MSCs (Figure 4(a)) In spite of differenttypes of MSCs showing similar reduction in migration levels(50 of control) towards HuH7 TCM after the blockage withanti-AMF antibody (data not shown) preincubation of HC-PT-5 TCM with anti-AMF antibody (AMF-ab) resulted ina further reduction in HUCPVCs migration capacity (54of control) when compared to BM-MSCs (67 of control)(Figure 4(b))
4 Discussion
The aim of this study was to assess the capability of differentsources of MSCs to migrate towards HCC released factorsin order to select those showing higher tumor recruitmentcapacity for future therapeutic applications Our resultsindicated that although both types of MSCs share simi-lar phenotypic characteristics as MSCs HUCPVCs have ahigher potential to migrate in vitro towards the conditionedmedium derived not only from HCC cells but also from
BioMed Research International 7
BM-MSCsHUCPVCs
lowastlowastlowast
0 050 075
rAMF (120583gmL)
300
250
200
150
100
50
0
Con
trol (
)
(a)
BM-MSCsHUCPVCs
lowast
Con
trol (
)
110
90
100
70
50
30
10
80
60
40
20
0+ minus + minus
+minus +minus
IgG-abAMF-ab
(b)
Figure 4 HUCPVCs showed enhanced migration towards AMF in comparison with BM-MSCs (a) In vitromigration of BM-MSCs (blackbars) or HUCPVCs (grey bars) towards rAMF 119875 lt 005 versus DMEM (0 120583gmL rAMF) lowast119875 lt 005 versus BM-MSCs (b) In vitromigrationof BM-MSCs (black bars) or HUCPVCs (grey bars) towards HC-PT-5 TCM preincubated with anti-AMF antibody (AMF-ab) or controlisotype (IgG-ab) was evaluated 119875 lt 005 versus IgG-ab lowast119875 lt 005 versus BM-MSCs Bars represent the average of MSCsfield (10x) plusmn SEMfrom three representative visual fields Results are representative of 3 independent experiments
nontumoral components of the tumor microenvironmentlike activated hepatic stellate cells (LX-2) fibroblasts (WI-38) or tumor microendothelial cells (HMEC-1) MoreoverHUCPVCs also showed an enhanced adhesion capacityto HMEC-1 cells compared to BM-MSCs This enhancedadhesion to endothelial cells could allow a more efficientarrest in tumor microvasculature which is a required step forMSC recruitment into HCC In line with this we observedthat HUCPVCs also exhibited greater migration capacity invivo towards experimental HCC In contrast to that observedin the in vitro migration experiments HUCPVCs showed asimilar migration capability towards nontumoral tissues likeliver or lung and also showed a lower recruitment to thespleen AlthoughHUCPVCsmigrated better than BM-MSCstoward activated hepatic stellate cells in vitro no differenceswere observed in in vivo migration to normal liver wherehepatic stellate cells are quiescent In addition it should benoted that mice injected with HUCPVCs showed a lowertotal signal than those injected with BM-MSCs probablydue to differences in the DiR uptake for both kinds of cellsor in the number of inoculated cells These results are inagreement with previous works showing that MCSs derivedfrom umbilical cord are able tomigrate towards experimentaltumors such as glioma [28] breast cancer [29] and lung ade-nocarcinoma [30] Nonetheless to the best of our knowledgethis is the first report describing the comparative migrationcapacity of human umbilical cord perivascular cells towardshuman HCC
Tumor-homing is thought to be mediated by interactionsof cytokineschemokines with their specific receptors Anumber of cytokines growth factors and chemokines aresecreted by HCC cells and their microenvironment [17] Wehave recently found that AMF is critical for MSC migrationtoward HCC in vitro and in vivo Furthermore other factorsreleased by tumor stroma components like IL-8 GRO IL-6or CCL-2 could also be involved inMSC recruitment toHCCThus differences observed between HUCPVCs and BM-MSCs in their in vitro and in vivomigration capacity towardsHCC seem at least partially related to differences in theexpression of cytokinechemokine receptors Some studieshave screened for similarities or differences among BM-MSCs and UC-MSCs at the molecular level [31 32] In addi-tion a previous report demonstrated that migration of UCB-MSC towards glioma cells was higher when compared toBM-MSCs likely due to increased levels of the IL-8 receptorCXCR1 and CXCR2 in the former cell type [18] In contrastto these observations we found that HUCPVCs presentedlower levels of CXCR1 and CXCR2 expression compared toBM-MSCs and these differences could be explained partiallydue to differences in tissue source and culture conditions andmerit further investigationThe analysis of IL-6R and IL6-STexpression showed similar levels in both types ofMSCs whilelevels of MCP-1 receptor (CCR2) were slightly increased inHUCPVCs Interestingly we found that not only AMFR butalso CAV-1 and CAV-2 which are genes that regulate theavailability of AMFR on cell surface [33] were overexpressedin HUCPVCs compared to BM-MSCs In line with this a
8 BioMed Research International
reduced expression level of IGFBP3 a protein which canbind to AMF blocking its binding to AMFR [34] was foundin HUCPVCs when compared to BM-MSCs Moreover anincreased expression of MMP3 was observed in HUCPVCsThis matrix metalloproteinase is involved in AMF-inducedmigration and it is necessary for cell invasion and proteolysisof the extracellularmatrix [35] Finally and consistentwith anincrease in the expression of genes related to the AMFAMFRpathway not only HUCPVCs show an enhanced in vitromigration towards rAMF but also the specific blockageof AMF in HC-PT-5 TCM was able to reduce HUCPVCsmigration to a greater level when compared to BM-MSCs
The potential use of MSCs in cancer treatment as carriersof therapeutic genes has raised some concerns about thesafety of their use in the clinic There are some studiesthat indicated that MSCs have protumorigenic capacitydue to their immunosuppression properties modulation ofthe epithelial-to-mesenchymal transition and induction ofangiogenesis [36] However others have demonstrated thatMSCs decreased HCC growth [37 38] Similarly we havepreviously demonstrated that BM-MSCs did not modifytumor growth in vivo [25] Moreover we observed thatHUCPVCs did not induce tumor growth when injected ivin Huh7 sc tumor-bearing mice (data not shown) Theuse of HUCPVCs as cellular carriers could also have theadvantage of its allogeneic application because of its lowimmunogenicity [9] In this regard HUCPVCs could beuseful as carriers of therapeutic genes for cancer patientswhere isolation of BM or adipose tissue-derived MSCs couldbe less responsive to chemotactic factors and might dis-play higher immunosuppression capacities than MSCs fromhealthy donors [39] or even present genetic abnormalities
5 Conclusions
In summary our results demonstrate a greater migrationcapacity to HCC of HUCPVCs when compared to BM-MSCs not only in vitro but also in vivo likely due to anincrease in migratory response to AMF and to an enhancedadhesive capacity to tumor microvasculature Consideringtheir availability and that no invasive procedures are requiredto obtain HUCPVCs these cells have advantages over BM-MSCs as candidates for carriers of therapeutic genes for thetreatment of HCC
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors gratefully thank Guillermo Gaston MarcosCabrera and Vanina Ferreira for technical assistance andDr Mengshiao Meng (National Chung Hsing University(NCHU) Taichung Taiwan) for providing recombinantAMF This work was supported by Grants from Austral
University (for Juan Bayo I04-12 for Mariana G Garcia T13-12 and for Guillermo Mazzolini T13-11) and from AgenciaNacional de Promocion Cientıfica y Tecnologica (ANPCyT)PICTO 200800115 (Mariana G Garcia) PICT 200800123(Jorge B Aquino) PICTO 200800122 (Jorge B Aquino)and PICT 20102818 (Mariana G Garcia and GuillermoMazzolini)
References
[1] H B El-Serag J A Marrero L Rudolph and K R ReddyldquoDiagnosis and treatment of hepatocellular carcinomardquo Gas-troenterology vol 134 no 6 pp 1752ndash1763 2008
[2] J M Llovet S Ricci V Mazzaferro et al ldquoSorafenib inadvanced hepatocellular carcinomardquo The New England Journalof Medicine vol 359 no 4 pp 378ndash390 2008
[3] A J Friedenstein J F Gorskaja andN N Kulagina ldquoFibroblastprecursors in normal and irradiated mouse hematopoieticorgansrdquo Experimental Hematology vol 4 no 5 pp 267ndash2741976
[4] D J Prockop and J Y Oh ldquoMedical therapies with adultstemprogenitor cells (MSCs) a backward journey from dra-matic results in vivo to the cellular andmolecular explanationsrdquoJournal of Cellular Biochemistry vol 113 no 5 pp 1460ndash14692012
[5] J B Aquino M F Bolontrade M G Garcıa O L Podhajcerand GMazzolini ldquoMesenchymal stem cells as therapeutic toolsand gene carriers in liver fibrosis and hepatocellular carcinomardquoGene Therapy vol 17 no 6 pp 692ndash708 2010
[6] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[7] H Yoshimura T Muneta A Nimura A Yokoyama H Kogaand I Sekiya ldquoComparison of rat mesenchymal stem cellsderived from bone marrow synovium periosteum adiposetissue and musclerdquo Cell and Tissue Research vol 327 no 3 pp449ndash462 2007
[8] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popu-lations and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[9] R Sarugaser D Lickorish D Baksh M M Hosseini and J EDavies ldquoHuman umbilical cord perivascular (HUCPV) cells asource of mesenchymal progenitorsrdquo Stem Cells vol 23 no 2pp 220ndash229 2005
[10] D Baksh R Yao and R S Tuan ldquoComparison of proliferativeand multilineage differentiation potential of human mesenchy-mal stem cells derived from umbilical cord and bone marrowrdquoStem Cells vol 25 no 6 pp 1384ndash1392 2007
[11] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[12] O K Lee T K Kuo W M Chen K D Lee S L Hsieh andT H Chen ldquoIsolation of multipotent mesenchymal stem cellsfrom umbilical cord bloodrdquo Blood vol 103 no 5 pp 1669ndash16752004
[13] HWang S Hung S Peng et al ldquoMesenchymal stem cells in theWhartonrsquos jelly of the human umbilical cordrdquo StemCells vol 22no 7 pp 1330ndash1337 2004
BioMed Research International 9
[14] J Kim J M Shin Y J Jeon H M Chung and J Chae ldquoPro-teomic validation of multifunctional molecules in mesenchy-mal stem cells derived from human bone marrow umbilicalcord blood and peripheral bloodrdquo PLoS ONE vol 7 no 5Article ID e32350 2012
[15] J-Y Hsieh H-W Wang S-J Chang et al ldquoMesenchymalstem cells from human umbilical cord express preferentiallysecreted factors related to neuroprotection neurogenesis andangiogenesisrdquo PLoS ONE vol 8 no 8 Article ID e72604 2013
[16] V Hernandez-Gea S Toffanin S L Friedman and J MLlovet ldquoRole of the microenvironment in the pathogenesis andtreatment of hepatocellular carcinomardquo Gastroenterology vol144 no 3 pp 512ndash527 2013
[17] J Bayo M Marrodan J B Aquino M Silva M G Garciaand G Mazzolini ldquoThe therapeutic potential of bone marrow-derived mesenchymal stromal cells on hepatocellular carci-nomardquo Liver International vol 34 no 3 pp 330ndash334 2014
[18] D Kim J H Kim J Kwon Lee et al ldquoOverexpression ofCXC chemokine receptors is required for the superior glioma-tracking property of umbilical cord blood-derived mesenchy-mal stem cellsrdquo Stem Cells and Development vol 18 no 3 pp511ndash519 2009
[19] S M Kim D Kim C H Jeong et al ldquoCXC chemokinereceptor 1 enhances the ability of human umbilical cord blood-derived mesenchymal stem cells to migrate toward gliomasrdquoBiochemical and Biophysical Research Communications vol 407no 4 pp 741ndash746 2011
[20] S A Park C H Ryu S M Kim et al ldquoCXCR4-transfectedhuman umbilical cord blood-derived mesenchymal stem cellsexhibit enhanced migratory capacity toward gliomasrdquo Interna-tional Journal of Oncology vol 38 no 1 pp 97ndash103 2011
[21] R S Rachakatla F Marini M L Weiss M Tamura and DTroyer ldquoDevelopment of human umbilical cord matrix stemcell-based gene therapy for experimental lung tumorsrdquo CancerGene Therapy vol 14 no 10 pp 828ndash835 2007
[22] C Yan S Li Z Li et al ldquoHuman umbilical cord mesenchymalstem cells as vehicles of CD20-specific TRAIL fusion proteindelivery a double-target therapy against non-Hodgkinrsquos lym-phomardquo Molecular Pharmaceutics vol 10 no 1 pp 142ndash1512013
[23] J Hu A Dong V Fernandez-Ruiz et al ldquoBlockade of Wntsignaling inhibits angiogenesis and tumor growth in hepatocel-lular carcinomardquo Cancer Research vol 69 no 17 pp 6951ndash69592009
[24] L Xu A Y Hui E Albanis et al ldquoHuman hepatic stellate celllines LX-1 and LX-2 new tools for analysis of hepatic fibrosisrdquoGut vol 54 no 1 pp 142ndash151 2005
[25] M G Garcia J Bayo M F Bolontrade et al ldquoHepatocellularcarcinoma cells and their fibrotic microenvironment modulatebone marrow-derived mesenchymal stromal cell migration invitro and in vivordquo Molecular Pharmaceutics vol 8 no 5 pp1538ndash1548 2011
[26] M Dominici K le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells theInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[27] J Bayo E Fiore J B Aquino et al ldquoIncreased migration ofhumanmesenchymal stromal cells byAutocrineMotility Factor(AMF) resulted in enhanced recruitment towards hepatocellu-lar carcinomardquo PLoS ONE vol 9 no 4 Article ID e95171 2014
[28] M K Seong Y L Jung I P Sang et al ldquoGene therapyusing TRAIL-secreting human umbilical cord blood-derived
mesenchymal stem cells against intracranial gliomardquo CancerResearch vol 68 no 23 pp 9614ndash9623 2008
[29] R Ayuzawa C Doi R S Rachakatla et al ldquoNaıve humanumbilical cord matrix derived stem cells significantly attenuategrowth of human breast cancer cells in vitro and in vivordquoCancerLetters vol 280 no 1 pp 31ndash37 2009
[30] T Matsuzuka R S Rachakatla C Doi et al ldquoHuman umbilicalcord matrix-derived stem cells expressing interferon-120573 genesignificantly attenuate bronchioloalveolar carcinomaxenograftsin SCID micerdquo Lung Cancer vol 70 no 1 pp 28ndash36 2010
[31] G Li X Zhang H Wang et al ldquoComparative proteomicanalysis of mesenchymal stem cells derived from human bonemarrow umbilical cord and placenta implication in themigra-tionrdquo Proteomics vol 9 no 1 pp 20ndash30 2009
[32] H C Miranda R H Herai C H Thome et al ldquoA quan-titative proteomic and transcriptomic comparison of humanmesenchymal stem cells from bone marrow and umbilical cordveinrdquo Proteomics vol 12 no 17 pp 2607ndash2617 2012
[33] P U Le G Guay Y Altschuler and I R Nabi ldquoCaveolin-1 isa negative regulator of caveolae-mediated endocytosis to theendoplasmic reticulumrdquo The Journal of Biological Chemistryvol 277 no 5 pp 3371ndash3379 2002
[34] S Mishra A Raz and L J Murphy ldquoInsulin-like growthfactor binding protein-3 interacts with autocrine motility fac-torphosphoglucose isomerase (AMFPGI) and inhibits theAMFPGI functionrdquo Cancer Research vol 64 no 7 pp 2516ndash2522 2004
[35] F L Yu M H Liao J W Lee and W L Shih ldquoInductionof hepatoma cells migration by phosphoglucose isomeraseautocrine motility factor through the upregulation of matrixmetalloproteinase-3rdquo Biochemical and Biophysical ResearchCommunications vol 314 no 1 pp 76ndash82 2004
[36] A H Klopp A Gupta E Spaeth M Andreeff and F MariniIII ldquoConcise review dissecting a discrepancy in the literaturedomesenchymal stem cells support or suppress tumor growthrdquoStem Cells vol 29 no 1 pp 11ndash19 2011
[37] L Qiao Z Xu T Zhao et al ldquoSuppression of tumorigenesisby human mesenchymal stem cells in a hepatoma modelrdquo CellResearch vol 18 no 4 pp 500ndash507 2008
[38] Y R Lu Y Yuan X J Wang et al ldquoThe growth inhibitory effectof mesenchymal stem cells on tumor cells in vitro and in vivordquoCancer Biology andTherapy vol 7 no 2 pp 245ndash251 2008
[39] Z-G Zhao W Xu L Sun W-M Li Q-P Li and P Zou ldquoThecharacteristics and immunoregulatory functions of regulatorydendritic cells induced by mesenchymal stem cells derivedfrom bone marrow of patient with chronic myeloid leukaemiardquoEuropean Journal of Cancer vol 48 no 12 pp 1884ndash1895 2012

BioMed Research International 3
24 In Vitro Migration Assays In vitro migration was per-formed using a 48-Transwell microchemotaxis BoydenChamber unit (Neuroprobe Inc) as previously described[25] MSCs (12 times 103 cellswell) were placed in the upperchamber and DMEM CCM TCM or rAMF were appliedto the lower chamber of the transwell unit For blockingexperiments TCM were preincubated for 60min with anti-AMF antibody or isotype control IgG All systems were left at37∘C in a 5 CO
2humidified atmosphere for 4 hours except
for experiments involving rAMF that were maintained for18 hours Cells attached to the lower side of the membranewere fixed in 2 formaldehyde stained with 410158406-diamidino-2-phenylindole dihydrochloride (DAPI Sigma-Aldrich) andcounted using fluorescent-fieldmicroscopy at 100x Capturedimages from three representative visual fields were analyzedusing Cell Profiler software (httpwwwcellprofilercom)and the mean number of cellsfield plusmn SEM was calculated
25 Cell Adhesion Assays For analyses of MSC adhesionto endothelial cells 2 times 105 HMEC-1 were seeded in 96-well microplates and cultured for 1 day prior to the assayCoated wells were incubated for 5 minutes with 01mLof 5 times 104 cellsmL of Fast-DiO prelabelled MSCs Cellsuspension was discarded and attached cells were fixed with2 paraformaldehyde Cells were counted using fluorescent-field microscopy at 200x pictures taken from ten represen-tative visual fields were analyzed using Cell Profiler soft-ware (httpwwwcellprofilercom) and the mean number ofcellsfield plusmn SEM was calculated
26 Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Total RNA of MSCs was extracted using TrizolReagent (Sigma-Aldrich Co St Louis MO) Total RNA(4 120583g) was reverse-transcribed with 200U of SuperScriptII Reverse Transcriptase (Invitrogen Carlsbad CA) using500 ng of Oligo (dT) primers cDNAs were subjected toreal-time polymerase chain reaction (qPCR) (StratageneMx3005p Stratagene La Jolla CA USA) For qRT-PCRthe mRNA levels of CXCR1 CXCR2 CCR2 IL-6 receptor(IL-6R) IL-6 signal transducer (IL-6ST) AMF receptor(AMFR) metalloproteinase 3 (MMP3) insulin-like growthfactor-binding protein 3 (IGFBP3) caveolin-1 (CAV-1) andcaveolin-2 (CAV-2) were quantified by SYBR Green (Invitro-gen) using the following primers CXCR1 forward 51015840-TTT-TCCGCCAGGCTTACCAT-31015840 and reverse 51015840-AACACC-ATCCGCCATTTTGC-31015840 CXCR2 forward 51015840-TAAGTG-GAGCCCCGTGGGG-31015840 and reverse 51015840-TGGGCTCAG-GGGCAGGATG-31015840 CCR2 forward 51015840-CGAGAGCGG-TGAAGAAGTCA-31015840 and reverse 51015840-AGCATGTTGCCC-ACAAAACC-31015840 IL-6R forward 51015840-GCACTTGCTGGT-GGATGTTC-31015840 and reverse 51015840-AGCCTTTGTCGTCAG-GGATG-31015840 IL-6ST forward 51015840-CCCACCTCATGCACT-GTTGA-31015840 and reverse 51015840-TTATGTGGCGGATTCGGCTT-31015840 AMFR forward 51015840-ACAAGATGTGGGCCTTGCAAG-A-31015840 and reverse 51015840-AAAACGCAGTGCTCCCAGGATA-31015840 MMP3 forward 51015840-ACGCCAGCCAACTGTGATCCT-31015840 and reverse 51015840-ATATGCGGCATCCACGCCTGAA-31015840IGFBP3 forward 51015840-ACTGTGGCCATGACTGAG-31015840 and
reverse 51015840-AGAGTCTCCCTGAGCCTGA-31015840 CAV-1 for-ward 51015840-AATCCAAGCATCCCTTTGCCCA-31015840 and reverse51015840-ACCAGGCAGCTTTCTGTACGA-31015840 CAV-2 forward51015840-GAGAGACAGGGGAGTTGTCAACTT-31015840 and reverse51015840-GCCCGGCCCAGAAATAATGAGAT-31015840 PCR ampli-fications were carried out using a cycle of 95∘C for 10 minutesand 45 cycles under the following parameters 95∘C for 30seconds 58∘C for 60 seconds 72∘C for 30 seconds At the endof PCR reaction the temperature was increased from 60∘C to95∘C at a rate of 2∘Cmin and the fluorescence was measuredevery 15 seconds to construct the melting curve Values werenormalized to levels of glyceraldehyde-3-phosphate dehy-drogenase (GAPDH used as housekeeping) transcript (for-ward 51015840-CATCTCTGCCCCCTCTGCTG-31015840 and reverse 51015840-GCCTGCTTCACCACCTTCTTG-31015840) Data were processedby theΔΔCtmethodThe relative amount of the PCR productamplified fromBM-MSCswas set as 1 A nontemplate control(NTC) was run in every assay and all determinations wereperformed as triplicates in three separated experiments
27 Mice and In Vivo Experiments Six- to eight-week-old male nude mice (NuNu) were purchased from CNEA(Comision Nacional de Energıa Atomica Ezeiza BuenosAires Argentina) Subcutaneous HuH7 tumors (2 times 106cells)were established and 10 days later BM-MSCs or HUCPVCswere intravenously (iv) injected Tumor growthwas assessedby calliper measurement and tumor volume (mm3) wascalculated by the formula 1205876 times larger diameter times (smallerdiameter)2 For in vivo migration studies MSCs (5 times 105)were prelabeled with CM-DiI for histological analysis andDiR (Molecular Probes Invitrogen) for fluorescence imag-ing (FI) FI was performed using the Xenogen In VivoImaging System (IVIS Caliper Life Sciences HopkintonMA USA) 1 hour after MSC injection and every day untilexperimental end point At day 3 mice were sacrificedand isolated tissues were exposed to FI Images representthe radiant efficiency and were analyzed with IVIS LivingImage (Caliper Life Sciences) software Regions of inter-est (ROI) were automatically drawn around the isolatedorgans to assess the fluorescence signal emitted For thetotal signal present in mice results were expressed as totalradiant efficiency in units of photonssecond within theregion of interest [ps][120583Wcm2] Signal present in tumorliver spleen or lungs was expressed as percentage of totalsignal
28 Detection of BM-MSCs by Fluorescence To detect CM-DiI+ cells within tumors frozen sections were mounted inmounting media with DAPI (Vector Laboratories Inc) andobserved under a fluorescence microscope at 200x
29 Statistical Analyses Unpaired 2-sided Studentrsquos 119905-test andone-way analysis of variance followed by posttests orKruskal-Wallis and Dunnrsquos posttests (GraphPad Prism Software) wereused for statistical analyses 119875 values lower than 005 wereconsidered as statistically significant
4 BioMed Research International
HUCPVCs
BM-MSCs
CD73 CD90 CD105 CD44 CD14 CD34 CD799723 9290 9692 9772 066 074 090
9759 9833 9410 9621 095 073 096
FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H
FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H
(a)
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
BM-MSCsHUCPVCs
DMEM HuH7 HC-PT-5 LX-2 WI-38 HMEC-1
700
600
500
400
300
200
100
0
Num
ber o
f cel
lsfie
ld
(b)
lowastlowastlowast
BM-MSCsHUCPVCs
BM-MSCs HUCPVCs
Num
ber o
f cel
lsfie
ld
25
20
15
10
5
0
(c)
Figure 1 Characterization of BM-MSCs andHUCPVCs (a) Flow cytometry analysis of cell surfacemarkers of both types ofMSCs Grey areaindicates background fluorescence with IgG isotype control One representative experiment is shown (b) In vitro migration of BM-MSCs(black bars) or HUCPVCs (grey bars) towards CCM from HCC (HuH7 and HC-PT-5) hepatic stellate cells (LX-2) fibroblasts (WI-38)or endothelial cells (HMEC-1) Bars represent the average of MSCsfield (10x) plusmn SEM from three representative visual fields Results arerepresentative of 3 independent experiments 119875 lt 0001 versus DMEM lowastlowastlowast119875 lt 0001 versus BM-MSCs (c) Adhesion towards endothelialcells of BM-MSCs (black bars) or HUCPVCs (grey bars) Results are representative of 3 independent experiments lowastlowastlowast119875 lt 0001 versusBM-MSCs
3 Results
31 Characterization of BM-MSCs and HUCPVCs In accor-dance with the criteria for definingMSCs of the InternationalSociety for Cellular Therapy (ISCT) [26] surface markerexpression of BM-MSCs and HUCPVCs was evaluated byflow cytometry Both types of MSCs showed similar pheno-typic characteristics and were found to express CD73 CD90CD105 and CD44 but not to express the hematopoieticmarkersCD14CD34 orCD79 (Figure 1(a))Wenext decidedto evaluate the in vitro migration capacity of MSCs towardscell culture conditioned media (CCM) obtained from HCCcell lines (HuH7 and HC-PT-5) hepatic stellate cells (LX-2) fibroblasts (WI-38) and endothelial cells (HMEC-1)Interestingly a higher migratory capacity towards all theCCM was found for HUCPVCs when compared to BM-MSCs (Figure 1(b)) Moreover in contrast to our previousresults observed with BM-MCSs [25] HUCPVCs showedcapability to migrate to CCM derived from nontumoral
components (fibroblast and endothelial cells) Besides theircapacity to migrate toward factors secreted by HCC thearrest of MSCs within the microvasculature is considered acritical step for an efficient homing and anchorage to tumorsTherefore we next decided to evaluate adhesion ability ofMSCs and observed that HUCPVCs showed an increased invitro adhesion to HMEC-1 endothelial cells in comparisonwith BM-MSCs (Figure 1(c))
32 In Vivo Migration of BM-MSCs and HUCPVCs towardsHCC To further characterize MSC behavior in vivo non-invasive migration assay was performed CM-DiI and DiRprelabelled BM-MSCs or HUCPVCs were iv injected inHCC tumor-bearing mice in order to evaluate MSC recruit-ment Similar to our previous observation with BM-MSCs[25] at 3 days after cell transplantation a positive signal corre-sponding to HUCPVCs was found in liver lungs spleen andsc tumors (Figure 2(a)) Despite the fact that total signal was
BioMed Research International 5
50
40
30
20
10
times108
Radi
ant e
ffici
ency
(ps
cm
2s
r)(120583
Wc
m2)
(a)
lowastlowastlowast
BM-MSCsHUCPVCs
BM-MSCs HUCPVCsTota
l rad
iant
effici
ency
(ps
) (120583
Wc
m2) times1013
24
23
22
21
20
19
18
17
16
15
14
(b)
BM-MSCs HUCPVCs50
40
30
20
10
times108Ra
dian
t effi
cien
cy(p
sc
m2s
r)(120583
Wc
m2)
(c)
BM-MSCsHUCPVCs
Tota
l sig
nal (
) 90
8086420
Liver Lung Spleen Tumor
lowast
lowast
(d)
DAPI CM-DiI
(e)
BM-MSCsHUCPVCs
250
200
150
100
50
0HuH7 HC-PT-5
Num
ber o
f cel
lsfie
ld
lowastlowastlowast
lowastlowastlowast
(f)
Figure 2 In vivomigration of BM-MSCs and HUCPVCs CM-DiI and DiR prelabeled MSCs were iv injected in sc HuH7 tumor-bearingmice At day 3 mice were sacrificed and organs were removed lungs livers spleen (a) and tumors (c) were exposed to obtain FI Imagesrepresent the radiant efficiency Representative images are shown (b) Total FI for injected BM-MSCs or HUCPVCs was calculated bymeasuring the region of interest (ROI) for all the tissues isolated and results were expressed as total radiant efficiency [ps][120583Wcm2]lowastlowastlowast119875 lt 0001 (d) Signal present in the isolated liver spleen lungs and tumors was represented as percentage of total signal for BM-MSCs or
HUCPVCs-injected mice lowast119875 lt 005 versus BM-MSCs (e) Microscopic analysis of transplanted CM-DiI-labeled MSCs (red signal indicatedby arrows) andDAPI staining in frozen sections of tumors times200magnification (f) In vitromigration ofMSCs to TCMderived fromHuH7 orHC-PT-5 sc tumors Bars represent the average of MSCsfield (10x) plusmn SEM from three representative visual fields Results are representativeof 3 independent experiments lowastlowastlowast119875 lt 0001 versus BM-MSCs
6 BioMed Research International
BM-MSCsHUCPVCs
lowastlowastlowast
lowastlowastlowast
lowast20
15
10
05
00
Fold
chan
ge (2
ΔΔ
ct)
CXCR1 CXCR2 CCR2 IL-6R IL-6ST
(a)
BM-MSCsHUCPVCs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
300
2705
4
3
2
1
0
Fold
chan
ge (2
ΔΔ
ct)
AMFR MMP3 CAV-1 CAV-2 IGFBP3
(b)
Figure 3 Differential expression of cytokineschemokines receptors and AMFAMFR pathway in MSCs Expression of cytokines andchemokines receptors (a) and AMFAMFR axis proteins (b) was evaluated in BM-MSCs (black bars) or HUCPVCs (grey bars) by qPCRlowastlowastlowast119875 lt 0001 versus BM-MSCs
lower in mice injected with HUCPVCs compared to thoseinjected with BM-MSCs (Figure 2(b)) the percentage of totalsignal corresponding to sc tumor locations was increasedin mice administered with HUCPVCs in comparison withanimals that received BM-MSCs (Figures 2(c) and 2(d))indicating an enhanced engraftment of HUCPVCs into HCCtumors In the other evaluated tissues signal intensity wassimilar for BM-MSC or HUCPVCs in lung and liver andit was comparatively reduced in the spleen of HUCPVCs-injected mice (Figure 2(d)) Presence of MSCs in the sctumors was also confirmed by fluorescence microscopy(Figure 2(e)) Finally we evaluated whether MSCs mightpresent differential migratory capacity towards CM obtainedfrom sc tumors (TCM) Interestingly a greater in vitromigratory capacity towards TCM from HCC was observedfor HUCPVCs when compared to BM-MSCs (Figure 2(f))
33 AMFR Was Highly Expressed in HUCPVCs In orderto evaluate mechanisms partially explaining the differentialmigratory capacity of HUCPVCs compared to BM-MSCstowards tumor released factors we analyzed the expressionof some chemokinersquos receptor likely involved inMSC recruit-ment towards HCC Taking into account several reportsdemonstrating that interleukin- (IL-) 8 GRO chemokine(C-C motif) ligand (CCL)-2 and IL-6 are among the mostrelevant factors in HCC [17] we decided to evaluate by qPCRthe expression of CXCR1 CXCR2 CCR2 IL-6R and IL-6ST Interestingly constitutive CXCR1 and CXCR2 mRNAexpression was found to be lower and CCR2 slightly higherin HUCPVCs when compared to BM-MSCs while IL-6Rand IL-6ST expression was similar in both MSCs sources(Figure 3(a)) Next we decided to evaluate the axis of theautocrinemotility factor (AMF) a cytokine with chemotacticeffect on MSCs as recently observed by our group [27]By qPCR a significantly higher expression of the AMFreceptor (AMFR) was found in HUCPVCs when comparedto BM-MSCs Similarly genes known to be related to the
availability of the receptor in the cell surface such as caveolin-1 (CAV-1) and caveolin-2 (CAV-2) were also highly expressedin HUCPVCs as well as the metalloproteinase 3 (MMP3)necessary to the transmigration process In contrast expres-sion levels of insulin-like growth factor-binding protein 3(IGFBP3) a protein that negatively regulates AMFAMFRpathway were found to be reduced in HUCPVCs whencompared to BM-MSCs (Figure 3(b))
34 HUCPVCs Displayed Enhanced Migration towards AMFIn Vitro We have previously demonstrated that AMF acytokine produced by HCC cells plays a critical role inMSC migration [27] Due to the increased AMFR and AMF-AMFR-related proteins expression in HUCPVCs we decidedto test the in vitro migration response to the recombinantAMF (rAMF) of both types ofMSCsusing a chemotaxis assayA significantly higher migration to different doses of rAMF(05 and 075 120583gmL) was observed for HUCPVCs whencompared to BM-MSCs (Figure 4(a)) In spite of differenttypes of MSCs showing similar reduction in migration levels(50 of control) towards HuH7 TCM after the blockage withanti-AMF antibody (data not shown) preincubation of HC-PT-5 TCM with anti-AMF antibody (AMF-ab) resulted ina further reduction in HUCPVCs migration capacity (54of control) when compared to BM-MSCs (67 of control)(Figure 4(b))
4 Discussion
The aim of this study was to assess the capability of differentsources of MSCs to migrate towards HCC released factorsin order to select those showing higher tumor recruitmentcapacity for future therapeutic applications Our resultsindicated that although both types of MSCs share simi-lar phenotypic characteristics as MSCs HUCPVCs have ahigher potential to migrate in vitro towards the conditionedmedium derived not only from HCC cells but also from
BioMed Research International 7
BM-MSCsHUCPVCs
lowastlowastlowast
0 050 075
rAMF (120583gmL)
300
250
200
150
100
50
0
Con
trol (
)
(a)
BM-MSCsHUCPVCs
lowast
Con
trol (
)
110
90
100
70
50
30
10
80
60
40
20
0+ minus + minus
+minus +minus
IgG-abAMF-ab
(b)
Figure 4 HUCPVCs showed enhanced migration towards AMF in comparison with BM-MSCs (a) In vitromigration of BM-MSCs (blackbars) or HUCPVCs (grey bars) towards rAMF 119875 lt 005 versus DMEM (0 120583gmL rAMF) lowast119875 lt 005 versus BM-MSCs (b) In vitromigrationof BM-MSCs (black bars) or HUCPVCs (grey bars) towards HC-PT-5 TCM preincubated with anti-AMF antibody (AMF-ab) or controlisotype (IgG-ab) was evaluated 119875 lt 005 versus IgG-ab lowast119875 lt 005 versus BM-MSCs Bars represent the average of MSCsfield (10x) plusmn SEMfrom three representative visual fields Results are representative of 3 independent experiments
nontumoral components of the tumor microenvironmentlike activated hepatic stellate cells (LX-2) fibroblasts (WI-38) or tumor microendothelial cells (HMEC-1) MoreoverHUCPVCs also showed an enhanced adhesion capacityto HMEC-1 cells compared to BM-MSCs This enhancedadhesion to endothelial cells could allow a more efficientarrest in tumor microvasculature which is a required step forMSC recruitment into HCC In line with this we observedthat HUCPVCs also exhibited greater migration capacity invivo towards experimental HCC In contrast to that observedin the in vitro migration experiments HUCPVCs showed asimilar migration capability towards nontumoral tissues likeliver or lung and also showed a lower recruitment to thespleen AlthoughHUCPVCsmigrated better than BM-MSCstoward activated hepatic stellate cells in vitro no differenceswere observed in in vivo migration to normal liver wherehepatic stellate cells are quiescent In addition it should benoted that mice injected with HUCPVCs showed a lowertotal signal than those injected with BM-MSCs probablydue to differences in the DiR uptake for both kinds of cellsor in the number of inoculated cells These results are inagreement with previous works showing that MCSs derivedfrom umbilical cord are able tomigrate towards experimentaltumors such as glioma [28] breast cancer [29] and lung ade-nocarcinoma [30] Nonetheless to the best of our knowledgethis is the first report describing the comparative migrationcapacity of human umbilical cord perivascular cells towardshuman HCC
Tumor-homing is thought to be mediated by interactionsof cytokineschemokines with their specific receptors Anumber of cytokines growth factors and chemokines aresecreted by HCC cells and their microenvironment [17] Wehave recently found that AMF is critical for MSC migrationtoward HCC in vitro and in vivo Furthermore other factorsreleased by tumor stroma components like IL-8 GRO IL-6or CCL-2 could also be involved inMSC recruitment toHCCThus differences observed between HUCPVCs and BM-MSCs in their in vitro and in vivomigration capacity towardsHCC seem at least partially related to differences in theexpression of cytokinechemokine receptors Some studieshave screened for similarities or differences among BM-MSCs and UC-MSCs at the molecular level [31 32] In addi-tion a previous report demonstrated that migration of UCB-MSC towards glioma cells was higher when compared toBM-MSCs likely due to increased levels of the IL-8 receptorCXCR1 and CXCR2 in the former cell type [18] In contrastto these observations we found that HUCPVCs presentedlower levels of CXCR1 and CXCR2 expression compared toBM-MSCs and these differences could be explained partiallydue to differences in tissue source and culture conditions andmerit further investigationThe analysis of IL-6R and IL6-STexpression showed similar levels in both types ofMSCs whilelevels of MCP-1 receptor (CCR2) were slightly increased inHUCPVCs Interestingly we found that not only AMFR butalso CAV-1 and CAV-2 which are genes that regulate theavailability of AMFR on cell surface [33] were overexpressedin HUCPVCs compared to BM-MSCs In line with this a
8 BioMed Research International
reduced expression level of IGFBP3 a protein which canbind to AMF blocking its binding to AMFR [34] was foundin HUCPVCs when compared to BM-MSCs Moreover anincreased expression of MMP3 was observed in HUCPVCsThis matrix metalloproteinase is involved in AMF-inducedmigration and it is necessary for cell invasion and proteolysisof the extracellularmatrix [35] Finally and consistentwith anincrease in the expression of genes related to the AMFAMFRpathway not only HUCPVCs show an enhanced in vitromigration towards rAMF but also the specific blockageof AMF in HC-PT-5 TCM was able to reduce HUCPVCsmigration to a greater level when compared to BM-MSCs
The potential use of MSCs in cancer treatment as carriersof therapeutic genes has raised some concerns about thesafety of their use in the clinic There are some studiesthat indicated that MSCs have protumorigenic capacitydue to their immunosuppression properties modulation ofthe epithelial-to-mesenchymal transition and induction ofangiogenesis [36] However others have demonstrated thatMSCs decreased HCC growth [37 38] Similarly we havepreviously demonstrated that BM-MSCs did not modifytumor growth in vivo [25] Moreover we observed thatHUCPVCs did not induce tumor growth when injected ivin Huh7 sc tumor-bearing mice (data not shown) Theuse of HUCPVCs as cellular carriers could also have theadvantage of its allogeneic application because of its lowimmunogenicity [9] In this regard HUCPVCs could beuseful as carriers of therapeutic genes for cancer patientswhere isolation of BM or adipose tissue-derived MSCs couldbe less responsive to chemotactic factors and might dis-play higher immunosuppression capacities than MSCs fromhealthy donors [39] or even present genetic abnormalities
5 Conclusions
In summary our results demonstrate a greater migrationcapacity to HCC of HUCPVCs when compared to BM-MSCs not only in vitro but also in vivo likely due to anincrease in migratory response to AMF and to an enhancedadhesive capacity to tumor microvasculature Consideringtheir availability and that no invasive procedures are requiredto obtain HUCPVCs these cells have advantages over BM-MSCs as candidates for carriers of therapeutic genes for thetreatment of HCC
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors gratefully thank Guillermo Gaston MarcosCabrera and Vanina Ferreira for technical assistance andDr Mengshiao Meng (National Chung Hsing University(NCHU) Taichung Taiwan) for providing recombinantAMF This work was supported by Grants from Austral
University (for Juan Bayo I04-12 for Mariana G Garcia T13-12 and for Guillermo Mazzolini T13-11) and from AgenciaNacional de Promocion Cientıfica y Tecnologica (ANPCyT)PICTO 200800115 (Mariana G Garcia) PICT 200800123(Jorge B Aquino) PICTO 200800122 (Jorge B Aquino)and PICT 20102818 (Mariana G Garcia and GuillermoMazzolini)
References
[1] H B El-Serag J A Marrero L Rudolph and K R ReddyldquoDiagnosis and treatment of hepatocellular carcinomardquo Gas-troenterology vol 134 no 6 pp 1752ndash1763 2008
[2] J M Llovet S Ricci V Mazzaferro et al ldquoSorafenib inadvanced hepatocellular carcinomardquo The New England Journalof Medicine vol 359 no 4 pp 378ndash390 2008
[3] A J Friedenstein J F Gorskaja andN N Kulagina ldquoFibroblastprecursors in normal and irradiated mouse hematopoieticorgansrdquo Experimental Hematology vol 4 no 5 pp 267ndash2741976
[4] D J Prockop and J Y Oh ldquoMedical therapies with adultstemprogenitor cells (MSCs) a backward journey from dra-matic results in vivo to the cellular andmolecular explanationsrdquoJournal of Cellular Biochemistry vol 113 no 5 pp 1460ndash14692012
[5] J B Aquino M F Bolontrade M G Garcıa O L Podhajcerand GMazzolini ldquoMesenchymal stem cells as therapeutic toolsand gene carriers in liver fibrosis and hepatocellular carcinomardquoGene Therapy vol 17 no 6 pp 692ndash708 2010
[6] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[7] H Yoshimura T Muneta A Nimura A Yokoyama H Kogaand I Sekiya ldquoComparison of rat mesenchymal stem cellsderived from bone marrow synovium periosteum adiposetissue and musclerdquo Cell and Tissue Research vol 327 no 3 pp449ndash462 2007
[8] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popu-lations and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[9] R Sarugaser D Lickorish D Baksh M M Hosseini and J EDavies ldquoHuman umbilical cord perivascular (HUCPV) cells asource of mesenchymal progenitorsrdquo Stem Cells vol 23 no 2pp 220ndash229 2005
[10] D Baksh R Yao and R S Tuan ldquoComparison of proliferativeand multilineage differentiation potential of human mesenchy-mal stem cells derived from umbilical cord and bone marrowrdquoStem Cells vol 25 no 6 pp 1384ndash1392 2007
[11] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[12] O K Lee T K Kuo W M Chen K D Lee S L Hsieh andT H Chen ldquoIsolation of multipotent mesenchymal stem cellsfrom umbilical cord bloodrdquo Blood vol 103 no 5 pp 1669ndash16752004
[13] HWang S Hung S Peng et al ldquoMesenchymal stem cells in theWhartonrsquos jelly of the human umbilical cordrdquo StemCells vol 22no 7 pp 1330ndash1337 2004
BioMed Research International 9
[14] J Kim J M Shin Y J Jeon H M Chung and J Chae ldquoPro-teomic validation of multifunctional molecules in mesenchy-mal stem cells derived from human bone marrow umbilicalcord blood and peripheral bloodrdquo PLoS ONE vol 7 no 5Article ID e32350 2012
[15] J-Y Hsieh H-W Wang S-J Chang et al ldquoMesenchymalstem cells from human umbilical cord express preferentiallysecreted factors related to neuroprotection neurogenesis andangiogenesisrdquo PLoS ONE vol 8 no 8 Article ID e72604 2013
[16] V Hernandez-Gea S Toffanin S L Friedman and J MLlovet ldquoRole of the microenvironment in the pathogenesis andtreatment of hepatocellular carcinomardquo Gastroenterology vol144 no 3 pp 512ndash527 2013
[17] J Bayo M Marrodan J B Aquino M Silva M G Garciaand G Mazzolini ldquoThe therapeutic potential of bone marrow-derived mesenchymal stromal cells on hepatocellular carci-nomardquo Liver International vol 34 no 3 pp 330ndash334 2014
[18] D Kim J H Kim J Kwon Lee et al ldquoOverexpression ofCXC chemokine receptors is required for the superior glioma-tracking property of umbilical cord blood-derived mesenchy-mal stem cellsrdquo Stem Cells and Development vol 18 no 3 pp511ndash519 2009
[19] S M Kim D Kim C H Jeong et al ldquoCXC chemokinereceptor 1 enhances the ability of human umbilical cord blood-derived mesenchymal stem cells to migrate toward gliomasrdquoBiochemical and Biophysical Research Communications vol 407no 4 pp 741ndash746 2011
[20] S A Park C H Ryu S M Kim et al ldquoCXCR4-transfectedhuman umbilical cord blood-derived mesenchymal stem cellsexhibit enhanced migratory capacity toward gliomasrdquo Interna-tional Journal of Oncology vol 38 no 1 pp 97ndash103 2011
[21] R S Rachakatla F Marini M L Weiss M Tamura and DTroyer ldquoDevelopment of human umbilical cord matrix stemcell-based gene therapy for experimental lung tumorsrdquo CancerGene Therapy vol 14 no 10 pp 828ndash835 2007
[22] C Yan S Li Z Li et al ldquoHuman umbilical cord mesenchymalstem cells as vehicles of CD20-specific TRAIL fusion proteindelivery a double-target therapy against non-Hodgkinrsquos lym-phomardquo Molecular Pharmaceutics vol 10 no 1 pp 142ndash1512013
[23] J Hu A Dong V Fernandez-Ruiz et al ldquoBlockade of Wntsignaling inhibits angiogenesis and tumor growth in hepatocel-lular carcinomardquo Cancer Research vol 69 no 17 pp 6951ndash69592009
[24] L Xu A Y Hui E Albanis et al ldquoHuman hepatic stellate celllines LX-1 and LX-2 new tools for analysis of hepatic fibrosisrdquoGut vol 54 no 1 pp 142ndash151 2005
[25] M G Garcia J Bayo M F Bolontrade et al ldquoHepatocellularcarcinoma cells and their fibrotic microenvironment modulatebone marrow-derived mesenchymal stromal cell migration invitro and in vivordquo Molecular Pharmaceutics vol 8 no 5 pp1538ndash1548 2011
[26] M Dominici K le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells theInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[27] J Bayo E Fiore J B Aquino et al ldquoIncreased migration ofhumanmesenchymal stromal cells byAutocrineMotility Factor(AMF) resulted in enhanced recruitment towards hepatocellu-lar carcinomardquo PLoS ONE vol 9 no 4 Article ID e95171 2014
[28] M K Seong Y L Jung I P Sang et al ldquoGene therapyusing TRAIL-secreting human umbilical cord blood-derived
mesenchymal stem cells against intracranial gliomardquo CancerResearch vol 68 no 23 pp 9614ndash9623 2008
[29] R Ayuzawa C Doi R S Rachakatla et al ldquoNaıve humanumbilical cord matrix derived stem cells significantly attenuategrowth of human breast cancer cells in vitro and in vivordquoCancerLetters vol 280 no 1 pp 31ndash37 2009
[30] T Matsuzuka R S Rachakatla C Doi et al ldquoHuman umbilicalcord matrix-derived stem cells expressing interferon-120573 genesignificantly attenuate bronchioloalveolar carcinomaxenograftsin SCID micerdquo Lung Cancer vol 70 no 1 pp 28ndash36 2010
[31] G Li X Zhang H Wang et al ldquoComparative proteomicanalysis of mesenchymal stem cells derived from human bonemarrow umbilical cord and placenta implication in themigra-tionrdquo Proteomics vol 9 no 1 pp 20ndash30 2009
[32] H C Miranda R H Herai C H Thome et al ldquoA quan-titative proteomic and transcriptomic comparison of humanmesenchymal stem cells from bone marrow and umbilical cordveinrdquo Proteomics vol 12 no 17 pp 2607ndash2617 2012
[33] P U Le G Guay Y Altschuler and I R Nabi ldquoCaveolin-1 isa negative regulator of caveolae-mediated endocytosis to theendoplasmic reticulumrdquo The Journal of Biological Chemistryvol 277 no 5 pp 3371ndash3379 2002
[34] S Mishra A Raz and L J Murphy ldquoInsulin-like growthfactor binding protein-3 interacts with autocrine motility fac-torphosphoglucose isomerase (AMFPGI) and inhibits theAMFPGI functionrdquo Cancer Research vol 64 no 7 pp 2516ndash2522 2004
[35] F L Yu M H Liao J W Lee and W L Shih ldquoInductionof hepatoma cells migration by phosphoglucose isomeraseautocrine motility factor through the upregulation of matrixmetalloproteinase-3rdquo Biochemical and Biophysical ResearchCommunications vol 314 no 1 pp 76ndash82 2004
[36] A H Klopp A Gupta E Spaeth M Andreeff and F MariniIII ldquoConcise review dissecting a discrepancy in the literaturedomesenchymal stem cells support or suppress tumor growthrdquoStem Cells vol 29 no 1 pp 11ndash19 2011
[37] L Qiao Z Xu T Zhao et al ldquoSuppression of tumorigenesisby human mesenchymal stem cells in a hepatoma modelrdquo CellResearch vol 18 no 4 pp 500ndash507 2008
[38] Y R Lu Y Yuan X J Wang et al ldquoThe growth inhibitory effectof mesenchymal stem cells on tumor cells in vitro and in vivordquoCancer Biology andTherapy vol 7 no 2 pp 245ndash251 2008
[39] Z-G Zhao W Xu L Sun W-M Li Q-P Li and P Zou ldquoThecharacteristics and immunoregulatory functions of regulatorydendritic cells induced by mesenchymal stem cells derivedfrom bone marrow of patient with chronic myeloid leukaemiardquoEuropean Journal of Cancer vol 48 no 12 pp 1884ndash1895 2012
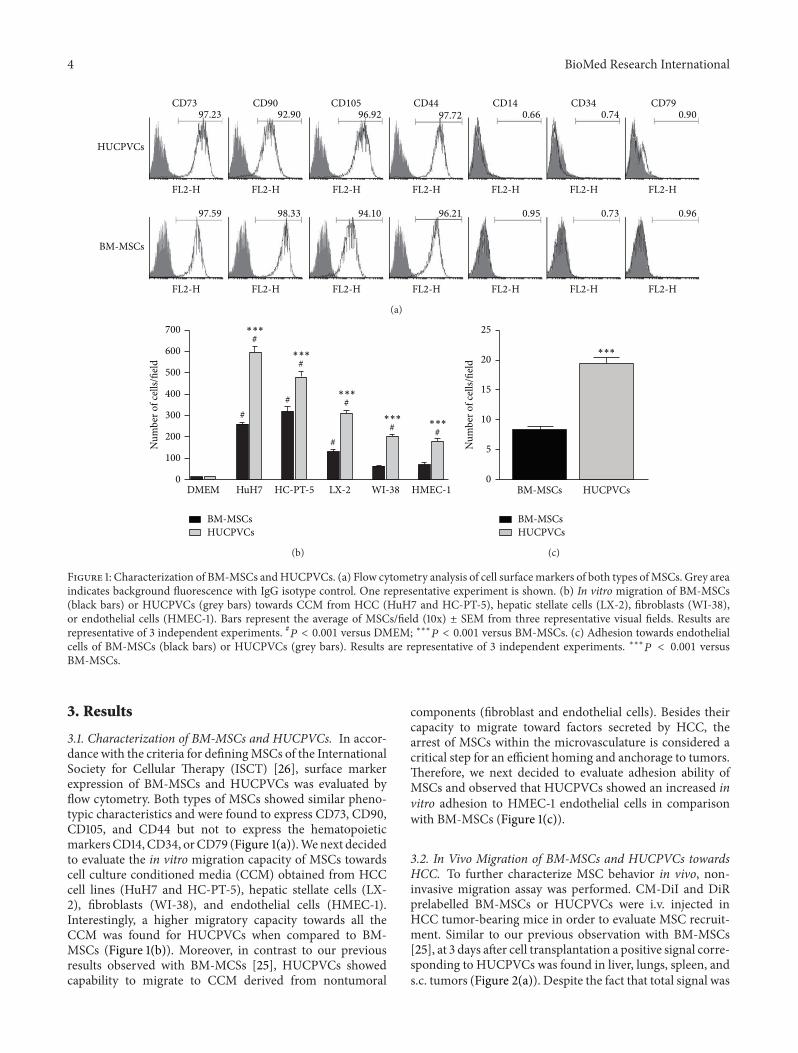
4 BioMed Research International
HUCPVCs
BM-MSCs
CD73 CD90 CD105 CD44 CD14 CD34 CD799723 9290 9692 9772 066 074 090
9759 9833 9410 9621 095 073 096
FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H
FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H FL2-H
(a)
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
BM-MSCsHUCPVCs
DMEM HuH7 HC-PT-5 LX-2 WI-38 HMEC-1
700
600
500
400
300
200
100
0
Num
ber o
f cel
lsfie
ld
(b)
lowastlowastlowast
BM-MSCsHUCPVCs
BM-MSCs HUCPVCs
Num
ber o
f cel
lsfie
ld
25
20
15
10
5
0
(c)
Figure 1 Characterization of BM-MSCs andHUCPVCs (a) Flow cytometry analysis of cell surfacemarkers of both types ofMSCs Grey areaindicates background fluorescence with IgG isotype control One representative experiment is shown (b) In vitro migration of BM-MSCs(black bars) or HUCPVCs (grey bars) towards CCM from HCC (HuH7 and HC-PT-5) hepatic stellate cells (LX-2) fibroblasts (WI-38)or endothelial cells (HMEC-1) Bars represent the average of MSCsfield (10x) plusmn SEM from three representative visual fields Results arerepresentative of 3 independent experiments 119875 lt 0001 versus DMEM lowastlowastlowast119875 lt 0001 versus BM-MSCs (c) Adhesion towards endothelialcells of BM-MSCs (black bars) or HUCPVCs (grey bars) Results are representative of 3 independent experiments lowastlowastlowast119875 lt 0001 versusBM-MSCs
3 Results
31 Characterization of BM-MSCs and HUCPVCs In accor-dance with the criteria for definingMSCs of the InternationalSociety for Cellular Therapy (ISCT) [26] surface markerexpression of BM-MSCs and HUCPVCs was evaluated byflow cytometry Both types of MSCs showed similar pheno-typic characteristics and were found to express CD73 CD90CD105 and CD44 but not to express the hematopoieticmarkersCD14CD34 orCD79 (Figure 1(a))Wenext decidedto evaluate the in vitro migration capacity of MSCs towardscell culture conditioned media (CCM) obtained from HCCcell lines (HuH7 and HC-PT-5) hepatic stellate cells (LX-2) fibroblasts (WI-38) and endothelial cells (HMEC-1)Interestingly a higher migratory capacity towards all theCCM was found for HUCPVCs when compared to BM-MSCs (Figure 1(b)) Moreover in contrast to our previousresults observed with BM-MCSs [25] HUCPVCs showedcapability to migrate to CCM derived from nontumoral
components (fibroblast and endothelial cells) Besides theircapacity to migrate toward factors secreted by HCC thearrest of MSCs within the microvasculature is considered acritical step for an efficient homing and anchorage to tumorsTherefore we next decided to evaluate adhesion ability ofMSCs and observed that HUCPVCs showed an increased invitro adhesion to HMEC-1 endothelial cells in comparisonwith BM-MSCs (Figure 1(c))
32 In Vivo Migration of BM-MSCs and HUCPVCs towardsHCC To further characterize MSC behavior in vivo non-invasive migration assay was performed CM-DiI and DiRprelabelled BM-MSCs or HUCPVCs were iv injected inHCC tumor-bearing mice in order to evaluate MSC recruit-ment Similar to our previous observation with BM-MSCs[25] at 3 days after cell transplantation a positive signal corre-sponding to HUCPVCs was found in liver lungs spleen andsc tumors (Figure 2(a)) Despite the fact that total signal was
BioMed Research International 5
50
40
30
20
10
times108
Radi
ant e
ffici
ency
(ps
cm
2s
r)(120583
Wc
m2)
(a)
lowastlowastlowast
BM-MSCsHUCPVCs
BM-MSCs HUCPVCsTota
l rad
iant
effici
ency
(ps
) (120583
Wc
m2) times1013
24
23
22
21
20
19
18
17
16
15
14
(b)
BM-MSCs HUCPVCs50
40
30
20
10
times108Ra
dian
t effi
cien
cy(p
sc
m2s
r)(120583
Wc
m2)
(c)
BM-MSCsHUCPVCs
Tota
l sig
nal (
) 90
8086420
Liver Lung Spleen Tumor
lowast
lowast
(d)
DAPI CM-DiI
(e)
BM-MSCsHUCPVCs
250
200
150
100
50
0HuH7 HC-PT-5
Num
ber o
f cel
lsfie
ld
lowastlowastlowast
lowastlowastlowast
(f)
Figure 2 In vivomigration of BM-MSCs and HUCPVCs CM-DiI and DiR prelabeled MSCs were iv injected in sc HuH7 tumor-bearingmice At day 3 mice were sacrificed and organs were removed lungs livers spleen (a) and tumors (c) were exposed to obtain FI Imagesrepresent the radiant efficiency Representative images are shown (b) Total FI for injected BM-MSCs or HUCPVCs was calculated bymeasuring the region of interest (ROI) for all the tissues isolated and results were expressed as total radiant efficiency [ps][120583Wcm2]lowastlowastlowast119875 lt 0001 (d) Signal present in the isolated liver spleen lungs and tumors was represented as percentage of total signal for BM-MSCs or
HUCPVCs-injected mice lowast119875 lt 005 versus BM-MSCs (e) Microscopic analysis of transplanted CM-DiI-labeled MSCs (red signal indicatedby arrows) andDAPI staining in frozen sections of tumors times200magnification (f) In vitromigration ofMSCs to TCMderived fromHuH7 orHC-PT-5 sc tumors Bars represent the average of MSCsfield (10x) plusmn SEM from three representative visual fields Results are representativeof 3 independent experiments lowastlowastlowast119875 lt 0001 versus BM-MSCs
6 BioMed Research International
BM-MSCsHUCPVCs
lowastlowastlowast
lowastlowastlowast
lowast20
15
10
05
00
Fold
chan
ge (2
ΔΔ
ct)
CXCR1 CXCR2 CCR2 IL-6R IL-6ST
(a)
BM-MSCsHUCPVCs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
300
2705
4
3
2
1
0
Fold
chan
ge (2
ΔΔ
ct)
AMFR MMP3 CAV-1 CAV-2 IGFBP3
(b)
Figure 3 Differential expression of cytokineschemokines receptors and AMFAMFR pathway in MSCs Expression of cytokines andchemokines receptors (a) and AMFAMFR axis proteins (b) was evaluated in BM-MSCs (black bars) or HUCPVCs (grey bars) by qPCRlowastlowastlowast119875 lt 0001 versus BM-MSCs
lower in mice injected with HUCPVCs compared to thoseinjected with BM-MSCs (Figure 2(b)) the percentage of totalsignal corresponding to sc tumor locations was increasedin mice administered with HUCPVCs in comparison withanimals that received BM-MSCs (Figures 2(c) and 2(d))indicating an enhanced engraftment of HUCPVCs into HCCtumors In the other evaluated tissues signal intensity wassimilar for BM-MSC or HUCPVCs in lung and liver andit was comparatively reduced in the spleen of HUCPVCs-injected mice (Figure 2(d)) Presence of MSCs in the sctumors was also confirmed by fluorescence microscopy(Figure 2(e)) Finally we evaluated whether MSCs mightpresent differential migratory capacity towards CM obtainedfrom sc tumors (TCM) Interestingly a greater in vitromigratory capacity towards TCM from HCC was observedfor HUCPVCs when compared to BM-MSCs (Figure 2(f))
33 AMFR Was Highly Expressed in HUCPVCs In orderto evaluate mechanisms partially explaining the differentialmigratory capacity of HUCPVCs compared to BM-MSCstowards tumor released factors we analyzed the expressionof some chemokinersquos receptor likely involved inMSC recruit-ment towards HCC Taking into account several reportsdemonstrating that interleukin- (IL-) 8 GRO chemokine(C-C motif) ligand (CCL)-2 and IL-6 are among the mostrelevant factors in HCC [17] we decided to evaluate by qPCRthe expression of CXCR1 CXCR2 CCR2 IL-6R and IL-6ST Interestingly constitutive CXCR1 and CXCR2 mRNAexpression was found to be lower and CCR2 slightly higherin HUCPVCs when compared to BM-MSCs while IL-6Rand IL-6ST expression was similar in both MSCs sources(Figure 3(a)) Next we decided to evaluate the axis of theautocrinemotility factor (AMF) a cytokine with chemotacticeffect on MSCs as recently observed by our group [27]By qPCR a significantly higher expression of the AMFreceptor (AMFR) was found in HUCPVCs when comparedto BM-MSCs Similarly genes known to be related to the
availability of the receptor in the cell surface such as caveolin-1 (CAV-1) and caveolin-2 (CAV-2) were also highly expressedin HUCPVCs as well as the metalloproteinase 3 (MMP3)necessary to the transmigration process In contrast expres-sion levels of insulin-like growth factor-binding protein 3(IGFBP3) a protein that negatively regulates AMFAMFRpathway were found to be reduced in HUCPVCs whencompared to BM-MSCs (Figure 3(b))
34 HUCPVCs Displayed Enhanced Migration towards AMFIn Vitro We have previously demonstrated that AMF acytokine produced by HCC cells plays a critical role inMSC migration [27] Due to the increased AMFR and AMF-AMFR-related proteins expression in HUCPVCs we decidedto test the in vitro migration response to the recombinantAMF (rAMF) of both types ofMSCsusing a chemotaxis assayA significantly higher migration to different doses of rAMF(05 and 075 120583gmL) was observed for HUCPVCs whencompared to BM-MSCs (Figure 4(a)) In spite of differenttypes of MSCs showing similar reduction in migration levels(50 of control) towards HuH7 TCM after the blockage withanti-AMF antibody (data not shown) preincubation of HC-PT-5 TCM with anti-AMF antibody (AMF-ab) resulted ina further reduction in HUCPVCs migration capacity (54of control) when compared to BM-MSCs (67 of control)(Figure 4(b))
4 Discussion
The aim of this study was to assess the capability of differentsources of MSCs to migrate towards HCC released factorsin order to select those showing higher tumor recruitmentcapacity for future therapeutic applications Our resultsindicated that although both types of MSCs share simi-lar phenotypic characteristics as MSCs HUCPVCs have ahigher potential to migrate in vitro towards the conditionedmedium derived not only from HCC cells but also from
BioMed Research International 7
BM-MSCsHUCPVCs
lowastlowastlowast
0 050 075
rAMF (120583gmL)
300
250
200
150
100
50
0
Con
trol (
)
(a)
BM-MSCsHUCPVCs
lowast
Con
trol (
)
110
90
100
70
50
30
10
80
60
40
20
0+ minus + minus
+minus +minus
IgG-abAMF-ab
(b)
Figure 4 HUCPVCs showed enhanced migration towards AMF in comparison with BM-MSCs (a) In vitromigration of BM-MSCs (blackbars) or HUCPVCs (grey bars) towards rAMF 119875 lt 005 versus DMEM (0 120583gmL rAMF) lowast119875 lt 005 versus BM-MSCs (b) In vitromigrationof BM-MSCs (black bars) or HUCPVCs (grey bars) towards HC-PT-5 TCM preincubated with anti-AMF antibody (AMF-ab) or controlisotype (IgG-ab) was evaluated 119875 lt 005 versus IgG-ab lowast119875 lt 005 versus BM-MSCs Bars represent the average of MSCsfield (10x) plusmn SEMfrom three representative visual fields Results are representative of 3 independent experiments
nontumoral components of the tumor microenvironmentlike activated hepatic stellate cells (LX-2) fibroblasts (WI-38) or tumor microendothelial cells (HMEC-1) MoreoverHUCPVCs also showed an enhanced adhesion capacityto HMEC-1 cells compared to BM-MSCs This enhancedadhesion to endothelial cells could allow a more efficientarrest in tumor microvasculature which is a required step forMSC recruitment into HCC In line with this we observedthat HUCPVCs also exhibited greater migration capacity invivo towards experimental HCC In contrast to that observedin the in vitro migration experiments HUCPVCs showed asimilar migration capability towards nontumoral tissues likeliver or lung and also showed a lower recruitment to thespleen AlthoughHUCPVCsmigrated better than BM-MSCstoward activated hepatic stellate cells in vitro no differenceswere observed in in vivo migration to normal liver wherehepatic stellate cells are quiescent In addition it should benoted that mice injected with HUCPVCs showed a lowertotal signal than those injected with BM-MSCs probablydue to differences in the DiR uptake for both kinds of cellsor in the number of inoculated cells These results are inagreement with previous works showing that MCSs derivedfrom umbilical cord are able tomigrate towards experimentaltumors such as glioma [28] breast cancer [29] and lung ade-nocarcinoma [30] Nonetheless to the best of our knowledgethis is the first report describing the comparative migrationcapacity of human umbilical cord perivascular cells towardshuman HCC
Tumor-homing is thought to be mediated by interactionsof cytokineschemokines with their specific receptors Anumber of cytokines growth factors and chemokines aresecreted by HCC cells and their microenvironment [17] Wehave recently found that AMF is critical for MSC migrationtoward HCC in vitro and in vivo Furthermore other factorsreleased by tumor stroma components like IL-8 GRO IL-6or CCL-2 could also be involved inMSC recruitment toHCCThus differences observed between HUCPVCs and BM-MSCs in their in vitro and in vivomigration capacity towardsHCC seem at least partially related to differences in theexpression of cytokinechemokine receptors Some studieshave screened for similarities or differences among BM-MSCs and UC-MSCs at the molecular level [31 32] In addi-tion a previous report demonstrated that migration of UCB-MSC towards glioma cells was higher when compared toBM-MSCs likely due to increased levels of the IL-8 receptorCXCR1 and CXCR2 in the former cell type [18] In contrastto these observations we found that HUCPVCs presentedlower levels of CXCR1 and CXCR2 expression compared toBM-MSCs and these differences could be explained partiallydue to differences in tissue source and culture conditions andmerit further investigationThe analysis of IL-6R and IL6-STexpression showed similar levels in both types ofMSCs whilelevels of MCP-1 receptor (CCR2) were slightly increased inHUCPVCs Interestingly we found that not only AMFR butalso CAV-1 and CAV-2 which are genes that regulate theavailability of AMFR on cell surface [33] were overexpressedin HUCPVCs compared to BM-MSCs In line with this a
8 BioMed Research International
reduced expression level of IGFBP3 a protein which canbind to AMF blocking its binding to AMFR [34] was foundin HUCPVCs when compared to BM-MSCs Moreover anincreased expression of MMP3 was observed in HUCPVCsThis matrix metalloproteinase is involved in AMF-inducedmigration and it is necessary for cell invasion and proteolysisof the extracellularmatrix [35] Finally and consistentwith anincrease in the expression of genes related to the AMFAMFRpathway not only HUCPVCs show an enhanced in vitromigration towards rAMF but also the specific blockageof AMF in HC-PT-5 TCM was able to reduce HUCPVCsmigration to a greater level when compared to BM-MSCs
The potential use of MSCs in cancer treatment as carriersof therapeutic genes has raised some concerns about thesafety of their use in the clinic There are some studiesthat indicated that MSCs have protumorigenic capacitydue to their immunosuppression properties modulation ofthe epithelial-to-mesenchymal transition and induction ofangiogenesis [36] However others have demonstrated thatMSCs decreased HCC growth [37 38] Similarly we havepreviously demonstrated that BM-MSCs did not modifytumor growth in vivo [25] Moreover we observed thatHUCPVCs did not induce tumor growth when injected ivin Huh7 sc tumor-bearing mice (data not shown) Theuse of HUCPVCs as cellular carriers could also have theadvantage of its allogeneic application because of its lowimmunogenicity [9] In this regard HUCPVCs could beuseful as carriers of therapeutic genes for cancer patientswhere isolation of BM or adipose tissue-derived MSCs couldbe less responsive to chemotactic factors and might dis-play higher immunosuppression capacities than MSCs fromhealthy donors [39] or even present genetic abnormalities
5 Conclusions
In summary our results demonstrate a greater migrationcapacity to HCC of HUCPVCs when compared to BM-MSCs not only in vitro but also in vivo likely due to anincrease in migratory response to AMF and to an enhancedadhesive capacity to tumor microvasculature Consideringtheir availability and that no invasive procedures are requiredto obtain HUCPVCs these cells have advantages over BM-MSCs as candidates for carriers of therapeutic genes for thetreatment of HCC
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors gratefully thank Guillermo Gaston MarcosCabrera and Vanina Ferreira for technical assistance andDr Mengshiao Meng (National Chung Hsing University(NCHU) Taichung Taiwan) for providing recombinantAMF This work was supported by Grants from Austral
University (for Juan Bayo I04-12 for Mariana G Garcia T13-12 and for Guillermo Mazzolini T13-11) and from AgenciaNacional de Promocion Cientıfica y Tecnologica (ANPCyT)PICTO 200800115 (Mariana G Garcia) PICT 200800123(Jorge B Aquino) PICTO 200800122 (Jorge B Aquino)and PICT 20102818 (Mariana G Garcia and GuillermoMazzolini)
References
[1] H B El-Serag J A Marrero L Rudolph and K R ReddyldquoDiagnosis and treatment of hepatocellular carcinomardquo Gas-troenterology vol 134 no 6 pp 1752ndash1763 2008
[2] J M Llovet S Ricci V Mazzaferro et al ldquoSorafenib inadvanced hepatocellular carcinomardquo The New England Journalof Medicine vol 359 no 4 pp 378ndash390 2008
[3] A J Friedenstein J F Gorskaja andN N Kulagina ldquoFibroblastprecursors in normal and irradiated mouse hematopoieticorgansrdquo Experimental Hematology vol 4 no 5 pp 267ndash2741976
[4] D J Prockop and J Y Oh ldquoMedical therapies with adultstemprogenitor cells (MSCs) a backward journey from dra-matic results in vivo to the cellular andmolecular explanationsrdquoJournal of Cellular Biochemistry vol 113 no 5 pp 1460ndash14692012
[5] J B Aquino M F Bolontrade M G Garcıa O L Podhajcerand GMazzolini ldquoMesenchymal stem cells as therapeutic toolsand gene carriers in liver fibrosis and hepatocellular carcinomardquoGene Therapy vol 17 no 6 pp 692ndash708 2010
[6] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[7] H Yoshimura T Muneta A Nimura A Yokoyama H Kogaand I Sekiya ldquoComparison of rat mesenchymal stem cellsderived from bone marrow synovium periosteum adiposetissue and musclerdquo Cell and Tissue Research vol 327 no 3 pp449ndash462 2007
[8] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popu-lations and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[9] R Sarugaser D Lickorish D Baksh M M Hosseini and J EDavies ldquoHuman umbilical cord perivascular (HUCPV) cells asource of mesenchymal progenitorsrdquo Stem Cells vol 23 no 2pp 220ndash229 2005
[10] D Baksh R Yao and R S Tuan ldquoComparison of proliferativeand multilineage differentiation potential of human mesenchy-mal stem cells derived from umbilical cord and bone marrowrdquoStem Cells vol 25 no 6 pp 1384ndash1392 2007
[11] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[12] O K Lee T K Kuo W M Chen K D Lee S L Hsieh andT H Chen ldquoIsolation of multipotent mesenchymal stem cellsfrom umbilical cord bloodrdquo Blood vol 103 no 5 pp 1669ndash16752004
[13] HWang S Hung S Peng et al ldquoMesenchymal stem cells in theWhartonrsquos jelly of the human umbilical cordrdquo StemCells vol 22no 7 pp 1330ndash1337 2004
BioMed Research International 9
[14] J Kim J M Shin Y J Jeon H M Chung and J Chae ldquoPro-teomic validation of multifunctional molecules in mesenchy-mal stem cells derived from human bone marrow umbilicalcord blood and peripheral bloodrdquo PLoS ONE vol 7 no 5Article ID e32350 2012
[15] J-Y Hsieh H-W Wang S-J Chang et al ldquoMesenchymalstem cells from human umbilical cord express preferentiallysecreted factors related to neuroprotection neurogenesis andangiogenesisrdquo PLoS ONE vol 8 no 8 Article ID e72604 2013
[16] V Hernandez-Gea S Toffanin S L Friedman and J MLlovet ldquoRole of the microenvironment in the pathogenesis andtreatment of hepatocellular carcinomardquo Gastroenterology vol144 no 3 pp 512ndash527 2013
[17] J Bayo M Marrodan J B Aquino M Silva M G Garciaand G Mazzolini ldquoThe therapeutic potential of bone marrow-derived mesenchymal stromal cells on hepatocellular carci-nomardquo Liver International vol 34 no 3 pp 330ndash334 2014
[18] D Kim J H Kim J Kwon Lee et al ldquoOverexpression ofCXC chemokine receptors is required for the superior glioma-tracking property of umbilical cord blood-derived mesenchy-mal stem cellsrdquo Stem Cells and Development vol 18 no 3 pp511ndash519 2009
[19] S M Kim D Kim C H Jeong et al ldquoCXC chemokinereceptor 1 enhances the ability of human umbilical cord blood-derived mesenchymal stem cells to migrate toward gliomasrdquoBiochemical and Biophysical Research Communications vol 407no 4 pp 741ndash746 2011
[20] S A Park C H Ryu S M Kim et al ldquoCXCR4-transfectedhuman umbilical cord blood-derived mesenchymal stem cellsexhibit enhanced migratory capacity toward gliomasrdquo Interna-tional Journal of Oncology vol 38 no 1 pp 97ndash103 2011
[21] R S Rachakatla F Marini M L Weiss M Tamura and DTroyer ldquoDevelopment of human umbilical cord matrix stemcell-based gene therapy for experimental lung tumorsrdquo CancerGene Therapy vol 14 no 10 pp 828ndash835 2007
[22] C Yan S Li Z Li et al ldquoHuman umbilical cord mesenchymalstem cells as vehicles of CD20-specific TRAIL fusion proteindelivery a double-target therapy against non-Hodgkinrsquos lym-phomardquo Molecular Pharmaceutics vol 10 no 1 pp 142ndash1512013
[23] J Hu A Dong V Fernandez-Ruiz et al ldquoBlockade of Wntsignaling inhibits angiogenesis and tumor growth in hepatocel-lular carcinomardquo Cancer Research vol 69 no 17 pp 6951ndash69592009
[24] L Xu A Y Hui E Albanis et al ldquoHuman hepatic stellate celllines LX-1 and LX-2 new tools for analysis of hepatic fibrosisrdquoGut vol 54 no 1 pp 142ndash151 2005
[25] M G Garcia J Bayo M F Bolontrade et al ldquoHepatocellularcarcinoma cells and their fibrotic microenvironment modulatebone marrow-derived mesenchymal stromal cell migration invitro and in vivordquo Molecular Pharmaceutics vol 8 no 5 pp1538ndash1548 2011
[26] M Dominici K le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells theInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[27] J Bayo E Fiore J B Aquino et al ldquoIncreased migration ofhumanmesenchymal stromal cells byAutocrineMotility Factor(AMF) resulted in enhanced recruitment towards hepatocellu-lar carcinomardquo PLoS ONE vol 9 no 4 Article ID e95171 2014
[28] M K Seong Y L Jung I P Sang et al ldquoGene therapyusing TRAIL-secreting human umbilical cord blood-derived
mesenchymal stem cells against intracranial gliomardquo CancerResearch vol 68 no 23 pp 9614ndash9623 2008
[29] R Ayuzawa C Doi R S Rachakatla et al ldquoNaıve humanumbilical cord matrix derived stem cells significantly attenuategrowth of human breast cancer cells in vitro and in vivordquoCancerLetters vol 280 no 1 pp 31ndash37 2009
[30] T Matsuzuka R S Rachakatla C Doi et al ldquoHuman umbilicalcord matrix-derived stem cells expressing interferon-120573 genesignificantly attenuate bronchioloalveolar carcinomaxenograftsin SCID micerdquo Lung Cancer vol 70 no 1 pp 28ndash36 2010
[31] G Li X Zhang H Wang et al ldquoComparative proteomicanalysis of mesenchymal stem cells derived from human bonemarrow umbilical cord and placenta implication in themigra-tionrdquo Proteomics vol 9 no 1 pp 20ndash30 2009
[32] H C Miranda R H Herai C H Thome et al ldquoA quan-titative proteomic and transcriptomic comparison of humanmesenchymal stem cells from bone marrow and umbilical cordveinrdquo Proteomics vol 12 no 17 pp 2607ndash2617 2012
[33] P U Le G Guay Y Altschuler and I R Nabi ldquoCaveolin-1 isa negative regulator of caveolae-mediated endocytosis to theendoplasmic reticulumrdquo The Journal of Biological Chemistryvol 277 no 5 pp 3371ndash3379 2002
[34] S Mishra A Raz and L J Murphy ldquoInsulin-like growthfactor binding protein-3 interacts with autocrine motility fac-torphosphoglucose isomerase (AMFPGI) and inhibits theAMFPGI functionrdquo Cancer Research vol 64 no 7 pp 2516ndash2522 2004
[35] F L Yu M H Liao J W Lee and W L Shih ldquoInductionof hepatoma cells migration by phosphoglucose isomeraseautocrine motility factor through the upregulation of matrixmetalloproteinase-3rdquo Biochemical and Biophysical ResearchCommunications vol 314 no 1 pp 76ndash82 2004
[36] A H Klopp A Gupta E Spaeth M Andreeff and F MariniIII ldquoConcise review dissecting a discrepancy in the literaturedomesenchymal stem cells support or suppress tumor growthrdquoStem Cells vol 29 no 1 pp 11ndash19 2011
[37] L Qiao Z Xu T Zhao et al ldquoSuppression of tumorigenesisby human mesenchymal stem cells in a hepatoma modelrdquo CellResearch vol 18 no 4 pp 500ndash507 2008
[38] Y R Lu Y Yuan X J Wang et al ldquoThe growth inhibitory effectof mesenchymal stem cells on tumor cells in vitro and in vivordquoCancer Biology andTherapy vol 7 no 2 pp 245ndash251 2008
[39] Z-G Zhao W Xu L Sun W-M Li Q-P Li and P Zou ldquoThecharacteristics and immunoregulatory functions of regulatorydendritic cells induced by mesenchymal stem cells derivedfrom bone marrow of patient with chronic myeloid leukaemiardquoEuropean Journal of Cancer vol 48 no 12 pp 1884ndash1895 2012
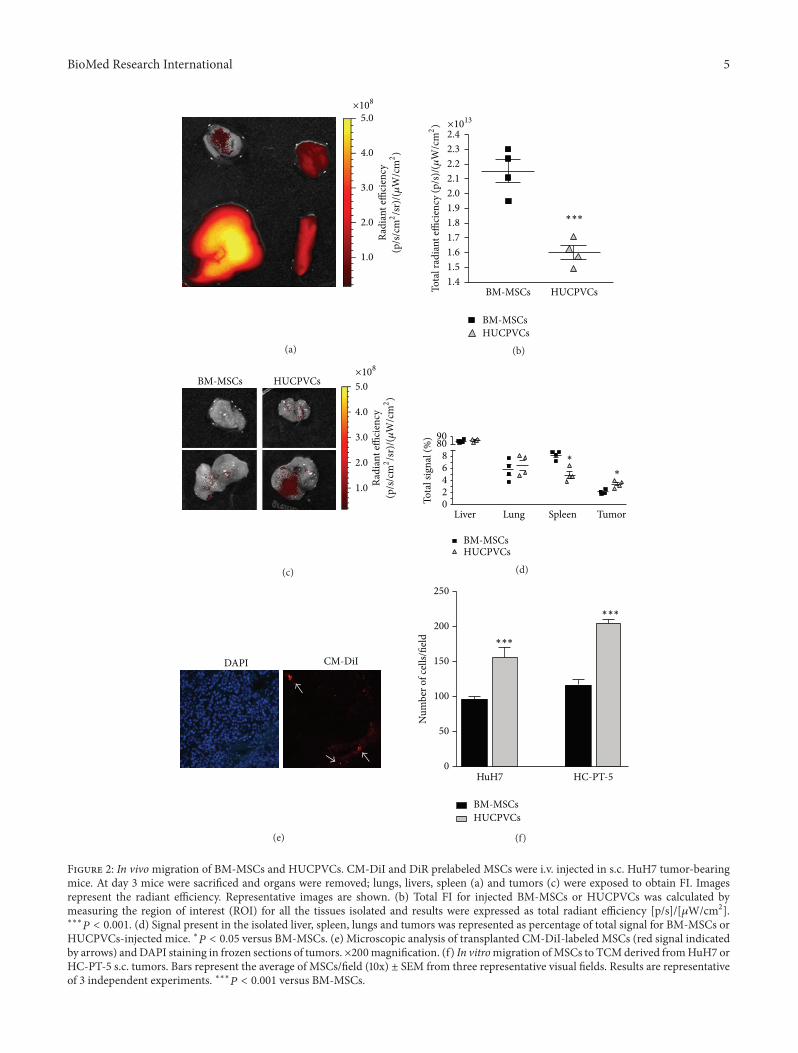
BioMed Research International 5
50
40
30
20
10
times108
Radi
ant e
ffici
ency
(ps
cm
2s
r)(120583
Wc
m2)
(a)
lowastlowastlowast
BM-MSCsHUCPVCs
BM-MSCs HUCPVCsTota
l rad
iant
effici
ency
(ps
) (120583
Wc
m2) times1013
24
23
22
21
20
19
18
17
16
15
14
(b)
BM-MSCs HUCPVCs50
40
30
20
10
times108Ra
dian
t effi
cien
cy(p
sc
m2s
r)(120583
Wc
m2)
(c)
BM-MSCsHUCPVCs
Tota
l sig
nal (
) 90
8086420
Liver Lung Spleen Tumor
lowast
lowast
(d)
DAPI CM-DiI
(e)
BM-MSCsHUCPVCs
250
200
150
100
50
0HuH7 HC-PT-5
Num
ber o
f cel
lsfie
ld
lowastlowastlowast
lowastlowastlowast
(f)
Figure 2 In vivomigration of BM-MSCs and HUCPVCs CM-DiI and DiR prelabeled MSCs were iv injected in sc HuH7 tumor-bearingmice At day 3 mice were sacrificed and organs were removed lungs livers spleen (a) and tumors (c) were exposed to obtain FI Imagesrepresent the radiant efficiency Representative images are shown (b) Total FI for injected BM-MSCs or HUCPVCs was calculated bymeasuring the region of interest (ROI) for all the tissues isolated and results were expressed as total radiant efficiency [ps][120583Wcm2]lowastlowastlowast119875 lt 0001 (d) Signal present in the isolated liver spleen lungs and tumors was represented as percentage of total signal for BM-MSCs or
HUCPVCs-injected mice lowast119875 lt 005 versus BM-MSCs (e) Microscopic analysis of transplanted CM-DiI-labeled MSCs (red signal indicatedby arrows) andDAPI staining in frozen sections of tumors times200magnification (f) In vitromigration ofMSCs to TCMderived fromHuH7 orHC-PT-5 sc tumors Bars represent the average of MSCsfield (10x) plusmn SEM from three representative visual fields Results are representativeof 3 independent experiments lowastlowastlowast119875 lt 0001 versus BM-MSCs
6 BioMed Research International
BM-MSCsHUCPVCs
lowastlowastlowast
lowastlowastlowast
lowast20
15
10
05
00
Fold
chan
ge (2
ΔΔ
ct)
CXCR1 CXCR2 CCR2 IL-6R IL-6ST
(a)
BM-MSCsHUCPVCs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
300
2705
4
3
2
1
0
Fold
chan
ge (2
ΔΔ
ct)
AMFR MMP3 CAV-1 CAV-2 IGFBP3
(b)
Figure 3 Differential expression of cytokineschemokines receptors and AMFAMFR pathway in MSCs Expression of cytokines andchemokines receptors (a) and AMFAMFR axis proteins (b) was evaluated in BM-MSCs (black bars) or HUCPVCs (grey bars) by qPCRlowastlowastlowast119875 lt 0001 versus BM-MSCs
lower in mice injected with HUCPVCs compared to thoseinjected with BM-MSCs (Figure 2(b)) the percentage of totalsignal corresponding to sc tumor locations was increasedin mice administered with HUCPVCs in comparison withanimals that received BM-MSCs (Figures 2(c) and 2(d))indicating an enhanced engraftment of HUCPVCs into HCCtumors In the other evaluated tissues signal intensity wassimilar for BM-MSC or HUCPVCs in lung and liver andit was comparatively reduced in the spleen of HUCPVCs-injected mice (Figure 2(d)) Presence of MSCs in the sctumors was also confirmed by fluorescence microscopy(Figure 2(e)) Finally we evaluated whether MSCs mightpresent differential migratory capacity towards CM obtainedfrom sc tumors (TCM) Interestingly a greater in vitromigratory capacity towards TCM from HCC was observedfor HUCPVCs when compared to BM-MSCs (Figure 2(f))
33 AMFR Was Highly Expressed in HUCPVCs In orderto evaluate mechanisms partially explaining the differentialmigratory capacity of HUCPVCs compared to BM-MSCstowards tumor released factors we analyzed the expressionof some chemokinersquos receptor likely involved inMSC recruit-ment towards HCC Taking into account several reportsdemonstrating that interleukin- (IL-) 8 GRO chemokine(C-C motif) ligand (CCL)-2 and IL-6 are among the mostrelevant factors in HCC [17] we decided to evaluate by qPCRthe expression of CXCR1 CXCR2 CCR2 IL-6R and IL-6ST Interestingly constitutive CXCR1 and CXCR2 mRNAexpression was found to be lower and CCR2 slightly higherin HUCPVCs when compared to BM-MSCs while IL-6Rand IL-6ST expression was similar in both MSCs sources(Figure 3(a)) Next we decided to evaluate the axis of theautocrinemotility factor (AMF) a cytokine with chemotacticeffect on MSCs as recently observed by our group [27]By qPCR a significantly higher expression of the AMFreceptor (AMFR) was found in HUCPVCs when comparedto BM-MSCs Similarly genes known to be related to the
availability of the receptor in the cell surface such as caveolin-1 (CAV-1) and caveolin-2 (CAV-2) were also highly expressedin HUCPVCs as well as the metalloproteinase 3 (MMP3)necessary to the transmigration process In contrast expres-sion levels of insulin-like growth factor-binding protein 3(IGFBP3) a protein that negatively regulates AMFAMFRpathway were found to be reduced in HUCPVCs whencompared to BM-MSCs (Figure 3(b))
34 HUCPVCs Displayed Enhanced Migration towards AMFIn Vitro We have previously demonstrated that AMF acytokine produced by HCC cells plays a critical role inMSC migration [27] Due to the increased AMFR and AMF-AMFR-related proteins expression in HUCPVCs we decidedto test the in vitro migration response to the recombinantAMF (rAMF) of both types ofMSCsusing a chemotaxis assayA significantly higher migration to different doses of rAMF(05 and 075 120583gmL) was observed for HUCPVCs whencompared to BM-MSCs (Figure 4(a)) In spite of differenttypes of MSCs showing similar reduction in migration levels(50 of control) towards HuH7 TCM after the blockage withanti-AMF antibody (data not shown) preincubation of HC-PT-5 TCM with anti-AMF antibody (AMF-ab) resulted ina further reduction in HUCPVCs migration capacity (54of control) when compared to BM-MSCs (67 of control)(Figure 4(b))
4 Discussion
The aim of this study was to assess the capability of differentsources of MSCs to migrate towards HCC released factorsin order to select those showing higher tumor recruitmentcapacity for future therapeutic applications Our resultsindicated that although both types of MSCs share simi-lar phenotypic characteristics as MSCs HUCPVCs have ahigher potential to migrate in vitro towards the conditionedmedium derived not only from HCC cells but also from
BioMed Research International 7
BM-MSCsHUCPVCs
lowastlowastlowast
0 050 075
rAMF (120583gmL)
300
250
200
150
100
50
0
Con
trol (
)
(a)
BM-MSCsHUCPVCs
lowast
Con
trol (
)
110
90
100
70
50
30
10
80
60
40
20
0+ minus + minus
+minus +minus
IgG-abAMF-ab
(b)
Figure 4 HUCPVCs showed enhanced migration towards AMF in comparison with BM-MSCs (a) In vitromigration of BM-MSCs (blackbars) or HUCPVCs (grey bars) towards rAMF 119875 lt 005 versus DMEM (0 120583gmL rAMF) lowast119875 lt 005 versus BM-MSCs (b) In vitromigrationof BM-MSCs (black bars) or HUCPVCs (grey bars) towards HC-PT-5 TCM preincubated with anti-AMF antibody (AMF-ab) or controlisotype (IgG-ab) was evaluated 119875 lt 005 versus IgG-ab lowast119875 lt 005 versus BM-MSCs Bars represent the average of MSCsfield (10x) plusmn SEMfrom three representative visual fields Results are representative of 3 independent experiments
nontumoral components of the tumor microenvironmentlike activated hepatic stellate cells (LX-2) fibroblasts (WI-38) or tumor microendothelial cells (HMEC-1) MoreoverHUCPVCs also showed an enhanced adhesion capacityto HMEC-1 cells compared to BM-MSCs This enhancedadhesion to endothelial cells could allow a more efficientarrest in tumor microvasculature which is a required step forMSC recruitment into HCC In line with this we observedthat HUCPVCs also exhibited greater migration capacity invivo towards experimental HCC In contrast to that observedin the in vitro migration experiments HUCPVCs showed asimilar migration capability towards nontumoral tissues likeliver or lung and also showed a lower recruitment to thespleen AlthoughHUCPVCsmigrated better than BM-MSCstoward activated hepatic stellate cells in vitro no differenceswere observed in in vivo migration to normal liver wherehepatic stellate cells are quiescent In addition it should benoted that mice injected with HUCPVCs showed a lowertotal signal than those injected with BM-MSCs probablydue to differences in the DiR uptake for both kinds of cellsor in the number of inoculated cells These results are inagreement with previous works showing that MCSs derivedfrom umbilical cord are able tomigrate towards experimentaltumors such as glioma [28] breast cancer [29] and lung ade-nocarcinoma [30] Nonetheless to the best of our knowledgethis is the first report describing the comparative migrationcapacity of human umbilical cord perivascular cells towardshuman HCC
Tumor-homing is thought to be mediated by interactionsof cytokineschemokines with their specific receptors Anumber of cytokines growth factors and chemokines aresecreted by HCC cells and their microenvironment [17] Wehave recently found that AMF is critical for MSC migrationtoward HCC in vitro and in vivo Furthermore other factorsreleased by tumor stroma components like IL-8 GRO IL-6or CCL-2 could also be involved inMSC recruitment toHCCThus differences observed between HUCPVCs and BM-MSCs in their in vitro and in vivomigration capacity towardsHCC seem at least partially related to differences in theexpression of cytokinechemokine receptors Some studieshave screened for similarities or differences among BM-MSCs and UC-MSCs at the molecular level [31 32] In addi-tion a previous report demonstrated that migration of UCB-MSC towards glioma cells was higher when compared toBM-MSCs likely due to increased levels of the IL-8 receptorCXCR1 and CXCR2 in the former cell type [18] In contrastto these observations we found that HUCPVCs presentedlower levels of CXCR1 and CXCR2 expression compared toBM-MSCs and these differences could be explained partiallydue to differences in tissue source and culture conditions andmerit further investigationThe analysis of IL-6R and IL6-STexpression showed similar levels in both types ofMSCs whilelevels of MCP-1 receptor (CCR2) were slightly increased inHUCPVCs Interestingly we found that not only AMFR butalso CAV-1 and CAV-2 which are genes that regulate theavailability of AMFR on cell surface [33] were overexpressedin HUCPVCs compared to BM-MSCs In line with this a
8 BioMed Research International
reduced expression level of IGFBP3 a protein which canbind to AMF blocking its binding to AMFR [34] was foundin HUCPVCs when compared to BM-MSCs Moreover anincreased expression of MMP3 was observed in HUCPVCsThis matrix metalloproteinase is involved in AMF-inducedmigration and it is necessary for cell invasion and proteolysisof the extracellularmatrix [35] Finally and consistentwith anincrease in the expression of genes related to the AMFAMFRpathway not only HUCPVCs show an enhanced in vitromigration towards rAMF but also the specific blockageof AMF in HC-PT-5 TCM was able to reduce HUCPVCsmigration to a greater level when compared to BM-MSCs
The potential use of MSCs in cancer treatment as carriersof therapeutic genes has raised some concerns about thesafety of their use in the clinic There are some studiesthat indicated that MSCs have protumorigenic capacitydue to their immunosuppression properties modulation ofthe epithelial-to-mesenchymal transition and induction ofangiogenesis [36] However others have demonstrated thatMSCs decreased HCC growth [37 38] Similarly we havepreviously demonstrated that BM-MSCs did not modifytumor growth in vivo [25] Moreover we observed thatHUCPVCs did not induce tumor growth when injected ivin Huh7 sc tumor-bearing mice (data not shown) Theuse of HUCPVCs as cellular carriers could also have theadvantage of its allogeneic application because of its lowimmunogenicity [9] In this regard HUCPVCs could beuseful as carriers of therapeutic genes for cancer patientswhere isolation of BM or adipose tissue-derived MSCs couldbe less responsive to chemotactic factors and might dis-play higher immunosuppression capacities than MSCs fromhealthy donors [39] or even present genetic abnormalities
5 Conclusions
In summary our results demonstrate a greater migrationcapacity to HCC of HUCPVCs when compared to BM-MSCs not only in vitro but also in vivo likely due to anincrease in migratory response to AMF and to an enhancedadhesive capacity to tumor microvasculature Consideringtheir availability and that no invasive procedures are requiredto obtain HUCPVCs these cells have advantages over BM-MSCs as candidates for carriers of therapeutic genes for thetreatment of HCC
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors gratefully thank Guillermo Gaston MarcosCabrera and Vanina Ferreira for technical assistance andDr Mengshiao Meng (National Chung Hsing University(NCHU) Taichung Taiwan) for providing recombinantAMF This work was supported by Grants from Austral
University (for Juan Bayo I04-12 for Mariana G Garcia T13-12 and for Guillermo Mazzolini T13-11) and from AgenciaNacional de Promocion Cientıfica y Tecnologica (ANPCyT)PICTO 200800115 (Mariana G Garcia) PICT 200800123(Jorge B Aquino) PICTO 200800122 (Jorge B Aquino)and PICT 20102818 (Mariana G Garcia and GuillermoMazzolini)
References
[1] H B El-Serag J A Marrero L Rudolph and K R ReddyldquoDiagnosis and treatment of hepatocellular carcinomardquo Gas-troenterology vol 134 no 6 pp 1752ndash1763 2008
[2] J M Llovet S Ricci V Mazzaferro et al ldquoSorafenib inadvanced hepatocellular carcinomardquo The New England Journalof Medicine vol 359 no 4 pp 378ndash390 2008
[3] A J Friedenstein J F Gorskaja andN N Kulagina ldquoFibroblastprecursors in normal and irradiated mouse hematopoieticorgansrdquo Experimental Hematology vol 4 no 5 pp 267ndash2741976
[4] D J Prockop and J Y Oh ldquoMedical therapies with adultstemprogenitor cells (MSCs) a backward journey from dra-matic results in vivo to the cellular andmolecular explanationsrdquoJournal of Cellular Biochemistry vol 113 no 5 pp 1460ndash14692012
[5] J B Aquino M F Bolontrade M G Garcıa O L Podhajcerand GMazzolini ldquoMesenchymal stem cells as therapeutic toolsand gene carriers in liver fibrosis and hepatocellular carcinomardquoGene Therapy vol 17 no 6 pp 692ndash708 2010
[6] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[7] H Yoshimura T Muneta A Nimura A Yokoyama H Kogaand I Sekiya ldquoComparison of rat mesenchymal stem cellsderived from bone marrow synovium periosteum adiposetissue and musclerdquo Cell and Tissue Research vol 327 no 3 pp449ndash462 2007
[8] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popu-lations and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[9] R Sarugaser D Lickorish D Baksh M M Hosseini and J EDavies ldquoHuman umbilical cord perivascular (HUCPV) cells asource of mesenchymal progenitorsrdquo Stem Cells vol 23 no 2pp 220ndash229 2005
[10] D Baksh R Yao and R S Tuan ldquoComparison of proliferativeand multilineage differentiation potential of human mesenchy-mal stem cells derived from umbilical cord and bone marrowrdquoStem Cells vol 25 no 6 pp 1384ndash1392 2007
[11] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[12] O K Lee T K Kuo W M Chen K D Lee S L Hsieh andT H Chen ldquoIsolation of multipotent mesenchymal stem cellsfrom umbilical cord bloodrdquo Blood vol 103 no 5 pp 1669ndash16752004
[13] HWang S Hung S Peng et al ldquoMesenchymal stem cells in theWhartonrsquos jelly of the human umbilical cordrdquo StemCells vol 22no 7 pp 1330ndash1337 2004
BioMed Research International 9
[14] J Kim J M Shin Y J Jeon H M Chung and J Chae ldquoPro-teomic validation of multifunctional molecules in mesenchy-mal stem cells derived from human bone marrow umbilicalcord blood and peripheral bloodrdquo PLoS ONE vol 7 no 5Article ID e32350 2012
[15] J-Y Hsieh H-W Wang S-J Chang et al ldquoMesenchymalstem cells from human umbilical cord express preferentiallysecreted factors related to neuroprotection neurogenesis andangiogenesisrdquo PLoS ONE vol 8 no 8 Article ID e72604 2013
[16] V Hernandez-Gea S Toffanin S L Friedman and J MLlovet ldquoRole of the microenvironment in the pathogenesis andtreatment of hepatocellular carcinomardquo Gastroenterology vol144 no 3 pp 512ndash527 2013
[17] J Bayo M Marrodan J B Aquino M Silva M G Garciaand G Mazzolini ldquoThe therapeutic potential of bone marrow-derived mesenchymal stromal cells on hepatocellular carci-nomardquo Liver International vol 34 no 3 pp 330ndash334 2014
[18] D Kim J H Kim J Kwon Lee et al ldquoOverexpression ofCXC chemokine receptors is required for the superior glioma-tracking property of umbilical cord blood-derived mesenchy-mal stem cellsrdquo Stem Cells and Development vol 18 no 3 pp511ndash519 2009
[19] S M Kim D Kim C H Jeong et al ldquoCXC chemokinereceptor 1 enhances the ability of human umbilical cord blood-derived mesenchymal stem cells to migrate toward gliomasrdquoBiochemical and Biophysical Research Communications vol 407no 4 pp 741ndash746 2011
[20] S A Park C H Ryu S M Kim et al ldquoCXCR4-transfectedhuman umbilical cord blood-derived mesenchymal stem cellsexhibit enhanced migratory capacity toward gliomasrdquo Interna-tional Journal of Oncology vol 38 no 1 pp 97ndash103 2011
[21] R S Rachakatla F Marini M L Weiss M Tamura and DTroyer ldquoDevelopment of human umbilical cord matrix stemcell-based gene therapy for experimental lung tumorsrdquo CancerGene Therapy vol 14 no 10 pp 828ndash835 2007
[22] C Yan S Li Z Li et al ldquoHuman umbilical cord mesenchymalstem cells as vehicles of CD20-specific TRAIL fusion proteindelivery a double-target therapy against non-Hodgkinrsquos lym-phomardquo Molecular Pharmaceutics vol 10 no 1 pp 142ndash1512013
[23] J Hu A Dong V Fernandez-Ruiz et al ldquoBlockade of Wntsignaling inhibits angiogenesis and tumor growth in hepatocel-lular carcinomardquo Cancer Research vol 69 no 17 pp 6951ndash69592009
[24] L Xu A Y Hui E Albanis et al ldquoHuman hepatic stellate celllines LX-1 and LX-2 new tools for analysis of hepatic fibrosisrdquoGut vol 54 no 1 pp 142ndash151 2005
[25] M G Garcia J Bayo M F Bolontrade et al ldquoHepatocellularcarcinoma cells and their fibrotic microenvironment modulatebone marrow-derived mesenchymal stromal cell migration invitro and in vivordquo Molecular Pharmaceutics vol 8 no 5 pp1538ndash1548 2011
[26] M Dominici K le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells theInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[27] J Bayo E Fiore J B Aquino et al ldquoIncreased migration ofhumanmesenchymal stromal cells byAutocrineMotility Factor(AMF) resulted in enhanced recruitment towards hepatocellu-lar carcinomardquo PLoS ONE vol 9 no 4 Article ID e95171 2014
[28] M K Seong Y L Jung I P Sang et al ldquoGene therapyusing TRAIL-secreting human umbilical cord blood-derived
mesenchymal stem cells against intracranial gliomardquo CancerResearch vol 68 no 23 pp 9614ndash9623 2008
[29] R Ayuzawa C Doi R S Rachakatla et al ldquoNaıve humanumbilical cord matrix derived stem cells significantly attenuategrowth of human breast cancer cells in vitro and in vivordquoCancerLetters vol 280 no 1 pp 31ndash37 2009
[30] T Matsuzuka R S Rachakatla C Doi et al ldquoHuman umbilicalcord matrix-derived stem cells expressing interferon-120573 genesignificantly attenuate bronchioloalveolar carcinomaxenograftsin SCID micerdquo Lung Cancer vol 70 no 1 pp 28ndash36 2010
[31] G Li X Zhang H Wang et al ldquoComparative proteomicanalysis of mesenchymal stem cells derived from human bonemarrow umbilical cord and placenta implication in themigra-tionrdquo Proteomics vol 9 no 1 pp 20ndash30 2009
[32] H C Miranda R H Herai C H Thome et al ldquoA quan-titative proteomic and transcriptomic comparison of humanmesenchymal stem cells from bone marrow and umbilical cordveinrdquo Proteomics vol 12 no 17 pp 2607ndash2617 2012
[33] P U Le G Guay Y Altschuler and I R Nabi ldquoCaveolin-1 isa negative regulator of caveolae-mediated endocytosis to theendoplasmic reticulumrdquo The Journal of Biological Chemistryvol 277 no 5 pp 3371ndash3379 2002
[34] S Mishra A Raz and L J Murphy ldquoInsulin-like growthfactor binding protein-3 interacts with autocrine motility fac-torphosphoglucose isomerase (AMFPGI) and inhibits theAMFPGI functionrdquo Cancer Research vol 64 no 7 pp 2516ndash2522 2004
[35] F L Yu M H Liao J W Lee and W L Shih ldquoInductionof hepatoma cells migration by phosphoglucose isomeraseautocrine motility factor through the upregulation of matrixmetalloproteinase-3rdquo Biochemical and Biophysical ResearchCommunications vol 314 no 1 pp 76ndash82 2004
[36] A H Klopp A Gupta E Spaeth M Andreeff and F MariniIII ldquoConcise review dissecting a discrepancy in the literaturedomesenchymal stem cells support or suppress tumor growthrdquoStem Cells vol 29 no 1 pp 11ndash19 2011
[37] L Qiao Z Xu T Zhao et al ldquoSuppression of tumorigenesisby human mesenchymal stem cells in a hepatoma modelrdquo CellResearch vol 18 no 4 pp 500ndash507 2008
[38] Y R Lu Y Yuan X J Wang et al ldquoThe growth inhibitory effectof mesenchymal stem cells on tumor cells in vitro and in vivordquoCancer Biology andTherapy vol 7 no 2 pp 245ndash251 2008
[39] Z-G Zhao W Xu L Sun W-M Li Q-P Li and P Zou ldquoThecharacteristics and immunoregulatory functions of regulatorydendritic cells induced by mesenchymal stem cells derivedfrom bone marrow of patient with chronic myeloid leukaemiardquoEuropean Journal of Cancer vol 48 no 12 pp 1884ndash1895 2012
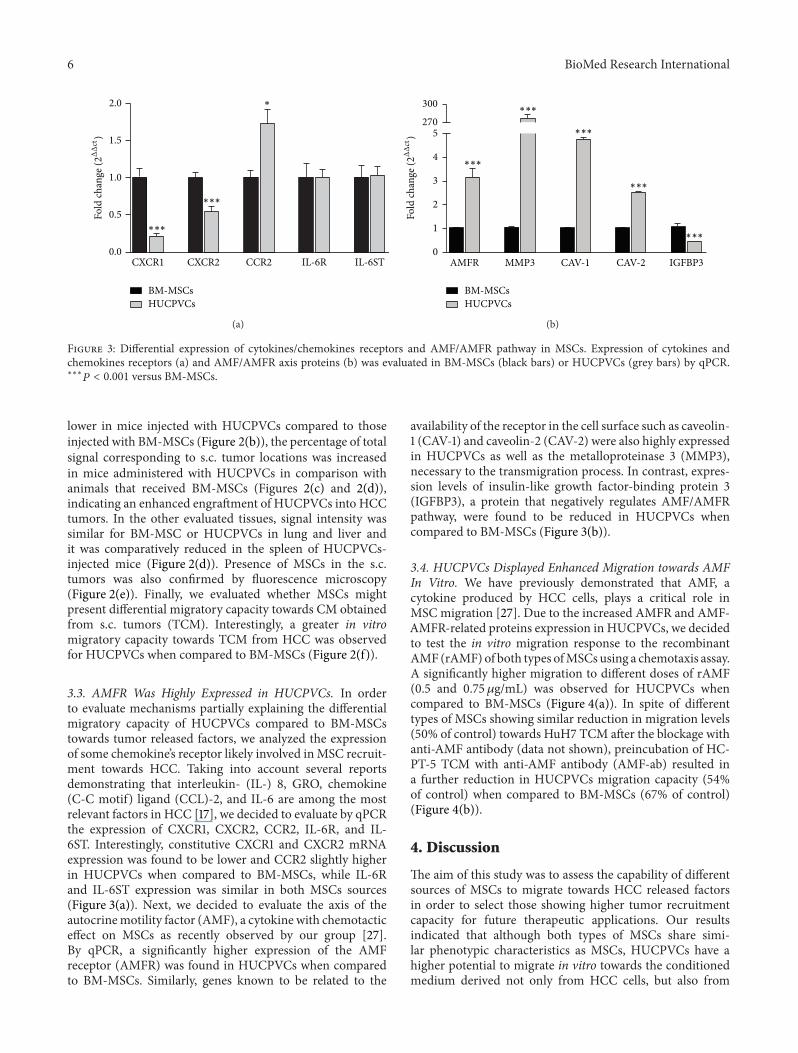
6 BioMed Research International
BM-MSCsHUCPVCs
lowastlowastlowast
lowastlowastlowast
lowast20
15
10
05
00
Fold
chan
ge (2
ΔΔ
ct)
CXCR1 CXCR2 CCR2 IL-6R IL-6ST
(a)
BM-MSCsHUCPVCs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
300
2705
4
3
2
1
0
Fold
chan
ge (2
ΔΔ
ct)
AMFR MMP3 CAV-1 CAV-2 IGFBP3
(b)
Figure 3 Differential expression of cytokineschemokines receptors and AMFAMFR pathway in MSCs Expression of cytokines andchemokines receptors (a) and AMFAMFR axis proteins (b) was evaluated in BM-MSCs (black bars) or HUCPVCs (grey bars) by qPCRlowastlowastlowast119875 lt 0001 versus BM-MSCs
lower in mice injected with HUCPVCs compared to thoseinjected with BM-MSCs (Figure 2(b)) the percentage of totalsignal corresponding to sc tumor locations was increasedin mice administered with HUCPVCs in comparison withanimals that received BM-MSCs (Figures 2(c) and 2(d))indicating an enhanced engraftment of HUCPVCs into HCCtumors In the other evaluated tissues signal intensity wassimilar for BM-MSC or HUCPVCs in lung and liver andit was comparatively reduced in the spleen of HUCPVCs-injected mice (Figure 2(d)) Presence of MSCs in the sctumors was also confirmed by fluorescence microscopy(Figure 2(e)) Finally we evaluated whether MSCs mightpresent differential migratory capacity towards CM obtainedfrom sc tumors (TCM) Interestingly a greater in vitromigratory capacity towards TCM from HCC was observedfor HUCPVCs when compared to BM-MSCs (Figure 2(f))
33 AMFR Was Highly Expressed in HUCPVCs In orderto evaluate mechanisms partially explaining the differentialmigratory capacity of HUCPVCs compared to BM-MSCstowards tumor released factors we analyzed the expressionof some chemokinersquos receptor likely involved inMSC recruit-ment towards HCC Taking into account several reportsdemonstrating that interleukin- (IL-) 8 GRO chemokine(C-C motif) ligand (CCL)-2 and IL-6 are among the mostrelevant factors in HCC [17] we decided to evaluate by qPCRthe expression of CXCR1 CXCR2 CCR2 IL-6R and IL-6ST Interestingly constitutive CXCR1 and CXCR2 mRNAexpression was found to be lower and CCR2 slightly higherin HUCPVCs when compared to BM-MSCs while IL-6Rand IL-6ST expression was similar in both MSCs sources(Figure 3(a)) Next we decided to evaluate the axis of theautocrinemotility factor (AMF) a cytokine with chemotacticeffect on MSCs as recently observed by our group [27]By qPCR a significantly higher expression of the AMFreceptor (AMFR) was found in HUCPVCs when comparedto BM-MSCs Similarly genes known to be related to the
availability of the receptor in the cell surface such as caveolin-1 (CAV-1) and caveolin-2 (CAV-2) were also highly expressedin HUCPVCs as well as the metalloproteinase 3 (MMP3)necessary to the transmigration process In contrast expres-sion levels of insulin-like growth factor-binding protein 3(IGFBP3) a protein that negatively regulates AMFAMFRpathway were found to be reduced in HUCPVCs whencompared to BM-MSCs (Figure 3(b))
34 HUCPVCs Displayed Enhanced Migration towards AMFIn Vitro We have previously demonstrated that AMF acytokine produced by HCC cells plays a critical role inMSC migration [27] Due to the increased AMFR and AMF-AMFR-related proteins expression in HUCPVCs we decidedto test the in vitro migration response to the recombinantAMF (rAMF) of both types ofMSCsusing a chemotaxis assayA significantly higher migration to different doses of rAMF(05 and 075 120583gmL) was observed for HUCPVCs whencompared to BM-MSCs (Figure 4(a)) In spite of differenttypes of MSCs showing similar reduction in migration levels(50 of control) towards HuH7 TCM after the blockage withanti-AMF antibody (data not shown) preincubation of HC-PT-5 TCM with anti-AMF antibody (AMF-ab) resulted ina further reduction in HUCPVCs migration capacity (54of control) when compared to BM-MSCs (67 of control)(Figure 4(b))
4 Discussion
The aim of this study was to assess the capability of differentsources of MSCs to migrate towards HCC released factorsin order to select those showing higher tumor recruitmentcapacity for future therapeutic applications Our resultsindicated that although both types of MSCs share simi-lar phenotypic characteristics as MSCs HUCPVCs have ahigher potential to migrate in vitro towards the conditionedmedium derived not only from HCC cells but also from
BioMed Research International 7
BM-MSCsHUCPVCs
lowastlowastlowast
0 050 075
rAMF (120583gmL)
300
250
200
150
100
50
0
Con
trol (
)
(a)
BM-MSCsHUCPVCs
lowast
Con
trol (
)
110
90
100
70
50
30
10
80
60
40
20
0+ minus + minus
+minus +minus
IgG-abAMF-ab
(b)
Figure 4 HUCPVCs showed enhanced migration towards AMF in comparison with BM-MSCs (a) In vitromigration of BM-MSCs (blackbars) or HUCPVCs (grey bars) towards rAMF 119875 lt 005 versus DMEM (0 120583gmL rAMF) lowast119875 lt 005 versus BM-MSCs (b) In vitromigrationof BM-MSCs (black bars) or HUCPVCs (grey bars) towards HC-PT-5 TCM preincubated with anti-AMF antibody (AMF-ab) or controlisotype (IgG-ab) was evaluated 119875 lt 005 versus IgG-ab lowast119875 lt 005 versus BM-MSCs Bars represent the average of MSCsfield (10x) plusmn SEMfrom three representative visual fields Results are representative of 3 independent experiments
nontumoral components of the tumor microenvironmentlike activated hepatic stellate cells (LX-2) fibroblasts (WI-38) or tumor microendothelial cells (HMEC-1) MoreoverHUCPVCs also showed an enhanced adhesion capacityto HMEC-1 cells compared to BM-MSCs This enhancedadhesion to endothelial cells could allow a more efficientarrest in tumor microvasculature which is a required step forMSC recruitment into HCC In line with this we observedthat HUCPVCs also exhibited greater migration capacity invivo towards experimental HCC In contrast to that observedin the in vitro migration experiments HUCPVCs showed asimilar migration capability towards nontumoral tissues likeliver or lung and also showed a lower recruitment to thespleen AlthoughHUCPVCsmigrated better than BM-MSCstoward activated hepatic stellate cells in vitro no differenceswere observed in in vivo migration to normal liver wherehepatic stellate cells are quiescent In addition it should benoted that mice injected with HUCPVCs showed a lowertotal signal than those injected with BM-MSCs probablydue to differences in the DiR uptake for both kinds of cellsor in the number of inoculated cells These results are inagreement with previous works showing that MCSs derivedfrom umbilical cord are able tomigrate towards experimentaltumors such as glioma [28] breast cancer [29] and lung ade-nocarcinoma [30] Nonetheless to the best of our knowledgethis is the first report describing the comparative migrationcapacity of human umbilical cord perivascular cells towardshuman HCC
Tumor-homing is thought to be mediated by interactionsof cytokineschemokines with their specific receptors Anumber of cytokines growth factors and chemokines aresecreted by HCC cells and their microenvironment [17] Wehave recently found that AMF is critical for MSC migrationtoward HCC in vitro and in vivo Furthermore other factorsreleased by tumor stroma components like IL-8 GRO IL-6or CCL-2 could also be involved inMSC recruitment toHCCThus differences observed between HUCPVCs and BM-MSCs in their in vitro and in vivomigration capacity towardsHCC seem at least partially related to differences in theexpression of cytokinechemokine receptors Some studieshave screened for similarities or differences among BM-MSCs and UC-MSCs at the molecular level [31 32] In addi-tion a previous report demonstrated that migration of UCB-MSC towards glioma cells was higher when compared toBM-MSCs likely due to increased levels of the IL-8 receptorCXCR1 and CXCR2 in the former cell type [18] In contrastto these observations we found that HUCPVCs presentedlower levels of CXCR1 and CXCR2 expression compared toBM-MSCs and these differences could be explained partiallydue to differences in tissue source and culture conditions andmerit further investigationThe analysis of IL-6R and IL6-STexpression showed similar levels in both types ofMSCs whilelevels of MCP-1 receptor (CCR2) were slightly increased inHUCPVCs Interestingly we found that not only AMFR butalso CAV-1 and CAV-2 which are genes that regulate theavailability of AMFR on cell surface [33] were overexpressedin HUCPVCs compared to BM-MSCs In line with this a
8 BioMed Research International
reduced expression level of IGFBP3 a protein which canbind to AMF blocking its binding to AMFR [34] was foundin HUCPVCs when compared to BM-MSCs Moreover anincreased expression of MMP3 was observed in HUCPVCsThis matrix metalloproteinase is involved in AMF-inducedmigration and it is necessary for cell invasion and proteolysisof the extracellularmatrix [35] Finally and consistentwith anincrease in the expression of genes related to the AMFAMFRpathway not only HUCPVCs show an enhanced in vitromigration towards rAMF but also the specific blockageof AMF in HC-PT-5 TCM was able to reduce HUCPVCsmigration to a greater level when compared to BM-MSCs
The potential use of MSCs in cancer treatment as carriersof therapeutic genes has raised some concerns about thesafety of their use in the clinic There are some studiesthat indicated that MSCs have protumorigenic capacitydue to their immunosuppression properties modulation ofthe epithelial-to-mesenchymal transition and induction ofangiogenesis [36] However others have demonstrated thatMSCs decreased HCC growth [37 38] Similarly we havepreviously demonstrated that BM-MSCs did not modifytumor growth in vivo [25] Moreover we observed thatHUCPVCs did not induce tumor growth when injected ivin Huh7 sc tumor-bearing mice (data not shown) Theuse of HUCPVCs as cellular carriers could also have theadvantage of its allogeneic application because of its lowimmunogenicity [9] In this regard HUCPVCs could beuseful as carriers of therapeutic genes for cancer patientswhere isolation of BM or adipose tissue-derived MSCs couldbe less responsive to chemotactic factors and might dis-play higher immunosuppression capacities than MSCs fromhealthy donors [39] or even present genetic abnormalities
5 Conclusions
In summary our results demonstrate a greater migrationcapacity to HCC of HUCPVCs when compared to BM-MSCs not only in vitro but also in vivo likely due to anincrease in migratory response to AMF and to an enhancedadhesive capacity to tumor microvasculature Consideringtheir availability and that no invasive procedures are requiredto obtain HUCPVCs these cells have advantages over BM-MSCs as candidates for carriers of therapeutic genes for thetreatment of HCC
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors gratefully thank Guillermo Gaston MarcosCabrera and Vanina Ferreira for technical assistance andDr Mengshiao Meng (National Chung Hsing University(NCHU) Taichung Taiwan) for providing recombinantAMF This work was supported by Grants from Austral
University (for Juan Bayo I04-12 for Mariana G Garcia T13-12 and for Guillermo Mazzolini T13-11) and from AgenciaNacional de Promocion Cientıfica y Tecnologica (ANPCyT)PICTO 200800115 (Mariana G Garcia) PICT 200800123(Jorge B Aquino) PICTO 200800122 (Jorge B Aquino)and PICT 20102818 (Mariana G Garcia and GuillermoMazzolini)
References
[1] H B El-Serag J A Marrero L Rudolph and K R ReddyldquoDiagnosis and treatment of hepatocellular carcinomardquo Gas-troenterology vol 134 no 6 pp 1752ndash1763 2008
[2] J M Llovet S Ricci V Mazzaferro et al ldquoSorafenib inadvanced hepatocellular carcinomardquo The New England Journalof Medicine vol 359 no 4 pp 378ndash390 2008
[3] A J Friedenstein J F Gorskaja andN N Kulagina ldquoFibroblastprecursors in normal and irradiated mouse hematopoieticorgansrdquo Experimental Hematology vol 4 no 5 pp 267ndash2741976
[4] D J Prockop and J Y Oh ldquoMedical therapies with adultstemprogenitor cells (MSCs) a backward journey from dra-matic results in vivo to the cellular andmolecular explanationsrdquoJournal of Cellular Biochemistry vol 113 no 5 pp 1460ndash14692012
[5] J B Aquino M F Bolontrade M G Garcıa O L Podhajcerand GMazzolini ldquoMesenchymal stem cells as therapeutic toolsand gene carriers in liver fibrosis and hepatocellular carcinomardquoGene Therapy vol 17 no 6 pp 692ndash708 2010
[6] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[7] H Yoshimura T Muneta A Nimura A Yokoyama H Kogaand I Sekiya ldquoComparison of rat mesenchymal stem cellsderived from bone marrow synovium periosteum adiposetissue and musclerdquo Cell and Tissue Research vol 327 no 3 pp449ndash462 2007
[8] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popu-lations and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[9] R Sarugaser D Lickorish D Baksh M M Hosseini and J EDavies ldquoHuman umbilical cord perivascular (HUCPV) cells asource of mesenchymal progenitorsrdquo Stem Cells vol 23 no 2pp 220ndash229 2005
[10] D Baksh R Yao and R S Tuan ldquoComparison of proliferativeand multilineage differentiation potential of human mesenchy-mal stem cells derived from umbilical cord and bone marrowrdquoStem Cells vol 25 no 6 pp 1384ndash1392 2007
[11] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[12] O K Lee T K Kuo W M Chen K D Lee S L Hsieh andT H Chen ldquoIsolation of multipotent mesenchymal stem cellsfrom umbilical cord bloodrdquo Blood vol 103 no 5 pp 1669ndash16752004
[13] HWang S Hung S Peng et al ldquoMesenchymal stem cells in theWhartonrsquos jelly of the human umbilical cordrdquo StemCells vol 22no 7 pp 1330ndash1337 2004
BioMed Research International 9
[14] J Kim J M Shin Y J Jeon H M Chung and J Chae ldquoPro-teomic validation of multifunctional molecules in mesenchy-mal stem cells derived from human bone marrow umbilicalcord blood and peripheral bloodrdquo PLoS ONE vol 7 no 5Article ID e32350 2012
[15] J-Y Hsieh H-W Wang S-J Chang et al ldquoMesenchymalstem cells from human umbilical cord express preferentiallysecreted factors related to neuroprotection neurogenesis andangiogenesisrdquo PLoS ONE vol 8 no 8 Article ID e72604 2013
[16] V Hernandez-Gea S Toffanin S L Friedman and J MLlovet ldquoRole of the microenvironment in the pathogenesis andtreatment of hepatocellular carcinomardquo Gastroenterology vol144 no 3 pp 512ndash527 2013
[17] J Bayo M Marrodan J B Aquino M Silva M G Garciaand G Mazzolini ldquoThe therapeutic potential of bone marrow-derived mesenchymal stromal cells on hepatocellular carci-nomardquo Liver International vol 34 no 3 pp 330ndash334 2014
[18] D Kim J H Kim J Kwon Lee et al ldquoOverexpression ofCXC chemokine receptors is required for the superior glioma-tracking property of umbilical cord blood-derived mesenchy-mal stem cellsrdquo Stem Cells and Development vol 18 no 3 pp511ndash519 2009
[19] S M Kim D Kim C H Jeong et al ldquoCXC chemokinereceptor 1 enhances the ability of human umbilical cord blood-derived mesenchymal stem cells to migrate toward gliomasrdquoBiochemical and Biophysical Research Communications vol 407no 4 pp 741ndash746 2011
[20] S A Park C H Ryu S M Kim et al ldquoCXCR4-transfectedhuman umbilical cord blood-derived mesenchymal stem cellsexhibit enhanced migratory capacity toward gliomasrdquo Interna-tional Journal of Oncology vol 38 no 1 pp 97ndash103 2011
[21] R S Rachakatla F Marini M L Weiss M Tamura and DTroyer ldquoDevelopment of human umbilical cord matrix stemcell-based gene therapy for experimental lung tumorsrdquo CancerGene Therapy vol 14 no 10 pp 828ndash835 2007
[22] C Yan S Li Z Li et al ldquoHuman umbilical cord mesenchymalstem cells as vehicles of CD20-specific TRAIL fusion proteindelivery a double-target therapy against non-Hodgkinrsquos lym-phomardquo Molecular Pharmaceutics vol 10 no 1 pp 142ndash1512013
[23] J Hu A Dong V Fernandez-Ruiz et al ldquoBlockade of Wntsignaling inhibits angiogenesis and tumor growth in hepatocel-lular carcinomardquo Cancer Research vol 69 no 17 pp 6951ndash69592009
[24] L Xu A Y Hui E Albanis et al ldquoHuman hepatic stellate celllines LX-1 and LX-2 new tools for analysis of hepatic fibrosisrdquoGut vol 54 no 1 pp 142ndash151 2005
[25] M G Garcia J Bayo M F Bolontrade et al ldquoHepatocellularcarcinoma cells and their fibrotic microenvironment modulatebone marrow-derived mesenchymal stromal cell migration invitro and in vivordquo Molecular Pharmaceutics vol 8 no 5 pp1538ndash1548 2011
[26] M Dominici K le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells theInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[27] J Bayo E Fiore J B Aquino et al ldquoIncreased migration ofhumanmesenchymal stromal cells byAutocrineMotility Factor(AMF) resulted in enhanced recruitment towards hepatocellu-lar carcinomardquo PLoS ONE vol 9 no 4 Article ID e95171 2014
[28] M K Seong Y L Jung I P Sang et al ldquoGene therapyusing TRAIL-secreting human umbilical cord blood-derived
mesenchymal stem cells against intracranial gliomardquo CancerResearch vol 68 no 23 pp 9614ndash9623 2008
[29] R Ayuzawa C Doi R S Rachakatla et al ldquoNaıve humanumbilical cord matrix derived stem cells significantly attenuategrowth of human breast cancer cells in vitro and in vivordquoCancerLetters vol 280 no 1 pp 31ndash37 2009
[30] T Matsuzuka R S Rachakatla C Doi et al ldquoHuman umbilicalcord matrix-derived stem cells expressing interferon-120573 genesignificantly attenuate bronchioloalveolar carcinomaxenograftsin SCID micerdquo Lung Cancer vol 70 no 1 pp 28ndash36 2010
[31] G Li X Zhang H Wang et al ldquoComparative proteomicanalysis of mesenchymal stem cells derived from human bonemarrow umbilical cord and placenta implication in themigra-tionrdquo Proteomics vol 9 no 1 pp 20ndash30 2009
[32] H C Miranda R H Herai C H Thome et al ldquoA quan-titative proteomic and transcriptomic comparison of humanmesenchymal stem cells from bone marrow and umbilical cordveinrdquo Proteomics vol 12 no 17 pp 2607ndash2617 2012
[33] P U Le G Guay Y Altschuler and I R Nabi ldquoCaveolin-1 isa negative regulator of caveolae-mediated endocytosis to theendoplasmic reticulumrdquo The Journal of Biological Chemistryvol 277 no 5 pp 3371ndash3379 2002
[34] S Mishra A Raz and L J Murphy ldquoInsulin-like growthfactor binding protein-3 interacts with autocrine motility fac-torphosphoglucose isomerase (AMFPGI) and inhibits theAMFPGI functionrdquo Cancer Research vol 64 no 7 pp 2516ndash2522 2004
[35] F L Yu M H Liao J W Lee and W L Shih ldquoInductionof hepatoma cells migration by phosphoglucose isomeraseautocrine motility factor through the upregulation of matrixmetalloproteinase-3rdquo Biochemical and Biophysical ResearchCommunications vol 314 no 1 pp 76ndash82 2004
[36] A H Klopp A Gupta E Spaeth M Andreeff and F MariniIII ldquoConcise review dissecting a discrepancy in the literaturedomesenchymal stem cells support or suppress tumor growthrdquoStem Cells vol 29 no 1 pp 11ndash19 2011
[37] L Qiao Z Xu T Zhao et al ldquoSuppression of tumorigenesisby human mesenchymal stem cells in a hepatoma modelrdquo CellResearch vol 18 no 4 pp 500ndash507 2008
[38] Y R Lu Y Yuan X J Wang et al ldquoThe growth inhibitory effectof mesenchymal stem cells on tumor cells in vitro and in vivordquoCancer Biology andTherapy vol 7 no 2 pp 245ndash251 2008
[39] Z-G Zhao W Xu L Sun W-M Li Q-P Li and P Zou ldquoThecharacteristics and immunoregulatory functions of regulatorydendritic cells induced by mesenchymal stem cells derivedfrom bone marrow of patient with chronic myeloid leukaemiardquoEuropean Journal of Cancer vol 48 no 12 pp 1884ndash1895 2012
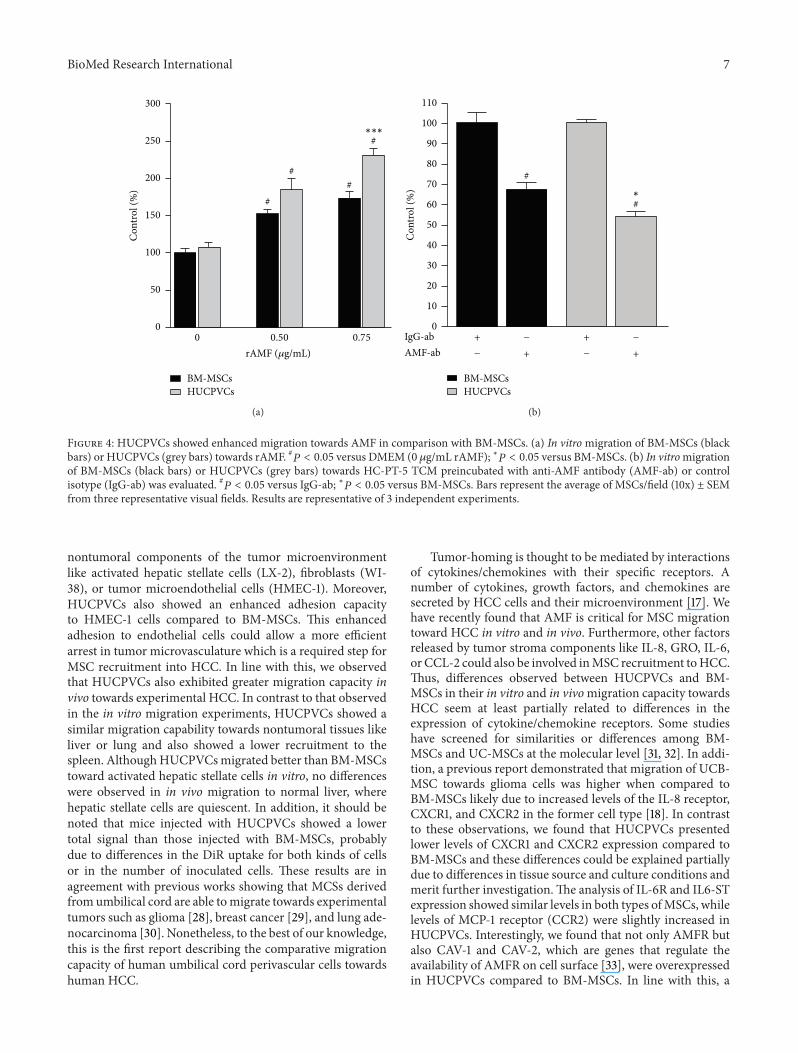
BioMed Research International 7
BM-MSCsHUCPVCs
lowastlowastlowast
0 050 075
rAMF (120583gmL)
300
250
200
150
100
50
0
Con
trol (
)
(a)
BM-MSCsHUCPVCs
lowast
Con
trol (
)
110
90
100
70
50
30
10
80
60
40
20
0+ minus + minus
+minus +minus
IgG-abAMF-ab
(b)
Figure 4 HUCPVCs showed enhanced migration towards AMF in comparison with BM-MSCs (a) In vitromigration of BM-MSCs (blackbars) or HUCPVCs (grey bars) towards rAMF 119875 lt 005 versus DMEM (0 120583gmL rAMF) lowast119875 lt 005 versus BM-MSCs (b) In vitromigrationof BM-MSCs (black bars) or HUCPVCs (grey bars) towards HC-PT-5 TCM preincubated with anti-AMF antibody (AMF-ab) or controlisotype (IgG-ab) was evaluated 119875 lt 005 versus IgG-ab lowast119875 lt 005 versus BM-MSCs Bars represent the average of MSCsfield (10x) plusmn SEMfrom three representative visual fields Results are representative of 3 independent experiments
nontumoral components of the tumor microenvironmentlike activated hepatic stellate cells (LX-2) fibroblasts (WI-38) or tumor microendothelial cells (HMEC-1) MoreoverHUCPVCs also showed an enhanced adhesion capacityto HMEC-1 cells compared to BM-MSCs This enhancedadhesion to endothelial cells could allow a more efficientarrest in tumor microvasculature which is a required step forMSC recruitment into HCC In line with this we observedthat HUCPVCs also exhibited greater migration capacity invivo towards experimental HCC In contrast to that observedin the in vitro migration experiments HUCPVCs showed asimilar migration capability towards nontumoral tissues likeliver or lung and also showed a lower recruitment to thespleen AlthoughHUCPVCsmigrated better than BM-MSCstoward activated hepatic stellate cells in vitro no differenceswere observed in in vivo migration to normal liver wherehepatic stellate cells are quiescent In addition it should benoted that mice injected with HUCPVCs showed a lowertotal signal than those injected with BM-MSCs probablydue to differences in the DiR uptake for both kinds of cellsor in the number of inoculated cells These results are inagreement with previous works showing that MCSs derivedfrom umbilical cord are able tomigrate towards experimentaltumors such as glioma [28] breast cancer [29] and lung ade-nocarcinoma [30] Nonetheless to the best of our knowledgethis is the first report describing the comparative migrationcapacity of human umbilical cord perivascular cells towardshuman HCC
Tumor-homing is thought to be mediated by interactionsof cytokineschemokines with their specific receptors Anumber of cytokines growth factors and chemokines aresecreted by HCC cells and their microenvironment [17] Wehave recently found that AMF is critical for MSC migrationtoward HCC in vitro and in vivo Furthermore other factorsreleased by tumor stroma components like IL-8 GRO IL-6or CCL-2 could also be involved inMSC recruitment toHCCThus differences observed between HUCPVCs and BM-MSCs in their in vitro and in vivomigration capacity towardsHCC seem at least partially related to differences in theexpression of cytokinechemokine receptors Some studieshave screened for similarities or differences among BM-MSCs and UC-MSCs at the molecular level [31 32] In addi-tion a previous report demonstrated that migration of UCB-MSC towards glioma cells was higher when compared toBM-MSCs likely due to increased levels of the IL-8 receptorCXCR1 and CXCR2 in the former cell type [18] In contrastto these observations we found that HUCPVCs presentedlower levels of CXCR1 and CXCR2 expression compared toBM-MSCs and these differences could be explained partiallydue to differences in tissue source and culture conditions andmerit further investigationThe analysis of IL-6R and IL6-STexpression showed similar levels in both types ofMSCs whilelevels of MCP-1 receptor (CCR2) were slightly increased inHUCPVCs Interestingly we found that not only AMFR butalso CAV-1 and CAV-2 which are genes that regulate theavailability of AMFR on cell surface [33] were overexpressedin HUCPVCs compared to BM-MSCs In line with this a
8 BioMed Research International
reduced expression level of IGFBP3 a protein which canbind to AMF blocking its binding to AMFR [34] was foundin HUCPVCs when compared to BM-MSCs Moreover anincreased expression of MMP3 was observed in HUCPVCsThis matrix metalloproteinase is involved in AMF-inducedmigration and it is necessary for cell invasion and proteolysisof the extracellularmatrix [35] Finally and consistentwith anincrease in the expression of genes related to the AMFAMFRpathway not only HUCPVCs show an enhanced in vitromigration towards rAMF but also the specific blockageof AMF in HC-PT-5 TCM was able to reduce HUCPVCsmigration to a greater level when compared to BM-MSCs
The potential use of MSCs in cancer treatment as carriersof therapeutic genes has raised some concerns about thesafety of their use in the clinic There are some studiesthat indicated that MSCs have protumorigenic capacitydue to their immunosuppression properties modulation ofthe epithelial-to-mesenchymal transition and induction ofangiogenesis [36] However others have demonstrated thatMSCs decreased HCC growth [37 38] Similarly we havepreviously demonstrated that BM-MSCs did not modifytumor growth in vivo [25] Moreover we observed thatHUCPVCs did not induce tumor growth when injected ivin Huh7 sc tumor-bearing mice (data not shown) Theuse of HUCPVCs as cellular carriers could also have theadvantage of its allogeneic application because of its lowimmunogenicity [9] In this regard HUCPVCs could beuseful as carriers of therapeutic genes for cancer patientswhere isolation of BM or adipose tissue-derived MSCs couldbe less responsive to chemotactic factors and might dis-play higher immunosuppression capacities than MSCs fromhealthy donors [39] or even present genetic abnormalities
5 Conclusions
In summary our results demonstrate a greater migrationcapacity to HCC of HUCPVCs when compared to BM-MSCs not only in vitro but also in vivo likely due to anincrease in migratory response to AMF and to an enhancedadhesive capacity to tumor microvasculature Consideringtheir availability and that no invasive procedures are requiredto obtain HUCPVCs these cells have advantages over BM-MSCs as candidates for carriers of therapeutic genes for thetreatment of HCC
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors gratefully thank Guillermo Gaston MarcosCabrera and Vanina Ferreira for technical assistance andDr Mengshiao Meng (National Chung Hsing University(NCHU) Taichung Taiwan) for providing recombinantAMF This work was supported by Grants from Austral
University (for Juan Bayo I04-12 for Mariana G Garcia T13-12 and for Guillermo Mazzolini T13-11) and from AgenciaNacional de Promocion Cientıfica y Tecnologica (ANPCyT)PICTO 200800115 (Mariana G Garcia) PICT 200800123(Jorge B Aquino) PICTO 200800122 (Jorge B Aquino)and PICT 20102818 (Mariana G Garcia and GuillermoMazzolini)
References
[1] H B El-Serag J A Marrero L Rudolph and K R ReddyldquoDiagnosis and treatment of hepatocellular carcinomardquo Gas-troenterology vol 134 no 6 pp 1752ndash1763 2008
[2] J M Llovet S Ricci V Mazzaferro et al ldquoSorafenib inadvanced hepatocellular carcinomardquo The New England Journalof Medicine vol 359 no 4 pp 378ndash390 2008
[3] A J Friedenstein J F Gorskaja andN N Kulagina ldquoFibroblastprecursors in normal and irradiated mouse hematopoieticorgansrdquo Experimental Hematology vol 4 no 5 pp 267ndash2741976
[4] D J Prockop and J Y Oh ldquoMedical therapies with adultstemprogenitor cells (MSCs) a backward journey from dra-matic results in vivo to the cellular andmolecular explanationsrdquoJournal of Cellular Biochemistry vol 113 no 5 pp 1460ndash14692012
[5] J B Aquino M F Bolontrade M G Garcıa O L Podhajcerand GMazzolini ldquoMesenchymal stem cells as therapeutic toolsand gene carriers in liver fibrosis and hepatocellular carcinomardquoGene Therapy vol 17 no 6 pp 692ndash708 2010
[6] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[7] H Yoshimura T Muneta A Nimura A Yokoyama H Kogaand I Sekiya ldquoComparison of rat mesenchymal stem cellsderived from bone marrow synovium periosteum adiposetissue and musclerdquo Cell and Tissue Research vol 327 no 3 pp449ndash462 2007
[8] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popu-lations and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[9] R Sarugaser D Lickorish D Baksh M M Hosseini and J EDavies ldquoHuman umbilical cord perivascular (HUCPV) cells asource of mesenchymal progenitorsrdquo Stem Cells vol 23 no 2pp 220ndash229 2005
[10] D Baksh R Yao and R S Tuan ldquoComparison of proliferativeand multilineage differentiation potential of human mesenchy-mal stem cells derived from umbilical cord and bone marrowrdquoStem Cells vol 25 no 6 pp 1384ndash1392 2007
[11] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[12] O K Lee T K Kuo W M Chen K D Lee S L Hsieh andT H Chen ldquoIsolation of multipotent mesenchymal stem cellsfrom umbilical cord bloodrdquo Blood vol 103 no 5 pp 1669ndash16752004
[13] HWang S Hung S Peng et al ldquoMesenchymal stem cells in theWhartonrsquos jelly of the human umbilical cordrdquo StemCells vol 22no 7 pp 1330ndash1337 2004
BioMed Research International 9
[14] J Kim J M Shin Y J Jeon H M Chung and J Chae ldquoPro-teomic validation of multifunctional molecules in mesenchy-mal stem cells derived from human bone marrow umbilicalcord blood and peripheral bloodrdquo PLoS ONE vol 7 no 5Article ID e32350 2012
[15] J-Y Hsieh H-W Wang S-J Chang et al ldquoMesenchymalstem cells from human umbilical cord express preferentiallysecreted factors related to neuroprotection neurogenesis andangiogenesisrdquo PLoS ONE vol 8 no 8 Article ID e72604 2013
[16] V Hernandez-Gea S Toffanin S L Friedman and J MLlovet ldquoRole of the microenvironment in the pathogenesis andtreatment of hepatocellular carcinomardquo Gastroenterology vol144 no 3 pp 512ndash527 2013
[17] J Bayo M Marrodan J B Aquino M Silva M G Garciaand G Mazzolini ldquoThe therapeutic potential of bone marrow-derived mesenchymal stromal cells on hepatocellular carci-nomardquo Liver International vol 34 no 3 pp 330ndash334 2014
[18] D Kim J H Kim J Kwon Lee et al ldquoOverexpression ofCXC chemokine receptors is required for the superior glioma-tracking property of umbilical cord blood-derived mesenchy-mal stem cellsrdquo Stem Cells and Development vol 18 no 3 pp511ndash519 2009
[19] S M Kim D Kim C H Jeong et al ldquoCXC chemokinereceptor 1 enhances the ability of human umbilical cord blood-derived mesenchymal stem cells to migrate toward gliomasrdquoBiochemical and Biophysical Research Communications vol 407no 4 pp 741ndash746 2011
[20] S A Park C H Ryu S M Kim et al ldquoCXCR4-transfectedhuman umbilical cord blood-derived mesenchymal stem cellsexhibit enhanced migratory capacity toward gliomasrdquo Interna-tional Journal of Oncology vol 38 no 1 pp 97ndash103 2011
[21] R S Rachakatla F Marini M L Weiss M Tamura and DTroyer ldquoDevelopment of human umbilical cord matrix stemcell-based gene therapy for experimental lung tumorsrdquo CancerGene Therapy vol 14 no 10 pp 828ndash835 2007
[22] C Yan S Li Z Li et al ldquoHuman umbilical cord mesenchymalstem cells as vehicles of CD20-specific TRAIL fusion proteindelivery a double-target therapy against non-Hodgkinrsquos lym-phomardquo Molecular Pharmaceutics vol 10 no 1 pp 142ndash1512013
[23] J Hu A Dong V Fernandez-Ruiz et al ldquoBlockade of Wntsignaling inhibits angiogenesis and tumor growth in hepatocel-lular carcinomardquo Cancer Research vol 69 no 17 pp 6951ndash69592009
[24] L Xu A Y Hui E Albanis et al ldquoHuman hepatic stellate celllines LX-1 and LX-2 new tools for analysis of hepatic fibrosisrdquoGut vol 54 no 1 pp 142ndash151 2005
[25] M G Garcia J Bayo M F Bolontrade et al ldquoHepatocellularcarcinoma cells and their fibrotic microenvironment modulatebone marrow-derived mesenchymal stromal cell migration invitro and in vivordquo Molecular Pharmaceutics vol 8 no 5 pp1538ndash1548 2011
[26] M Dominici K le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells theInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[27] J Bayo E Fiore J B Aquino et al ldquoIncreased migration ofhumanmesenchymal stromal cells byAutocrineMotility Factor(AMF) resulted in enhanced recruitment towards hepatocellu-lar carcinomardquo PLoS ONE vol 9 no 4 Article ID e95171 2014
[28] M K Seong Y L Jung I P Sang et al ldquoGene therapyusing TRAIL-secreting human umbilical cord blood-derived
mesenchymal stem cells against intracranial gliomardquo CancerResearch vol 68 no 23 pp 9614ndash9623 2008
[29] R Ayuzawa C Doi R S Rachakatla et al ldquoNaıve humanumbilical cord matrix derived stem cells significantly attenuategrowth of human breast cancer cells in vitro and in vivordquoCancerLetters vol 280 no 1 pp 31ndash37 2009
[30] T Matsuzuka R S Rachakatla C Doi et al ldquoHuman umbilicalcord matrix-derived stem cells expressing interferon-120573 genesignificantly attenuate bronchioloalveolar carcinomaxenograftsin SCID micerdquo Lung Cancer vol 70 no 1 pp 28ndash36 2010
[31] G Li X Zhang H Wang et al ldquoComparative proteomicanalysis of mesenchymal stem cells derived from human bonemarrow umbilical cord and placenta implication in themigra-tionrdquo Proteomics vol 9 no 1 pp 20ndash30 2009
[32] H C Miranda R H Herai C H Thome et al ldquoA quan-titative proteomic and transcriptomic comparison of humanmesenchymal stem cells from bone marrow and umbilical cordveinrdquo Proteomics vol 12 no 17 pp 2607ndash2617 2012
[33] P U Le G Guay Y Altschuler and I R Nabi ldquoCaveolin-1 isa negative regulator of caveolae-mediated endocytosis to theendoplasmic reticulumrdquo The Journal of Biological Chemistryvol 277 no 5 pp 3371ndash3379 2002
[34] S Mishra A Raz and L J Murphy ldquoInsulin-like growthfactor binding protein-3 interacts with autocrine motility fac-torphosphoglucose isomerase (AMFPGI) and inhibits theAMFPGI functionrdquo Cancer Research vol 64 no 7 pp 2516ndash2522 2004
[35] F L Yu M H Liao J W Lee and W L Shih ldquoInductionof hepatoma cells migration by phosphoglucose isomeraseautocrine motility factor through the upregulation of matrixmetalloproteinase-3rdquo Biochemical and Biophysical ResearchCommunications vol 314 no 1 pp 76ndash82 2004
[36] A H Klopp A Gupta E Spaeth M Andreeff and F MariniIII ldquoConcise review dissecting a discrepancy in the literaturedomesenchymal stem cells support or suppress tumor growthrdquoStem Cells vol 29 no 1 pp 11ndash19 2011
[37] L Qiao Z Xu T Zhao et al ldquoSuppression of tumorigenesisby human mesenchymal stem cells in a hepatoma modelrdquo CellResearch vol 18 no 4 pp 500ndash507 2008
[38] Y R Lu Y Yuan X J Wang et al ldquoThe growth inhibitory effectof mesenchymal stem cells on tumor cells in vitro and in vivordquoCancer Biology andTherapy vol 7 no 2 pp 245ndash251 2008
[39] Z-G Zhao W Xu L Sun W-M Li Q-P Li and P Zou ldquoThecharacteristics and immunoregulatory functions of regulatorydendritic cells induced by mesenchymal stem cells derivedfrom bone marrow of patient with chronic myeloid leukaemiardquoEuropean Journal of Cancer vol 48 no 12 pp 1884ndash1895 2012

8 BioMed Research International
reduced expression level of IGFBP3 a protein which canbind to AMF blocking its binding to AMFR [34] was foundin HUCPVCs when compared to BM-MSCs Moreover anincreased expression of MMP3 was observed in HUCPVCsThis matrix metalloproteinase is involved in AMF-inducedmigration and it is necessary for cell invasion and proteolysisof the extracellularmatrix [35] Finally and consistentwith anincrease in the expression of genes related to the AMFAMFRpathway not only HUCPVCs show an enhanced in vitromigration towards rAMF but also the specific blockageof AMF in HC-PT-5 TCM was able to reduce HUCPVCsmigration to a greater level when compared to BM-MSCs
The potential use of MSCs in cancer treatment as carriersof therapeutic genes has raised some concerns about thesafety of their use in the clinic There are some studiesthat indicated that MSCs have protumorigenic capacitydue to their immunosuppression properties modulation ofthe epithelial-to-mesenchymal transition and induction ofangiogenesis [36] However others have demonstrated thatMSCs decreased HCC growth [37 38] Similarly we havepreviously demonstrated that BM-MSCs did not modifytumor growth in vivo [25] Moreover we observed thatHUCPVCs did not induce tumor growth when injected ivin Huh7 sc tumor-bearing mice (data not shown) Theuse of HUCPVCs as cellular carriers could also have theadvantage of its allogeneic application because of its lowimmunogenicity [9] In this regard HUCPVCs could beuseful as carriers of therapeutic genes for cancer patientswhere isolation of BM or adipose tissue-derived MSCs couldbe less responsive to chemotactic factors and might dis-play higher immunosuppression capacities than MSCs fromhealthy donors [39] or even present genetic abnormalities
5 Conclusions
In summary our results demonstrate a greater migrationcapacity to HCC of HUCPVCs when compared to BM-MSCs not only in vitro but also in vivo likely due to anincrease in migratory response to AMF and to an enhancedadhesive capacity to tumor microvasculature Consideringtheir availability and that no invasive procedures are requiredto obtain HUCPVCs these cells have advantages over BM-MSCs as candidates for carriers of therapeutic genes for thetreatment of HCC
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors gratefully thank Guillermo Gaston MarcosCabrera and Vanina Ferreira for technical assistance andDr Mengshiao Meng (National Chung Hsing University(NCHU) Taichung Taiwan) for providing recombinantAMF This work was supported by Grants from Austral
University (for Juan Bayo I04-12 for Mariana G Garcia T13-12 and for Guillermo Mazzolini T13-11) and from AgenciaNacional de Promocion Cientıfica y Tecnologica (ANPCyT)PICTO 200800115 (Mariana G Garcia) PICT 200800123(Jorge B Aquino) PICTO 200800122 (Jorge B Aquino)and PICT 20102818 (Mariana G Garcia and GuillermoMazzolini)
References
[1] H B El-Serag J A Marrero L Rudolph and K R ReddyldquoDiagnosis and treatment of hepatocellular carcinomardquo Gas-troenterology vol 134 no 6 pp 1752ndash1763 2008
[2] J M Llovet S Ricci V Mazzaferro et al ldquoSorafenib inadvanced hepatocellular carcinomardquo The New England Journalof Medicine vol 359 no 4 pp 378ndash390 2008
[3] A J Friedenstein J F Gorskaja andN N Kulagina ldquoFibroblastprecursors in normal and irradiated mouse hematopoieticorgansrdquo Experimental Hematology vol 4 no 5 pp 267ndash2741976
[4] D J Prockop and J Y Oh ldquoMedical therapies with adultstemprogenitor cells (MSCs) a backward journey from dra-matic results in vivo to the cellular andmolecular explanationsrdquoJournal of Cellular Biochemistry vol 113 no 5 pp 1460ndash14692012
[5] J B Aquino M F Bolontrade M G Garcıa O L Podhajcerand GMazzolini ldquoMesenchymal stem cells as therapeutic toolsand gene carriers in liver fibrosis and hepatocellular carcinomardquoGene Therapy vol 17 no 6 pp 692ndash708 2010
[6] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[7] H Yoshimura T Muneta A Nimura A Yokoyama H Kogaand I Sekiya ldquoComparison of rat mesenchymal stem cellsderived from bone marrow synovium periosteum adiposetissue and musclerdquo Cell and Tissue Research vol 327 no 3 pp449ndash462 2007
[8] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popu-lations and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[9] R Sarugaser D Lickorish D Baksh M M Hosseini and J EDavies ldquoHuman umbilical cord perivascular (HUCPV) cells asource of mesenchymal progenitorsrdquo Stem Cells vol 23 no 2pp 220ndash229 2005
[10] D Baksh R Yao and R S Tuan ldquoComparison of proliferativeand multilineage differentiation potential of human mesenchy-mal stem cells derived from umbilical cord and bone marrowrdquoStem Cells vol 25 no 6 pp 1384ndash1392 2007
[11] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[12] O K Lee T K Kuo W M Chen K D Lee S L Hsieh andT H Chen ldquoIsolation of multipotent mesenchymal stem cellsfrom umbilical cord bloodrdquo Blood vol 103 no 5 pp 1669ndash16752004
[13] HWang S Hung S Peng et al ldquoMesenchymal stem cells in theWhartonrsquos jelly of the human umbilical cordrdquo StemCells vol 22no 7 pp 1330ndash1337 2004
BioMed Research International 9
[14] J Kim J M Shin Y J Jeon H M Chung and J Chae ldquoPro-teomic validation of multifunctional molecules in mesenchy-mal stem cells derived from human bone marrow umbilicalcord blood and peripheral bloodrdquo PLoS ONE vol 7 no 5Article ID e32350 2012
[15] J-Y Hsieh H-W Wang S-J Chang et al ldquoMesenchymalstem cells from human umbilical cord express preferentiallysecreted factors related to neuroprotection neurogenesis andangiogenesisrdquo PLoS ONE vol 8 no 8 Article ID e72604 2013
[16] V Hernandez-Gea S Toffanin S L Friedman and J MLlovet ldquoRole of the microenvironment in the pathogenesis andtreatment of hepatocellular carcinomardquo Gastroenterology vol144 no 3 pp 512ndash527 2013
[17] J Bayo M Marrodan J B Aquino M Silva M G Garciaand G Mazzolini ldquoThe therapeutic potential of bone marrow-derived mesenchymal stromal cells on hepatocellular carci-nomardquo Liver International vol 34 no 3 pp 330ndash334 2014
[18] D Kim J H Kim J Kwon Lee et al ldquoOverexpression ofCXC chemokine receptors is required for the superior glioma-tracking property of umbilical cord blood-derived mesenchy-mal stem cellsrdquo Stem Cells and Development vol 18 no 3 pp511ndash519 2009
[19] S M Kim D Kim C H Jeong et al ldquoCXC chemokinereceptor 1 enhances the ability of human umbilical cord blood-derived mesenchymal stem cells to migrate toward gliomasrdquoBiochemical and Biophysical Research Communications vol 407no 4 pp 741ndash746 2011
[20] S A Park C H Ryu S M Kim et al ldquoCXCR4-transfectedhuman umbilical cord blood-derived mesenchymal stem cellsexhibit enhanced migratory capacity toward gliomasrdquo Interna-tional Journal of Oncology vol 38 no 1 pp 97ndash103 2011
[21] R S Rachakatla F Marini M L Weiss M Tamura and DTroyer ldquoDevelopment of human umbilical cord matrix stemcell-based gene therapy for experimental lung tumorsrdquo CancerGene Therapy vol 14 no 10 pp 828ndash835 2007
[22] C Yan S Li Z Li et al ldquoHuman umbilical cord mesenchymalstem cells as vehicles of CD20-specific TRAIL fusion proteindelivery a double-target therapy against non-Hodgkinrsquos lym-phomardquo Molecular Pharmaceutics vol 10 no 1 pp 142ndash1512013
[23] J Hu A Dong V Fernandez-Ruiz et al ldquoBlockade of Wntsignaling inhibits angiogenesis and tumor growth in hepatocel-lular carcinomardquo Cancer Research vol 69 no 17 pp 6951ndash69592009
[24] L Xu A Y Hui E Albanis et al ldquoHuman hepatic stellate celllines LX-1 and LX-2 new tools for analysis of hepatic fibrosisrdquoGut vol 54 no 1 pp 142ndash151 2005
[25] M G Garcia J Bayo M F Bolontrade et al ldquoHepatocellularcarcinoma cells and their fibrotic microenvironment modulatebone marrow-derived mesenchymal stromal cell migration invitro and in vivordquo Molecular Pharmaceutics vol 8 no 5 pp1538ndash1548 2011
[26] M Dominici K le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells theInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[27] J Bayo E Fiore J B Aquino et al ldquoIncreased migration ofhumanmesenchymal stromal cells byAutocrineMotility Factor(AMF) resulted in enhanced recruitment towards hepatocellu-lar carcinomardquo PLoS ONE vol 9 no 4 Article ID e95171 2014
[28] M K Seong Y L Jung I P Sang et al ldquoGene therapyusing TRAIL-secreting human umbilical cord blood-derived
mesenchymal stem cells against intracranial gliomardquo CancerResearch vol 68 no 23 pp 9614ndash9623 2008
[29] R Ayuzawa C Doi R S Rachakatla et al ldquoNaıve humanumbilical cord matrix derived stem cells significantly attenuategrowth of human breast cancer cells in vitro and in vivordquoCancerLetters vol 280 no 1 pp 31ndash37 2009
[30] T Matsuzuka R S Rachakatla C Doi et al ldquoHuman umbilicalcord matrix-derived stem cells expressing interferon-120573 genesignificantly attenuate bronchioloalveolar carcinomaxenograftsin SCID micerdquo Lung Cancer vol 70 no 1 pp 28ndash36 2010
[31] G Li X Zhang H Wang et al ldquoComparative proteomicanalysis of mesenchymal stem cells derived from human bonemarrow umbilical cord and placenta implication in themigra-tionrdquo Proteomics vol 9 no 1 pp 20ndash30 2009
[32] H C Miranda R H Herai C H Thome et al ldquoA quan-titative proteomic and transcriptomic comparison of humanmesenchymal stem cells from bone marrow and umbilical cordveinrdquo Proteomics vol 12 no 17 pp 2607ndash2617 2012
[33] P U Le G Guay Y Altschuler and I R Nabi ldquoCaveolin-1 isa negative regulator of caveolae-mediated endocytosis to theendoplasmic reticulumrdquo The Journal of Biological Chemistryvol 277 no 5 pp 3371ndash3379 2002
[34] S Mishra A Raz and L J Murphy ldquoInsulin-like growthfactor binding protein-3 interacts with autocrine motility fac-torphosphoglucose isomerase (AMFPGI) and inhibits theAMFPGI functionrdquo Cancer Research vol 64 no 7 pp 2516ndash2522 2004
[35] F L Yu M H Liao J W Lee and W L Shih ldquoInductionof hepatoma cells migration by phosphoglucose isomeraseautocrine motility factor through the upregulation of matrixmetalloproteinase-3rdquo Biochemical and Biophysical ResearchCommunications vol 314 no 1 pp 76ndash82 2004
[36] A H Klopp A Gupta E Spaeth M Andreeff and F MariniIII ldquoConcise review dissecting a discrepancy in the literaturedomesenchymal stem cells support or suppress tumor growthrdquoStem Cells vol 29 no 1 pp 11ndash19 2011
[37] L Qiao Z Xu T Zhao et al ldquoSuppression of tumorigenesisby human mesenchymal stem cells in a hepatoma modelrdquo CellResearch vol 18 no 4 pp 500ndash507 2008
[38] Y R Lu Y Yuan X J Wang et al ldquoThe growth inhibitory effectof mesenchymal stem cells on tumor cells in vitro and in vivordquoCancer Biology andTherapy vol 7 no 2 pp 245ndash251 2008
[39] Z-G Zhao W Xu L Sun W-M Li Q-P Li and P Zou ldquoThecharacteristics and immunoregulatory functions of regulatorydendritic cells induced by mesenchymal stem cells derivedfrom bone marrow of patient with chronic myeloid leukaemiardquoEuropean Journal of Cancer vol 48 no 12 pp 1884ndash1895 2012

BioMed Research International 9
[14] J Kim J M Shin Y J Jeon H M Chung and J Chae ldquoPro-teomic validation of multifunctional molecules in mesenchy-mal stem cells derived from human bone marrow umbilicalcord blood and peripheral bloodrdquo PLoS ONE vol 7 no 5Article ID e32350 2012
[15] J-Y Hsieh H-W Wang S-J Chang et al ldquoMesenchymalstem cells from human umbilical cord express preferentiallysecreted factors related to neuroprotection neurogenesis andangiogenesisrdquo PLoS ONE vol 8 no 8 Article ID e72604 2013
[16] V Hernandez-Gea S Toffanin S L Friedman and J MLlovet ldquoRole of the microenvironment in the pathogenesis andtreatment of hepatocellular carcinomardquo Gastroenterology vol144 no 3 pp 512ndash527 2013
[17] J Bayo M Marrodan J B Aquino M Silva M G Garciaand G Mazzolini ldquoThe therapeutic potential of bone marrow-derived mesenchymal stromal cells on hepatocellular carci-nomardquo Liver International vol 34 no 3 pp 330ndash334 2014
[18] D Kim J H Kim J Kwon Lee et al ldquoOverexpression ofCXC chemokine receptors is required for the superior glioma-tracking property of umbilical cord blood-derived mesenchy-mal stem cellsrdquo Stem Cells and Development vol 18 no 3 pp511ndash519 2009
[19] S M Kim D Kim C H Jeong et al ldquoCXC chemokinereceptor 1 enhances the ability of human umbilical cord blood-derived mesenchymal stem cells to migrate toward gliomasrdquoBiochemical and Biophysical Research Communications vol 407no 4 pp 741ndash746 2011
[20] S A Park C H Ryu S M Kim et al ldquoCXCR4-transfectedhuman umbilical cord blood-derived mesenchymal stem cellsexhibit enhanced migratory capacity toward gliomasrdquo Interna-tional Journal of Oncology vol 38 no 1 pp 97ndash103 2011
[21] R S Rachakatla F Marini M L Weiss M Tamura and DTroyer ldquoDevelopment of human umbilical cord matrix stemcell-based gene therapy for experimental lung tumorsrdquo CancerGene Therapy vol 14 no 10 pp 828ndash835 2007
[22] C Yan S Li Z Li et al ldquoHuman umbilical cord mesenchymalstem cells as vehicles of CD20-specific TRAIL fusion proteindelivery a double-target therapy against non-Hodgkinrsquos lym-phomardquo Molecular Pharmaceutics vol 10 no 1 pp 142ndash1512013
[23] J Hu A Dong V Fernandez-Ruiz et al ldquoBlockade of Wntsignaling inhibits angiogenesis and tumor growth in hepatocel-lular carcinomardquo Cancer Research vol 69 no 17 pp 6951ndash69592009
[24] L Xu A Y Hui E Albanis et al ldquoHuman hepatic stellate celllines LX-1 and LX-2 new tools for analysis of hepatic fibrosisrdquoGut vol 54 no 1 pp 142ndash151 2005
[25] M G Garcia J Bayo M F Bolontrade et al ldquoHepatocellularcarcinoma cells and their fibrotic microenvironment modulatebone marrow-derived mesenchymal stromal cell migration invitro and in vivordquo Molecular Pharmaceutics vol 8 no 5 pp1538ndash1548 2011
[26] M Dominici K le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells theInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[27] J Bayo E Fiore J B Aquino et al ldquoIncreased migration ofhumanmesenchymal stromal cells byAutocrineMotility Factor(AMF) resulted in enhanced recruitment towards hepatocellu-lar carcinomardquo PLoS ONE vol 9 no 4 Article ID e95171 2014
[28] M K Seong Y L Jung I P Sang et al ldquoGene therapyusing TRAIL-secreting human umbilical cord blood-derived
mesenchymal stem cells against intracranial gliomardquo CancerResearch vol 68 no 23 pp 9614ndash9623 2008
[29] R Ayuzawa C Doi R S Rachakatla et al ldquoNaıve humanumbilical cord matrix derived stem cells significantly attenuategrowth of human breast cancer cells in vitro and in vivordquoCancerLetters vol 280 no 1 pp 31ndash37 2009
[30] T Matsuzuka R S Rachakatla C Doi et al ldquoHuman umbilicalcord matrix-derived stem cells expressing interferon-120573 genesignificantly attenuate bronchioloalveolar carcinomaxenograftsin SCID micerdquo Lung Cancer vol 70 no 1 pp 28ndash36 2010
[31] G Li X Zhang H Wang et al ldquoComparative proteomicanalysis of mesenchymal stem cells derived from human bonemarrow umbilical cord and placenta implication in themigra-tionrdquo Proteomics vol 9 no 1 pp 20ndash30 2009
[32] H C Miranda R H Herai C H Thome et al ldquoA quan-titative proteomic and transcriptomic comparison of humanmesenchymal stem cells from bone marrow and umbilical cordveinrdquo Proteomics vol 12 no 17 pp 2607ndash2617 2012
[33] P U Le G Guay Y Altschuler and I R Nabi ldquoCaveolin-1 isa negative regulator of caveolae-mediated endocytosis to theendoplasmic reticulumrdquo The Journal of Biological Chemistryvol 277 no 5 pp 3371ndash3379 2002
[34] S Mishra A Raz and L J Murphy ldquoInsulin-like growthfactor binding protein-3 interacts with autocrine motility fac-torphosphoglucose isomerase (AMFPGI) and inhibits theAMFPGI functionrdquo Cancer Research vol 64 no 7 pp 2516ndash2522 2004
[35] F L Yu M H Liao J W Lee and W L Shih ldquoInductionof hepatoma cells migration by phosphoglucose isomeraseautocrine motility factor through the upregulation of matrixmetalloproteinase-3rdquo Biochemical and Biophysical ResearchCommunications vol 314 no 1 pp 76ndash82 2004
[36] A H Klopp A Gupta E Spaeth M Andreeff and F MariniIII ldquoConcise review dissecting a discrepancy in the literaturedomesenchymal stem cells support or suppress tumor growthrdquoStem Cells vol 29 no 1 pp 11ndash19 2011
[37] L Qiao Z Xu T Zhao et al ldquoSuppression of tumorigenesisby human mesenchymal stem cells in a hepatoma modelrdquo CellResearch vol 18 no 4 pp 500ndash507 2008
[38] Y R Lu Y Yuan X J Wang et al ldquoThe growth inhibitory effectof mesenchymal stem cells on tumor cells in vitro and in vivordquoCancer Biology andTherapy vol 7 no 2 pp 245ndash251 2008
[39] Z-G Zhao W Xu L Sun W-M Li Q-P Li and P Zou ldquoThecharacteristics and immunoregulatory functions of regulatorydendritic cells induced by mesenchymal stem cells derivedfrom bone marrow of patient with chronic myeloid leukaemiardquoEuropean Journal of Cancer vol 48 no 12 pp 1884ndash1895 2012
Related Documents











