Human Semilunar Cardiac Valve Remodeling by Activated Cells From Fetus to Adult Implications for Postnatal Adaptation, Pathology, and Tissue Engineering Elena Aikawa, MD, PhD; Peter Whittaker, PhD; Mark Farber, MS; Karen Mendelson, MS; Robert F. Padera, MD, PhD; Masanori Aikawa, MD, PhD; Frederick J. Schoen, MD, PhD Background—The evolution of cell phenotypes and matrix architecture in cardiac valves during fetal maturation and postnatal adaptation through senescence remains unexplored. Methods and Results—We hypothesized that valvular interstitial (VIC) and endothelial cell (VEC) phenotypes, critical for maintaining valve function, change throughout life in response to environmental stimuli. We performed quantitative histological assessment of 91 human semilunar valves obtained from fetuses at 14 to 19 and 20 to 39 weeks’ gestation; neonates minutes to 30 days old; children aged 2 to 16 years; and adults. A trilaminar architecture appeared by 36 weeks of gestation but remained rudimentary compared with that of adult valves. VECs expressed an activated phenotype throughout fetal development. VIC density, proliferation, and apoptosis were significantly higher in fetal than adult valves. Pulmonary and aortic fetal VICs showed an activated myofibroblast-like phenotype (-actin expression), abundant embryonic myosin, and matrix metalloproteinase-collagenases, which indicates an immature/activated phenotype engaged in matrix remodeling versus a quiescent fibroblast-like phenotype in adults. At birth, the abrupt change from fetal to neonatal circulation was associated with a greater number of -actin–positive VICs in neonatal aortic versus pulmonary valves. Collagen content increased from early to late fetal stages but was subsequently unchanged, whereas elastin significantly increased postnatally. Collagen fiber color analysis revealed a progressive temporal decrease in thin fibers and a corresponding increase in thick fibers. Additionally, collagen fibers were more aligned in adult than fetal valves. Conclusions—Fetal valves possess a dynamic/adaptive structure and contain cells with an activated/immature phenotype. During postnatal life, activated cells gradually become quiescent, whereas collagen matures, which suggests a progressive, environmentally mediated adaptation. (Circulation. 2006;113:1344-1352.) Key Words: cells collagen remodeling tissue engineering valves A n understanding of extracellular matrix (ECM) architec- ture and cellular changes that occur in cardiac valves during fetal development, maturation, and aging would pro- vide mechanistic insights into the pathogenesis of congenital and acquired valve abnormalities, aid assessment of thera- peutic strategies for valve disease, and assist in the develop- ment of regenerative and tissue-engineered approaches to valve repair and replacement. Although the mechanisms of valvulogenesis in early cardiac development have been elu- cidated, 1–5 subsequent cellular and ECM maturation, remod- eling, and growth processes, particularly in humans, remain unexplored. ECM in general and collagen in particular play a crucial role in valve function and durability. Indeed, valves must accommodate substantial changes in hemodynamic environ- ment and structure throughout a lifetime. Furthermore, val- vular interstitial (VIC) and endothelial cell (VEC) functions likely influence ECM synthesis and remodeling. For exam- ple, activated myofibroblast-like VICs are critical to collagen metabolism and are altered in some valve diseases, including myxomatous degeneration of the mitral valve. 6 We recently showed that large populations of VICs undergo phenotypic modulation to become activated myofibroblasts and return to quiescent fibroblasts during adaptive remodeling in response to changing environmental conditions, which we have ob- served in long-term pulmonary autografts and in tissue- engineered valves. 7–9 We therefore tested the hypothesis that phenotypic changes in valvular cells, determined by altered Received September 27, 2005; revision received December 24, 2005; accepted January 9, 2006. From the Department of Pathology (E.A., M.F., K.M., R.F.P., F.J.S.) and Cardiovascular Division, Department of Medicine (M.A.), Brigham and Women’s Hospital, Harvard Medical School, Boston, Mass; Departments of Emergency Medicine and Anesthesiology (P.W.), University of Massachusetts Medical School, Worcester, Mass; and Center for Molecular Imaging Research (E.A.), Massachusetts General Hospital, Boston, Mass. Guest Editor for this article was William C. Roberts, MD. Correspondence to Frederick J. Schoen, MD, PhD, Department of Pathology, Brigham and Women’s Hospital, 75 Francis St, Boston, MA 02115 (e-mail [email protected]), or Elena Aikawa, MD, PhD, Center for Molecular Imaging Research, Massachusetts General Hospital, 149 13th St, Charlestown, MA 02129 (e-mail [email protected]). © 2006 American Heart Association, Inc. Circulation is available at http://www.circulationaha.org DOI: 10.1161/CIRCULATIONAHA.105.591768 1344 Valvular Heart Disease by guest on July 20, 2015 http://circ.ahajournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Human Semilunar Cardiac Valve Remodeling by ActivatedCells From Fetus to Adult
Implications for Postnatal Adaptation, Pathology, and Tissue Engineering
Elena Aikawa, MD, PhD; Peter Whittaker, PhD; Mark Farber, MS; Karen Mendelson, MS;Robert F. Padera, MD, PhD; Masanori Aikawa, MD, PhD; Frederick J. Schoen, MD, PhD
Background—The evolution of cell phenotypes and matrix architecture in cardiac valves during fetal maturation andpostnatal adaptation through senescence remains unexplored.
Methods and Results—We hypothesized that valvular interstitial (VIC) and endothelial cell (VEC) phenotypes, critical formaintaining valve function, change throughout life in response to environmental stimuli. We performed quantitativehistological assessment of 91 human semilunar valves obtained from fetuses at 14 to 19 and 20 to 39 weeks’ gestation;neonates minutes to 30 days old; children aged 2 to 16 years; and adults. A trilaminar architecture appeared by 36 weeksof gestation but remained rudimentary compared with that of adult valves. VECs expressed an activated phenotypethroughout fetal development. VIC density, proliferation, and apoptosis were significantly higher in fetal than adultvalves. Pulmonary and aortic fetal VICs showed an activated myofibroblast-like phenotype (�-actin expression),abundant embryonic myosin, and matrix metalloproteinase-collagenases, which indicates an immature/activatedphenotype engaged in matrix remodeling versus a quiescent fibroblast-like phenotype in adults. At birth, the abruptchange from fetal to neonatal circulation was associated with a greater number of �-actin–positive VICs in neonatalaortic versus pulmonary valves. Collagen content increased from early to late fetal stages but was subsequentlyunchanged, whereas elastin significantly increased postnatally. Collagen fiber color analysis revealed a progressivetemporal decrease in thin fibers and a corresponding increase in thick fibers. Additionally, collagen fibers were morealigned in adult than fetal valves.
Conclusions—Fetal valves possess a dynamic/adaptive structure and contain cells with an activated/immature phenotype.During postnatal life, activated cells gradually become quiescent, whereas collagen matures, which suggests aprogressive, environmentally mediated adaptation. (Circulation. 2006;113:1344-1352.)
Key Words: cells � collagen � remodeling � tissue engineering � valves
An understanding of extracellular matrix (ECM) architec-ture and cellular changes that occur in cardiac valves
during fetal development, maturation, and aging would pro-vide mechanistic insights into the pathogenesis of congenitaland acquired valve abnormalities, aid assessment of thera-peutic strategies for valve disease, and assist in the develop-ment of regenerative and tissue-engineered approaches tovalve repair and replacement. Although the mechanisms ofvalvulogenesis in early cardiac development have been elu-cidated,1–5 subsequent cellular and ECM maturation, remod-eling, and growth processes, particularly in humans, remainunexplored.
ECM in general and collagen in particular play a crucialrole in valve function and durability. Indeed, valves must
accommodate substantial changes in hemodynamic environ-ment and structure throughout a lifetime. Furthermore, val-vular interstitial (VIC) and endothelial cell (VEC) functionslikely influence ECM synthesis and remodeling. For exam-ple, activated myofibroblast-like VICs are critical to collagenmetabolism and are altered in some valve diseases, includingmyxomatous degeneration of the mitral valve.6 We recentlyshowed that large populations of VICs undergo phenotypicmodulation to become activated myofibroblasts and return toquiescent fibroblasts during adaptive remodeling in responseto changing environmental conditions, which we have ob-served in long-term pulmonary autografts and in tissue-engineered valves.7–9 We therefore tested the hypothesis thatphenotypic changes in valvular cells, determined by altered
Received September 27, 2005; revision received December 24, 2005; accepted January 9, 2006.From the Department of Pathology (E.A., M.F., K.M., R.F.P., F.J.S.) and Cardiovascular Division, Department of Medicine (M.A.), Brigham and
Women’s Hospital, Harvard Medical School, Boston, Mass; Departments of Emergency Medicine and Anesthesiology (P.W.), University ofMassachusetts Medical School, Worcester, Mass; and Center for Molecular Imaging Research (E.A.), Massachusetts General Hospital, Boston, Mass.
Guest Editor for this article was William C. Roberts, MD.Correspondence to Frederick J. Schoen, MD, PhD, Department of Pathology, Brigham and Women’s Hospital, 75 Francis St, Boston, MA 02115
(e-mail [email protected]), or Elena Aikawa, MD, PhD, Center for Molecular Imaging Research, Massachusetts General Hospital, 149 13th St,Charlestown, MA 02129 (e-mail [email protected]).
© 2006 American Heart Association, Inc.
Circulation is available at http://www.circulationaha.org DOI: 10.1161/CIRCULATIONAHA.105.591768
1344
Valvular Heart Disease
by guest on July 20, 2015http://circ.ahajournals.org/Downloaded from

expression of cytoskeletal and surface proteins and by pro-teolytic enzymes, regulate age-associated structural remodel-ing and adaptation of the ECM. Specifically, we character-ized phenotypes and turnover of VIC and VEC in humanfetal, neonatal, child, and adult semilunar valves and corre-lated cellular changes with alterations in valvular ECM.
MethodsMorphological CharacterizationWe studied 91 human semilunar valves: fetal valves in the secondtrimester (14 to 19 weeks’ gestation; n�11) and third trimester (20to 39 weeks’ gestation; aortic n�10, pulmonary n�10); neonatal(early neonatal with minutes to hours of survival: aortic n�5,pulmonary n�5; late neonatal [up to 30 days of survival]: aorticn�5, pulmonary n�5); valves from children (aged 6.0�1.6 years;aortic n�10, pulmonary n�10); and valves from normal adults (aged50.1�2.5 years; aortic n�10, pulmonary n�10) obtained at autopsyaccording to a protocol for tissue use approved by the HumanResearch Committee at Brigham and Women’s Hospital. No patienthad documented cardiac disease or conditions known to predisposeto heart valve disease. Examination revealed no evidence of valvedisease, and the fetal heart morphology appeared appropriate for thegestational age.
Valves were fixed in 10% buffered formalin, cut radially in thecentral cusp region through the adjacent arterial wall, and embeddedin paraffin. Sections cut at 6-�m thickness were stained withhematoxylin and eosin for general morphological assessment. Weused Movat pentachrome stain to differentiate connective tissueelements (collagen, elastin, and proteoglycans) and picrosirius redviewed with circularly polarized light to assess collagen architec-ture.6,10 All 3 cusps from each aortic and pulmonary valve wereanalyzed.
Characterization of Cell Phenotype and TurnoverMarkers of cell phenotype and synthetic products were identified byimmunohistochemistry. We defined myofibroblasts like VICs ascells that had antibody reaction to �-smooth muscle actin (�-SMA,1A4, Dako, Carpenteria, Calif).6,11 Mouse monoclonal antibodiesagainst human metalloproteinase-1 (MMP-1/collagenase-1) andMMP-13/collagenase-3 (Calbiochem, San Diego, Calif) were used todetermine proteolytic enzyme expression. Cell activation was alsodemonstrated by a monoclonal antibody against human SMemb12 (anonmuscle myosin also known as MHC-B, expressed by embryonicor activated mesenchymal cells). Phenotypic changes in valvularendothelium were assessed by antibodies to CD31 (Dako), intercel-lular adhesion molecule-1 (ICAM-1; US Biological, Swampscott,Mass), and vascular cell adhesion molecule-1 (VCAM-1; US Bio-logical). Immunohistochemistry methods used avidin-biotin-peroxidase after antigen unmasking pretreatment with proteinase Kfor 5 minutes at room temperature. Adjacent sections treated withnonimmune IgG provided controls for antibody specificity. Cellproliferation was demonstrated with the Ki-67 (MIB-1, Dako)antigen, which is present during active phases of the cell cycle butabsent in resting cells, and cell apoptosis was demonstrated byterminal dUTP nick end-labeling (TUNEL) assay (Intergen, NewYork, NY). Commercially available human lymph node and thymussections provided positive controls. Images were captured andanalyzed with a digital camera (Nikon DXM1200-F, Nikon Inc,Melville, NY) and imaging software (ACT-1 version 2.63, Nikon).
Cell density was measured as mean number of cells per 10high-power fields (�400) and expressed as cell number per squaremillimeter of the tissue section in each cusp. Cell proliferation andapoptosis indices were calculated as the ratio of Ki-67 and TUNEL-positive cells, respectively, to the total number of valve cusp cellnuclei. The ratio of �-SMA, MMP-1, MMP-13, and SMemb-positivecells to the total cell number was averaged over 5 representativehigh-power fields. The ratio of ICAM-1– and VCAM-1–positivecells to total CD31-positive cells was calculated for each cusp. Onlycells expressing an antigen of interest (defined as red reaction
product associated with blue hematoxylin nuclear counterstain) werecounted. All measurements were done by 2 independent researchers.
Collagen and Elastin AssessmentWe used picrosirius red staining in conjunction with polarized lightmicroscopy to assess collagen fiber content, thickness, and organi-zation and Movat pentachrome to detect elastin (Olympus BX51microscope, Olympus America Inc, Melville, NY).
Collagen Content and Fiber ColorWith this microscopy combination, collagen fiber color is deter-mined by thickness; the color changes from green to yellow toorange to red as thickness increases.13 We assessed the proportion ofdifferent colored fibers using published methods.14,15 Briefly, werecorded images viewed with circularly polarized light16 using adigital camera (DP11, Olympus) and analyzed 2 regions (260�200�m) per valve. Images were resolved, using an automated softwarefunction (SigmaScan Pro; SPSS Inc, Chicago, Ill), into their hue,saturation, and value components. We retained only the hue compo-nent, and a histogram of hue frequency was obtained from the 8-bithue images that contained 256 possible colors, defined as follows;red 2 to 9 and 230 to 256, orange 10 to 38, yellow 39 to 51, and green52 to 128.14 All other hue values corresponded to interstitial space,confirmed by inspection. We determined the number of pixels withineach hue range and expressed this as a percentage of the total numberof collagen pixels, which in turn was expressed as a percentage of thetotal number of pixels in the regions analyzed to give collagencontent.
Collagen OrientationThe optical properties of birefringent materials, such as collagen, canbe exploited to determine their 2D orientation.17 We used a recentlydeveloped automated system (PolScope, CRI, Inc, Woburn, Mass) tomeasure collagen fiber orientation at 100 locations in each valve,with locations selected by construction of a 10�5-point rectangulargrid overlying 2 regions in each valve. An orientation frequencydistribution was obtained for each region. Briefly, the system uses anelectronically controlled liquid crystal compensator to acquire andrecord images, with a CCD camera, at 4 predetermined compensatorsettings. Algorithms are used to calculate the orientation angle of theslow optical axis of the birefringent collagen from the data obtainedin these images18 given the property that the slow optical axis ofcollagen corresponds to the long-axis orientation of the fibers. Eachdistribution was analyzed with circular statistics, which are methodsdesigned to examine directional data.19 The degree of fiber align-ment was assessed by calculation of the angular deviation of thedistribution, the circular statistics’ equivalent of standard deviation;the smaller the angular deviation, the more aligned the fibers.17
Elastin ContentMovat pentachrome stains elastin black, collagen yellow, and glycos-aminoglycans blue-green. Elastin-positive areas were measured in 3high-power fields of each cusp in the ventricularis layer with imagingsoftware (IPLab version 3.9.3; Scanalytics, Inc, Rockville, Md).
Statistical AnalysesData are presented as mean�SEM. Group differences were evalu-ated with ANOVA followed by pairwise comparisons with theTukey-Kramer post hoc test; however, t tests were used when 2groups were compared. Probability values less than 0.05 wereconsidered significant.
The authors had full access to the data and take full responsibilityfor its integrity. All authors have read and agree to the manuscript aswritten.
ResultsFetal Valves Have an IncompleteMorphological DifferentiationAt 14 weeks of gestational age, the ECM of fetal valves waspredominantly composed of glycosaminoglycans, as indi-
Aikawa et al Human Cardiac Valve Maturation 1345
by guest on July 20, 2015http://circ.ahajournals.org/Downloaded from
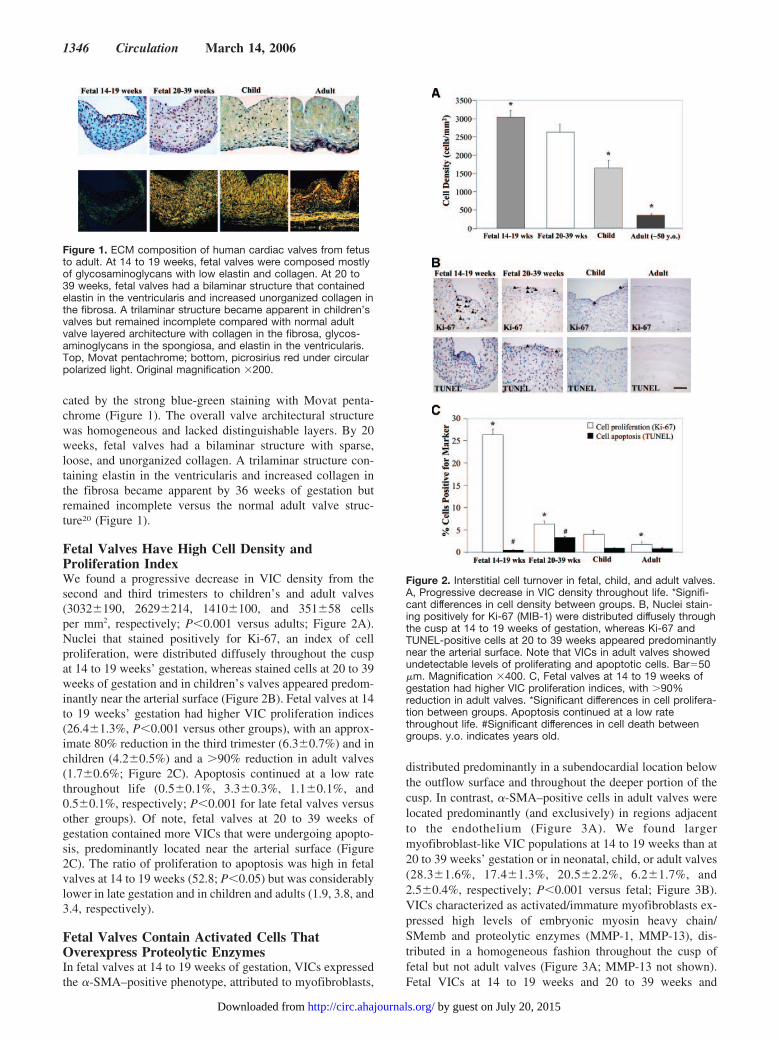
cated by the strong blue-green staining with Movat penta-chrome (Figure 1). The overall valve architectural structurewas homogeneous and lacked distinguishable layers. By 20weeks, fetal valves had a bilaminar structure with sparse,loose, and unorganized collagen. A trilaminar structure con-taining elastin in the ventricularis and increased collagen inthe fibrosa became apparent by 36 weeks of gestation butremained incomplete versus the normal adult valve struc-ture20 (Figure 1).
Fetal Valves Have High Cell Density andProliferation IndexWe found a progressive decrease in VIC density from thesecond and third trimesters to children’s and adult valves(3032�190, 2629�214, 1410�100, and 351�58 cellsper mm2, respectively; P�0.001 versus adults; Figure 2A).Nuclei that stained positively for Ki-67, an index of cellproliferation, were distributed diffusely throughout the cuspat 14 to 19 weeks’ gestation, whereas stained cells at 20 to 39weeks of gestation and in children’s valves appeared predom-inantly near the arterial surface (Figure 2B). Fetal valves at 14to 19 weeks’ gestation had higher VIC proliferation indices(26.4�1.3%, P�0.001 versus other groups), with an approx-imate 80% reduction in the third trimester (6.3�0.7%) and inchildren (4.2�0.5%) and a �90% reduction in adult valves(1.7�0.6%; Figure 2C). Apoptosis continued at a low ratethroughout life (0.5�0.1%, 3.3�0.3%, 1.1�0.1%, and0.5�0.1%, respectively; P�0.001 for late fetal valves versusother groups). Of note, fetal valves at 20 to 39 weeks ofgestation contained more VICs that were undergoing apopto-sis, predominantly located near the arterial surface (Figure2C). The ratio of proliferation to apoptosis was high in fetalvalves at 14 to 19 weeks (52.8; P�0.05) but was considerablylower in late gestation and in children and adults (1.9, 3.8, and3.4, respectively).
Fetal Valves Contain Activated Cells ThatOverexpress Proteolytic EnzymesIn fetal valves at 14 to 19 weeks of gestation, VICs expressedthe �-SMA–positive phenotype, attributed to myofibroblasts,
distributed predominantly in a subendocardial location belowthe outflow surface and throughout the deeper portion of thecusp. In contrast, �-SMA–positive cells in adult valves werelocated predominantly (and exclusively) in regions adjacentto the endothelium (Figure 3A). We found largermyofibroblast-like VIC populations at 14 to 19 weeks than at20 to 39 weeks’ gestation or in neonatal, child, or adult valves(28.3�1.6%, 17.4�1.3%, 20.5�2.2%, 6.2�1.7%, and2.5�0.4%, respectively; P�0.001 versus fetal; Figure 3B).VICs characterized as activated/immature myofibroblasts ex-pressed high levels of embryonic myosin heavy chain/SMemb and proteolytic enzymes (MMP-1, MMP-13), dis-tributed in a homogeneous fashion throughout the cusp offetal but not adult valves (Figure 3A; MMP-13 not shown).Fetal VICs at 14 to 19 weeks and 20 to 39 weeks and
Figure 1. ECM composition of human cardiac valves from fetusto adult. At 14 to 19 weeks, fetal valves were composed mostlyof glycosaminoglycans with low elastin and collagen. At 20 to39 weeks, fetal valves had a bilaminar structure that containedelastin in the ventricularis and increased unorganized collagen inthe fibrosa. A trilaminar structure became apparent in children’svalves but remained incomplete compared with normal adultvalve layered architecture with collagen in the fibrosa, glycos-aminoglycans in the spongiosa, and elastin in the ventricularis.Top, Movat pentachrome; bottom, picrosirius red under circularpolarized light. Original magnification �200.
Figure 2. Interstitial cell turnover in fetal, child, and adult valves.A, Progressive decrease in VIC density throughout life. *Signifi-cant differences in cell density between groups. B, Nuclei stain-ing positively for Ki-67 (MIB-1) were distributed diffusely throughthe cusp at 14 to 19 weeks of gestation, whereas Ki-67 andTUNEL-positive cells at 20 to 39 weeks appeared predominantlynear the arterial surface. Note that VICs in adult valves showedundetectable levels of proliferating and apoptotic cells. Bar�50�m. Magnification �400. C, Fetal valves at 14 to 19 weeks ofgestation had higher VIC proliferation indices, with �90%reduction in adult valves. *Significant differences in cell prolifera-tion between groups. Apoptosis continued at a low ratethroughout life. #Significant differences in cell death betweengroups. y.o. indicates years old.
1346 Circulation March 14, 2006
by guest on July 20, 2015http://circ.ahajournals.org/Downloaded from

children’s VICs had larger numbers of SMemb-, MMP-1–,and MMP-13–positive cells versus the negligible numbers ofVICs that expressed these proteins in adult valves (SMemb80.9�3.1%, 75.2�4.1%, and 38�4.5% versus 7�1.0%;MMP-1 45.7�5.9%, 65.9�6.7%, and 61�5.6% versus5.5�0.7%; MMP-13 18.5�2.5%, 47.3�4.8%, and26.0�3.1% versus 3.5�0.7%, respectively; P�0.001 versusadult; Figure 3B).
At birth, after separation of the single fetal circulation intoisolated pulmonary and systemic circulations, pulmonaryarterial pressure decreases and aortic pressure increases.21
These hemodynamic changes were associated with decreasedVIC activation in late neonatal (1 to 30 days of survival)pulmonary valves (6.3�0.8%; n�5), whereas aortic valveVICs remained activated (20.5�2.2%, n�5; P�0.05; Figure4). Nevertheless, this change did not occur immediately,because the numbers of activated VICs in early neonatalvalves with minutes to hours of survival were identical inboth pulmonary (20.5%; n�5) and aortic valves (20.5%;n�5). Activated VICs continued to decrease after adaptation,because valves from individuals aged 2 to 16 years hadsimilar numbers of activated VICs in pulmonary (6%; n�10)and aortic (6%; n�10) valves (Figure 4). Fetal VECs in thesecond and third trimesters and children’s valves consistentlyexpressed the cell adhesion molecules ICAM-1 (99.0�7.2%,98.7�4.6%, and 47.6�9.5% versus 5.0�0.5%) andVCAM-1 (54.0�9.4%, 68.2�13.9%, and 16.0�3.7% versus
0%), as well as SMemb (99.2�1.5%, 99.3�0.5%, and71.3�10.3% versus 1.0�0.5%), MMP-1 (99.3�0.8%,97.0�1.3%, and 71.7�9.1% versus 1.0�0.5%), andMMP-13 (98.4�2.5%, 95.3�1.7%, and 43.1�7.0% versus0.9�0.3%), whereas normal adult VECs exhibited negligibleexpression levels of these proteins (P�0.001 fetal versusadult; Figure 5A and 5B).
Figure 3. Activated interstitial cells undergo evolution to a morequiescent phenotype during postnatal life. A, Larger VIC popula-tions at 14 to 19 weeks expressed the �-SMA–positive pheno-type, attributed to myofibroblasts and high levels of SMemb andMMP-1, markers of activation, compared with negligible expres-sion of these proteins in adult valves. Bar�50 �m. Magnification�400. B, Percent of interstitial cells positive for marker. *Signifi-cant differences in protein expression in adult valves. y.o. indi-cates years old.
Figure 4. Changes in VIC activation at birth. Abrupt hemody-namic changes at birth were associated with decreased VICactivation in late neonatal (days of survival) pulmonary valves(PV), whereas aortic valve (AV) VICs remained activated(P�0.05). Numbers of activated VICs in early neonatal valves(minutes to hours of survival) were identical in both pulmonaryand aortic valves, which suggests that this change did notoccur immediately.
Figure 5. Endothelial cells of fetal valves have an immature/acti-vated phenotype. A, CD31-positive fetal VECs in the secondand third trimesters and VECs in children’s valves consistentlyexpressed SMemb, MMP-1, MMP-13, and cell adhesion mole-cules ICAM-1 and VCAM-1, whereas normal adult VECs mostlyhad negligible expression levels of these proteins. Bar�50 �m.Magnification �400. B, Percent of endothelial cells positive formaker. *Significant differences in protein expression in adultvalves (P�0.001).
Aikawa et al Human Cardiac Valve Maturation 1347
by guest on July 20, 2015http://circ.ahajournals.org/Downloaded from
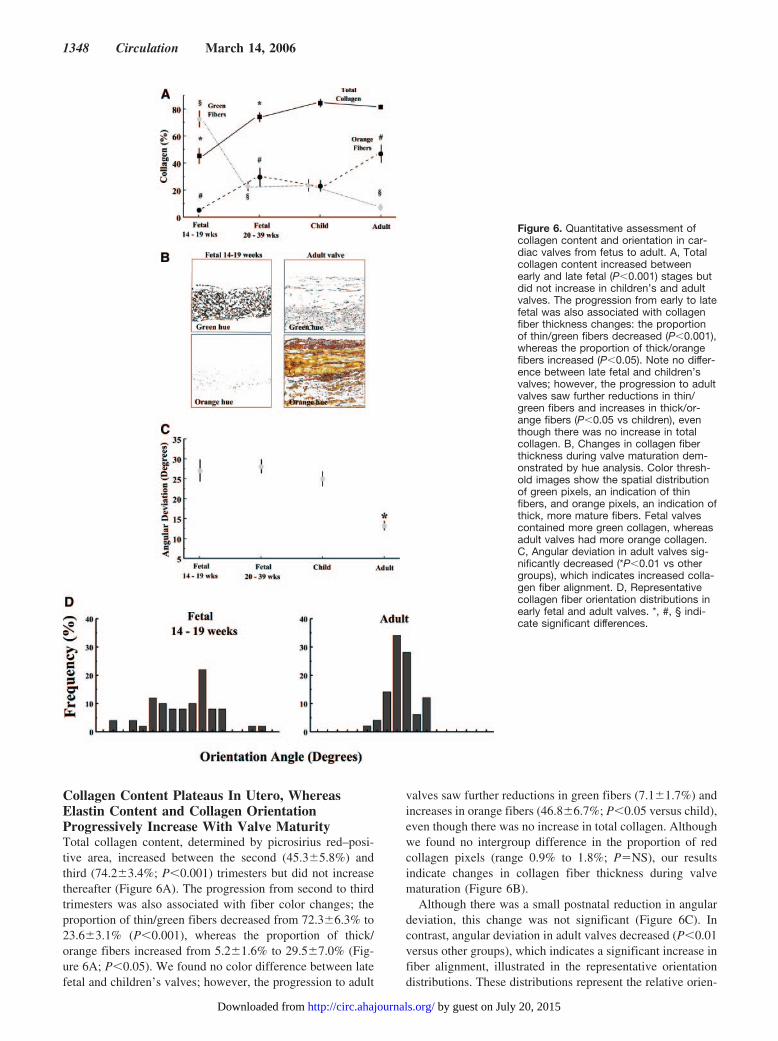
Collagen Content Plateaus In Utero, WhereasElastin Content and Collagen OrientationProgressively Increase With Valve MaturityTotal collagen content, determined by picrosirius red–posi-tive area, increased between the second (45.3�5.8%) andthird (74.2�3.4%; P�0.001) trimesters but did not increasethereafter (Figure 6A). The progression from second to thirdtrimesters was also associated with fiber color changes; theproportion of thin/green fibers decreased from 72.3�6.3% to23.6�3.1% (P�0.001), whereas the proportion of thick/orange fibers increased from 5.2�1.6% to 29.5�7.0% (Fig-ure 6A; P�0.05). We found no color difference between latefetal and children’s valves; however, the progression to adult
valves saw further reductions in green fibers (7.1�1.7%) andincreases in orange fibers (46.8�6.7%; P�0.05 versus child),even though there was no increase in total collagen. Althoughwe found no intergroup difference in the proportion of redcollagen pixels (range 0.9% to 1.8%; P�NS), our resultsindicate changes in collagen fiber thickness during valvematuration (Figure 6B).
Although there was a small postnatal reduction in angulardeviation, this change was not significant (Figure 6C). Incontrast, angular deviation in adult valves decreased (P�0.01versus other groups), which indicates a significant increase infiber alignment, illustrated in the representative orientationdistributions. These distributions represent the relative orien-
Figure 6. Quantitative assessment ofcollagen content and orientation in car-diac valves from fetus to adult. A, Totalcollagen content increased betweenearly and late fetal (P�0.001) stages butdid not increase in children’s and adultvalves. The progression from early to latefetal was also associated with collagenfiber thickness changes: the proportionof thin/green fibers decreased (P�0.001),whereas the proportion of thick/orangefibers increased (P�0.05). Note no differ-ence between late fetal and children’svalves; however, the progression to adultvalves saw further reductions in thin/green fibers and increases in thick/or-ange fibers (P�0.05 vs children), eventhough there was no increase in totalcollagen. B, Changes in collagen fiberthickness during valve maturation dem-onstrated by hue analysis. Color thresh-old images show the spatial distributionof green pixels, an indication of thinfibers, and orange pixels, an indication ofthick, more mature fibers. Fetal valvescontained more green collagen, whereasadult valves had more orange collagen.C, Angular deviation in adult valves sig-nificantly decreased (*P�0.01 vs othergroups), which indicates increased colla-gen fiber alignment. D, Representativecollagen fiber orientation distributions inearly fetal and adult valves. *, #, § indi-cate significant differences.
1348 Circulation March 14, 2006
by guest on July 20, 2015http://circ.ahajournals.org/Downloaded from
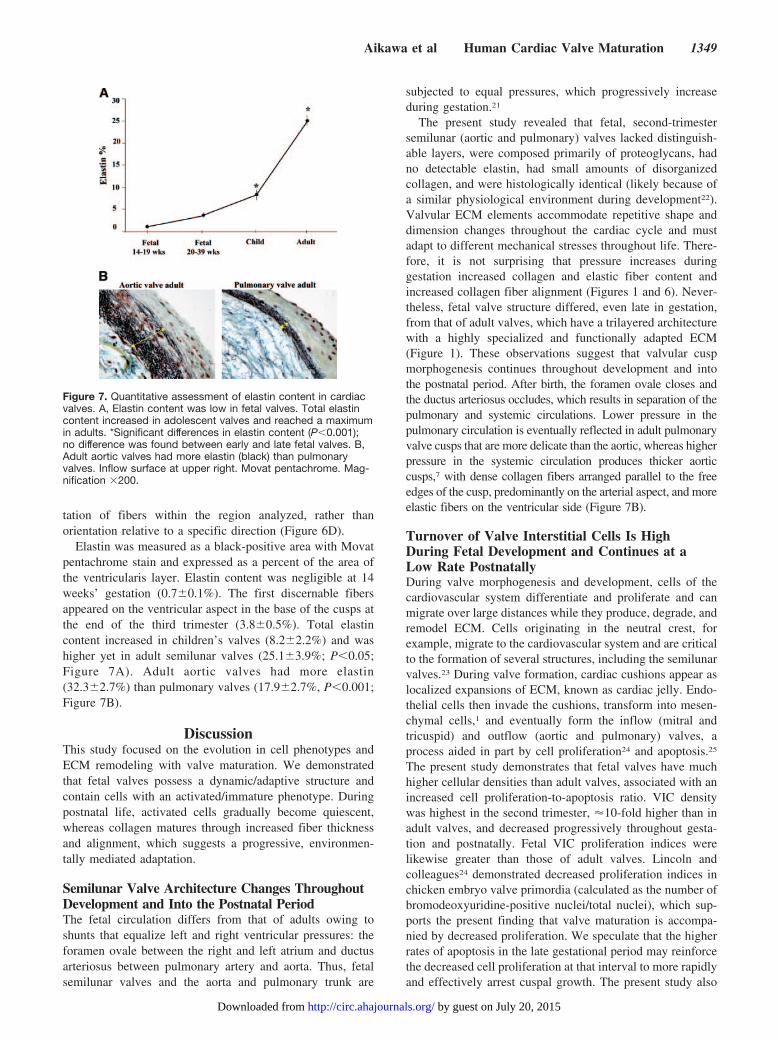
tation of fibers within the region analyzed, rather thanorientation relative to a specific direction (Figure 6D).
Elastin was measured as a black-positive area with Movatpentachrome stain and expressed as a percent of the area ofthe ventricularis layer. Elastin content was negligible at 14weeks’ gestation (0.7�0.1%). The first discernable fibersappeared on the ventricular aspect in the base of the cusps atthe end of the third trimester (3.8�0.5%). Total elastincontent increased in children’s valves (8.2�2.2%) and washigher yet in adult semilunar valves (25.1�3.9%; P�0.05;Figure 7A). Adult aortic valves had more elastin(32.3�2.7%) than pulmonary valves (17.9�2.7%, P�0.001;Figure 7B).
DiscussionThis study focused on the evolution in cell phenotypes andECM remodeling with valve maturation. We demonstratedthat fetal valves possess a dynamic/adaptive structure andcontain cells with an activated/immature phenotype. Duringpostnatal life, activated cells gradually become quiescent,whereas collagen matures through increased fiber thicknessand alignment, which suggests a progressive, environmen-tally mediated adaptation.
Semilunar Valve Architecture Changes ThroughoutDevelopment and Into the Postnatal PeriodThe fetal circulation differs from that of adults owing toshunts that equalize left and right ventricular pressures: theforamen ovale between the right and left atrium and ductusarteriosus between pulmonary artery and aorta. Thus, fetalsemilunar valves and the aorta and pulmonary trunk are
subjected to equal pressures, which progressively increaseduring gestation.21
The present study revealed that fetal, second-trimestersemilunar (aortic and pulmonary) valves lacked distinguish-able layers, were composed primarily of proteoglycans, hadno detectable elastin, had small amounts of disorganizedcollagen, and were histologically identical (likely because ofa similar physiological environment during development22).Valvular ECM elements accommodate repetitive shape anddimension changes throughout the cardiac cycle and mustadapt to different mechanical stresses throughout life. There-fore, it is not surprising that pressure increases duringgestation increased collagen and elastic fiber content andincreased collagen fiber alignment (Figures 1 and 6). Never-theless, fetal valve structure differed, even late in gestation,from that of adult valves, which have a trilayered architecturewith a highly specialized and functionally adapted ECM(Figure 1). These observations suggest that valvular cuspmorphogenesis continues throughout development and intothe postnatal period. After birth, the foramen ovale closes andthe ductus arteriosus occludes, which results in separation of thepulmonary and systemic circulations. Lower pressure in thepulmonary circulation is eventually reflected in adult pulmonaryvalve cusps that are more delicate than the aortic, whereas higherpressure in the systemic circulation produces thicker aorticcusps,7 with dense collagen fibers arranged parallel to the freeedges of the cusp, predominantly on the arterial aspect, and moreelastic fibers on the ventricular side (Figure 7B).
Turnover of Valve Interstitial Cells Is HighDuring Fetal Development and Continues at aLow Rate PostnatallyDuring valve morphogenesis and development, cells of thecardiovascular system differentiate and proliferate and canmigrate over large distances while they produce, degrade, andremodel ECM. Cells originating in the neutral crest, forexample, migrate to the cardiovascular system and are criticalto the formation of several structures, including the semilunarvalves.23 During valve formation, cardiac cushions appear aslocalized expansions of ECM, known as cardiac jelly. Endo-thelial cells then invade the cushions, transform into mesen-chymal cells,1 and eventually form the inflow (mitral andtricuspid) and outflow (aortic and pulmonary) valves, aprocess aided in part by cell proliferation24 and apoptosis.25
The present study demonstrates that fetal valves have muchhigher cellular densities than adult valves, associated with anincreased cell proliferation-to-apoptosis ratio. VIC densitywas highest in the second trimester, �10-fold higher than inadult valves, and decreased progressively throughout gesta-tion and postnatally. Fetal VIC proliferation indices werelikewise greater than those of adult valves. Lincoln andcolleagues24 demonstrated decreased proliferation indices inchicken embryo valve primordia (calculated as the number ofbromodeoxyuridine-positive nuclei/total nuclei), which sup-ports the present finding that valve maturation is accompa-nied by decreased proliferation. We speculate that the higherrates of apoptosis in the late gestational period may reinforcethe decreased cell proliferation at that interval to more rapidlyand effectively arrest cuspal growth. The present study also
Figure 7. Quantitative assessment of elastin content in cardiacvalves. A, Elastin content was low in fetal valves. Total elastincontent increased in adolescent valves and reached a maximumin adults. *Significant differences in elastin content (P�0.001);no difference was found between early and late fetal valves. B,Adult aortic valves had more elastin (black) than pulmonaryvalves. Inflow surface at upper right. Movat pentachrome. Mag-nification �200.
Aikawa et al Human Cardiac Valve Maturation 1349
by guest on July 20, 2015http://circ.ahajournals.org/Downloaded from

demonstrates that valvular cell turnover is high during fetaldevelopment and continues at a low rate postnatally (Figure2C) and that the total number of cells decreases substantiallythroughout life (Figure 2A), a fact that might account forslower or incomplete valve remodeling in older adults andpredisposition to clinically important valve degeneration.
Activated Myofibroblasts Mediate CollagenMetabolism During Valve MaturationWe also demonstrated VIC plasticity throughout gestationalvalve maturation, in contrast to the resting VIC population ofnormal adult valves and their predominantly fibroblast-likephenotype.8 We observed that a large number of fetal VICsexhibited an �-SMA–positive phenotype attributed tomyofibroblast-like cells (Figure 3). Diseased valves alsocontain numerous myofibroblast-like VICs that express�-SMA, which suggests a role for myofibroblasts in valvularconnective tissue remodeling.7,8 Moreover, we found thatfetal VICs had abundant SMemb (nonmuscle myosin pro-duced by embryonic or activated cells), MMP-1/collage-nase-1, and MMP-13/collagenase-3, which indicates an acti-vated/immature phenotype compatible with matrixremodeling. Despite the well-established role played byMMPs in pathological processes, including myxomatousmitral valve degeneration6 and atherosclerosis,10,15 their rolein remodeling normal valves and in fetal valve developmentremains unknown. We found more MMP-1 and MMP-13 infetal valves, particularly late in gestation, than in adult valves,whereas collagen content increased during valve maturation.This inverse relationship suggests a critical role for MMP-family collagenases in collagen metabolism in cardiac valvedevelopment. A significant increase in collagen fiber thick-ness and alignment occurred between childhood and adult-hood, whereas collagenase expression was negligible; how-ever, whether this is due to prolonged exposure to mechanicalforces or reflects completion of cell- and enzyme-associatedremodeling is unknown.
Activated Myofibroblasts Undergo PhenotypicModulation at Birth and Become a QuiescentFibroblast-Like Cell Type During AdulthoodPrevious studies reported that altered mechanical forces8,26
and injury27 are associated with phenotypic changes ofvalvular cells. Of particular interest in the present study wasthe way in which valves responded to abrupt change in bloodpressures and other hemodynamic conditions at birth afterseparation of the single fetal circulation into isolated pulmo-nary and systemic circulations. Therefore, we also focused oncomparison between fetal and neonatal pulmonary and aorticvalves. Blood pressure in the fetal aorta and pulmonary arteryis approximately 50/15 mm Hg. At birth, pulmonary pressurefalls to 30/15 mm Hg; however, arterial pressure increases to70/40 mm Hg. We found that the change from fetal toneonatal circulation correlated with reduced VIC activation(�6%) in neonatal pulmonary valves versus aortic valves(�21%; Figure 4).28 Whether these changes might also bestimulated by changes in local blood oxygen content isunknown. In children’s valves, the numbers of activated VICwere similar in both pulmonary and aortic locations, whichsuggests tissue adaptation to pulmonary and systemiccirculations.
Endothelial Cells Express an Activated PhenotypeThroughout Fetal Development, a PhenotypeDistinct From Adult ValvesObservations suggest that valvular endothelium is uniquecompared with other types of endothelial cells (particularlyaortic and cardiac microvascular endothelium).29 The endo-thelium regulates vascular tone, inflammation, thrombosis,and vascular remodeling.30 Structurally intact endothelialcells can respond to pathophysiological stimuli by adjustingtheir usual functions and by expressing newly acquiredproperties, a process termed “endothelial activation.” Acti-vated endothelial cells produce a variety of biologicallyactive products, including cytokines, growth factors, proteo-lytic enzymes, and adhesion molecules. Normal endothelial
Figure 8. Schematic representation ofnatural history of cardiac valve remodel-ing by interstitial cells. Schematic sum-mary of cardiac valve remodeling in fetalvalves according to the findings of thepresent study. Activated/immature cellphenotypes and collagen remodeling ingestation were followed after birth bydecreased cell activation and eventually,in adults, by quiescence. These cellularchanges were accompanied byincreased collagen fiber thickness andalignment. Thin lines represent thin,immature collagen fibers; thick lines rep-resent thick, mature collagen fibers.
1350 Circulation March 14, 2006
by guest on July 20, 2015http://circ.ahajournals.org/Downloaded from

function is characterized by a balance of these factors and thecells’ ability to respond appropriately to stimuli. The presentstudy demonstrated physiological activation of endothelialcells that consistently expressed high levels of SMemb,MMP-1, MMP-13, ICAM-1, and VCAM-1 in fetal andchildren’s valves. In contrast, adult valves had negligibleexpression of these molecules (Figure 5).
Clinical and experimental evidence suggests that valveleaflet endothelium is protected from inflammatory andimmune-mediated injury by reduced expression of adhesionmolecules, including VCAM-1, which is abundant in cardiacmicrovascular endothelium of rejected orthotopic heart trans-plants and in pig hearts after transplantation into baboons.31
The present study demonstrates consistent ICAM-1 andVCAM-1 expression by human fetal VECs during mid to lategestation. Furthermore, ICAM-1 and VCAM-1 expressionhas been found in surgically removed diseased valves32,33 andin vitro,34 which suggests that other factors, such as hemo-dynamic forces, may contribute to protection from inflamma-tory cell infiltration. Nevertheless, understanding endothelialadhesion molecule function in vivo in the presence ofcomplex blood flow patterns requires further investigation.
Study ImplicationsThe present study provides a natural history of cell and matrixchanges in valve development, maturation, and adaptation(summarized in Figure 8) and extends the paradigm, previ-ously suggested by us, that cardiac valvular tissue can adaptto environmental conditions, particularly mechanical load-ing.8 Specifically, under equilibrium conditions, VICs arequiescent and ECM is well adapted. We hypothesize thatwhen stimulated by mechanical loading, VICs become acti-vated and mediate connective tissue remodeling to restore anormal stress profile in the tissue. When equilibrium isrestored, cells return to quiescence. The present study dem-onstrates that fetal VIC activation occurs throughout devel-opment, analogous to the valve changes that occur in patho-logical conditions and after surgical substitution.6,7 Thus,analogous molecular mechanisms likely direct both physio-logical and pathological interstitial cell activation. Further-more, our characterization of human semilunar valve VIC andVEC phenotypes and dynamic ECM changes from fetus toadult also indicates potential markers of cell activation/maturation and matrix alterations (eg, MMP-1, MMP-13,embryonic myosin, VCAM-1, and ICAM-1). These mole-cules are overexpressed during fetal valve development,several valvular diseases, and surgical transplantation and intissue-engineered valves7–9 and therefore may represent tar-gets for molecular imaging to noninvasively monitor biolog-ical processes during healing and remodeling. Moreover, theprogressive age-associated decrease in cell number and activecapacity for remodeling may be an important clue in under-standing senile valve degeneration.
ConclusionsWe demonstrated that fetal valves possess a dynamic/adap-tive structure and contain activated/immature cells. More-over, the valvular cells that were activated in utero undergophenotypic changes at birth and gradually become quiescent,whereas collagen matures through increased fiber thickness
and alignment. This suggests a progressive adaptation to theprevailing hemodynamic environment. Knowledge of celland matrix changes during fetal valve maturation may aid inthe development of therapy for valve disease and assist in thedesign and evaluation of tissue-engineered valves.35–37
AcknowledgmentsThis study was supported in part by a grant from the Donald W.Reynolds Foundation (to Peter Libby, MD, Cardiovascular Division,Department of Medicine, Brigham and Women’s Hospital). Theauthors thank Vincent M. Lok, BA, and Matthias Nahrendorf, MD(Center for Molecular Imaging Research, Massachusetts GeneralHospital), for assistance with quantitative image analyses.
DisclosuresNone.
References1. Eisenberg LM, Markwald RR. Molecular regulation of atrioventricular
valvuloseptal morphogenesis. Circ Res. 1995;77:1–6.2. Schroeder JA, Jackson LF, Lee DC, Camenisch TD. Form and function of
developing heart valves: coordination by extracellular matrix and growthfactor signaling. J Mol Med. 2003;81:392–403.
3. Hurle JM. Scanning and light microscope studies of the development ofthe chick embryo semilunar heart valves. Anat Embryol (Berl). 1979;157:69–80.
4. Hurle JM, Colvee E, Blanco AM. Development of mouse semilunarvalves. Anat Embryol (Berl). 1980;160:83–91.
5. Hurle JM, Colvee E. Changes in the endothelial morphology of thedeveloping semilunar heart valves: a TEM and SEM study in the chick.Anat Embryol (Berl). 1983;167:67–83.
6. Rabkin E, Aikawa M, Stone JR, Libby P, Schoen FJ. Activated interstitialmyofibroblasts express catabolic enzymes and mediate matrix remodelingin myxomatous heart valves. Circulation. 2001;104:2525–2532.
7. Rabkin-Aikawa E, Aikawa M, Farber M, Kratz JR, Garcia-Cardena G,Kouchoukos NT, Mitchell MB, Jonas RA, Schoen FJ. Clinical pulmonaryautograft valves: pathologic evidence of adaptive remodeling in the aorticsite. J Thorac Cardiovasc Surg. 2004;128:552–561.
8. Rabkin-Aikawa E, Farber M, Aikawa M, Schoen FJ. Dynamic andreversible changes of interstitial cell phenotype during remodeling ofcardiac valves. J Heart Valve Dis. 2004;13:841–847.
9. Rabkin E, Hoerstrup SP, Aikawa M, Schoen FJ. Evolution of cell phe-notype and extracellular matrix in tissue-engineered heart valves duringin-vitro maturation and in-vivo remodeling. J Heart Valve Dis. 2002;11:308–314.
10. Aikawa M, Rabkin E, Okada Y, Voglic SJ, Clinton SK, Brinckerhoff CE,Sukhova GK, Libby P. Lipid lowering by diet reduces matrix metallo-proteinase activity and increases collagen content of rabbit atheroma: apotential mechanism of lesion stabilization. Circulation. 1998;97:2433–2444.
11. Schurch W, Seemayer TA, Gabbiani G. The myofibroblast: a quartercentury after its discovery. Am J Surg Pathol. 1998;22:141–147.
12. Aikawa M, Sivam PN, Kuro-o M, Kimura K, Nakahara K, Takewaki S,Ueda M, Yamaguchi H, Yazaki Y, Periasamy M. Human smooth musclemyosin heavy chain isoforms as molecular markers for vascular devel-opment and atherosclerosis. Circ Res. 1993;73:1000–1012.
13. Hiss J, Hirshberg A, Dayan DF, Bubis JJ, Wolman M. Aging of woundhealing in an experimental model in mice. Am J Forensic Med Pathol.1988;9:310–312.
14. Rich L, Whittaker P. Collagen and picrosirius red staining: a polarizedlight assessment of fibrillar hue and spatial distribution. Br J Morphol Sci.2005;22:97–104.
15. Deguchi J, Aikawa E, Libby P, Vachon JR, Inada M, Krane SM,Whittaker P, Aikawa M. MMP-13/collagenase-3 deletion promotescollagen accumulation and organization in mouse atherosclerotic plaques.Circulation. 2005;112:2708–2715.
16. Whittaker P, Kloner RA, Boughner DR, Pickering JG. Quantitativeassessment of myocardial collagen with picrosirius red staining andcircularly polarized light. Basic Res Cardiol. 1994;89:397–410.
17. Whittaker P, Canham PB. Demonstration of quantitative fabric analysisof tendon collagen using two-dimensional polarized light microscopy.Matrix. 1991;11:56–62.
Aikawa et al Human Cardiac Valve Maturation 1351
by guest on July 20, 2015http://circ.ahajournals.org/Downloaded from

18. Oldenbourg R, Mei G. New polarized light microscope with precisionuniversal compensator. J Microsc. 1995;180:140–147.
19. Batschelet E, Hillman D, Smolensky M, Halberg F. Angular-linear cor-relation coefficient for rhythmometry and circannually changing humanbirth rates at different geographic latitudes. Int J Chronobiol. 1973;1:183–202.
20. Schoen FJ. Aortic valve structure-function correlations: role of elasticfibers no longer a stretch of the imagination. J Heart Valve Dis. 1997;6:1–6.
21. Johnson P, Maxwell DJ, Tynan MJ, Allan LD. Intracardiac pressures inthe human fetus. Heart. 2000;84:59–63.
22. Maron BJ, Hutchins GM. The development of the semilunar valves in thehuman heart. Am J Pathol. 1974;74:331–344.
23. Poelmann RE, Mikawa T, Gittenberger-de Groot AC. Neural crest cells inoutflow tract septation of the embryonic chicken heart: differentiation andapoptosis. Dev Dyn. 1998;212:373–384.
24. Lincoln J, Alfieri CM, Yutzey KE. Development of heart valve leafletsand supporting apparatus in chicken and mouse embryos. Dev Dyn.2004;230:239–250.
25. Fisher SA, Langille BL, Srivastava D. Apoptosis during cardiovasculardevelopment. Circ Res. 2000;87:856–864.
26. Weston MW, Yoganathan AP. Biosynthetic activity in heart valve leafletsin response to in vitro flow environments. Ann Biomed Eng. 2001;29:752–763.
27. Tamura K, Jones M, Yamada I, Ferrans VJ. Wound healing in the mitralvalve. J Heart Valve Dis. 2000;9:53–63.
28. Shepherd JV, Vanhoutte PM. The Human Cardiovascular System: Factsand Concepts. New York, NY: Raven Press; 1979: 270–274.
29. Butcher JT, Penrod AM, Garcia AJ, Nerem RM. Unique morphology andfocal adhesion development of valvular endothelial cells in static andfluid flow environments. Arterioscler Thromb Vasc Biol. 2004;24:1429–1434.
30. Schoen FJ. Blood vessels. In: Kumar V, Fausto N, Abbas A, eds. Robbinsand Cotran Pathologic Basis of Disease. 7th ed. Philadelphia, Pa: WBSaunders; 2004:511–554.
31. Mitchell RN, Jonas RA, Schoen FJ. Pathology of explanted cryopreservedallograft heart valves: comparison with aortic valves from orthotopicheart transplants. J Thorac Cardiovasc Surg. 1998;115:118–127.
32. Muller AM, Cronen C, Kupferwasser LI, Oelert H, Muller KM,Kirkpatrick CJ. Expression of endothelial cell adhesion molecules onheart valves: up-regulation in degeneration as well as acute endocarditis.J Pathol. 2000;191:54–60.
33. Ghaisas NK, Foley JB, O’Briain S, Crean P, Kelleher D, Walsh M.Adhesion molecules in nonrheumatic aortic valve disease: endothelialexpression, serum levels and effects of valve replacement. J Am CollCardiol. 2000;36:2257–2262.
34. Dvorin EL, Jacobson J, Roth SJ, Bischoff J. Human pulmonary valveendothelial cells express functional adhesion molecules for leukocytes.J Heart Valve Dis. 2003;12:617–624.
35. Rabkin E, Schoen FJ. Cardiovascular tissue engineering. CardiovascPathol. 2002;11:305–317.
36. Rabkin-Aikawa E, Mayer JE Jr, Schoen FJ. Heart valve regeneration. AdvBiochem Eng Biotechnol. 2005;94:141–179.
37. Mol A, Bouten CV, Baaijens FP, Zund G, Turina MI, Hoerstrup SP.Tissue engineering of semilunar heart valves: current status and futuredevelopments. J Heart Valve Dis. 2004;13:272–280.
1352 Circulation March 14, 2006
by guest on July 20, 2015http://circ.ahajournals.org/Downloaded from

Aikawa and Frederick J. SchoenElena Aikawa, Peter Whittaker, Mark Farber, Karen Mendelson, Robert F. Padera, Masanori
Implications for Postnatal Adaptation, Pathology, and Tissue EngineeringHuman Semilunar Cardiac Valve Remodeling by Activated Cells From Fetus to Adult:
Print ISSN: 0009-7322. Online ISSN: 1524-4539 Copyright © 2006 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation doi: 10.1161/CIRCULATIONAHA.105.591768
2006;113:1344-1352Circulation.
http://circ.ahajournals.org/content/113/10/1344World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circ.ahajournals.org//subscriptions/
is online at: Circulation Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer this process is available in the
click Request Permissions in the middle column of the Web page under Services. Further information aboutOffice. Once the online version of the published article for which permission is being requested is located,
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the EditorialCirculationin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on July 20, 2015http://circ.ahajournals.org/Downloaded from
Related Documents











