Human TRIM71 and Its Nematode Homologue Are Targets of let-7 MicroRNA and Its Zebrafish Orthologue Is Essential for Development You-Chin Lin,* 1 Li-Ching Hsieh,* 1 Ming-Wei Kuo,à John Yu,*à Huan-Hsien Kuo,* Wan-Lin Lo,* Ruey-Jen Lin,* Alice L. Yu,*§ 1 and Wen-Hsiung Li* 1 k *Genomics Research Center, Academia Sinica, Taipei, Taiwan; Institute of Information Science, Academia Sinica, Taipei, Taiwan; àInstitute of Cellular and Organismic Biology, Academia Sinica, Taipei, Taiwan; §Department of Pediatrics Hematology-Oncology, University of California, San Diego; and kDepartment of Ecology and Evolution, University of Chicago Animal microRNAs (miRNAs) are short RNAs that function as posttranscriptional regulators of gene expression by binding to the target mRNAs. Noting that some miRNAs are highly conserved in evolution, we explored the possibility of evolutionary conservation of their targets. We identified human orthologues of experimentally verified let-7 miRNA target genes in Caenorhabditis elegans and used the luciferase reporter system to examine whether these human genes are still the targets of let-7 miRNA. We found that in some cases, the miRNA–target relationship has indeed been conserved in human. Interestingly, human TRIM71, an orthologue of C. elegans let-7–target lin-41 gene, can be repressed by hsa-let-7a and hsa-let-7c. This repression was abolished when both predicted let-7 target sites of TRIM71 were mutated. Moreover, the zebrafish lin-41 orthologue was also repressed by let-7 to a similar degree as was TRIM71. When the expression of zebrafish lin-41 orthologue was silenced by microinjection of RNA interference or morpholino into zebrafish zygotes, retarded embryonic development was observed, providing direct evidence for an essential role of lin-41 in zebrafish development. Taken together, our results suggest that the regulation of TRIM71 expression by let-7 has been evolutionarily conserved and that TRIM71 likely plays an important role in development. Introduction MicroRNAs (miRNAs) are endogenous noncoding RNA molecules of ;22 nt that negatively regulate the ex- pression of target genes in animals by binding to the 3# un- translated regions (UTRs) of the target mRNAs. The binding generally includes strong base pairing between the 5# end region of an miRNA and the complementary se- quence of its target, although extensive base pairing in the 3# end of the miRNA may compensate for insufficient base pairing of the 5# end (reviewed in Carrington and Ambros 2003; Lai 2003; Ambros 2004; Bartel 2004; Kim and Nam 2006; Niwa and Slack 2007). The importance of miRNAs in animals is underscored by the facts that they are esti- mated to comprise 1–5% of animal genes (Altschul et al. 1990; Lim et al. 2003), appear to regulate a large proportion of protein-coding genes (Brennecke et al. 2005; Krek et al. 2005; Lewis et al. 2005; Lim et al. 2005), and are involved in the control of a variety of physiological processes includ- ing development, cell proliferation, apoptosis, tissue differ- entiation, and metabolism (Johnston et al. 2001; Lim et al. 2003; Ambros 2004; Plasterk 2006; Valencia-Sanchez et al. 2006). A number of miRNAs are evolutionarily highly con- served (Pasquinelli et al. 2000; Johnston et al. 2001; Pasquinelli et al. 2003; Ambros 2004; Valencia-Sanchez et al. 2006). For example, the let-7 miRNA gene, which was originally identified as an important regulator involved in the heterochronic pathway controlling developmental timing in Caenorhabditis elegans (Reinhart et al. 2000), is conserved from nematodes to primates. Supplementary figure S1 (Supplementary Material online) shows the se- quence alignment of the elements of the let-7 family found in C. elegans, zebra fish, and human. Because each miRNA regulates multiple genes, the binding regions of miRNAs are under tight structural constraints to avoid losing the reg- ulation. This might have contributed to the high conserva- tion of binding regions of these miRNAs across species. In contrast, the binding sites of their targets would not be un- der such strong constraints and might have more freedom to mutate without affecting many other genes. Moreover, gene duplication and alternative splicing are 2 important sources of gene function diversity and are prevalent in complex or- ganisms (Mironov et al. 1999; Li et al. 2001; Kan et al. 2002; Waterston et al. 2002). Duplicate genes may diverge in their 3# UTRs, and alternatively spliced transcripts of a gene may give rise to different 3# UTRs. These 2 mech- anisms may accelerate the evolution of the target sites on 3# UTRs, making the miRNA–target interactions more complex. In this study, we are interested in the evolution of the target genes of highly conserved miRNAs. We selected all experimentally verified miRNA–target pairs of C. elegans that have orthologues for both miRNA and target genes in human and used the luciferase reporter system to examine whether the human orthologues and their cognate miRNAs remain interacting pairs. In addition, if any of the C. elegans target genes have multiple orthologous copies in human, we also examined these paralogous copies with their cognate miRNAs. For this purpose, we focused on the 4 experimen- tally verified C. elegans let-7–target pairs that have orthol- ogous pairs in human. For the pair of RAS and let-7, its orthologous pairs have been shown to function in human (Johnson et al. 2005). Our result showed that in the remain- ing 3 pairs, only the orthologue TRIM71 (human orthologue of lin-41) is still the target of let-7 in human. Thus, all to- gether, 2 of the 4 let-7–target pairs studied have been con- served, suggesting that the interaction relationship of a miRNA–target pair can be conserved during a very long period of evolution. Because the function of TRIM71 has not been documented yet, we silenced the lin-41 orthologue in a zebrafish model and demonstrated developmental de- fects similar to the effects of let-7 oligo injection in zebra- fish embryos reported previously (Kloosterman et al. 2004). 1 Equal contribution to this work. Key words: TRIM71, lin-41, let-7, microRNA target. E-mail: [email protected]. Mol. Biol. Evol. 24(11):2525–2534. 2007 doi:10.1093/molbev/msm195 Advance Access publication September 21, 2007 Ó The Author 2007. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution. All rights reserved. For permissions, please e-mail: [email protected] by guest on June 2, 2013 http://mbe.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Human TRIM71 and Its Nematode Homologue Are Targets of let-7 MicroRNAand Its Zebrafish Orthologue Is Essential for Development
You-Chin Lin,*1 Li-Ching Hsieh,*�1 Ming-Wei Kuo,� John Yu,*� Huan-Hsien Kuo,* Wan-Lin Lo,*Ruey-Jen Lin,* Alice L. Yu,*§1 and Wen-Hsiung Li*1k*Genomics Research Center, Academia Sinica, Taipei, Taiwan; �Institute of Information Science, Academia Sinica, Taipei, Taiwan;�Institute of Cellular and Organismic Biology, Academia Sinica, Taipei, Taiwan; §Department of Pediatrics Hematology-Oncology,University of California, San Diego; and kDepartment of Ecology and Evolution, University of Chicago
Animal microRNAs (miRNAs) are short RNAs that function as posttranscriptional regulators of gene expression bybinding to the target mRNAs. Noting that some miRNAs are highly conserved in evolution, we explored the possibilityof evolutionary conservation of their targets. We identified human orthologues of experimentally verified let-7 miRNAtarget genes in Caenorhabditis elegans and used the luciferase reporter system to examine whether these human genesare still the targets of let-7 miRNA. We found that in some cases, the miRNA–target relationship has indeed beenconserved in human. Interestingly, human TRIM71, an orthologue of C. elegans let-7–target lin-41 gene, can berepressed by hsa-let-7a and hsa-let-7c. This repression was abolished when both predicted let-7 target sites of TRIM71were mutated. Moreover, the zebrafish lin-41 orthologue was also repressed by let-7 to a similar degree as was TRIM71.When the expression of zebrafish lin-41 orthologue was silenced by microinjection of RNA interference or morpholinointo zebrafish zygotes, retarded embryonic development was observed, providing direct evidence for an essential role oflin-41 in zebrafish development. Taken together, our results suggest that the regulation of TRIM71 expression by let-7has been evolutionarily conserved and that TRIM71 likely plays an important role in development.
Introduction
MicroRNAs (miRNAs) are endogenous noncodingRNA molecules of ;22 nt that negatively regulate the ex-pression of target genes in animals by binding to the 3# un-translated regions (UTRs) of the target mRNAs. Thebinding generally includes strong base pairing betweenthe 5# end region of an miRNA and the complementary se-quence of its target, although extensive base pairing in the3# end of the miRNA may compensate for insufficient basepairing of the 5# end (reviewed in Carrington and Ambros2003; Lai 2003; Ambros 2004; Bartel 2004; Kim and Nam2006; Niwa and Slack 2007). The importance of miRNAsin animals is underscored by the facts that they are esti-mated to comprise 1–5% of animal genes (Altschul et al.1990; Lim et al. 2003), appear to regulate a large proportionof protein-coding genes (Brennecke et al. 2005; Krek et al.2005; Lewis et al. 2005; Lim et al. 2005), and are involvedin the control of a variety of physiological processes includ-ing development, cell proliferation, apoptosis, tissue differ-entiation, and metabolism (Johnston et al. 2001; Lim et al.2003; Ambros 2004; Plasterk 2006; Valencia-Sanchez et al.2006).
A number of miRNAs are evolutionarily highly con-served (Pasquinelli et al. 2000; Johnston et al. 2001;Pasquinelli et al. 2003; Ambros 2004; Valencia-Sanchezet al. 2006). For example, the let-7 miRNA gene, whichwas originally identified as an important regulator involvedin the heterochronic pathway controlling developmentaltiming in Caenorhabditis elegans (Reinhart et al. 2000),is conserved from nematodes to primates. Supplementaryfigure S1 (Supplementary Material online) shows the se-quence alignment of the elements of the let-7 family foundin C. elegans, zebra fish, and human. Because each miRNA
regulates multiple genes, the binding regions of miRNAsare under tight structural constraints to avoid losing the reg-ulation. This might have contributed to the high conserva-tion of binding regions of these miRNAs across species. Incontrast, the binding sites of their targets would not be un-der such strong constraints and might have more freedom tomutate without affecting many other genes. Moreover, geneduplication and alternative splicing are 2 important sourcesof gene function diversity and are prevalent in complex or-ganisms (Mironov et al. 1999; Li et al. 2001; Kan et al.2002; Waterston et al. 2002). Duplicate genes may divergein their 3# UTRs, and alternatively spliced transcripts ofa gene may give rise to different 3# UTRs. These 2 mech-anisms may accelerate the evolution of the target sites on3# UTRs, making the miRNA–target interactions morecomplex.
In this study, we are interested in the evolution of thetarget genes of highly conserved miRNAs. We selected allexperimentally verified miRNA–target pairs of C. elegansthat have orthologues for both miRNA and target genes inhuman and used the luciferase reporter system to examinewhether the human orthologues and their cognate miRNAsremain interacting pairs. In addition, if any of the C. eleganstarget genes have multiple orthologous copies in human, wealso examined these paralogous copies with their cognatemiRNAs. For this purpose, we focused on the 4 experimen-tally verified C. elegans let-7–target pairs that have orthol-ogous pairs in human. For the pair of RAS and let-7, itsorthologous pairs have been shown to function in human(Johnson et al. 2005). Our result showed that in the remain-ing 3 pairs, only the orthologue TRIM71 (human orthologueof lin-41) is still the target of let-7 in human. Thus, all to-gether, 2 of the 4 let-7–target pairs studied have been con-served, suggesting that the interaction relationship ofa miRNA–target pair can be conserved during a very longperiod of evolution. Because the function of TRIM71 hasnot been documented yet, we silenced the lin-41 orthologuein a zebrafish model and demonstrated developmental de-fects similar to the effects of let-7 oligo injection in zebra-fish embryos reported previously (Kloosterman et al. 2004).
1 Equal contribution to this work.
Key words: TRIM71, lin-41, let-7, microRNA target.
E-mail: [email protected].
Mol. Biol. Evol. 24(11):2525–2534. 2007doi:10.1093/molbev/msm195Advance Access publication September 21, 2007
� The Author 2007. Published by Oxford University Press on behalf ofthe Society for Molecular Biology and Evolution. All rights reserved.For permissions, please e-mail: [email protected]
by guest on June 2, 2013http://m
be.oxfordjournals.org/D
ownloaded from

Taken together, our results suggest that TRIM71 and let-7are an evolutionarily conserved miRNA–target pair andthat TRIM71 likely plays an important role in embryonicdevelopment.
Materials and MethodsVerified miRNA–Target Pairs
Information regarding the miRNA–target pairs veri-fied by experiment was obtained from TarBase version 3(Sethupathy et al. 2006). A total of 14 experimentally sup-ported miRNA–target pairs were identified in C. elegans.
Identification of Human Orthologues of C. elegansmiRNA–Target Pairs
For each of the verified miRNA–target pairs in C. el-egans, we first looked for its miRNA orthologues and targetorthologues in human. We obtained the family informationon each miRNA of the pairs from miRBase release 9 (Steinet al. 2003) to identify whether the miRNA had orthologuesin human or not. Moreover, every C. elegans target proteinsequence of the pairs was used as a query to search againstthe whole set of human protein sequences from Ensembldatabase release 42.36d (Waterston et al. 2002) usingBlastP program (Altschul et al. 1990). If there existed Blasthits with E value , 10�20, we took the hits with the small-est E values as the human orthologues of the target gene.
Definition of Alignable Length and Alignable LengthRatio of a Protein
The ‘‘alignable length’’ between the 2 proteins ofa Blast query was defined as the sum of all the lengthsof the Blast hits of the query minus the gap lengths. The‘‘alignable length ratio of a protein’’ was defined as thealignable length divided by the protein length.
Identification of Putative miRNA Target Sites
In each of the human orthologous pairs identified bythe above procedure, we used a bioinformatics approach topredict putative miRNA binding sites of the cognate mi-RNA in the 3# UTR sequence. The 3# UTR sequences wereretrieved from the Ensembl database (Homo sapiens coreversion 42.36d) by the Ensembl Core API (Waterstonet al. 2002). We considered 2 factors for judging whethera segment of the 3# UTR sequence of the gene is a potentialtarget site or not. The first factor was the degree of affinity,based on binding energy, between a segment of 3# UTRsequence and miRNA sequence. We used a sliding windowof 25 nt to slide over the 3# UTR sequence of the gene andcalculated the free energy between the miRNA sequenceand the segment sequence within the window. The 2-statehybridization function of the UNAFold package (version3.0) (Dimitrov and Zuker 2004; Markham and Zuker2005) was used to perform the core of the calculation.For a segment of interest and the miRNA sequence to have
high affinity for each other, we used the criterion that thefree energy between them is ��10 kcal/mol. The secondfactor was pairing quality between the segment of interestand the miRNA sequence. A number of miRNA–targetpairing rules have been deduced in the literature (Lewiset al. 2003; Doench and Sharp 2004; Kiriakidou et al.2004; Brennecke et al. 2005; Didiano and Hobert 2006).We chose the following rules that are considered to be im-portant for pairing constraints. 1) Extensive base pairing atthe 5# end ‘‘seed’’ region of the miRNA is important for targetsite function (Lewis et al. 2003; Doench and Sharp 2004;Kiriakidou et al. 2004; Brennecke et al. 2005; Didianoand Hobert 2006); the seed region is defined as a stretchof 7 consecutive nucleotides starting from second nucleotideat the 5# end of an miRNA (Lewis et al. 2003). 2) The seedsequence or the seed-match target sequence can have a singlenucleotide bulge that is positioned symmetrically in the pair-ing region (Kiriakidou et al. 2004; Brennecke et al. 2005).3) The seed pairing region may have G:U base pairs(Kiriakidou et al. 2004; Brennecke et al. 2005; Didianoand Hobert 2006), although its presence in the seed regionis usually detrimental (Brennecke et al. 2005). 4) If the seedregion is imperfectly base paired or has a shorter stretch ofbase pairings, extensive base pairings around the 3# end ofthe miRNA should be present to compensate for the insuf-ficient pairing of the 5# end (Brennecke et al. 2005).Combining the computational results of pairing qualityand binding affinity between an miRNA sequence andthe segments of the 3#UTR sequence of a gene, we can iden-tify putative target sites for each miRNA–target pair.
Cell Culture and Plasmids
Human hepatocellular carcinoma HepG2 cell line wascultured in modified Eagle medium supplemented with10% fetal bovine serum and 0.1 mM nonessential aminoacids in 5% CO2. As described previously (Kiriakidouet al. 2004; Boutz et al. 2007), we used phRG-TK plasmid(coding for Renilla luciferase; Promega, Madison, WI) toconstruct the let-7 target site reporters and used thepGL3-control plasmid as a control plasmid (coding for fire-fly luciferase; Promega) for normalizing the transfection ef-ficiency. Primer sequences are listed in supplementary tableS4 (Supplementary Material online). To construct targetsite reporter plasmids, various target fragments, which werecloned by polymerase chain reaction (PCR) from a cDNApool of HRK 293 cells, were inserted at the XbaI site, down-stream of the luciferase gene in the phRG-TK vector. Thetarget fragment of Vitamin D3 Receptor (VDR) includingnt1922 to nt4232 of human VDR cDNA (accession no.NM_001017535) was amplified by PCR with primersVDR1922 and VDR4232R. The target fragment ofNR1I2 including nt3535 to nt4366 of human NR1I2 gene(accession no. NM_003889) was amplified by PCR withprimers NR1I2 F and NR1I2 R from the human genomicDNA of peripheral blood monocytes. The target fragmentof FOXA1 including nt1814 to nt2850R of human FOXA1cDNA (accession no. NM_004496) was amplified by PCRwith primers FOXA1814 and FOXA2850R. The targetfragment of TRIM71 including nt2810 to nt2958 of human
2526 Lin et al.
by guest on June 2, 2013http://m
be.oxfordjournals.org/D
ownloaded from

TRIM71 cDNA (accession no. NM_001039111) was am-plified by PCR with primers TRIM2810 and TRIM2958R.The target fragment of Danio rerio lin-41 (DLIN41) includ-ing nt2222 to nt2389 of zebra fish lin-41 cDNA (accessionno. XM_685160) was amplified by PCR with primersDLIN2222 and DLIN2389R from zebra fish genomicDNA. PCR was performed by using Phusion DNA poly-merase (Finnzymes Oy, Espoo, Finland) following manu-facturer’s instruction. The TRIM71 mutants with deletion ofthe first target site (nt2828 to nt2849), the second target site(nt2908 to nt2929), or both sites (nt2828 to nt2849 and2908 to nt2929) were generated by QuickChange II XLSite-directed Mutagensis Kit (Stratagene, La Jolla, CA)with the combination of following primer pairs: TRIM71mt1 F and TRIM71 mt1 R for the first site; TRIM71mt2 F and TRIM71 mt2 R for the second site, accordingto the instruction. The plet7AS reporter plasmid, whichwas inserted only with the antisense sequence of has-let-7a,was generated by inserting the ds-oligo, which wereannealed from primer pairs let7AS F and let7AS R intothe XbaI site of phRG-TK vector.
Transfection and Luciferase Reporter Assay
Transient transfection and luciferase activity assayswere performed as described previously with modifications(Kiriakidou et al. 2004; Boutz et al. 2007). miRNA precur-sors for hsa-let-7a (pre-let7a), hsa-let-7c (pre-let7c), andthe negative control precursor #1 (pre-ctrl) were purchasedfrom Ambion (Austin, TX). HepG2 cells were seeded in 12-well plates 24 h before transfection. Cells were transfectedwith indicated amounts of precursor miRNA oligo and re-porter plasmids (0.2 lg of target reporter and 0.2 lg ofpGL3-control) in the presence of Lipofectamine2000 re-agent (Invitrogen, Carlsbad, CA) following manufacturer’smanual. Three days posttransfection, the luciferase activitywas determined using the Dual-luciferase assay system(Promega) according to the manufacturer’s instruction.The Renilla luciferase activity was normalized by dividingthe firefly luciferase activity of the same transfection. Thenormalized luciferase activity of pre-ctrl in vector transfec-tion was set to 1.0, and all others were expressed relative to it.
let-7a and let-7c miRNA Detection and Quantification
Total RNA was extracted by the Trizol reagent (Invi-trogen), and quantitative reverse transcriptase–polymerasechain reaction for let-7a (or let-7c) and U6 RNA was per-formed with the TaqMan Real-time PCR kit (Applied Bio-systems, Foster City, CA) according to the manufacturer’sinstructions and detected with an ABI Prism 7000 sequencedetection system (Applied Biosystems). The raw data wereanalyzed by ABI Prism 7000 SDS software (Applied Biosys-tems). The cycle threshold, Ct, of each sample was generatedwith thedefault setting.The let-7expression levelofeachsam-ple was normalized to the expression level of U6 in the samesample by the formula: let-7/U6 5 2�(Ct of let-7a-Ct of U6). Thelet-7/U6 ratio of the 2 nM pre-ctrl oligo transfection was setto 1.0, and the values of all others relative to it were calculatedaccordingly. The result represents the average of 3 indepen-dent experiments with standard deviations.
lin-41 Silencing in the Zebrafish Model
To silence the lin-41 expression in zebrafish with (smallinterfering RNA [siRNA]), we used the pcDNA6.2-GW/EmGFP-miR of the Block-iT Pol II miR RNAi ExpressionVector Kits (Invitrogen) to construct engineered siRNA-expressing plasmids according to the user manual. Two re-gions of zebrafish lin-41 orthologue cDNA were chosen forthe engineered siRNAs: one from nt716 to nt736 and theother from nt1611 to nt1631. The pcDNA6.2-GW/EmGFP-neg control plasmid, which expresses the engi-neered siRNA with random sequence, served as a negativecontrol. These 3 resulting plasmids were used as templatesfor PCR amplifications with 2 primers, siGW F and siGWR, to obtain the inserts for the next transposon plasmid con-struction. These amplified inserts were digested with ClaIand ligated into the ClaI sites of pretreated transposonpT2KXIG plasmid (Kawakami et al. 2004), which has beendigested with ApaI and then self-ligated first, to obtainthe engineered siRNA-expressing transposon plasmids,716RNA interference (RNAi), 1611RNAi, or RNAi con-trol. One hundred picograms of the final transposonplasmid was comicroinjected with 50 pg of pCS-TP plas-mid, which expressed the transposase (Kawakami et al.2004), into 1-cell stage of zebrafish embryos. For eachtransposon plasmid injection, at least 150 eggs were in-jected and the abnormal embryos were counted under thedissection microscope.
In addition to the RNAi strategy, morpholino knock-down experiments were also carried out to silence lin-41expression. Morpholinos are chemically modified oligonu-cleotides (Nasevicius and Ekker 2000), which have beenwidely used in zebrafish and frog studies to specificallyknockdown gene expression by blocking translation. Weinjected lin-41 morpholino or control morpholino intothe yolk region at the 1- to 4-cell stage of zebrafish em-bryos. lin-41 morpholino (5#-ATTGAGCATGGAGAAG-CTGTTGTGG-3#) and nonspecific control morpholino(5#-CCTCTTACCTCAGTTACAATTTATA-3#) werepurchased from Gene Tools, Philomath, OR.
ResultsCaenorhabditis elegans let-7 Targets and Their HumanOrthologues Have Similar Functions
There were 14 experimentally verified miRNA–targetpairs in C. elegans (supplementary table S1, SupplementaryMaterial online), and we found only 4 of them to have or-thologues for both miRNA and target gene in human (seeMaterials and Methods). These consisted of let-7 miRNAand its 4 target genes in C. elegans.
These 4 verified C. elegans target genes of let-7 aredaf-12 (Grosshans et al. 2005), pha-4 (Grosshans et al.2005), let-60 (Johnson et al. 2005), and lin-41 (Slacket al. 2000). Their human orthologues found by amino acidsequence Blast are listed in table 1, and the Blast results aresummarized in supplementary table S2 (SupplementaryMaterial online). It was noted that some of the orthologousgenes had paralogues (duplicates). In the case of worm daf-12, both VDR (also termed NR1I1) and NR1I2 (also termedPXR, the pregnane X receptor [Kliewer et al. 1998] and
TRIM71 Is a Conserved let-7 Target in Evolution 2527
by guest on June 2, 2013http://m
be.oxfordjournals.org/D
ownloaded from

SXR, the steroid and xenobiotic receptor [Blumberg et al.1998]) are its human orthologues. VDR and NR1I2 are bothnuclear receptors (Evans 1988; Kliewer et al. 1998); VDRfunctions as a ligand-dependent transcription factor tomaintain calcium metabolism through the regulation of tar-get genes in response to vitamin D3 (Jones et al. 1998).NR1I2 is also a ligand-dependent transcription factor in-volved in the regulation of a number of xenobiotic-metab-olizing genes (Bertilsson et al. 1998; Blumberg et al. 1998;Kliewer et al. 1998; Lehmann et al. 1998). Similarly, theworm daf-12, which regulates the dauer diapause and de-velopmental age (Antebi et al. 1998, 2000), is also a nuclearreceptor (Yeh 1991) and has been seen as being closely re-lated to vertebrate VDR and NR1I2 (Antebi et al. 2000). Theworm pha-4, which encodes a protein that most closely re-sembles a forkhead box A (FOXA) transcription factor(Horner et al. 1998; Kalb et al. 1998), has 3 human ortho-logues FOXA1, FOXA2, and FOXA3. pha-4 functions inorganogenesis of the C. elegans pharynx (Azzaria et al.1996; Kalb et al. 1998; Gaudet and Mango 2002). In mam-mals, the FOXA family has been shown to be critical inmultiple developmental stages, beginning with early devel-opment, continuing during organogenesis, and finally in me-tabolism and homeostasis in the adult (reviewed in Friedmanand Kaestner 2006). let-60, a GTP-binding RAS proto-oncogene (Han and Sternberg 1990), is theC. elegans ortho-logue of human HRAS, KRAS, and NRAS. Because humanRAS genes have recently been verified to be the target genesof let-7miRNA (Johnson et al. 2005), they were not pursuedin this study.
The C. elegans heterochronic gene lin-41, which en-codes a Ring finger–B box–coiled coil (RBCC) protein(Slack et al. 2000), seems to have only one human ortho-logue, that is, TRIM71. TRIM71 is a novel gene withunknown function in human; however, its mouse orthologueis also a member of the RBCC protein family (Kanamotoet al. 2006). During mouse embryogenesis, reciprocal ex-pression of the lin-41 orthologue and let-7 has been reported(Schulman et al. 2005). Moreover, in chicken and mouse, thelin-41 orthologues seem to be important in developing limbbuds, branchial arches, and tail buds (Lancman et al. 2005;Schulman et al. 2005; Kanamoto et al. 2006).
From our survey, the 4 target genes of the let-7 mi-RNA in C. elegans and their human orthologues have, tosome degree, similar functions. This observation lends fur-ther support for these human genes as the orthologues of theC. elegans genes.
Putative Target Sites for let-7 on Human Orthologues
The predicted target sites of let-7 on human ortho-logues are listed in supplementary table S3 (SupplementaryMaterial online). The putative target sites were identified byconsidering the pairing quality and the degree of bindingaffinity between an miRNA sequence and the segmentsof the 3# UTR sequence of an orthologous gene (see Ma-terials and Methods). We did not impose the cross-speciesconservation requirement, which has been used in many tar-get prediction programs (Enright et al. 2002; Lewis et al.2003, 2005; John et al. 2004; Kiriakidou et al. 2004; Grunet al. 2005; Krek et al. 2005) because we tried to identify notonly the evolutionarily conserved target sites but also thehuman-specific target sites. In addition, as our pairing con-straints were loose (see Materials and Methods), these pu-tative target sites likely include most or all of the targetsites. This is useful for identifying a sufficient portion ofa 3# UTR sequence that can be tested in the followingexperiments.
For each of the C. elegans genes we studied, we choseone of its human orthologues that has most putative targetsites on its 3# UTR for the first round reporter assay. Aftersurveying the putative target sites of the human genes, wechose VDR, FOXA1, and TRIM71 to examine whether theyare targets of let-7. We then chose the paralogues of thehuman genes used in the first round test for the secondround reporter assay. As shown in table 1, FOXA2 andFOXA3 are paralogues of FOXA1 and NR1I2 is a paralogueof VDR, and these 3 genes had not been tested in the firstround. By surveying the putative target sites of these 3genes, we found that NR1I2 had a perfect seed-match site,but FOXA2 and FOXA3 did not have good putative targetsites. Therefore, NR1I2 was selected for the second roundtest. The locations of the putative target sites of these 4 se-lected human genes are shown in figure 1A. It is noteworthythat 2 of the putative target sites of TRIM71 have perfect basepairing with the seed region of let-7 and have been evolution-arily conserved (see supplementary fig. S2, SupplementaryMaterial online). On the other hand, NR1I2 has a perfectseed-match site, but the site is not evolutionarily conserved(supplementary fig. S2, Supplementary Material online).
Both hsa-let-7a and hsa-let-7c Repress Human TRIM71
To examine the connection between the target ortho-logues and their cognate miRNAs, the luciferase reporterassay provides a reliable solution and has been used inmany studies. Because the sequences of the let-7 familyhave been highly conserved from C. elegans to human (sup-plementary fig. S1, Supplementary Material online), weused hsa-let-7a to perform the reporter assay for 2 reasons.First, the sequences of hsa-let-7a and C. elegans let-7 areidentical. Second, hsa-let-7a has been shown to repressRAS effectively (Johnson et al. 2005). We first constructed
Table 1The 4 Caenorhabditis elegans Target Genes and TheirHuman Orthologues
C. elegans Gene Orthologous Gene (in Human)
daf-12: Nuclear receptorVDR (NR1I1), NR1I2:Nuclear receptors
pha-4: Forkheadtranscription factor
FOXA1, FOXA2, FOXA3:Forkhead transcription factors
let-60: GTP-binding RASproto-oncogene
HRAS, KRAS, NRAS:GTP-binding RASproto-oncogenes
lin-41: RBCC heterochronic gene TRIM71: RBCC protein gene
NOTE.—The name and the functional or structural information of the 4 C.
elegans target genes are listed in the left column and that of their human orthologues
are listed in the right column. All the 4 C. elegans genes are experimentally verified
targets of let-7. Except for lin-41, the other 3 C. elegans target genes have multiple
human orthologues. Each of the C. elegans target genes and its human orthologues
have similar function or structure.
2528 Lin et al.
by guest on June 2, 2013http://m
be.oxfordjournals.org/D
ownloaded from
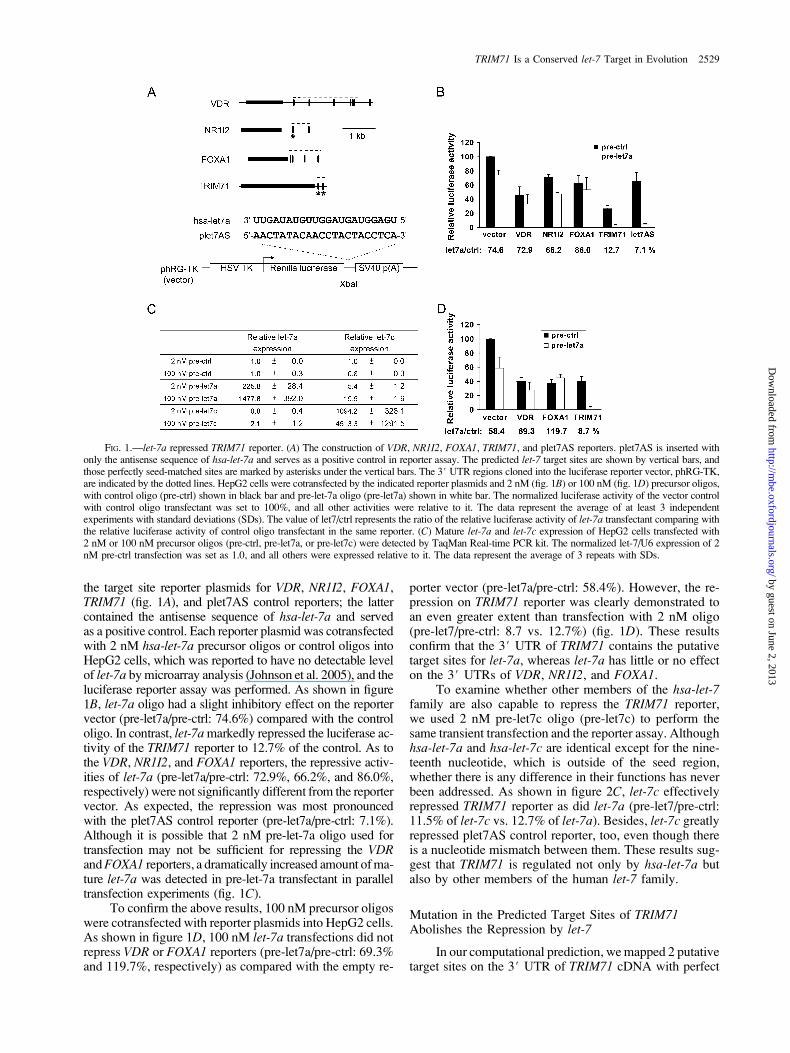
the target site reporter plasmids for VDR, NR1I2, FOXA1,TRIM71 (fig. 1A), and plet7AS control reporters; the lattercontained the antisense sequence of hsa-let-7a and servedas a positive control. Each reporter plasmid was cotransfectedwith 2 nM hsa-let-7a precursor oligos or control oligos intoHepG2 cells, which was reported to have no detectable levelof let-7a by microarray analysis (Johnson et al. 2005), and theluciferase reporter assay was performed. As shown in figure1B, let-7a oligo had a slight inhibitory effect on the reportervector (pre-let7a/pre-ctrl: 74.6%) compared with the controloligo. In contrast, let-7a markedly repressed the luciferase ac-tivity of the TRIM71 reporter to 12.7% of the control. As tothe VDR, NR1I2, and FOXA1 reporters, the repressive activ-ities of let-7a (pre-let7a/pre-ctrl: 72.9%, 66.2%, and 86.0%,respectively) were not significantly different from the reportervector. As expected, the repression was most pronouncedwith the plet7AS control reporter (pre-let7a/pre-ctrl: 7.1%).Although it is possible that 2 nM pre-let-7a oligo used fortransfection may not be sufficient for repressing the VDRandFOXA1 reporters, a dramatically increased amount of ma-ture let-7a was detected in pre-let-7a transfectant in paralleltransfection experiments (fig. 1C).
To confirm the above results, 100 nM precursor oligoswere cotransfected with reporter plasmids into HepG2 cells.As shown in figure 1D, 100 nM let-7a transfections did notrepress VDR or FOXA1 reporters (pre-let7a/pre-ctrl: 69.3%and 119.7%, respectively) as compared with the empty re-
porter vector (pre-let7a/pre-ctrl: 58.4%). However, the re-pression on TRIM71 reporter was clearly demonstrated toan even greater extent than transfection with 2 nM oligo(pre-let7/pre-ctrl: 8.7 vs. 12.7%) (fig. 1D). These resultsconfirm that the 3# UTR of TRIM71 contains the putativetarget sites for let-7a, whereas let-7a has little or no effecton the 3# UTRs of VDR, NR1I2, and FOXA1.
To examine whether other members of the hsa-let-7family are also capable to repress the TRIM71 reporter,we used 2 nM pre-let7c oligo (pre-let7c) to perform thesame transient transfection and the reporter assay. Althoughhsa-let-7a and hsa-let-7c are identical except for the nine-teenth nucleotide, which is outside of the seed region,whether there is any difference in their functions has neverbeen addressed. As shown in figure 2C, let-7c effectivelyrepressed TRIM71 reporter as did let-7a (pre-let7/pre-ctrl:11.5% of let-7c vs. 12.7% of let-7a). Besides, let-7c greatlyrepressed plet7AS control reporter, too, even though thereis a nucleotide mismatch between them. These results sug-gest that TRIM71 is regulated not only by hsa-let-7a butalso by other members of the human let-7 family.
Mutation in the Predicted Target Sites of TRIM71Abolishes the Repression by let-7
In our computational prediction, we mapped 2 putativetarget sites on the 3# UTR of TRIM71 cDNA with perfect
FIG. 1.—let-7a repressed TRIM71 reporter. (A) The construction of VDR, NR1I2, FOXA1, TRIM71, and plet7AS reporters. plet7AS is inserted withonly the antisense sequence of hsa-let-7a and serves as a positive control in reporter assay. The predicted let-7 target sites are shown by vertical bars, andthose perfectly seed-matched sites are marked by asterisks under the vertical bars. The 3# UTR regions cloned into the luciferase reporter vector, phRG-TK,are indicated by the dotted lines. HepG2 cells were cotransfected by the indicated reporter plasmids and 2 nM (fig. 1B) or 100 nM (fig. 1D) precursor oligos,with control oligo (pre-ctrl) shown in black bar and pre-let-7a oligo (pre-let7a) shown in white bar. The normalized luciferase activity of the vector controlwith control oligo transfectant was set to 100%, and all other activities were relative to it. The data represent the average of at least 3 independentexperiments with standard deviations (SDs). The value of let7/ctrl represents the ratio of the relative luciferase activity of let-7a transfectant comparing withthe relative luciferase activity of control oligo transfectant in the same reporter. (C) Mature let-7a and let-7c expression of HepG2 cells transfected with2 nM or 100 nM precursor oligos (pre-ctrl, pre-let7a, or pre-let7c) were detected by TaqMan Real-time PCR kit. The normalized let-7/U6 expression of 2nM pre-ctrl transfection was set as 1.0, and all others were expressed relative to it. The data represent the average of 3 repeats with SDs.
TRIM71 Is a Conserved let-7 Target in Evolution 2529
by guest on June 2, 2013http://m
be.oxfordjournals.org/D
ownloaded from
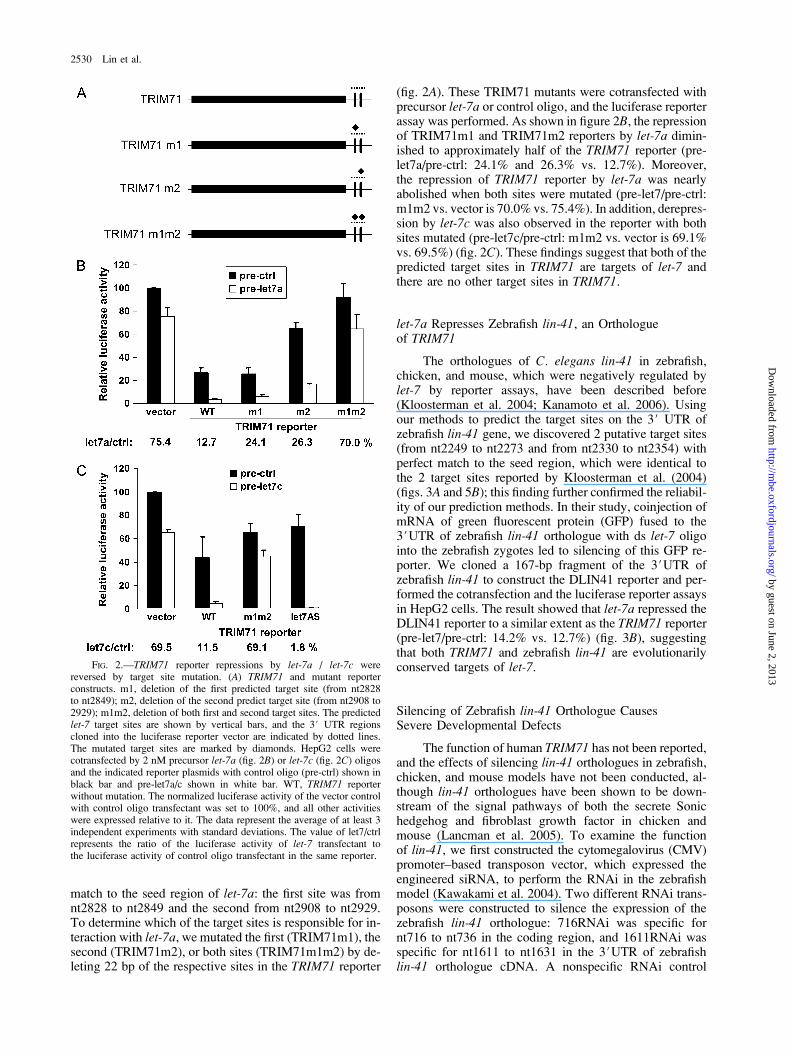
match to the seed region of let-7a: the first site was fromnt2828 to nt2849 and the second from nt2908 to nt2929.To determine which of the target sites is responsible for in-teraction with let-7a, we mutated the first (TRIM71m1), thesecond (TRIM71m2), or both sites (TRIM71m1m2) by de-leting 22 bp of the respective sites in the TRIM71 reporter
(fig. 2A). These TRIM71 mutants were cotransfected withprecursor let-7a or control oligo, and the luciferase reporterassay was performed. As shown in figure 2B, the repressionof TRIM71m1 and TRIM71m2 reporters by let-7a dimin-ished to approximately half of the TRIM71 reporter (pre-let7a/pre-ctrl: 24.1% and 26.3% vs. 12.7%). Moreover,the repression of TRIM71 reporter by let-7a was nearlyabolished when both sites were mutated (pre-let7/pre-ctrl:m1m2 vs. vector is 70.0% vs. 75.4%). In addition, derepres-sion by let-7c was also observed in the reporter with bothsites mutated (pre-let7c/pre-ctrl: m1m2 vs. vector is 69.1%vs. 69.5%) (fig. 2C). These findings suggest that both of thepredicted target sites in TRIM71 are targets of let-7 andthere are no other target sites in TRIM71.
let-7a Represses Zebrafish lin-41, an Orthologueof TRIM71
The orthologues of C. elegans lin-41 in zebrafish,chicken, and mouse, which were negatively regulated bylet-7 by reporter assays, have been described before(Kloosterman et al. 2004; Kanamoto et al. 2006). Usingour methods to predict the target sites on the 3# UTR ofzebrafish lin-41 gene, we discovered 2 putative target sites(from nt2249 to nt2273 and from nt2330 to nt2354) withperfect match to the seed region, which were identical tothe 2 target sites reported by Kloosterman et al. (2004)(figs. 3A and 5B); this finding further confirmed the reliabil-ity of our prediction methods. In their study, coinjection ofmRNA of green fluorescent protein (GFP) fused to the3#UTR of zebrafish lin-41 orthologue with ds let-7 oligointo the zebrafish zygotes led to silencing of this GFP re-porter. We cloned a 167-bp fragment of the 3#UTR ofzebrafish lin-41 to construct the DLIN41 reporter and per-formed the cotransfection and the luciferase reporter assaysin HepG2 cells. The result showed that let-7a repressed theDLIN41 reporter to a similar extent as the TRIM71 reporter(pre-let7/pre-ctrl: 14.2% vs. 12.7%) (fig. 3B), suggestingthat both TRIM71 and zebrafish lin-41 are evolutionarilyconserved targets of let-7.
Silencing of Zebrafish lin-41 Orthologue CausesSevere Developmental Defects
The function of human TRIM71 has not been reported,and the effects of silencing lin-41 orthologues in zebrafish,chicken, and mouse models have not been conducted, al-though lin-41 orthologues have been shown to be down-stream of the signal pathways of both the secrete Sonichedgehog and fibroblast growth factor in chicken andmouse (Lancman et al. 2005). To examine the functionof lin-41, we first constructed the cytomegalovirus (CMV)promoter–based transposon vector, which expressed theengineered siRNA, to perform the RNAi in the zebrafishmodel (Kawakami et al. 2004). Two different RNAi trans-posons were constructed to silence the expression of thezebrafish lin-41 orthologue: 716RNAi was specific fornt716 to nt736 in the coding region, and 1611RNAi wasspecific for nt1611 to nt1631 in the 3#UTR of zebrafishlin-41 orthologue cDNA. A nonspecific RNAi control
FIG. 2.—TRIM71 reporter repressions by let-7a / let-7c werereversed by target site mutation. (A) TRIM71 and mutant reporterconstructs. m1, deletion of the first predicted target site (from nt2828to nt2849); m2, deletion of the second predict target site (from nt2908 to2929); m1m2, deletion of both first and second target sites. The predictedlet-7 target sites are shown by vertical bars, and the 3# UTR regionscloned into the luciferase reporter vector are indicated by dotted lines.The mutated target sites are marked by diamonds. HepG2 cells werecotransfected by 2 nM precursor let-7a (fig. 2B) or let-7c (fig. 2C) oligosand the indicated reporter plasmids with control oligo (pre-ctrl) shown inblack bar and pre-let7a/c shown in white bar. WT, TRIM71 reporterwithout mutation. The normalized luciferase activity of the vector controlwith control oligo transfectant was set to 100%, and all other activitieswere expressed relative to it. The data represent the average of at least 3independent experiments with standard deviations. The value of let7/ctrlrepresents the ratio of the luciferase activity of let-7 transfectant tothe luciferase activity of control oligo transfectant in the same reporter.
2530 Lin et al.
by guest on June 2, 2013http://m
be.oxfordjournals.org/D
ownloaded from
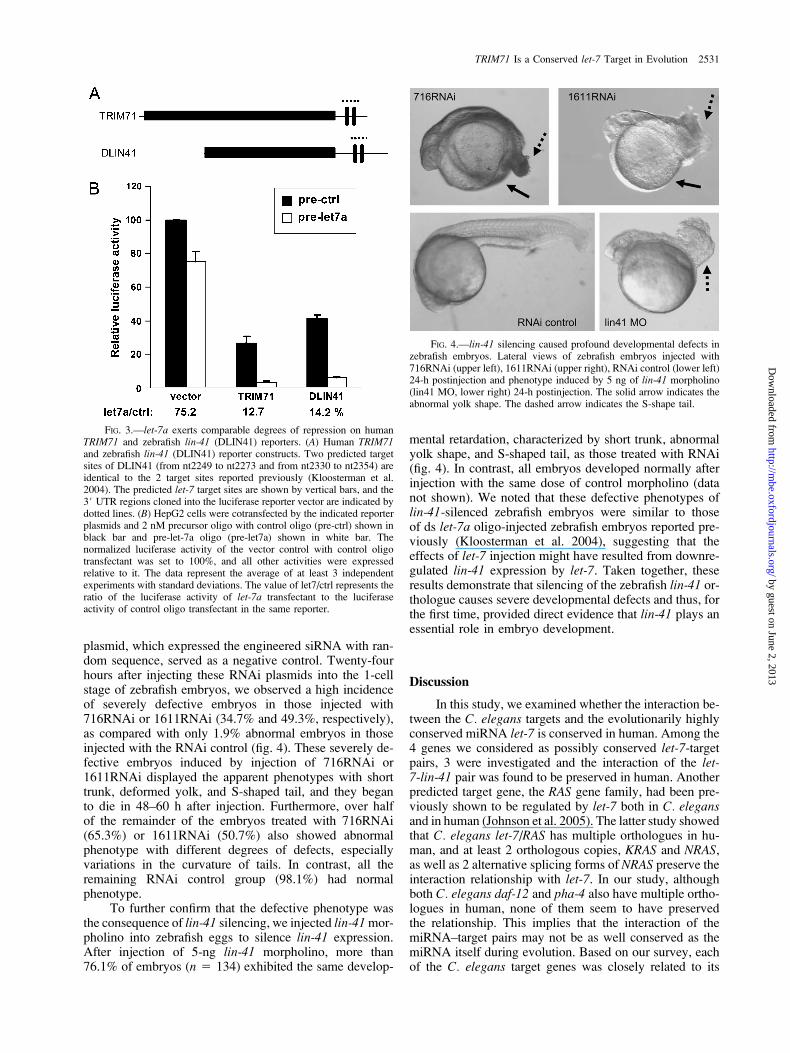
plasmid, which expressed the engineered siRNA with ran-dom sequence, served as a negative control. Twenty-fourhours after injecting these RNAi plasmids into the 1-cellstage of zebrafish embryos, we observed a high incidenceof severely defective embryos in those injected with716RNAi or 1611RNAi (34.7% and 49.3%, respectively),as compared with only 1.9% abnormal embryos in thoseinjected with the RNAi control (fig. 4). These severely de-fective embryos induced by injection of 716RNAi or1611RNAi displayed the apparent phenotypes with shorttrunk, deformed yolk, and S-shaped tail, and they beganto die in 48–60 h after injection. Furthermore, over halfof the remainder of the embryos treated with 716RNAi(65.3%) or 1611RNAi (50.7%) also showed abnormalphenotype with different degrees of defects, especiallyvariations in the curvature of tails. In contrast, all theremaining RNAi control group (98.1%) had normalphenotype.
To further confirm that the defective phenotype wasthe consequence of lin-41 silencing, we injected lin-41 mor-pholino into zebrafish eggs to silence lin-41 expression.After injection of 5-ng lin-41 morpholino, more than76.1% of embryos (n 5 134) exhibited the same develop-
mental retardation, characterized by short trunk, abnormalyolk shape, and S-shaped tail, as those treated with RNAi(fig. 4). In contrast, all embryos developed normally afterinjection with the same dose of control morpholino (datanot shown). We noted that these defective phenotypes oflin-41-silenced zebrafish embryos were similar to thoseof ds let-7a oligo-injected zebrafish embryos reported pre-viously (Kloosterman et al. 2004), suggesting that theeffects of let-7 injection might have resulted from downre-gulated lin-41 expression by let-7. Taken together, theseresults demonstrate that silencing of the zebrafish lin-41 or-thologue causes severe developmental defects and thus, forthe first time, provided direct evidence that lin-41 plays anessential role in embryo development.
Discussion
In this study, we examined whether the interaction be-tween the C. elegans targets and the evolutionarily highlyconserved miRNA let-7 is conserved in human. Among the4 genes we considered as possibly conserved let-7-targetpairs, 3 were investigated and the interaction of the let-7-lin-41 pair was found to be preserved in human. Anotherpredicted target gene, the RAS gene family, had been pre-viously shown to be regulated by let-7 both in C. elegansand in human (Johnson et al. 2005). The latter study showedthat C. elegans let-7/RAS has multiple orthologues in hu-man, and at least 2 orthologous copies, KRAS and NRAS,as well as 2 alternative splicing forms of NRAS preserve theinteraction relationship with let-7. In our study, althoughboth C. elegans daf-12 and pha-4 also have multiple ortho-logues in human, none of them seem to have preservedthe relationship. This implies that the interaction of themiRNA–target pairs may not be as well conserved as themiRNA itself during evolution. Based on our survey, eachof the C. elegans target genes was closely related to its
FIG. 3.—let-7a exerts comparable degrees of repression on humanTRIM71 and zebrafish lin-41 (DLIN41) reporters. (A) Human TRIM71and zebrafish lin-41 (DLIN41) reporter constructs. Two predicted targetsites of DLIN41 (from nt2249 to nt2273 and from nt2330 to nt2354) areidentical to the 2 target sites reported previously (Kloosterman et al.2004). The predicted let-7 target sites are shown by vertical bars, and the3# UTR regions cloned into the luciferase reporter vector are indicated bydotted lines. (B) HepG2 cells were cotransfected by the indicated reporterplasmids and 2 nM precursor oligo with control oligo (pre-ctrl) shown inblack bar and pre-let-7a oligo (pre-let7a) shown in white bar. Thenormalized luciferase activity of the vector control with control oligotransfectant was set to 100%, and all other activities were expressedrelative to it. The data represent the average of at least 3 independentexperiments with standard deviations. The value of let7/ctrl represents theratio of the luciferase activity of let-7a transfectant to the luciferaseactivity of control oligo transfectant in the same reporter.
FIG. 4.—lin-41 silencing caused profound developmental defects inzebrafish embryos. Lateral views of zebrafish embryos injected with716RNAi (upper left), 1611RNAi (upper right), RNAi control (lower left)24-h postinjection and phenotype induced by 5 ng of lin-41 morpholino(lin41 MO, lower right) 24-h postinjection. The solid arrow indicates theabnormal yolk shape. The dashed arrow indicates the S-shape tail.
TRIM71 Is a Conserved let-7 Target in Evolution 2531
by guest on June 2, 2013http://m
be.oxfordjournals.org/D
ownloaded from

human orthologues either in structure or in function. In anattempt to explain why the interaction is not always pre-served, we compared the conserved systems (the lin-41and let-60 systems) and the nonconserved systems (thedaf-12 and pha-4 systems). It was noted that the alignablelength ratios (see Materials and Methods and supplementarytable S2 [Supplementary Material online]) of the 2 con-served systems were much higher than that of the 2 noncon-served systems (0.61 ; 1.00 for the conserved systems,whereas 0.20 ; 0.41 for the nonconserved systems). Thissuggests that the orthologous proteins in the conserved sys-tems may have greater similarities in protein domains andtheir structure and function, so that the interaction with thehighly conserved miRNA may be better preserved. In con-trast, the orthologous proteins in the nonconserved systemsmay have less similar protein domains; some of their ances-tral functions may have been lost during evolution and themiRNA–target regulation may have been replaced or com-pensated by other regulatory pathways. To sum up, it seemsthat a system that has a higher alignable length ratio may bebetter conserved. However, a definitive conclusion awaitsstudies of a larger number of cases.
We noticed that NR1I2 has a perfect let-7 seed-matchsite in the 3#UTR (supplementary fig. S2, SupplementaryMaterial online), and yet our NR1I2 reporter construct didnot provide clear-cut evidence that it could be regulatedby let-7. This suggests that the perfect seed-match rule isnot the only criterion for the miRNA–target binding. It isknown that miRNAs can act in a combinatorial fashion (Kreket al. 2005), and multiple miRNAs may contribute togetherto an important effect. Thus, we could not exclude the pos-
sibility that these genes are regulated by let-7 in concert withother miRNAs acting simultaneously. In contrast, TRIM71,which has 2 evolutionarily conserved perfect seed-matchsites in the 3#UTR (supplementary fig. S2, SupplementaryMaterial online), was shown to be the target of let-7. More-over, it is intriguing that even though the locations of the 2sites in the 3#UTR in human are different from that in zebrafish, the distance between the 2 sites in human (81 nt) is aboutthe same as that in zebra fish (80 nt) (fig. 5). Further, wenotice that the 2 let-7 target sites of lin-41 (the worm ortho-logoue of TRIM71) (Reinhart et al. 2000; Slack et al. 2000;Vella et al. 2004) are not perfect match to the seed region buthave high binding energy (fig. 5 and supplementary fig. S2[Supplementary material online]). This suggests that even ifthe interaction relationship of these orthologous miRNA–target pairs is conserved during evolution, the pairing con-dition for these pairs may be different.
Downregulation of target genes by miRNA may in-volve one of the 2 posttranscriptional suppression mecha-nisms: translational repression or mRNA cleavage(reviewed in Bartel 2004). It has been reported that the deg-radation of lin-41 mRNA is induced by let-7 in C. elegans(Bagga et al. 2005). However, in zebrafish, the plasmid-containing GFP gene fused with the 3#UTR of the lin-41zebra fish orthologue was translationally repressed bylet-7, but its mRNA level was unchanged (Kloostermanet al. 2004). This implies that even though the orthologousmiRNA–target pairs are conserved in evolution, their reg-ulatory mechanisms may have changed.
We demonstrated that human TRIM71 is a specifictarget of let-7a and let-7c by the luciferase reporter assay.
FIG. 5.—The locations in 3# UTRs and the pairing information of the let-7 target sites of the TRIM71: let-7 (or lin-41: let-7) orthologous pairs inhuman (A), zebrafish (B) and nematode (C). Cartoon shows the 3# UTRs of human TRIM71, zebrafish lin-41 orthologue, and Caenorhabditis eleganslin-41. The locations of the target sites are indicated by arrows, and the pairing information for each of the target site is showed in a rectangle box linkedto the site. The pairing information is based on the results generated by the UNAFold package. For each pairing information, let-7 sequence is shown atthe bottom with 5# ribonucleotide to the right and seed region in bold, and the 25 nt putative target site sequence is shown on top with 5# ribonucleotideto the left. The target sites with perfect seed match are denoted by solid arrows and those without were indicated by hollow arrows. Each of the 3 targetgenes has 2 let-7 target sites in their 3# UTRs, and the distance between the 2 sites of the target gene is 80, 81, and 49 nt in human, zebrafish, andnematode, respectively.
2532 Lin et al.
by guest on June 2, 2013http://m
be.oxfordjournals.org/D
ownloaded from

Besides the pair of RAS and let-7, the TRIM7 and let-7 pairis another target–miRNA pair that showed evolutionaryconservation in both C. elegans and human. In additionto C. elegans and human, the evolutionarily conserved pairof lin-41 and let-7 has also been verified in zebrafish(Kloosterman et al. 2004), chicken, and mouse (Kanamotoet al. 2006), providing additional evidence to support thatthe regulation of lin-41 mediated by let-7 miRNA is con-served in animals, from C. elegans to human.
Why has this target–miRNA interaction been con-served in distantly related animals? The answer may belinked to the importance of the lin-41 gene in animal devel-opment. The lin-41 protein family, from nematodes tomammals, belongs to the RBCC protein family and sharesmost of the functional domains (Lancman et al. 2005;Schulman et al. 2005; Kanamoto et al. 2006). In C. elegans,late larval activation of let-7 expression downregulates lin-41 to relieve inhibition of the expression of the adult spec-ification transcription factor lin-29 (Slack et al. 2000), andnull mutation of let-7 caused precocious expression of adultfates at larval stages. Another study demonstrated a role forthe lin-41 gene in C. elegans male tail tip morphogenesis(Del Rio-Albrechtsen et al. 2006). Although the above stud-ies provide no direct in vivo evidence in vertebrate animals,the results of reporter assay and the reciprocal expressionpattern of lin-41 and let-7 in chicken and mouse suggestthat the negative regulation of lin-41 by let-7 is importantin their limb development (Lancman et al. 2005; Schulmanet al. 2005;Kanamotoet al. 2006). Moreover, in ourzebrafishmodel, silencing of lin-41 by siRNA or morpholino resultedin developmentally defective embryos, thus providing directin vivo evidence for the first time that lin-41 plays an im-portant role in the development of vertebrates. To put theseresults together, we propose that downregulation of lin-41by let-7 plays a fundamental role in animal development,which may be the reason why the lin-41-let-7 pair has beenwell preserved during animal evolution.
Supplementary Material
Supplementary figures S1 and S2 and tables S1–S3 areavailable at Molecular Biology and Evolution online (http://www.mbe.oxfordjournals.org/).
Acknowledgments
We thank Arthur Shih for suggestions. This studywas supported by grants from National Science Council(NSC95-3114-P-002-005-Y to W.-H.L. and A.L.Y., andNSC 93005P to L.-C.H.) and by Academia Sinica.
Literature Cited
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. 1990.Basic local alignment search tool. J Mol Biol. 215:403–410.
Ambros V. 2004. The functions of animal microRNAs. Nature.431:350–355.
Antebi A, Culotti JG, Hedgecock EM. 1998. daf-12 regulatesdevelopmental age and the dauer alternative in Caenorhabditiselegans. Development. 125:1191–1205.
Antebi A, Yeh WH, Tait D, Hedgecock EM, Riddle DL. 2000.daf-12 encodes a nuclear receptor that regulates the dauerdiapause and developmental age in C. elegans. Genes Dev.14:1512–1527.
Azzaria M, Goszczynski B, Chung MA, Kalb JM, McGhee JD.1996. A fork head/HNF-3 homolog expressed in the pharynxand intestine of the Caenorhabditis elegans embryo. Dev Biol.178:289–303.
Bagga S, Bracht J, Hunter S, Massirer K, Holtz J, Eachus R,Pasquinelli AE. 2005. Regulation by let-7 and lin-4 miRNAsresults in target mRNA degradation. Cell. 122:553–563.
Bartel DP. 2004. MicroRNAs: genomics, biogenesis, mechanism,and function. Cell. 116:281–297.
Bertilsson G, Heidrich J, Svensson K, Asman M, Jendeberg L,Sydow-Backman M, Ohlsson R, Postlind H, Blomquist P,Berkenstam A. 1998. Identification of a human nuclear recep-tor defines a new signaling pathway for CYP3A induction.Proc Natl Acad Sci USA. 95:12208–12213.
Blumberg B, Sabbagh W Jr, Juguilon H, Bolado J Jr, vanMeter CM, Ong ES, Evans RM. 1998. SXR, a novel steroidand xenobiotic-sensing nuclear receptor. Genes Dev. 12:3195–3205.
Boutz PL, Chawla G, Stoilov P, Black DL. 2007. MicroRNAsregulate the expression of the alternative splicing factor nPTBduring muscle development. Genes Dev. 21:71–84.
Brennecke J, Stark A, Russell RB, Cohen SM. 2005. Principlesof microRNA-target recognition. PLoS Biol. 3:e85.
Carrington JC, Ambros V. 2003. Role of microRNAs in plantand animal development. Science. 301:336–338.
Del Rio-Albrechtsen T, Kiontke K, Chiou SY, Fitch DH. 2006.Novel gain-of-function alleles demonstrate a role for theheterochronic gene lin-41 in C. elegans male tail tip mor-phogenesis. Dev Biol. 297:74–86.
Didiano D, Hobert O. 2006. Perfect seed pairing is not a generallyreliable predictor for miRNA-target interactions. Nat StructMol Biol. 13:849–851.
Dimitrov RA, Zuker M. 2004. Prediction of hybridization andmelting for double-stranded nucleic acids. Biophys J. 87:215–226.
Doench JG, Sharp PA. 2004. Specificity of microRNA targetselection in translational repression. Genes Dev. 18:504–511.
Enright AJ, Van Dongen S, Ouzounis CA. 2002. An efficientalgorithm for large-scale detection of protein families. NucleicAcids Res. 30:1575–1584.
Evans RM. 1988. The steroid and thyroid hormone receptorsuperfamily. Science. 240:889–895.
Friedman JR, Kaestner KH. 2006. The Foxa family of transcriptionfactors in development and metabolism. Cell Mol Life Sci.63:2317–2328.
Gaudet J, Mango SE. 2002. Regulation of organogenesis by theCaenorhabditis elegans FoxA protein PHA-4. Science. 295:821–825.
Grosshans H, Johnson T, Reinert KL, Gerstein M, Slack FJ.2005. The temporal patterning microRNA let-7 regulatesseveral transcription factors at the larval to adult transition inC. elegans. Dev Cell. 8:321–330.
Grun D, Wang YL, Langenberger D, Gunsalus KC, Rajewsky N.2005. microRNA target predictions across seven Drosophilaspecies and comparison to mammalian targets. PLoS ComputBiol. 1:e13.
Han M, Sternberg PW. 1990. let-60, a gene that specifies cellfates during C. elegans vulval induction, encodes a ras protein.Cell. 63:921–931.
Horner MA, Quintin S, Domeier ME, Kimble J, Labouesse M,Mango SE. 1998. pha-4, an HNF-3 homolog, specifies pha-ryngeal organ identity in Caenorhabditis elegans. Genes Dev.12:1947–1952.
TRIM71 Is a Conserved let-7 Target in Evolution 2533
by guest on June 2, 2013http://m
be.oxfordjournals.org/D
ownloaded from

John B, Enright AJ, Aravin A, Tuschl T, Sander C, Marks DS.2004. Human microRNA targets. PLoS Biol. 2:e363.
Johnson SM, Grosshans H, Shingara J, Byrom M, Jarvis R,Cheng A, Labourier E, Reinert KL, Brown D, Slack FJ. 2005.RAS is regulated by the let-7 microRNA family. Cell.120:635–647.
Johnston WK, Unrau PJ, Lawrence MS, Glasner ME, Bartel DP.2001. RNA-catalyzed RNA polymerization: accurate and generalRNA-templated primer extension. Science. 292:1319–1325.
Jones G, Strugnell SA, DeLuca HF. 1998. Current understandingof the molecular actions of vitamin D. Physiol Rev. 78:1193–1231.
Kalb JM, Lau KK, Goszczynski B, Fukushige T, Moons D,Okkema PG, McGhee JD. 1998. pha-4 is Ce-fkh-1, a fork head/HNF-3alpha,beta,gamma homolog that functions in organo-genesis of the C. elegans pharynx. Development. 125:2171–2180.
Kan Z, States D, Gish W. 2002. Selecting for functionalalternative splices in ESTs. Genome Res. 12:1837–1845.
Kanamoto T, Terada K, Yoshikawa H, Furukawa T. 2006.Cloning and regulation of the vertebrate homologue of lin-41that functions as a heterochronic gene in Caenorhabditiselegans. Dev Dyn. 235:1142–1149.
Kawakami K, Takeda H, Kawakami N, Kobayashi M,Matsuda N, Mishina M. 2004. A transposon-mediated genetrap approach identifies developmentally regulated genes inzebrafish. Dev Cell. 7:133–144.
Kim VN, Nam JW. 2006. Genomics of microRNA. TrendsGenet. 22:165–173.
Kiriakidou M, Nelson PT, Kouranov A, Fitziev P, Bouyioukos C,Mourelatos Z, Hatzigeorgiou A. 2004. A combinedcomputational-experimental approach predicts human micro-RNA targets. Genes Dev. 18:1165–1178.
Kliewer SA, Moore JT, Wade L, et al. (12 co-authors). 1998. Anorphan nuclear receptor activated by pregnanes defines a novelsteroid signaling pathway. Cell. 92:73–82.
Kloosterman WP, Wienholds E, Ketting RF, Plasterk RH. 2004.Substrate requirements for let-7 function in the developingzebrafish embryo. Nucleic Acids Res. 32:6284–6291.
Krek A, Grun D, Poy MN, et al. (11 co-authors). 2005. Com-binatorial microRNA target predictions. Nat Genet. 37:495–500.
Lai EC. 2003. microRNAs: runts of the genome assertthemselves. Curr Biol. 13:R925–R936.
Lancman JJ, Caruccio NC, Harfe BD, Pasquinelli AE,Schageman JJ, Pertsemlidis A, Fallon JF. 2005. Analysis ofthe regulation of lin-41 during chick and mouse limbdevelopment. Dev Dyn. 234:948–960.
Lehmann JM, McKee DD, Watson MA, Willson TM, Moore JT,Kliewer SA. 1998. The human orphan nuclear receptor PXRis activated by compounds that regulate CYP3A4 gene expres-sion and cause drug interactions. J Clin Invest. 102:1016–1023.
Lewis BP, Burge CB, Bartel DP. 2005. Conserved seed pairing,often flanked by adenosines, indicates that thousands ofhuman genes are microRNA targets. Cell. 120:15–20.
Lewis BP, Shih IH, Jones-Rhoades MW, Bartel DP, Burge CB.2003. Prediction of mammalian microRNA targets. Cell. 115:787–798.
Li WH, Gu Z, Wang H, Nekrutenko A. 2001. Evolutionaryanalyses of the human genome. Nature. 409:847–849.
Lim LP, Glasner ME, Yekta S, Burge CB, Bartel DP. 2003.Vertebrate microRNA genes. Science. 299:1540.
Lim LP, Lau NC, Garrett-Engele P, Grimson A, Schelter JM,Castle J, Bartel DP, Linsley PS, Johnson JM. 2005.Microarray analysis shows that some microRNAs down-regulate large numbers of target mRNAs. Nature. 433:769–773.
Markham NR, Zuker M. 2005. DINAMelt web server fornucleic acid melting prediction. Nucleic Acids Res. 33:W577–W581.
Mironov AA, Fickett JW, Gelfand MS. 1999. Frequent alter-native splicing of human genes. Genome Res. 9:1288–1293.
Nasevicius A, Ekker SC. 2000. Effective targeted gene‘knockdown’ in zebrafish. Nat Genet. 26:216–220.
Niwa R, Slack FJ. 2007. The evolution of animal microRNAfunction. Curr Opin Genet Dev. 17:145–150.
Pasquinelli AE, McCoy A, Jimenez E, Salo E, Ruvkun G,Martindale MQ, Baguna J. 2003. Expression of the 22nucleotide let-7 heterochronic RNA throughout the Metazoa:a role in life history evolution? Evol Dev. 5:372–378.
Pasquinelli AE, Reinhart BJ, Slack F, et al. (19 co-authors). 2000.Conservation of the sequence and temporal expression of let-7heterochronic regulatory RNA. Nature. 408:86–89.
Plasterk RH. 2006. Micro RNAs in animal development. Cell.124:877–881.
Reinhart BJ, Slack FJ, Basson M, Pasquinelli AE, Bettinger JC,Rougvie AE, Horvitz HR, Ruvkun G. 2000. The 21-nucleotidelet-7 RNA regulates developmental timing in Caenorhabditiselegans. Nature. 403:901–906.
Schulman BR, Esquela-Kerscher A, Slack FJ. 2005. Reciprocalexpression of lin-41 and the microRNAs let-7 and mir-125during mouse embryogenesis. Dev Dyn. 234:1046–1054.
Sethupathy P, Corda B, Hatzigeorgiou AG. 2006. TarBase:a comprehensive database of experimentally supported animalmicroRNA targets. RNA. 12:192–197.
Slack FJ, Basson M, Liu Z, Ambros V, Horvitz HR, Ruvkun G.2000. The lin-41 RBCC gene acts in the C. elegansheterochronic pathway between the let-7 regulatory RNAand the LIN-29 transcription factor. Mol Cell. 5:659–669.
Stein LD, Bao Z, Blasiar D, et al. (36 co-authors). 2003. Thegenome sequence of Caenorhabditis briggsae: a platform forcomparative genomics. PLoS Biol. 1:E45.
Valencia-Sanchez MA, Liu J, Hannon GJ, Parker R. 2006.Control of translation and mRNA degradation by miRNAsand siRNAs. Genes Dev. 20:515–524.
Vella MC, Choi EY, Lin SY, Reinert K, Slack FJ. 2004. The C.elegans microRNA let-7 binds to imperfect let-7 complemen-tary sites from the lin-41 3#UTR. Genes Dev. 18:132–137.
Waterston R, Lindblad-Toh HK, Birney E, et al. (222 co-authors). 2002. Initial sequencing and comparative analysis ofthe mouse genome. Nature. 420:520–562.
Yeh WH. 1991. Genes acting late in the signalling pathway forCaenorhabditis elegans dauer larval development. [Ph.D.thesis]. Columbia (MO): University of Missouri.
Takashi Gojobori, Associate Editor
Accepted September 3, 2007
2534 Lin et al.
by guest on June 2, 2013http://m
be.oxfordjournals.org/D
ownloaded from
Related Documents











